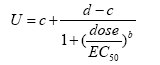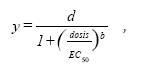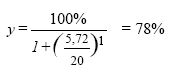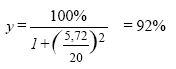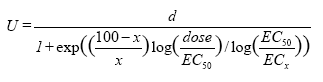|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 89, 2004 Pesticiders påvirkning af planter og alger i vandmiljøetIndholdsfortegnelse3 Giftigheden af herbicider i vand 4 Artspecifikke forskelle i følsomhed 5 Giftighedens afhængighed af tid, effekt variabel og naturlige stressfaktorer
8 Vækststimulering ved lave pesticid-koncentrationer
ForordDenne publikation søger at belyse effekterne af de pesticider, der findes i det danske vandmiljø på den akvatiske flora. Arbejdet er udført på Den Kongelige Veterinær- og Landbohøjskole (KVL) på Institut for Jordbrugsvidenskab, sektion for Ukrudtslære og på Center for Dansk JordbrugsForskning, Flakkebjerg. Vi vil gerne takke medlemmerne af projektets følgegruppe for godt selskab og konstruktive kommentarer gennem tiden, og Anja Weibel Stubbe for hendes engagement og omhyggeligt udførte arbejde i laboratoriet. Nina Cedergreen, Jens C. Strebig og Niels Henrik Spliid Sammenfatning og konklusionerVi har i dette projekt undersøgt pesticiders påvirkning af planter og alger i vandmiljøet. Vi har primært fokuseret på herbiciderne, ukrudtsmidlerne, da det er disse stoffer, der formodes at have den største effekt på planter. For at få det bredest mulige billede af giftigheden af herbicider i vand lavede vi et database-studie på omkring 150 forskellige herbicider tilhørende 15 forskellige fysiologiske/biokemiske virkemåder. Den akvatiske toksisitet, givet som EC50 på alger, blev sammenlignet med den terrestriske toksisitet, estimeret ud fra den anbefalede markdosis. Generelt set, var herbicider relativt uvirksomme i vand. For mere end halvdelen af de herbicider, der indgik i studiet, ville en fuld markdosis udsprøjtet på 30 cm vand ikke kunne reducere algers vækst med mere end 50%. Toksisiteten i databasestudiet var bekræftet i laboratorieforsøg med standard algetests og Lemna (andemad)-tests på 10 repræsentative herbicider. For 7 af de 10 herbicider var toksisiteten overfor Lemna dog 10-1000 gange højere end toksisiteten overfor alger, hvilket understøtter vigtigheden af også at indrage højere planter i risikovurderingen af pesticider. To af de potentielt mest toksiske herbicider, som forhandles og bruges i Danmark, blev udvalgt til yderligere undersøgelse. Dette var sulfonylureaherbicidet metsulfuron-methyl og s-trazinen terbuthylazin. På disse to stoffer blev den artspecifikke forskel i følsomhed mellem 10-12 forskellige arter af vandplanter undersøgt under to vækstvilkår, og arts-følsomheds-fordelinger (SSD's) udregnet. Resultaterne viste, at forskellen i følsomhed mellem arterne var relativt lille, en faktor 56 og 34, sammenlignet med forskellen mellem terrestriske arter, der kan variere med op til en faktor 1000. Forsøgene viste også, at standard-testen med Lemna minor var representativ for vandplanterne, og at standard-testen med algen Pseudokirchneriella subcapitata er en god repræsentant for følsomheden af et naturligt udviklet epifytsamfund. Hvis man bruger en sikkerhedsfaktor på 10 mellem de højest forventede koncentrationer i miløet (Predicted Environmental Concentrations (PEC) udregnet som en fuld markdosis opblandet i 30 cm vand) og NOEC, defineret som EC10, eller en faktor 100 mellem PEC og EC50 på Lemna-standard testen, vil man beskytte >95% af vandplante-arterne. Da de højeste koncentrationer af herbicider formodes at forekomme i pulse af kortere varighed i forbindelse med sprøjtning eller nedbørshændelser, blev effekten af en pulseksponering med forskellige herbicider på Lemna minor undersøgt. Herbiciderne repræsenterede 3 virkemåder og havde forskellig Kow (et udtryk for fedtopløselighed, der også påvirker hastigheden, hvormed stoffer optages og frigives fra planter) indenfor hver virkemåde. Forsøgene viste, at planternes vækst efter 4 dage var upåvirket af en 3-timers pulseksponering med koncentrationer, der var dødelige under længerevarende (4 dage) eksponering for de to fotosyntesehæmmere diquat og terbuthylazin. For ALS-hæmmerne imazamox og metsulfuron-methyl og mikrotubuli-hæmmerne propyzamid og pendimethalin havde pulsen en effekt, og EC50 for pulseksponeringern var omkring en faktor 10 højere end EC50 for den langvarige eksponering. Der var ingen forskel mellem stoffer med samme virkemåde, men med forskellig Kow. Årsagen til den lille effekt af en pulseksponering for fotosyntesehæmmerne kunne skyldes, at planterne har et apparat til udbedring af skader forårsaget af oxygen-radikaler, som er den type skader, disse herbicider forårsager. En hæmning af celledelingen er tilsyneladende sværere at komme sig over, og for metsulfuron-methyl viste det sig, at det tog ca. 4 dage for planter, eksponeret til en næsten dødelig puls, at opnå vækstrater sammenlignelige med ikke-eksponerede planter. Den samlede konklusion på samtlige forsøg var, at herbicider i de koncentrationer der er målt i dansk overfladevand (<10 μg l-1), ikke vil nedsætte væksten mærkbart eller ændre væsentligt på artssammensætningen af neddykkede vandplanter. Vi observerede i flere forsøg, at lave doser af herbicider havde en stimulerende effekt på plantevæksten. Et fænomen, der betegnes hormesis. Nogle svampemidler er også rapporteret at kunne stimulere effekten af væksten af terrestriske planter, og vi så en tilsvarende effekt for svampemidlet prochloraz tilsat i lave doser (10 og 20 μg l-1) til Lemna, dyrket under suboptimale vækstforhold. Så snart vækstforholdene blev optimeret, forsvandt effekten. Prochloraz (< 100 μg l-1) påvirkede ikke toksisiteten af hverken metsulfuron-methyl eller terbuthylazin. Selvom de herbicidkoncentrationer, der er blevet målt i dansk overfladevand, alene ikke vil kunne påvirke den akvatiske flora betydeligt, er det mere usikkert, hvilken effekt tilstedeværelsen af flere stoffer sammen kan have. Vi præsenterer her den Additive Model, som kan bruges til at estimere effekten af flere herbicider, når de er til stede sammen under forudsætning af, at man har toksisitetsdata for de enkelte stoffer. I et eksempel, der tager udgangspunkt i en puls-hændelse med tre fotosyntesehæmmere, viser vi, at de tre stoffer tilsammen potentielt kan nå niveauer, der ifølge den additive model kunne have en effekt. Den Additive Model er et godt redskab til at identificiere potentielt kritiske eksponeringssituationer, som kræver yderligere undersøgelser. Et blandingsforsøg med metsulfuron-methyl og terbuthylazin viste, at de to stoffer tilsammen giver en mindre effekt end forventet på baggrund af den additive model. Dette ses ofte og understøtter brugen af den additive model til at estimere den værst tænkelige situation. Da formålet med vores projekt var at fokusere på lave effektkoncentrationer, har vi diskuteret definitionen af "ingen effekt". Vi har valgt at bruge EC10 og argumenterer for brugen af ECx niveauer fremfor de meget brugte termer NOEC og LOEC, da ECx værdier er mere konsistente på tværs af testsystemer. Variansen af ECx giver desuden et indtryk af experimenternes kvalitet. Der hvor der er størst usikkerhed omkring virkningen af pesticider på den akvatiske flora i søer og vandløb er i situationerne, hvor pesticiderne findes i blandinger. En større viden om sammensætningen og tidsforløbet af blandinger og deres effekt på floraen ville kunne bidrage til at udpege steder og situationer, hvor pesticid-blandinger kunne opstå i koncentrationer over tidsforløb, der kunne påvirke den akvatiske flora væsentligt. Vi mener heller ikke, at mekanismerne for optagelsen af pesticider i vand er belyst tilfredsstillende, da der er indikationer på, at optagelsen i vand er væsentlig forskellig fra optagelsen i det terrestriske miljø. En bedre forståelse af mekanismerne for optagelse af pesticider i vand kunne lede til en højere grad af forudsigelse af pesticiders og andre kemiske stoffers akvatiske toksisitet. Overordnet set kan det dog konkluderes, at de koncentrationer af herbicider, der måles i Danmark, enkeltvis ikke vil påvirke væksten og artssammensætningen af floraen i det danske vandmiljø væsentligt. Summary and conclusionsThe aim of this study has been to investigate the effect of pesticide residues on aquatic plants and algae in the Danish environment. Our primary focus has been on herbicides, as they are products designed to kill plants and therefore are believed to be the group of pesticides with the largest potential effect on the aquatic flora. To get a broad view of the aquatic toxicity of herbicides we did a database study on approximately 150 different herbicides, representing 15 different modes of action. The aquatic toxicity, expressed as the EC50 of algae, was compared to the terrestrial toxicity, estimated as the average recommended field dose. Generally, herbicides were very inefficient in water. For more than half the herbicides included in this study, a full field dose sprayed on a 30 cm water column could not reduce algal growth by more than 50%. The low aquatic toxicity was verified in laboratory experiments on both Lemna minor (Duckweed) and the algae Pseudokirchneriela subcapitata with 10 selected herbicides. Two of the potentially most toxic herbicides sold and used frequently in Denmark were chosen for further investigation. These were the sulfonylurea herbicide metsulfuron-methyl and the s-triazine terbuthylazine. For these two herbicides, species specific sensitivity was investigated on 10-12 aquatic plant species and a naturally evolving epiphyte community under two growth conditions, and Species Sensitivity Distributions (SSD's) were constructed. The results revealed a relatively small variance in sensitivity between species, varying by a factor of 56 and 34 between the most and the least sensitive species for metsulfuron-methyl and terbuthylazine, respectively. This should be compared with the selectivity between terrestrial species, which can vary by a factor of up to 1000. The experiments also showed that the standard Lemna and algae tests represented the sensitivity of the aquatic plants and the epiphyte community well, despite the rather different growth conditions in the different test systems. If a safety factor of 10 was used between the maximal allowable environmental concentration and No Observable Effects Concentration (NOEC), defined as EC10, or a factor of 100 between the maximal allowable environmental concentration and EC50 on the Lemna test, more than 95% of the species would be protected. The toxicity of metsulfuron-methyl was independent of the growth conditions in these experiments, while the toxicity of terbuthylazine was enhanced as irradiance increased. As the highest herbicide concentrations in the environment are expected to occur in pulses associated with spray and/or precipitation events, pulse exposure with different herbicides was investigated on Lemna minor. The six herbicides selected represented three modes of action and had different Kow within the same mode of action. The experiments showed that a three-hour pulse, with concentrations that were lethal at chronic exposure, had no effect on Lemna growth measured over 4 days when the herbicides were photosynthetic inhibitors, diquat and terbuthylazine. For the ALS-inhibitors, imazamox and metsulfuron-methyl, and the inhibitors of microtubule assembly, propyzamid and pendimethalin, a three-hour pulse did affect growth. The EC50 of the pulse-exposed plants was approximately a factor of 10 higher than the EC50 of the plants experiencing chronic exposure. There was no detectable difference between herbicides with the same mode of action, but different Kow. The reason why a pulse exposure with photosynthetic inhibitors was so relatively inefficient in reducing plant growth compared to a pulse exposure with the other herbicides could be that damage caused by reactive oxygen species, which is the immediate effect of photosynthetic inhibiting herbicides, is a common phenomenon in all plants. Hence, an efficient repair system already exists to ameliorate this kind of damage. It seems to be harder to recover from direct or indirect inhibition of cell division, and for metsulfuron-methyl it was shown that it took about four days to recover from an almost lethal pulse exposure. The general conclusion of all experiments was that herbicides in the concentrations measured in Danish surface waters (< 10 μg l-1) would generelly neither reduce growth nor change the composition of submerged species significantly,- at least not if the effect of the herbicides was assessed for each herbicide individually. On the contrary, a stimulation of plant growth at low herbicide concentrations was observed in several experiments, a phenomenon called hormesis. Some fungicides have been reported to stimulate plant growth in the field, and we saw a similar effect on Lemna grown under sub-optimal light conditions when exposed to low doses of prochloraz (10 and 20 μg l-1). As soon as growth conditions were optimized, however, the growth stimulating effect disappeared. The presence of prochloraz (<100 μg l-1) did not affect the toxicity of either metsulfuron-methyl or terbuthylazine. Even though the presence of herbicides in the concentrations measured in Denmark is unlikely to have a significant effect on the aquatic flora if present alone, it is more uncertain what the effect of several simultaneously occurring herbicides would have. Here, we present the Additive Dose Model, also called Concentration Addition, which can be used to assess the effect of mixtures of herbicides, if comparable toxicity data for all the single substances are available. We show an example where several photosynthetic inhibitors have been measured in a pulse event associated with high precipitation. The example shows, that the cumulative effect of the three herbicides could potentially affect the growth of algae. We consider the Additive Dose Model to be a good tool to identify critical situations that would need further investigation. A mixture experiment with metsulfuron-methyl and terbuthylazine performed on Lemna and algae showed that the cumulative effect of the mixture was less than expected from the Additive Dose Model. This proved to be a common pattern and supports the use of the Additive Dose Model as conservative estimates of a worst-case situation. As one of the goals of the present project was to focus on low concentrations and determine No Effect Concentrations, we have discussed the definition of "no effect". We chose to use EC10 and in general we prefer the use of ECx values instead of the often used NOEC and Lowest Observable Effect Concentration (LOEC), as ECx values are more consistent across test-systems. Also, the variance on the ECx gives an impression of the experimental quality that is not reflected in either NOEC or LOEC. It is our opinion that the largest uncertainty about the effect of pesticides on the aquatic flora is to be found in the situations where pesticides occur in mixtures. More extensive knowledge of the composition, concentrations and time courses of mixtures and their effect on the flora could help to define places and situations where potentially harmful mixtures could occur. We also believe that the mechanisms of pesticide uptake by aquatic plants are not very well described as there are indications that these are very different from uptake mechanisms in the terrestrial environment, especially for aerial herbicides. A better understanding of the mechanisms of pesticide uptake could facilitate better predictions of the aquatic toxicity of pesticides and other chemical compounds. The general picture, however, is that the individual pesticide concentrations measured in Danish surface waters will not affect the growth and species composition of the aquatic flora significantly. 1. IndledningDanmark er et landbrugsland, hvor ca. 70% af jorden er kategoriseret som dyrket landbrugsareal (Windolf et al., 1996). På dette areal bliver der i gennemsnit sprøjtet med pesticider omkring 2 gange årligt og den samlede mængde af udspredt aktivstof kommer op på omkring 2800 ton (Miljøstyrelsen, 2003). Dertil kommer forbruget i gartnerier, frugtplantager, skovbruget, offentlige institutioner og af private. Det er derfor ikke overraskende, at man kan finde rester af pesticider i miljøet omkring de dyrkede arealer. Spørgsmålet er i hvor høj grad pesticiderne i miljøet påvirker organismer udenfor målgruppen. Da 75-80% af den udspredte mængde pesticider er herbicider, ukrudtsmidler, der virker specifikt på planters biokemi, vil man forvente, at især vilde planter udgør en følsom gruppe organismer udenfor målgruppen. I denne rapport fokuserer vi på planter, der vokser i søer og vandløb. 1.1 Vandplanter og algerPlantelivet i søer og vandløb består af flydende og mere eller mindre neddykkede vandplanter, større alger som kransnålalger samt af fastsiddende og frit flydende mikroalger, epifytter og fytoplankton. Mængden og fordelingen af større planter og mikroalger afhænger for en stor del af vandets næringssaltindhold, bund- og strømforhold. Udbredelsen af neddykkede arter i søer vil oftest være begrænset af næringssalt indholdet, da høje næringssaltkoncentrationer medfører en øget vækst af fytoplankton, der skygger for de rodfæstede planter, men også bundforholdene kan spille en rolle. I vandløbene spiller næringssalt-tilgængeligheden også en rolle, men især muligheden for rodfæste samt strømhastighed er af betydning for de rodfæstede planters udbredelse, og specielt har eutroficiering og den hyppige grødeskæring ændret voldsomt på sammensætningen af arter i danske vandløb gennem de sidste 100 år (Baattrup-Pedersen, 2000; Baattrup-Pedersen et al., 2003; Sand-Jensen et al., 2000). Vandplanter spiller en stor rolle i den fysiske stabilisering af bundforholdene i både søer og vandløb for sedimentationshastigheder og strømningsforhold og for optagelsen og recirkuleringen af næringssalte. Derudover fungerer de som skjul for insekter, krebsdyr og fisk og udgør et substrat for overflade-levende mikroorganismer, snegle og andre græssere (Baattrup-Pedersen, 2000; Sand-Jensen, 1997). En forringelse af vandplanternes vækst og overlevelse vil derfor påvirke hele det akvatiske økosystem kraftigt. 1.2 Pesticider i vandmiljøetPesticider kan ende i vandmiljøet ad forskellige veje: ved vinddrift, oversprøjtning ved kørsel for tæt på vandløbet, deposition fra luften ved fjerntransport i atmosfæren, udvaskning fra marker via drænvand eller grundvand, overfladisk afstrømning eller fra punktkilder som f.eks. vaskepladser eller affaldsdepoter (Løkke, 2000; Mogensen et al., 1999)lsesprodukter man har testet for i Fyns Amt, har man fundet 33 af stofferne i koncentrationer over detektionsgrænsen. Generelt ligger niveauet for pesticider i overfladevand på under 1 μg l-1. Ud af over 7000 målinger af omkring 60 pesticider og pesticid-nedbrydningsprodukter målt i perioden 2000-2002 på 8 målestationer gælder dette for 99.9% af prøverne. I de 9 prøver, der ligger over 1 μg l-1, er den højeste koncentration 15 g glyfosat l-1. I ca. 3% af målingerne er der målt koncentrationer på over 0,1 μg l-1, som er den fastsatte grænseværdi for drikkevand. (Data fra det Nationale program for Overvågning af Vandmiljøet (NOVA) fra 2000-2002 venligst stillet til rådighed af DMU, Afdeling for Ferskvandsøkologi, Jytte Erfurt. Herefter refereret som NOVA-data). I et vandløb har man i løbet af et døgn kunnet måle op til 18 forskellige pesticider (Løkke, 2000), og NOVA-dataene viser, at det ikke er usædvanligt at finde mellem 10 og 20 forskellige pesticider i koncentrationer over 0,01 μg l-1 på en lokalitet i løbet af en dag. Ikke overraskende finder man specielt pesticiderne i vandmiljøet i sprøjteperioden. De højeste koncentrationer forekommer ofte i pulse af timers varighed i forbindelse med nedbørshændelser (Styccen et al., 2003). Pulsenes størrelse og længde afhænger af, hvor langt nedstrøms fra afstrømshændelsen man befinder sig, da opblanding og fortynding gør pulsene længere og fladere. Forekomsten af pesticider i søer og stillestående vand er ikke så udførligt efterforsket, som forekomsten i vandløb. De data, der findes på mindre vandhuller, indikerer dog niveauer, der er sammenlignelige med dem, der findes i vandløbene (Løkke, 2000; NOVA-data). I større søer må man forvente, der sker en opblanding, der vil føre til en yderligere fortynding. 