Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 60, 2002
Effekter af sprøjtemiddelafdrift på bærbærende buske og træer som indikator for biodiversitetsforandringer
Pilotforsøg som oplæg til forsøgsplanlægning
Indholdsfortegnelse
Forord
Dette projekt blev iværksat som et pilotforsøg. Pilotforsøget skulle understøtte planlægningen af en undersøgelse af effekten af simuleret afdrift af et ukrudtsmiddel til tjørnehegn. Dette indebar at bestemme om det er sandsynligt at der vil være en effekt, hvor mange prøver der i givet fald skulle tages, og hvilken prøvetagningsstrategi der var det bedste økonomiske valg. Rapporten beskriver resultatet af disse overvejelser og deres grundlag.
Vi vil gerne takke Birgit Kristensen og Knud Erik Nielsen for hjælp i projektet. Birgit Kristensen med artsbestemmelsen af insekter og råd vedrørende relevant litteratur samt Knud Erik Nielsen for at have udført en jordbundskarakterisering af forsøgshegnene.
Vi vil endvidere meget gerne takke styregruppen for et godt samarbejde og konstruktiv kritik gennem alle projektets faser. Styregruppen var sammensat af følgende medlemmer:
Jørn Kirkegaard, Miljøstyrelsen (formand)
Claus Hansen, Miljøstyrelsen
Jens Mossin, Miljøstyrelsen
Heidi Buur Holbeck, Landbrugets Rådgivningscenter
Niels Lindemark, Dansk Planteværn
Kristian Kristensen, DJF, Forskningscenter Foulum
Ditte Olrik, Forskningscenter for Skov- og Landskab
Knud Tybirk, Danmarks Miljøundersøgelser
Christian Kjær, Danmarks Miljøundersøgelser
Tirsdag d. 7. maj 2002
Christian Kjær
Morten Strandberg
Niels Elmegaard
Søren Larsen
Sammenfatning og konklusioner
Det er kendt at tætheden og biodiversiteten af ukrudt, insekter og fugle på markfladerne påvirkes af brugen af sprøjtemidler. Sprøjtemidlerne påvirker imidlertid også omkringliggende habitater såsom ruderater, hegn, skovkanter, markkanter etc. Der findes ikke undersøgelser af effekter af sprøjtemiddelafdrift på træer og buske i hegn.
I nærværende projekt anvendes tjørn som repræsentant for hegnets træer, idet tjørn er hyppigt forekommende i danske læhegn og desuden udgør en del af fødegrundlaget for mange leddyr. Endvidere ved man fra England at tjørnebær er af stor betydning som fødekilde for fugle.
Formålet med projektet
Vi ønsker at undersøge og kvantificere, hvorledes herbicidet metsulfuron-methyl umiddelbart påvirker flerårige bærbærende planters reproduktion. For at designe et forsøg optimalt er det nødvendigt at have kendskab til variationen på de udvalgte måleparametre for hermed at kunne estimere det nødvendige prøveantal ved hjælp af statistiske styrkeberegninger.
Hovedformålet med pilotprojektet har således været at bestemme den nødvendige prøvestørrelse for statistisk at detektere en given effekt på produktionen af bladbiomasse samt for reproduktionen udtrykt ved blomster- og frugtproduktion. Disse to mål fungerer som udtryk for tilgængelighed af føde for herbivore arter, f.eks. insekter og fugle.
Det var endvidere målet at bestemme effekten af afdrift af herbicid til hegn af engriflet hvidtjørn ved begyndende frugtsætning. Dette blev gjort vha. kontrollerede sprøjteforsøg i felten hvor effekten af metsulfuron på antallet af bær blev bestemt.
Endelig blev der gennem sæsonen indsamlet insekter tilknyttet tjørnehegn.
Alle disse resultater danner baggrund for planlægning af undersøgelser af mulige konsekvenser af herbicidafdrift til tjørnehegn.
Resultater
Prøvestørrelser
De statistiske styrkeberegninger viste at der er store forskelle mellem forskellige endpoints (blade, knopper, blomster, grønne bær, modne bær) i hvor mange sidegrene per træ og hvor mange træer per hegn det er nødvendigt at indsamle for at opnå en statistisk styrke på 80%. Det var tydeligt for at det nødvendige prøveantal vil være lavest ved indsamling af få sidegrene per træ men med lidt flere træer per hegn. Det var mere variabelt hvorledes et øget antal hegn påvirkede antallet af prøver.
De modne bær er klart den mest ressourcekrævende måleparameter, men også den mest relevante for bærædende fugle. For de modne bær er det kun muligt at opnå en statistisk styrke på 80% for vægten af bærrene i sideskuddet, hvis der indsamles 20 sideskud fra 26 træer i et enkelt hegn, hvilket ligeledes vil være ressourcemæssigt uforsvarligt. Samtidig er det også oplagt at inddragelsen af flere hegn vil øge undersøgelsens generelle værdi. Det blev derfor for alle endpoints besluttet at indsamle fra mindst 4 hegn. Dette betød samtidig at der kun skulle indsamles en prøve af hver responsvariabel pr. træ. Til gengæld skulle der så indsamles fra mange træer, hvorfor der skulle findes lange homogene tjørnehegn. En samlet oversigt over det endelige valg af forsøgsdesign kan ses nedenfor i Tabel 1.
Tabel 1
Oversigt over det valgte design til prøveindsamling i 2002 ved henholdsvis
forårs- og sommersprøjtning, samt andet års indsamlinger fra begge sprøjtninger. De
trufne valg er baseret på ønsket om at kunne påvise effekter �
20% ved et signifikansniveau på 5% og en styrke på 80%.
Blade |
Knopper |
Blomster |
Grønne bær |
Modne bær |
|
Forår 2002 |
|||||
Hegn |
4 |
4 |
4 |
8 |
8 |
Træer/hegn |
4 |
10 |
13 |
13 |
13 |
Sideskud/træ |
1 |
1 |
1 |
1 |
1 |
Sommer 2002 |
|||||
Hegn |
4 |
8 |
8 |
||
Træer/hegn |
4 |
13 |
13 |
||
Sideskud/træ |
1 |
1 |
1 |
||
Forår 2003 |
|||||
Hegn |
4 |
4 |
4 |
8 |
8 |
Træer/hegn |
4 |
10 |
13 |
13 |
13 |
Sideskud/træ |
1 |
1 |
1 |
1 |
1 |
Sommer 2003 |
|||||
Hegn |
4 |
4 |
4 |
8 |
8 |
Træer/hegn |
4 |
10 |
13 |
13 |
13 |
Sideskud/træ |
1 |
1 |
1 |
1 |
1 |
Økonomiske beregninger
For at opnå den størst mulige sandsynlighed for en høj generel værdi af pilot-
forsøgets efterfølger, sigtes der som nævnt mod indsamlinger i så mange hegn som
praktisk og økonomisk muligt. Derfor blev de ressourcer der skal til for at måle en 20%
effekt ved et signifikansniveau på 0,05 og en statistisk styrke på 80% beregnet for
indsamlinger i op til 16 hegn.
Beregningen viste at indsamlinger i et enkelt hegn var meget tidskrævende fordi der skulle indsamles prøver fra et meget stort antal træer. Der ville i givet fald gå 9329 timer på at indsamle data. Tidsmæssigt ville indsamlinger i henholdsvis 4 og 8 hegn ikke adskille sig væsentligt fra hinanden, og de ville begge ligger under antallet af indsamlinger i 1, 2 eller 16 hegn. I sidste ende blev en kombination af 4 og 8 hegn valgt således at nogle endpoints skulle indsamles i 4 hegn og andre i 8 hegn (Tabel 1). Det antal timer der skal bruges for at gennemføre det forsøgsdesign der er skitseret i Tabel 1 er vist i Tabel 2.
Tabel 2
Tidsforbrug (timer) knyttet til forsøgsdesignet i Tabel 1.
|
2002 |
2003 |
Blade |
72 |
72 |
Knopper |
|
180 |
Blomster |
140 |
234 |
Umodne bær |
489 |
468 |
Modne bær |
489 |
468 |
Sprøjtning |
52 |
|
Chop |
|
100 |
Insekter |
291 |
|
Insekticid |
|
72 |
Udbytte |
|
59 |
Kørsel |
80 |
160 |
I alt |
1613 |
1813 |
Sprøjteforsøget
Der blev fundet en klar negativ effekt ved at eksponere engriflet hvidtjørn for metsulfuron i doser op til 40% af anbefalet markdosis. Sammenhængen kan beskrives ved følgende ligning:
Andel tilbageblevne bær = 0,287 - 0,0067 x Dosis
hvor "Dosis" er % af den anbefalede markdosis og "Andel tilbageblevne bær" er den relative mængde af bær der var tilbage ved høst 22 dage efter sprøjtning. Ovenstående relation er blevet etableret på basis af optælling af både beskadigede og ubeskadigede bær. Nedenstående figur viser hvorledes sammenhængen mellem eksponering og andelen af bær der ikke falder af afhænger af om de insektangrebne bær indregnes eller ej.
Figur 1
Sprøjteforsøgets resultater præsenteret som andel tilbageblevne bær ved forsøgets
afslutning som funktion af dosis.
Dataanalysen af pilotforsøgets sprøjteforsøg viste at det er vigtigt at opgøre antallet af angrebne bær så de kan udelades i nogle analyser. Det var således nødvendigt at korrigere for de frugter der var angrebet på sprøjtetidspunktet da der for disse var et mindre nedfald end for de tilsvarende bær uden insekter. Samlet kan man sige at resultaterne af dette meget begrænsede sprøjteforsøg viser at:
| Vi kan forvente at finde udtalte effekter på bærmængden ved doser ned til 10 procent af markdosis | |
| Mængden af insektangreb er af stor betydningen for tolkning af resultater |
Insekter
Der blev fundet 26 arter af insekter i de udvalgte forsøgshegn. Specielt var fundet af fire billearter med tilknytning til hvidtjørn interessant på grund af den effekt de har på tjørn og på grund af deres relativt høje hyppighed. Anthonomus pedicularius og A. sorbi er begge snudebiller som angriber tjørnens blomster og frugtknopper i maj-juni. En anden snudebille, Rhynchites aequatus, angriber den grønne frugt og frugten falder til jorden, hvor larven overvintrer. Den voksne bille klækker næste forår. Endelig er der tjørnebladbillen, Lochmaea crataegi, hvis larver gnaver af bladene. Den yngler så vidt vides udelukkende på hvidtjørn. De voksne biller træffes på hvidtjørn fra april – juni.
Pilotforsøgets indsamlinger viser aktuelt at der er en række insekter der angriber blomsterknopper og bær. De forårsager ofte at blomster og bær aborteres. Arterne angriber på forskellige tidspunkter og derfor skal indsamlingerne af tjørnens reproduktive dele til bestemmelse af insektmængden ligge med relativt korte intervaller for at opnå en realistisk bestemmelse. Da nedfaldet af knopper, blomster og bær tildels kan være betinget af insekter, er det vigtigt at bestemme hvor stor en andel af de reproduktive enheder der er angrebet og hvor mange af de nedfaldne blomster og bær der er angrebet. Der skal altså indsamles knopper, blomster og bær gennem sæsonen. Disse dissekeres efterfølgende for at bestemme om de er eller har været angrebet af insekter. Derudover opsættes bakker for at opsamle nedfaldne knopper, blomster og bær.
Summary and conclusions
It is known from studies in arable fields that the density and biodiversity of weeds, insects, and birds are affected by the use of pesticides. However, pesticides do not only affect species within the fields, but also organisms in adjacent habitats like hedgerows, field edges, and forest lots. The effect of spray drift on trees and shrubs is not known.
This project uses hawthorn as a representative of hedgerow trees as it is commonly found in Danish hedgerows, and supports a wide range of insect species. It is known from English studies that hawthorn berries are an important food item for birds.
Aim of the project
We want to study and quantify the effects of herbicide spray drift onto perennial berry-producing plants. In order to design such an experiment optimally it is necessary to know the variability of the selected response variables. This measure is used in a statistical power analysis in order to calculate the necessary sample size.
Therefore, the primary aim of the project is to assess the necessary sample size in order to document statistically a specified effect level on the production of leaf biomass and reproductive endpoints (i.e. flowers and berries). These two endpoints represent the availability of food for herbivorous insects and birds.
In addition the pilot experiments should establish the effects of herbicide treatment on hawthorn at the point of berry production. A controlled spray trial with the herbicide metsulfuron assessed the effects on the berries.
Finally insects associated to the hawthorn hedgerows were collected throughout the season.
All results were used in the planning of an experiment to establish the possible consequences of herbicide spray drift on biodiversity.
Results
The statistical power analyses showed that samples needed were highly variable for different endpoints (leaves, buds, flowers, green berries and mature berries). For leaves, buds and flowers it was possible to obtain a high statistical power with relatively few trees, even in a single hedgerow. For juvenile berries it was also possible to limit the set-up to only one hedgerow. Mature berries were the most variable endpoint and therefore the most resource demanding. A summary of the results of statistical power analyses is presented in Table 1.
Table 1
Summary of the minimum number of samples needed for a statistical power
of at least 80% when the number of hedgerows, developmental stage of the
hawthorn and the response variable is given. The figures are the result of
the statistical power analyses performed in Chapter 2.
|
Leaves |
Flower buds |
Flowers |
Green berries |
Ripe berries |
Spring 2002 |
|
|
|
|
|
Hedgerow |
4 |
4 |
4 |
8 |
8 |
Trees/hedgerow |
4 |
10 |
13 |
13 |
13 |
Shootlets/tree |
1 |
1 |
1 |
1 |
1 |
Summer 2002 |
|
|
|
|
|
Hedgerow |
4 |
|
|
8 |
8 |
Trees/hedgerow |
4 |
|
|
13 |
13 |
Shootlets/tree |
1 |
|
|
1 |
1 |
Spring 2003 |
|
|
|
|
|
Hedgerow |
4 |
4 |
4 |
8 |
8 |
Trees/hedgerow |
4 |
10 |
13 |
13 |
13 |
Shootlets/tree |
1 |
1 |
1 |
1 |
1 |
Summer 2003 |
|
|
|
|
|
Hedgerow |
4 |
4 |
4 |
8 |
8 |
Trees/hedgerow |
4 |
10 |
13 |
13 |
13 |
Shootlets/tree |
1 |
1 |
1 |
1 |
1 |
Economy
We wanted to have the highest confidence in the conclusions drawn from a new experiment. Therefore we aimed to sample as many hedgerows as practically and economically feasible. Consequently, we calculated the necessary resources needed to measure a 20% reduction with a significance level of 0.05 and a statistical power of 80% for both types of response variables.
The calculation showed that sampling a single hedgerow would be much more time consuming than collecting samples from several hedgerows because sampling then should be made from a high number of trees. The resulting design is presented in Table 1 and the cost (hours) associated with this design is presented in Table 2.
