|
Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetation 2 Vegetation og leddyr i hegn på økologiske og konventionelle bedrifter2.1 Materiale og metoder Dette kapitel beskriver læhegnsundersøgelserne på sandjord i år 2001 og på lerjord i år 2002. 2.1 Materiale og metoderHerbiciders utilsigtede påvirkning af naturindholdet i småbiotoper belyses i en komparativ undersøgelse af læhegnsvegetation og tilhørende leddyr på økologiske hhv. konventionelle bedrifter. Med henblik på at sikre maksimal sammenligning har en række kriterier været gældende for udvælgelsen af læhegn. Kriterierne for de undersøgte læhegn på sandjord var, at hegnet skulle
De økologiske hegn opfyldte yderligere flg. kriterium
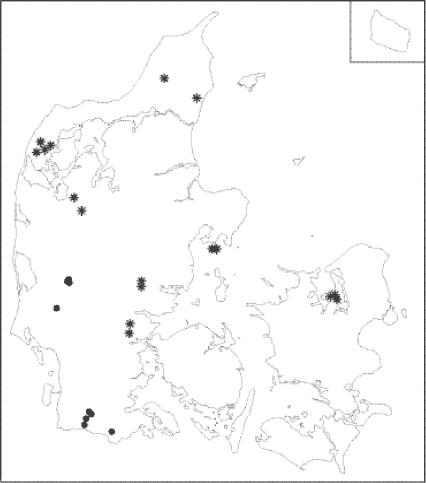
Seksogtyve (2x13) hegn blev undersøgt på konventionelle og økologiske bedrifter i 2001 syd og vest for israndslinien (dvs. JB 1-3(4)). De økologiske hegn blev udvalgt først og derefter blev tilsvarende konventionelle hegn i samme postdistrikt udvalgt. En detaljeret beskrivelse af de undersøgte hegn findes i Bilag A. I år 2002 blev der på baggrund af resultaterne fra 2001 udvalgt hegn efter de samme kriterier, blot med den forskel at hegnene skulle findes nord og øst for israndslinien og jordbundene skulle have et vist lerindhold, baseret på telefoninterview med ejeren (dvs. JB 3-7). Det blev dog i 2002 nødvendigt at udvide alderskriteriet (6-22 år, langt de fleste dog 10-15 år) i enkelte tilfælde for at opnå de 2x15 hegn der var målsat på baggrund af poweranalyser, da det var umuligt at opdrive et tilstrækkeligt antal hegn, der opfyldte samtlige kriterier. Eksempelvis blev der i enkelte tilfælde valgt konventionelle hegn i nabo-postdistrikter. Disse justeringer har været nødvendige for at opretholde minimum 10 års økologisk mark i omdrift på begge sider af hegnet som det væsentligste kriterium i udvælgelsen. Undersøgelsen omfatter dermed stort set samtlige eksisterende danske hegn, der opfylder de opstillede kriterier. Der er i alt i 2001 og 2002 inkluderet 28 hegn på 15 økologiske bedrifter og 28 hegn på 19 konventionelle bedrifter. Det typiske hegn var 14 år og 2,7 m bredt, 3-7 m højt og orienteret nord/syd. Den geografiske placering af hegn kan ses af Figur 2.1.
Figur 2.1. Placeringen af de undersøgte hegn i Danmark . Hegn markeret med cirkel er undersøgt på sandjord i 2001, hegn markeret med stjerne på lerjord i 2002. 2.1.1 Feltundersøgelse2.1.1.1 Prøvefelter 2.1.1.2 Hegnskarakteristika 2.1.1.3 Lysmåling 2.1.1.4 Jordbundskarakteristika Jordprøver indsamledes i oktober 2001 og august 2002. Fem delprøver af jord 0-10 cm under A0 horisonten blev poolet til én prøve. Jorden blev sigtet i laboratoriet gennem 2 mm sigte for at fjerne rødder, sten, mv. pH og ledningsevne blev målt i vandig opløsning. Frisk jord ektraheredes for uorganisk N og P i 1 time i 50 ml 0,5 molær K2SO4. Ekstraktet blev filtreret gennem Whatman GF-D-filter og nedfrosset til analyse for NH4+-N med indophenol metoden, NO3--N med cadmium reduktionsmetoden og PO4-P med molybdæn-blå-metoden (Allen, 1989). PO4-P ekstrationen var meget lav og er derfor udeladt. En delmænge af jorden blev tørret ved 70 grader C og knust. 4 gram heraf brændtes ved 550 grader C. En anden fraktion (200 mg tørvægt) blev opløst i svovlssyre i 1 time og analyseret for total N og P (Kedrowski 1983) med Hitachi U-2000 spektophotometer. C-total måltes med LECO CNS-1000 med metoden efter Sørensen & Bülow-Olsen (1984). Kornstørrelsesfordeling blev foretaget af Forskningscenter Foulum og de kemiske analyser blev foretaget af Økologisk Kemisk Laboratorium, Botanisk Institut, Københavns Universitet. 2.1.1.5 Vegetationsanalyse Til identifikation af planter er anvendt Hansen (1993), for græsser Hubbard (1984), for underarter af Rubus fruticosus (L.) Pedersen & Schou (1988) og for mosser Andersen m.fl. (1976). Der er ikke skelnet mellem underarter af mælkebøtte og rød svingel. Ud fra plantelisterne blev der udregnet en række indirekte hegnskarakteristiska, såsom total diversitet og vægtet gennemsnit af Ellenberg-værdier (Ellenberg m.fl. 1991) for næringsstofniveau (N), temperatur (T), kontinentalitet (K), pH (R), lys (L) og fugtighed (F) for alle prøvefelter. Gennemsnitlige vægtede C-S-R værdier (Grime 1979) blev tilsvarende udregnet for samtlige hegnsprøvefelter med metoden anbefalet af Ejrnæs & Bruun (2000). Total artsliste pr. hegn er fundet ved at registrere samtlige karplanter og mosser på én side i 0,5x100m felt placeret 20 cm fra afgrødekant. Hvert prøvefelt har således i alt 15 direkte og indirekte miljøvariable inklusive jordbundskarakteristika. Habitatpræference for arter er baseret på Hansen (1993) og Smith (1978). Derudover anvendes Grime's (1979) klassifikationssystem, hvor de enkelte arter under britiske forhold tildeles point for hhv C, S og R-strategi. Hansens (1993) klassifikation er enten-eller, mens Grimes er graduereret. Derfor kan en ruderat- eller en ager-art i Hansen godt være delvis C- eller S- strateg i Grimes system. Ukrudtsarter kan forekomme i agerjord men også findes i øvrige naturtyper. Ruderatarter er i øvrige naturtyper beskrevet som også forekommende i ruderate biotoper. Arter klassificeret som halv-natur-arter er ikke i ovennævnte kategorier, men i øvrige lysåbne biotoper. Tilsvarende gælder for skovarter. 