1.2.1 ProblemstillingerPesticiderne findes altså i danske søer og vandløb, og størstedelen af de fundne stoffer er herbicider (Løkke, 2000; NOVA-data). Herbiciderne formodes at kunne påvirke planterne og algerne i det akvatiske miljø. Formålet med det pågældende arbejde er derfor at undersøge, hvor vidt det er sandsynligt, at dette også sker. Dette har vi valgt at gøre ved at opstille en række problemstillinger, der hver især belyser dele eller aspekter af den samlede problemstilling: Påvirker pesticider i vandmiljøet væksten af akvatiske planter og alger? 1.2.1.1 Hvad er ingen effekt?Da vores studie skulle fokusere på lave koncentrationer og effektniveauer, har vi fundet det relevant at diskutere, hvad man mener med "ingen effekt". Lidt filosofisk kan man sige, at "Man kan altid vise, at der er en effekt, men man kan aldrig vise, at der ikke er en effekt". Der vil altid være ting man ikke har målt på, niveauer man ikke kan måle og tidsperspektiver, man ikke har taget i betragtning. Vi må derfor definere, hvad vi mener med "ingen effekt". Dette indebærer at vores "ingen-effekt-niveau" kan variere afhængigt af vores definition. Nogle af de mest brugte definitioner vil her blive vurderet. 1.2.1.2 Giftigheden af herbicider i vandVi valgte derefter at vurdere giftigheden af herbicider i vand i forhold til deres giftighed i det terrestriske miljø. I modsætning til det terrestriske miljø, hvor planterne kun bliver påført herbicidet en enkelt gang, kan planterne i vandmiljøet være omgivet af herbicider i en længere periode. Specielt hvis planterne vokser i langsomt flydende eller i stillestående vand. Til gengæld bliver stofferne meget fortyndede i det akvatiske miljø sammenlignet med de koncentrationer, der findes i sprøjtebeholderen og i de afsatte dråber på planterne, hvilket kunne nedsætte deres optagsrate dramatisk og gøre det muligt for planterne at afgifte stofferne, inden de når kritiske niveauer. 1.2.1.3 Artsspecifikke forskelle i følsomhedMange herbicider er selektive i det terrestriske miljø. Det er netop en af deres attraktive egenskaber, at de lader afgrødeplanten stå, mens ukrudtsplanterne dør. Spørgsmålet er, om de også er selektive overfor planter i vandmiljøet og derved kan forskubbe den naturlige balance mellem arterne? Hvis nogle arter er mindre følsomme overfor et hyppigt forekommende herbicid end andre, ville disse kunne overtage og komme til at dominere på steder, hvor de ellers ville være blevet holdt i skak af andre mere følsomme arter. 1.2.1.4 Giftighedens afhængighed af tid, effekt variable og naturlige stressfaktorerHvad betyder valget af forsøgstid og effektvariabel for de giftighedsparametre, man bestemmer og som danner grundlag for lovgivningen om brug, sprøjtegrænser etc. for pesticider? Nogler herbicider virker hurtigt, andre langsomt og de påvirker forskellige fysiologiske processer i planterne. Dette betyder, at den målte giftighed af et stof kan afhænge af forsøgstiden og af hvilken effekt, man måler. Også naturlige vækstfaktorer kan påvirke et stofs giftighed. Vi har i dette studie valgt at se på effekten af lys på virkningen af to herbicider med forskellig virkemåde. Lyskvantiteten og -kvaliteten ændres hurtigt ned igennem vandsøjlen, og da mange herbicider virker ved at påvirke fotosynteseapparatet, kunne det formodes, at lysintensiteten påvirker virkningen af disse herbicider. Også pH kan muligtvis påvirke effekten af herbicider, der er syrer eller baser og derfor skifter ladning, afhængigt af pH. Da pH i danske ferske vande kan variere fra pH 5 i sure tørvegrave til over pH 10 i vande med høj fotosyntese aktivitet (Sand-Jensen, 2001), kan denne problemstilling være højst relevant. 1.2.1.5 Effekter af pulseksponeringSom beskrevet optræder de højeste og derfor potentielt mest toksiske koncentrationer af herbicider i pulse. Hvad er effekten af en puls i forhold til en længerevarende eksponering, og hvor lang tid tager det for planterne at regenerere efter en væksthæmmende puls? Er nogle herbicider mere effektive i pulsform end andre, sammenlignet med deres effekt efter længere tids eksponering? Man kunne forestille sig, at herbicider med en evne til at bioakkumulere ville have en større relativ pulseffekt, da de ville blive i planterne i længere tid end mere vandopløselige stoffer. Planternes evne til at afgifte og reparere de skader, de er blevet påført af herbicider under en puls, kunne også være af betydning for effekterne af en pulseksponering. 1.2.1.6 KombinationseffekterSom tidligere nævnt kan man finde flere pesticider i vandmiljøet på en gang (Løkke, 2000; NOVA-data). Ud af de 140 NOVA-prøver, hvor der blev målt pesticider i koncentrationer over detektionsgrænsen, var der mere end 7 stoffer tilstede i over halvdelen af prøverne. Amerikanske undersøgelser af over 80 stoffer i overfladevand viste tilsvarende resultater (Martin et al., 2003). I Fyns Amt blev der målt op til 18 forskellige stoffer på et døgn (Løkke, 2000). Så hvad er effekten af tilstedeværelsen af flere stoffer på en gang? 1.2.1.7 Vækststimulering ved lave pesticid-koncentrationerDer ses ind imellem en vækststimulering hos planter ved lave pesticid-koncentrationer, ligesom nogle svampemidler er kendte for at påvirke væksten af terrestriske planter (Gerhard et al., 1998). Hvordan påvirker en eventuel vækststimulering toksisiteten af herbicider? Og er vækststimulering et fænomen, der har relevans under naturlige forhold? 2 Hvad er ingen effekt?"Man kan altid bevise, at der er en effekt, men man kan aldrig bevise, at der ikke er en effekt", som man lidt filosofiskt kan udtrykke det. Vi må derfor lave en definition af, hvad vi mener med "ingen effekt". Den første begrænsning ligger i valget af testorganisme, fysiske testforhold og tidsforløb af testen og ikke mindst hvilken responsvariabel (Vækst, overlevelse, reproduktion, bevægelses adfærd etc.), vi vælger at se på. Når disse rammer er sat, kommer vi til forsøgsdesign og statistisk behandling af vores data. Den mest brugte ramme for et forsøgsdesign er at teste et vist antal af ens forsøgsorganismer overfor en stigende koncentration af det formodede giftstof og så måle den valgte respons-variabel efter et givet tidsrum. Den højeste koncentration i et sådant design, der ikke udviser en signifikant forskel i responsvariablen i forhold til kontrol, benævnes ofte No Observable Effect Concentration (NOEC) og den laveste koncentration, der udviser en signifikant respons, kaldes Lowest Observable Effect Concentration (LOEC). Kender man lidt til statistik, kan man hurtigt se, at hvis man formindsker antallet af replika eller har en stor varians på sine målinger, vil NOEC og LOEC stige og det er tilmed vist, at NOEC kan variere meget afhængigt af, hvilken test man bruger for at teste for signifikans (Chapman & Caldwell, 1996). Et alternativ til termerne NOEC og LOEC er et effekt niveau. Man kan i et givet system sige, at man tolererer effekter på under f.eks 1, 10 eller 25%. Ved at bruge effektniveau antager man, at giftigheden af et stof på en organisme følger en bestemt model. De mest brugte er den logistiske og Weibull (Nyholm, 1990). Indbygget i langt de fleste modeller er også, at stoffet altid har en effekt. Undtagelsen er Kooijmans model (Kooijman, 1996). Man kan derfor aldrig tale om "ingen effekt", men er nødt til at bestemme sig for et effektniveau, som man vurderer er lavt nok til, at det ikke spiller nogen rolle sammenlignet med den variation organismerne oplever i deres naturlige miljø. Der kommer altså en biologisk vurdering af effektniveau ind, ligesom man i statistik må tage en beslutning om signifikansniveau. Valget af effektniveau varierer typisk fra 10% til 25%, hvor EC25 bl.a. bruges af miljømyndighederne i U.S.A. (US EPA) (Bruce & Versteg, 1992; Hoekstra &Van Ewijk, 1993; Chapman & Caldwell,1996), mens Miljøstyrelsen i Danmark i flere tilfælde bruger EC10 (Miljøstyrelsen, 1999). 2.1 NOEC og LOEC versus Effekt Niveau2.1.1 Metode og ResultaterFølgende eksempel viser afhængigheden af parametrene No Effect Concentration (NOEC) og Lowest Effect Concentration (LOEC) sammenholdt med EC10 bestemt med non-liniær regression. Udgangspunktet er en dosis-responskurve på den relative arealspecifikke vækstrate (RGR) af Lemna minor som funktion af koncentrationen af herbicidet mesotrion (Figur 1a). Kurven består af 8 doseringer med tre replika i hver og en Coefficient of Variation (CV) på kontrollen, der indeholder 12 replika på 5%. I kurven Figur 1b, er der taget udgangspunkt i gennemsnittet af vækstraterne fra figur 1a, og nye data er genereret fra en normalfordeling hvor variansen er øget fra en CV på kontrollen på 5% til en CV på 20% af kontrollen. En CV på 20% er stadig regnet for udmærket for kontroller i et testsystem, og er et krav man overvejer at indføre for tests i Tyskland (Fiebig et al., 2003). Data i figur 1c er genereret på tilsvarende måde, men antallet af doseringer er halveret fra 8 doser til 4, mens antallet af replika er fordoblet fra 3 til 6. Variansen på data svarer til en CV på kontrollen på 5%. I figur 1d er variancen øget til at udgøre 20% af CV på kontrollen, men er ellers tilsvarende Figur 1c. Alle data er beskrevet med en log-logistiske dosis-respons model, der angiver både EC50 og EC10 med 95% konfidensintevaller. NOEC og LOEC er beregnet ud fra t-tests på kontrol og de enkelte doser med en signifikansgrænse på p< 0,05. Figur 1. Dosis-responskurver genereret ud fra figur a, med varierende varians og antal af doser. Alle kurver er beskrevet med en logistisk dosis-respons model. 2.1.2 Vurdering af NOEC og LOEC overfor ECxSom det kan ses på figuren, varierer NOEC mellem udefinerbar (1c) og 3,13 μg l-1 (1b), svarende til EC-værdier fra "ikke-bestemmelige" til EC33, mens LOEC varierer fra 1,56 μg l-1 (1c) til 6,25 μg l-1 (1d), svarende til EC6 og EC58. Disse variationer er ikke store sammenlignet med f.eks. den faktor 9, der blev fundet mellem to NOEC's af to certificerede firmaer på den samme prøve med den samme test (Chapman & Caldwell, 1996). Alle NOEC og LOEC værdier er imidlertid faste værdier, hvor usikkerheden på data ikke kan ses. Beskriver man derimod data med en log-logistisk dosis-responskurve eller en anden tilsvarende model, får man meget ens kurver med en lille variation på en hvilken som helst effekt koncentration. Den største variation er på usikkerhederne på ECx parametrene, hvilket afspejler kvaliteten af forsøgs-design og data (Tabel 1). Vælger man f.eks EC10 som et udtryk for NOEC, varierer den fra 1,94±0,22 μg l-1 (18d) til 3,60±0,94 μg l-1 (18c), hviket er under en faktor 2 (Tabel 1). Tabel 1. Nogle parametre der beskriver dosis-responskurverne i figur 1. Værdierne er angivet som middel asymptotisk standard variation. 95% konfidensintervaller for EC10 er angivet i sidste kolonne.
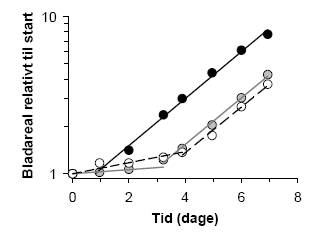
Værdimæssigt set, er der i det angivne eksempel ikke den store forskel på NOEC (udefinerbar – 3,13), LOEC (1,56 – 6,25) og EC10 værdierne, da de præsenterede data stadig er relativt pæne. Den største forskel ligger i regressionens mulighed for at udtrykke variationen på en værdi og dermed gøre en kvalitativ vurdering af en lav effektkoncentration mulig, hvilket især er vigtigt, hvor original-data ikke er tilgængelige. Vi mener derfor, sammen med en række andre (Bruce & Versteg, 1992; Hoekstra &Van Ewijk, 1993; Chapman & Caldwell,1996), at NOEC og LOEC værdier bør erstattes med ECx værdier, da de er meget mere konsistente og uafhængige af forsøgsdesign og giver et mål for datas kvalitet, hvis 95% konfidensintervaller eller lignende usikkerhedsparametre angives. Vi vil derfor i det følgende anvende EC10 som et udtryk for koncentrationer, der ikke vil have en signifikant effekt under naturlige forhold. 3 Giftigheden af herbicider i vandHvor giftige er herbicider i vandmiljøet i forhold til det terrestriske miljø? Som nævnt, kunne man forestille sig at de havde en større effekt i vand end på land, da eksponeringen kan vare i længere tid, og da hele den neddykkede del af planten er omgivet af herbicidet modsat situationen i det terrestriske miljø, hvor kun dele af planten rammes af sprøjtevæsken. Omvendt bliver herbicidet meget fortyndet, når det udsprøjtes på en vandoverflade. Man kunne derfor forestille sig, at optagshastigheden, som er diffusiv og dermed bestemt af bl.a. herbicid-koncentrationsforskellen mellem vand/dråbe og plantevæv, i vandmiljøet vil være så langsom, at planten kan nå at afgifte det optagne herbicid, inden det når at påføre nogen skade. 3.1 Materiale og MetoderVi testede den akvatiske toksisitet af 10 forskellige herbicider, der repræsenterer fire forskellige fysiologiske virkemåder, på flydebladsplanten Lemna minor, liden andemad, og grønalgen Pseudokirchneriella subcapitata, tidligere kendt som Selenastrum capricornutum. Vi testede både de tekniske produkter og de formulerede produkter, som bruges og usprøjtes af jordbruget. Dosis-responskurverne blev sammenlignet med de målte herbicid-koncentrationer i vandmiljøet. Derudover lavede vi en litteratursøgning i The e-Pesticide Manual (Tomlin, 2002) på de ca. 150 herbicider, hvor der fandtes akvatiske toksisitetsdata. 3.1.1 Andemads-testLemna minor stammer fra en dam på Frederiksberg og er blevet steriliseret med hypochlorit (Landolt & Kandeler, 1987). Testen tager udgangspunkt i OECD's guideline for Lemna-tests (OECD guideline for testing of chemicals, 2000). Den sterile kultur dyrkes i autoklaveret K-medie (Maeng & Khudairi, 1973), ved pH 5, i Ehrlenmeyer kolber lukket med en tot vat ved 24°C og en konstant lysintensitet på ca. 100 mol m-2 s-1 (PAR). Ved forsøgets start overføres en plante, frond, til 10 ml medie tilsat herbicid. Planterne fotograferes med et digitalt kamera og placeres i vækstskab under overnævnte vækstforhold (Figur 2). Efter 7 dage fotograferes planterne atter, og start og slut overfladeareal bestemmes ved digital billedbehandling i Photoshop 7.0. Plantenes relative vækstrate bestemmes ved: (ln Aslut-ln Astart)/T, hvor Astart er arealet ved forsøgets start, Aslut er arealet ved forsøgets afslutning og T er det tidsrum hvori planterne har vokset angivet i dage. I disse forsøg har vi anvendt 6 kontrolplanter og 6 til 8 doseringer i 3-6 replika. Planternes vækstrate som funktion af herbicid-koncentrationen beskrives med en log-logistisk dosis-respons model (Streibig et al., 1993):
Hvor U er plantens respons, i dette tilfælde vækstrate, d og c er den øvre og nedre grænse for plantens respons ved henholdsvis nul og en uendelig stor herbicid koncentration, EC50 er den herbicid koncentration, der skal til for at halvere responsen mellem den øvre og nedre grænse, og b er proportionel med hældningen i punktet EC50. 
Figur 2. a) Placering af bakker med Lemna minor i klimaskab. Hver bakke indeholder 6 uafhængige brønde med hver en plante. b) Fotografering af start-planter sammen med en standard cm2. 3.1.2 Alge-testAlgetesten er beskrevet af Arensberg et al., 1995 og Mayer et al., 1997 og er i overensstemmelse med ISO's standard test for alger (International Organization for standardization, 1989). Forsøget startes ved, at man tilsætter alger fra en stamkultur til 4 ml ISO Standard 8692 vækstmedie (pH 8) i en 20 ml vial tilsat herbicider. Der er i hvert forsøg anvendt 2 replika af 8 herbicid koncentrationer og 6 kontrol uden herbicid. Algerne tilsættes i en koncentration på ca. 10 000 alger ml-1. Koncentrationen på stamkulturen er bestemt spektrofotometriskt. Til måling af start pigmentindhold udtages der straks 400 l af algesuspensionen (Figur 3) til et reagensglas med skruelåg, hvortil der tilsættes 1,6 ml acetone mættet med MgSO4 (12 g l-1). Prøverne stilles mørkt. Vialene med alger og vækstmedie placeres på et rystebord belyst nedefra med en intensitet på ca. 80 μmol m-2 s-1 (PAR). Rystebordet indstilles til 300 omdrejninger min-1, hvilket sikrer god omrøring af algerne. Der udtages prøver på dag 1 og dag 2, som beskrevet, hvorefter forsøget afsluttes og slut pH måles. pH varierer aldrig mere end 0,5 pH enheder. På dag tre kan algernes pigment fluorescens (primært klorofyl a) måles på et fluorometer med en exitations-bølgelængde på 420 nm og en emmisionsbølgelængde på 670 nm. Algernes vækstrate udregnes ved at udføre en liniær regresion på ln til fluorescensen på dag 0, 1 og 2 og angives i enheden d-1. Kontrol vækstraterne ligger mellem 1,6 og 2,0 d-1. Dosis respons forløbet beskrives med en log-logistisk model (Streibig et al., 1993.)
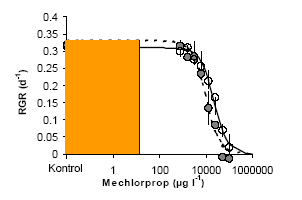
Figur 3. Der udtages prøver fra algesuspensionen ved start og de to følgende dage, så væksten i algernes biomasse kan følges. Algernes biomasse estimeres ud fra fluorescensen af pigmenterne, (primært klorophyl a) ekstraheret i acetone. Der er et lille hul i låget til vialen, der sikrer kulstofligevægt mellem luft og algemediet. 3.1.3 Litteratur-studieSamtlige herbicider i The e-Pesticide Manual med toksisitetsdata på alger blev udvalgt. Udover toksisiteten på alger blev stoffernes virkemåde (klassificeret efter The Herbicide Resistance Action Committee (HRAC) classification (2000) (Boutsalis, 2000)), molekylvægt, Kow, opløselighed, pKa, anbefalet minimum og maximum markdosis og optagelses sted (jord versus blad-herbicid) noteret. I de tilfælde, hvor Kow og opløselighed var opgivet ved forskellige pH, blev værdier ved pH 7 eller tæt ved valgt. I de tilfælde, hvor toksisitetsdata for flere forskellige arter var opgivet, blev data på Selenastrum anvendt og ellers Scenedesmus, da langt de fleste tests, hvor kun en art var opgivet, var udført på Selenastrum. Toksisiteten var normalt opgivet som EC50 og i enkelte tilfælde (<5 %) som NOEC (No Observable Effect Level). NOEC-værdier blev konverteret til EC50 ved at gange med en faktor 10. Dette er et konservativt estimat for EC50, da forholdet mellem mellem NOEC defineret som EC10 eller EC25 og EC50 varierer mellem 2 og 9 for dosis-responskurver med hældnings koefficienter på mellem 1 og 2 for den logistiske responskurve. Hældningskoefficienter i dette interval var dækkende for over 85% af 106 stoffer testet på alger (Faust & Scholze, 2004). Sammenhængen mellem herbicidernes toksisitet overfor alger og virkemåde blev testet med en variansanalyse (ANOVA) efterfulgt af en Tukey-test. Sammenhængen mellem toksisitet, Kow og opløselighed blev testet med en simpel regressionsmodel. SAS blev anvendt for samtlige analyser. For at kunne vurdere toksisiteten af herbicider i vand sammenlignet med toksisiteten på terrestriske planter, har vi indført begrebet: Herbicidets Relative Toksisitet. Dette er herbicidets akvatiske toksisitet relativt til stoffets terrestriske toksisitet angivet som den gennemsnitlige markdosis. Vi ved at markdosis i grove træk er bestemt ud fra et kriterie om at slå ca. 90% af målgruppe-ukrudtet ned (kudsk, 1993). Markdosis er derfor et groft udtryk for stoffernes terrestriske toksisitet overfor målgruppe-arter. For at kunne sammenligne en dosis i det terrestriske miljø med en koncentration i det akvatiske miljø, er man nødt til at omregne den terrestriske dosis per overfladeareal til en koncentration i vandmiljøet. Dette gøres ved at antage, at man sprøjter en fuld markdosis ud på 30 cm vand, som er den dybde, man i EU-sammenhæng har bestemt er standard for et vandløb eller en grøft. Denne koncentration er også hvad man i risikovurderings-sammenhæng kalder den værst tænkelige koncentration i miljøet (Predicted Environmental Concentration, PEC)(Miljøstyrelsen, 1999). Herbicidets Relative Toksisitet har vi defineret ved: EC50 akvatiske planter/ markdosis omregnet til koncentration (PEC). En relativ toksisitet på 1 betyder derfor, at en fuld markdosis opblandet i 30 cm vand vil kunne reducere væksten af test-alger med 50%. Er den relative toksisitet >1, vil en fuld markdosis reducere væksten med mindre end 50%, og er den <1, vil en fuld markdosis reducere væksten med mere end 50%. I reguleringssammenhæng kaldes EC50/PEC for Toxicity Exposure Ratio (TER) (Miljøstyrelsen, 1999). Sammenhængen mellem herbicidernes relative toksisitet og parametrene i databasen blev undersøgt parallelt med den faktiske toksisitet opgivet som EC50. Adgang til databasen kan fås ved henvendelse til Nina Cedergreen. 3.2 Resultater og Diskussion3.2.1 Laboratorie tests på 10 herbiciderIngen af de testede herbicider havde en virkning på væksten af hverken Lemna eller alger ved de herbicidkoncentrationer, som er blevet målt i Danmark (Løkke, 2000; NOVA-data). Heller ikke en afdriftshændelse på omkring 10% af markdosis, hvilket ikke er urealistiskt (Asman et al., 2003), vil kunne forårsage væksthæmninger på over 50% (Tabel 2). For 6 af de 10 herbicider vil end ikke en fuld markdosis på 30 cm vand kunne forårsage den vækstnedsættelse. Et typisk kurveforløb for mechlorprop, som er det pesticid, der er fundet i højeste koncentrationer, er vist i Figur 4, og alle resultaterne er opsummerede i Tabel 2.