Table 2.
Cost (man-hours) associated with the design presented in Table
1.
|
2002 |
2003 |
Leaves |
72 |
72 |
Flowering buds |
|
180 |
Flowers |
140 |
234 |
Green berries |
489 |
468 |
Ripe berries |
489 |
468 |
Herbicide application |
52 |
|
Chopping |
|
100 |
Insect sampling |
291 |
|
Insecticide application |
|
72 |
Production |
|
59 |
Transport |
80 |
120 |
Total |
1613 |
1813 |
Spraying experiment
It was found that the herbicide metsulfuron caused a clear negative effect on hawthorn when the plant was exposed to dosages between 10 and 40% of the recommended field rate. The dose � response relationship can be described by means of the following equation:
Proportion of berries present = 0.287 � 0.0067 x Dose
where �Dose� is the percentage of the recommended field rate, and �Proportion of berries present� is the relative amount of berries remaining on the tree 22 days after spraying. The present dose � response relationship was established on the basis of counting both berries attacked by insects and undamaged berries. Figure 1 (below) shows that the results are highly dependent on whether the berries were attacked.
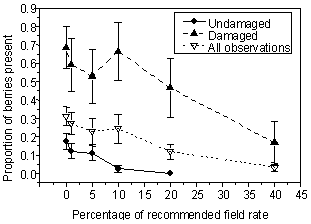
Figure 1
Relationship between the spray dose and the proportion of
berries remaining on hawthorn 22 days after spraying with metsulfuron.
This experiment shows us that:
| Effects on berry production can be expected at dosages of 10 percent of the recommended field rate and higher | |
| The herbivore impact is highly important for the assessment of the results |
Insects
Twenty-six species of insects were collected. In particular three of these are interesting due to their effect on hawthorn and because there were large numbers of them. Both Anthonomus pedicularius and A. sorbi are weevils that attack flowers and early berries in May and June. It is not known to what extent these insects provoke abortion of berries; however, our data show that there is a high loss of flowers. Another weevil, Rhynchites aequatus, attacks the green berries and as a result the berries are lost.
The pilot experiment shows that there is a number of insect species, which damage early flowers and berries. The different species attack at different times of the season. Therefore it is necessary to sample buds, flowers and berries throughout the season. Furthermore lost units should be sampled in order to assess the proportion damaged by insects.
1. Biodiversitet i hegn og sprøjtemiddeleffekter
| 1.1 | Baggrund |
| 1.2 | Forsøgsorganisme |
| 1.3 | Sprøjtemidler |
| 1.4 | Formålet med projektet |
1.1 Baggrund
Det er kendt at tætheden og biodiversiteten af ukrudt, insekter og fugle på markfladerne påvirkes af brugen af sprøjtemidler. Der findes en række danske undersøgelser (primært i kornafgrøder) der direkte såvel som indirekte har undersøgt sprøjtemidlers indvirken på markens dyre- og planteliv samt sammenlignet konventionel og økologisk dyrkning (for eksempel: samt det igangværende projekt "Monitering af effekten af Pesticidhandlingsplan II på naturindholdet i agerlandet 2000-2003").
Sprøjtemidlerne påvirker imidlertid ikke kun organismer der opholder sig på markfladen men også organismer i de omkringliggende habitater såsom ruderater, hegn, skovkanter, markkanter etc. . Der findes ikke undersøgelser af effekter af sprøjtemiddelafdrift på træer og buske i hegn (Maudsley 2000). Hegnene udgør et relativt uforanderligt element i et ellers meget omskifteligt landskab og fungerer derfor dels som en slags refugium for de vilde arter i agerlandet dels som leverandør af føde i vinterhalvåret. Specielt buske og træer er i kraft af deres lange levetid pålidelige opholds- og fourageringssteder for såvel insekter som fugle i agerlandet (. Ydermere muliggør hegnene på grund af deres anderledes fysiske struktur tilstedeværelsen af dyr som ellers ikke kunne eksistere i landbrugslandskabet. Hegnene har således på flere måder en positiv effekt på biodiversiteten i agerlandet. Denne funktion af hegnene kan dog reduceres ved (uhensigtsmæssig) brug af pesticider. ) fandt flere skovarter og en større biodiversitet i bunden af hegn ved ekstensivt dyrkede marker end i tilsvarende hegn ved konventionelt dyrkede marker. Ligeledes observerede en faldende biodiversitet i simuleret hegnsvegetation ved lave herbiciddoser. ) fandt negative effekter af sulfonylurea-midler og glyphosat på vækst og frugtsætning i kirsebær (Prunus avium) i forsøg der simulerede herbiciddrift. I nogle tilfælde blev der observeret negative effekter ved doser der svarer til 1% af anbefalet markdosis. Der er ikke lavet tilsvarende undersøgelser af herbicidafdrift på bærbærende buske og træer i Danmark eller Europa, men det kan have stor betydning for de mange fødekæder, der er knyttet til netop blomstring, frugtsætning og frøspredning.
I nærværende projekt anvendes tjørn som repræsentant for hegnets træer, idet tjørn er hyppigt forekommende i danske læhegn og desuden udgør en del af fødegrundlaget for mange leddyr . Endvidere ved man fra England at tjørnebær er af stor betydning som fødekilde for fugle (Snow og Snow 1988).
1.2 Forsøgsorganisme
Hos engriflet hvidtjørn Crataegus monogyna finder løvspring sted fra slutningen af april til begyndelsen af maj afhængig af vejret. Blomstring finder normalt sted fra slutningen af maj til første halvdel af juni. I slutningen af juni er blomstringen overstået. Frugten udvikles i løbet af sommeren og den begynder at blive rød i slutningen af august og modnes i løbet af september.
Der findes ikke danske undersøgelser af tjørns betydning som fødekilde for fugle. Fra England ved man at den har betydning for overlevelsen af mange lokale fugle og trækfugle i vinterhalvåret, specielt drosselarterne solsort, sjagger og vindrossel . Frugten spises sjældent i større mængde førend bærspisende trækfugle som for eksempel sjagger dukker op, typisk i løbet af oktober. Afhængig af fuglemængde, bærmængde og tilgængeligheden af andre arter af frugt kan tjørnens bær blive hængende hele vinteren. Buske som forsvares af bærspisende fugle vil typisk bære bær i længere tid end buske der ikke forsvares. Sådanne buske er i England set med mange bær helt ind i februar .
1.3 Sprøjtemidler
"Minimidler", herunder sulfonylurea-produkterne, anvendes i stigende mængder i dansk landbrug . Sulfonylurea-midlerne virker ved at blokere dannelsen af de grenede aminosyrer, valin, leucin og isoleucin. Det er kendt at minimidlerne, som en indirekte effekt, kan hæmme transporten af fotosynteseprodukter ud af behandlede blade . Dermed er det også sandsynligt, at der kan forekomme effekter på for eksempel frugtsætningen, hvis sprøjtningen finder sted på et kritisk tidspunkt. Det er ikke kendt om sulfonylurea-midlerne har en effekt på planterne året efter at de er blevet eksponeret.
1.4 Formålet med projektetVi ønsker at undersøge og kvantificere hvorledes udvalgte herbicider umiddelbart påvirker flerårige bærbærende planters reproduktion. Men vi vil endvidere også gerne undersøge om sprøjtetidspunktet er af betydning, om der kan spores effekter året efter behandling og om mængden af insekter er af betydning. Før vi kan designe vores forsøg optimalt er det nødvendigt at have kendskab til variationen på de udvalgte måleparametre, for at kunne estimere det nødvendige prøveantal udfra den effektstørrelse der skal kunne påvises statistisk.
Hovedformålet for pilotprojektet har således været at bestemme den nødvendige prøvestørrelse for statistisk at detektere en given effekt på de udvalgte responsvariable. Vi ønsker derfor at få et mål for variationen i bladbiomasse og antal reproduktive plantedele i usprøjtede hegn. Disse to mål fungerer som udtryk for tilgængelighed af føde for herbivore arter, f.eks. insekter og fugle.
Det var endvidere målet med det et-årige pilotprojekt at undersøge effekten af afdrift af sprøjtemidler (herbicider) til hegn af engriflet hvidtjørn ved begyndende frugtsætning. I kontrollerede sprøjteforsøg i felten bestemmes effekten af metsulfuron på bærproduktionen.
Endelig indsamles insekter tilknyttet tjørnehegn gennem sæsonen. Alle disse resultater skal danne baggrund for design af efterfølgende undersøgelser med henblik på at indikere mulige konsekvenser af herbicidafdrift for biodiversiteten i hegn.
2. Prøvestørrelser
2.1 Introduktion
Det er intentionen i dette afsnit at estimere det nødvendige prøveantal for forskellige målvariable ved hjælp af statistiske styrkeberegninger. Som forberedelse til et forsøg kan man udfra en kendt variation på måleparameteren, det ønskede signifikansniveau og den ønskede statistiske styrke beregne hvor mange prøver der skal indsamles. En forudsætning for at dette også fører til et godt statistisk funderet materiale er, at alle betydende faktorer for variansen er kendt. Et sådant variansskøn vil være et estimat, og det er derfor muligt at man til trods for en udregning af det nødvendige prøveantal ender med en lav statistisk styrke. Dette vil ske hvis enten forforsøget eller hovedforsøget gennemføres under atypiske forhold (abiotiske såvel som biotiske). Den forberedende styrkeberegning vil øge sandsynligheden for at få en statistisk god test. Det er vigtig at fastslå at statistisk signifikans ikke nødvendigvis er ensbetydende med biologisk relevans. I enhver test er det vigtigt at den der planlægger og gennemfører forsøget forholder sig til den "relevante effektstørrelse".
2.2 Metode
2.2.1 Indsamlinger
Der blev indsamlet blade, blomster og bær af tjørn fire gange gennem plantens fænologiske udvikling. Første indsamling blev fortaget den 30. maj hvor tjørnen havde udfoldet årsskuddets blade og der var blomsterknopper til stede. Anden høst blev foretaget 14. juni, hvor alle blomsterne var færdigudviklede. Første bærhøst blev indsamlet den 6. august, og der blev siden foretaget en efterårsindsamling af bær. Sidst i august blev de grene der skulle høstes i efterårsperioden udvalgt tilfældigt og en netpose (maskestørrelse på 1mm) blev monteret for at forhindre fugle i at spise bærrene. Disse bær blev høstet den 22. oktober.
Ved hver høst blev der indsamlet blad-, blomster- eller bærskud fra fire tjørnehegn beliggende nord for Silkeborg (jordbundsbeskrivelse findes i Bilag A). Der blev indsamlet fra fem tilfældigt udvalgte træer i hvert hegn. I hvert træ blev ovennævnte skudtyper indsamlet fra grene i 1, 3 og 5 meters højde. For hver højde blev der høstet mellem 1 og 10 sideskud. De lave antal indsamlinger forekom når den givne skudtype ikke var at finde i det givne træ i den givne højde. Indsamlingsprogrammet er gengivet i Tabel 2.1. Tjørns sideskud kan enten være langskud eller kortskud. Kortskuddene kan bære både blomster og blade, mens langskuddene udelukkende bærer blade.
Tabel 2.1
Tabel over indsamlinger i usprøjtede hegn.
Tidspunkt |
Respons- |
Skud- |
Loka- |
Træ- |
Grene/ |
Grene/ |
Bemærk- |
30. maj |
Blade |
Kortskud |
B |
4 |
5-10 |
25 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Langskud |
B |
5 |
5-15 |
55 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Kortskud |
C |
4 |
5-10 |
25 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Langskud |
C |
5 |
5-15 |
60 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Kortskud |
D |
5 |
15 |
75 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Langskud |
D |
1 |
1 |
1 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Kortskud |
E |
5 |
10-15 |
70 |
Blade tælles, afklippes og vejes |
30. maj |
Blade |
Langskud |
E |
2 |
1-5 |
6 |
Blade tælles, afklippes og vejes |
|
|
|
|
|
|
|
|
30. maj |
Knopper |
|
B |
5 |
1-15 |
45 |
Knopper tælles afklippes og vejes |
30. maj |
Knopper |
|
C |
5 |
5-15 |
54 |
Knopper tælles afklippes og vejes |
30. maj |
Knopper |
|
D |
5 |
10-15 |
62 |
Knopper tælles afklippes og vejes |
30. maj |
Knopper |
|
E |
5 |
11-15 |
67 |
Knopper tælles afklippes og vejes |
|
|
|
|
|
|
|
|
14. juni |
Blomster |
|
B |
5 |
10-19 |
75 |
Blomster tælles |
14. juni |
Blomster |
|
C |
5 |
16-23 |
88 |
Blomster tælles |
14 juni |
Blomster |
|
D |
5 |
15-23 |
86 |
Blomster tælles |
14. juni |
Blomster |
|
E |
5 |
18-23 |
104 |
Blomster tælles |
|
|
|
|
|
|
|
|
6. august |
Grønne frugter |
|
B |
5 |
9-20 |
79 |
Frugter tælles og vejes |
6. august |
Grønne frugter |
|
C |
5 |
9-20 |
81 |
Frugter tælles og vejes |
6. august |
Grønne frugter |
|
D |
5 |
20-30 |
121 |
Frugter tælles og vejes |
6. august |
Grønne frugter |
|
E |
5 |
26-30 |
143 |
Frugter tælles og vejes |
|
|
|
|
|
|
|
|
22. oktober |
Modne frugter |
|
B |
5 |
10 |
50 |
Frugter tælles og vejes |
22. oktober |
Modne frugter |
|
C |
5 |
10-30 |
100 |
Frugter tælles og vejes |
22. oktober |
Modne frugter |
|
D |
5 |
20-30 |
111 |
Frugter tælles og vejes |
22. oktober |
Modne frugter |
|
E |
5 |
30-40 |
170 |
Frugter tælles og vejes |
2.2.2 Forsøgsområder
Der blev udvalgt fem hegn til at bestemme variationen i blad, blomster og bærproduktion i hegn der ikke var eksponeret for sprøjtemidler. Det viste sig dog at en af markerne til trods for dette var blevet sprøjtet. Dette hegn, benævnt A, blev herefter taget ud af undersøgelserne. De resterende hegn i henholdsvis Buskelund og på Sinding Hedevej (nordlige Silkeborg) er afmærket på nedenstående luftfotos taget i 1995 (Figur 2.1- 2.3).
Figur 2.1
Luftfoto af de to hegn (B og C) fra Buskelund. (Kilde: Danmarks Digitale
Orthofoto 1995/Kampsax 1995).
Figur 2.2
Luftfoto af hegn D på Sinding Hedevej. (Kilde: Danmarks Digitale Orthofoto
1995/Kampsax 1995).
Figur 2.3
Luftfoto af hegn E på Sinding Hedevej. (Kilde: Danmarks Digitale Orthofoto
1995/Kampsax 1995).
