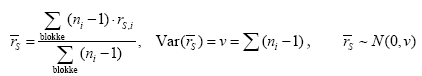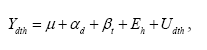|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 92, 2004 Effekten af sprøjtemiddelafdrift på buske og træer i læhegn- Bær som indikator for biodiversitetsforandringerIndholdsfortegnelse
Bilag A: Forhold under forårssprøjtning Bilag B: Forhold under sommersprøjtning Bilag C: Datagrundlaget for eksponeringsåret ForordDenne rapport er slutrapporteringen af projektet " Bærbærende buske og træer som indikatorer for biodiversitetsforandringer ved sprøjtemiddelafdrift II ". Projektet er efterfølger for pilotprojektet " Påvirkes tjørnehegns biodiversitet af afdrift af ukrudtsmidler fra marksprøjtning? – et pilotprojekt" som blev udført i 2001 og afrapporteret i 2002. Projektet har modtaget støtte fra den del af pesticidafgiften, som udgøres af pesticidforskningsmidlerne, der administreres af Miljøstyrelsen (J.nr. MST 7041-0471). Projektet har undervejs været fulgt af følgende følgegruppemedlemmer: Allan Prang, GEUS, Bo Svenning Petersen, Hedeselskabet, Christian Kjær, DMU, Claus Hansen, Miljøstyrelsen, Heidi Buur Holbeck, Dansk Landbrugsrådgivning, Ib Johnsen, Københavns Universitet, Jens Erik Jensen, Dansk Landbrugsrådgivning, Jens Erik Ørum, Fødevareøkonomisk Institut, Jesper Rasmussen, KVL, Knud Tybirk, DMU, Kristian Kristensen, DJF, Lene Gravesen, Miljøstyrelsen, Lise Nistrup Jørgensen, DJF, Marianne Bruus Pedersen, DMU, Mette Wier, Fødevareøkonomisk Institut, Mogens Erlandsen, Århus Universitet, Morten Strandberg, DMU, Niels Lindemark, Dansk Planteværn, Peter Esbjerg, KVL, Peter Odderskær, DMU, Peter Sandøe, KVL, Poul Henning Petersen, Dansk Landbrugsrådgivning, Søren Navntoft, KVL, Tommy Dalgaard, DJF og Jørn Kirkegaard (formand og referent), Miljøstyrelsen. Følgegruppen har undervejs skiftet navn fra "Biodiversitet i Agerlandet" til "Jordbrug og Pesticider". Alle følgegruppemedlemmer takkes hermed for kommentarer til projektet. Udover forfatterne har følgende DMU-medarbejdere bidraget til projektet: Lise Lauridsen, Inger Møller, Trine Guldager Sørensen, Marianne Hilligsøe Pedersen, Susanne Breuner Meldgaard, Lene Birksø Bødskov, Bodil Thestrup og Niels Elmegaard, som alle takkes for deres indsats. Projektet har haft Signe Gammeltoft og Kristian Michaelsen tilknyttet som specialestuderende. Endelig takkes følgende ejere af tjørnehegn for at have medvirket til projektets gennemførelse ved at give tilladelse til at lade deres hegn indgå i projektet: Harm Oenema, Inger Jensen, Inger Strandberg, Jørn Mortensen, Mogens og Jane Kortnum samt Frederiksborg Amt. SammenfatningDenne rapport beskriver effekter af simuleret afdrift af herbicidet metsulfuron til tjørnehegn i eksponeringsåret og året efter. Baggrund Det er kendt, at tætheden og diversiteten af ukrudt, insekter og fugle på markfladerne påvirkes af brugen af sprøjtemidler. Det er forventeligt, at afdrift af herbicider også vil kunne påvirke de træer og buske, der i form af skel eller læhegn findes imellem markerne. Imidlertid er der i dag nationalt og internationalt meget begrænset viden om utilsigtede effekter af sprøjtemidler på træer og buske, både hvad angår direkte effekter, forsinkede effekter og indirekte effekter på den biologi som er knyttet til træer og buske i form af for eksempel insekter og fugle. Al-Khatib et al. 1992, Bhatti et al. 1995 og Fletcher et al. 1996 fandt negative effekter af sulfonylurea-midler og glyfosat på vækst og frugtsætning i fuglekirsebær (Prunus avium) i forsøg, der simulerede herbicidafdrift. I nogle tilfælde blev der observeret negative effekter ved doser, der svarer til 1% af den af producenten anbefalede maksimaldosis. Så vidt vides, er der ikke tidligere lavet tilsvarende undersøgelser af herbicidafdrift på bærbærende buske og træer i Danmark eller Europa. Der er dog næppe tvivl om, at herbicidafdrift kan have stor betydning for de fødekæder, der er knyttet til netop blomstring, frugtsætning og frøspredning. Formål Projektets overordnede formål er at fremskaffe viden om potentielle effekter af herbicider på træer og buske i eksponeringsåret og året efter, eksemplificeret ved herbicidet metsulfuron-methyl og arten engriflet hvidtjørn, Crataegus monogyna. Projektet vil opfylde det overordnede formål ved at besvare følgende spørgsmål:
Design Projektet er designet på baggrund af et pilotprojekt. Pilotprojektet undersøgte variationen i antal af blade, knopper, blomster, grønne bær og modne bær i de enkelte sideskud (Kjær et al. 2002). Statistiske styrkeberegninger og logistiske forhold førte til, at vi inddrog 8 tjørnehegn fordelt med 5 i Jylland og 3 på Sjælland. I hvert hegn blev det besluttet at sprøjte med fire doser på to tidspunkter for at undersøge betydningen af sprøjtetidspunkt. De nominelle doser var 5%, 10%, 20% og 40% af den af producenten angivne maksimaldosis, samt en kontrol. Ved hvert sprøjtetidspunkt blev der for hver dosis sprøjtet 13 træer per hegn. Kontrollerne blev ikke sprøjtet. Aktiviteter Sprøjtning Forårssprøjtning blev gennemført medio maj 2002 under noget varierende vejrforhold. Sommersprøjtning blev gennemført primo juni under mere ensartede vejrforhold. Ved begge sprøjtninger blev sprøjtemidlet tilsat glycin, hvis afsætning på bladene blev bestemt ved indsamling af blade umiddelbart efter sprøjtning. Afsætning af glycin blev derefter anvendt til omregning fra nomi-nel dosis til afsat dosis. Dataindsamling 1.år Efter forårssprøjtningen blev der indsamlet blade, blomster, grønne bær og modne bær. Efter sommersprøjtningen blev der indsamlet blade, grønne bær og modne bær. Ved alle prøver blev antallet af den enkelte målvariabel bestemt, og for alle målvariable undtagen blomster blev tørvægten bestemt efter frysetørring. Nedfaldne knopper, blomster, frugtanlæg og bær blev indsamlet i spande under træerne for at bestemme sæsonvariationen i nedfaldet og hvilket reproduktivt stadie, der er mest udsat for tab som følge af herbivori og abortering. 2. år For begge sprøjtninger blev alle målvariable indsamlet, dvs. blade, knopper, blomster, grønne bær og modne bær. I det andet år blev tilfældigt udvalgte træer i den del af hegn 5, der ikke indgik i herbicideksperimentet, sprøjtet med insekticidet cypermethrin med det formål at fjerne effekten af herbivore insekter. Betydningen heraf blev opgjort ved at tælle mængden af knopper, blomster og bær på i forvejen udvalgte skud. Bærmængden blev yderligere opgjort i rammer. Resultater Sprøjteeffekter i eksponeringsåret:
Sprøjteeffekter året efter eksponering:
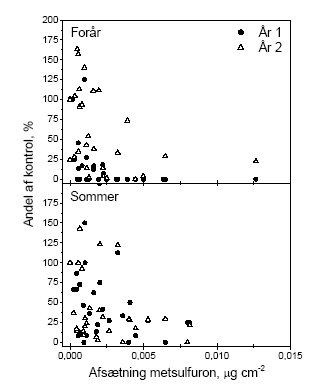
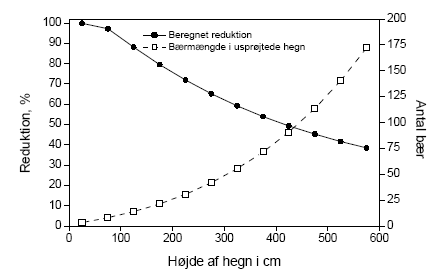
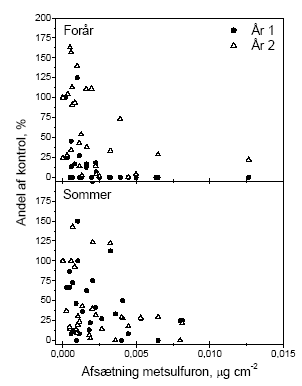
Figur 1. Sammenligning af den procentuelle effekt på bærmængden af metsulfuronsprøjtning i eksponeringsåret og året efter. Data benyttet er antallet af bær i sideskud i år 1 og antal bær i en ramme i år 2 . Biologiske observationer:
Resultaternes relevans i forhold til regulering Reguleringen af pesticider tager for nærværende ikke hensyn til nogle af de effekter, der er påvist eller sandsynliggjort i dette projekt. Det gælder det forhold at flerårige planter kan være påvirket ud over det år, hvor de har været udsat for sprøjtemidlet. Dernæst inddrager de testmetoder, der bruges ved vurderingen af sprøjtemidler, ikke mulige effekter på frugtsætning. Resultaterne taler for at undersøgelser af effekter på frugtsætning og undersøgelser, der inddrager flerårige planter er tilrådelig udfra et naturbeskyttelseshensyn. SummaryIn this report, effects of simulated herbicide spray drift on hedgerows consisting of hawthorn were studied. Observations were made both within the spraying season, as well as in the following year. Background It is known that the density and diversity of weeds, insects and birds in the field are affected by the use of pesticides. It is furthermore expected that herbicide spray drift might affect trees and shrubs forming the property line or hedgerows between fields. However, there is only limited knowledge of the unintended effects of pesticides on trees and shrubs, both nationally and internationally. This includes direct toxic effects as well as indirect effects on the organisms associated with the trees and shrubs such as insects and birds. Experiments, which simulated herbicide spray drift, all found negative effects from sulfonylurea-herbicides and from glyphosate on the growth and fruit set of bird cherry (Prunus avium) (Al-Khatib et al. 1992, Bhatti et al. 1995 and Fletcher et al. 1996). Negative effects were observed down to dosages equivalent to 1% of the maximum dosage given on the label. According to our knowledge similar studies of herbicide spray drift have not been conducted in Denmark or in the rest of Europe. However, herbicide spray drift is most likely to be important for food chains associated with flowering, fruit set and seed dispersal. Aim The primary aim of the project was to provide knowledge of potential effects of herbicides on trees and shrubs in the year of exposure as well as in the year after. Potential effects on these species were exemplified with the herbicide metsulfuron-methyl and with hawthorn (Crataegus monogyna). The project sought to fulfil the aim by answering the following questions:
Design The project was designed on the basis of a pilot project. The pilot project estimated the variability of biomass and number of leaves, buds, flowers, green and mature fruits (Kjær et al. 2002). Based on statistical power analyses and logistic constrains, the design included eight hedgerows dominated by hawthorn. The experimental hedgerows were geographically distributed so that five of the hedgerows were positioned in Jutland and three on Zealand. Each hedgerow was sprayed with four dosages on two occasions in order to study the importance of spray timing. The nominal dosages were 5%, 10%, 20% and 40% of maximum label rate, and a control. For each spray event, 13 trees were sprayed per dosage per hedgerow. The control trees were not sprayed. Activities Spraying The early (spring) spraying was carried out in the middle of May 2002 under variable weather conditions. The late (summer) spraying was done in the beginning of June under fairly stable weather conditions. Glycine was added to the spray solution in order to estimate the spray deposition onto leaves sampled immediately after the actual spraying. This measure was used to convert nominal spray dosages to spray solution deposited on the leaves. Sampling Year one Leaves, buds, flowers, green berries and mature berries were sampled from trees subjected to the early spraying. Sampling from trees subjected to the late spraying included leaves, green and mature berries. For all samples except flowers, the number and weight of the single end-point were measured. Flowers were only counted. The fall-down of buds, flowers, early and mature fruits were sampled in buckets placed under the experimental trees in order to measure the variability over the season, and to determine the life stage most susceptible to loss due to herbivory and other causes. Second year All end-points were sampled in the second year, i.e. leaves, buds, flowers green berries and mature berries. In the year after herbicide treatment, untreated trees in a single hedgerow were randomly selected and repeatedly sprayed with the insecticide cypermethrin in order to estimate the effect of insect herbivory on the fruit set. This was done by counting the number of buds, flowers and berries on selected shoots. The amount of berries was also counted as present within an area of 35 x 35 cm2. Results Pesticide effects in the year of exposure:
Pesticide effects the year after exposure:
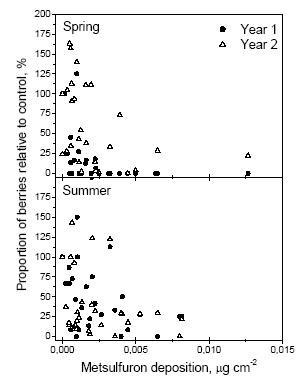
Figure 1. Comparison of percentage effects on the amount of berries in the year of exposure and the year after. The figure is based on the number of berries on side shoots the first year and the number counted in 0.1225m2 frames in the second year Biological observations:
Relevance of the data in connection with pesticide regulation The pesticide regulation does not, at present, account for any of the effects observed or made probable in the present study. This applies for the circumstance that perennial plants can be affected longer than the year of exposure. Furthermore, the standard test methods used for ecotoxicity testing do not measure effects on reproduction. The results suggest that effects on reproduction and tests that involve perennial species are recommended to consider ecosystem protection. 1 IntroduktionDenne rapport beskriver resultater fra undersøgelser af effekter af simuleret afdrift af herbicidet metsulfuron-methyl (herefter metsulfuron) til tjørnehegn i eksponeringsåret og året efter. Derudover bygger rapporten på resultater fra et pilotprojekt udført i 2001 (Kjær et al. 2002), der havde det formål at skabe fundamentet for planlægning af prøveindsamlingen i nærværende projekt. I pilotprojektet blev variationen i produktion af blade, knopper, blomster og bær undersøgt i 5 tjørnehegn, og sprøjtemidlets effekt på frugtsætningen blev undersøgt ved 4 doser og en kontrol. Hver dosis blev sprøjtet på 1 træ, hvor 10 sidegrene på forhånd var blevet afmærket, i kontroltræet blev der ligeledes afmærket 10 sideskud. Pilotstudiet viste, at der kunne forventes effekter ved doser i det mindste ned til 10% af den på etiketten anbefalede sprøjtedosering. 1.1 BaggrundDet er kendt at tætheden og diversiteten af ukrudt, insekter og fugle på markfladerne påvirkes af brugen af sprøjtemidler. Der findes en række danske undersøgelser (primært i kornafgrøder), der direkte såvel som indirekte har undersøgt sprøjtemidlers indvirken på markens dyre- og planteliv samt sammenlignet konventionel dyrkning med økologisk, for eksempel Hald & Reddersen 1990, Hald et al. 1994, Reddersen et al. 1998, Christensen et al. 1996, Esbjerg og Petersen 2002, samt (Topping & al. In press). Sprøjtemidlerne påvirker imidlertid ikke kun organismer, der opholder sig på markfladen, men også organismer knyttet til omkringliggende habitater, såsom ruderater, hegn, skovkanter, markkanter etc. (Freemark &Boutin 1995). Kleijn & Snoeijing 1997, Mars & Frost 1997 og de Snoo& van der Poll 1999 observerede en faldende biodiversitet ved lave herbiciddoser. Al-Khatib et al. 1992, Bhatti et al. 1995 og Fletcher et al. 1996 fandt negative effekter af sulfonylurea-midler og glyfosat på vækst og frugtsætning i fuglekirsebær (Prunus avium) i forsøg, der simulerede herbicidafdrift. I nogle tilfælde blev der observeret negative effekter ved doser, der svarer til 1% af den på etiketten anbefalede sprøjtedosering. En gennemgang af litteraturen på området antyder, at der ikke tidligere er lavet tilsvarende undersøgelser af herbicidafdrift på bærbærende buske og træer i Danmark eller Europa. Der er dog næppe tvivl om, at afdrift af herbicid til hegn kan have stor betydning for de mange fødekæder, der er knyttet til netop blomstring, frugtsætning og frøspredning. I nærværende projekt anvendes tjørn som repræsentant for hegnets træer, idet tjørn er hyppigt forekommende i danske læhegn og desuden udgør en del af fødegrundlaget for mange arthropoder (Kennedy & Soutwood 1984). I USA anvendes minimidler, deriblandt metsulfuron, til bekæmpelse af uønsket opvækst af bl.a. Crataegus spp., hvor man også har iagttaget at effekten rækker ud over det første år (Sprague & Hager 2003). Det er endvidere kendt, at tjørnebær sammen med en række andre bær er et vigtigt fødeemne for overvintrende og trækkende fugle (Snow & Snow 1988). "Minimidler", herunder sulfonylurea-produkterne, anvendes i stigende mængder i dansk landbrug. Sulfonylurea-midlerne virker ved at blokere dannelsen af de grenede aminosyrer, valin, leucin og isoleucin. Det er kendt, at minimidlerne kan hæmme transporten af fotosynteseprodukter ud af behandlede blade (Bestman et al. 1990, Vanden Borne et al. 1988). Dermed er det også sandsynligt, at der kan forekomme effekter på for eksempel frugtsætningen, hvis sprøjtningen finder sted på et kritisk tidspunkt. Påvirkning af hegn forventes at kunne ske på forskellige årstider og med både herbicider og insekticider. I frugt- og frøproducerende hegn kan påvirkningen potentielt medføre effekter på de fleste af de dyr, der er knyttet til hegnet enten i forbindelse med fødesøgning eller det sted de lever. Påvirkning af hegnet som levested kræver dog formentlig en ret drastisk herbicideksponering af hegnet, som mere eller mindre afløver dele af hegnet. Påvirkningen af den tilgængelige fødemængde kan derimod ske ved langt mindre eksponering. Fuglekirsebær er et eksempel på en art, som er akut følsom for små herbicidmængder. Bhatti et al. 1995 fandt således at koncentrationer af chlorsulfuron på helt ned til 3,1 x 10-7 M nedsatte udbyttet signifikant. Det vil sige at træerne gav cirka 50% færre bær. 1.2 FormålDet er formålet med dette projekt at etablere kvantitative data for herbicidet metsulfurons påvirkning af vegetativ vækst, blomsterproduktion og bærproduktion hos engriflet hvidtjørn i samme sæson og sæsonen efter sprøjtemiddeleksponering. Projektet ønsker således at besvare følgende spørgsmål:
2 Materiale og metoder
Effekter af afdrift af metsulfuron fra marksprøjtning blev undersøgt ved at udføre sprøjteforsøg med reducerede doser, hvor der sprøjtes direkte på træet. De valgte doser var 0, 5, 10, 20 og 40% af den af producenten angivne maksimaldosis. Disse doser blev valgt på baggrund af pilotforsøg og litteraturundersøgelser af afdriftens størrelse. Dette sikrede, at doserne både var relevante for afdrift under markforhold og at der blev inkluderet doser med stor effekt, i det mindste på frugtsætningen. Projektet etablerer således kvantitative data for herbicidet metsulfurons effekt på vegetativ vækst, blomsterproduktion og bærproduktion hos engriflet hvidtjørn. Projektet belyser endvidere betydningen af tidspunktet for eksponering ved at lade sprøjtning finde sted på to tidspunkter (sent forår henholdsvis tidlig sommer). Projektet inkluderer undersøgelse af effekter i eksponeringsåret og effekter året efter eksponering. 2.1 DesignDer blev foretaget prøvetagning i 8 hegn. Hegn 8 viste sig at være klippet meget hårdt i 2001. Dette gjorde, at der ikke var så mange egnede træer i hegnet. Derfor blev der ikke plads til sommersprøjtning. Da det i mange tilfælde ikke var muligt at indsamle nok sidegrene fra de sprøjtede træer, valgte vi helt at udelade dette hegn fra dataanalyserne. I hegn 1 til 7 blev 117 træer i hvert hegn afmærket med fortløbende numre. De 117 træer blev fordelt på fire blokke. I hver blok var der kontroltræer og træer til sprøjtning med de fire valgte doser både forår og sommer. Træerne var placeret fortløbende i hegnet. Hver blok startede med alle kontroltræerne, derefter kom i nævnte rækkefølge 5% forår, 5% sommer, 10% forår, 10% sommer, 20% forår, 20% sommer, 40% forår og 40% sommer. Der var et mellemrum på mindst 5 m mellem hver nye dosis, mellem forårs- og sommersprøjtede træer og mellem blokke. De hegn der lå op af dyrkede arealer blev ikke sprøjtet i en zone på minimum 6 m fra hegnet. 2.2 SprøjtningForud for sprøjtning blev der for hvert hegn lavet 1 stamopløsning med 1 g Ally per 500 ml vand. Der blev tilsat 10 ml 10% Citowett per 2 l sprøjteopløsning. Citowett blev tilsat sprøjtevæsken, fordi det i nogen tilfælde er anbefalet at tilsætte det. Det må antages at effekten øges herved og derfor er at opfatte som et "worst case scenarie". Ved marksprøjtning bliver der sprøjtet med 200 l opløsning ha-1 svarende til 0,1 g Ally l-1. Umiddelbart før sprøjtningen blev stamopløsningen fortyndet til den ønskede koncentration (5%, 10%, 20% og 40% af den af producenten anbefalede dosering for vårbyg (20 g Ally ha-1)). Disse doser blev valgt på baggrund af range-finding forsøg udført i forbindelse med pilotforsøget (Kjær et al. 2002). Planterne blev sprøjtet med en azo-sprøjte, hvor sprøjtebommen blev holdt lodret, således at planterne sprøjtedes med den ønskede dosis i et "bælte" fra jordoverfladen og op til ca. 2,5 m. Der blev kun sprøjtet på den ene side af hegnet. Der blev foretaget verifikation af den eksponering grenene var udsat for. Væskemængden, der blev udsprøjtet svarede til 200 l ha-1. Denne mængde blev opnået ved at justere ganghastigheden i forhold til den væskemængde som azosprøjten leverer ved 2 atm arbejdstryk og med Hardi 4110-16 dyser monteret. Der blev foretaget to sprøjtninger i alle hegn; en forårs- og en sommersprøjtning. Forårssprøjtningen blev foretaget over en relativ lang periode (fra den 13. til den 22. maj). Det skyldes først og fremmest, at der var meget regn og blæst i perioden (Bilag A). Sommersprøjtningen blev foretaget under gunstigere vejrforhold (stille vejr uden regn) i perioden fra den 19. juni til den 21. juni (Bilag B). I hegn 8 blev der kun blev sprøjtet i maj (forårssprøjtning). Hegnet havde, på grund af beskæring, for få træer med fertile sideskud til at gennemføre begge sprøjtninger. 2.2.1.1 Afsætning af sprøjtemiddelFor at være i stand til at relatere observerede effekter til den sprøjtemiddelafsætning der forekommer, når man bruger normalt sprøjteudstyr monteret på traktorer blev der tilsat et sporstof til sprøjtemiddelopløsningerne. Der blev tilsat glycin til alle opløsninger i en koncentration på 10 g glycin l-1. Herefter kunne vi bestemme afsætningen af sprøjtemidlet ved at måle hvor meget glycin, der blev afsat på tre blade udvalgt omkring de på forhånd afmærkede sideskud (Babcock et al. 1990). 2.2.1.2 Omregning til metsulfurondoserMetsulfuron udsprøjtes, ved den af producenten angivne maksimaldosis for vårsæd, i en mængde på 4 g ha-1. Disse 4 g bliver fortyndet i 200 L vand. Med en koncentration på 10 g glycin l-1 ville der ved anvendelse af 200 L per ha blive udspredt 2000 g glycin ha-1. Det vil sige, at koncentrationen af glycin i sprøjtevæsken ved 100% dosis er 2000/4 = 500 gange koncentrationen af metsulfuron. Den på etiketten anbefalede sprøjtedosering svarer til 40 ng metsulfuron cm-2, denne værdi opnås ved 20 g glycin cm-2, hvis der sprøjtes med 100% af den af producenten angivne maksimaldosis. En afsætning på 1 g glycin cm-2 svarer altså til 0,002 g metsulfuron cm-2 hvis der sprøjtes med 100% af den af producenten angivne maksimaldosis. Imidlertid sprøjtes der med henholdsvis 5%, 10%, 20% og 40% heraf. For at omregne fra glycinafsætning til aktuel afsætning af sprøjtemidlet skal der altså for de 4 anvendte doser ganges med henholdsvis 0,05; 0,1; 0,2 og 0,4 (Tabel 2.1). Tabel 2.1 Omregningstabel fra afsætning af glycin til afsætning af metsulfuron.