Figur 4. Den relative vækstrate (RGR) som funktion af Mechlorprop-koncentrationen for Lemna minor. De åbne symboler er det tekniske stof og de lukkede, det formulerede produkt. Det grå område angiver det i Danmark målte koncentrationsinterval. En anden væsentlig observation er, at der stort set ikke er forskel på de tekniske og formulerede produkter for samtlige herbicider, undtaget glyfosat, hvor den anvendte formulering var Roundup (Monsanto). Det additiv, der bliver brugt i denne formulering, er i sig selv phytotoksiskt, hvilket har ført til udviklingen af mindre toksiske formuleringer som Roundup-bio (Cheminova, personlig kommunikation). De syntetiske auxiner (plantehormoner) påvirker ikke væksten af alger selv i koncentrationer, der nærmer sig dem der findes i sprøjtebeholderen. Dette skyldes sandsynligvis, at alger er encellede organismer, der ikke påvirkes af de hormoner, der virker på celledifferentieringen i højerestående planter. Tilsvarende resultater er blevet fundet i mesokosmosforsøg, hvor også makrofytter indgik (Brock et al., 2002). Der er stor forskel mellem resultaterne fra Lemna og algetesten med omkring en faktor 1000 større tolerance hos alger overfor metsulfuron, triasulfuron og mesotrion. Disse tre stoffer er, sammen med bentazon, svage syrer med en pKa på <5. Dette betyder, at der vil være omkring en faktor 1000 flere molekyler på negativ ladet form ved pH 8, som er det pH algetesten udføres ved, sammenlignet med pH 5, som er Lemnas vækst pH. Dette kunne indikere, at herbiciderne og andre produkter med tilsvarende egenskaber hovedsageligt tages op på den uladede form, hvilket også er vist for alger (Fahl et al.; 1995). For den svage syre bentazon var forskellen i toksisitet mellem de to testsystemer kun en faktor 10. Vi havde forventet en større forskel, hvis toksisiteten udelukkende afhang af pH-afhængige optagsrater, men planternes følsomhed spiller selvfølgelig også en rolle. Tabel 2. EC50 for Lemna minor og algen Pseudokirchneriella subcapitata for en række tekniske og formulerede (Form.) herbicider tilhørende fem overordnede virkemåder (Syntetiske auxiner: O, Aminosyresyntese hæmmende stoffer: B og G, Fotosyntesehæmmere: C og D, hæmmere af carotenoid dannelsen: F samt Inhibitorer af celledeling: K (Boutsalis, 2000)). EC50 værdierne er sammenlignede med de fundne koncentrationer af herbicider (Løkke, 2000; NOVA-data)og med den koncentration, som en afdriftshændelse på 10% af en gennemsnitlig markdosis vil give på 30 cm vand. Alle koncentrationer er angivet i μg l-1, og de tilhørende usikkerheder er standardfejlen på kurve-fittet. Fungicidet prochloraz er medtaget, da det indgår i senere undersøgelser.
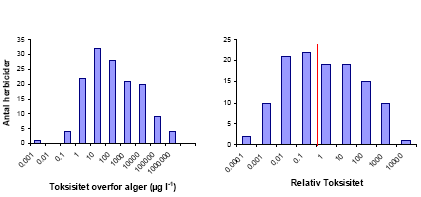
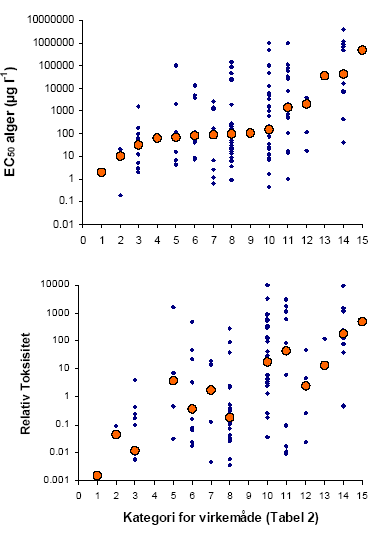
a Diquat fåes kun som Diquat-bromid, som er den opløsning der anvendes, og derfor her betegnes som det formulerede produkt. b Emulsionsformuleringen Stomp-pentagon 3.2.2 Database-analyseDer var 146 herbicider der opfyldte de stillede krav om toksisitetsdata på alger. For 55 af disse var testene udført på Selenastrum sp., for 16 var Scenedesmus sp. anvendt, 6 opgav Chlorella sp., 2 Anabaena sp. og 1 Navicula sp. For de resterende tests er arten ikke angivet ligesom usikkerhederne på de bestemte EC50 værdier aldrig angives. Dette giver, sammen med varierende testbetingelser, hvoraf vi har set at bl.a. pH kan spille en stor rolle, større usikkerhed på toksisitetsdataene sammenlignet med laboratorieforsøgene. Den store mængde data gør det dog muligt at kunne udrede nogle generelle tendenser. 3.2.2.1 Fordelingen af EC50 og Relativ ToksisitetSer man på fordelingen af EC50 for alger og deres Relative Toksisitet på en logaritmisk skala, får man i begge tilfælde en klokkeformet kurve (Figur 5).
For giftighed, angivet som EC50, kan man se, at <20% af det totale antal herbicider har en EC50 værdi på <10 μg l-1, som er omkring det højeste niveau man har målt i Danmark. Af disse 27 herbicider er kun fire kendt i Danmark og éet tilladt (www.lr.dk/middeldatabasen/Middelvalg.asp d. 21/10 2003). De fire er: 1) Pyraflufen-ethyl (EC50: 0,65 μg l-1), som er under afprøvning men endnu ikke tilladt til afgrøder, 2) Dichlorbenil (EC50: 2 g 1–1), som nu er udgået, men kendt for nedbrydningsproduktet BAM, 3) Oxadiargyl (EC50: 1,2 μg l-1), der som Raft 800 WG har været afprøvet i Landsforsøg, men som der ikke er blevet søgt godkendelse til og 4) Aclonifen (EC50: 6,9 μg l-1), der sælges under navnet Fenix. Aclonifen er, så vidt vi er orienterede, ikke målt i dansk overfladevand. Vender man sig mod den Relative Toksisitet (RT), har over halvdelen af herbiciderne en Relativ Toksisitet på >1. Det vil sige, at en fuld markdosis på 30 cm vand ikke vil kunne reducere algernes vækst med mere end 50%, hvis algerne i det naturlige miljø reagerer som algerne i alge-testen. Betragter vi en mere realistisk eksponering som f.eks. en 10% afdriftshændelse, vil 27% af de 146 herbider potentielt kunne reducere test-algevæksten med mere end 50%. Ud af disse, er 7 af stofferne kendt i Danmark og 3 af dem forhandles. Disse er: 1) Terbuthylazin (RT: 0,03), der indgår i 6 markedsførte produkter, 2)Diquat bromide (RT: 0,09), der sælges som Reglone og 3) Prosulfocarb (RT: 0,09), der sælges som Boxer EC. De fire udgåede eller endnu ikke markedsførte stoffer er Propachlor, Isoproturon, Dimethachlor og Oxadiargyl. Af disse 7 stoffer er både Terbuthylazin og Isoproturon jævnligt fundet i overfladevand (Løkke 2000, NOVA-data). Den generelt lavere toksisitet af herbicider i vand sammenlignet med det terrestriske miljø kan skyldes flere ting. Vores hypotese er, at den store fortynding af herbiciderne nedsætter optagelseshastigheden af herbiciderne til et niveau, hvor planten i højere grad kan nå at afgifte det optagne stof end det er tilfældet i det terrestriske miljø. Dette begrunder vi med udgangspunkt i Ficks lov, der beskriver principperne for diffusion, som er den måde de fleste herbicider optages i planter. Ficks lov siger: F = D (C0-C1)/L, hvor F er den arealspecifikke flux, D er diffusions koefficienten, L diffusionslængden og C0 – C1 er koncentrationsforskellen mellem herbicidopløsningen og plantevævet. En toksisk koncentration af herbicid i plantevævet vil derfor afhænge dels af herbicideksponeret overflade/volumenforhold, som er 2-5 gange større for akvatiske planter sammenlignet med terrestriske, hvis hele planten er eksponeret, og endnu større hvis der er tale om alger eller kun en delvis eksponering af den terrestriske plante. Diffusionskoefficienten er molekylafhængig og derfor ens i de to miljøer, så længe diffusionen foregår i et vandigt medie. Diffusionslængden gennem cuticula, cellevæg og –membran er kortere for vandplanter og alger med en skønnet faktor 2-20 (Frost-Christensen et al., 2003; Kirkwood, 2003). Både overflade volumenforholdet og den lavere diffusionslængde hos vandplanter peger altså på et øget optag hos vandplanter fremfor landplanter. Men sammenlignes koncentrationen af herbicidopløsningen i sprøjtebeholderen (C0) med den koncentration en fuld markdosis på 30 cm vand og en sprøjtevæskemængde på 150 l ha-1 giver, viser det sig, at der finder en fortynding på en faktor 20 000 sted. D.v.s. at koncentrationsgradienten (C0 – C1) er 20 000 gange lavere i det akvatiske miljø end i det terrestriske for herbicider, der sprøjtes direkte på planterne. For jordherbicider vil forskellen være noget lavere, da sprøjtevæsken vil blive delvist fortyndet af jordvand og herbicider, der imobiliseres ved adsorbtion til jordpartikler. Vi mener derfor, at hovedårsagen til den lavere toksisitet af herbicider i vand, sammenlignet med den terrestriske toksisitet, ligger i den kraftige fortynding og dermed optagshastigheden af stofferne. Udover at vurdere den akvatiske toksisitet af et herbicid sammenlignet med den terrestriske toksisitet, fortæller den Relative Toksisitet også noget om sandsynligheden for, at et bestemt herbicid vil udgøre en fare for vandmiljøet. Det er den tolkning af termen Relativ Toksisitet man bruger, når man vurderer TER-værdier. Men en fuld markdosis på 30 cm vand er ikke kun et udtryk for den værst tænkelige koncentration i miljøet, det er også den parameter, der bedst forklarer den herbicidkoncentration (og frekvens), man reelt finder i vandmiljøet (Sørensen et al., 2003). Den fundne sammenhæng kan bl.a. forklares med afdriftshændelser og punktkildeforurening fra vaskepladser, hvor man må forvente en korrelation mellem koncentrationen af herbicid i sprøjtebeholderen og udledningen til vandmiljøet. De mere komplicerede interaktioner, der finder sted så snart herbiciderne har været i kontakt med jord, hvortil nogle herbicider binder sig kraftigt, synes at være af mindre betydning for det overordnede billede. Den Relative Toksisitet - TER er altså et meget anvendeligt udtryk for den potentielle toksisitet af et stof i vandmiljøet, da den kombinerer toksisitet og sandsynlighed for forekomst og koncentrationsstørrelse. Når man som i dette litteraturstudie kun har EC50-værdier fra en enkelt art per herbicid, vil man, når man skal vurdere risikoen for alger og planter i miljøet bruge en sikkerhedsfaktor på mindst 100 (Miljøstyrelsen, 1999), for at tage højde for de forskelle, der måtte være mellem arter og mellem laboratorie og virkelige forhold. Vores laboratorieforsøg viste, at forskellen i toksisitet mellem arter (L. minor og P. supcapitata) i sig selv kunne variere med op til en faktor 1000. En faktor 100 mellem EC50 på alger og PEC vil altså for visse herbicider være for lav til også at kunne beskytte højere planter. Vender man tilbage til figur 5 og vurderer den ud fra en risikovurderings-synsvinkel vil man se, at langt størstedelen af herbiciderne har en Relativ Toksisitet på <100 nemlig ialt 78%, og 90% har en Relativ Toksisitet på <1000. Langt de fleste herbicider vil derfor blive klassificeret som potentielt toksiske for den akvatiske flora. Dette indebærer, at man vil foretage yderligere undersøgelser, hvor man tager hensyn til nedbrydning og adsorption af herbicidet i miljøet og undersøger eksponeringen ved introduktion af sprøjtebræmmer og evt. introducerer mikrokosmos- og felt-forsøg (Miljøstyrelsen, 1999). 3.2.2.2 Sammenhænge mellem akvatisk toksisitet og fysisk/kemiske og fysiologiske egenskaberVi sammenlignede den akvatiske toksisitet, både som algernes EC50 og som Relativ Toksisitet, med herbicidernes Kow og vandopløselighed. Den eneste signifikante sammenhæng var mellem logaritmen til den Relative Toksisitet og logaritmen til Kow (Regression: R2 = 0,07; p = 0,003, n =120, hældning = –0,21) og mellem gennemsnitlig markdosis og Kow (Regression: R2 = 0,09; p = 0,001, n =120, hældning = 0,11). Begge korrelationer forklarer dog så lidt af den samlede variation, at deres kemiske og biologiske signifikans er begrænset. Man finder ofte en positiv sammenhæng mellem stoffers narkotiske toksisitet og Kow (skønt knap så overbevisende for alger som for dyr), da stoffer med en høj fedtopløselighed let opløses i organismers cellemembraner, og ad den vej forårsager toksiske effekter (ECETOC,1998). Denne sammenhæng var ikke fundet i dette studie med herbicider, som er stoffer, der er kendetegnet ved at virke specifikt på forskellige fysiologiske processer i planter. Dette understreger deres specifikke virkning, der er uafhængig af deres fedtopløselighed. Ser man på herbicidernes fysiologiske virkning og opdeler dem efter deres virkemåde (Boutsalis, 2000), får man følgende billede (Figur 6). Virkemåderne er angivet med et nummer og forklarede i Tabel 3. Tabel 3. Virkemåde og den kategori de forskellige virkemåder får ifølge "The Herbicide Resistance Action Comittee (HRAC)" og det nummer, de er givet i Figur 6.
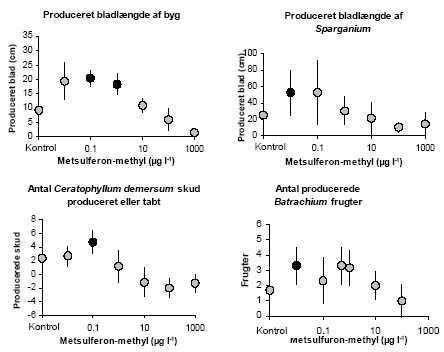
Figur 6. Spredningen i toksisitet angivet som EC50 (n = 135) og Relativ Toksisitet (n = 120) overfor alger mellem herbicider med forskellig virkemåde. Virkemåden er angivet med et tal som er forklaret i Tabel 3. Alle herbicider er angivet med små symboler og meridianen for hver virkemåde er angivet med et stort symbol. Det er markant at glyfosat (nummer 15) og de syntetiske auxiner (nummer 14) generelt er meget uvirksomme overfor alger. For de syntetiske auxiner gælder det, at algerne ikke har de hormoner, der påvirker celledifferentieringen, og for glyfosat er det vores hypostese, at dette herbicid simpelthen er svært at optage. Glyfosat er en zwitter-ion, hvilket vil sige, at den ved neutral pH har både en positiv og en negativ ladning (Knuuttila & Knuuttila, 1979). Det eneste andet herbicid med to pKa værdier under pH 7 er Imazethapyr, der ligeledes har en meget høj EC50 for Selenastrum på 500 000 μg l-1. Der er signifikant forskel i EC50 mellem de syntetiske auxiner og celledelings hæmmerne, mikrotubuli samlings hæmmerne, PS II hæmmerne og ALS hæmmerne (Tukey HSD). For den Relative Toksisitet er der ingen signifikant forskel mellem nogen af grupperne. 4 Artspecifikke forskelle i følsomhedI det forrige afsnit blev det vist, at det er de færreste herbicider, der er meget toksiske overfor alger i koncentrationer, der måles i overfladevand i Danmark. Ud over alger er Lemna sp., andemad, den eneste akvatiske plante som herbicider testes overfor på en rutinemæsig basis, og som der findes standard OECD-guidelines for (OECD guideline for testing of chemicals, 2000). Ud af de 486 herbicider, der er med i The e-Pesticide Manual fra 2002-2003 (Tomlin, 2002), findes der toksisitetsdata for alger for 156 produkter og for andemad på 36 produkter. Disse data er, som laboratorieforsøgene beskrevet i afsnit 3 også viste, langtfra ens. Vi ved, at mange herbicider virker selektivt, d.v.s. at nogle, helst ukrudtsplanter, er meget følsomme overfor et herbicid, mens afgrøden er tolerant og derfor overlever en sprøjtning, der slår ukrudtet ned. Forskelle i følsomhed mellem arter kan dels skyldes forskellig eksponering: nyspiret tokimbladet ukrudt får en større del af planten eksponeret, inklusiv meristemet, end en nyspiret græsafgrøde. Men den vigtigste årsag til artsspecifik forskel i følsomhed er metabolsk. Nogle planter kan simpelthen deaktivere herbicider hurtigere end andre og kan derfor tolerere en større dosis (Devine et al., 1993). Der kan være op til en faktor 1000 forskel i følsomhed mellem følsomme og tolerante terrestriske planter (Boutin et al., 2000). Spørgsmålet er, om noget tilsvarende gælder for vandplanter, der har en utrolig varieret morfologi og levemåde og stammer fra mange forskellige ikke nært beslægtede familier. Kan vi basere lovgivningen, der skal beskytte den akvatiske flora, på laboratorieresultater fra alger og en lille flydebladsplante? For at undersøge denne problemstillig valgte vi at se på artsspecifik følsomhed hos 10-12 forskellige danske vandplanter i forhold til to herbicider: Det ene er metsulfuron-methyl, et sulfornylureaherbicid som godt nok ikke, så vidt vi er orienteret, er målt i overfladevand, men som har et nedsivningspotentiale. Metsulfuron-methyl er også et af de herbicider, hvor der er vist en stor artspecifik forskel i følsomhed hos terrestriske planter (Boutin et al., 2000). Det andet herbicid er terbuthylazin, som var et af de herbicider, der havde en lav Relativ Toksisitet (afsnit 2.2.2.1), og som er målt i høje koncentrationer i overfladevandet i Danmark. Vi forventede at selektiviteten hos akvatiske planter afhang af de samme to primære faktorer som hos terrestriske planter: eksponering samt evnen til at deaktivere herbiciderne. Eksponeringen hos en akvatisk plante vil være proportionel med den herbicid-eksponerede overflade i forhold til plantevolumen. Man kunne derfor forvente af arter med et stort overflade/volumenforhold ville være mere følsomme end arter med en mindre overflade. Evnen til at deaktivere herbicider blev ikke målt i dette studie. 4.1 Materiale og MetoderDe forskellige vandplantearter blev testet i en opstilling med flere arter og med et kontinuert flow af medietilsat herbicid. Disse resultater blev sammenlignet med resultater fra standardtest beskrevet i afsnit 2.1.1 og 2.1.2. 4.1.1 Forsøg med flere arterNeddykkede vegetative skud fra Elodea canadensis (Almindelig Vandpest), Callitriche platycarpa (Fladfrugtet Vandstjerne), Potamogeton crispus, (Kruset Vandaks), Ceratophyllum demersum (Tornfrøet Hornblad), Ceratophyllum submersum (Tornløs Hornblad), Myriophyllum spicatum (Aks-Tusindblad), Batrachium trichophyllum (Hårfliget Vandranunkel), Berula erecta (Smalbladet Mærke/Sideskærm), Sparganium emersum (Grenet Pindsvineknop), og Lemna trisulca (Kors-andemad) samt en flydebladsplante Spirodela polyrrhiza (Stor Andemad) blev indsamlet i vandløb og damme på Nord-Sjælland. Planterne blev renset og placeret i akvarier i et vækstkammer med en dag/nat vandtemperatur på 18°C/15°C og 16 timers lys. Vækstmediet var et Smart & Barko medie (Smart & Barko, 1985) tilsat en mikronæringsblanding bestående af: 3 mmol m-3 H3BO3, 0.08 mmol m-3 ZnSO4, 0.06 mmol m-3 NaMoO4, 0.03 mmol m-3 CuSO4, 3 mmol m-3 MnCl2, 2 mmol m-3 FeCl2 and 2 mmol m-3 EDTA samt tilsat kvælstof og fosfor (Tabel 4). I det første forsøg blev kvælstof- og fosforkoncentrationerne bestemt ud fra de gennemsnitlige koncentrationer fra 218 lokaliteter i danske vandløb (www.dmu.dk1_Viden/2_Miljoe-tilstand/3_vand/4_vandkemi/start.asp, April 2001). Da vi imidlertid havde mistanke om fosformangel, justerede vi koncentrationerne i de efterfølgende forsøg. Start pH var pH 7.7 og pH steg aldrig til over pH 8.5 i løbet af dagen. Tabel 4. Lys, kvælstof og forforkoncentrationer i de 4 forsøg med flere arter.