2.1.1.6 Arthropod indsamling Leddyrene fra 2001 blev sorteret i henhold til sorteringsskemaet (Bilag C1). På grund af det uventet store antal dyr har vi kun optalt de grupper, som omfatter herbivore arter. De indsamlede prøver fra 2002 er sorteret som prøverne fra 2001, bortset fra, at antallet af bladlus og trips er talt og ikke kun estimeret, mens løbebiller og rovbiller er opgjort i to størrelsesgrupper (Bilag C2). Leddyrene er identificeret vha. Danmarks Fauna (Jensen-Haarup 1912, Hansen 1927; 1951, 1952, 1954, 1965, 1968, Gaun 1974), og nomenklaturen følger denne, bortset fra nogle af tægearterne, som er identificeret vha. Wagner (1961). 2.1.2 DatabehandlingFor vegetation, leddyr og jordbundskarakteristika er effekten af dyrkningsform og placering i hegnene testet i variansanalyse med SAS-proceduren MIXED, idet dyrkningsform og placering i hegn betragtes som systematiske (”fixed”) variable, mens hegnnummer og bloknummer betragtes som tilfældige (”random”) variable (beskrevet i afsnit 1.3.1). Samme type test er brugt til at teste for effekten af andre variable, fx hegnenes ejerforhold. Denne form for analyse forudsætter, at data er normalfordelte, evt. efter passende transformation. Hvis dette ikke kan opfyldes, fx pga. mange nuller i de observerede data, er i stedet anvendt en analyse, som baserer sig på poissonfordelingen (proc GENMOD i SAS), idet antalsdata med mange nuller med rimelighed kan antages at være poissonfordelte. Denne analyse kan ikke på samme måde som proc mixed kvantificere bidraget fra de tilfældige variable til variationen i responsdata. Proc GENMOD er anvendt på alle leddyrdata som supplement til proc MIXED, idet nogle leddyrarter kunne opfylde kravet om normalfordeling for nogle hegn, men ikke for andre. For at tage hensyn til, at der er taget flere prøver i hvert hegn, er anvendt et REPEATED-statement. De sjældneste arter er på grund af de mange nuller ikke egnet til statistisk analyse. I første omgang er data fra sandjord og fra lerjord analyseret hver for sig for at teste effekten af dyrkningsformen i de to geografiske områder. Efterfølgende er de to datasæt slået sammen, og effekten af jordbund er testet som systematisk variabel sammen med dyrkningsform, med hegnnummer hierarkisk til jordbund som tilfældig variabel (se afsnit 1.3.1). I fortolkningen af alle test er anvendt et signifikansniveau på 5 %. Vegetationens diversitet blev beregnet på tre niveauer: α (arter på prøvefelt-niveau), β-diversiteten (artsudskiftningen) og γ-diversiteten (antal arter i hele hegnet). β-diversiteten beregnes ved at udvælge 12 tilfældige sæt af 10 vegetationsprøvefelt fra hver driftstype og beregnes som total artsantal i de 10 vegetationsprøvefelt divideret med gennemsnitligt antal arter i de 12 sæt (Ejrnæs m.fl. 2002). 2.1.1.7 Supplerende analyser af vegetationsdata Plantede træer og buske ligesom dyrkede arter fra landbruget (kulturgræsarter, samt rød- og hvidkløver) blev udeladt af analyserne, men spontane vedagtige arter (kristtjørn, enebær, brombær, hindbær etc.) blev inddraget. Mosser er endvidere udeladt fra denne analyse. 2.1.1.8 Multivariat databehandling Ordinationerne af det totale vegetationsdatasæt (inklusive andre naturtyper) blev foretaget i PCord (McCune & Mefford 1999) uden nedvægtning af sjældne arter. Euclidiske afstande blev beregnet mellem prøvefeltscoren for alle økologiske og konventionelle hegn i dette studie (2x28) i det tre-dimensionelle ordinationsrum i forhold til den gennemsnitlige score for de øvrige naturtyper. Statistiske forskelle mellem Euclidiske afstande blev testet med t-test. Produkt-moment korrelationer beregnedes mellem ordinationsakserne og forklaringsvariable på såvel egne data som det totale datasæt. Signifikante forskelle i forekomsten af enkeltarter blev testet med en indikatorartsanalyse (McCune & Mefford 1999, Dufrene & Legendre 1997). Forskelle i leddyrenes artssammensætning mellem de to dyrkningsformer blev analyseret vha. ANOSIM-analysen i Primer (se 1.3.1.), og arternes bidrag til forskelle mellem dyrkningsformerne blev undersøgt vha. SIMPER. 2.2 Resultater2.2.1 Hegnenes jordbund og strukturAf de i alt 56 hegn der er undersøgt er 25 med græsmark i omdrift som nabo, heraf er 6 konventionelle og 19 økologiske græsmarker. 31 hegn har andre énårige afgrøder (fx. korn, grøntsager) som nabo, heraf 22 konventionelle og 9 økologiske. Hegnene er udvalgt for at ligne hinanden så meget som muligt med pesticidfravær som den væsentligste forskel på de to driftsformer. Forskelle i naboafgrøder er uden tvivl udtryk for de reelle forhold, da det var lettere at finde økologiske hegn der opfylder vore kriterier hos mælkeproducenter med meget kløvergræs. Hegnene var orienteret mod sydøst og bredden af fodposen var i gennemsnit 1,35m. De økologiske hegn var gennemsnitligt 14,7 år gamle og de konventionelle var 13,6. Data for de enkelte hegn forefindes i Bilag A. 2.2.1.1 Hegn på sandjord Tabel 2.1. Middelværdier for jordfysiske og –kemiske målinger i fodposen i hegnene på sandjord samt resultater af variansanalysen af effekter på de fysik-kemiske karakteristika af hegnnummer, blok (tilfældige variabel), dyrkning, position (fodpose eller centralt i hegnet) samt interaktionen mellem disse (systematiske variable). For tilfældige variable er det estimerede procentvise bidrag til den samlede ”tilfældige” variation opgivet. For systematiske variable fremgår signifikansnivauet af variansanalysen. I de tilfælde, hvor der er interaktion mellem de systematiske variable, kan effekten af de enkelte variable ikke bedømmes ud fra p-værdierne.