2.3 PrøveindsamlingI pilotprojektet (Kjær et al. 2002), som blev gennemført i 5 hegn i 2001 udførtes der styrkeberegninger med det formål at estimere hvor mange hegn, træer per hegn og sidegrene per træ, der skulle indgå i hovedprojektet for med 80% sandsynlighed at kunne detektere en 20% effekt ved et signifikansniveau på 0,05. Resultatet blev, at der i hvert af 8 hegn for hver dosis skulle indsamles 1 sideskud fra hvert af 13 træer for bær; fra 10 træer for blomster og endelig fra 4 træer for blade. Så snart det blev muligt at skelne mellem fertile og sterile skud blev de fertile sideskud afmærket med mærkater der angav hvilken dosis, der skulle sprøjtes med og hvilken type indsamling, der skulle foretages. 2.3.1 Indsamling og prøvebehandling i 20022.3.1.1 Indsamling af prøver til bestemmelse af sprøjtemiddeleffektEfter forårssprøjtningen blev alle bladene i de udvalgte sideskud indsamlet i perioden fra den 4. til den 6. juni. Efter sommersprøjtningen blev blade indsamlet den 29. – 31. juli. Prøverne blev indsamlet i papirposer og frosset ved hjemkomst. Senere blev de frysetørret, vejet og optalt. Da alle knopper var sprunget ud inden indsamling blev påbegyndt (ca. 1 uge efter sprøjtning), blev der ikke indsamlet knopper i 2002, da en reaktion på sprøjtemidlet ikke kunne forventes inden for en uge efter sprøjtning. Blomster blev udelukkende indsamlet efter forårssprøjtning, idet de var afblomstret kort efter sommersprøjtning. Blomsterne blev hjembragt i papirposer, frosset og senere optalt. Blomsterne blev ikke vejet. Grønne bær blev indsamlet fra den 30. – 31. juli og de modne bær blev indsamlet fra den 23. til den 26. september. De blev behandlet på samme måde som blad- og knopprøver. 2.3.1.2 Indsamling af prøver til bestemmelse af sprøjtemiddelafsætningTre blade fra hvert træ blev plukket umiddelbart efter, at alle træer i hegnet var sprøjtet. Ved plukning blev bladene anbragt i 10 ml demineraliseret vand i 1 dl beholdere med låg. Ved hjemkomst blev prøverne sat på køl. Dagen efter blev bladene taget op til bladarealmåling. Beholderne med det afvaskede glycin blev frosset. Senere blev væskens koncentration af glycin bestemt. 2.3.1.3 Indsamling af nedfaldne knopper, blomster og bærI hvert hegn blev der efter endt afmærkning medio maj (uge 20) stillet 80 spande op til opsamling af nedfald af knopper, blomster og bær. Der blev stillet 20 spande per blok dvs. 4 spande for hver af de fem anvendte doser inklusive kontrol. Nedfaldet blev opgjort i felten. Data blev delt op i nedfald med synligt biologisk angreb og nedfald af anden årsag. Der blev ikke skelnet mellem sommer- og forårssprøjtning. 2.3.2 Indsamling og prøvebehandling i 2003Fra alle hegn blev blade fra fire afmærkede sideskud per hegn, knopper og blomster fra 10 afmærkede sideskud per hegn, samt grønne og modne bær fra 13 afmærkede sideskud per hegn indsamlet indenfor samme uge. Prøverne blev efterfølgende frosset, frysetørret, optalt og endelig blev tørvægten bestemt. Blomsterne blev ikke vejet, kun talt, dels fordi de nogle gange taber kronblade og støvdragere under indsamlingen, dels fordi deres vægt må forventes at være tæt korreleret med knoppernes. Ved indsamlingen af de grønne bær fik vi det indtryk, at der var færre bær på sideskuddene end i 2001 og 2002. På denne baggrund forventede vi at antallet af nul-prøver kunne blive for stort. Derfor besluttede vi at supplere indsamlingen af de modne bær med en optælling af antallet af modne bær i en 35 x 35 cm2 ramme som havde det afmærkede sideskud som centrum. Eventuelle modne bær fra det enkelte sideskud indgår altså i antallet i rammen. Bærrene fra sideskuddene blev både talt og vejet, bærrene fra rammerne blev kun talt. 2.4 LokalitetsbeskrivelseDer indgår 8 tjørnehegn i undersøgelsen. De 5 er placeret omkring Silkeborg i Midtjylland. De resterende ligger i et område nord for Birkerød i Nordsjælland. Tabel 2.2 præsenterer hegnene. Tabel 2.2 Generel beskrivelse af forsøgshegnene og deres omgivelser
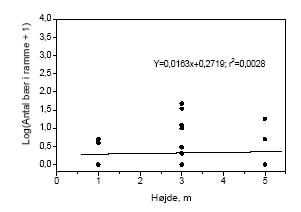
2.5 Bærproduktion med højdenTil bestemmelse af biomassen af træets reproduktive enheder og hvorledes de er fordelt, blev mængden af bær i en ramme med et areal på 0,1225 cm2 talt. Der blev lavet en optælling i tre højder henholdsvis 1, 3 og 5 m. Der var 5 replikater i hvert hegn og optællingen blev foretaget i alle hegn. Desuden blev antallet af bær på 5 grene på ca. 3/4 meters længde, i højderne 1, 3 og 5 m opgjort. Der taltes grene fra 2 træer i hvert hegn og indsamlingen blev foretaget både på sprøjtesiden og læsiden. Optællingen foregik 5. - 6. august 2003. Resultaterne muliggør en bedømmelse af hvor meget eksponering af hegn med metsulfuron betyder for den tilgængelige fødemængde i form af bær, idet afsætningen af sprøjtemidlet ved afdrift fra mark aftager med hegnets højde (Weisser et al. 2002). 2.6 InsekticidsprøjtningBetydningen af herbivori for frugtsætningen i tjørn blev bestemt ved at fjerne de herbivore insekter gennem gentagne insekticidsprøjtninger. I hegn fem blev nogle træer således sprøjtet med cypermethrin, et insekticid af pyrethroidtypen, hvorefter effekten på insekter og tjørnen blev registreret. Der var 5 replikater per behandling og hvert replikat bestod af 3 træer. Behandling og kontrol var placeret skiftevis langs hegnet og adskilt af en sikkerhedszone på ca. 5 meter. Der blev sprøjtet med cypermethrin med den af producenten anbefalede maksimaldosering (0,2 l ha-1) hver fjortende dag. Første sprøjtning blev gennemført i uge 20. Sprøjtningen ophørte på det tidspunkt, hvor der var grønne bær til stede. De gentagne sprøjtninger blev foretaget for at sikre at de herbivore insekter blev holdt væk, medens tjørnen var i knop, blomst og frugtanlægsstadierne. Inden første sprøjtning blev i hvert træ markeret 5 tilfældigt udvalgte blomsterstande, hvor der ikke er tegn på insektangreb. Antallet af blomsterknopper blev registreret og udviklingen fulgt ved registreringer hver fjortende dag gennem sæsonen. Insekter blev indsamlet hver 14. dag. Indsamlingen bestod af 5 ketcherslag i hvert træ i 150-180 cm højde og 3 bankeprøver på grenafsnit. De indsamlede insekter blev puljet til en prøve per behandling. Optællingen af de modne bær blev efterfølgende opgjort i 35 x 35 cm2 rammer. Der blev optalt 3 tilfældige steder inden for hvert replikat. 2.7 Databehandling og statistiske analyser2.7.1 20022.7.1.1 SprøjtemiddeleffektHverken biomassedata eller antalsdata kunne med rimelighed beskrives ved en normalfordeling. Antalsdata kunne heller ikke beskrives ved en Poissonfordeling. Derfor benyttede vi en ikke-parametrisk test til at undersøge, om der var forskel på de 5 doser. Med det valgte randomiserede blok-design kan det gøres på flere måder, f.eks. Jonckheere-Terpstra test med stratifikation efter blokke eller Page' test (der svarer til en 2-sidet variansanalyse med blokke og doser men uden interaktion). Vi har dog valgt at benytte Spearman's rang korrelation (rS) stratificeret efter blokke til at vurdere, om der er en monoton sammenhæng mellem respons og dosis. Der er næppe den store forskel i statistisk styrke på de 3 tests, men rent regneteknisk er Spearmans rang korrelation nemmere at lave i en stratificeret version. Det skyldes, at under nulhypotesen om ingen sammenhæng mellem respons og dosis (indenfor en blok) gælder at Var(rS) = 1/(n-1), uanset om der er ties eller ej (n = antal observationer i blokken, ties = sammenfald af observationer). Det bemærkes, at der er det samme antal blokke pr. hegn. For at lave den stratificerede test benyttes de reciprokke varianser som vægte på de blok-specifikke korrelationer. Det vil sige