2.2.3 Beregning af varianskomponenter
Der blev opstillet følgende hierakiske statistiske model som indeholder alle komponenter af variationen for de enkelte undersøgte responsvariable:
, (1)hvor Y er den analyserede responsvariabel. Indeks i står for niveau (i=1, 2, 3), j for læhegn (j=1, 2, 3, 4), k for trænummer i et givent læhegn (k=1, 2,..., tj) og l for sideskud i et givet træ (l = 1, 2, ...., sjk). Bj er varianskomponenten for læhegn, Cjk er varianskomponenten for træer i et givent hegn og Djki er varianskomponenten for niveauer i et givet træ og et givet læhegn. Endelig er Eijkl fejlleddet (det vil sige variansen mellem sideskud). Vi antager, at
Bj ~ N (0,) , Cjk ~ N (0,) , Dijk ~ N (0, ), samt Eijkl ~ N (0,).
Desuden antages, at de enkelte stokastiske variable er indbyrdes uafhængige. Med disse antagelser er:
hvor står for gennemsnittet af Y, det vil sige
.
Vi har bestemt os for at udføre styrkeberegningerne i kun et niveau nemlig mellemhøjden, idet det vil være mere resourcekrævende at indsamle i højeste niveau og fordi hovedparten af sprøjtemiddelafdriften afsættes i de nederste dele af hegnet.
Dermed anvendes følgende model :
, (2),hvor Bj ~ N (0, ) , Cjk ~ N (0, ) , Ejkl ~ N (0, ) og Ejkl står for fejlleddet.
I denne model er
,Varianskomponenterne , og estimeres ved REML-metoden ved anvendelse af Proc Mixed i SASÓ (Verbela & Molenberghs 1997).
2.2.4 Beregning af styrke
Den statistiske model som er blevet benyttet ved styrkeberegningerne er
(3),hvor vi antager at Cijk ~ N (0,) , Eijkl~ N (0,), og indeks i står for behandling , j for læhegn, k for trænummer i et givent læhegn og l for sideskud i et givet træ (i=1,..., nb, j=1,..., nl k=1,..., nt og l=1,..., ns). Det vil sige at modellen ikke indeholder niveau og vi antager at effekten af læhegn ikke er tilfældig. Det vil sige at inddeling efter behandling og læhegn udgør et 2-sidet skema hvor der er nt � ns observationer i hver celle.
I vores rapport har vi antaget at hegn er en systematisk effekt. I den statistiske analyse er det muligt både at antage en systematisk eller en tilfældig effekt af hegn. Argumentet for at vælge hegn som en tilfældig faktor er, at de hegn som medtages i projektet er et tilfældigt udtag (random sample) af alle mulige hegn af en bestemt type. Generalisering til alle hegn af denne type er mulig men kun ved et udtag af et større antal hegn. Argumentet for at betragte hegn som en systematisk faktor kan være at man er interesseret i at teste forskelle mellem de enkelte hegn. Denne angrebsvinkel egner sig bedst til undersøgelser med få hegn. Generalisering er ikke direkte opnåelig. Konsekvensen af at vælge at effekten af hegn er tilfældig er at variansen på forskellen mellem to behandliger øges og hermed øges også kravet til antal prøver.
Ved styrkeberegningerne ønskes at kunne påvise en forskel på d mellem to givne behandlinger på et 5% signifikansniveau ved en ensidig test. Den ene behandling kan i dette tilfælde være kontrolbehandlingen, det vil sige ingen sprøjtning. Vi har valgt d = 20% i de udførte beregninger.
Antager vi ingen vekselvirkning mellem læhegn og behandlinger (abij = 0), så er spredningen på differensen mellem to behandlinger lig
, (4),hvor
nl: antal læhegn
nt: antal træer
ns: antal sideskud
Interaktioner mellem hegn og behandling kan forekomme fordi: 1) hegnene har forskellig struktur og hermed er der forskelle i sprøjtemiddelafsætningen, 2) de abiotiske forhold er forskellige mellem sprøjtningerne i de enkelte hegn eller 3) fordi hegnene er genetisk og aldersmæssigt forskellige. Antagelsen om ingen vekselvirkning mellem læhegn og behandlinger bygger på følgende: 1) forskellen i afsætning vil ikke påvirke analysen idet vi måler afsætningen det samme sted som prøvetagningen foregår. 2) de forskelle der vil være ved sprøjtningen er negligerbare i forhold til de effekter vi ønsker at dokumentere og endelig vil genetiske og aldersmæssige effekter være beskrevet af den systematiske effekt.
Størrelsen s diff kan beregnes ved at anvende estimater forog fundet ved anvendelse af formel (2).
På baggrund af beregninger som præsenteret ovenfor ønsker vi at beregne det antal prøver der er nødvendige for statistisk at kunne dokumentere en 20% forandring af den enkelte responsvariabel.
Projektet sigter på at etablere en dosis – respons sammenhæng for de enkelte responsvariable, men vælger at basere styrkeberegningerne på en t-test for ikke at være afhængig af en række doser der rammer inden for intervallet af 20 til 90% effekt. Hermed bliver vi, med det valgte prøveantal, i stand til at opnå en høj styrke selv med kun en brugbar testdose. Skulle testdoserne alle ligge i effektområdet mellem 20 og 90%, vil en regressionsanalyse, der har samme eller større statistisk styrke end en t-test, blive anvendt.
Styrkeberegningen blev gennemført for en ensidig t-test, fordi vi kun interesserer os for negative effekter af herbicidafdrift. Beregningen tog udgangspunkt i at vi ønsker at indsamle så mange prøver at vi kan detektere effekter (d ) på 20% eller derover med en statistisk styrke på 80% eller mere.
Styrken beregnes ved sandsynligheden:
styrke = P(tu,t � tu (0,05))
Det vil sige at sandsynligheden for at tu,t i en ikke-central t-fordeling med parametrene t og u er større end den kritiske t-værdi med u frihedsgrader.
Parametrene u og t er:
u = 2 nl (nt-1), bemærk at træ er forsøgsenheden,
, er ikke centralitetsparameter i den ikke-centrale t-fordelingVed beregningen af styrken er rådata transformeret på følgende måde:
| vægt transformeres med den naturlige logaritme | |
| antal transformeres med kvadratroden |
Dette har betydning for beregningen af d på følgende måde:
| logaritmetransformation: |
| kvadratrodstransformation: |
Vi har valgt at anvende nl: 1, 2, 4, 8 eller 16; nt: 2, 3, 4,....29; ns: 1, 3, 5, 10 eller 20.
2.3 Resultater
2.3.1 Varianskomponenter
I nedenstående Tabel 2.2 findes varianskomponent-estimaterne til ovenstående model (2).
Tabel 2.2
Tabel over de estimerede varianskomponenter for formel (2) for forskellige
måleparametre. Data er transformeret; ved antal anvendes og ved vægt anvendes log(x).
Tabelværdier på 0 fremkommer fordi nogle analyser gav negative varianskomponenter og
dette er ikke logisk muligt.
Måleparameter og enhed |
|
|
|
Blade på kortskud, antal |
0 |
0,00112 |
0,05057 |
Blade på kortskud, vægt |
0,04688 |
0,08041 |
0,14630 |
Blade på langskud, antal |
0 |
0,04672 |
0,02948 |
Blade på langskud, vægt |
0,01051 |
0,06068 |
0,03832 |
Knopper, antal |
0,00150 |
0,01192 |
0,08984 |
Knopper, vægt |
0 |
0,01645 |
0,09879 |
Blomster, antal |
0,04637 |
0,13423 |
0,19666 |
Bær, antal |
0,02054 |
0,02563 |
0,15479 |
Bær, vægt |
0,09276 |
0,10019 |
0,34209 |
Modne bær, antal |
0 |
0,03902 |
0,12721 |
Modne bær, vægt |
0 |
0,11474 |
0,25231 |
Tabel 2.3
Tabel over middelværdien af forskellige måleparametre i mellemniveau. n er antallet
af obervationer. im betyder: ikke målt.
Måleparameter og enhed |
n |
Antal |
Vægt, g |
Blade på kortskud |
50 |
5,540 |
0,140 |
Blade på langskud |
50 |
9,580 |
0,238 |
Knopper |
84 |
2,519 |
0,097 |
Blomster |
127 |
2,202 |
im |
Bær |
149 |
2,383 |
0,239 |
Modne bær |
140 |
2,286 |
0,338 |
2.3.2 Statistisk styrke
Den statistiske styrke blev beregnet for en række måleparametre ved forskellige prøvetagningsscenarier. Som tidligere nævnt har vi valgt følgende scenarier: Hegn (nl): 1, 2, 4, 8 eller 16; træer (nt): 1, 2, 3, 4,....29; og sidegrene (ns): 1, 3, 5, 10 eller 20 gennemført i alle kombinationer. Nedenfor er resultatet for sådanne styrkeberegninger præsenteret i Figur 2.3. Figuren er lavet på basis af data for grønne bær og vi har udvalgt de kombinationer hvor antallet af træer er 1, 2, 3, 4, 5, 6, 10, 15, 20, 25 og 29 for at forbedre læsbarheden af figuren.
Figur 2.4
Relationen mellem det antal sidegrene der høstes fra hver træ og den resulterende
statistiske styrke for antal grønne bær. Delfigurerne (A, B, C, D og E) repræsenterer
prøvetagninger i henholdsvis 1, 2, 4, 8 eller 16 hegn.
Nedenfor findes tabeller over det nødvendige prøveantal for at vi kan detektere effekter (d ) på 20% eller derover med en statistisk styrke på mindst 80% for forskellige responsvariable. Tabellerne 2.1 til 2.4 præsenteres således at hvert enkelt måleparameter præsenteres i forhold til deres placering i den fænologiske udvikling (blad, knopper, blomster, bær og modne bær). I tabellerne er der også opgjort det samlede antal prøver der skal indsamles og bearbejdes.
2.3.2.1 Blade
Bladene blev høstet fra både kort- og langskud. Da disse skudtyper er morfologisk forskellige holdes de adskilt i styrkeberegningen. Der var blevet målt både antal blade i et sideskud og den samlede vægt af bladene i sideskuddet (biomasse). For responsvariablen "Antal" (antallet af blade i det enkelte sideskud) var det muligt at opfylde de givne kriterier (påvise en effekt på 20% med et 5% signifikansniveau og en statistisk styrke på 80%) ved indsamlinger i 1,2,4, 8 eller 16 hegn (Tabel 2.4). For både kortskud og langskud viste det sig at der skulle samles færre prøver hvis der kun bliver høstet en sidegren per træ. Endvidere krævede kortskud flere prøver end langskud. For responsvariablen "Biomasse" kunne der ikke inden for vores scenarier opnås en statistisk styrke på 80% for kortskud, hvis der kun blev indsamlet prøver i et enkelt hegn. Det samme gjorde sig gældende for 2 hegn og 1 sideskud. For de øvrige kombinationer var det muligt at opnå en statistisk styrke på 80%. Igen var det tydeligt at det samlede prøveantal var lavest med de færreste høstede sidegrene, men der var ingen større forskelle mellem antal af testhegn.
Tabel 2.4
Tabel over antal træer der er nødvendige i det enkelte hegn for at kunne
påvise en effekt på 20% med et 5% signifikansniveau og en statistisk styrke på 80% når
antallet af sidegrene og antallet af hegn er givet. Måleparametrene er antallet eller
biomassen af blade på henholdsvis kortskud og langskud. im betyder at det ikke var muligt
at opnå en statistisk styrke på 80% med de kombinationer af #hegn, #træer og #sidegrene
vi har valgt. "Prøver" er det antal prøver der i alt skal analyseres (#hegn � # træer � # sidegrene). De skraverede
felter angiver den kombination der resulterer i færrest prøvetagninger.
Antal hegn |
Antal side- |
Antal |
Biomasse |
|||||||||
Kortskud |
Langskud |
Kortskud |
Langskud |
|||||||||
Træ- |
Prø- |
Træ- |
Prø- |
Træ- |
Prø- |
Træ- |
Prø- |
|||||
1 |
1 |
15 |
15 |
11 |
11 |
im |
im |
- |
- |
|||
1 |
3 |
7 |
21 |
8 |
24 |
im |
im |
27 |
81 |
|||
1 |
5 |
6 |
30 |
8 |
40 |
im |
im |
25 |
125 |
|||
1 |
10 |
4 |
40 |
7 |
70 |
im |
im |
24 |
240 |
|||
1 |
20 |
4 |
80 |
7 |
140 |
im |
im |
23 |
460 |
|||
|
|
|
|
|
|
|
|
|
|
|||
2 |
1 |
7 |
14 |
5 |
10 |
im |
im |
13 |
26 |
|||
2 |
3 |
3 |
18 |
4 |
24 |
24 |
144 |
13 |
78 |
|||
2 |
5 |
3 |
30 |
4 |
40 |
20 |
200 |
12 |
120 |
|||
2 |
10 |
2 |
40 |
3 |
60 |
17 |
340 |
12 |
240 |
|||
2 |
20 |
2 |
80 |
3 |
120 |
16 |
640 |
11 |
440 |
|||
|
|
|
|
|
|
|
|
|
|
|||
4 |
1 |
4 |
16 |
2 |
8 |
20 |
80 |
9 |
36 |
|||
4 |
3 |
2 |
24 |
2 |
24 |
12 |
144 |
6 |
72 |
|||
4 |
5 |
1 |
20 |
2 |
40 |
10 |
200 |
6 |
120 |
|||
4 |
10 |
1 |
40 |
2 |
80 |
8 |
320 |
6 |
240 |
|||
4 |
20 |
1 |
80 |
2 |
160 |
8 |
640 |
5 |
400 |
|||
|
|
|
|
|
|
|
|
|
|
|||
8 |
1 |
2 |
16 |
1 |
8 |
10 |
80 |
4 |
32 |
|||
8 |
3 |
1 |
24 |
1 |
24 |
6 |
144 |
3 |
72 |
|||
8 |
5 |
1 |
40 |
1 |
40 |
5 |
200 |
3 |
120 |
|||
8 |
10 |
1 |
80 |
1 |
80 |
4 |
320 |
3 |
240 |
|||
8 |
20 |
1 |
160 |
1 |
160 |
4 |
640 |
2 |
320 |
|||
|
|
|
|
|
|
|
|
|
|
|||
16 |
1 |
1 |
16 |
1 |
16 |
5 |
80 |
2 |
32 |
|||
16 |
3 |
1 |
48 |
1 |
48 |
3 |
144 |
1 |
48 |
|||
16 |
5 |
1 |
80 |
1 |
80 |
2 |
160 |
1 |
80 |
|||
16 |
10 |
1 |
160 |
1 |
160 |
2 |
320 |
1 |
160 |
|||
16 |
20 |
1 |
320 |
1 |
320 |
2 |
640 |
1 |
320 |
|||
2.3.2.2 Knopper
For at påvise en 20% effekt på enten antallet eller biomassen af knopper i et sideskud viser de styrkeberegninger der er præsenteret nedenfor (Tabel 2.5), at det ikke er muligt at nøjes med at høste 1 sidegren per træ i 1 hegn. For alle de øvrige kombinationer var det muligt at opnår den ønskede styrke. Antallet af prøver er lavest når der høstes få sidegrene i hvert træ. Responsvariablen "Biomasse" var generelt mere variabel og krævede dermed også flere prøver. Der var kun små forskelle mellem antal testhegn.