2.2.1.2 Hegn på lerjord De fleste strukturelle, jordkemiske og jordfysiske karakteristika var ikke signifikant forskellige for de to dyrkningsformer (Tabel 2.2), dog var total-N signifikant højere i de økologiske hegn end i de konventionelle. Variansbidraget fra hegnnummer i forhold til residualbidraget varierede meget mellem de målte responsvariable (Tabel 2.2), men for de fleste bidrog hegnnummeret langt mere til variationen, hvilket viser at der er meget store forskelle mellem hegnene i forhold til variationen inden for hegnene. Tabel 2.2. Resultater af variansanalyse for forskelle i middelværdier for strukturelle, jordfysiske og –kemiske målinger samt effekt af dyrkningsform i hegnene på lerjord. For tilfældige variable er det estimerede procentvise bidrag til den samlede tilfældige variation opgivet. P – Dyrkning = p-værdien for test af forskel mellem økologiske og konventionelle hegn.
2.2.2 Vegetationen2.2.2.1 Artsdiversitet i hegn Af Tabel 2.3 fremgår endvidere, at frekvensen af agerarter og antal arter pr. prøve var en signifikant effekt af dyrkningsformen på sandjorden. Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne, CSR-indikatorerne og antal skovarter. Antallet af arter med dokumenteret viden om herbicidfølsomhed er så lavt (0 i konventionelle hegn, 0,013 pr. prøve i økologiske), at analysen ikke isoleret set giver mening (se dog afsnit 2.2.2.5). Der var klart flere plantearter og højere tæthed i fodposen end midt i hegnene, og forskellen mellem driftsformerne skyldtes forskel i fodposens diversitet. De tilfældige varianskomponenters bidrag fremgår også af tabellen. Variansen mellem hegnene bidrog med mellem 23 og 83 % af den tilfældige variation. Tabel 2.3. Resultater af variansanalyse af udvalgte vegetationsdata for fodposen i hegn på sandjord. Ellenbergværdier er udregnet på basis af tilstede-/ikke tilstedeværelse af arter, CSR-værdier er vægtet med arternes frekvens, opdelt efter forekomst i forskeliige habitattyper, samt antal arter pr. prøvefelt (10m²). For tilfældige variable (hegn og residual) er det estimerede procentvise bidrag til den samlede “tilfældige” variation. For dyrkningsformen fremgår p-værdien af variansanalysen.
På lerjorden fandt vi i gennemsnitligt 36,1 arter i de økologiske hegn mod 24,1 arter i de konventionelle. Artsantallet pr prøvefelt samt antallet af naturarter, ruderatarter og agerarter var signifikant højere i de økologiske hegn på lerjord (Tabel 2.4). Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne, CSR-indikatorerne og antal skovarter. Variansbidraget fra hegnnummer er generelt ikke mere end maksimalt tre gange residualbidraget, hvilket betyder, at variationen mellem hegnene ikke er meget større end variationen inden for hegnene. Tabel 2.4. Resultater af variansanalyse af udvalgte vegetationsdata for fodposen i hegn på lerjord. Ellenbergværdier er udregnet på basis af tilstede-/ikke tilstedeværelse af arter, CSR-værdier er vægtet med arternes frekvens, opdelt efter forekomst i forskeliige habitattyper, samt antal arter pr. prøvefelt (10m²). For tilfældige variable (hegn og residual) er det estimerede procentvise bidrag til den samlede tilfældige variation opgivet. For dyrkningsformen fremgår p-værdien af variansanalysen.
2.2.2.2 Vegetationens sammensætning
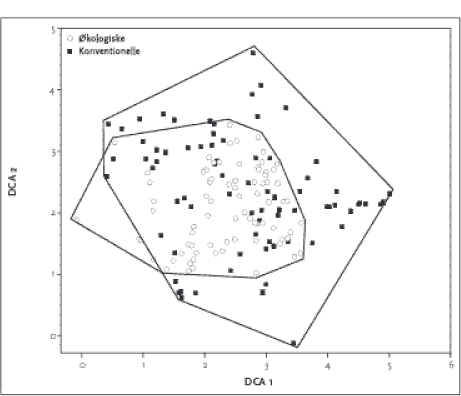
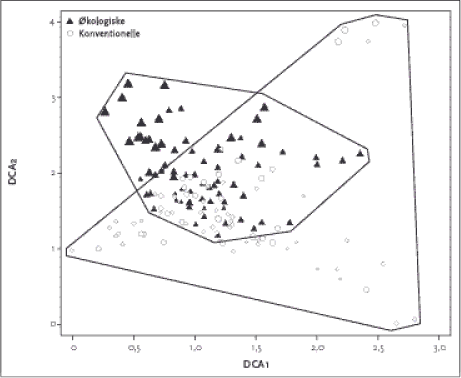
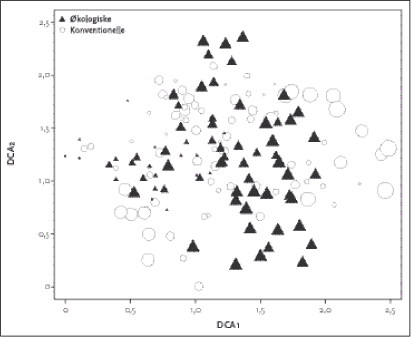
Figur 2.2. Placering af de 155 hegnsprøvefelter på sandjord langs de to vigtigste gradienter i datamaterialet. Polygonerne indikerer det totale areal omfattet af de to dyrkningsformer. Værdierne på DCA1 aksen korrellerede bedst med næringssttofstatus i jorden (Ellenberg N-værdi; P-total), med kornstørrelsesfordeling (mængde grovsand og fint sand) og lys, hvilket indikerer at DCA 1 afspejler en væsentlig gradient i såvel jordbund som lysforhold i artssammensætningen. DCA 2 havde højst korrelation med jordbundsvariable (finsand, P-total) og kanthøjden over markniveau. En DCA analyse af sammensætningen af vegetationen på lerjorden kan ses i Figur 2.3. Førsteaksen er ikke så lang som hos hegn på sandjorden, hvilket delvist skyldes, at der på lerjorden ikke er taget prøver centralt i hegnene. Der var et ganske stort overlap i artssammensætningen hos hhv. økologisk og konventionel drift. Det fremgår også, at der var en længere 1. og 2. akse og dermed større artsforskelle (beta-diversitet) mellem de konventionelle hegn. Produkt-moment korrellationer (Tabel 2.5) viser at DCA 1 på lerjorden primært var korrelleret med Ellenberg-Lys mens andenaksen var bedst korrelleret med Ellenberg-Næringsindhold, som afspejler jordbundsvariation. På sandjord var Ellenberg-Næringsindhold den vigtigste variabel.