2.7.1.2 Antal nedfaldne knopper, blomster og bærData kunne beskrives med en normalfordeling. Mængden af nedfaldsfrugt samt den andel af disse, der var angrebet af insekter, blev analyseret ved en variansanalyse, der medtog både systematiske og tilfældige variansfaktorer. I analysen blev sprøjtemiddeldosen (i % af den af producenten angivne maksimaldosis) samt tidspunkt indsat som systematiske effekter, hvorimod hegn er at opfatte som en tilfældig effekt. Varianskomponenten for hegn (dvs. variationen mellem hegn) vil blive opgjort som den procentvise andel af den samlede variation (=hegn + residual). Den statistiske model er:
hvor Ydth er responsvariablen målt under træer i hegn h, med behandling d og til tidspunkt t. μ, αd og βt betegner middelværdi og de systematiske effekter af henholdsvis sprøjtemiddelbehandling d og tidspunkt t. Den tilfældige effekt af hegn h betegnes Eh og residualleddet betegnes Udth. De tilfældige effekter og residualleddene antages uafhængige og normalfordelte. 2.7.2 20032.7.2.1 SprøjtemiddeleffektMålvariable for knopper, blomster og blade viste sig at kunne antages at være normaltfordelte. Derfor blev der lavet en variansanalyse af disse. I analysen blev sprøjtemiddeldosen (i μg metsulfuron cm-2) indsat som systematisk effekt, hvorimod hegn og blok begge opfattes som tilfældige effekter. Blokfaktoren er 'nested' inden for hegnsfaktoren. De to varianskomponenter vil blive opgjort som deres andel af den samlede variation (hegn + 'blok inden for hegn'+residual). Modellen er:
hvor Ydthb er responsvariablen målt under træer i hegn h, blok b, med behandling d og til tidspunkt t. μ, αd og βt betegner middelværdi og de systematiske effekter af henholdsvis sprøjtemiddelbehandling d og tidspunkt t. De tilfældige effekter af hegn h og af blok b i hegn h betegnes Eh og Fh,b residualleddet betegnes Udthb. Alle de tilfældige effekter og residualleddene antages uafhængige og normalfordelte. Det viste sig, at bærdata ikke kunne antages at være normalfordelte. Mængden af bær var simpelthen så lille, at der næsten kun var nul-prøver, og hermed var det også irrelevant at gennemføre nogen statistisk analyse. I stedet blev den supplerende optælling af bær inden for en ramme testet med en Spearman rang korrelation (jf. 2.6.1.1). 2.7.2.2 Bærproduktion med højden i hegnetDisse data blev analyseret ved lineær regressionsanalyse på logaritmetransformerede data (Log(antal bær+1)). 2.7.2.3 InsekterForskelle i antallet af reproduktive enheder i henholdsvis ubehandlede (kontrol) og behandlede (insekticidsprøjtet) træer blev analyseret ved hjælp af en variansanalyse med gentagne målinger (de samme 15 sideskud målt på de samme 5 tidspunkter) for effekten af sprøjtebehandling. Det viste sig, at der var større variation i kontrolgruppen end i den behandlede gruppe, og at der var forskel i variationen på de 5 forskellige tidspunkter. Desuden var det ikke muligt at opnå en simpel reduktion af kovariansstrukturen (korrelationsstrukturen) mellem de 5 tidspunkter, der derfor blev fittet som en ustrukturet 5x5 kovariansmatrix (pr.behandling). For at vurdere behandlingseffekten pr. tidspunkt blev der lavet Bonferoni korrektion af disse test (det var de samme sideskud, der blev talt op gentagne gange). Modellen er:
hvor Ybts er den "målte respons" ved behandling b til tidspunkt t i sample s. αt angiver den systematiske effekt til tidspunkt t, og βbt angiver interaktionen mellem behandling b og tidspunkt t (dvs. behandlingseffekten til tidspunkt t). Ebts er residualleddet. Residualerne for sample s der har fået behandling b antages at følge en multivariat normalfordeling, dvs. hvis
så skal
3 Resultater
3.1 Sprøjtemiddeleffekter i eksponeringsåretDer var et signifikant fald i mængden af blade, blomster, grønne bær og modne bær efter forårssprøjtning med metsulfuron (Tabel 3.1). Ved sommersprøjtningen var der ikke nogen effekt af sprøjtningen på antallet og vægten af blade i sideskud. Blomsterne blev ikke høstet, idet de på det tidspunkt hvor sprøjtningen blev gennemført var næsten afblomstret. Der var en signifikant reduktion i vægten og antallet af både grønne og modne bær per sideskud som følge af sommersprøjtningen (Tabel 3.1). Tabel 3.1 Statistiske analyser af effekten i 2002 af metsulfuron-behandling på antal og vægt af blade, blomster, grønne bær og modne bær i sideskud af tjørn. Analysen blev gennemført med en Spearman rang test vægtet for antallet af prøver per blok. N er antallet af blokke, Z er testværdien for testen. p-værdier fremhævet med fed er statistisk signifikante med en på 0,05.
*) dette endpoint er ikke beregnet idet hegnet var afblomstret ved sommersprøjtningen 3.1.1 Relevans af effektstørrelser3.1.1.1 Sammenhæng mellem nominel dosis og målt afsætningPå grund af vores manipulerede sprøjtning kan analyserne, der blev præsenteret i forrige afsnit, ikke umiddelbart sammenlignes med den eksponering som tjørnehegn udsættes for ved en drifthændelse ved en normal marksprøjtning. For at omsætte disse data, må vi omsætte den nominelle dosis til en mængde sprøjtemiddel afsat på blade af tjørn. En regressionsanalyse af den nominelle dosis over for den målte afsætning viste, at der var en klar sammenhæng mellem det, det var intentionen at udsprøjte, og det der faktisk blev afsat på bladene af tjørnene. Analyserne viste også at afsætningen var meget forskellig mellem forskellige hegn (Tabel 3.2). En tovejs variansanalyse viste, at der var signifikant (F=13,4 df=7 p<0,0001) interaktion mellem sprøjtemiddeldose og hegn. I de efterfølgende analyser vil afsætningen i det enkelte hegn derfor være udregnet ud fra de regressionslinier, der er præsenteret i Tabel 3.2. Tabel 3.2. Regressionsanalyser af sprøjtemiddelafsætning (μg cm-2) i forhold til den ønskede dosering (% af den af producenten angivne standard dose). Data blev beskrevet ved følgende ligning: Y = β x dosis, hvor Y er den målte sprøjtemiddelafsætning. β er hældningen på regressionslinien. Dosis er den procentvise andel af den af producenten angivne standarddose. N er antallet af observationer. SE angiver standardfejlen på .
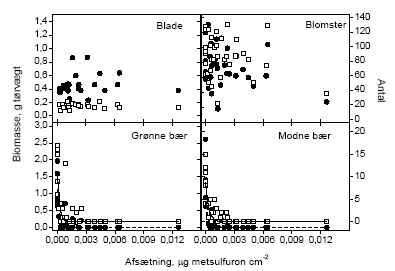
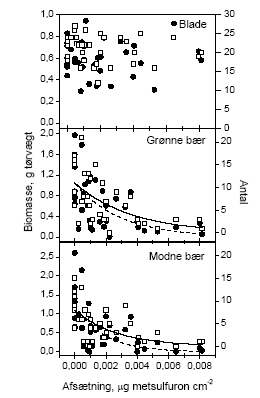
3.1.1.2 Effekt i forhold til faktuel eksponeringDe mange observationer med en vægt/antal på nul gør, at vi ikke kan fæste lid til at almindelige forudsætninger for regressionsværktøjer er opfyldt. I stedet har vi, for at kunne håndtere data, valgt at pulje data for hvert hegn således at der fremkommer ét tal for sprøjtemidlets effekt på produktionen af den valgte målvariabel. Måleparameteren er "antal" eller "biomasse". Disse data kunne antages at være normaltfordelte, hvorfor en almindelig regressionsanalyse blev benyttet. Det blev fundet, at der ikke var nogen signifikant sammenhæng mellem sprøjtemiddeldosis og mængden (antal og biomasse) af henholdsvis blade og blomster ved en forårssprøjtning. Der var signifikant sammenhæng mellem sprøjtedosis og mængden af både grønne og modne bær (Tabel 3.3 og Figur 3.1).
Figur 3.1 Forholdet mellem den målte afsætning af sprøjtemiddel i det enkelte hegn og antal og biomasse af blade, blomster, grønne bær og modne bær efter en forårssprøjtning i 2002. De åbne symboler indikerer antallet af den givne målevariabel og de fyldte symboler viser biomassen af den givne målevariabel. Tabel 3.3 Regressionsanalyse af sprøjtemiddelafsætning (μg metsulfuron cm-2) i relation til de to måleparametre; biomasse i g tørvægt og antal. Data for blade og blomster blev beskrevet ved en lineær sammenhæng med funktionen: Y = •β x depos + α•. Data for bærprøverne blev beskrevet ved en eksponentiel førsteordens henfaldsfunktion med ligningen: Y = γx E(-depos/λ). Y er måleparameteren og depos er den beregnede sprøjtemiddelafsætning (μg metsulfuron cm-2). N er antallet af observationer.
Blade på tjørn der var blevet eksponeret for metsulfuron ved en sommersprøjtning var ikke påvirket af sprøjtningen. Det vil sige, at bladene ikke var signifikant mindre med stigende dosis, og at der ikke var færre blade. Blomster blev ikke høstet efter sommersprøjtningen, idet de var afblomstret på dette tidspunkt. Både grønne og modne bær blev påvirket signifikant af sprøjtningen (Tabel 3.3 og Figur 3.2).