Tabel 2.5
Tabel over antal træer der er nødvendige for at kunne påvise en effekt på
20% med et 5% signifikansniveau og en statistisk styrke på 80% når antallet af sidegrene
og antallet af hegn er givet. Måleparameteren er antallet eller biomassen af
blomsterknopper. im betyder at det ikke var muligt at opnå en statistisk styrke på 80%
med de kombinationer af #hegn, #træer og #sidegrene vi har valgt "Prøver" er
det antal prøver der i alt skal analyseres (#hegn � # træer � # sidegrene). De skraverede felter angiver den kombination der
resulterer i færrest prøvetagninger.
Hegn |
Sidegrene |
Antal |
Biomasse |
||
Træer |
Prøver |
Træer |
Prøver |
||
1 |
1 |
im |
im |
im |
im |
3 |
16 |
48 |
18 |
54 |
|
5 |
11 |
55 |
14 |
70 |
|
10 |
8 |
80 |
10 |
100 |
|
20 |
6 |
120 |
8 |
160 |
|
2 |
1 |
19 |
38 |
21 |
42 |
3 |
8 |
48 |
9 |
54 |
|
5 |
5 |
50 |
7 |
70 |
|
10 |
4 |
80 |
5 |
100 |
|
20 |
3 |
120 |
4 |
160 |
|
4 |
1 |
10 |
40 |
10 |
40 |
3 |
4 |
48 |
4 |
48 |
|
5 |
2 |
40 |
3 |
60 |
|
10 |
2 |
80 |
2 |
80 |
|
20 |
1 |
80 |
2 |
160 |
|
8 |
1 |
4 |
32 |
5 |
40 |
3 |
2 |
48 |
2 |
48 |
|
5 |
1 |
40 |
1 |
40 |
|
10 |
1 |
80 |
1 |
80 |
|
20 |
1 |
160 |
1 |
160 |
|
16 |
1 |
2 |
32 |
2 |
32 |
3 |
1 |
48 |
1 |
48 |
|
5 |
1 |
80 |
1 |
80 |
|
10 |
1 |
160 |
1 |
160 |
|
20 |
1 |
320 |
1 |
320 |
|
2.3.2.3 Blomster
Det var kun antallet af blomster der blev registreret. Vægten kunne ikke registreres fordi mange af blomsterne havde tabt deres kronblade under prøvetagning og prøvebehandling. Ud fra data på antal blomster viste den statistiske styrkeberegning, at det ikke var muligt at opnå en statistisk styrke på 80% hvis der indsamles hhv. 1 eller 3 sideskud fra træer i et enkelt hegn (Tabel 2.6). For de øvrige scenarier kunne den statistiske styrke opnås. Antallet af indsamlinger var lavest ved indsamlinger et enkelt sideskud per træ og sammenlignelig mellem 2, 4, 8 eller 16 testhegn.
Tabel 2.6
Tabel over antal træer der er nødvendige for at kunne påvise en effekt på
20% med et 5% signifikansniveau og en statistisk styrke på 80% når antallet af sidegrene
og antallet af hegn er givet. Målepareameteren er antallet blomster. im betyder at det
ikke var muligt at opnå en statistisk styrke på 80% med de kombinationer af #hegn,
#træer og #sidegrene vi har valgt. "Prøver" er det antal prøver der i alt
skal analyseres (#hegn � # træer �
# sidegrene). De skraverede felter angiver den kombination der resulterer i færrest
prøvetagninger.
Hegn |
Sidegrene |
Træer |
Prøver |
1 |
1 |
im |
im |
1 |
3 |
im |
im |
1 |
5 |
28 |
140 |
1 |
10 |
25 |
250 |
1 |
20 |
24 |
480 |
2 |
1 |
27 |
54 |
2 |
3 |
16 |
96 |
2 |
5 |
14 |
140 |
2 |
10 |
12 |
240 |
2 |
20 |
12 |
480 |
4 |
1 |
13 |
52 |
4 |
3 |
8 |
96 |
4 |
5 |
7 |
140 |
4 |
10 |
6 |
240 |
4 |
20 |
6 |
480 |
8 |
1 |
6 |
48 |
8 |
3 |
4 |
96 |
8 |
5 |
3 |
120 |
8 |
10 |
3 |
240 |
8 |
20 |
3 |
480 |
16 |
1 |
3 |
48 |
16 |
3 |
2 |
96 |
16 |
5 |
1 |
80 |
16 |
10 |
1 |
160 |
16 |
20 |
1 |
320 |
Grønne bær
Denne styrkeberegning (Tabel 2.7) viser, at for at påvise en 20% effekt på antallet af bær i et sideskud skal vi for at opnå en statistisk styrke på 80% for begge responsvariable indsamle mindst 10 sideskud i hvert af 26 træer i et enkelt hegn. I 2 hegn skal der mindst indsamles 3 sideskud per træ (Tabel 2.6). Ellers er mønsteret som for de øvrige endpoints (blade, knopper og blomster) at det giver færrest indsamlinger at høste en sidegren per træ. Som det også fremgår af tabellen skal der tages færrest prøver ved at indsamle 1 sidegren fra hvert af 13 træer i hvert af 8 hegn.
Tabel 2.7
Tabel over antal træer der er nødvendige for at kunne påvise en effekt på
20% med et 5% signifikansniveau og en statistisk styrke på 80% når antallet af sidegrene
og antallet af hegn er givet. Måleparameteren er antallet eller biomassen af grønne
bær. im betyder at det ikke var muligt at opnå en statistisk styrke på 80% med de
kombinationer af #hegn, #træer og #sidegrene vi har valgt. "Prøver" er det
antal prøver der i alt skal analyseres (#hegn � # træer � # sidegrene). De skraverede felter angiver den kombination der
resulterer i færrest prøvetagninger.
|
|
Antal |
Biomasse |
||
Hegn |
Sidegrene |
Træer |
Prøver |
Træer |
Prøver |
1 |
1 |
- |
- |
- |
- |
1 |
3 |
- |
- |
- |
- |
1 |
5 |
- |
- |
- |
- |
1 |
10 |
26 |
260 |
- |
- |
1 |
20 |
21 |
420 |
- |
- |
2 |
1 |
- |
- |
- |
- |
2 |
3 |
24 |
144 |
23 |
138 |
2 |
5 |
13 |
130 |
21 |
210 |
2 |
10 |
13 |
260 |
19 |
380 |
2 |
20 |
10 |
400 |
18 |
720 |
4 |
1 |
27 |
108 |
17 |
68 |
4 |
3 |
12 |
144 |
11 |
132 |
4 |
5 |
8 |
160 |
10 |
200 |
4 |
10 |
6 |
240 |
9 |
360 |
4 |
20 |
5 |
400 |
9 |
720 |
8 |
1 |
13 |
104 |
8 |
64 |
8 |
3 |
6 |
144 |
5 |
120 |
8 |
5 |
4 |
160 |
5 |
200 |
8 |
10 |
3 |
240 |
4 |
320 |
8 |
20 |
2 |
320 |
4 |
640 |
16 |
1 |
7 |
112 |
4 |
64 |
16 |
3 |
3 |
144 |
2 |
96 |
16 |
5 |
2 |
160 |
2 |
160 |
16 |
10 |
1 |
160 |
2 |
320 |
16 |
20 |
1 |
320 |
2 |
640 |
2.3.2.5 Modne bær
Med de begrænsninger vi har sat for vores styrkeberegninger, dvs. for at opnå en statistisk styrke på 80% for vægten af de modne bær i sideskuddet skal der i et enkelt hegn indsamles 20 sideskud per træ i 29 træer. I 2 hegn skal der mindst indsamles 3 sideskud per træ i 26 træer. For 4, 8, og 16 hegn er alle kombinationer mulige. Prøvetallet minimeres ved at høste så få sideskud per træ som muligt. Responsvariablen "Antal" er er den mest variable for de modne bær. For de modne bær angiver beregningerne at der skal samles færrest prøver hvis der bliver indsamlet 1 sidegren fra hver af 13 træer i hvert af 8 hegn eller 1 sidegren fra hvert af 26 træer i hvert af 4 hegn.
Tabel 2.8
Tabel over antal træer der er nødvendige for at kunne påvise en effekt på
20% med et 5% signifikansniveau og en statistisk styrke på 80% når antallet af sidegrene
og antallet af hegn er givet. Måleparameteren er antallet og biomassen af modne bær. im
betyder at det ikke var muligt at opnå en statistisk styrke på 80% med de kombinationer
af #hegn, #træer og #sidegrene vi har valgt. "Prøver" er det antal prøver der
i alt skal analyseres (#hegn � # træer �
# sidegrene). De skraverede felter angiv er den kombination der resultere i færrest
prøvetagninger.
|
|
Antal |
Biomasse |
||
Hegn |
Sidegrene |
Træer |
Prøver |
Træer |
Prøver |
1 |
1 |
- |
- |
- |
- |
1 |
3 |
- |
- |
- |
- |
1 |
5 |
- |
- |
23 |
115 |
1 |
10 |
- |
- |
14 |
140 |
1 |
20 |
29 |
580 |
9 |
180 |
2 |
1 |
- |
- |
- |
- |
2 |
3 |
26 |
156 |
17 |
102 |
2 |
5 |
20 |
200 |
11 |
110 |
2 |
10 |
16 |
320 |
7 |
140 |
2 |
20 |
14 |
560 |
4 |
160 |
4 |
1 |
26 |
104 |
24 |
96 |
4 |
3 |
13 |
156 |
8 |
96 |
4 |
5 |
10 |
200 |
5 |
100 |
4 |
10 |
8 |
320 |
4 |
160 |
4 |
20 |
7 |
560 |
2 |
160 |
8 |
1 |
13 |
104 |
5 |
40 |
8 |
3 |
6 |
144 |
3 |
72 |
8 |
5 |
5 |
200 |
2 |
80 |
8 |
10 |
4 |
320 |
1 |
80 |
8 |
20 |
3 |
480 |
1 |
160 |
16 |
1 |
6 |
96 |
6 |
96 |
16 |
3 |
3 |
144 |
2 |
96 |
16 |
5 |
2 |
160 |
1 |
80 |
16 |
10 |
2 |
320 |
1 |
160 |
16 |
20 |
1 |
320 |
1 |
320 |
2.4 Konklusioner for kommende forsøg
De ovenstående styrkeberegninger viser entydigt at antallet af prøver bliver lavest når der høstes få sideskud per træ, men at det selvfølgelig kræver tjørnehegn med flere træer hvilket selvsagt begrænser antallet af egnede lokaliteter. Derudover er der andre omkostninger ved at tage prøver i flere hegn. Der følger en omkostningsberegning i Kapitel 6.
3. Biologisk relevans af effektniveau
3.1 Introduktion
I kapitel 2 valgte vi et effektniveau på 20% for vores styrkeberegninger. Formålet med nærværende kapitel er at evaluere om dette niveau er biologisk betydende for bærspisende fugle. Dette gør vi ved hjælp af viden og antagelser om danske tjørnehegns produktion af bær, bærrenes betydning for fugles vinterfouragering, og herbiciders påvirkning af frugtsætning hos bærbærende buske og træer. Afsnittet er opbygget således at det først gennemgår de forskellige delforudsætninger der indgår i ovennævnte vurdering. Dette gøres dels argumenterende, dels ved hjælp af den viden der findes rapporteret fra området. I tilfælde, hvor den eksisterende viden ikke er afklaret, har vi måttet træffe et argumenteret valg mellem de foreliggende resultater. Argumentet kan være at undersøgelsen er lavet under betingelser der er sammenlignelige med de danske forhold eller en vurdering af resultaternes pålidelighed.
3.1.1 Mængden af tjørnehegn
Det har ikke været muligt nøjagtigt at få fastslået hvor meget tjørn der er i Danmark. Vi ved det er en almindelig hegnsplante, både i markskel og omkring haver, og at den er almindelig i krat, specielt langs kyster. Et gæt baseret på hyppigheden af tjørnehegn i det område hvor vi fandt vores forsøgslokaliteter vil lyde på ca. 20.000 km tjørnehegn i Danmark eller ca. 500 m km-2. Dette svarer igen til at hvis Danmarks areal på ca. 42.000 km2 var en klods på 140 km x 300 km, så skulle man i gennemsnit støde på et tjørnehegn for hver anden kilometer hvis man gik på tværs af klodsen ~ 140/2 x 300 km = 21.000 km tjørnehegn. Til sammenligning findes der i Storbritannien angiveligt 430.000 km tjørnehegn, svarende til næsten 2.000 m km-2 (Sparks et al. 1996) eller en tæthed af tjørnehegn som er 4 gange vores estimat for Danmark. Dertil kommer så hvad der findes rundt omkring i krat og på overdrev m.m.
Antagelse: Der er 20.000 km tjørnehegn i Danmark
3.1.2 Bedømmelse af produktionen af tjørnebær i tjørnehegn i Danmark
Vi antager altså at der er 20.000 km tjørnehegn i Danmark. Med produktionen i Storbritannien in mente ved vi at der i 1 km tjørnehegn kan produceres mellem 47 kg og 3.488 kg tjørnebær afhængigt af hvor intensivt det klippes og beskæres (. Da de fleste danske tjørnehegn er høje (4 – 7 m) og oftest kun bliver klippet i op til 2-3 meters højde (dog ikke hvert år) og kun sjældent bliver skåret ned må vi formode at den gennemsnitlige bærproduktion er væsentligt højere end 47 kg km-1.
I vore beregninger har vi antaget at danske tjørnehegn som gennemsnit producerer 300 kg km-1. Dermed har vi en produktion på 300 kg km-1 � 20.000 km = 6.000.000 kg eller 6.000 tons tjørnebær produceret af danske tjørnehegn.
Antagelse: De 20.000 km tjørnehegn producerer i et gennemsnitsår 6.000 tons bær
3.1.3 Tjørnebær som fødekilde
Når man skal vurdere betydningen af at tjørns frugtsætning reduceres er det vigtigt at kende tjørns værdi som fødekilde for fugle. Til belysning af dette foreligger der materiale først og fremmest fra England (Tabel 3.1). Vi har ikke fundet danske undersøgelser som kan supplere dette.