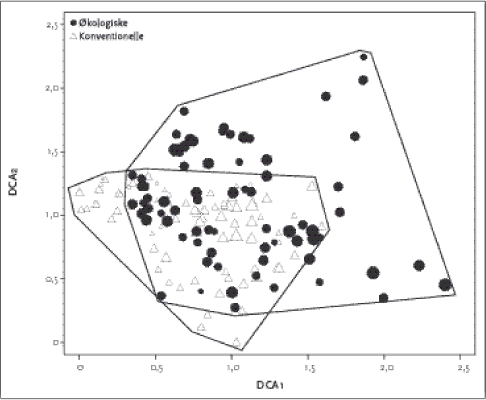
Figur 2.3. DCA plot for vegetationssammensætningen for økologisk (trekant) og konventionelle hegn på lerjord. Størrelsen af symbolet angiver artsdiversiteten af de enkelte hegn. Tabel 2.5. Produkt-momentkorrelationskoefficienter mellem kalibrerede Ellenbergværdier og ordinationsscorene (DCA1 & DCA2) fra sand- og lerjorder.
2.2.2.3 Funktionel fortolkning af vegetationen i alle hegn I alt fandt vi 169 arter af urter, heraf var 153 karplanter inklusive Rubus-arter og 16 mosser. I de 28 økologiske hegn blev der fundet 162 arter og tilsvarende blev der fundet 115 arter i de 28 konventionelle hegn. Antallet af arter pr. prøvefelt var signifikant højere (P<0,0001) i økologiske hegn (19,1) sammenlignet med konventionelle (13,2) (Tabel 2,6). Selvom der var flere græsmarker langs de økologiske hegn, gav græsmarkerne ikke i sig selv nogen signifikant højere diversitet sammenlignet med andre afgrøder. Forskellen mellem driftsformerne viste også ved antallet af ruderatarter, naturarter og agerarter og C-strateger er signifikant højere i de økologiske hegn (Tabel 2.6). En uvægtet analyse med CSR-indikatorerne viste signifikant flere S-strateger i de økologiske hegn. Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne og antal skovarter. Variansanalysen viste endvidere at der var signifikant effekt af jordbunden på en række af Ellenberg-indikatorerne, C-strateger samt antallet af ruderat-arter. Variansbidraget fra hegnnummer var op til tre gange residualbidraget, hvilket betyder, at variationen mellem hegnene var større end variationen inden for hegnene. Tabel 2.6. Resultater af variansanalyse i alle økologiske (n=84) og konventionelle (n=84) hegn. For hegnnummer inden for jordtype og residualeffekt fremgår det estimerede procentuelle bidrag til den samlede tilfældige variation. P-værdien for effekt af dyrkningsform (økologisk/konventionel) samt effekt af jordbund (sandjord/lerjord) er opgivet Interaktionsleddet mellem dyrkning og jordbund er ikke medtaget i variansanalysen, da det ikke var signifikant.
2.2.2.4 Vegetationens sammensætning og relationer til andre naturtyper Tabel 2.7 Produkt-moment korrelations koefficienter mellem kalibrerede Ellenberg værdier og ordinations scorer (n =1661) for DCA1-DCA3. Fede tal angiver værdier > 0.5.
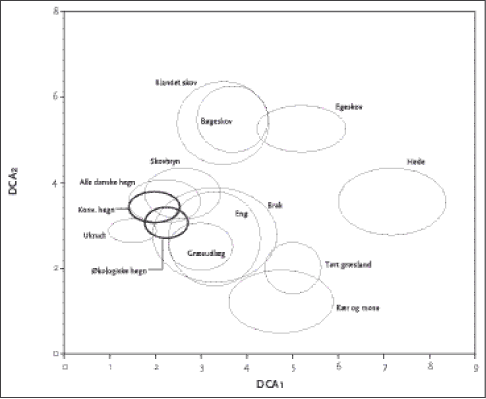
Figur 2.4. DCA figur der viser de undersøgte hegnsdata (de to fede ringe) i relation til en række udvalgte naturtyper. Det ses af Tabel 2.8, at de økologiske hegn havde signifikant større affinitet end konventionelle hegn til lysåbne kultiverede og stærkt påvirkede elementer som ukrudtsflora på marker, græsmark, dyrket eng og mose, samt i mindre grad til tørt græsland og hede. Endvidere havde økologiske hegn større affinitet til skovbryn end konventionelle. Affiniteterne til skovtyperne er relativt perifere, og vi fandt kun meget få obligate skovarter i hegnene, især i de konventionelle. Tabel 2.8. Gennemsnitlig euclidisk afstand mellem ordinationsscoren for hhv. økologiske (n=28) og konventionelle (n=28) og 12 naturtyper. T-test er anvendt til at beregne statistisk forskel mellem middelværdierne. Rækkerne er sorteret efter nærmeste Euclidiske afstand til økologiske hegn N.S.= ikke signifikant, ** = 0.001 <P<0.01, *** = P < 0.001. Fede tale indikerer, hvilken af de to hegnstyper der har signifikant affinitet til naturtyperne.