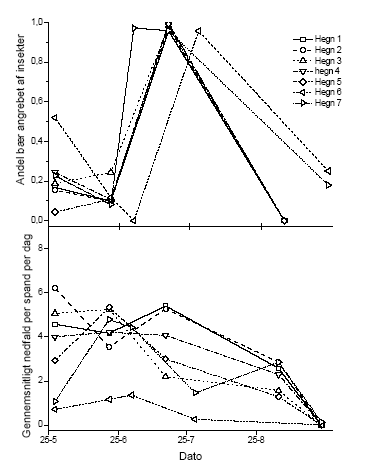
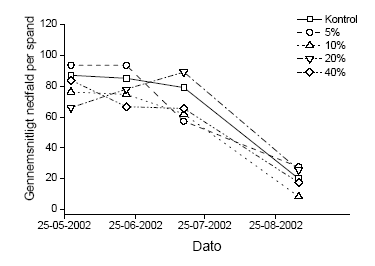
Figur 3.2 Forholdet mellem den målte afsætning af sprøjtemiddel i det enkelte hegn og antal eller biomasse af blade, blomster, grønne bær og modne bær efter en sommersprøjtning. De åbne symboler indikerer antallet af enheder af den givne målvariabel og de fyldte symboler viser biomassen af samme. 3.1.2 Nedfald af knopper, blomster og bærDe reproduktive enheder, der blev opsamlet i spande under forsøgshegnene blev analyseret for de systematiske effekter af tidspunkt og sprøjtemiddeldosis. Da der ikke blev fundet signifikante interaktioner, blev data kun testet for disse to faktorer. For både forårs- og sommersprøjtningen var den mængde af reproduktive enheder, der faldt af tjørnehegnene, uafhængig af den sprøjtemiddeldosis, hegnet var sprøjtet med. Der var dog en signifikant forandring i nedfaldet med tiden (Tabel 3.4). Den totale variation blev opdelt i den variation, der kunne henføres til forskelle mellem hegn og den øvrige tilfældige variation (residualvariation). 40,7 og 46,7% af totalvariationen kunne henføres til variation mellem hegn for henholdsvis forårs- og sommersprøjtning. Det blev endvidere opgjort, hvor mange af de nedfaldne reproduktive enheder der var påvirket/angrebet af insekter (antal angrebne/antal i alt). Denne målvariabel var også afhængig af tidspunkt, men ikke af dosis. I dette tilfælde viste det sig, at 22,1 og 18,3% af totalvariationen skyldtes variation mellem hegn for henholdsvis forårs- og sommersprøjtning. Figur 3.3 og 3.4 illustrerer, at der var et nedfald af knopper, blomster og bær gennem hele opsamlingsperioden, som dog tog af mod periodens slutning (Figur 3.3). Afbildningen af den andel af nedfaldet, der viste tegn på insektangreb, viser at der var en periode i slutningen af maj, hvor der var skader efter insekter. Efterfølgende periode var stort set fri for herbivori, hvorefter der var en periode i slutningen af juli, hvor samtlige bær var angrebet (gnavskader). Herefter var der ikke nogen videre tegn på herbivori. Sammenligningen mellem kontrol og behandling i Figur 3.4 viser, at der ved denne indsamlingsmetode ikke var nogen effekt af vores sprøjtning med metsulfuron i en højde op til 2,5 m. Tabel 3.4 Variansanalyse af henholdsvis nedfald og andel insektangrebne reproduktive enheder fra tjørnetræer i forhold til sprøjtemiddeldose i de nederste 2,5 m og tidspunkt på sæsonen. Den totale variation blev opdelt i den del, der kunne beskrives ved variation mellem hegn og restvariationen.
Figur 3.3 Nedfald af frugtanlæg og frugter i spande under usprøjtede træer. Øverst vises den andel, der er angrebet af insekter og nederst det samlede nedfald af reproduktive enheder i forhold til tidspunkt på sæsonen.
Figur 3.4 Nedfald af frugtanlæg og frugter i spande under hegn 2 over tid for hver anvendte sprøjtemiddeldose. 3.2 Sprøjtemiddeleffekter året efter eksponeringDe indsamlede data for knopper, blomster og blade i året efter sprøjtning kunne antages at være normaltfordelte, hvorfor vi gennemførte en variansanalyse med sprøjtemiddeldosis som den systematiske faktor. Den totale variation blev opdelt i den variation, der kan henføres til hegnet og den øvrige tilfældige variation. Som det fremgår af Tabel 3.5, var der en stærkt signifikant negativ sammenhæng mellem sprøjtedosis og antallet og vægten af alle målvariable for både forårssprøjtning og sommersprøjtning. Hældningen og skæring med y-aksen var sammenlignelige for den enkelte målvariabel mellem forårssprøjtning og sommersprøjtning. Det fremgår endvidere af Tabel 3.6, at variationen mellem blokke maksimalt bidrog til 4,7% af den totale variation. Den variation der kan henføres til forskelle mellem hegn varierede mellem 2,6 og 22,8%. Tabel 3.5 Regressionsanalyse af vægt og antal knopper, blomster og blade året efter exponering i forhold til sprøjtemiddeleksponering (μg metsulfuron cm-2) efter henholdsvis en forårs- og en sommersprøjtning. Data blev beskrevet ved en lineær sammenhæng med funktionen: Y = depos + ß. Y er målevariablen og depos er den beregnede sprøjtemiddelafsætning (μg metsulfuron cm-2).
Tabel 3.6 Oversigt over fordelingen af den totale variation på de tilfældige faktorer hegn og blok. Den totale variation blev opdelt i den del, der kunne beskrives ved variation mellem hegn, varians mellem blokke og restvariationen.
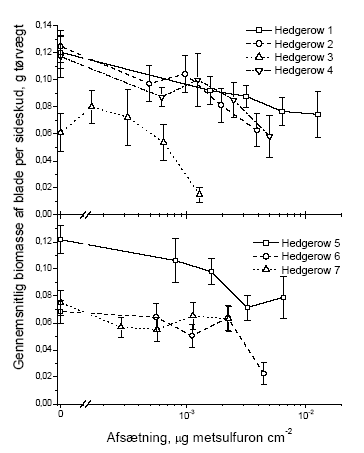
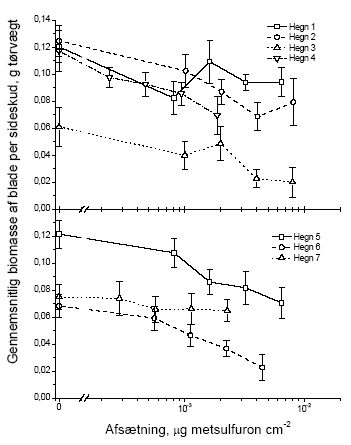
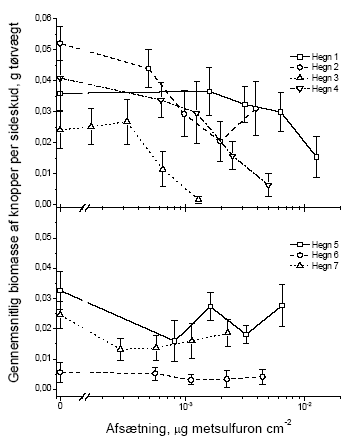
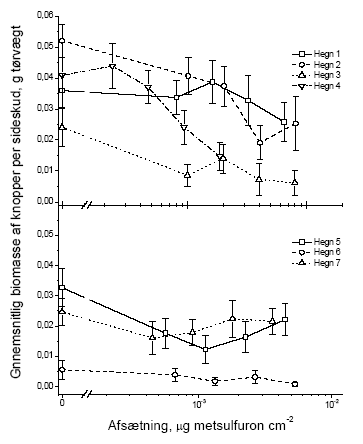
I dette afsnit har vi valgt kun at vise figurer for vægt, idet data for antal fuldstændigt følger tendenserne i de præsenterede vægtdata. De følgende figurer illustrerer datagrundlaget for resultaterne i tabellerne 3.5 og 3.6. 3.2.1.1 Blade
Figur 3.4 Sammenhængen mellem sprøjtemiddeldosis og gennemsnitsvægten af bladene i et sideskud året efter en forårssprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet.
Figur 3.5 Sammenhængen mellem sprøjtemiddeldosis og gennemsnitsvægten af bladene i et sideskud året efter en sommersprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet. 3.2.1.2 Knopper
Figur 3.6 Sammenhængen mellem sprøjtemiddeldosis og gennemsnitsvægten af knopper i et sideskud året efter en forårssprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet.
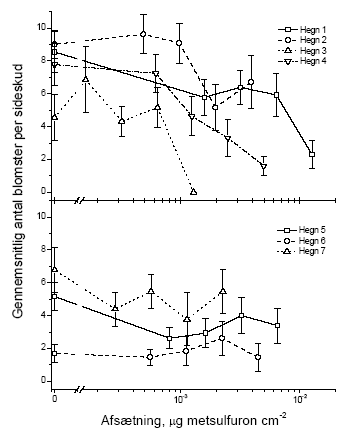
Figur 3.7 Sammenhængen mellem sprøjtemiddeldosis og gennemsnitsvægten af knopper i et sideskud året efter en sommersprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet. 3.2.1.3 BlomsterBlomsterne blev kun talt, ikke vejet. Dette skyldes at kronbladene ofte falder af ved indsamling, hvorfor vægten ikke er et repræsentativt mål.
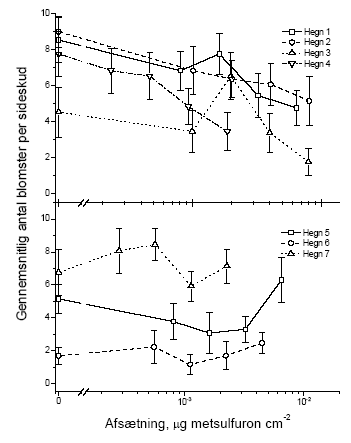
Figur 3.8 Sammenhængen mellem sprøjtemiddeldosis og antallet af blomster i et sideskud året efter en forårssprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet.
Figur 3.9 Sammenhængen mellem sprøjtemiddeldosis og antallet af blomster i et sideskud året efter en sommerssprøjtning med metsulfuron. Data er præsenteret hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet. 3.2.1.4 Grønne bærDer var for få tilbageværende grønne bær både i kontroltræerne og i de behandlede træer til at det gav nogen mening at lave en statistisk analyse. 3.2.1.5 Modne bærDer var ligeledes for få modne bær til at det gav nogen mening at lave en statistisk analyse. Den supplerende optælling af bær indenfor en ramme øgede observationsmængden så meget at der kunne testes for effekter af sprøjtningen. Idet der stadig var mange nulprøver blev data analyseret ved hjælp af Spearman rang korrelationstest vægtet for antallet af prøver per blok. Der var signifikant effekt i en række hegn både ved forårs- og sommersprøjtning (Tabel 3.7 og Figur 3.10). Tabel 3.1 Statistiske analyser af effekten i 2003 af metsulfuron-behandling på antal modne bær i sideskud af tjørn (ramme). Analysen blev gennemført med en Spearman rang korrelationstest vægtet for antallet af prøver per blok. N er antallet af blokke, Z er testværdien for testen. p-værdier fremhævet med fed er statistisk signifikante med en α på 0,05.
Figur 3.10 Sammenhængen mellem sprøjtemiddeldosis og antallet af modne bær inden for en ramme på 35 x 35 cm2 året efter forårs- eller sommersprøjtning med metsulfuron. Data er præsenteret for hvert hegn for sig. Stolperne repræsenterer ± standardfejlen på middeltallet. Bemærk at skala på y-aksen varierer mellem delfigurerne. 3.3 Bærproduktionen med højden i hegnetPå sprøjtesiden af træerne var der en positiv sammenhæng mellem bærmængden og højden i hegnet ved begge opgørelsesmetoder (Figur 3.11 og 3.12). Tendensen var nogenlunde den samme hvad enten optællingen foregik i rammer eller på grene, men det var kun i rammerne den var signifikant (p 0,05). På læsiden var produktionen væsentlig lavere end på sprøjtesiden, og der var ingen sammenhæng mellem højde og bærmængde (Figur 3.13). Dette skyldes sandsynligvis, at sprøjtesiden generelt faldt sammen med den side af hegnet som får mest sol. Ved studiet af figurerne 3.11 – 3.13 er det vigtigt at være opmærksom på, at et enkelt punkt på figurerne kan dække over flere ens observationer. Dette gælder særligt for nul-observationerne.