Tabel 3.1
Frugtkød- og kerneandele af tjørnefrugter Crataegus monogyna ifølge , Herrera
(1987), og Eriksson og Ehrlen (1991).
|
Eriksson |
Sorensen |
Herrera |
Snow |
Diameter, mm |
8,6 |
|
9,3 |
10,7 |
Volumen, cm3 |
0,73 |
|
|
|
Længde, mm |
|
|
12,1 |
11,5 |
Frisk vægt, g |
0,476 |
0,3 |
0,68 |
0,63 |
Tørvægt, g |
|
|
0,29 |
|
Antal frø |
1 |
1 |
1 |
1 |
Frøets tørvægt, g |
|
|
0,12 |
|
Frøets vægt, g |
0,096 |
0,13 |
|
0,13 |
Frøets andel i % |
|
43 |
18 |
21 |
Vandindhold i pulp, % |
75,7 |
80 |
|
73 |
Pulp tørvægt, g |
|
0,05 |
0,17 |
|
Pulp tørvægt i % af total frisk vægt |
|
21 |
25 |
21 |
Fuentes (1994) målte et forhold på 1,4 til 1 mellem frugtkød og frø på tjørnebær
fra middelhavsområdet eller 42% frø og 58% pulp på vådvægtsbasis. Dette svarer til
den frøvægtsandel på 43% der er rapporteret for engelske tjørn (Sorensen 1984).
Andre undersøgelser i England har vist, at fra begyndelsen af oktober begynder bærmængden at falde (. Med udgangen af december var bærrene væk i den pågældende vinter (1998-99). De bær der blev spist først var de største så da man nåede frem til januar, var der kun de mindste bær tilbage. Vægten af 50 bær faldt således fra 20,4 g i starten af oktober til 15,3 g den 17. december, medens frugtkødets tørstofandel steg fra 24,2% til 29,3% i samme periode. Nogle få bær blev hængende helt til løvspring. Forfatterne formoder at dette skyldes at de var af ringe værdi for fuglene eller at deres ringe antal gjorde fødesøgningen ineffektiv. Ligeledes må det formodes at en del af den observerede ændring i bærrenes totalvægt og tørstofandel skyldes en lettere udtørring af frugtkødet i perioden.
Tabel 3.2
Energi og næringsindhold i tjørnebær (efter ,Herrera (1987), Eriksson og
Ehrlen (1991).
|
Snow - Herrera |
Snow - Herrera |
Eriksson |
Lipid |
9,3 |
2,30 |
1,0 |
Protein |
5,2 |
2,45 |
|
Nitrogen |
|
|
0,7 |
Kulhydrat |
4,0 |
72,40 |
9,0 |
Energi g tør pulp-1 |
3,24 |
|
|
Energi g frisk frugt-1 |
0,68 |
|
|
Sorensen (1984) har beregnet energimængden i 100 g tør pulp til at være 4,3 kcal
hvilket er ca. 2 størrelsesordner mindre end værdien i Tabel 3.2. Vi formoder at der må
foreligge en fejlberegning idet denne energimængde i pulpen ville betyde at en solsort
dagligt skulle æde ca. 1200 g tør pulp (se Tabel 3.5 og afsnit. 3.1.6
"Drosselfugles energibehov i dansk vinter"). Ud fra forsøg med solsorte angiver
at 66,2 % af energiindholdet i tjørnebær er metaboliserbart. I følge citater fra andre
undersøgelser ( varierer udnyttelsesgraden hos 5 drosselarter mellem 62 og 85 %.
Antagelse: Et gennemsnitsbær har en frisk vægt på 0,476 g, heraf udgør pulpen 0,38 g hvoraf 75,7% er vand og 24,3% eller 0,092 g er tørstof. Yderligere har vi i vore beregninger valgt at regne med at 75% af energien i tjørnens frugtkød kan udnyttes og at der er et energiindhold på 3,24 kcal g-1 tør pulp eller 0,68 kcal g-1 frisk frugt.
3.1.4 Hvilke fugle spiser tjørnebær
Det er primært drosselfugle som spiser tjørnebær (, men de spises også af andre fuglearter (Tabel 3.3). Nogle fugle spiser hele frugten (frugtædere), medens andre kun tager frugtkødet (pulpædere), endelig er der nogen få som spiser kernen (frøædere). I følge Hartley (1954) udgør tjørnefrugter den dominerende føde for sjagger, vindrossel og solsort, medens misteldrossel og sangdrossel foretrækker taks efterfulgt af tjørn og kristtorn.
Tabel 3.3
Fugle der spiser tjørnefrugter.
Fugleart |
Kategori |
Reference |
Vindrossel Turdus iliacus |
Frugtæder |
Hartley (1954), Sorensen (1981) Guitian (1992) |
Solsort Turdus merula |
Frugtæder |
(Hartley, (1954), Sorensen (1981) Fuentes (1994) |
Sjagger Turdus pilaris |
Frugtæder |
Sorensen (1981) Guitian (1992) |
Misteldrossel Turdus viscivorous |
Frugtæder |
|
Sangdrossel Turdus philomelos |
Frugtæder |
|
Ringdue Columba palumbus |
Frugtæder (frøæder) |
Sorensen (1981) |
Stær Sturnus vulgaris |
Frugtæder |
|
Rødhals Erithacus rubecula |
Frugtæder |
|
Silkehale Bombycilla garrulus |
Frugtæder |
|
Blåmejse Parus caeruleus |
Pulpæder |
|
Kernebider Coccothraustes coccothraustes |
Frøæder |
|
Collinge (1941) angiver at vild frugt udgør fra 26,5 til 30,5% af føden hos solsort.
Dette er set over hele året ,og det er derfor klart at den vilde frugt må have stor
betydning i efterår og vinter. Da solsort samtidig regnes som specialist i tjørnefrugter
(, er det sandsynligt at størstedelen af dens fødeindtag i perioden fra oktober til
januar er tjørnebær.
Antagelse: Det er hovedsagelig drosselfugle der spiser tjørnebær. Blandt drosselfuglene er solsorten den eneste som ikke trækker væk fra Danmark. Vi antager at den mængde bær der er tilbage når de øvrige drosler trækker væk er af betydning for solsortens vinteroverlevelse.
3.1.5 Mængden af fugle der spiser tjørnebær
Det er hovedsageligt drosselfugle som fouragerer i tjørnehegn. Derfor omfatter den følgende estimering af antallet af fugle der søger deres føde i tjørnehegn kun drosselfuglene.
Drosselfuglene i Danmark kan deles i to hovedgrupper standfugle og trækgæster. Solsorten er en standfugl, medens de øvrige drosler alt overvejende kommer igennem Danmark på deres træk fra Skandinavien til deres overvintringsområder. I følge Dansk Ornitologisk Forening er der ca. 2,25 mill. ynglepar af solsorte i Danmark. Med de unger de får og den dødelighed der finder sted, er der formentlig mindst fem millioner solsorte i den danske vinter. Det er dog langt fra alle solsortene der lever i tjørnehegn. I dagens Danmark er solsorten i høj grad blevet til en havefugl, og det er kun en mindre del af populationen som findes i mere naturlige habitater som hegn, krat og skov. Vi anslår at ca. 500.000 solsorte i en periode er afhængige af tjørnebær.
Antallet af trækfugle er det endnu sværere at anslå. Men der findes formentlig mindst 20 millioner andre drosler end solsorte i Sverige, Norge og Finland tilsammen (Asbirk 1997) (Tabel 3.4). Mange af disse passerer Danmark på deres trækrute. Derfor er det sandsynligt at ca. 10 mill. trækkende drosler passerer Danmark. Det vides ikke hvor længe de opholder sig her. Der er vel også tale om stor variation mellem år. På baggrund af antallet af potentielle trækfugle antager vi at der i gennemsnit er en halv mill. trækdrosler til stede hver dag fra midten af oktober til midten af november hvorefter antallet formodes at falde til nær nul omkring udgangen af november.
Tabel 3.4.
Ynglefugle af drosler i Skandinavien (antal mill. par) efter .
|
Sverige |
Finland |
Norge |
Danmark |
Sjagger |
0,75 – 1,5 |
0,8 – 1,2 |
1,0 – 3,0 |
|
Sangdrossel |
1,5 – 3,0 |
1,0 – 1,5 |
0,5 – 1,0 |
|
Vindrossel |
1,0 – 2,0 |
1,5 – 3,0 |
1,0 – 1,5 |
|
Misteldrossel |
0,075 – 0,2 |
0,05 – 0,08 |
0,01 – 0,05 |
|
Solsort |
1,0 – 2,0 |
0,15 – 0,2 |
0,1 – 1,0 |
2,25 |
Antagelse: Der passerer 10 mill. drosselfugle gennem Danmark i løbet af efteråret. Dertil kommer at der findes ca. 5 mill. danske solsorte som normalt bliver i landet.
3.1.6 Drosselfugles energibehov i dansk vinter
En fugl på ca. 100 g skal ved 0 ° C bruge ca. 50 kcal døgn-1 for at opretholde sin kropsvægt ved et lavt aktivitetsniveau . Tilsvarende skal en fugl på 130 g bruge 55 kcal døgn-1. Drosselfugle vejer typisk omkring 100 g i den del af sæsonen hvor der er modne tjørnebær (Tabel 3.5).
Tabel 3.5
Nogle bærspisende fugles vægt (efter Snow & Snow 1988).
Art |
Vægt (g) |
Sæson |
Solsort |
113,9 |
November – januar |
Sangdrossel |
95,9 |
November – februar |
Misteldrossel |
144,0 |
November – februar |
Sjagger |
117,9 |
September – januar |
Vindrossel |
64,9 |
September – november |
Stær |
92,1 |
November – februar |
Antagelse: Som gennemsnit skal drosselfugle bruge 50 kcal døgn-1 for at opretholde livet ved en lav aktivitet ved danske efterårstemperaturer.
3.1.7 Fødebehov i form af tjørnebær
Ved en udnyttelse af energien i føden på 75% og et energiindhold i tjørnebærs pulpdel på 3,24 kcal g-1 tør pulp kan følgende fødebehov i form af tjørnebær beregnes (Tabel 3.6).
Tabel 3.6
Bærspisende fugles energi- og fødebehov per døgn under forudsætning af at
energien skaffes fra tjørnebær (baseret på opgivelser hos (Snow og Snow 1988, Herrera
1998 og Eriksson, 1991).
Art
|
Energi- |
Tør pulp |
Frisk frugt |
Tør pulp |
Frisk pulp |
Frisk bær |
Energi udbytte |
Behov |
Behov |
Solsort |
52 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
102 |
214 |
Sang- |
50 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
98 |
206 |
Mistel- |
57 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
112 |
235 |
Sjag- |
52 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
102 |
214 |
Vin- |
45 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
88 |
185 |
Stær |
50 |
3,24 |
0,68 |
0,092 |
0,38 |
0,476 |
75 |
98 |
206 |
Det gennemsnitlige energibehov per døgn for en drossel udtrykt i gram tjørnebær er
således 100 g tjørnebær. Denne mængde benævnes et drosseldøgnækvivalent.
Dermed kan det beregnes at tjørnehegnenes 6.000 tons tjørnebær svarer til 60.000.000 drosseldøgn.
Antagelser: 100 g tjørnebær kan forsyne en gennemsnitsdrosselfugl med den nødvendige energi døgn-1. Denne mængde kaldes herefter et drosseldøgnækvivalent eller blot drosseldøgn. Mængden af tjørnebær i de danske tjørnehegn svarer til 60 millioner drosseldøgnsækvivalenter.
3.2. Beregninger
En kritisk størrelse i de kommende beregninger er hvor stor en andel af den tilstedeværende bærmængde der spises af trækgæsterne. Mængden er sandsynligvis variabel fra år til år. Den måde vi har valgt at anskueliggøre problemstillingen på, er ved at se på antallet af ubrugte drosseldøgn som funktion af reduktioner i bærmængden og som funktion af hvor mange bær de trækkende drosler fortærer. Dette gør vi på tre måder:
- I Figur 3.1 viser vi hvordan mængden af drosseldøgn ændrer sig hvis trækgæsterne fortærer en fast procentdel af de bær der er til rådighed. Det vil sige at reduktionen i bærmængden modsvares fuldstændig af en reduktion i fortæringen (20% færre bær medfører 20% mindre fortæring).
- I Figur 3.2 viser vi hvordan mængden af drosseldøgn ændrer sig hvis trækgæsterne fortærer en fast mængde af bær uanset hvor meget bærmængden initielt er reduceret.
- Da Figur 3.1 og 3.2 viser hver sin ekstreme situation, viser vi i Figur 3.3 gennemsnittet af Figur 3.1 og Figur 3.2. Vi mener at gennemsnitsscenariet af de tre præsenterede er det der kommer virkeligheden nærmest, idet det præsenterer en model der forudsiger at med faldende bærmængde til rådighed vil der i den periode hvor trækfuglene er i landet blive spist en større og større andel. Dog med den begrænsning at den totale mængde fortærede bær i modsætning til situationen i Figur 3.2 også falder med faldende bærmængde.
Figur 3.1
Trækdroslerne og solsortene fortærer en fast andel (0 – 1; vist vha.
kurverne i spring af 0,1) af de bær der er til rådighed i den periode hvor
trækdroslerne er i landet. x-aksen angiver reduktionen i bærmængden. På y-aksen vises
den mængde drosseldøgn der vil være til rådighed for standfuglene efter at
trækdroslerne har forladt landet.
Figur 3.2
Trækdroslerne og solsortene fortærer en fast mængde tjørnebær (0 –
1; vist vha. kurverne i spring af 0,1) i den tid trækdroslerne er i landet. x-aksen
angiver reduktionen i bærmængden. På y-aksen vises den til reduktionen svarende mængde
drosseldøgn der vil være til rådighed for standfuglene efter at trækdroslerne har
forladt landet.
Figur 3.3
Trækdroslerne og solsortene fortærer en forøget andel af bærrene når
bærmængden reduceres. Kurverne er fremkommet som gennemsnittet af Figur 3.1 og Figur
3.2. Ellers aflæses kurverne ligesom i Figur 1 og 2.
Hvis trækdroslerne og solsortene initialt fortærer hvad der svarer til 65% af de drosseldøgn der er til rådighed, så er der 21 mill. drosseldøgn tilbage til standfuglene. Hvis der samtidig er en 20 procents reduktion i den bærmængde der initialt er til rådighed vil vi ved aflæsning (Figur 3.3) se at mængden af drosseldøgn til rådighed for standfuglene falder fra 21,0 millioner drosseldøgn til 12, 9 millioner drosseldøgn, svarende til en nedgang på 38,6% (Figur 3.4). Ved henholdsvis en fast fortæring og en andelsmæssig fortæring ville de respektive nedgange være fra 21 til 9 mill. drosseldøgn ~ 57,1% og fra 21 til 16,8 mill. drosseldøgn ~ 20,0%. Når kurven går under nul betyder det ikke at droslerne begynder at spytte bærrene ud igen. Det betyder derimod at alle tjørnebærrene er fortæret og at trækdroslerne enten forlader landet eller begynder at fortære andre ressourcer f. eks. æbler eller andre bær.