I tillæg til de indsamlede planter anvendte vi også den Lemna minor klon, vi bruger til standard Lemna-testene. Efter 5 dages tilvænning til forholdene i vækstkammeret blev planterne placeret i tests-systemet (Figur 7a,b,c). Test-systemet bestod af 7 stk 25 liters glas-flasker indeholdende vækstmedium tilsat herbicid i koncentrationerne 0,00; 0,01; 0,10; 0,50; 1,0; 10 og 100 g metsulfuron-methyl l-1 for forsøg I, 0,00; 0,01; 0,10; 1,0; 10; 100 og 1000 μg metsulfuron-methyl l-1 for forsøg II og 0; 8 ; 23; 64; 184; 525 og 1500 μg terbuthylazin l-1 for forsøg III og IV. Flaskerne var forbundet til hver 2 akvarier (L:B:H = 45:30:30 cm) med en 1 mm (indre diameter) Teflon slange. Mediet blev tilført akvarierne via teflon-slangen v.h.a. hævert princippet. Ialt blev der tilført 12.5 liter medie til hvert akvarie hver- (Forsøg I og II) eller hver anden dag (Forsøg III og IV) (Figur 7a). Hvert akvarie indeholdt 25 liter og var forsynet med overløb. Tre afvejede 5 cm apicale skud af langskudsarterne Elodea, Callitriche, Batrachium, Myriophyllum og de to Ceratophyllum arter samt tre grundskud af grundskudsplanterne Berula og Sparganium og Lemna trisulca på 5-bladsstadiet blev placeret i et net i hvert af de ialt 14 akvarier (Figur 7b). Tre planter af de to flydebladsplanter Lemna minor og Spirodela blev placeret i hver deres flydende ramme tæt på akvariets kant (Figur 7c). På grund af de forskellige tider på sommeren eksperimenterne blev udført (2001 og 2002), var det ikke altid muligt at skaffe alle de nævnte arter, men et minimum af 10 arter indgik i alle forsøg. Ud over planterne blev der placeret 3 bånd 11 mm bredt grønt Max tape (Maxco, Ltd, Tyskland) vertikalt i akvarierne som kunstigt substrat for fastsiddende alger. Der blev udtaget 5 skud/planter identiske med de inkuberede af hver art til bestemmelse af friskvægt/tørvægtsforholdet ved forsøgets start. For flydebladsplanterne blev overflade/tørvægt forholdet målt ud fra overflademålinger som beskrevet i afsnit 2.1.1.
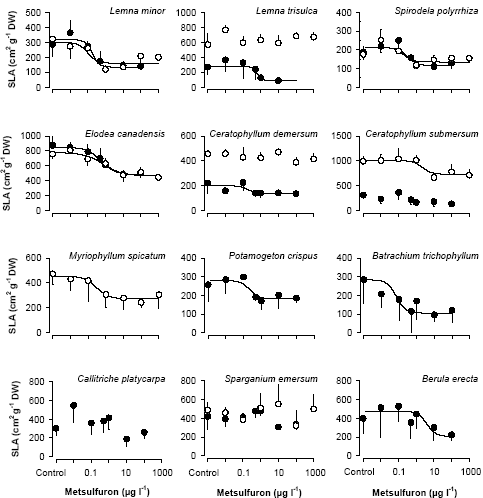
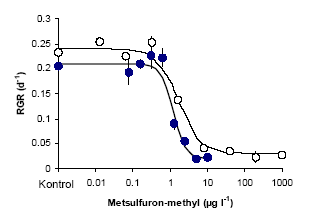
Figur 7. Opsætning af vandplanter i Mikro-kosmosforsøg med vandgennemstrømning: a) Selve forsøgsopsætningen, b) placering af de neddykkede vandplanter I net og c) placering af flydebladsplanterne. Efter en vækstperiode på 14 dage blev alle planter høstet, frysetørret og vejet, og flydebladsplanterne fotograferet. Planternes relative vækstrate blev udregnet som: (ln Wslut-ln Wstart)/T, hvor Wstart er tørvægten ved forsøgets start, Wslut er tørvægten ved forsøgets afslutning, og T er det tidsrum, hvori planterne har vokset angivet i dage. Båndene med epifytbelægning blev placeret i et reagensglas med 96% ethanol og ekstraherede et døgn i mørke inden klorofyl koncentrationen blev målt spektrofotometriskt ved 665nm. Klorofyl a koncentrationen blev udregnet efter: ((Abs665 – Abs750)E)/AL, hvor Abs665 og Abs750 er absorbtionen ved henholdsvis 665 og 750 nm, E er ekstraktions volumen i ml, A absorbtionskoefficienten af klorofyl a i ethanol (83,4 l g-1 cm-1) og L arealet af båndets overflade (Wintemans & De Mots,1965). Ud over vækstraten, blev planternes specifikke overfladeareal (SLA) af frysetørret materiale også målt. Dette blev gjort på de første fuldt udviklede blade på hvert skud. Bladene blev vejet og placeret i vand, så de svulmede op til friskvægts-størrelse. De blev derefter placeret på et gennemsigtigt stykke plastik (et overhead-ark) og skannet sammen med et standard areal, hvorefter arealet blev beregnet i Photoshop 7.0. Det herbicid-eksponerede overfladeareal blev udregnet som: et-sidet overfladeareal for flydebladsplanter, to-sidet overfladeareal for neddykkede planter og for planter med nålelignende blade som f.eks de to Ceratophyllum arter som ½(målt areal * π). 4.1.2 Kemiske analyserDer blev udtaget vandprøver fra det nyblandede medie i flaskerne samt akvarierne 3-4 gange i løbet af hver forsøgsperiode til herbicidanalyse. Analyserne blev udført på Dansk Jordbrugs Forskning, Flakkebjerg. Analyserne af metsulfuron-methyl blev foretaget efter metoden beskrevet i (Køppen & Spliid, 1998). Terbuthylazin var analyseret med HPLC (Hewlett Packard HPLC 1100; Thermo Hypersil BDS column 250 mm x 2.1 mm - 5m; Applied Biosystems Sciex API 2000 MSMS detector) ved at benytte en gradient med A-eluenten bestående af 99% 10 mM ammonium acetat i vand og 1% methanol og B-eluenten bestående af 90% methanol og 10% mM ammonium acetat i vand. Retentionstiden for terbuthylazin var omkring 10 minutter. Detektionen blev udført efter elektro-spray ionisering i positivt mode med MRM, en Q1 m/z på 230 og Q3 m/z på 174. "Declustering potential" var 60 V, kollisionsenergien 16 V og den kollisionsaktiverede dissociering (CAD) 4. Detektionsgrænsen var omkring 0.02 terbuthylazin μg l-1. 4.1.3 Statistiske analyserAlle data blev først testet med en variansanalyse, for at se om der var forskel mellem herbicid-doseringerne. Hvis der var en signifikant forskel, blev data beskrevet med den logistiske dosis-respons model, som beskrevet i afsnit 2.1.1. Variansen var stabiliseret ved at bruge "Transform-both-sides" (Streibig et al., 1993) og modellens validitet sammenlignet med variansanalysen var testet med en lack-of-fit F-test (Seefeldt et al., 1995). I de tilfælde hvor hældnings parameteren ikke kunne bestemmes med en varians <b, blev b fikseret til gennemsnittet af de resterende kurver som lå rimelig konstant omkring 1,5 – 2,5 (Cedergreen et al., 2003). EC10, som er det niveau, vi her definerer som et økologiskt relevant "ikke-effekt-niveau", blev beregnet ud fra dosis-respons kurvernes parametre. For at vurdere fordelingen af arternes følsomhed overfor de to herbicider, kontruerede vi art-følsomheds-fordelinger (Species Sensitivity Distributions, SSD's). Fordelingerne blev lavet ved at sortere ECx værdierne for hvert forsøg efter størrelse (Rang) og estimere deres centiler som (100 x rang)/(n+1), hvor n er antal arter inkluderet i fordelingen (Klaine et al., 1996). Arts-følsomheds-fordelingen blev beskrevet med en logistisk model analogt til dosis-respons kurverne. Sparganium emersum var inkluderet i samtlige fordelinger, men dens ECx var ikke inkluderet i fitting proceduren, da den i alle tilfælde lå udenfor det testede koncentrationsinterval. 4.2 Resultater og diskussionFørst beskrives og diskuteres resultaterne for metsulfuron eksperimenterne og dernæst for terbuthylazin eksperimenterne. Til sidst sammenholdes arts-følsomheds-fordelingerne med de ECx niveauer der er opgivet i litteraturen, og som vi har målt, for at vurdere om standardtestene på Lemna og alger er repræsentative for et større antal akvatiske arter. 4.2.1 Akvatiske planter og algers følsomhed overfor metsulfuron-methylDet viste sig at den tørvægtsspecifikke vækst af akvatiske planter over 14 dage ikke var nogen særlig følsom parameter til at måle effekten af metsulfuron-methyl. Kun 4 af de i alt 19 kurver kunne fittes med en dosis-responsmodel (Tabel 5). Tabel 5. Dosis-responskurvens parameter (se afsnit 2.1.1) for tørvægtsspecifik relativ vækstrate i forsøg med metsulfuron-methyl (middel asymptotisk stdev)
* b var fikseret til gennemsnittet af de kendte b-værdier. Dette skyldes, at metsulfuron-methyl påvirker aminosyre-syntesen, og da planterne langt hen ad vejen kan kompensere for denne virkning ved at sætte omsætningsraten af proteiner i vejret, går der forholdsvis lang tid, inden forstyrrelserne i aminosyresyntesen påvirker fotosyntesen og derved kulstoffikseringen og tørvægtstilvæksten. Celledelingen bliver, af uvis årsag, dog ramt allerede efter få timer (Cobb, 1992). Når celledelingen stopper, mens tørvægtstilvæksten stiger, bliver bladene tykkere. Dette sker sandsynligvis p.g.a. stivelses akkumulering, som det er set for andre planter udsat for ALS-hæmmere (Rouuela et al., 2000). Hvis vi derfor ser på data for specifikt bladareal (SLA), som er et indirekte mål for bladtykkelse, får vi fine dosis-responskurver for 13 ud af de 19 kurver (Tabel 6 og Figur 8). Sammenligner vi EC50 værdierne for den tørvægtsspecifikke vækstrate med EC50 værdierne for SLA, finder vi, at SLA er en ca. 10 gange mere følsom parameter end tørvægtsspecifik vækst. Det er dog muligt, at denne forskel vil udligne sig, hvis forsøget fortsatte i længere tid. For Lemna-forsøg, hvor udviklingen i EC50 er fulgt over 15 dage for en række forskellige responsvariable, stabiliserer EC50 for SLA sig dog på et niveau, der en faktor 3 lavere end EC50 for tørvægtsspecifik vækst (Ikke viste resultater). Tabel 6. Dosis-responskurvens parameter (se afsnit 2.1.1) for specifikt bladareal i forsøg med metsulfuron-methyl (middel asymptotisk stdev)
a b var fikseret til gennemsnittet af de kendte b-værdier.
Figur 8. Specifikt bladareal (SLA) for det første fuldt udviklede blad for 12 arter vandplanter som en funktion af metsulfuron-methyl koncentrationen i to forsøg: Forsøg 1, lukkede symboler, Forsøg 2, åbne symboler. Data er middel ±asymptotisk standard afvigelse (n = 5 eller 6). Udviklingen af epifytbelægningen var uafhængig af metsulfuron-methyl koncentrationen (ANOVA: p > 0,05)(Cedergreen et al., 2003). Dette er i overensstemmelse med resultaterne fra algetesten udført i vores laboratorie ved pH 8 (Tabel 2), hvor EC50 værdien lå omkring 700-800 μg l-1 og med litteraturværdien i Tomlin (2002), der angiver en NOEC på 100 000 g l-1. Også andre danske undersøgelser af følsomheden af naturlige epifyt-samfund overfor metsulfuron-methyl viste lignende resultater (Møhlenberg et al., 2004). 4.2.1.1 Kemiske analyserAnalyser af metsulfuron-methyl koncentrationen i vandet viste ingen signifikant forskel mellem det til-ledte vand og vandet i akvarierne (ANOVA: p = 0,94). De målte koncentrationer var en smule lavere (<10%), men passede ellers fint overens med de beregnede (Regression: r2 = 0,96, slope = 0,94 på log-transformerede data). 4.2.2 Akvatiske planters og algers følsomhed overfor terbuthylazinDen tørvægtsspecifikke vækstrate var en meget følsom parameter i forsøgene med terbuthylazin. I disse forsøg opstod imidlertid et andet problem: Efter inkuberingen af planterne, tabte en del planter blade, hvorefter nye skud og blade begyndte at vokse ud. Dette kunne skyldes stress i forbindelse med "dubning" inden bestemmelse af skuddenes friskvægt, eller noget helt andet. Det havde imidlertid den konsekvens, at de tørvægtsspecifikke vækstrater, som jo er et udtryk for netto-tilvækst hvor tabsraten er inkluderet, blev negativ i tilfælde, hvor der var en klar tilvækst af nye skud og blade. Den negative vækst kan gøre det svært at bestemme den nedre grænse for en dosis-responskurve, og det blev derfor besluttet at anvende slut-tørvægt som responsvariabel for terbuthylazinforsøgene (Tabel 7, Figur 9). Variansen er lidt større end for vækstrateresultaterne, men EC50 værdierne var ikke væsentligt forskellige mellem de to responsvariable. Tabel 7. Dosis-responskurvens parameter for slut-tørvægt i forsøg med terbuthylazin (middel ±asymptotisk stdev)
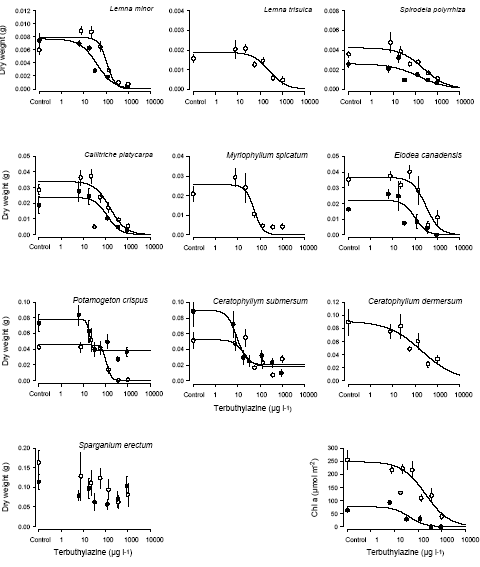
a b var fikseret til gennemsnittet af de kendte b-værdier.
Terbuthylazin havde en klar effekt på udviklingen af epifytbelægningen med EC50 værdier på henholdsvis 42±15 μg l-1 og 206±74 μg l-1 i forsøgene med lav- og høj lysintensitet. Disse værdier ligger indenfor det samme interval som EC50 værdierne for de akvatiske makrofytter og er en smule højere end Tomlins (2002) værdier for Scenedesmus subspicatus på 16-24 μg l-1. 4.2.2.1 Kemiske analyserAnalyser af terbuthylazin i vandet viste, at koncentrationen i det tilførte vand var ca. 7212% af den beregnede koncentration. Dette skyldes sandsynligvis en kombination af adsorption af herbicidet til overflader og ukomplet opløsning af det ikke særligt let opløselige herbicid (Opløselighed: 8500 μg l-1), der blev tilsat direkte til de tre højeste koncentrationer. Ca. 87±12% af den terbuthylazin, der blev tilsat akvarierne, kunne genfindes efter 2-3 dage, og der var ikke nogen variation i de målte koncentrationer over tid (ANOVA: p = 0.86; p = 0.99 for henholdsvis det første og det andet forsøg). 4.2.3 Arts-følsomheds-fordelinger og "naturlige" koncentrationerArts-følsomheds-fordelingerne for de to herbicider kan ses i Figur 10. Arts-følsomheds-fordelinger er lavet ved at sortere arterne efter deres følsomhed overfor et givet stof, angivet ved deres ECx. Hver art gives derefter en placering i fordelingen efter formelen: (100* rang nummer)/(1+n), hvor n er antallet af arter (Klaine et al., 1996). Rangen afbilledes herefter som funktion af følsomheden og beskrives med en sigmoid kurve (Van Straalen, 2002). Stejle kurver er betegnende for grupper med meget ens følsomhed, hvorimod flade kurver viser en stor spredning i følsomhed mellem arterne.
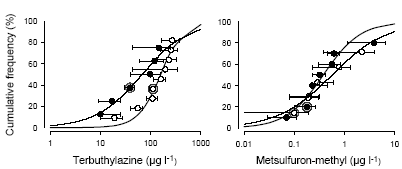
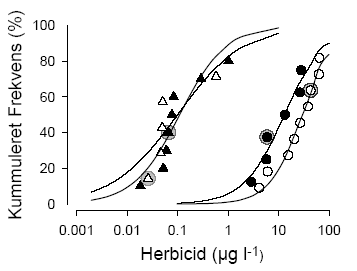
Figur 10. Arts-følsomheds-fordelinger for terbuthylazin og metsulfuron-methyl baseret på EC50 værdierne (Tørvægts- og overflade/vægt-baserede) for de to forsøg: lav lysintensitet, lukkede symboler, fed kurve og høj lysintensitet, åbne symboler, tynd kurve. Data for standardtest organismen Lemna minor er markerede med en grå cirkel. Værdierne er middel den asymptotiske standard afvigelse. Data er beskrevet med en logistisk model. Sammenlignet med terrestriske planter er variationen i følsomhed mellem arterne overraskende lille. For terbuthylazin var der en faktor 34 mellem EC50 for den mest og mindst følsomme art, og for metsulfuron-methyl var faktoren 56, hvilket for metsulfuron skal sammenlignes med en faktor >1000 for terrestriske planter og en faktor 350 på 10 grønalgearter (Boutin et al., 2000; Nyström et al., 1999). Den forholdsvis lille spredning er også observeret for andre herbicider testet i mikrokosmos eksperimenter (Brock et al., 2003). I ovenstående analyse af forskellen i følsomhed er Sparganium, som var ufølsom overfor begge herbicider indenfor det koncentrationsinterval, vi anvendte, ikke indregnet. Sparganiums tolerance kan enten skyldes plantens morfologi, hvor meristemet og de inderste blade på den ålegræs-lignende plante er godt beskyttede af de yderste blade, hvilket kunne betyde, at det eksponerede overfladeareal i forhold til biomassen er forholdsvist lille; eller det kan skyldes at Sparganium immobiliserer og metaboliserer herbicider hurtigere end andre akvatiske planter. Da Sparganium sp. også er en af de planter, der er mest robust overfor grødeskæring (Baattrup-Pedersen, 2000), vil en evt. kraftig herbicid belastning, der vil sænke væksten af andre arter, kunne fremme dens dominans. Der var ingen korrelation mellem EC50 og herbicid eksponeret specifikt bladareal (SLA) eller vækstrate for terbuthylazin, mens der var en signifikant korrelation mellem logaritmen til begge parametre og logaritmen til EC50 for metsulfuron-methyl (Korrelation: r = 0,69; p = 0,03 for SLA og r = -0,79; p = 0,01 for Relativ Vækstrate). Den positive korrelation mellem EC50 og SLA var overraskende, da den betyder, at planter med et stort eksponeret overfladeareal pr. tørvægtsenhed er mindre følsomme overfor metsulfuron-påvirkningen end planter med et lavt eksponeret overfladeareal. Vækstraten og SLA var imidlertid også korrelerede, så det var de planter med den laveste SLA, der var de hurtigst voksende. Den negative korrelation mellem EC50 og vækstrate viser, at de hurtigst voksende planter var de mest følsomme. Da det har været vist i andre studier, at hurtigt voksende planter er mere følsomme over for sulfonylurea herbicider end langsomtvoksende planter (Cobb, 1992), mener vi, at det er de metabolske forskelle fremfor de morfologiske, der i største grad kan forklare forskellene i følsomhed overfor metsulfuron-methyl hos de testede akvatiske planter.