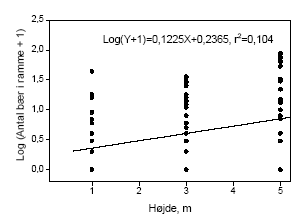
Figur 3.11. Bærmængdens fordeling over højden i tjørnehegn opgjort ved hjælp af optælling i rammer i kontroltræer på hegnets sprøjteside. Bemærk at et enkelt punkt kan repræsentere flere observationer, dette gælder specielt 0-observationer.
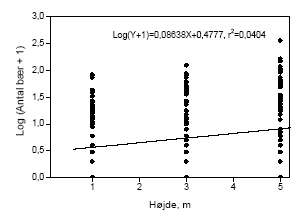
Figur 3.12. Bærmængdens fordeling over højden i tjørnehegn opgjort ved hjælp af optælling på grene i kontroltræer på hegnets sprøjteside. Bemærk at et enkelt punkt kan repræsentere flere observationer, dette gælder specielt 0-observationer.
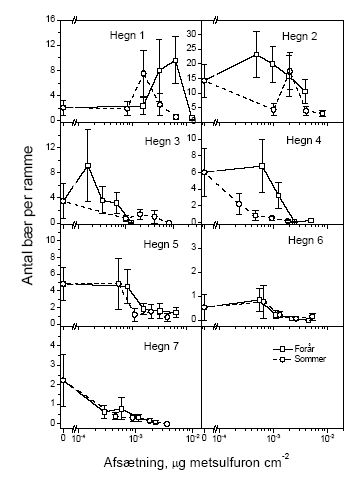
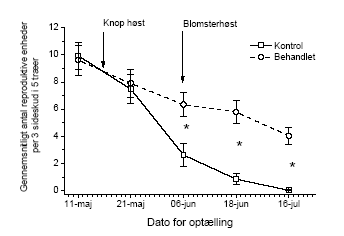
Figur 3.13. Bærmængdens fordeling over højden i tjørnehegn opgjort ved hjælp af optælling i rammer på den usprøjtede læside af hegnet. Bemærk at et enkelt punkt kan repræsentere flere observationer, dette gælder specielt 0-observationer. 3.4 Insekter3.4.1 Bærproduktion når insekterne bortsprøjtesBortsprøjtning af insekterne med cypermethrin viste, at insekterne har stor betydning for frugtsætningen. Således var antallet af bær på de usprøjtede skud allerede den 16. juli faldet til nul, medens der på de sprøjtede i gennemsnit var godt fire bær per sideskud (Figur 3.14). Den store forskel blev endvidere dokumenteret ved optællinger af bær i rammer på 35 x 35 cm2 (Tabel 3.8).
Figur 3.14. Antal frugter/frugtanlæg per sideskud opgjort i hegn sprøjtet med cypermethrin gentagne gange indtil frugtsætning. Der er signifikant forskel i antallet af reproduktive enheder på de tidspunkter som er markeret med *. Tabel 3.8. Gennemsnitligt antal bær i 35*35 cm2 rammer på træer med eller uden bortsprøjtning af insekter.
3.4.2 Forekomst af insekter og deres betydning for frugtsætningenRegistreringen af insekter forekommende i hegn 5 i perioden fra den 23. maj til 17. juli i henholdsvis insekticidsprøjtede og usprøjtede træer er præsenteret i Tabel 3.9. Disse indsamlinger viser, at der kun var få arter, der var almindeligt forekommende i alle indsamlinger i hegnet. Af snudebiller, der er kendt for at angribe tjørnens reproduktive enheder, blev de to Anthonomus arter A. sorbi og A. pedicularis registreret. Den sidste blev set både i en første og en anden generation medens A. sorbi overvejende blev registreret i anden generation. Dette indikerer, at arten har været til stede tidligere uden at blive registreret. Begge arter angriber tjørnens blomsterknopper hvori larven udvikles og falder til jorden med knoppen, her finder forpupning og forvandling sted. De adulte anden-generationsbiller blev registreret den 17. juli i ret store antal på de usprøjtede tjørn. Den tredje og sidste art Rhynchites aequatus angriber tjørnens frugtanlæg. Denne art blev kun set få gange, og det må formodes at den i dette hegn i år 2003 har spillet en mindre rolle for den formindskede frugtsætning i de usprøjtede tjørn. I andre hegn har vi specielt i år 2001 og 2002 observeret R. aequatus meget hyppigt, så der er næppe tvivl om at denne art også kan have betydning for tabet af reproduktive enheder. Tjørnens bladbille Lochmaea crataegi var hyppig på de usprøjtede træer den 23. maj (Tabel 3.9). Hyppigst blev den voksne bille set spisende af blomsterknopperne, hvor det sikkert er det proteinrige pollen i støvknapperne, som er målet for dens herbivori. Adfærden er givetvis en del af forklaringen på, at frugtsætningen var større i de insekticidsprøjtede områder end i de usprøjtede. Den tidlige tilstedeværelse af Anthonomus-arterne og tjørnenes bladbille er sammenfaldende med det største tab af reproduktive enheder; nemlig tab af blomsterknopper og blomster i perioden mellem 21. maj og 6. juni (Figur 3.14). Tjørnens bladloppe Psylla peregrina blev fundet i perioden mellem 26. juni og 17. juli. Antallet af nymfer og voksne individer steg frem gennem perioden både i de sprøjtede og i de usprøjtede træer. Mængden af insekter var dog betydeligt højere i de usprøjtede end i de sprøjtede (Tabel 3.9). Den eneste kendte effekt af disse insekter er en kortere afstand (reduceret længdevækst) mellem de enkelte sideskud (Sutton 1984). Vi antager derfor, at disse insekter kun spiller en minimal rolle for nedfaldet af reproduktive enheder. 3.4.3 Insekticideffekt på insekt-diversitetBåde individantal og artsantal var lavest i indsamlingerne fra de sprøjtede træer (Tabel 3.9). Artsantallet varierede således mellem 11 og 28 for de usprøjtede og mellem 4 og 12 for de sprøjtede træer. Procentuelt udgjorde artsantallet i de sprøjtede træer mellem 16 og 73% af arterne i de usprøjtede. Totalt set for alle registreringer var antallet af arter i de sprøjtede træer 43% af antallet af arter i de usprøjtede. For individantallet var der over perioden målt mellem 94 og 268 individer på de usprøjtede, medens der på de sprøjtede var mellem 25 og 58 individer pr.registrering. Procentuelt varierede antallet af individer for de sprøjtede mellem 14 og 33% af de usprøjtede. Totalt set udgjorde individantallet i de sprøjtede 20% af individantallet i de usprøjtede. Medens det ikke er så overraskende, at der er færre arter og individer der, hvor der blev sprøjtet, så er der dog nogle få "arter", der udviser den modsatte tendens. Dette gælder 2 grupper (blandt andet bladhvepsene) under Hymenoptera, hvis individantal generelt lå væsentligt højere i de sprøjtede end i de usprøjtede (Tabel 3.9). En mulig forklaring på dette er, at disse arter/grupper tiltrækkes af den større mængde af uangrebne reproduktive enheder, samtidig med at de i mindre grad end deres prædatorer påvirkes af insekticidet. Endelig kunne det også være en tilfældighed afstedkommet af de få observationer, der er af disse to arter. Tabel 3.9 Tabel over de insekter der blev indsamlet i hegn 5 i insekticidsprøjtede henholdsvis usprøjtede dele af hegnet. U og S indikerer optællinger fra henholdsvis usprøjtede og sprøjtede træer. De markerede insekter er de arter der er omtalt i teksten.
4 Diskussion
Dette afsnit er opbygget således, at i afsnit 4.1. præsenteres en kort diskussion af mulige årsager til de mange nul-observationer i datasættene for bær. Disse fik stor betydning for den måde, databehandlingen blev udført på, og de medførte også, at vi supplerede indsamlingen af de modne bær på sideskuddene med optællinger i rammer. Strukturen af data for bær har blandt andet gjort, at vi ikke har kunnet foretage en statistisk sammenligning af effekter på bær i første og andet år. I afsnit 4.2. diskuteres årsager til variation i den afsatte dosis der blev målt, samt litteraturværdier vedrørende afsætning af sprøjtemiddelafdrift på vertikale mål placeret i forskellig højde og afstand fra et eller flere sprøjtespor. Dette afsnit er medtaget for at relatere vores resultater for sprøjtemiddelafsætning ved direkte sprøjtning på hegnet til værdier, der er opnået i forbindelse med afdrift fra marksprøjtning. I afsnit 4.3 og 4.4 diskuteres effekter i henholdsvis eksponeringsåret og året efter eksponering. Diskussionerne, som har hovedfokus omkring effekterne på bærmængden, kombinerer de opnåede målinger af effekter, bærrenes fordeling med højden og litteraturværdier for afsætning af sprøjtemiddelafdrift fra marksprøjtning i et hegn. Derved bliver det muligt at give et bud på den samlede effekt af sprøjtemiddelafdrift ved marksprøjtning med den af producenten oplyste anbefalede maksimale dosis. I afsnit 4.4 foretages en procentuel sammenligning af effekterne året efter sprøjtning med effekterne i eksponeringsåret. Endelig følger i afsnit 4.5 og 4.6 diskussion af årsager til bærmængdens fordeling over højden i hegn (afsnit 4.5), en kort diskussion af projektets insektobservationer samt af insekternes betydning for frugtsætningen (afsnit 4.6). 4.1 BærtabDet første år viste der sig at være en meget høj andel af nul-observationer i datasættet for grønne bær og modne bær. Dette var både tilfældet blandt kontrollerne og behandlingerne. Årsagen til 0-observationerne i kontrollerne er ikke kendt med sikkerhed. Vi ved således ikke, om det er en normal situation, eller om vi har været udsat for en kombination af klima og herbivortryk, som har medført et usædvanligt stort tab af frugter. Fra vores observationer af nedfaldne enheder i spandene under hegnet kan vi se, at der har været et stort nedfald i knop- og afblomstringsstadiet, som både skyldes insektangreb og anden årsag, formentlig abortering. Året efter sprøjtning var der en endnu højere andel af nul-observationer i datasættene for grønne og modne bær, både for kontroller og behandlinger. Den høje andel af nul-observationer i kontrollerne indikerer, at det er normalt, at der forekommer et stort tab af bær forårsaget af andre årsager end afdrift af herbicider. Forsøget med insekticidsprøjtning i et enkelt hegn viser, at herbivorerne udgør en stor del af forklaringen, idet fjernelse af herbivorerne i det pågældende hegn i det pågældende år medførte, at ca. 40% af knopperne udvikledes til modne frugter. 4.2 Dosis og afsætningSammenhængen mellem nominel dosis og afsat dosis var meget klar for det enkelte hegn (Tabel 3.2). Der var dog stor variation mellem enkelthegn og mellem henholdsvis forårs- og sommersprøjtning. Der kan tænkes flere årsager til dette: Varierende vejr under udsprøjtning, hegnsmorfologisk variation eller tilfældig variation som følge af bladvalg. Stor variation i afsætning ved sprøjtning er kendt fra andre undersøgelser, eksempelvis fandt (Nordbo et al. 1993) en variation på en faktor 10 ved afsætning på vertikale mål ved sprøjtning med en konventionel marksprøjte, se iøvrigt også næste afsnit samt Tabel 4.1. Det er væsentligt for værdien af vores forsøg, at afsætningen ligger inden for det interval, der er observeret i studier, der benyttede vertikale mål til at estimere/måle afdrift. Hovedparten af disse studier er gennemført med konventionelle fladdyser. Nordby og Skuterud (1975) fandt, at afsætningen i en afstand på 2,5 m fra sprøjtebommen varierede mellem 0,8 og 22 % af den teoretiske afsætning under bommen. Den mindste værdi blev målt ved lav bomhøjde (40 cm), lav vindhastighed (1,15 m s-1) og lavt sprøjtetryk (2,5 bar), medens den højeste værdi blev målt ved høj bomhøjde (80 cm), høj vindhastighed (4 m s-1) og højt sprøjtetryk (10 bar). Generelt var afsætningen højere end de 0,8 %, idet typiske værdier for afsætningen lå mellem 2 og 5% af afsætningen under sprøjtebommen, når enten bomhøjde, sprøjtetryk eller vindhastighed steg. I en afstand på 25 m var afsætningen generelt mindre end 0,5% af afsætningen under sprøjtebommen, og 50 m fra sprøjtebommen var afsætningen mindre end 0,1%. Longley & Sotherton (1997) målte afsætninger af deltamethrin på mellem 1 og 7% af den nominelle afsætning i en afstand på 1 m fra sprøjtebommen, dvs. afsætningen hvis alt sprøjtemidlet fordeles ligeligt over en jævn flade, således at afsætningen per ha bliver lig med udspredningen per ha. Resultater fra andre undersøgelser viser sammen med resultater fra Nordby og Skuterud (1975), hvorledes afsætningen på lodrette mål varierer med afstanden fra sprøjtebommen (Tabel 4.1). Tabel 4.1 Undersøgelser af målinger af sprøjtemiddelafsætning på lodrette mål i forskellige højder over jorden ved afdrift fra marksprøjtning i et sprøjtespor målt som procent af udsprøjtet. Værdierne givet i parantes markere yderpunkterne for observationerne.