En indikatorartsanalyse viste, at tyve arter forekom signifikant hyppigere på en af driftstypene (på P<0,1-niveau) (Tabel 2.9). Mosset Brachythecium rutabulum (alm. kortkapsel) og urterne alm. hønsetarm, lav ranunkel, agertidsel, alm. svinemælk og lancetvejbred kan alle siges at være indikatorarter for økologiske hegn. Det er dog kun sidstnævnte der kun forekom i økologiske hegn, mens de øvrige blev også fundet i konventionelle hegn, men signifikant mindre hyppigt. Tabel 2.9. Indikatorarts-analyse af driftsform. Arterne er sorteret efter stigende p-værdi. Indikatorværdi og statistisk signifikans er beregnet med 1000 MonteCarlo permutationer. Arter med P ≥ 0.1 er ikke vist. Plantede hegnsarter og kulturplanter fra græsmarker er udeladt.
2.2.2.5 Herbicidfølsomhed Vi har kunnet konstatere, at der forekommer under 10 'glyphosat-følsomme' arter i vore hegn, og ingen af disse er særligt almindeligt forekommende. Derfor er der ikke foretaget en statistisk analyse af, om der er flere herbicidfølsomme plantearter i økologisk hegn jf. hypotese 2. Vi må konstatere, at den nødvendige viden pt. ikke er tilstede. 2.2.2.6 Sjældne arter 2.2.3 Leddyrfaunaen2.2.3.1 Data fra hegn på sandjord Som det fremgår af Tabel 2.11, var der på sandjord ikke signifikante forskelle mellem dyrkningsformerne på forekomsten af udvalgte arter og gruppe. Antal arter pr. prøve, antal dyr pr. prøve og diversiteten (Shannon-Wiener indeks) var heller ikke signifikant påvirket af dyrkningsformen. Derimod var der for de fleste responsvariable en signifikant forskel mellem prøver taget inde i hegnet og prøver taget i fodposen, og for alle disse gælder, at der gennemsnitligt var flere dyr i i fodposen (Tabel 2.10). Analysen af de tilfældige variable viser, at der ikke er nogen blokeffekt, hvorimod variansen mellem hegnene i de fleste tilfælde udgør en tredjedel af den tilfældige variation eller mere (Tabel 2.11). Numerisk set er variansen mellem hegnene i de fleste tilfælde meget større end variationen mellem dyrkningsformerne. DCA analysen på alle data med undtagelse af Stilbus sp., som lå uden for hovedsværmen, viste, at prøverne fra de konventionelle hegn havde en længere førsteakse end prøverne fra økologiske hegn (Figur 2.5). For andenaksen gør det modsatte sig gældende. Førsteaksen er bedst korreleret til finsand, total-P, jordbundstypen, og lysforholdene (Tabel 2.12), mens andenaksen er bedst korreleret til total-N, ledningsevnen og humusindholdet. At dømme efter størrelsesfordelingen af signaturerne i diagrammet er gradienten i finsand langs førsteaksen ret entydig for økologiske hegn, mens den er temmelig utydelig for de konventionelle hegn. Tabel 2.10. Gennemsnitlige antal af de forskellige leddyrgrupper pr. prøve (0,9 m²) i de to typer hegn på sandjord hhv. i fodposen (ude) og midt i hegnet (inde).
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. Tabel 2.11. Resultater af variansanalyse af udvalgte leddyrdata fra hegn på sandjord. For tilfældige variable er det procentvise bidrag til den samlede tilfældige variation opgivet. For effekten af systematiske variable, dvs. dyrkningsform, position i hegnet (inde/ude) samt interaktionen mellem de to variable, fremgår signifikansnivauet af variansanalysen. I de tilfælde, hvor interaktionen er signifikant, bør man se bort fra p-værdierne for de enkelte variable.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl.
Figur 2.5. DCA analyse på arthropod-data for samtlige 156 prøver på sandjord. Størrelsen af symbolet angiver forekomsten af finsand i feltet (som havde den stærkeste korrelation med 1. aksen, se Tabel 2.12). Hvis man kun ser på prøverne fra fodposerne (Tabel 2.13), er førsteaksen mest korreleret til lysforholdene og total-P og derefter til forekomsten af ruderat-planter. På andenaksen er finsand og grovsand vigtigst, men ellers er det de samme variable som for førsteaksen. Desuden viste ANOSIM-analysen af leddyrdata (se 1.3.1), at der var signifikante forskelle i sammensætningen af leddyrfaunaen mellem hegn på økologiske og hegn på konventionelle bedrifter (p<0,01). Analysen af, hvilke arter der betød mest for forskellene i artssammensætning mellem hegnene i de to dyrkningssystemer på sandjord, viste, at ingen arter eller gruppe bidrog med mere en 10 % (SIMPER, udregnet på baggrund af kvadratrodstransformation, se 1.3.1). Størst forklaringsværdi havde de meget talrige grupper (tæppespindere, stilkhvepse, fluer, myg, bænkebidere, delphacidae-nymfer), der tilsammen bidrog med 31,5 % og hver især med maks. 7,24 %. De enkeltarter, der bidrog mest til forskellen, var snudebillerne Apion flavipes og A. virens, som forklarede hhv. 2,48 og 2,45 %. Som det fremgår af Tabel 2.14, havde ingen af disse de plantearter, der adskilte de to hegnstyper (Tabel 2.9), som værtsplanter. En fjerderodstransformation, som ændrer vægtningen til fordel for sjældnere arter, gav ingen forskel i analysen af enkeltarters bidrag til forskellene i artssammensætning mellem hegnstyperne. Tabel 2.12. DCA-analyse af leddyr overfor udvalgte miljøvariable. ”fr.” = frekvens, ”ts.” = til stede/ikke til stede. Analysen er udført på det fulde datasæt, med undtagelse af Stilbus sp. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
* Udregnet på baggrund af artssammensætningen i feltet Tabel 2.13. DCA-analyse af leddyr overfor udvalgte miljøvariable. ”fr.” = frekvens, ”ts.” = tilstede/ikke tilstede. Analysen er udført udelukkende på data fra fodposen i hegn på sandjord. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
* Udregnet på baggrund af artssammensætningen i prøvefeltet Tabel 2.14. Fødekilder for udvalgte leddyr.