Figur 3.4
Sammenhængen for gennemsnitsscenariet mellem den mængde bær der fortæres
af trækfugle og standfugle i den periode trækfuglene er i landet og den reduktion denne
fortæring afstedkommer i den bærmængde der er til rådighed for standfuglene efter at
trækfuglene har forladt landet. Dette er præsenteret ved tre forskellige effektniveauer
(5, 20, 40 % reduktion i bærmængden, f. eks. som følge af sprøjtemiddelafdrift).
3.3 Konklusioner for kommende forsøg
Under forudsætning af at de trækkende drosselfugle sammen med solsortene som udgangspunkt fortærer 65% af tjørnebærrene, medens trækfuglene er i landet, viser de tre scenarietyper at en 20% reduktion i bærmængden bevirker at der bliver mellem 20 og 57% færre bær tilbage til de bærspisende standfugle (hovedsagelig solsorte). Dette må antages at have en signifikant biologisk effekt for standfuglene. Det er dog klart at antallet af tilbageblevne bær på tjørnen er følsomt overfor hvor mange drosselfugle der trækker gennem Danmark. Derudover er det også af stor betydning hvor stor en del af de trækkende fugles føde der udgøres af tjørn. Dette afhænger igen af sæson og hvilke alternative fødekilder der er. Disse alternativer vil typisk være andre bær, f.eks. slåen, hyld, rose og kristtorn.
4. Sprøjteforsøg
| 4.1 | Introduktion |
| 4.2 | Metode |
| 4.2.1 | Forsøgsdesign |
| 4.2.2 | Sprøjteprocedure |
| 4.2.3 | Prøvetagning |
| 4.3 | Resultater |
| 4.4 | Diskussion |
| 4.5 | Konklusioner for kommende forsøg |
4.1 Introduktion
Der blev gennemført et lille pilotsprøjteforsøg med metsulfuron. Dette forsøg, havde til formål at fungere som range-finding for et forsøg til belysning af tjørn som indikator for biodiversitetspåvirkning ved drift af herbicid fra mark til hegn.
4.2 Metode
4.2.1 Forsøgsdesign
Før eksponering med metsulfuron blev der tilfældigt udvalgt 6 træer i hegn C. I hvert af disse træer blev 10 blomsterskud mærket. Den efterfølgende dag blev et træ sprøjtet for hver af følgende sprøjtedoser: 0, 1, 5, 10, 20, 40 % af den anbefalede markdosis for metsulfuron (4 g a.i. ha-1) i kornafgrøder. Sprøjtetidspunktet blev valgt således at et formodet "worst case scenario" opnåedes. Det vil sige at der blev sprøjtet på det tidspunkt hvor planten begynder at føre ressourcer fra bladene til de reproduktive organer, jævnfør sprøjtemidlets virkning i sublethalt påvirkede planter hvor resultatet er at frøene bliver mindre .
4.2.2 Sprøjteprocedure
Planterne blev sprøjtet med en azo-sprøjte, hvor sprøjtebommen blev holdt lodret, således at planterne sprøjtedes med den ønskede dosis i et "bælte" fra jordoverfladen og op til ca. 2 m. Der blev kun sprøjtet på den ene side af hegnet. Sprøjtningen blev gennemført den 27. juni. Da forsøget var ment som et pilotforsøg blev der ikke foretaget verifikation af den eksponering grenene var udsat for. Væskemængden der blev udsprøjtet svarede til 200 l ha-1. Denne mængde blev opnået ved at justere ganghastigheden i forhold til den væskemængde som azosprøjten leverer ved 2 atm arbejdstryk og med Hardi 4110-16 dyser monteret.
4.2.3 Prøvetagning
Forud for sprøjtningen blev de enkelte frugtskud, der skulle høstes 22 dage senere, afmærket og antallet af blomster, frugter og insektangrebne frugter registreret. Ved høst blev disse "frugtklaser" atter optalt. De 10 skud høstet for hver sprøjtedose blev udvalgt i ca. en meters højde over jorden på et træ.
4.3 Resultater
Der blev fundet en klar negativ effekt ved at eksponere engriflet hvidtjørn for metsulfuron i doser op til 40% af anbefalet markdosis (se Figur 4.1). Sammenhængen kan beskrives ved følgende ligning.
Andel tilbageblevne bær = 0,287 - 0,0067 x Dosis; r2 = 0,232 N=59 ,
hvor "Dosis" er % af den anbefalede markdosis og "Andel tilbageblevne bær" er den relative mængde af bær der var tilbage ved høst 22 dage efter sprøjtning. Ved sprøjtning på dette tidspunkt viser disse data at den største effekt af en given øgning af dosis opnås mellem 10 og 30 % af anbefalet markdosis. Ovenstående relation er blevet etableret på basis af optælling af både beskadigede og ubeskadigede bær. De beskadigede bær blev observeret som sorte prikker der indikerer herbivorangreb eller som unormale bær i øvrigt. I nogle tilfælde kunne vi se af vores optællinger at antallet af angrebne bær var forøget ved slutningen af forsøget. Da det i starten af forsøget var vanskeligt at erkende de angrebne bær antog vi at de der var tilstede ved høst også havde været angrebet ved start. Vi havde bare ikke været i stand til at detektere dem.
Figur 4.1.
Sprøjteforsøgets resultater præsenteret som andel tilbageblevne bær ved
forsøgets afslutning som funktion af dosis. De præsenterede afvigelser er standardfejlen
på middeltallet.
4.4. Diskussion
Dataanalysen af pilotforsøgets sprøjteforsøg viste at det er vigtigt at opgøre antallet af angrebne bær, så de kan udelades i nogle analyser (Fig. 4.1). Det var således nødvendigt at korrigere for de frugter der var angrebet på sprøjtetidspunktet da der for disse var et mindre nedfald end for de tilsvarende bær uden insekter.
Det er dog muligt vi ville have set at de angrebne bær også var faldet hvis vi var fortsat med at observere de angrebne bær i en længere periode, eventuelt kun ved de højeste doser. Hvordan og i hvilken grad de angrebne bær påvirkes af metsulfuron er dog uklart. Muligheden for at de angrebne bær også falder af, blot senere, bygger vi primært på en ikke-systematisk observation nogle uger efter afslutningen af forsøget hvor stort set alle bær var væk i de sprøjtede områder. Vi har tidligere iagttaget at angrebne bær i ikke-herbicidpåvirkede hegn kan blive hængende helt ind til næste års løvspring. Vi tror at dette skyldes at bærrene ikke er attraktive for fugle og måske også at nogle arter af insektlarver inducerer en ændring i planten som gør at bærrenes stilk bliver hængende unormalt længe. Dette må betyde at de angrebne bær ikke påvirkes af sprøjtemidlet på samme måde som de uangrebne. Måske udsender insekterne i de angrebne bær en forbindelse der modvirker herbicidets hæmning af allokering fra blade til reproduktive dele, f. eks. er det vist at tilførsel af de forgrenede aminosyrer L-valine, L-leucine og L-isoleucine kan ophæve virkningen af chlorsulfuron hos Almindelig pengeurt Thlaspi arvense . Grunden til at træet aborterer de uangrebne bær så hurtigt skyldes uden tvivl sprøjtemidlets hæmning af translokation af fotosynteseprodukter til frugtanlæggene.
Det er vores forventning at tilsvarende sprøjteforsøg udført i andre niveauer i træet vil være sammenlignelige med det udførte forsøg på grund af at hæmningen af transporten af fotosynteseprodukter er lokal. Det er vigtigt at kende såvel variationen i sprøjtemidlets afsætning, som dets halveringstid under de givne forhold (lys/skygge, top/bund, kort/langskud, etc.). Derudover vil der eventuelt skulle tages højde for en forskel i mængden af insektangrebne bær mellem niveauer. Sådan en forskel er ikke usandsynlig, idet f. eks. snudebillerne pga. vind vil have svært ved at bevæge sig op i tjørnens øverste dele. I og med at vi tager højde for disse forhold, mener vi fortsat at vi kan nøjes med at gennemføre sprøjtninger i et niveau af træerne. Det er vigtigt at konstatere at også kontrolplanter tabte en vis andel blomster/frugter. Ved forsøgets slutning var der således kun 30% af de optalte bær tilbage på kontrolplanterne. De øvrige er formodentlig tabt på grund af herbivore insekter eller som et resultat af plantens "prioritering". Dette er i modsætning til det store tab af knopper og bær der forekommer tidligere (planten aborterer f. eks. på grund af dårlig bestøvning eller insekter i blomster/frugtknopper (snudebiller som Anthonomus pedicularius, A. sorbi og Rhyncites aequatus) se iøvrigt afsnit 5.3).
4.5 Konklusioner for kommende forsøg
Samlet kan man sige at resultaterne af dette meget begrænsede sprøjteforsøg oplyser os om at:
| For ikke insektangrebne frugtanlæg kan vi forvente at finde udtalte effekter på bærsætningen ved doser ned til 10 procent af markdosis | |
| Mængden af insektangrebne frugter er af stor betydning for tolkningen af resultater |
Vi ved fortsat ikke om:
| Effekterne er mere eller mindre udtalt ved tidligere sprøjtning | |
| Effekterne er begrænset til det første år | |
| Der er effekter af lavere doser, i det mindste ikke kvantitativt |
5. Insekter i tjørnehegn
| 5.1 | Introduktion |
| 5.2 | Metode |
| 5.3 | Resultater og diskussion |
| 5.4 | Konklusioner for kommende forsøg |
5.1 Introduktion
Når man skal planlægge et forsøg vedrørende bærbærende buske og træer (her tjørn) som indikatorer for biodiversitetsforandringer ved sprøjtemiddelafdrift er insekterne interessante af to grunde. For det første er de en del af biodiversiteten og for det andet kan de planteædende arter påvirke det vi måler på (bærmængden). Courtney og Manzur (1985) fandt at nedfaldet af bær i hegn domineret af tjørn var relateret til insektangreb, frugtmængde, nærmeste nabo (konkurrent) og bredde af hegnet. De insekter, der findes i hegnet, kan være der for at søge forskellige ressourcer. De kan udnytte urtevegetationen, søge byttedyr, nektar eller pollen. Endelig kan de udnytte hegnsplanten som føde. Insekternes effekt på plantens vækst og reproduktion er det vigtigste for planlægningen af forsøget. Vi har derfor undersøgt hvilke arter der er registreret som hjemmehørende i tjørnehegn og hvilke der forekommer i de hegn som vi har arbejdet i i pilotforsøget.
5.2 Metode
Insekterne blev indsamlet ved både såkaldte bankeprøver, altså en skærm bestående af en ramme på ca. 1 � 1 m hvori hvidt stof udspændes, holdes under en gren, som bankes og rystes grundigt; ved individuel indsamling på enkelt grene og ved ketcherslag. Indsamlingerne blev lavet flere gange gennem sæsonen for at repræsentere forskellige fænologiske perioder. Indsamlingsdatoerne var: 30. maj, 14. juni, og den 15. august. I laboratoriet blev dyrene sorteret og konserveret i sprit. En række blomster og knopper blev dissekeret for herbivore larver. De indsamlede arter vil blive sammenlignet med de registrerede arter på tjørn.
5.3 Resultater og diskussion
Der er registreret en lang række insektarter i tjørnehegn. Vores indsamlinger viste at arterne i de udvalgte hegn var at finde blandt de registrerede. Der registreredes meget naturligt en fænologisk forskydning mellem arter der henholdsvis udnytter knopper, blomster og bær (Tabel 5.1).
Indsamlingerne af blade og reproduktive enheder viste at der er et meget stort tab af reproduktive enheder gennem udviklingen (Figur 5.1). Tabet kan være plantens strategi for at optimere de ressourcer der er til rådighed, men det er også en respons på herbivori. Som det ses ud af figuren forekom nedfaldet i 2001 først og fremmest i knop- og blomsterstadiet.
Tabel 5.1
Oversigt over de arter der er almindeligt hjemmehørende i tjørnehegn i
Danmark samt en opgørelse af de arter der er fundet i forsøgshegnene hhv. den 30. maj,
14. juni og 15. august.
Figur 5.1
Det gennemsnitlige antal reproduktive enheder per sidegren afhængigt af
udviklingsstadie. Data er opsplittet efter højde i hegnet (Bund, Mellem, og Top). De
præsenterede afvigelser er standardfejlen på middeltallet.
Fire billearter med tilknytning til hvidtjørn blev identificeret. Anthonomus pedicularius og A. sorbi er begge snudebiller som angriber hvidtjørnens blomster og frugtknopper i maj-juni. A. pedicularius larver er kendt for at være årsag til fænomenet "springende knopper", dvs. knopper som bevæger sig i små spring, hvis man tager dem op i hånden eller lægger dem på et bord. Springene forårsages af snudebillelarverne inde i knoppen ved forstyrrelser. A. pedicularius kan undertiden angribe æble og pære. A. sorbi træffes også på slåen og hæg. Det er uvist i hvilket omfang larverne bevirker at knopperne på et tidspunkt aborteres og falder til jorden; men vores data viser at en meget stor andel af knopperne og blomsterne tabes (Figur 5.1). Hvad angår blomsterne skal det bemærkes at indsamlingstidspunktet betyder at vi reelt ikke kan sige om tabet finder sted i blomsterfasen eller i fasen mellem blomster og bær ~ frugtanlægsfasen. Larverne forpupper sig i jorden og de voksne biller klækker i løbet af sommeren og opsøger overvintringssteder i tjørnen.
En anden snudebille, Rhynchites aequatus, er ud over tjørn og slåen også fundet på blomme. Den angriber den grønne frugt og frugten falder til jorden hvor larven overvintrer. Imago klækker næste forår. Det ser ikke umiddelbart ud til at denne art har haft stor effekt på mængden af bær. Antallet af modne bær er af samme størrelse som antallet af grønne bær.
Tjørnebladbillen, Lochmaea crataegi, yngler i følge vores oplysninger udelukkende på hvidtjørn. De voksne biller træffes på hvidtjørn fra april til juni. Larverne gnaver af bladene. Vi fandt ingen larver af denne art.
De tre snudebiller nævnt ovenfor var klart de hyppigste insekter i alle hegnene.