Figur 11. Arts-følsomheds-fordelinger for terbuthylazin (cirkler) og metsulfuron-methyl (triangler) baseret på EC10 værdierne for de to forsøg: lav lysintensitet, lukkede symboler, fed kurve og høj lysintensitet, åbne symboler, tynd kurve. Data for standard-test organismen Lemna minor er markerede med en grå cirkel. Data er beskrevet med en logistisk model. Andemaden Lemna minor var for begge herbiciders vedkommende representativ for de resterende arter. For metsulfuron-methyl var den en af de mest følsomme, og for terbuthylazin lå den mere i midten af fordelingen (Figur 10). Følsomhederne opnået over 14 dage i forsøgene med flere arter, stemte også godt overens med resultaterne fra standardtestene (Tabel 2), hvilket støtter troværdigheden af standardtestene. Hvis man prøver at betragte arts-følsomheds-kurver baseret på EC10, som er det effektniveau, der ofte bruges som et økologisk "ingen effekt niveau", kan man beregne det koncentrations interval der teoretisk set vil beskytte 95% af arterne i det pågældende samfund (Van Straalen, 2002). For metsulfuron-methyl vil den koncentration, der vil beskytte 95% af makrofyt-samfundet (The 5% Hazard Concentration: HC5) være henholdsvis 1 og 5 ng l-1 for de to eksperimenter. For terbuthylazin er koncentrationerne 1 og 3 μg l-1. Beregnes HC5 værdierne for EC50-arts-følsomheds-fordelingerne er størrelserne ca. en faktor 10 større end for EC10-fordelingerne, nemlig 14 og 31 ng l-1 for metsulfuron og 3 og 33 μg l-1 for terbuthylazin. Dette svarer godt til den faktor 10, der ofte bruges, når der skal ekstrapoleres fra EC50 til NOEC. Også i forhold til EC10-følsomheds-fordelingen er Lemna minor representativ for de resterende arter og vil med den faktor 10 der bliver brugt mellem NOEC for Lemna og den højest tilladelige koncentration (Pesticidkontoret, 1999), kunne beskytte over 95% af vandplante-arterne. HC5 værdierne for metsulfuron-methyl er så lave, at de kun vanskeligt kan måles med de nuværende analysemetoder, hvor detektionsgrænsen ligger omkring 10 ng l-1 (Køppen & Spliid, 1998). På den anden side er de estimerede HC5 værdier lave i forhold til hvad der er underbygget af data, så de skal nok tages for værende et meget konservativt estimat. For terbuthylazin derimod ligger HC5-værdierne faktisk under de koncentrationer på omkring 10 μg l-1, der er blevet målt i Danmark. Det vil sige, at hvis de 10 g terbuthylazin l-1 der er blevet målt er nogenlunde konstante over tid, vil terbuthylazin kunne påvirke væksten og dermed muligvis også sammensætningen af arter af vandplanter i det danske vandmiljø. Det skal nævnes, at sensitiviteten af vandplanter overfor terbuthylazin svarer til de værdier, der findes på det langt bedre undersøgte herbicid, atrazin, der ligner terbuthylazin både fysisk/kemisk og m.h.t. virkemåde (Solomon et al., 1996). 5 Giftighedens afhængighed af tid, effekt variabel og naturlige stressfaktorer
Hvad betyder valget af forsøgstid og effekt variabel for de giftighedsparametre man bestemmer, og som der tages udgangspunkt i, når der lovgives omkring pesticider? Nogle herbicider virker hurtigt andre langsomt. Det kan tage uger, før virkningen af mange ALS-inhibitorer samt glyfosat viser sig under feltforhold (Cobb, 1992). Spørgsmålet er derfor, om den forsøgstid, der bruges til screening for akvatisk fytotoksisitet på 7 dage for Lemna, er tilstrækkelig til at kunne fastslå den fulde virkning af langsomt virkende herbicider som f.eks. glyfosat og sulfonylurea herbicider? Herbicider påvirker også forskellige processer i planterne. Nogle påvirker f.eks. celledelingen, mens andre virker direkte på fotosyntesen. Dette betyder bl.a., at den estimerede giftighed af et stof må afhænge af, hvilken effekt variabel man måler på, sådan som det også blev vist for metsulfuron-methyl i afsnit 4.2.1. Vi undersøgte variationen i toksisitet udtrykt som EC50 og EC10 over tid for 4 af de effekt variable der er mest anvendt for tests på standard organismen Lemna minor (OECD guideline for testing of chemicals, 2000) på to herbicider: 1) Metsulfuron-methyl, som er et langsomt virkende sulfonylurea-herbicid, der hæmmer dannelsen af de grenede aminosyrer, og 2) terbuthylazin, som er en hurtigt virkende hæmmer af PS II komplekset i fotosynteseapparatet. Naturlige vækstfaktorer kan også påvirke et stofs giftighed. Generelt undersøger man effekten af herbicider på testplanter, der vokser under optimale vækstbetingelser. Dette giver også mening, da nogle undersøgelser viser, at man i hvert fald for sulfonylurea herbicider får den hurtigste effekt på hurtigt voksende planter (Cobb, 1992). Mayer et al. har tilsvarende vist, at alger i hurtig vækst ved høje lysintensiteter er mere følsomme overfor kemisk stress end alger, hvis vækst er lysbegrænset (Mayer et al., 1998). Der er imidlertid en masse faktorer i det naturlige miljø, der både påvirker tilgængeligheden af herbiciderne og muligtvis også deres fysiologiske virkning. En af de faktorer, de tidligere beskrevne forsøg har vist, kunne være af betydning for tilgængeligheden og dermed for planternes optag af herbicider, er pH (afsnit 2.2.1). Forsøg med alger har vist, at det primært er den uladede ion, der optages (Fahl et al., 1995, Mayer et al., 1998). Hvis dette også er tilfældet for højere planter, kan det forklare den op til faktor 1000 store forskel i toksisitet mellem Lemna og alger, der dyrkes ved pH på helholdsvis 5 og 8. Da pH i dansk overfladevand kan variere mellem pH 5 i sure tørvegrave og pH 10 i eutrofe vande (Sand-Jensen, 2001), kan en mulig pH-effekt på toksisiteten være af stor betydning. På grund af herbiciders specifikke virkning på forskellige biokemiske processer, kunne man forestille sig, at miljømæsige forhold, der påvirker de samme processer som et givet herbicid, kunne forøge eller formindske et herbicids effekt. Vi vil i dette afsnit kort gøre rede for hvilke miljømæsige parametre, der kan være af betydning for herbiciders plantetilgængelighed og toksisitet i det akvatiske miljø, underbygget af enkelte forsøg. 5.1 Materiale og Metoder5.1.1 Tid og effekt variable hos LemnaLemna minor blev dyrket som beskrevet i afsnit 2.1.1. Fem fortyndingsrækker med koncentrationerne: 0,053; 0,15; 0,43; 1,2; 3,5 og 10 μg metsulfuron-methyl l-1 og 7,9; 23; 64; 184; 825 og 1500 μg terbuthylazin l-1 og 6 replika for hver koncentration inklusiv kontrol blev fotograferet og anbragt i klimaskabe ved 24°C og en konstant lysintensitet på ca. 100 mol m-2 s-1 (PAR). Stamopløsninger for samtlige fortyndingsrækker blev opbevaret i køleskab ved 5°C. Areal, frond antal, friskvægt og tørvægt blev bestemt på 6 Lemna populationer identiske med startmaterialet. Efter henholdsvis 3, 6, 9, 12 og 15 dage blev planterne fotograferede for at bestemme areal og frondantal, og en fortyndingsrække med tilhørende kontrol blev høstet og friskvægt og tørvægt bestemt. På dag 3, 6, 9 og 12 blev de planter, der ikke blev høstet, overflyttet til friskt medie, bestående af de originale stamopløsninger, for at undgå evt. næringsmangel. På dag 9 og 12 blev de hurtigst voksende planter delt i to brønde for at undgå tætheds-afhængig vækst. På baggrund af areal, frond antal, friskvægt og tørvægt blev de relative vækstrater for de 5 forskellige forsøgstider beregnet, og data blev beskrevet med en logistisk dosis-responskurve som beskrevet i afsnit 2.1.1. 5.1.2 Effekt variable hos ElodeaI forsøgene med flere arter blev ikke kun vækstrater og specifikt bladareal målt og beregnet på alle planter, men også morfologiske parametre som antal sideskud, skudlængde, bladantal og –længde, rodantal og –længde samt dannelsen af blomster og frugtlegemer ved forsøgs-afslutning blev talt og målt. Data var i mange tilfælde langt fra tilstrækkelige til, at det var muligt at beskrive dem med en dosis-respons kurve. I enkelte tilfælde kunne det dog lade sig gøre at få tilfredsstillende dosis-respons kurver på flere effektvariable på de samme planter. Nogle af disse vil blive præsenteret her. 5.1.3 Forsøg med pHFor at be- eller afkræfe om forskellen i toksisitet af svage syrer mellem Lemna og alger kan skyldes forskellen i forsøgs-pH, blev toksisiteten af metsulfuron-methyl målt på Lemna minor tilpasset og dyrket i algemedie pH 8 tilsat 12.8 mM NO3-, svarende til koncentrationerne i det medie standard testene med Lemna normalt udføres i. Bortset fra mediets sammensætning blev forsøgene udført som beskrevet i afsnit 2.1.1. 5.2 Variation som følge af forsøgsdesign5.2.1 Giftighedens afhængighed af tidParametrene EC50, som er det mest anvendte udtryk for toksisitet og EC10, der ofte bruges som et udtryk for No Observable Effect Concentrations, fra dosis-respons kurver baseret på relative vækstrater beregnet på baggrund af de fire forskellige effekt variable, er afbilledet som funktion af tid i Figur 12. Det ses, at der for effekt variablene frond antal og tørvægt ikke kunne beregnes tilfredsstillende dosis-responskurver for metsulfuron-methyl efter 3 dage. Efter 6 dage kunne man dog få tilfredsstillende parametre for samtlige effekt variable. Hvis den fulde toksisitet af et stof ikke var kommet til udtryk efter 6 dage, ville vi have forventet at ECx værdierne ville falde (Walker et al., 2001). Dette skete fra dag tre til dag 6 for en del af effektvariablene, men derefter var
Figur 12. Parametrene EC50 og EC10 fra dosis-responskurver baseret på relative vækstrater beregnet på baggrund af 4 effekt variable: Frond antal (lukket cirkel), bladareal (åben trekant), friskvægt (åben cirkel) og tørvægt (lukket trekant) afbilledet som funktion af tid. Usikkerhederne på bestemmelsen af parametrene er opgivet i figur 13. ECx-værdierne relativt konstante eller endda svagt stigende. Denne stigning kunne skyldes en tilpasning til herbiciderne eller at koncentrationen af herbicider faldt i løbet af forsøgstiden i de opbevarede stamopløsninger. Ser man på usikkerheden på bestemmelserne af ECx-værdierne (Figur 13), falder den klart som funktion af tid. Mest tydeligt ses det for terbuthylazin, hvor man allerede efter 6 dage har en CV på EC50 på under 20% for samtlige effekt variable.
Figur 13. Coefficient of variation (CV%) på EC50 og EC10 udregnet som assymptotisk standard fejl på parameteren * 100%/parameteren, som funktion af tid. Symboler som i figur 12. For metsulfuron-methyl er vækstrater baseret på areal eller friskvægt klart at foretrække, p.g.a. den mere sikre bestemmelse af EC50 og EC10. Usikkerheden på frond antal og tørvægt kan dog formindskes ved at øge antallet af planter per replika, men dette medfører en meget pladskrævende forsøgsopsætning. Forsøget viser, at selv for et meget langsomt virkende herbicid som metsulfuron-methyl, er den fulde giftighed udtrykt efter en forsøgstid på 7 dage, som er standard test-tiden (OECD guideline for testing of chemicals, 2000). 5.2.2 Giftighedens afhængighed af effekt variabelFor et stof som terbuthylazin, der hæmmer fotosyntesen og dermed indirekte kulstoffikseringen, havde vi forventet, at tørvægtsspecifik vækst ville være en følsom parameter. For metsulfuron-methyl, der har den indirekte effekt at stoppe celledelingen få timer efter applikation (Cobb, 1992), forventede vi, at areal specifik vækst ville være den mest følsomme parameter. Begge disse forventninger viste sig at holde stik (Figur 12). Mest markant for EC50 og mest markant ved korttids-forsøg. Hvis man ser på giftighedens afhængighed af effekt variabel efter 6 dage, som er det tidsrum, der ligger tættest op af standard test-tiden, varierer EC50 imidlertid ikke mere end maksimalt en faktor to. Så hvis man tager de sædvanligt anvendte sikkerhedsfaktorer i betragtning, er en faktor to ikke alamerende. Vender man sig mod makrofytterne, kan man finde langt større forskelle mellem forskellige effekt variable (men også langt større usikkerhed på bestemmelsen af en ECx-værdi). I figur 14 er udviklingen i fire morfologiske karakterer som funktion af metsulfuron-methyl koncentrationen i mediet vist sammen med en fittet eller visuelt estimeret EC50værdi. De tørvægtsbaserede vækstrater i det samme forsøg var ikke påvirkede af herbicidet (Cedergreen et al., 2003b), men morfologien ændrede sig tydeligt. Der var op til en faktor 50 forskel på de estimerede EC50-værdier. Disse resultater stemmer godt overens med forsøg på Myriophyllum aquaticum, hvor vækst, friskvægt forøgelse, længdevækst, total rodlængde og rodantal var målt sammen med pigmenterne klorofyl a, b og karotenoider på fortyndingsrækker af 17 forskellige herbicider (Turgut & Fomin, 2002). Disse forsøg viste, at for samtlige herbicider var der en eller flere af effektvariable, der ikke udviste nogen signifikant respons på herbicidet, mens andre gjorde. De eneste effektvariable hvor det altid var muligt at beregne EC50 var pigmentmålingerne. Forskellen i EC50 mellem effekt variablene var på en faktor 2-10. I forsøg med makrofytter anbefales det derfor altid at tage mere end en effekt variable i betragtning, og det synes at være en god idé at lade pigment koncentrationer være en af dem.
Figur 14. Udviklingen i fire forskellige morfologiske karakterer hos Elodea canadensis som funktion af koncentrationen af metsulfuron-methyl. Data er fra forsøg 2 med flere arter (Kapitel 4). 5.3 Naturlige faktorers påvirkning af giftighed5.3.1 Herbicid tilgængelighedVi så allerede i Kapitel 3 (Tabel 2), at vandets pH muligvis har en stor indvirkning på svage syrers toksisitet. Dette er vist for alger og skyldes, at den negative ion, som er den dominerende form ved pH højere end stoffets pKa har sværere ved at trænge ind i en celle. Planteceller har nemlig et negativt potentiale over cellemembranen (Ullrich et al., 1984). Det er derfor altid vigtigt at vide, under hvilken pH en test er foretaget, hvis man vil vide noget om den potentielle toksisitet af svage syrer. Af de 146 herbicider, der var med i litteraturstudiet, havde ca. 40% en pKa værdi på <pH 5, som er den pH mange Lemna test udføres ved. Toksisiteten ude i miljøet ved en pH 7-9, kan derfor nemt vise sig at være meget lavere. Forsøget med Lemna minor dyrket ved pH 8 viste imidlertid, at toksisiteten ved denne pH var tilsvarende toksisiteten ved pH 5 (Figur 15). Dette var også tilfældet i forsøget med flere arter (Kapitel 4), hvor forsøgs-pH svingede mellem pH 7.7 og pH 8.5. Dette kan enten være, fordi pH ikke spiller nogen rolle for optaget, men at Lemna er meget følsom overfor de herbicider, vi har testet, og som er svage syrer. Hvis vi imidlertid sammenligner EC50 værdier for de 27 herbicider, hvor EC50 er opgivet for både alger og Lemna i Tomlin (2002), viser det sig, at der stort set ikke er forskel i toksisiteten mellem de to arter for de 10 af herbiciderne, som er uladede (EC50 alger/EC50 Lemna = 1.48±2.64 (Interval: 0.01-8.0)). For de 17 herbicider, der har en pKa < 5, eller hvor Kow er opgivet ved flere pH værdier (indikere at molekylet kan dissociere), er EC50 for alger imidlertid i gennemsnit ca. 75000 gange højere en EC50 for Lemna (Interval: 1.3 – 625000). Der er altså noget, der kunne tyde på at svage syrer virker forskelligt på de to organismer.
Figur 15. Den relative vækstrate af Lemna minor som funktion af metsulfuron-methyl koncentrationen i mediet ved pH 8 (n=3). Forsøget er udført to gange (Åbne og lukkede cirkler). En anden faktor, der i høj grad kan påvirke mængden af plante-tilgængeligt herbicid, er vandets turbiditet, altså mængden af organisk materiale og sediment-partikler. Studier med glyfosat, et stof der binder hårdt til jord, fortyndet med flodvand har vist, at stoffets effektivitet afhænger af mængden af opløst materiale. Også mængden af kationer som Ca++ og Mg++ påvirker glyfosats effektivitet (Stahlman & Phillips, 1979; Thelen et al., 1995). Ofte optræder de største koncentrationer af herbicider i forbindelse med nedbørshændelser, og udvaskningen medfører typisk også store mængder af jordpartikler m.m. Spørgsmålet er derfor, hvor stor en del af den målte herbicidkoncentration, der rent faktisk er tilgængelig for planter og alger? En fremtidig differentiering mellem partikelbundne og opløste pesticider ville kunne nuacere undersøgelser af pesticiders effekt. 5.3.2 Fysiologisk virkningBegge fler-arts-forsøg blev udført under to forskellige vækstbetingelser, for at se, om herbicidets effektivitet blev påvirket af vækstbetingelserne. I forsøgene med metsulfuron-methyl var vækstraten generelt højere i forsøget med høj lysintensitet, på trods af det noget lavere fosfor-niveau, men toksisiteten forblev uændret (Figur 8 og 10). For et stof som metsulfuron-methyl, der hæmmer dannelsen af visse aminosyrer, kunne vi forvente, at f.eks. N-stress ville påvirke herbicidets effektivitet, men dette er ikke systematiskt undersøgt. For terbuthylazin forsøget var slutvægtene og dermed vækstraterne ikke markant forskellige mellem de to forsøg (Figur 9). Til gengæld var toksisiteten lavere under de høje lysintensiteter (Figur 10). Dette er overraskende, da terbuthylazin blokerer elektrontransporten i PSII-fotosystemet, hvilket fører til dannelsen af oxygene radikaler, der ødelægger membraner m.m. inde i cellen (Fedtke, 1982b). Højere lysintensiteter og faktorer, der generelt vil fremme fotoinhibering, så som lave temperaturer og næringsstress (Vestergaard, 2003; Osmond, 1994; Osmond, 1994), er derfor forventet også at forøge effekten af terbuthylazin og andre fotosyntese-hæmmere. Årsagen til, at vi ikke så dette i vores forsøg, mener vi kan skyldes den øgede vækst af epifytter under højere lysintensiteter. Et øget epifyt lag vil nedsætte kontakten mellem planten og herbicidet og dermed nedsætte optaget. Hvor vidt dette også vil gøre sig gældende i naturen er uvist, da der i disse systemer også findes græssere, der vil minimere væksten af epifytter. Det er derfor ikke usansynligt, at man under naturlige forhold vil kunne finde, at planter udsat for høj indstråling, vil være mere følsomme overfor fotosyntesehæmmere, end planter udsat for lavere indstråling. 6 Effekter af pulseksponeringDet blev i Kapitel 4 konkluderet, at terbuthylazin kunne have en effekt på væksten af op til flere vandplantearter, hvis de målte terbuthylazin koncentrationer på op til 10 μg l-1 er konstante over tid, sådan som det var tilfældet i forsøgene med flere arter. Høje og potentielt toksiske herbicid koncentrationer er dog, så vidt vi ved, oftest kun til stede i pulse associeret med nedbørshændelser, punktkildeforurening fra vaskepladser etc. (Styczen et al., 2003). Pulsenes længde kan variere fra få timer til dage, afhængigt af hvor tæt på kilden de måles. De kraftigste er dog typisk de korteste. Spørgsmålet er så, hvor kraftig effekten af en puls er i forhold til effekten af en længerevarende eksponering? Og hvor lang tid det tager en plante at komme sig over en ikke-dødelig herbicid-puls? Man kunne forvente, at effekten af en puls kunne afhænge af et stofs optagelse- og frigivelseshastighed, da stoffer med en langsom optagelse og en langsom frigivelse forventes at findes i planterne i længst tid efter en puls. På den anden side vil der ikke nås den samme høje interne koncentration som for et stof, der optages og frigives hurtigt. Effekten af en puls kunne også være relateret til plantens evne til at inaktivere herbicidet, samt reparere den påførte skade. For at undersøge denne problemstilling, udsatte vi andemad for både en 3-timers puls og en 4 dages eksponering til 6 udvalgte herbicider, der representerer tre virkemekanismer, men har forskellig Kow, som er et udtryk for et stofs fedtopløselighed, og som påvirker optagelse- og frigivelses hastigheden. 6.1 Materiale og MetoderForsøgene blev foretaget med Lemna minor i testsystemet beskrevet i afsnit 2.2.1. Alle stoffer blev omrørt i mindst et døgn, for at sikre fuld opløselighed af de svært opløselige tekniske produkter. Imazamox var det eneste anvendte formulerede produkt. Formuleringen indeholdt ingen organiske opløsningsmidler eller andre potentielt toksiske stoffer (Jørgen Lundsgård, BASF, personlig kommunikation). Stoffernes virkemåde samt fysisk/kemiske egenskaber er beskrevet i tabel 8. Tabel 8. Virkemåde (HRAC-klassifikation, se tabel 3) og fysisk/kemiske egenskaber af de anvendte herbicider.