2.2.3.2 Leddyrdata fra hegn på lerjord Variansanalysen viste, at hegnsnummerets bidrag til variationen i leddyrenes forekomst var lige så stor eller op til dobbelt så stort som det residuelle bidrag, dvs. variationen inden for de enkelte hegn (Tabel 2.16). Snudebillen Apion flavipes og det totale antal dyr per prøve var dog mere påvirket af hegnsnummeret. Der var signifikante forskelle mellem de to hegnstyper (økologisk og konventionel) for skumcikader (flest i konventionelle hegn), de tre tægearter Scolopostethus affinis, S. thomsoni og Stenoderma holsatum (flest i økologiske hegn), snudebillerne Apion flavipes, Ceutorrhynchus assimilis (flest i økologiske hegn) og Ceutorrhynchus contractus (flest i konventionelle hegn). Overordnet set var der signifikant flere (15 %) arter/grupper i de økologiske hegn end i de konventionelle. Tabel 2.15. Gennemsnitlige antal leddyr per prøve (0,9 m²) i hhv. økologiske og konventionelle hegn på lerjord.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. Tabel 2.16. Resultat af variansanalyse på udvalgte leddyr fra hegn på lerjord. For de tilfældige variable fremgår det estimerede procentvise bidrag til den samlede tilfældige variation. For den systematiske variabel, dyrkningsformen, er p-værdien for test af forskel mellem økologiske og konventionelle hegn i hhv. proc mixed og proc genmod opgivet.
DCA-analysen på leddyrdata fra lerjorden (Figur 2.6) viste, at både første- og andenaksen var længere for økologiske hegn end for konventionelle. Derved lå en hel del af punkterne for økologiske hegn og kun enkelte af punkterne for konventionelle hegn uden for den fælles punktsværm. Førsteaksen var bedst korreleret med total-P, pH, antal naturarter (planter), antal plantearter ialt og vandprocenten i jorden (Tabel 2.17), mens andenaksen var positivt korreleret med hegnets alder og antal agerarter (planter) og negativt korreleret med fodposebredde og antal skovarter (planter). Med andre ord var begge akser korreleret med både jordbundskarakteristika og planterelaterede variable. ANOSIM-analysen af leddyrdata (se 1.3.1) viste, at der var signifikante forskelle i sammensætningen af leddyrfaunaen mellem hegn på økologiske og hegn på konventionelle bedrifter (p<0.01) Trips, delphacidae-nymfer og små rovbiller, som alle var almindelige i hegnen, var de grupper, der forklarede mest af forskellen i artssammensætning mellem økologiske og konventionelle hegn, nemlig tilsammen 14,3 % og hver for sig maks. 6,45 % (analyseret vha. SIMPER efter kvadratrodstransformation af data, se 1.3.1). Ligesom på sandjorden var snudebillearterne Apion flavipes og A. virens de enkeltarter, der foklarede mest af forskellen, hhv. 3,37 og 2,88 %. Som det fremgår af Tabel 2.14, havde ingen af de plantearter, der adskilte de to hegnstyper (Tabel 2.9), som værtsplanter. En fjerderodstransformation, som ændrer vægtningen til fordel for sjældnere arter, gav ingen forskel i analysen af enkeltarters bidrag til forskellene i artssammensætning mellem hegnstyperne.
Figur 2.6. DCA analyse på arthropod-data for hegn på lerjord. Prøver afbilledet tæt på hinanden har ens sammensætning af leddyrfaunaen. Størrelsen af symbolet angiver artsdiversiteten. Tabel 2.17. DCA-analyse af leddyr fra hegn på lerjordoverfor udvalgte miljøvariable. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
2.2.3.3 Alle leddyr – sand- og lerjord Tabel 2.18. Effekt af dyrkningsform, år og hegnnummer på udvalgte leddyr. For de tilfældige variable (hegnsnummer inden for jordtypen og det residuelle bidrag) er angivet det estimerede procentvise bidrag til den samlede tilfældige variation.For de systematiske variable ses p-værdien fra variansanalysen. ’+’ indikerer flere dyr på lerjord end på sandjord, ’-’ færre dyr på lerjord.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. 2.2.3.4 Samspil mellem vegetation og leddyr – fødekæder En indikatorartsanalyse afslørede at nogle arter fandtes signifikant (P < 0.05) mere i hegn (kun fodposen) med græsmarker som naboafgrøde, og andre foretrak hegn der grænsede op til andre afgrøder (Tabel 2.20). En tilsvarende analyse mht. dyrkningsform er præsenteret i Tabel 2.21, som viser, at løbebiller samt en enkelt tægeart og kornbladbillen var hyppigere i hegn ved konventionelt dyrkede marker, men bladlopper, svirrefluer og nogle snudebiller foretrak hegn ved økologisk dyrkede marker. Tabel 2.19. Forekomsten af herbivore leddyr i relation til udvalgte strukturelle og planterelatede variable på sandjord.
*Prøver med færre end tre arter og arter som forekommer i færre end tre prøver er ikke medtaget. Tabel 2.20. Indikatorarters fordeling i forhold til om hegnet har græsmark eller andre afgrøder som nabo på sandjord.
Tabel 2.21. Indikatorartsanalyse af betydende arter/dyregruppers fordeling i forhold til dyrkningsform på de marker, der grænser op til hegnet på sandjord. De arter/grupper, der fandtes signifikant (P < 0.05) hyppigere i fodposen i hegn på hhv. økologiske og ikke økologiske bedrifter er vist.