Det er væsentligt at være opmærksom på, at der kan være en forskel på afsætningen i et hegn og afsætningen på curlere eller andre opsamlere placeret i luftstrømmen, idet luften delvis strømmer over hegnet delvis strømmer gennem hegnet (Davis et al. 1994). Således vil afsætning målt i hegnet typisk være reduceret i forhold til afsætning målt på opsamlere placeret i luftstrømmen. Jo mindre gennemtrængelig hegnet er, jo større del af luftstrømmen vil gå over hegnet, og afsætningen i hegnet vil blive tilsvarende reduceret (Davis et al. 1994). Weisser et al (2002) præsenterer målinger af afdrift fra marksprøjtning til hegn fordelt over højden (Tabel 4.1 og Figur 4.1). Ydermere har Parkin og Merritt (1988) ved målinger af sprøjtemiddelafsætning i højdeintervallerne 0-3, 3-6, 6-9 og 9-12 meter i forskellig afstand fra sprøjtebommen vist, at der allerede 10 m fra bommen er målbare mængder i 9-12 meters højde. Disse er i størrelsesordnen 10% af afsætningen i 3-6 meters højde. Mængden i 9-12 meters højde blev mere end femdoblet ud til en afstand på 30 m fra sprøjtebommen, hvorefter den ved 50 meters afstand aftog til ca. 4 gange mængden i 10 meters afstand. I de øvrige højder aftog mængden af sprøjtemiddel jævnt med afstanden, således at afsætningen i 50 meters afstand i de ovenfor nævnte højdeintervaller lå inden for en faktor 2, med 9-12 meter som minimum og 3-6 meter som maksimum. Den samlede afsætning (0-12 meter) i 50 meters afstand var ca. 40% af afsætningen i 5 meters afstand og 60% af afsætningen i 10 meters afstand fra sprøjtebommen. Den forholdsvis store afsætning med afstanden bekræfter, at der ved afsætning fra flere sprøjtespor kan være tale om en væsentlig forøgelse i forhold til afsætning fra et enkelt sprøjtespor, ligesom det ved flere sprøjtespor må forventes, at en større andel af den afsatte mængde sprøjtemiddel afsættes højere oppe i hegnet (Parkin & Merritt 1988). Til sammenligning var gennemsnitsafsætningen i vores forsøg ved 5% nominel dosis 1,6% af den på etiketten angivne dosis med et minimum på 0,4% og et maksimum på 2,5%. Ved 10%, 20% og 40% var den henholdsvis 3,2% (0,8-5,08%), 6,5% (1,61-15,81%) og 12,9% (3,22-31,62%)(Tabel 3.2). Disse værdier ligger alle inden for værdier for afsætninger, som kan måles i forbindelse med afdrift af pesticider under feltbetingelser, selvom de 12,9% ligger i den høje ende (Tabel 4.1). Det skal understreges, at værdierne i tabel 4.1 er målt i forbindelse med afdrift fra ét sprøjtespor. Et højere antal sprøjtespor forøger størrelsen af afdriften (Gilbert & Bell 1988, Yates et al. 1978); Maybank et al. 1978, Nordby & Skuterud 1975). I de tilfælde, hvor marken bliver sprøjtet med den maksimalt dose angivet på etiketten, er afsætningen ved de øvrige doser at betegne som normalt forekommende i forbindelse med udbringning af pesticider til marker med omgivende hegn. Det indikerer samtidigt, at der er grund til at forvente, at hegnene i det åbne land er påvirket af afdrift af pesticider fra marksprøjtninger. 4.3 Sprøjtemiddeleffekter i eksponeringsåretBærrenes store følsomhed for sprøjtning i eksponeringsåret (Figur 3.1 og 3.2) stemmer godt overens med observationer foretaget af Bhatti et al. (1995) på fuglekirsebær. Forskellen på forårs- og sommersprøjtning gik i retning af at effekten af sprøjtning var størst, når den fandt sted om foråret. Årsagen er formentlig, at jo tidligere man påvirker transporten af fotosynteseprodukter til frugtanlægget, jo større er muligheden for at udviklingen til færdig frugt kan forringes. Beregning af effekten på hegnsniveau forudsætter viden om følgende tre forhold:
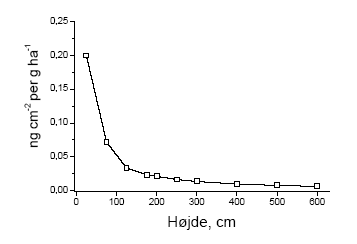
Det første forhold har vi selv etableret i forbindelse med sprøjteforsøget, det andet forhold har vi ikke undersøgt, men en undersøgelse fra Tyskland præsenterer data for sprøjtemidlets fordeling over højden 0 – 2 m (Weisser et al. 2002). Disse data giver mulighed for at ekstrapolere afsætningen til andre højder (Figur 4.1). Det tredje forhold har vi selv etableret ved optælling af bærmængden i rammer fordelt over højden (Fig. 3.12-3.14). Weisser et al. (2002) undersøgte afsætningens fordeling over højden i 50 cm's intervaller fra 0 – 2 m ved sprøjtning med konventionelle dyser (XR 110 03 med et arbejdstryk på 2 bar). Undersøgelsen angiver standardafvigelsen på middelværdien til at være 140% i 0-50 cm og jævnt aftagende med højden til 69,7% i 150 – 200 cm. Der er altså en stor variation i afsætningen selv inden for det samme hegn i forbindelse med en afdriftshændelse. Dertil kommer at undersøgelser af afsætningen under andre betingelser (andre hegn, andre sprøjtemidler, andet sprøjteudstyr, samt andre vejrforhold) viser at variationen kan være endnu større. Dertil kommer at selve opgørelsen af bærrene er behæftet med betydelig variation. Blandt andet af disse grunde er den følgende beregning mere et regneeksempel end et bud på et generelt billede. Ud over ovenstående tre forhold er der et fjerde forhold som vi ønsker at inddrage. Dette er effekten af flere sprøjtespor i marken hvorfra afdrift forekommer. De få undersøgelser der sammenligner afdriften fra 1 henholdsvis 10 sprøjtespor viser at afsætningen i omgivelserne stiger med en faktor 3-10 (Gilbert & Bell 1988, Yates et al. 1978, Maybank et al 1978, Norby & Skuterud 1975). I de følgende beregninger har vi valgt at anvende resultater fra en undersøgelse foretaget af Gilbert og Bell (1998), som viser hvorledes afdriften til et område der grænser op til marken stiger med antallet af sprøjtespor. Ved 10 sprøjtespor forøges afdriften således med en faktor 3,4 relativt til et enkelt sprøjtespor. Sammenlagt har vi altså en afsætning der aftager med højden, en produktion der tiltager med højden, stor effekt ved små doser og tiltagende afsætning ved flere sprøjtespor. Den samlede effekt af dette efter forårssprøjtning er vist i Figur 4.2.
Figur 4.1. Afsætningens fordelingen over højden i hegn ved sprøjtning grænsende umiddelbart op til hegnet. Højden fra 2 m og opefter er beregnet ved ekstrapolation baseret på de lavere højder (efter Weisser et al. 2002).