5.4 Konklusioner for kommende forsøg
Pilotforsøgets indsamlinger viser at der er en række insekter der angriber blomsterknopper og bær som kan forårsage at disse aborteres. Arterne angriber på forskellige tidspunkter og derfor skal indsamlingerne foretages med relativt korte intervaller for at få et realistisk skøn over insektmængden. Da nedfaldet af knopper, blomster og bær skyldes insekter, er det vigtigt at bestemme hvor stor en andel af de reproduktive enheder der er angrebet. Dette gælder både de der er på træet og de nedfaldne. Der skal altså indsamles knopper, blomster og bær gennem sæsonen. Disse skal efterfølgende dissekeres for at se om de er eller har været angrebet af insekter. Derudover opsættes bakker til at opsamle nedfaldne knopper, blomster og bær.
6. Begrundet forsøgsdesign
6.1 Introduktion
Det er formålet med dette projekt at etablere kvantitative data for herbicidet metsulfurons effekt på vegetativ vækst, blomsterproduktion og bærproduktion hos engriflet hvidtjørn. Afdrift af herbicider fra marker til omkringliggende hegn kan påvirke mængden af disse plantedele og hermed også fødemængden for den tilknyttede fauna såsom herbivore insekter og bærædende fugle. Ydermere kan en eventuel påvirkning af insekterne bevirke at de insektædende fugles antal reduceres. Vi har i de foregående kapitler gennemgået de data der er indsamlet i pilotforsøget for at understøtte planlægningen af et forsøg der skal dokumentere om der er effekt af herbicidet metsulfuron på tjørn i de udvalgte hegn. I kapitel 2 blev der gennemført en statistisk styrkeberegning for at beregne de prøvestørrelser der er nødvendige for med en styrke på 80% at detektere effekter der ligger over 20%. Kapitel 4 gennemgik resultatet af et lille sprøjteforsøg og i kapitel 5 blev resultatet af insektindsamlinger i fire tjørnehegn gennemgået. I nærværende kapitel vil vi på basis af de indsamlede data, litteraturen og økonomiske scenarieberegninger argumentere for et givent forsøgsdesign. Det design, som vi vil foreslå her i kapitlet, har som overordnet mål at sammenholde observerede effektniveauer med afsætning og relativ biomasse i regioner/højder af træerne for at beregne om det er sandsynligt at der vil forekomme effekter i hegn under normal sprøjtepraksis.
6.2 Prøveantal
De statistiske styrkeberegninger viste at der er store forskelle mellem forskellige endpoints (blade, knopper, blomster, grønne bær, modne bær) i hvor mange sidegrene per træ og hvor mange træer per hegn det er nødvendigt at indsamle for at opnå en statistisk styrke på 80%. Det var tydeligt for alle de valgte endpoints at det nødvendige prøveantal er lavest ved høst af få sidegrene per træ men med lidt flere træer per hegn. Det var mere variabelt hvorledes et øget antal hegn påvirkede antallet af prøver.
Når man skal vælge mellem de forskellige alternativer er der andre parametre end den statistiske styrke der skal inddrages. En vigtig diskussion er afvejningen af undersøgelsens generalitet, det vil sige antallet af hegn der laves undersøgelser i, i forhold til den nødvendige ressource der skal til for at gennemføre dataindsamlingen. Hvis der kun inddrages et enkelt hegn er det muligt at vi kan opnå den ønskede statistiske styrke, men i den efterfølgende diskussion af data vil konklusionerne kun være repræsentative for dette ene hegn. Hvis flere hegn, gerne af meget forskellig karakter, inddrages vil de biologiske konklusioner være af mere generel karakter. Det er derfor vores mål at inddrage flere hegn hvis dette ikke medfører for store omkostninger praktisk og økonomisk.
Nedenfor præsenteres de mulige scenarier for indsamlinger der opfylder kriteriet med en statistisk styrke på 80% (ud fra vores valg). Senere i dette kapitel vil vi beregne tidsforbruget ved at gennemføre det foreslåede forsøg i henholdsvis 1, 2, 4, 8 eller 16 hegn. Det betyder at vi har udvalgt de scenarier der har det laveste antal prøver for hver variabel (Tabel 6.1) og dernæst udvalgt den måleparameter hvor der skulle bruges flest træer således at begge måleparametre var dækket ind.
I pilotforsøget blev der indsamlet de sidegrene der som den mindste enhed indeholder blade eller knopper, blomster og bær. Disse indsamlinger giver imidlertid ikke data for den samlede bærbiomasse (udbytte) i hegnet hvorfor data skal suppleres med areal-relaterede data. Det vil sige at der skal tælles skud inden for et givent areal fordelt på forskellige positioner i træet . Disse tællinger vil gøre det muligt at omsætte effekter til hele hegnet når man kender afsætningens fordeling i et hegn som funktion af højden. Der foretages kun en enkelt måling på et tidspunkt hvor det er klart om sideskuddene er fertile. Forandringer i de enkelte sideskud måles jo andetsteds. Målingerne bliver yderligere understøttet af en massebestemmelse af 2 træer per forsøgshegn. Oplysninger om sprøjtemidlers afsætningsmønster i hegn findes i litteraturen .
Som tidligere nævnt vides det ikke om der er effekter året efter at planterne har været udsat for sprøjtning, derfor skal der høstes i sprøjtede træer både det år der sprøjtes og året efter.
6.3 Sprøjteprocedure
Forsøgene gennemføres som sprøjteforsøg i hegn, hvor engriflet hvidtjørn er dominerende og hvor der ikke sprøjtes i det yderste sprøjtespor på de tilstødende marker (12-18 m). I forsøgene sprøjtes sektioner af 3 til 4 træer. Der sprøjtes med fem doser henholdsvis 0%, 5%, 10%, 20% og 40% af anbefalet markdosis for kornmarker. Disse doser er blevet valgt dels fordi pilotforsøget viste at der var en målbar effekt mellem 10 og 30% af anbefalet markdosis og fordi højere doser ville være væsentlig højere end de doser der vil kunne observeres under normale driftforhold. Vi holder fast i den højeste dosis på 40% selvom pilotforsøget viste en 100% effekt allerede ved 20% for de bær der ikke var angrebet af insekter. Det gør vi fordi pilotforsøget kun er gennemført for et enkelt udviklingsstadie på et enkelt sprøjtetidspunkt. Der sprøjtes på to tidspunkter, nemlig forår (maj) og tidlig sommer (juni).
Kontrolplanterne sprøjtes ikke da vi ønsker at undersøge den samlede effekt af sprøjtning af tjørnehegn. Som en yderligere fordel medfører ovennævnte synsvinkel at den samme kontrol kan bruges ved begge sprøjtninger. Dette reducerer prøveantallet i det enkelte hegn hvilket gør det lettere at finde egnede hegn.
Idet den valgte sprøjtemetode er kunstig i forhold til en naturlig driftsituation og fordi det ikke umiddelbart er muligt at omsætte udsprøjtet mængde til mængde afsat sprøjtemiddel, er det nødvendigt at få et mål for sprøjtemiddelafsætningen. Disse afsætningsmålinger skal både bruges til at relatere effektmålingerne til en "normal" drifthændelse og til at få en direkte dosis-effekt sammenhæng. Ved sprøjtningen bestemmes den væskemængde der afsættes i hegnet ved hjælp af glycin afsætning. Idet træets/hegnets form er af betydning for afsætningen (for eksempel kan de yderste grene skygge for blade siddende længere inde i træet) skal afsætningen måles på en sidegren lige ved siden af de sidegrene der skal bruges til indsamling.
6.4 Insektindsamlinger
Pilotforsøgets indsamlinger viser at der er en række insekter der angriber blomsterknopper og bær med den mulige konsekvens at disse aborteres. Da pilotforsøget yderligere viste at ukrudtsmidlet metsulfuron også forårsagede nedfald er det vigtigt at isolere herbicideffekten fra andre årsager til nedfald. Udover insektprovokeret nedfald kan det ikke udelukkes at nogle af bærrene aborteres fordi bladædende insekter eller andre energi- og næringsbegrænsende faktorer "udsulter" de bærbærende sideskud (Manzur og Courtney 1984). Derfor er det vigtigt at fastslå hvor mange af de knopper, blomster og bær der aborteres der er angrebet af insekter. Vi vil derfor indsamle både knopper, blomster og bær på 8 tidspunkter gennem sæsonen og dissekere dem for insekter, ligeledes vil vi opsamle nedfaldne knopper, blomster og bær således at en eventuel sprøjtemiddeleffekt kan adskilles fra andre årsager til nedfald af frugter.
6.5 Pilotforsøg af insekticideffekt på frugtsætning
Ifald insekterne betyder meget for mængden af tjørnefrugter og blomster der falder ned før modenhed kan man forestille sig at drift af et insekticid ind i hegnet betyder at færre generative enheder falder ned og at der bliver flere frugter. Vi ønsker som nævnt i Kapitel 1 at bruge tjørn som indikator for biodiversiteten i hegn. Selvom bærmængden faktisk stiger er det ikke ensbetydende med at biodiversiteten også stiger idet insektmidlet har en direkte effekt på insekterne. Derfor vil vi lave et pilotforsøg for at etablere effekten af en insekticidsprøjtning på bærproduktionen af tjørn. Pyrethroider er de mest benyttede insektmidler i Danmark (Miljøstyrelsen 2001), og derfor udvælger vi en af de mest benyttede af disse som repræsentant for drift af insektmidler ind i hegn.
Fem enkelttræer sprøjtes med et insekticid i starten af juni. Doserne vælges i spændet mellem 0 og 40% af anbefalet markdosis. Inden sprøjtning afmærkes 10 sidegrene og antallet af blomster opgøres. Endelig anslås det, hvor mange af blomsterne der er angrebet af insekter. Efterfølgende optælles sideskuddene en gang om ugen i 6 uger.
6.6 Økonomiske betragtninger
6.6.1 Kriterier
Som nævnt tidligere ønsker vi at have den størst mulige sikkerhed i de konklusioner vi når frem til i et efterfølgende forsøg. Derfor sigter vi på at indsamle i så mange hegn som praktisk og økonomisk muligt. Det er dog også vigtigt at konstatere at mængde af træer som skal sprøjtes også bestemmes af den normale størrelse af tjørnehegn. Hvis der for eksempel skal behandles 30 træer per dose vil det betyde at der skal bruges 300 træer (30 træer x 5 doser x 2 sprøjtetidspunkter). Med bufferzoner mellem de sprøjtede træer betyder dette at alle hegn skulle mindst være 1 km lange. Vi har lavet en beregning af de nødvendige ressourcer til at kunne måle en 20% effekt ved et signifikansniveau på 0,05 og en statistisk styrke på 80% for begge måleparametre (vægt af sideskud og antal af enheder per sideskud). Beregningerne af tidsforbruget for høst af blade er kun lavet på antallet af blade i kortskud idet langskud overvejende forekommer i klippede hegn.
6.6.2 Beregninger
Beregningen af tidsforbruget for de forskellige scenarier er lavet udfra den tid der går med: indsamling af sidegrene (herunder også prøve behandling), sprøjtning, kørsel, afmærkning, insektprøver, massebestemmelse af træer og sideskudsoptælling på træer per areal. For beregningen er der gjort visse antagelser baseret på erfaringer fra pilotforsøgene. Disse er gennemgået nedenfor.
Prøvebehandling
Prøvebehandling af høstede plantedele inkluderer mærkning, tælling, tørring, vejning og dataindtastning som anslået beløber sig til 6 min. per sideskud.
Sprøjtning
I forbindelse med sprøjtning er der en del forberedelser der samlet set beløber sig til 1656 min.. Heri er indregnet en kalibrering af sprøjteudstyr, sprøjteopløsninger, opsætning, opbevaring og måling af afsætning såvel som oprydning. Derudover er der indregnet 8 min. til at sprøjte det enkelte træ og opfyldning af sprøjte mm., 15 min. ekstra per dosis fordi der skal skiftes sprøjtebeholder mm. og endelig er der beregnet en time ekstra per hegn til for eksempel af- og påføring af sprøjteudstyr. I Tabel 6.1 er der kun beregnet tid til sprøjtning for et enkelt udviklingsstadie (det der kræver flest træer) idet træerne genbruges ved alle øvrige indsamlinger.
Kørsel
Til kørsel er der beregnet 50, 70, 100, 200 og 220 min. per person ved hhv 1, 2, 4, 8, og 16 hegn. Disse tal er skønnet udfra at vi regner med at kunne finde hegn tæt ved Silkeborg og at disse ikke ligger for langt fra hinanden (men nok til at de er forskellige). I beregningerne indgår det at tre personer deltager per gang. Det er, som tidligere nævnt, planlagt at der skal laves en massebestemmelse af nogle hele træer fra forsøgshegnene (to per hegn). Tidsforbruget hertil er skønnet udfra tidligere erfaringer med opskæring og tælling af egetræer.
Opskæring
Vi skønner at det vil tager i alt 250 min. per træ (fældning 30 min., opdeling 120 min., transport 10 min., vejning 20 min., tælling 60 min. og indtastning 10 min..). Opskæringen finder sted mellem tidspunktet for de to forsøgsprøjtninger.
Udbytte-måling
’Udbytte-målingen’ det vil sige den arealrelaterede optælling af sideknopper, er skønnet til at koste en dags arbejde per hegn. Dette er baseret på køretid, at der skal to afsted for at kunne arbejde i flere højder af træet og at der skal tælles i tyve træer og en ramme per højde i træet.
Insektindsamlinger
Insektindsamlingerne laves på otte tidspunkter gennem sæsonen med fem træer per hegn. Fra hvert træ indsamles 30 enheder af det dominerede udviklingsstadie (knopper, blomster, bær). I alt vil dette give 1200 prøver. Herudover vil vi om muligt opsamle 320 nedfaldne enheder. Det vil sige at der i alt bliver 1520 prøver. Vi skønner at det i gennemsnit tager 1 minut at konstatere om der er insekter i og hvilken type. Det giver derfor 25 timer + 8 timer til indsamling + 4 timers ekstra køretid for tre personer.
Insekticidbehandling
Pilotforsøget til at undersøge om en insekticidbehandling har effekt på bærproduktionen gennemføres i år 2. Vi planlægger at behandle tjørnetræer med 5 doser i 3 replikater (træer). Fra hvert træ høstes 20 sidegrene. Alt i alt betyder det at der per indsamling vil være 300 prøver til registrering og dissektion. Der skal indsamles sideskud 6 gange gennem sæsonen, altså 1800 prøver. Vi skønner at dette vil tage 40 timer. Sprøjtningen tager 15 timer inklusive forberedelse, og kørsel tager samlet ca. 7 timer (8 ture af 50 min.).