2.3 Diskussion2.3.1 Hegn og jordbundDer var ingen forskelle mellem dyrkningsformen, hvad angår de strukturelle og jordbundsfysiske og kemisk karakteristika i hegn på sandjord. Derimod var der en del forskel mellem fodpose og den centrale del af hegnene, som formentlig skyldes vindaflejret materiale og gødningsspredning i markkanten (Tabel 2.1). På de bedre jorder fandt vi, at total-N var signifikant højest i økologiske hegn (Tabel 2.1 og 2.2), men det er uvist, hvordan denne forskel kan relateres til driftsformen. 2.3.2 Vegetation2.3.2.1 Artsdiversitet Generelt må det forventes, at artspuljen potentielt er næsten den samme for alle de inkluderede hegn, fordi jordbunds- og hegnsvariable ikke viser store forskelle. Der er dog i en række studier fundet flere arter på økologiske marker (Reganolds m.fl. 1993, Hald 1999, Stolze m.fl. 2000, Mäder 2002, Rydberg & Milberg 2000), så puljen af arter, der kan etablere sig fra marker, er lidt større på økologiske bedrifter. Der er ikke tidligere konstateret så klare forskelle mellem konventionelle hegn og økologiske hegns vegetation (Boutin & Jobin 1998, 2001). Andre studier har undersøgt markdriftens indflydelse på hegnsvegetationen over en kort årrække (Hald 1998, Marshall m.fl. 2002), men da hegnsvegetationen har vanskelige vilkår for spredning og etablering i et fragmenteret landskab kan man ikke forvente store effekter på få år (Milchunas & Lauenroth, 1995). Da de undersøgte hegn endvidere var relativt unge, er sandsynligheden, for at der er sket omfattende indvandring af mange arter fra halvnaturtyper og skov, ikke stor og formentligt ikke væsentligt forskellig på de to driftsformer. Udryddelsesraten kan formodes at værre større på konventionelle bedrifter som følge af herbicidanvendelsen, da selv små doser i afdrift kan påvirke den naturlige vegetation (Kleijn & Verbeek 2000, Boutin & Jobin 1998, 2001). Forskellene i artsantal kan således forklares ved forskelle i indvandringsrate af markarter (flest på økologiske bedrifter) og forskelle i udryddelsesrater af herbicidfølsomme arter ved de to driftsformer (Zobel m.fl., 1998). En række studier i England og Frankrig har tidligere vist, at landbrugsdriften har indflydelse på hegnsvegetation (Boatman m.fl., 1994; Cummins & French, 1994; Hegarty m.fl., 1994; McAdam m.fl., 1994; Le Coeur m.fl., 2002), således at mere ekstensive driftsformer giver højere diversitet. De vigtigste påvirkninger er afdrift af pesticider og ammoniak samt fejlplaceret kunstgødning på konventionelle bedrifter (Baudry m.fl., 2000; McCollin m.fl., 2000). Det er generelt accepteret, at en af de væsentlige påvirkninger af markkantvegetation er tilførsel af store næringsmængder (Hegarty m.fl., 1994, Barr m.fl. 1995, Kleijn & Snoeijing 1997, Kleijn & Verbek, 2000; Tybirk m.fl., 2001), som resulterer i dominans af nitrofile C-strateger med eliminering af S-strateger (sensu Grime 1979). Eutrofieringen er også markant i de økologiske hegn, og selvom vi kan konstatere forskelle mellem de to driftsformer er alle hegn eutrofe. I nærværende studie har antagelsen derfor været, at der ikke er mangel på næringsstoffer i de inkluderede hegn, så de alle forventedes at være domineret af mere eller mindre nitrofil vegetation. Driftsformerne var i princippet lige intensive i dette studie mht. næringsstofanvendelse, omend ammoniaktabet er lidt lavere på økologiske bedrifter (Kristensen m.fl. 2003). Driftsformerne var derimod forskellige mht. anvendelsen af pesticider gennem minimum 10 år, så det vi ser kan fortolkes som langtidseffekter af pesticidfravær. Et nyligt afsluttet studie inspireret af tesen om økologisk inerti i agerlandet (Milchunas & Lauenroth 1995, Reddersen m.fl. 1999) har dokumenteret en signifikant ændring i vegetationen og fluefaunaen i gamle skel og hegn på økologiske kvægbedrifter på sandjord allerede 3,5 år efter omlægning til økologisk drift (Petersen 2003, Petersen m.fl. 2003). Metoderne er meget sammenlignelige med vore, og resultaterne var ganske overbevisende på gamle skel efter få års omlægning. For vegetationens vedkommende forstærkes effekterne efter 7 års omlægning. Man kan således konstatere, at herbicidfravær giver effekter meget hurtigere end tidligere forventet. Det kan forklares ved, at hegnsvegetationen løbende rekrutterer arter fra den eksisterende frøbank og via langdistancespredning fra omkringliggende biotoper. I begge tilfælde har de indvandrende planter større overlevelse på økologiske bedrifter pga herbicidfravær, så ekstinktionsraten er lavere. 2.3.2.2 Funktionel fortolkning På trods af en del ligheder i sammenligning med andre naturtyper i landskabet, fandt vi dog også klare forskelle mellem de to hegnstyper. De økologiske hegn havde således flere mosser, flere ukrudts- og ruderatplanter og klarere affiniteter til lysåbne og mere forstyrrelsesprægede landskabselementer som ukrudtssamfund på marker, græsmark og dyrket eng (Tabel 2.3, 2.4 og 2.8, Figur 2.4). Endvidere havde de økologiske hegn flere arter fra de lysåbne halvnaturtyper, og derfor er de økologiske hegn et bedre levested for halvnaturarter. I studiet, som fokuserede på økologiske kvægbedrifter (Petersen 2003), fandtes signifikant flere S-strateger i hegn og skel på de økologiske bedrifter og flere Ruderatstrateger og højere Ellenberg N-værdier i hegn og skel på konventionelle. Billedet i nærværende undersøgelse var ikke så entydigt, både som følge af at jordbunden er meget mere variabel, og fordi vi har inkluderet forskellige bedriftstyper for at opnå tilstrækkelige hegn til sammenligningen. De arter, der fandtes hyppigere i økologiske hegn, var primært natur-, agerlands- og ruderatarter (Tabel 2.6). Dette stemmer fint overens med konklusionerne af Petersen (2003), at arter der har relativt let ved at kolonisere (de fleste ruderater og visse langdistancespredte naturarter), klarer sig bedre uden herbicidafdrift på de økologiske bedrifter. 