Ved forsøgene, der skulle simulere en puls, blev planterne overflyttet til rent medie 3 timer efter inkubering. Forsøgene forløb over 4 dage. Planterne blev tillige fotograferet dagligt i op til 7 dage. Alle forsøg blev gentaget mindst to gange. Data blev analyseret som tidligere beskrevet i afsnit 2.2.2. 6.2 Resultater og Diskussion6.2.1 Puls- versus langvarig-eksponeringDosis-respons kurverne for puls- versus langvarig-eksponering kan ses i figur 16, og værdierne for kurvefittene i tabel 9.
Figur 16. Den relative arealspecifikke vækstrate (RGR) af Lemna minor udsat for enten puls-(Åbne symboler) eller langvarig-eksponering (Lukkede symboler) til forskellige herbicider. Data er udvalgte 4-dages forsøg, middelstdev (n = 3-6, kontrol n =12) og koncentrationerne er angivet i M, for bedre at kunne sammenligne herbicidernes aktivitet. Data er beskrevet med en logistisk dosis-responsmodel (Tabel 9). Tabel 9. Parametre fra den logistiske dosis-respons model (Streibig et al., 1993) for Puls- og langvarig-eksponering af Lemna minor til forskellige herbicider, samt forholdet mellem EC50 for de to eksponeringstyper.
Resultaterne tyder på, at det er herbicidernes virkemåde, mere end deres Kow, der bestemmer effekten af en pulseksponering. Stoffer som terbuthylazin og propyzamid har næsten ens Kow, og man kunne derfor forvente et parallelt optags- og frigivelses-forløb. Ikke desto mindre ser man en signifikant effekt af en puls for propyzamid, men ingen for terbuthylazin. Heller ikke overfor diquat har en 3 timers puls af en størrelse, der er dødelig ved langvarig eksponering, nogen signifikant effekt (Figur 16, tabel 9). Dette viser, at pulseksponeringer til fotosyntesehæmmere som diquat og terbuthylazin er relativt ineffektive. Vi mener ikke, at det skyldes forskelle i optag mellem fotosyntesehæmmerne og de resterende stoffer, men det er vores hypotese, at det snarere skyldes planternes evne til at reparere de skader, der opstår som følge af fotosyntesehæmmere. Som beskrevet i forrige afsnit (4.2), forårsager fotosyntesehæmmere en øget dannelse af oxygen-radikaler, der går ind og ødelægger membraner og andre celle-komponenter. Denne form for skade sker hele tiden i planter, men med en lavere hastighed. Planter har derfor systemer, der kan inaktivere de oxygene radikaler, og udbedre de skader de forårsager (Osmond, 1994). En forholdsvis kort eksponering til fotosyntesehæmmere vil derfor ikke forårsage større skader, end de hurtigt kan repareres. Forsøg med pulseksponering til atrazin har vist lignende resultater (Solomon et al., 1996). Modsat forholder det sig åbenbart for stoffer, der inhiberer syntesen af aminosyrer (med den side-effekt at celledelingen stopper efter få timer (Cobb, 1992)) og stoffer, der hæmmer samlingen af mikrotubuli og forhindrer celledeling ad den vej. For alle disse stoffer ser vi en effekt af en tre timers puls ved koncentrationer, der er ca. en faktor 10 højere end de koncentrationer, der giver en effekt ved langvarig eksponering (Tabel 9). 6.2.2 "Recovery"Hvor lang tid tager det for en plante at vende tilbage til fuld vækstrate efter at have været udsat for en ikke dødelig dosis herbicid? Vi fulgte alle planter i op til 7 dage, og figur 17 viser dosis-responsforløbet for imazamox og metsulfuron-methyl efter både 4 og 7 dage. 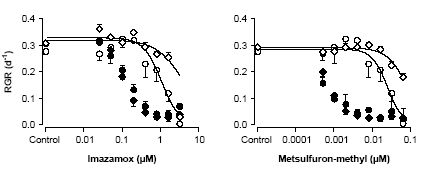
Figur 17. Den relative arealspecifikke vækstrate (RGR) af Lemna minor udsat for enten puls-(Åbne symboler) eller langvarig-eksponering (Lukkede symboler) til imazamox og metsulfuron. Data er vækstrater efter 4 dage (Cirkler), eller 7 dage (Ruder) og er beskrevet med en logistisk dosis-respons model (middel stdev, n = 3). Den højeste puls-koncentration for imazamox er ikke medtaget i fittet for 7 dages forsøget, da planterne var døde. Det ses, at der ikke er den store forskel på vækstraterne efter 4 og 7 dage for en længerevarende eksponering. De planter, der ikke er udsat for en dødelig dosis, har tilsyneladende en reduceret men rimelig konstant vækst. De puls-eksponerede planter derimod kommer sig hurtigt, hvis puls-dosen altså ikke er dødelig, som det er tilfældet for den højeste dosis imazamox. Ser man på udviklingen i bladareal over tid for de planter, der har været udsat for puls-eksponering ved de to højeste doser metsulfuron-methyl(Figur 18), er det tydeligt, at væksten er hæmmet for alle planter det første døgn, sikkert på grund af håndterings-stress. Derefter har kontrolplanterne en relativ vækstrate på omkring 0.31 d-1. Væksten af de planter, der har været udsat for en puls på 33 nM/12,5 μg l-1 metsulfuron-methyl, er hæmmet i ca. 3 dage, hvorefter de begynder at vokse med en vækstrate analog til kontrol-vækstraten. For planter, der har været udsat for en puls på 66 nM/25 μg l-1 metsulfuron, tager det ca. en dag længere inden væksten atter er oppe på kontrol-vækstrater.
Figur 18. Udviklingen i bladareal over til for kontrolplanter (Sorte symboler), og planter udsat for en metsulfuron-methyl puls på henholdsvis 33 nM/12,5 μg l-1 (Grå symboler) og 66 nM/25 μg l-1 (Hvide symboler). Bladarealet er relativt til arealet af planten ved start, som er sat til 1 (n = 3, for kontrol n = 12). Notér log-skala. Den tid det tager at komme sig over en ikke-dødelig puls, afhænger altså af puls-koncentrationen, men overstiger ikke 4 dage i forsøgene med metsulfuron-methyl. Mønsteret var det samme for imazamox, og planter udsat for propyzamid og pendimethalin udviste samme tendens. I de sidste to tilfælde kunne vi dog ikke følge udviklingen i planter, som var "næsten-døde" af en puls, da opløseligheden af stofferne satte en øvre grænse for de koncentrationer, vi kunne bruge i forsøgene. En maksimal "recovery-time" på 4 dage kan synes lang, taget i betragtning, at pulsen kun var af 3 timers varighed. Sammenlignet med frekvensen af estimerede pulse, som det ikke er sandsynligt, ofte vil forekomme med mindre end 4 dages mellemrum, kommer planterne sig dog hurtigt (Liess et al. , 1999; Solomon et al., 1996)Det er dog langt fra sikkert, at "recovery" hos den hurtigt voksende Lemna minor er repræsentativ for andre mere langsomt voksende akvatiske arter. 7 Kombinationseffekter
Resultaterne indtil videre har vist, at ingen af de i Danmark målte herbicider i søer og vandløb optræder i koncentrationer, der kan betragtes at udgøre en risiko for vandplanter og alger. Undtagelsen var terbuthylazin, men da de højeste og potentielt væksthæmmende koncentrationer formodes at forekomme i pulse, som for terbuthylazins vedkommende har en meget lille effekt, er det heller ikke sandsynligt, at målte terbuthylazin-koncentrationer vil påvirke den akvatiske flora radikalt. Herbicider og andre potentielt væksthæmmende stoffer findes imidlertid sjældent enkeltvis (Løkke, 2000). Ud af de 140 NOVA-prøver, hvor der blev målt pesticider i koncentrationer over detektionsgrænsen, var der mere end 7 stoffer tilstede i over halvdelen af prøverne. Amerikanske undersøgelser af over 80 stoffer i overfladevand viste tilsvarende resultater (Martin et al., 2003). I Fyns Amt blev der målt op til 18 forskellige stoffer på et døgn (Løkke, 2000). Så hvad er effekten af tilstedeværelsen af flere stoffer på en gang? Og findes der metoder, hvormed man kan estimere effekten af flere stoffer, hvis man kender koncentrationerne af de enkelte? 7.1 Hvad ved vi om kombinationseffekterSelvom problematikken har været kendt længe, er gode systematiske undersøgelser af kombinationseffekter af pesticider i miljøet først for alvor dukket op i løbet af de sidste 10 til 20 år. Der er mange "skoler" og forskellige teorier og terminologier, som er godt opsummeret i reviews (Altenburger et al., 2003; Greco et al., 1995; Berenbaum, 1989). I store træk kan det koges ned til to modeller: 1) Addition af koncentrationer (Concentration Addition, CA, også kaldet Additive Dose Model. ADM) og 2) Uafhængig virkning (Independent Action, IA også kaldet multiplicative survival model (MSM)). CA forudsætter, at de stoffer man regner på, har samme virkemåde. IA forudsætter, at de virker på forskellige mål i organismen. Begge modeller har vist sig at kunne bruges til at beregne effekten af blandinger, hvor mange stoffer er tilstede i meget lave koncentrationer (Faust et al., 2001; Faust et al., 2003; Walter et al., 2002) EC1 har været anvendt for at vise, at selv koncentrationer, hvis effekt vil være umulig at måle, hvis stofferne var testet enkeltvis, har en effekt samlet set. Hvis man sammenligner de to modeller finder man, af CA som regel forudsiger en større effekt end IA. Man finder også, at selv om den teoretiske baggrund bag modellerne er forskellig, vil man sjældent finde en større forskel end en faktor 4 mellem forudsigelserne af de to (Faust & Scholze, 2004). Set i en regulatorisk sammenhæng, hvor man under alle omstændigheder vil benytte sig af en sikkerhedsfaktor på 10, er en faktor 4 lille og mere af akademisk interesse. Og da CA som regel altid vil forudsige en effekt, der er større end effekten forudset med IA, kan man for at være på den sikre side bruge CA som et redskab til at vurdere den mulige maksimale effekt af en blanding. Vi vil i det følgende afsnit beskrive den additive model og v.h.a. et eksempel vise, hvordan den kan bruges til at beregne effekten af en blanding. Eksemplet kan også findes i Cedergreen et al. (2003a). 7.2 Den additive modelVirkningen af et stof på en organisme afhænger af dosis, og i langt de fleste tilfælde kan denne sammenhæng beskrives tilfredsstillende med en logistisk doseringsmodel, som beskrevet i afsnit 2.2.1. og hos Streibig et al. (1993). Kender man et stofs doseringskurve overfor en bestemt organisme, kan man beregne virkningen af en hvilken som helst koncentration af stoffet overfor den givne organisme. Når man bruger den additive model (Additive Dose Model også kaldet Concentration Addition), lægger man doserne eller koncentrationerne af de forskellige stoffer sammen, efter at man har korrigeret dem i forhold til deres relative styrker, hvorefter man beregner virkningen af hele blandingen. Man kan sige, at man omregner doserne/koncentrationerne af pesticider til samme "valuta", hvor vekselkursen bestemmes af forholdet mellem deres giftighed angivet ved deres EC50-værdi. EC50 værdier for en lang række stoffer kan hentes i databaser og opslagsværker (Cedergreen et al., 2003a). Når man har "vekslet" de forskellige koncentrationer af pesticider i en blanding til samme "valuta", lægger man dem sammen og udregner den samlede virkning ved brug af den logistiske doseringsmodel:
Hvor d er den maximale respons (vækst, overlevelse, fertilitet etc.), EC50 er den dosis der giver 50% af den maximale respons og b er proportional med hældningen på kurven i EC50. I beregningen sætter vi den maximale respons, d, til 100%, og anvender den EC50, man har brugt som sin "valuta". Derefter mangler vi bare kurvens hældningsparameter, b, for at kunne beregne virkningen af summen af stofferne. For herbiciders virkning på både alger, vand- og landplanter varierer b som regel mellem 1 og 3 (Cedergreen et al., 2003a). Da de koncentrationer, vi vil beregne virkningen af i en miljømæsig sammenhæng, som regel er lavere end EC50, er det stoffer med en lav b, der vil have den største virkning, hvis de har den samme EC50. Hvis vi derfor antager, at b er lig 1 i de tilfælde, hvor vi ikke har kendskab til den målte b, får vi et konservativt estimat for virkningen af et stof ved koncentrationer < EC50. Tyske undersøgelser med algetests af over 106 forskellige kemiske stoffer viser, at b-værdier <1 findes for mindre end 20% af stofferne (Faust & Scholze, 2004). 7.2.1 BeregningFor at give et eksempel, hvor man kunne have mistanke om en væsentlig miljøpåvirkning, har vi valgt et datasæt, hvor DHI og DMU i forbindelse med en afstrømningshændelse har målt koncentrationer af terbuthylazin, diuron og atrazin på op til henholdsvis 4; 1,8 og 0,5 μg/L (Styczen et al., 2003). Vi finder EC50 værdierne for atrazin og terbuthylazin målt på algen Pseudokirchneriella subcapitata, som er opgivet til at være henholdsvis 45 g/L og ca. 20 μg/L (Tomlin, 2002). For diuron findes der data på algen Chlorella pyrenoidosa, hvor EC50 er opgivet til at være 24 μg/L (Miljøstyrelsen, pers. comm.). På baggrund af EC50 værdierne beregnes den relative styrke af de målte herbicidkoncentrationer og koncentrationerne lægges sammen (Tabel 10): Tabel 10. Toksikologiske parametre og udregning af den relative toksisitet for de målte herbicider.
Dernæst beregnes algernes relative vækstrate, når de udsættes for det, der svarer til en terbuthylazin koncentration på 5,72 g/L og under antagelsen af, at hældningsparameteren på dosis-responskurven er 1:
En beregnet vækst på 78% betyder, at den pågældende blanding af terbuthylazin, diuron og atrazin maximalt vil kunne hæmme algernes vækst med 22%, hvis kurverne har en hældningsparameter på 1. Sættes hældningsparameteren derimod til 2, sådan som det er målt i adskillige forsøg med P. supcabitata, får man følgende resultat:
Brugen af en mere realistisk hældningsparameter sænker altså den potentielle væksthæmmende virkning fra 22% til 8%. Denne blanding ligger altså på grænsen af, hvad der kunne være kritisk. Da de målte koncentrationer er pulskoncentrationer og herbiciderne fotosyntesehæmmere, er det dog usandsynligt, at denne blanding alene vil have en stor effekt på floraen (j.f. Kapitel 6). Eksemplet viser, at det er muligt, på en forholdsvis enkel måde, at estimere den maximalt mulige virkning af en pesticid-blanding på forskellige organisme-grupper, hvis der findes toksikologiske data på enkeltstofferne. Dette estimat kan så bruges i en samlet vurdering af, om denne blanding kan have en signifikant miljømæssig virkning, og til identifikation af hvilke undersøgelser det vil være relevante at fortage for at belyse en sådan mulig virkning. 7.3 Metsulfuron-methyl og terbuthylazinDa vi i dette studie har koncentreret os meget om metsulfuron og terbuthylazin, har vi lavet et binært blandingsforsøg med disse to herbicider. 7.3.1 Materiale og MetoderDe to testsystemer med Lemna og alger beskrevet i afnit 2.1.1. og 2.1.2. blev anvendt. Herbicider blandes i forholdene: 0:100, 17:83, 33:67, 50:50, 67:33, 83:17, 100:0 relative doser, hvilket i praksis vil sige, at der blev lavet to stamopløsninger på en faktor 10 gange stoffets EC50. Disse opløsninger blev blandet med 17 ml af det ene stof plus 83 ml af det andet etc.etc. Hver blanding blev derefter fortyndet med en faktor 2 i alt 7 gange. Dette gav 7 dosis-responskurver med hver 8 fortyndinger, samt kontrol. Lemna forsøgene blev udført med 3 replika og 12 kontrol, alge forsøgene med 2 replika og 10 kontrol. Efter forsøgets udførelse blev hver af de 7 dosis-responskurver beskrevet med en logistisk dosis-responsmodel, der inkluderer både ECx og EC50 (Streibig et al., 1995):
Vi valgte ECx niveauet til EC10. De fittede EC10 og EC50 værdier blev omregnet til koncentrationer af de to enkeltstoffer ud fra kendskabet til deres respektive andel i de forskellige blandinger. Variansen på den fittede værdi blev fordelt proportionalt med andelen af stof i blandingen. Disse koncentrationer blev indtegnet som funktion af hinanden, hvilket giver det, der kaldes et isobologram (Gessner, 1995). Gessner har i en artikel i tidsskriftet Toxicology givet en udmærket illustration af, hvad det er, et isobologram viser (Figur 19a). Det skal dog i den sammenhæng bemærkes, at skalaen på x- og y-akserne skal være logaritmisk, hvis formen på dosis-responskurverne skal være sigmoid. Men princippet er det samme, uanset skala. Figur 19a. En konceptuel model af et isobologram. a) viser en dosis-responskurve for stof 1, samt en dosis-responskurve for en 1:4 blanding af stof 1 og stof 2. b) viser dosis responskurver for stof 1 og stof 2, samt af blandinger af de to stoffer. c) Her er dosis-respons-fladen illustreret ved at forbinde punkter af blandinger, der giver den samme effekt. Kurverne er forbundet med deres projektion på x-y planet. Dette er analogt til højdekurverne på et kort. d) z-aksen er fjernet og et isobologram med kurver for 4 effektniveauer er tilbage. Figuren er lånt fra Gessner (1995). 7.3.2 Resultater og DiskussionIsobolerne for både EC10 og EC50 er vist i figur 19b. Hvis punkterne havde ligget på den rette linie, der forbinder de to enkeltstoffer, havde de fulgt modellen for koncentrations addition (CA). EC50 værdierne for størstedelen af blandingerne er imidlertid større end den forudset v.h.a. modellen for CA. Dette betyder, at blandingerne generelt er mindre giftige end forventet. Da metsulfuron-methyl og terbuthylazin er herbicider med to forskellige virkemåder, ville man forvente, at de fulgte modellen for uafhængig virkning. Denne model er indtegnet som den buede isobol på EC50-figurerne. I dette tilfælde adskiller IA sig ikke særskilt fra CA, og hvis man inddrager usikkerhederne på parameterestimaterne og på estimaterne af isobolerne, er det ikke muligt at sige, hvilke af de to modeller, der statistisk set passer bedst. En additiv eller mindre end additiv virkning er imidlertid et resultat som ofte ses (Christensen et al., 2001; Faust & Scholze, 2004; Faust et al., 1994). Synergistiske effekter på vandplanter og alger er utroligt sjælne, og det har ikke været os muligt at finde et eneste veldokumenteret eksempel (Faust & Scholze, 2004; Kloepper-Sams et al., 2001). På den baggrund og med baggrund i et større EU-projekt, der har indraget data for flere hundrede stoffer testet i forskellige testsystemer på både alger og dyr, er det konkluderet, at CA er en konservativ model, der kan forudsige effekter af blandinger indenfor en sikkerheds-faktor på 5-10 (Faust & Scholze, 2004).