6.6.3 Resultater
Beregningerne af tidsforbruget viste at det var mest omkostningskrævende kun at indsamle prøver fra et enkelt hegn, nemlig 9329 timer(Tabel 6.1). Tidsforbruget faldt med stigende antal hegn op til fire hegn (3034 timer). Ved at indsamle i 8 hegn skulle der bruges 3209 timer og ved indsamlinger i 16 hegn steg tidsforbruget yderligere til 3587. Dermed er indsamlinger i 4 hegn den optimale løsning for forbruget af tid. Herved kræves der mindst 27 træer per dose per hegn. Det vil sige at det er nødvendigt at finde 4 hegn domineret af tjørn der har en længde på cirka 1 km. Det har ikke været muligt. Vi vælger derfor i stedet at øge antallet af hegn til 8 for de to endpoints "grønne bær" og "modne bær". Herved reduceres antallet af træer per dose til 13. At vi ikke også øger antallet af hegn til 8 for blade, knopper og blomster beror på flere forhold:
| at vi anser bærrene som det vigtigste endpoint | |
| at vi ønsker at inddrage en større geografisk variabilitet. De hegn vi har kunnet finde ligger på Sjælland, derfor bliver der større udgift til køretid end medregnet i Tabel 6.1 og også øgede udgifter til transport. For at holde udgiften nede nøjes vi derfor med bær-prøverne på Sjælland. |
For hegnene på Sjælland har vi regnet med en køretid på 4 timer hver vej for hver person der deltager i turen. For at spare køretid og bropenge inkluderes der overnatninger som gør at der kan indsamles fra alle hegn på en tur. Til gengæld skal der tillægges hoteludgifter. Der skal gennemføres 10 prøvetagninger på Sjælland, hvilket vil give et ekstra forbrug på 240 timer per år i forhold til beregningerne i Tabel 6.1.
Tabel 6.1
Beregninger af tidsforbruget (timer) ved at lave undersøgelser i 1, 2 , 4, 8
eller 16 hegn. Antallet af træer og sideskud er bestemt ud fra statistiske
styrkeberegninger. Søjlen benævnt "Tid" er den tid der skal bruges på
indsamlede prøver i år 1.’Chop’ er den tid der går med at skære 2 træer op
per hegn og tælle antal sideskud i forskellige dele af træet. ’Insekter’ er
den tid der går med at indsamle knopper, blomster og bær og dissekere dem for indhold af
insekter. ’Insekticid’ er tidsforbruget til pilotforsøg vedr. insekticiders
effekt på bærmængden. ’Udbytte’ er tiden brugt til at lave et arealrelateret
mål for bærmængden. ’Total’ er den beregnede tid der skal bruges hvis der
bliver indsamlet i henholdsvis 1, 2 , 4, 8 eller 16 hegn.
7. Referencer
Khatib, K., Parker, R. and Fuerst, E.P., 1992. Sweet Cherry (Prunus-Avium) Response to Simulated Drift from Selected Herbicides. Weed Technology, 6(4): 975-979.
Asbirk, S., Berg, L., Hardeng, G., Koskimies, P. and Petersen, A., 1997. Population sizes and trends of birds in the Nordic countries 1978 - 1994. TemaNord, 614: 1-88.
Bestman, H.D., Devine, M.D. and Vandenborn, W.H., 1990. Herbicide Chlorsulfuron Decreases Assimilate Transport Out of Treated Leaves of Field Pennycress (Thlaspi arvense L.) Seedlings. Plant Physiol., 93: 1441-1448.
Bhatti, M.A., Alkhatib, K., Felsot, A.S., Parker, R. and Kadir, S., 1995. Effects of Simulated Chlorsulfuron Drift on Fruit Yield and Quality of Sweet Cherries (Prunus-Avium L). Environmental Toxicology and Chemistry, 14(3): 537-544.
Boutin, C. and Jobin, B., 1998. Intensity of agricultural practices and effects on adjacent habitats. Ecological Applications, 8(2): 544-557.
Boutin, C., Lee, H.B., Peart, E.T., Batchelor, P.S. and Maguire, R.J., 2000. Effects of the sulfonylurea herbicide metsulfuron methyl on growth and reproduction of five wetland and terrestrial plant species. Environmental Toxicology and Chemistry, 19(10): 2532-2541.
Boutin, C. and Rogers, C.A., 2000. Pattern of sensitivity of plant species to various herbicides - An analysis with two databases. Ecotoxicology, 9(4): 255-271.
Christensen, H. et al., 1998. Effekter af reduceret pesticidforbrug på flora og fauna i agerlandet. Ikke-publiceret statusrapport for 1997.
Collinge, W.E., 1941. The food of the blackbird Turdus merula in succesive years. Ibis, 1941: 610-613.
Courtney, S.P. and Manzur, M.I., 1985. Fruiting and finess in Crataegus monogynae: the effects of frugivores and seed predators. OIKOS, 44: 398-406.
Davis, B.N.K., Brown, M.J., Frost, A.J., Yates, T.J. and Plant, R.A., 1994. The Effects of Hedges on Spray Deposition and on the Biological Impact of Pesticide Spray Drift. Ecotoxicology and Environmental Safety, 27(3): 281-293.
de Snoo, G.R., 1999. Unsprayed field margins: effects on environment, biodiversity and agricultural practice. Landscape and Urban Planning, 46(1-3): 151-160.
de Snoo, G.R. and de Wit, P.J., 1998. Buffer zones for reducing pesticide drift to ditches and risks to aquatic organisms. Ecotoxicology and Environmental Safety, 41(1): 112-118.
de Snoo, G.R. and van der Poll, R.J., 1999. Effect of herbicide drift on adjacent boundary vegetation. Agriculture Ecosystems & Environment, 73(1): 1-6.
Eriksson, O. and Ehrlen, J., 1991. Phenological variation in fruit characteristics in vertebrate dispersed plants. Oecologia, 86: 463-470.
Fletcher, J.S., Pfleeger, T.G., Ratsch, H.C. and Hayes, R., 1996. Potential impact of low levels of chlorsulfuron and other herbicides on growth and yield of nontarget plants. Environmental Toxicology and Chemistry, 15(7): 1189-1196.
Freemark, K. and Boutin, C., 1995. Impacts of Agricultural Herbicide Use on Terrestrial Wildlife in Temperate Landscapes - a Review with Special Reference to North-America. Agriculture Ecosystems & Environment, 52(2-3): 67-91.
Fuentes, M., 1994. Diets of Fruit-Eating Birds - What Are the Causes of Interspecific Differences. Oecologia, 97(1): 134-142.
Guitian, J. and Fuentes, M., 1992. Reproductive-Biology of Crataegus-Monogyna in Northwestern Spain. Acta Oecologica-International Journal of Ecology, 13(1): 3-11.
Hald, A.B., Reddersen, J. and Elbek-Pedersen, H., 1994. Sprøjtefri randzoner i sædskiftemarker - Plante- og insektliv samt udbytte. Landsforsøg 1987-92. Bekæmpelsesmiddelforskning fra Miljøstyrelsen, nr 6. Miljøstyrelsen, København.
Hald, A.B., Reddersen, J., 1990. Fugleføde i kornmarker - insekter og vilde planter. Undersøgelser på konventionelle og økologiske landbrug 1987-88, Miljøstyrelsen, Copenhagen.
Hartley, P.H.T., 1954. Wild fruits in the diet of British trhrushes. A study in the ecology of closely allied species. British Birds, 47(4): 97-107.
Herrera, C.M., 1987. Vertebrate dispersed plants of the Iberian peninsula: A study of fruit characteristics. Ecological Monographs, 57(4): 305-331.
Herrera, C.M., 1998. Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits: A 12-year study. Ecological Monographs, 68(4): 511-538.
Kendeigh, S.C., 1970. Energy requirements for existence in relation to size of bird. Condor, 72: 60-65.
Kennedy, C.E.J. and Southwood, T.R.E., 1984. The number of species of insects associated with British trees: A reanalysis. Journal of Animal Ecology, 53: 455-478.
Kleijn, D. and Snoeijing, G.I.J., 1997. Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology, 34(6): 1413-1425.
Manzur, M.I. and Courtney, S.P., 1984. Influence of insect damage in fruits of hawthorn on bird foraging and seed dispersal. OIKOS, 43: 265-270.
Marrs, R.H. and Frost, A.J., 1997. A microcosm approach to the detection of the effects of herbicide spray drift in plant communities. Journal of Environmental Management, 50(4): 369-388.
Maudsley, M.J., 2000. A review of the ecology and conservation of hedgerow invertebrates in Britain. Journal of Environmental Management, 60(1): 65-76.
Miljøstyrelsen, 2001. Bekæmpelsesmiddelstatistik 2000 - Salg 1998, 1999 og 2000: Behandlingshyppighed 1999. Orientering fra Miljøstyrelsen, 10.
Reddersen, J., Elmholt, S. and Holm, S., 1998. Indirect Effects of Fungicide and Herbicides on Arthropods. 44, Danish Environmental Protection Agency, Copenhagen.
Skuterud R, Bjugstad N, Tyldum A and Torresen KS (1998) Effect of Herbicides Applied At Different Times of the Day. Crop Protection 17: 41-46.
Snedecor, G. W. and Cochran, W. G., 1989. Statistical Methods. Iowa State University Press. Ames Iowa.
Snow, B. and Snow, D., 1988. Birds and berries - a study of an ecological interaction. T & A D Poyser, Calton.
Sorensen, A.E., 1981. Interactions between birds and fruit in a temperate woodland. Oecologia, 50: 242-249.
Sorensen, A.E., 1984. Nutrition, energy and passage time: Experiments with fruit preference in European Blackbirds (Turdus merula). Journal of Animal ecology, 53: 545-557.
Sparks, T. et al., 1996. The effect of three hedge management treatments on the wildlife of a Cambridgeshire hedgerow. Aspects of applied Biology, 44: 277-284.
Sparks, T.H. and Martin, T., 1999. Yields of hawthorn Crataegus monogyna berries under different hedgerow management. Agriculture Ecosystems & Environment, 72(2): 107-110.
Sparks, T.H. and Robinson, K.A., 1999. Hawthorn berry availability in autumn. Aspects of Applied Ecology, 54: 241-244.
Vandenborn, W.H., Bestman, H.D. and Devine, M.D., 1988. The inhibition of assimilate translocation by chlorsulfuron as a component of its mechanism of action. Proc. EWRS symp.: 69-74.
Verbela, G and Molenberghs, G., 1997. Linear mixed models in practice. Springer Verlag, New York.
Bilag A
Beskrivelse af jordbunden i de enkelte hegn
Beskrivelse af jordbunden i de enkelte hegn
For begge lokaliteter er der tale om at udgangsmaterialet er en moræneaflejring der overvejende er sandet/leret og ikke indeholder kalk. Lerindholdet kan være op mod 30% men er generelt væsentligt lavere i de øverste 20 cm. Som i alle moræneaflejringer er der tale om stor geologisk variation i sammensætningen af udgangsmaterialet. Der er imidlertid intet der tyder på at jordbundens udgangsmateriale på de to lokaliteter adskiller sig fra hinanden. Profiler af denne type har generelt en høj vandforsyningsgrad og en rigelig forsyning med næringsstoffer. I forhold til den omgivende vegetation på åbent land vil planter der vokser nær eller i et læhegn få en forsyning af kalk, kvælstof, fosfor og andre næringsstoffer der er flere gange større.
Jordbunden på begge lokaliteter er kun beskrevet visuelt og der er ikke hjemtaget prøver. I alt er der gravet 10 profiler til ca. 60-70 cm�s dybde. Generelt for alle profiler gælder, at der er tale om sandede morænejorde med et lerindhold på mellem 10 og 30%. Profilerne er beskrevet i meget tør tilstand.
De øverste 15 –20 cm er alle beskrevet som en mørkfarvet A horisont som er tæt gennemvævet med græsrødder. Dette øverste lag har formentlig været pløjet i en periode. For en del af de undersøgte profiler er der således en meget skarp grænse mellem A-horisonten og den underliggende horisont. Generelt for alle profiler gælder, at A-horisonten er mere sandet end de underliggende horisonter. På grund af rodaktiviteten hos græsserne og regnormes aktivitet er horisonten endvidere løs. Indholdet af sten er lille. Tykkelsen af den øverste horisont varierer noget afhængig af hvor i terrænet profilet er gravet. Typisk er A-horisonten tykkest i terrænlavninger som følge af pløjning som flytter materiale fra toppe mod lavninger samt på grund af at lavningerne altid vil være mere produktive som følge af lateral vand-/og stofbevægelse.
Under de mørkfarvede A-horisonter følger en forbrunet og mere humusfattig B-horisont. For flere af de undersøgte profiler er der tale om blandingshorisonter Ap/B. Forbruningen skyldes forvitring af jernmineraler, der oxideres og derfor er rød/brunfarvede. B-horisonten er alle steder meget kompakt og nærmest cementeret. Der er kun ganske få store rødder fra tjørnen og ingen synlige finrødder. Lerindholdet i morænejordene omkring Silkeborg ligger generelt på mellem 10 – 20% og kan i overgangen mellem A og B horisonten være større på grund af lernedslemning fra oven. I våde vintermåneder vil der formentlig kunne optræde vandstuvning i perioder.
Tabel A.1
Beskrivelse af jordbundsprofiler for de enkelte forsøgshegn. De billeder der er
refereret til findes i figur A.1.
Lokalitet |
Horisont (dybde) |
Kortfattet horisontbeskrivelse |
Buskelundvej |
|
|
Hegn C |
Ap (0 – 20 cm): Mørk rødbrun (5 YR 3/2) |
løs sandet med få små sten; humusrig og med mange fine græssrøder; regnormegange talrige. Overgang mod B er ikke jævn. |
B (20 – cm): Gulbrun (10 YR 5/4) |
kompakt humusfattigt lerblandet sand; ingen synlige røder |
|
Hegn B |
Ap1 ( 0 – 16 cm) Meget mørk gråbrun (10 YR 3/2) |
som profil 1; skarp overgang |
Ap2/B (17 – 50 cm) Mørk brun (10YR 4/3) |
mere kompakt end ovenliggende horisont med få større sten |
|
B/C (50 - ) Mørk gulbrun (10YR 4/4) |
gulligt stenet finsand/ler |
|
Sinding Hedevej |
|
|
Hegn D |
Ap (0-15 cm) Mørk brun (7.5 YR 3/2) |
løs sandet med få små sten; specielt i toppen humusrig og aftagende nedefter.Mange fine græssrøder. Enkelte klumper jordbrugskalk. Overgang mod B meget urolig |
Bt (25 - ) Gul brun (10YR 5/4) |
meget kompakt ler/sand |
|
Hegn E |
Ap (0-20 cm) Mørk brun (7.5 YR 3/2) |
løs sandet med få små sten; humusrig. Mange fine græssrøder. Enkelte klumper jordbrugskalk. Overgang mod B meget skarp |
A/B (20 - ) Mørk brun (10YR 4/3) |
lerblandet sand, meget kompakt |
Figur A.1
Billeder af udvalgte jordbundsprofiler fra henholdsvis hegn B, C, D og E