2.3.2.3 Indikatorarter og refugier Alle hegn i vort studie var eutrofe og temmelig forstyrrede set fra en økologisk vinkel. Langt de fleste arter er meget almindelige og relativt uinteressante i relation til national og international naturbeskyttelse, men diversiteten er naturligvis væsentlig for bredden af de fødekæder der kan etableres. I England har man konstateret en reduktion i hegnsvegetationens diversitet på 14 % mellem 1978 og 1990 (Bunce m.fl. 1999). Vi kender ikke den historiske udvikling i hegnsdiversiteten i Danmark, men man kunne forestille sig et tilsvarende billede med stigende intensitet af landbruget. Der er interessante ligheder mellem de arter der er gået tilbage i England og de arter vi har fundet flere af i de økologiske hegn. 11 af 16 arter, der forekom signifikant mere i danske økologiske hegn, er gået tilbage i England i nævnte periode. Det gælder for alm. røllike, alm. hvene, alm. hønsetarm, agertidsel, rød svingel, fløjlsgræs, lancetvejbred, lav ranunkel, rødknæ, mælkebøtte og mark-ærenpris. Andreasen m.fl. (1996) har desuden fundet signifikant tilbagegang i Danmark for alm. hønsetarm, der var en indikatorart for økologiske hegn. Derimod er burresnerre, som var indikatorart for konventionelle hegn i denne undersøgelse fundet i fremgang i England (Bunce m.fl. 1999). Resultaterne i nærværende studie tyder således på, at økologiske hegn kan spille en væsentlig rolle som et bedre refugium for relativt almindelige arter, som dog er i tilbagegang i agerlandet i dag. Man kan endvidere forestille sig at ældre økologiske hegn kan spille en betydeligt større rolle i denne sammenhæng som refugium for bl.a. skovarter (Stidsen 2004). 2.3.3 LeddyrFor læhegnene på sandjorder (2001) kunne der ikke påvises signifikante effekter af dyrkningsformen på leddyrene via variansanalyserne. Dette skyldes meget små forskelle imellem dyrkningsformerne i de undersøgte variable (Tabel 2.10). På den anden side viste de multivariate analyser, at artssammensætningen var signifikant forskellig i hegnene på de to bedriftstyper, og at variationen i leddyrmaterialet kunne forklares lige så godt eller bedre end variationen i plantedata. Det faktum, at dyrene fordeler sig nogenlunde efter samme miljøvariable som planterne, inklusive dyrkningsformen, er formentlig udtryk for at dyrene fordeler sig efter planterne og ikke efter jordbundskarakteristika i sig selv. Leddyrfaunaen i de to år, dvs. på de to jordtyper, var forbavsende ens, men der var dog større forskelle mellem de to år end mellem de to dyrkningsformer. På lerjorden var der signifikante effekter af dyrkningsformen på artsantallet og på forekomsten af visse arter, hvilket delvis kunne genfindes i den samlede analyse af leddyrene. Forskellen mellem årene (og mellem hegnene) følger forskellen i jordbundsforhold og plantediversitet, og i hegnene på lerjord var der også en signifikant forskel i leddyrenes artssammensætning mellem de to dyrkningsformer. Analyserne af fordelingen af leddyr i forhold til planter og strukturelle variable viste, at de plantefordelende variable (primært jordbund) generelt var de vigtigste for de planteædende leddyr, men især for snudebiller var de omgivende markers afgrøde (græsning – ikke græsning) og dyrkningsform også vigtige variable. Disse to variable er ikke totalt uafhængige, idet frekvensen af hegn med græsmark som nabo var højere på de økologiske brug end på de konventionelle. Visse plantefamilier syntes også at påvirke forekomsten af tæger og snudebiller, uden at der dog nødvendigvis var tale om dyrenes værtsplanter. Snudebillernes præference for græsmarker som nabo (Tabel 2.19 og 2.20) kan evt. hænge sammen med forekomsten af kløver på disse marker, idet kløver er værtsplante for en del snudebillearter (Tabel 2.14). Sammenhængen mellem forekomsten af tæger og græs (poaceae) samt skærmplanter (Apiaceae) i hegnene synes rimelig, idet flere tægearter lever på disse typer af planter. 2.3.3.1 Fødekæder- samspil mellem planter og leddyr 2.3.4 HypoteserneDen første hypotese for hegnene var, at driftsformen ville påvirke artsantallet og artssammensætningen. Hypotesen kan bekræftes for planterne i nærværende undersøgelse. For de herbivore leddyr var det ikke muligt direkte at påvise sådanne effekter på artsantallet på sandjord, hvorimod der på lerjord og i det samlede datasæt var signifikant flere arter i de økologiske hegn end i de konventionelle, ligesom der var en signifikant korrelation mellem artsfordelingen og dyrkningsformen i DCA og signifikante forskelle i leddyrenes artssammensætning mellem de to dyrkningsformer. Den anden hypotese var, at der ville være flere herbicidfølsomme plantearter og flere specialiserede (fx værtsspecifikke herbivore) leddyr i de økologiske hegn end i de konventionelle. Det er vi ikke i stand til at konkludere om, idet vi kun har identificeret få dokumenteret herbicidfølsomme arter blandt de fundne, og vidensgrundlaget generelt har vist sig mangelfuldt til en sådan analyse. Det øgede antal plantearter i økologiske hegn giver i princippet mulighed for flere speciealiserede leddyr, men dette er ikke undersøgt i detaljen. Tredje hypotese forudsagde, at antallet af skovarter ville være større midt i økologiske hegn end midt i konventionelle. På grundlag af denne undersøgelse kan hypotesen imidlertid ikke bekræftes, idet der kun blev fundet 13 skovarter i alt og kun ganske få individer. For plantearter delvist tilknyttet skov ('fakultative' skovarter) var der dog signifikant flere i midten af de økologiske hegn end i de konventionelle. De økologiske hegn viste desuden i den totale DCA analyse bedre korrelationer med skovbryn end konventionelle. Det er klart, at hegnene generelt har været for unge til, at der kan være indvandret ret mange skovarter, der typisk er karakteriseret ved langsom spredning. Det kan derfor ikke udelukkes, at der vil være signifikante forskelle på ældre hegnstyper.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||