Figur 19b. Isoboler for EC10 og EC50 på blandingen: metsulfuron-methyl og terbuthylazin for både alger og Lemna. Den rette linie mellem de to datapunkter på akserne udgør isobolen for CA, mens den buede linie på EC50 isobolerne udgør IA-isobolen. Punkterne er fittede værdier asymptotisk stdev. En anden konklusion, der kan drages ud fra figurerne er, at isobolerne for EC10 og EC50 er tilnærmelsesvis ens. D.v.s. at de interaktioner vi finder ved de høje koncentrationer der skal til for at nå EC50 også, i dette tilfælde, gælder for de lavere, mere miljørelevante koncentrationer. Det er også i andre forsøg vist, at modellerne CA og IA kan anvendes på blandinger med stoffer i koncentrationer, der giver meget lave effektniveauer (Faust et al., 2001; Faust et al., 2003; Junghans et al., 2003). Der er sammenfald mellem Lemna- og alge-testen, hvilket bekræfer den mindre end additive respons' generalitet på tværs af arter. 8 Vækststimulering ved lave pesticid-koncentrationerVi har i op til flere tilfælde stødt på vækststimulering ved lave doser af herbicider. Fænomenet kaldes hormesis, og er kendt fra så og sige alle systemer, hvor man måler effekten af en stressfaktor på en bestemt organisme (Calabrese & Baldwin, 2001). Denne vækststimulering bliver ofte nævnt, men de fysiologiske virkningsmekanismer bag er dårligt belyst. Spørgsmålet er, om den har nogen økologisk effekt? Nogle svampemidler er tillige kendt for at forøge væksten hos terrestriske planter (Gerhard et al., 1998). Man ved ikke, om de har samme effekt hos akvatiske planter, og hvad sådan vækststimulering evt. vil betyde for toksisiteten af forskellige herbicider? 8.1 HormesisDe forsøg, hvor vi har haft den mest tydelige hormetiske effekt, er i forsøgene med flere arter (Kapitel 4)(Figur 20). Rent visuelt så væksten i akvarierne med de to laveste koncentrationer betydeligt kraftigere og grønnere ud, og den kraftigere vækst var også signifikant målbar i flere tilfælde.
Figur 20. Forskellige eksempler på kurver, der udviser en hormetisk effekt. De markerede doseringer er signifikant højere end kontrollen (En-sidet t-test: p < 0,05). Byg blev, sammen med raps, dyrket med rødderne i akvarierne, for at kunne sammenligne de akvatiske planters følsomhed med mere kendte terrestriske (Cedergreen et al., 2003a). Undertiden har vi også observeret en vækststigning i Lemna- og algetestene (F.eks. Figur 21, terbuthylazin), men den har sjældent været så kraftig som i forsøgene med flere arter. En af forklaringerne kunne være, at planterne i standard-testene vokser nær den maksimalt mulige vækstrate. En vækst-øgning er derfor fysiologisk set ikke mulig. I fler-arts-forsøgene derimod er det mere sandsynligt, at vækstforholdene ikke har været optimale for en stor del af planterne, og de har således haft et potentiale for at øge i hvert fald nogle vækstparametre. Parametrene fra figur 20 er primært morfologiske parametre, og det er ikke sikkert, der er nogen direkte sammenhæng mellem f.eks. en forøgelse af bladlængden og vækstraten. I vores forsøg var en sådan sammenhæng i hvert fald ikke signifikant. Også det tidsmæsige perspektiv af vækststimuleringen kan være af betydning, da det af nogen er blevet observeret, at vækststimulering aftager over tid (Brock et al., 2003; Calabrese, 2001), mens andre har fundet en permanent vækstforøgelse frem til høst (Stebbing, 1997; Appleby, 2001). Da vækstvilkårene ude i naturen sjældent er optimale, kunne hormesis rent faktisk finde sted derude, men det ville kræve yderligere studier. Figur 21. Relative arealspecifikke vækstrater af Lemna minor som funktion af herbiciderne metsulfuron-methyl og terbuthylazin. Kontrol (Sort cirkel), 1 g prochloraz l-1 (Åben cirkel),10 g prochloraz l-1 (Åben firkant), 20 g prochloraz l-1 (Åben trekant) 100 g prochloraz l-1 (Åben rude), (n = 6). 8.2 Interaktioner med prochloraz8.2.1 Materiale og MetoderEffekten af svampemidlet prochloraz, både teknisk og formuleret produkt (Sportak), og dets interaktioner med toksisiteten af metsulfuron-methyl og terbuthylazin blev undersøgt på Lemna minor som beskrevet i afsnit 2.1.1. Et af forsøgene med effekten af prochloraz på toksisiteten af metsulfuron-methyl blev dog udført i et andet klimaskab ved lysintensiteter på 25-45 μmol m-1 s-1 (PAR), hvilket er noget lavere end standard intensiteterne på omkring 100 mol m-2 s-1 (PAR). 8.2.2 Resultater og diskussionDer var ingen effekt af prochloraz på væksten af Lemna minor i koncentrationsintervallet 31-1000 μg l-1 (ANOVA: F(6,47) = 3,9, p < 0,001), når Lemna blev dyrket under gode vækstbetingelser. Når Lemna blev dyrket under lave lysintensiteter med resulterende lavere vækst (Figur 21a), stimulerede 10 og 20 g prochloraz (Sportak) l-1 imidlertid væksten i forhold til kontrol. Toksisiteten af metsulfuron-methyl forblev dog relativ uændret med EC50 værdier på 0,76±0,08, 0,99±0,06 og 1,02±0,10 for metsulfuron-methyl koncentrationer på henholdsvis 0, 10 og 20 μg l-1. Så snart planterne blev flyttet til mere optimale vækstbetingelser, forsvandt denne vækststimulering dog, som det ses af figur 21a og b. På disse figurer ses det også, at prochloraz (tekniskt) under gode vækstforhold ingen effekt havde på toksisiteten af hverken metsulfuron-methyl eller terbuthylazin. Alle forsøg under høje lysintensiteter blev gentaget med samme resultat. De anvendte prochloraz koncentrationer blev valgt ud fra en hypotese om en afdrift-hændelse, da der ikke er rapporteret fund af prochloraz i overfladevand (Løkke, 2000, NOVA-DATA). Den maksimalt anbefalede prochloraz dosis på 600 g ha-1, vil, hvis den udsprøjtes på en 30 cm vandsøjle, give en koncentration på 200 g l-1. De anvendte 10, 20 og 100 μg prochloraz l-1 svarer derfor til henholdsvis 5, 10 og 50% afdrift. I felten kan en vækstforøgelse som følge af brug af fungicider skyldes nedkæmpelsen af patogene svampe. Laboratorieforsøgene med Lemna viser imidlertid, at der i tilfældet med prochloraz må være tale om en ren fysiologisk påvirkning, da planterne dyrkes under sterile forhold. 9 Overordnet diskussion
I indledningen stillede vi spørgsmålet: "Påvirker pesticider i vandmiljøet væksten af akvatiske planter og alger? Overordnet set mener vi, at tilstedeværelsen af pesticider i de koncentrationer, de er blevet målt i overfladevand i Danmark, næppe vil lægge vandløbsstrækninger øde, endsige ændre sammensætningen af den neddykkede vegetation markant. I hvert fald ikke, hvis effekten af stofferne vurderes enkeltvis for hvert pesticid. Ændringer i udbredelse og artssammensætning af neddykkede vandplanter vil være betydeligt mere påvirket af faktorer som fysiske vækstforhold, eutroficiering og grødeskæring (Baattrup-Pedersen et al., 2003; Baattrup-Pedersen et al., 2002; Riis et al, 2000; Sand-Jensen et al., 2000). Der er dog visse stoffer og situationer, der kan være problematiske og derfor kræver særlig opmærksomhed. 9.1 Problematiske stoffer og pulseksponeringerDe stoffer der er tilladte i Danmark, som kræver særlig opmærksomhed er: Aclonifen, Terbuthylazin, Diquat bromid og Prosulfucarb. For disse stoffer er effekterne på vandmiljøet velundersøgte, og en sprøjtegrænse på 10-20 meter fra nærmeste sø eller vandløb er påkrævet for at minimere afdrift og direkte afstrømning (http://www.lr.dk/middeldatabasen/MiddelValg.asp, November 2003). Sprøjteafstande påvirker dog ikke hyppigheden af gårdbidrag fra håndteringen af herbiciderne. Tilførsel via dræn er sandsynligvis også mindre afhængig af sprøjteafstandene og kan være årsag til, at man på trods af de relativt store sprøjteafstande finder terbuthylazin i overfladevandet, ligesom man i mange år har fundet dets søster-produkt atrazin (Løkke, 2000). Dette er problematisk, da s-triazinerne netop er nogle af de herbicider, der har stor akvatisk toksisitet. På den anden side, er s-trazinerne og andre fotosyntesehæmmere tilsyneladende også en stoftype, hvis skader planterne forholdsvis hurtigt kommer sig over (Solmon et al., 1996; Kersting & Van Wijngaarden, 1999; Van Geest et al., 1999). Der er derfor ikke nogen tegn på, at de stofkoncentrationer man har målt, kombineret med deres forekomst over tid, vil påvirke væksten af den akvatiske vegetation alvorligt. Det er dog en gruppe stoffer, vi mener man bør holde øje med. En gruppe stoffer, der kunne være problematisk, er mini-midlerne (ALS-inhibitore: sulfonylurea og imidazolioner), hvoraf nogle har et stort nedsivnings-potentiale (Black et al., 1999; Bossi et al., 1998; Günther et al., 1993). De måles sjældent, da de udbringes i lave doser, og koncentrationerne i vandmiljøet derfor må forventes at være tilsvarende lave (Sørensen et al., 2003) (Man har i en enkelt af prøverne fra NOVA målt en metsulfuron-methyl koncentration på 0,014 μg l-1). Men nogle mini-midler kan, som forsøgene med metsulfuron-methyl viste, være potentielt toksiske helt nede i ikke målbare koncentrationer. Vurderer man deres toksisitet i forhold til markdosis, er gruppen af mini-midler dog generelt ikke specielt toksisk. Til gengæld er effekten af selv kortvarige pulse af mini-midlerne imazamox og metsulfuron-methyl meget effektive i forhold til tilsvarende korte pulse af fotosyntesehæmmerne bentazon og terbuthylazin. Det er sandsynligt, at de højeste pesticidkoncentrationer vil stamme fra gårdbidrag (Frede et al., 1998, Styczen et al., 2003). Gårdbidrag, typisk fra vaskepladser, har i tyske undersøgelser vist sig at kunne forklare op til 90% af den mængde pesticider man finder i overfladevand (Frede et al., 1998). Og selvom landbrugspraksis i Danmark er forskellig fra den i Tyskland, hvor gårdene er forbundet til et fælles rensningsanlæg, der lukker spildevand ud i åerne, kan det ikke udelukkes, at gårdbidrag også i Danmark er af stor betydning. Da gårdbidrag typisk vil være kortvarige og ikke nødvendigvis blive opfanget af moniteringsprogrammerne, kan puls-eksponering meget vel være den eksponeringsform, der kan have den største umiddelbare effekt. Og her er mini-midlerne sammen med de to testede hæmmere af mikrotubuli-dannelsen, propyzamid og pendimethalin potente. En tre timers puls med metsulfuron-methyl koncentrationer der stoppede plantevæksten uden at slå planten ihjel, virkede i ca. 4 dage på Lemna minor, hvorefter planterne genoptog vækstrater parallelle til kontrolplanterne. Hvorvidt regenereringen vil være lige så hurtig blandt andre arter er uvist, ligesom sammenhængen mellem længden og koncentrationen på en puls heller ikke kendes. Frekvensen af kortvarige pulse er svær at monitere. Men hvis man antager, at de største herbicid-pulse korrelerer med kraftige nedbørshændelser, hvilket gælder for udvaskning og afstrømning, er det ikke sandsynligt, at der vil forekomme kraftige pulse med under 4 dages mellemrum ( Liess et al., 1999; Solomon et al., 1996). Frekvensen for gårdbidrag kendes ikke. 9.2 KombinationseffekterNogle af de situationer, som dette studie også peger på, kan være kritiske, er de situationer hvor flere pesticider optræder samtidig. NOVA-dataene viser med al tydelighed, at problematikken er meget aktuel, når der i over halvdelen af de prøver, hvor der kan måles pesticider, findes mere end syv og op til 18 forskellige stoffer. Der findes modeller, der kan estimere effekten af blandinger, hvis man kender toksisiteten af de individuelle komponenter (Cedergreen et al., 2003a; Backhaus et al., 2000; Faust et al., 2001; Junghans et al., 2003), men det er sjældent gjort. I et eksempel, der tog udgangspunkt i danske målinger af herbicider fra en puls-måling, viste det sig, at den kummulerede effekt af 3 herbicider kunne udøve en potentiel effekt på 8-22% på vandplanter. Sandsynligheden for at dette rent faktisk ville ske blev dog vurderet at være lille, da der var tale om en pulseksponering af primært fotosyntesehæmmere. Tyske undersøgelser har imidlertid vist, at 16-18 stoffer i koncentrationer, som enkeltvis ikke ville sænke væksten af alger med mere end 1%, sammenlagt kan give en effekt på 18-47% (Faust et al., 2001; Faust et al., 2003). Så kombinationseffekter under danske forhold kan bestemt ikke udelukkes. 9.3 Validitet af toksikologiske dataEn af forudsætningerne for at kunne ekstrapolere fra toksisitets-data på standard testorganismer til miljøet er, at testorganismerne opfører sig repræsentativt i forhold til den gruppe, de menes at repræsentere. Vores test af repræsentabiliteten af de to standard test-systemer: Lemna sp. og algen Pseudokirchneriella subcapitata i forhold til 10-12 vandplantearter og et naturligt epifytsamfund overfor herbiciderne metsulfuron-methyl og terbuthylazin, viste sig at være god. Hvis man dividerer med en faktor 10 på NOEC (defineret som EC10) eller en faktor 100 på EC50, og bruger disse koncentrationer som maksimalt tilladelige grænseværdier, vil man beskytte over 95% af vandplante-arterne og det samlede epifytsamfund. Man har fået tilsvarende resultater for algesamfund og mesokosmos forsøg, hvor det dog for algesamfundets vedkommende blev understreget, at de strukturelle parametre, så som ændringer i artssammensætning, var mere følsomme end funktionelle parametre som brutto fotosynteserater (Selck et al., 2002; Van Geest et al., 1999). Man kom ikke til samme konklusion i mesokosmos forsøgene, hvor fotosyntesehæmmeren Linuron blev testet (Van Geest et al., 1999; Kersting & Van Wijngaarden, 1999). Men som dette projekts forsøg også viser, varierer følsomheden overfor et giftstof ikke kun på tværs af arter, men er også afhængigt af, hvilken parameter man måler på, og hvilket stof planterne er udsat for. Det er derfor ikke overraskende, at for en fotosyntesehæmmer som Linuron vil den funktionelle parameter fotosyntese være en følsom effekt variabel, hvorimod tributyltin (TBT) og linear alkylbenzene sulfonate (LAS), hvis virkning er mindre specifik, kan virke mere artsspecifikt. Dette stemmer overens med resultaterne fra mesocosmosforsøg, der netop konkluderer, at de mest følsomme effektvariable for fotosyntesehæmmere er funktionelle, hvorimod de for vækst-hæmmere synes at være strukturelle (Brock et al., 2002). I dette projekts forsøg var forskellen mellem effekt variable størst for de morfologiske parametre hos vandplanter udsat for metsulfuron-methyl. Her sås en kraftig effekt på f.eks. roddannelse og skudvækst ved koncentrationer, hvor der indenfor forsøgstiden endnu ikke kunne ses effekter på den tørvægtsspecifikke vækstrate. De samme resultater blev fundet af Wendt-Rasch et al. (2003) for metsulfuron-methyl i felt- og laboratorieforsøg ved tilsvarende koncentrationer og for en lang række herbicider af Turgut &Fomin (2002). Variationen imellem effektvariable synes generelt at ligge indenfor en faktor 10-50 (Wendt-Rasch et al., Turgut & Fomin, 2002), så hvis man ikke har mulighed for at måle på mere end en effektvariabel, må man anbefale en sikkerhedsfaktor af den størrelsesorden. Forskellene i følsomheden af forskellige effektvariable kan til dels skyldes tidspunktet for måling. Netop for metsulfuron-methyl ved man, at celledelingen stopper få timer efter eksponering (Cobb, 1992), hvorfor man må forvente en hurtig effekt på netop dannelsen af nye rødder og skud. Hvis man ventede endnu et par uger, er det ikke usandsynligt, at fotosyntesen og væksten ville vise sig at være tilsvarende følsom. Forsøg med Lemna minor, hvor forskellige effektvariable blev målt over tid, viste, at for denne plante blev den fulde følsomhed af metsulfuron-methyl og terbuthylazin nået indenfor standard test-tiden på 7 dage. Det er dog ikke sikkert at det samme gælder for de mere langsomt voksende neddykkede vandplanter, testet i forsøgene med flere arter og hos Wendt-Rasch et al. (2003) og Turgut & Fomin (2002). 9.4 Forskelle mellem vandplanter og alger samt stimulerende effekterFor de herbicider, der er svage syrer med en pKa på under 5, var der stor forskel på toksisiteten mellem Lemna og algen P. subcapitata. For metsulfuron-methyl, der blev testet på 12 arter af vandplanter og et naturligt epifyt-samfund, blev den samme forskel mellem vandplanter generelt og epifyt-samfundet også fundet (Cedergreen et al., 2003b). Det er nærliggende at relatere forskellen, der var op til en faktor 1000, til forskellen i andelen af dissocierede molekyler ved den pH på henholdsvis pH 5 og pH 8, der bliver brugt i de to standard-forsøg. Forsøg med alger har nemlig vist, at det primært er stoffer på den uladede form, der optages (Fahl et al., 1995; Mayer et al., 1998), og denne andel vil netop være omkring en faktor 1000 større ved pH 5 sammenlignet med pH 8. Dette synes dog ikke umiddelbart at være tilfældet for Lemna, da forsøg med Lemna ved pH 7,5 – 8,5 viste en tilsvarende toksisitet som forsøg ved pH 5. Dette kan skyldes at Lemna er betydeligt mere følsom overfor svage syrer end P. subcapitata. En anden hypotese kunne være, at pH i apoplasten hos Lemna minor og andre højere planter er sur, ca. pH5-6, sådan som det er tilfældet for størstedelen af terrestriske planter (Salisbury & Ross, 1985). De enkelte celler hos Lemna vil i så tilfælde altid opleve det omgivende miljø som surt, uanset den eksterne pH. Store forskelle i følsomhed mellem højere planter og alger vil kunne forskubbe balancen mellem de to plantegrupper. Tilsvarende effekt ser man, når eutroficieringen af vandfasen forøges. Dette fandt man i mesokosmosforsøget hos Wendt-Rasch et al. (2002), hvor algevæksten øgedes ved de lave metsulfuron-methyl koncentrationer. Dette foreslog de kunne skyldes en øget næringsstoffrigivelse fra planter, der var påvirket af herbicidet. Det kunne imidlertid også skyldes en hormetisk effekt, d.v.s. en vækststimulering ved lave herbicid koncentrationer, sådan som det ofte er observeret for bl.a. metsulfuron-methyl (Wendt-Rasch et al., 2003, figur 8) samt i nærværende projekt. En vækststimulering forårsaget af lave doser af herbicider, der for produceret bladlængde hos byg og Sparganium var på 50-100%, kunne ændre konkurenceforholdene mellem arterne. Så for de lave herbicid koncentrationer, der måles i dansk overfladevand, kan effekter af vækststimulering måske spille en lige så stor eller større rolle for artssammensætningen, som effekter der nedsætter væksten hos planter og alger. 10 KonklusionGenerelt må det konkluderes, at de pesticid-koncentrationer, der er målt i det danske overfladevand, ikke udgør nogen umiddelbar alvorlig fare for væksten eller artssammensætningen af neddykkede vandplanter og alger i de danske søer og vandløb. I hvert fald ikke når effekten vurderes for stofferne enkeltvis. Der er dog få grupper af herbicider, der kan være problematiske. Blandt disse er s-trazinerne, der er meget potente i vand og som tillige findes i relativ høje koncentrationer (> 1 μg l-1). Hvis man skulle finde nogle effekter af herbicider på neddykkede vandplanter, er det derfor sandsynligt, at de skal findes indenfor denne gruppe. Det er imidlertid vist, at planter kommer sig meget hurtigt efter skader af fotosyntesehæmmere som s-triaziner, så en mulig skade bliver derfor næppe langvarig. En anden gruppe er mini-midlerne, hvoraf nogle har vist at have en effekt ved lave koncentrationer. En af disse er sulfonyl-urea herbicidet metsulfuron-methyl. Den artsspecifikke følsomhed overfor metsulfuron-methyl og s-trazinen terbuthylazin viste sig at være lille sammenlignet med diversiteten i det terrestriske miljø. De mest anvendte testplanter, flydebladsplanten Lemna sp. og algen Pseudokirchneriella subcapitata var repræsentative for et bredere spekter af vandplanter og alger. Anvender man derfor de anbefalede sikkerhedsfaktorer på 10 mellem de højeste tilladte koncentrationer og NOEC (EC10) og en faktor 100 mellem de højeste tilladte koncentrationer og EC50 på Lemna, vil man kunne beskytte over 95% af arterne i vandplantesamfundene. Der var stor forskel imellem effekten af en pulseksponering afhængigt af herbicidets virkemåde, men selv med de mest effektive kom Lemna minor sig efter fire dage efter en høj, men ikke dødelig dosis. Hvis andre arter kommer sig lige så hurtigt som Lemna, er det vores vurdering, at høje koncentrationer fra f.eks. en nedbørshændelse umiddelbart efter sprøjtning kun vil have en kortvarig effekt. En af de situationer hvor pesticider på trods af deres lave individuelle koncentrationer, alligevel kunne have en effekt, er når der optræder mange sammen på samme tid, hvilket ofte vil ske i forbindelse med f.eks. nedbørshændelser. Disse situationer bør evalueres nærmere. Overordnet set er konklusionen dog, at de niveauer af pesticider, der findes i det danske vandmiljø, kan anses for at være relativt uvirksomme overfor neddykkede vandplanter og alger. Effekter af brugen af herbicider på planter udenfor målgruppen skal derfor snarere findes i det terrestriske miljø, hvor effektiviteten af stofferne synes at være betydelig større. 11 PerspektiveringI løbet af projektforløbet er der fremkommet flere problemstillinger, som vi mener det kunne være interessant at undersøge nærmere, hvis man ønsker at få en større forståelse af virkningen af herbicider i det akvatiske miljø. Disse er:
12 ReferencerAltenburger R, Nenza M, Schüürmann G. (2003). Mixture toxicity and its modeling by quantitative structure-activity relationships. Environmental toxicology and Chemistry 22: 1900-1915. Appleby AP. (2001). The practical implications of hormetic effects of herbicides on plants. Belle Newsletter 011001: 1-3. Arensberg P, Hemmingsen VH, Nyholm N. (1995). A miniscale algal toxicity test. Chemosphere 30: 2103-2115. Asman W, Jørgensen A, Jensen PK. (2003). Dry deposition and spray drift of pesticides to nearby water bodies. 66, 1-171. Copenhagen, Danish Environmental Protection Agency. Pesticide Research. Baattrup-Pedersen A. (2000). Planter i vandløb -fortid, nutid og fremtid. 34/2000, 1-33. 2000. Silkeborg. Baattrup-Pedersen A, Larsen SE, Riis T. (2002). Long-term effects of stream management on plant communities in two Dansih lowland streams. Hydrobiologia 481: 33-45. Baattrup-Pedersen A, Larsen SE, Riis T. (2003). Composition and richmess of macrophyte communities in small Dansih streams - Influence of environmental factors and weed cutting. Hydrobiologia 495: 171-179. Backhaus T, Altenburger R, Boedeker W, Faust M, Scholze M, Grimme LH. (2000). Predictability of the toxicity of multiple mixture of dissimilarly acting chemicals to Vibrio Fischeri. Environmental toxicology and Chemistry 19: 2348- 2356. Berenbaum MC. (1989). What is synergy? Pharmacological Reviews 41: 93- 141. Black ID, Pederson RN, Flynn A, Moerkerk M, Dyson CB, Kookana R, Wilhelm N. (1999). Mobility and persistence of three sulfonylurea herbicides in alkaline cropping soils of south-eastern Australia. Australian Journal of Experimental Agriculture 39: 465-472. Bossi R, Koppen B, Spliid NH, Streibig JC. (1998). Analysis of sulfonylurea herbicides in soil water at sub-pert-per billion levels by electrospray negative ionization mass spectrometry followed by confirmatory tandem mass spectrometry. Journal of AOAC International 81. Boutin C, Lee HB, Peart ET, Batchelor PS, Maguire RJ. (2000). Effects of the sulfonylurea herbicide metsulfuron methyl on growth and reproduction of five wetland and terrestrial plant species. Environmental toxicology and Chemistry 19: 2532-2541. Boutsalis P. (2000). The World of Herbicides according to HRAC classification on mode of action. 1. England, AD+AM Design and Marketing. Map Brock TCM, Blake N, Maltby L, Van den Brink P. (2003). Linking laboratory derived species sensitivity distributions to treshold levels of pesticides observed in experimental freshwater ecosystems. In:Solutions to Pollution: Programme abstract book. Christchurch, New Zealand: The Socoety of Environmental Toxicology and Chemistry Asia/Pacific - The Australian Society of Ecotoxicology, 149. Brock TCM, Lahr J, van den Brink PJ. (2002). Ecological risks of pesticides in freshwater ecosystems. ISSN 1566-7197, 1-103. Wageningen, Alterra. Bruce RD, Versteeg DJ. (1992). A statistical procedure for modeling continuous toxicity data. Environmental toxicology and Chemistry 11: 1494. Calabrese EJ. (2001). Overcompensation stimulation: A mechanism for hormetic effects. Critical Reviews in Toxicology 31: 425-470. Calabrese EJ, Baldwin LA. (2001). Hormesis: A generalizable and unifying hypothesis. Critical Reviews in Toxicology 31: 353-424. Cedergreen N, Streibig JC, Kudsk P, Møhlenberg F, Kamper A. (2003a). Er det synergi, når 0+0=1? Vand og Jord 10: 88-91. Cedergreen N, Streibig JC, Spliid NH. (2003b). Sensitivity of Aquatic plants to the herbicide metsulfuron-methyl. Ecotoxicology and Environmental Safety In press. Chapman PM, Caldwell RS. (1996). A warning: NOECs are inapropriate for regulatory use. Environmental toxicology and Chemistry 5: 77-79. Christensen ER, Chen D, Nyholm N, Kusk OK. (2001). Joint action of chemicals in algal toxicity tests: Influence of response level adm dose response regression model. Environmental toxicology and Chemistry 20: 2361-2369. Cobb A. (1992). The inhibition of amino acid biosynthesis. In:Herbicides and Plant Physiology. London: Chapman & Hall, 126-143. Devine MD, Duke SO, Fedtke C. (1993). An introduction to herbicide action. In: Devine MD, Duke SO, Fedtke C, eds. Physiolohy of herbicide action. New Jersey: P T R Prentice-Hall, Inc., 1-13. ECETOC. 1998. QSARs in the assessment of the environmental fate and the effects of chemicals. 74, 1-136. (1998). Brussels, European Centre for Ecotoxicology and Toxicology of Chemicals. Technical Report. Fahl GM, Kreft L, Altenburger R, Faust M, Boedeker W, Grimme LH. (1995). pH-dependent sorption, bioconcentration and algal toxicity of sulfunylurea herbicides. Aquatic Toxicology 31: 175-187. Faust M, Altenburger R, Backhaus T, Blanck H, Boedeker W, Gramatica P, Hamer V, Scholze M, Vighi M, Grimme LH. (2001). Predicting the joint algal toxicity of multi-component s-triazine mixtures at low-effect concentrations of individual toxicants. Aquatic Toxicology 56: 13-32. Faust M, Altenburger R, Backhaus T, Blanck H, Boedeker W, Gramatica P, Hamer V, Scholze M, Vighi M, Grimme LH. (2003). Joint algal toxicity of 16 dissimilarly acting chemicals is predictable by the concept of independent action. Aquatic Toxicology 63: 43-63. Faust M, Altenburger R, Boedeker W, Grimme LH. (1994). Algal toxicity of binary combinations of pesticides. Bull.Environ.Contam.Toxicol. 53: 134-141. Faust M, Scholze M. (2004). Competing concepts for the prediction of mixture toxicity: Do the difference matter for regulatory purposes? EVKICT1999- 00012, 2-62. Fedtke C. (1982b). Photosynthesis. In:Biochemistry and physiology of herbicide action. Berlin: Springer-Verlag, 19-113. Fiebig S, McVean K, Makus P, Noack U. (2003). Non-target plant testing: Variation of plant growth in relation to the plant species. SETAC Europe 13th Annual Meeting, Hamburg, Germany, 27 April - 1 May Understanding the complexity of environmental issues - A way to sustainability, 118. Frede H-G, Fischer P, Bach M. (1998). Reduction of herbicide contamination in flowing waters. Zeitschrift für Planzenernährung und Bodenkunde 161: 395-400. Frost-Christensen H, Jørgensen LB, Floto F. (2003). Species specificity of resistance to oxygen diffusion in thin cuticular membranes from amphibious plants. Plant, Cell and Environment 26: 561-569. Gerhard M, Habermeyer J, Zinkernagel V. (1998). The impact of strobilurins on plant vitality on winter wheat under field conditions. In: Lyr H, Russell PE, Dehne H-W, Sisler HD, eds. Modern fungicides and antifungal compounds II. HANTS: Intercept Limited, 197-208. Gessner PK. (1995). Isobolographic analysis of interactions: an update on applications and utility. Toxicology 105: 161-179. Greco WR, Bravo G, Parsons JC. (1995). The search for synergy: A critical review from a response surface perspektive. Pharmacological Reviews 47: 332- 385. Günther P, Pestemer W, Rahman A, Nordmeyer H. (1993). A bioassay technique to study the leaching behaviour of sulfunylurea herbicides in different soils. Weed Research 33: 177-185. Hoekstra JA, Van Ewijk PH. (1993). Alternatives for the No-Observed-Effect level. Environmental toxicology and Chemistry 12: 187-194. International Organization for standardization. (1989). Water quality - Fresh water algal growth inhibition test with Scenedesmus subspicatus and Selenastrum capricornutum. In: Geneva: International Organization for standardization. Junghans M, Backhaus T, Faust M, Scholze M, Grimme LH. (2003). Predictability of combined effects of eight chloroacetanilide herbicides on algal reproduction. Pest Management Science In press. Kersting K, Van Wijngaarden RPA. (1999). Effects of a pulsed treatment with the herbicide afalon (Active ingredient linuron) on macrophytedominated mesocosms. I. Responses of ecosystem metabolism. Environmental toxicology and Chemistry 18: 2859-2865. Kirkwood RC. (1999). Recent developments in our understanding of the plant cuticle as a barrier to the foliar uptake of pesticides. Pesticide Science 55: 69-77. Klaine SJ, Cobb GP, Dickerson RL, Smith E.E., Dixon KR, Kendall RJ, Solomon KR. (1996). An ecological risk assessment for the use of the biocide, dibromonitrilopropionamide (DBNPA), in industrial cooling systems. Environmental toxicology and Chemistry 15: 21-30. Kloepper-Sams P, Beach S, Dyer S, Guhl W, McCahon P, Marshall S, Peterson D, Stephenson R, Holt M. (2001). Aquatic Toxicity of Mixtures. 80, 1-64. Brussels, European Centre for Ecotoxicology and Toxicology of Chemicals. ECETOC technical Reports. Knuuttila P, Knuuttila H. (1979). The crystal and molecular structure of N- (phosphonomethyl)-glysine (Glyphosate). Acta Chemica Scandinavica B 33: 623-626. Kooijman SALM. 1996. An alternative to NOEC exists, but the standard model has to be abandoned first. OIKOS 75: 310-316. Køppen B, Spliid NH. (1998). Determination of acid herbicides using liquid chromatography with pneumatically assisted electrospray ionization mass spectromtric and tandem spectrophotometric detection. Journal of Chromatography A 803: 157-168. Kudsk P. (1993). Aktivstoffers virkemåde og virkningsbetingelser. In: Wagn O, Graugaard S, eds. Ukrudsbekæmpelse i Landbruget. Copenhagen: Statens Planteavlsforsøg, 229-254. Landolt E, Kandeler R. (1987). Physiological characteristics. In: Landolt E, Kandeler R, eds. The family of Lemnaceae - a monographic study. Zürich: Stiftung Rübel, 54-113. Liess M, Schulz R, Liess MHD, Rother B, Kreuzig R. (1999). Determination of insecticide contamination in agricultural headwater streams. Water Research 33: 239-247. Løkke, H. (2000). Bichel-udvalget, Rapport fra underudvalget om miljø og sundhed. 1-83. Miljøstyrelsen. Maeng J, Khudairi AK. (1973). Studies on the flowering mechanism of Lemna I. Amino acid changes during flower induction. Physiologia Plantarum 28: 264-270. Martin JD, Crawford CG, Larson SJ. (2003). Pesticides in streams. 1-5. National Water Assessment Program (NAWQA). Mayer P, Cuhel R, Nyholm N. (1997). A simple in vitro fluorescence method for biomass measurements in algal growth inhibition tests. Water Research 31: 2525-2561. Mayer P, Frickmann J, Christensen ER, Nyholm N. (1998). Influence of growth conditions on the results obtained in algal toxicity tests. Environmental toxicology and Chemistry 17: 1091-1098. Miljøstyrelsen. (1999). Rammer for vurdering af plantebeskyttelsesmidler. 7016-0002, 1-53. 1999. København, Miljøstyrelsen. Miljøstyrelsen. (2003) Bekæmpelsesmiddelstatistik 2002. 1-48. København, Miljøstyrelsen. Mogensen BB, Spliid NH, Helweg A. (1999). Bekæmpelsesmidler - anvendelse og spredning i miljøet. 26/1999, 1-47. Roskilde, Miljø- og Energiministeriet, Danmarks Miljøundersøgelser. Møhlenberg F, Schlüter L, Gustavson K, Andersen TT, Cold A, Forbes VE, Friberg N. (2004). Effekt af bekæmpelsesmidler på flora og fauna i vandløb. Copenhagen, Danish Environmental Protection Agency. Pesticide Research. Nyholm N. (1990). Expression of results from growth inhibition toxicity tests with algae. Archives of Environmental Contamination and Toxicology 19: 518- 522. Nyström B, Björnsäter B, Blanck H. (1999). Effects of sulfunylurea herbicides on non-target aquatic micro-organisms. Growth inhibition of microalgae and short-term inhibition of adenine and thymine incorporation in periphyton communities. Aquatic Toxicology 47: 9-22. OECD guideline for testing of chemicals (2000). Lemna sp. growth inhibition test. 1-24. Osmond CB. (1994). What is photoinhibition? Some insight from comparisons of shade and sun plants. In: Baker NR, Bowyer JR, eds. Photoinhibition of photosynthesis, from molecular macanisms to the field. Oxford: Bios Scientific Publishers Limited, 1-24. Riis T, Sand-Jensen K, Vestergaard O. (2000). Plant communities in lowland Danish streams: species composition and environmental factors. Aquatic Botany 66: 255-272. Royuela M, Gonzalez A, Gonzalez EM, Arrese-Igor C, Aparicio-Tejo PM, Gonzalez-Murua C. (2000). Physiological consequences of continuous, sublethal imazethapyr supply to pea plants. Journal of Plant physiology 147: 345-354. Sabadie J. (2000). Behavior of four sulfonylurea herbicides in the presence of hydroxy compounds. Journal of Agriculture and Food Chemistry 48: 4752- 4756. Salisbury FB, Ross CW. (1985). Plant Physiology. Belmont: Wadsworth Publishing Company. Sand-Jensen K. (1997). Macrophytes as biological engineers in the ecology of Danish streams. In: Sand-Jensen K, Pedersen O, eds. Freshwater Biology. Copenhagen: Gad, 74-101. Sand-Jensen K, Riis T, Vestergaard O, Larsen SE. (2000). Macrophyte decline in Danish lakes and streams over the past 100 years. Journal of Ecology 88: 1030-1040. Sand-Jensen K. (2001) Søer - en beskyttet naturtype. Gads Forlag, København. Seefeldt SS, Jensen JE, Fuerst E. (1995). Log-logistic analysis of herbicide dose-response relationships. Weed Technology 9: 218-227. Selck H, Riemann B, Forbes VE, Christoffersen K, Gustavson K, Hansen BW, Jacobsen JA, Kusk OK, Petersen S. (2002). Comparing sensitivity of ecotoxicological effect endpoints between laboratory and field. Ecotoxicology and Environmental Safety 52: 97-112. Smart LE, Barko JW. (1985). Laboratory culture of submersed freshwater macrophytes on natural sediments. Aquatic Botany 21: 251-263. Solomon KR, Baker DB, Richards RP, Dixon DR, Klaine SJ, LaPoint TW, Kendall RJ, Weisskopf CP, Giddings J.M., Giesy JP, Hall LW, Williams WM. (1996). Ecological risk assessment of atrazine in North American surface waters. Environmental toxicology and Chemistry 15: 31-74. Sørensen PB, Brüggemann R, Carlsen L, Mogensen BB, Kreuger J, Pudenz S. (2003). Analysis of monitoring data of pesticide residues in surface waters using partial order ranking theory. Environmental toxicology and Chemistry 22: 661-670. Stahlman PW, Phillips WM. (1979). Effects of water quality and spray volumen on glyphosate phytotoxicity. Weed Science 27: 38-41. Stebbing ARD. (1997). A theory for growth hormesis. Belle Newsletter 6: 1- 11. Streibig JC, Rudemo M, Jensen JE. (1993). Dose-response curves and statistical models. In: Streibig JC, Kudsk P, eds. Herbicide Bioassay. Boca Raton: CRC Press, 29-55. Streibig JC, Walker A, Blair AM, Anderson-Taylor G, Eagle DJ, Friedländer H, Hacker E, Iwanzik W, Kudsk P, Labhart C, Luscomber BM, Madafiglio G, Nel PC, Pestemer W, Rahman A, Retzlaff G, Rola J, Stefanovic L, Straathof HJM, Thies EP. (1995). Variability of bioassays with metsulfuronmethyl in soil. Weed Research 35: 215-224. Styczen M, Wiberg-Larsen P, Aagaard A. (2003). Tag pulsen på pesticiderne i vandmiljøet. Vand og Jord 10: 84-87. Thelen KD, Jackson EP, Penner D. (1995). The basis for hard water antagonism of glyphosate activity. Weed Science 43: 541-548. Tomlin CDS. (2002). The e-Pesticide Manual. Tomlin, C. D. S. [2.2]. 2002. Surrey, British Crop Protection Council. Turgut C, Fomin A. (2002). Sensitivity of the rooted macrophyte Myriophyllum aquaticum (Vell.) Verdcourt to seventeen pesticides determined on the basis of EC50. Bulletin of Environmental Contamination and Toxicology 69: 601-608. Ullrich WR, Larsson M, Larsson CM, Lesch S, Novacky A. (1984). Ammonium uptake in Lemna gibba G 1, related membrane potential changes, and inhibition of anion uptake. Physiologia Plantarum 61: 369-376. Van Geest GJ, Zwaardemaker ND, Van Wijngaarden RPA, Cuppen JGM. (1999). Effects of a pulsed treatment with the herbicide afalon (Active ingredient linuron) on macrophyte-dominated mesocosms. II. Structural responses. Environmental toxicology and Chemistry 18: 2866-2874. van Straalen NM. (2002). Treshold models for species sensitivity distributions applied to aquatic risk assessment for zinc. Environmental Toxicology and Pharmacology 11: 167-172. Vestergaard A. (2003). Light acclimation in freshwater macrophytes -with emphasis on photoinhibition., Department of Plant Ecology, Institute of Biological Sciences, Aarhus University. Walker CH, Hopkin SP, Sibly RM, Peakall DB. (2001). Toxicity testing. In: Walker CH, ed. Principles of Ecotoxicology. London: Taylor & Francis, 107- 130. Walter H, Consolaro F, Gramatica P, Scholze M, Altenburger R. (2002). Mixture toxicity of priority pollutants at No Observed Effect Concentrations (NOECs). Ecotoxicology 11: 299-310. Wendt-Rasch L, Pirzadeh P, Woin P. (2003). Effects of metsulfuron methyl and cypermethrin exposure on freshwater model ecosystems. Aquatic Toxicology 63: 243-256. Wiedman SJ, Appleby AP. (1972). Plant growth stimulation by sublethal concentrations of herbicides. Weed Research 12: 65-74. Windolf J, Svendsen LM, Kronvang B, Skriver J, Oversen NB, Larsen SE, Baattrup-Pedersen A, Iversen HL, Erfurt J, Müller-Wohlfeil D, Jensen JP. (1996). Ferske Vandområder - Vandløb og kilder. Vandmiljøplanens overvågningsprogram 1996. Windolf, J. 214, 1-112. 1996. Silkeborg, Danmarks Miljøundersøgelser. Faglig rapport fra DMU nr. 214. Wintermans JFGM, De Mots A. (1965). Spectrophotometric characteristics of chloraphylls a and b and their phenophytins in ethanol. Biochimica et Biophysica Acta (BBA) 109: 448-453. Wright DA, Welbourn P. (2002). Factors affecting toxicity. In: Wright DA, Welbourn P, eds. Environmental Toxicology. Cambridge: University Press, 218-248.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||