Figur 4.2. Beregnet effekt (% reduktion i antallet af bær) af forårsprøjtning med metsulfuron i eksponeringsåret for hegn af varierende højde. Effekten er beregnet for den eksponerede side af hegnet på baggrund af afsætning som funktion af højden (efter Weisser et al 2002) og afdrift fra 10 sprøjtespor (efter Gilbert og Bell, 1988), egne målinger af effekten af metsulfuron på frugtsætningen ved sprøjtning medio maj samt måling af bærrenes fordeling over højden hos tjørn. Konsekvensen af pesticidforårsagede reduktioner i bærproduktionen kan endnu ikke relateres til konsekvenser for frugivore fugle, idet vi ikke kender omfanget af brugen af reducerede doser og dysetyper med et andet driftmønster. Udover afdrift af sprøjtemidler er der andre påvirkninger af hegnet, som kan medføre at den potentielle produktion nedsættes. Disse kan være naturlige eller menneskeskabte. De naturlige inkluderer klimatiske forhold og herbivore organismer. En af de væsentligste menneskeskabte påvirkninger er beskæring af hegnene. Hyppig beskæring af de fertile skud påvirker reproduktionen, da frugtsætning kun finder sted på 2-årige eller ældre skud. Croxton og Sparks (2002) fandt således i en systematisk undersøgelse af effekten af beskæring ved årlig, biannuel og ingen eller meget lille beskæring, at forskellen mellem de hyppigt beskårne og de ikke beskårne hegn var op til en faktor 50, med den største produktion i de ikke beskårne. Undersøgelserne fandt sted på tre lokaliteter i England med fem hegn af hver type på hver lokalitet. 4.4 Sprøjtemiddeleffekter året efter eksponeringAfdrift af metsulfuron medfører en reduktion af frugtsætningen hos tjørn, som rækker ud over det år (den sæson), hvor afdriften finder sted. Dette stemmer godt overens med amerikanske erfaringer fra bekæmpelse af opvækst af tjørn med stofferne imazapyr og metsulfuron. Her angives det, at stofferne kan bruges til kontrol af opvækst af buske, men at de ultimative effekter undertiden først ses året efter sprøjtning (Sprague & Hager 2003). Det er iøvrigt værd at bemærke, at der i andet år er negative effekter på alle målvariable. Dette er i modsætning til effekterne i eksponeringsåret, hvor der kun var betydende effekter på bærmængden. Figur 4.3 viser den relative effekt på bærmængden i henholdsvis år 1 og år 2. Grunden til at påvirkningen af knopper og blomster var lille det første år skyldes sikkert, at der på sprøjtetidspunktet allerede var allokeret tilstrækkeligt med ressourcer til disse organer, til at de kunne udvikles. Det modsatte var så tilfældet året efter eksponering, hvor effekten af det første års sprøjtning medførte en formindsket allokering til knopstadierne, så de dels fik en ringere start og formentlig også en formindsket tilførsel af fotosynteseprodukter ved udspring i det andet år. Det ses, at sommersprøjtningen giver effekter af samme størrelsesorden i både år 1 og år 2. Forårssprøjtningen er derimod meget mere effektfuld i eksponeringsåret end i året efter. Denne forskel betyder også at den effekt på hegnets bærmængde, som er præsenteret for forårssprøjtningen i Figur 4.2, bliver mindre efter sommersprøjtningen og året efter. Det, at der er et tab året efter eksponering, viser først og fremmest, at effekterne af sprøjtningen er langvarige. Graden af recovery antyder at effekterne varer mere end de to år, vi har kigget på, i det mindste ved afsætninger større end 0.004 μg cm-2, hvor der kun er ca. 25% af bærmængden relativt til kontrollen (Figur 4.3). I figur 4.3 er ikke medtaget data for hegn 1 i år 2. Det har vi valgt, fordi kontrollen i 10 ud af 13 tilfælde ikke havde nogen bær overhovedet, som det kan ses i figur 3.10. Vi kender ikke årsagen hertil, men at inddrage 2 punkter i hver graf med værdier omkring 400-500% giver et skævt billede af tendensen i data. Dette hænger i øvrigt også sammen med, at variationen aftager med stigende eksponering, idet der i kontrollen og ved de laveste eksponeringer kan optræde andre årsager end metsulfuron til at alle bær tabes. Denne variation forsvinder ved de høje doseringer på grund af påvirkningen fra metsulfuron.
Figur 4.3 Sammenligning af den procentuelle effekt på bærmængden af metsulfuronsprøjtning i eksponeringsåret og året efter. Data benyttet er antallet af bær i sideskud i år 1 og antal bær i en ramme i år 2. Der kan tænkes en længere række af årsager til, at der opstår effekter året efter sprøjtning. Det er meget muligt, at det ikke bare er en af dem, der er den korrekte, men at forklaringen snarere skal søges i den samtidige virkning af flere af påvirkningsmåderne. Disse er opremset i det følgende med en summarisk forklaring på, hvordan virkningen kan tænkes at opstå:
Varigheden af effekten afhænger af hvilken påvirkningsmåde, der er tale om. Rester af metsulfuron i vævet vil f.eks. have en begrænset levetid i forhold til forekomsten af dødt væv. I det sidstnævnte eksempel vil effekten vare, indtil det døde væv er erstattet af nyt væv i form af dannelse af nye grene. Det er klart, at varigheden derfor også afhænger af dosis, i og med at en større dosis både vil kunne føre til en større restmængde af aktivstoffet i vævet og en større mængde dødt eller beskadiget væv. Vi iagttog en tendens til, at effekten af sprøjtemidlet året efter eksponering var mere signifikant i de hegn, der støder op til landbrugsmæssigt drevne områder end i de hegn, der støder op til usprøjtede græsarealer. Umiddelbart kunne det få en til at tro at hegn, der ligger op til landbrugsområder, er mere følsomme. Forklaringen kan dog også være, at reduktionen som følge af herbivore insekter er større i de hegn, der ligger op af græsområder. En anden mulighed, som indeværende undersøgelse understøtter, er at de hegn, der ligger i umiddelbar nærhed af landbrugsområder, allerede er påvirket af afdrift af andre sprøjtemidler fra tidligere år. Derudover er det muligt, at spredning af kunstgødning på de landbrugsmæssigt drevne arealer kan have en positiv effekt på tjørns frugtsætning, hvis kunstgødningen spredes ud under hegnet. Herved vil træer, der ligger op til landbrugsarealer der har et overskud af næringsstoffer, have en større vækstrate end andre træer. Data for enårige planter viser, at en forøget vækst forårsaget af næringstilførelse vil afstedkomme større forskel mellem behandlinger, idet mængden/antallet af kontrollens målvariable forøges, hvorved forskellen til behandling kan blive større reelt såvel som relativt (Stephenson et al. 2001). 4.5 Bærproduktionen med højdenIagttagelsen af at bærmængden stiger med højden på sydligt og vestligt orienterede sider af tjørnehegn kan have flere forklaringer. Vi kan ikke udfra vores data bedømme hvilke af disse, der er de vigtigste, men forventer dog at de alle bidrager. Der vil afhængig af hegnets facon være mere lys jo højere man kommer op i hegnet. Øget lysmængde fører igen til øget produktion. Det er muligt, at der som udgangspunkt er lige mange frugtanlæg pr. arealenhed uanset placering i hegnet, men at herbivorpresset på knopper og blomster aftager med højden. Dernæst kan forskellene skyldes husdyr eller vildt som overvejende spiser af blade og knopper i hegnets nedre dele. Forskellen kan også skyldes vedligeholdelse af hegnet hvor det typisk er hegnets nedre dele som beskæres eller klippes. Det er en nærliggende tanke at en del af forskellen kunne skyldes afsætning af herbicider fra marksprøjtning, som netop aftager med højden. Endelig kan det rent udviklingsmæssigt tænkes, at der kan være tale om at det er en selektiv fordel at have bærrene højt oppe, hvor de ses lettere af fugle og derved har større chance for at blive spredt. 4.6 InsekterÅrsagen, til at der ikke kunne måles en effekt af herbicidsprøjtning på nedfaldet af reproduktive enheder i den første sæson, kan skyldes at det kun var en del af hegnet over de spande, hvori nedfaldet blev opsamlet, som blev sprøjtet. Dermed skete der en fortynding af nedfaldet, som skjuler eventuelle sprøjteeffekter, som ellers ville kunne være iagttaget i nedfaldet. Herudover, var der i perioder et stort nedfald som følge af herbivore insekter, der også kan have skjult herbicideffekter. Den gentagne sprøjtning med insekticid på træer i hegn 5, der blev gennemført i år 2, bidrager med oplysninger på to fronter: 1) det medvirker til at forbedre vurderingen af, hvor stor en del af bærtabet der skyldes insekter, 2) det giver et første bud på, hvad afdrift af insekticider til tjørnehegn betyder for mængde og diversitet af insekter i tjørnehegn. Vores observationer viser, dog med den begrænsning at de kun er insamlet i et enkelt hegn og et enkelt år, at insekter, specielt snudebiller af slægterne Rhyncites og Anthonomus samt bladbillen Lochmea crataegi, forårsager store tab af knopper og frugtanlæg. Men uanset omfanget af insektforårsaget reduktion i frugtsætning vil afdrift af herbicider forårsage en betydelig reduktion af den resterende bærmængde. Vi ved dog ikke om tabet af bær som følge af herbicidafdrift ville være procentuelt anderledes, hvis der ikke var tab som følge af insektangreb. 5 Konklusioner
5.1 Undersøgelsens resultatmæssige konklusionerSprøjtemiddeleffekter i eksponeringsåret:
Sprøjtemiddeleffekter året efter eksponering:
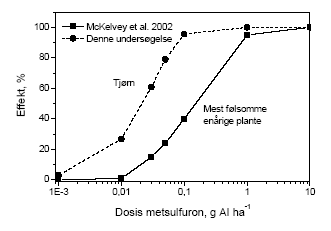
Engriflet hvidtjørn er en vigtig hegnsplante i Danmark, der producerer en stor mængde bær, som dels ædes af solsorte, dels af millioner af trækkende drosler fra Skandinavien (Snow & Snow 1988; Kjær et al. 2002). Det må formodes, at mængden af tjørnebær er af stor betydning både for trækfuglene og solsortenes overlevelse. Det er derfor sandsynligt, at afdrift af ukrudtsmidler ved at påvirke tjørns bærsætning har stor betydning for fuglenes overlevelse i vinterhalvåret. Yderligere er der mange arter af insekter, der er knyttet til tjørn (Kennedy & Southwood 1984). Blandt disse vil det først og fremmest være dem, der er knyttet til andre dele end bladene, der bliver påvirket. 5.2 Faglige perspektiver og forskningsbehovOmend resultatet ikke er overraskende, så er det forskningsmæssigt regi nyt, at metsulfurons virkning strækker sig til året efter eksponeringen har fundet sted. Derudover bekræftes tidligere rapporterede effekter af sulfonylureastoffer på fuglekirsebær med en anden art og et andet middel. Undersøgelsens resultater peger dermed på et behov for undersøgelse af langtidseffekter af flere forskellige typer af herbicider på flerårige bærbærende arter. Samtidig er der nogen begrænsninger ved den type af feltforsøg, som i nærværende rapport er blevet anvendt for at påvise at der potentielt kan være effekter af afdrift af herbicider til hegn. Således kan kontrollerede feltforsøg, som de i rapporten beskrevne, ikke umiddelbart beskrive konsekvenserne af en afdriftshændelse, da der endnu mangler feltmålinger af afdriftens størrelse i forbindelse med reel marksprøjtning under de varierende betingelser (mht. vindstyrke, vindretning, temperatur) sprøjtningen finder sted. For det første er de gennemført som manipulerede forsøg, hvor sprøjtemidlet er sprøjtet direkte ind i hegnet med en rygsæksprøjte. Dernæst eksponeres hegnet ved afdrift fra mark fra overlappende sprøjtefaner og eventuelt fra flere sider. Endelig bruger mange landmænd reducerede doseringer i forhold til den af producenten anbefalede maksimale dosering. Der er derfor behov for at undersøge om og i hvilken grad bærbærende hegnstræer påvirkes af afdrift af metsulfuron fra mark til hegn samt for at vurdere konsekvenser af forskellige måde at forvalte sprøjtningen på. For at bestemme omfanget af afdrift må man kunne måle eller beregne, hvor stor en mængde sprøjtemiddel, der rammer hegnet. Dette kunne gøres ved at måle sprøjtemiddelafsætningen i hegn efter en realistisk sprøjtning med flere overlappende sprøjtespor. Udfra disse data vil der kunne udvikles en model til at forudsige afsætningen under varierende betingelser. Efterfølgende vil en sådan model kunne udnyttes til at beregne effekter af metsulfuronsprøjtning. Udvidelse af modellen til at dække andre hegnsarter end tjørn vil kræve undersøgelser af disse. Derudover er der et behov for at undersøge effekten af sprøjtemidler med andre virkningsmekanismer både i eksponeringsåret og året efter eksponering. Resultater fra sådanne projekter vil kunne understøtte en vurdering af, i hvilket omfang ukrudtsmiddelafdrift påvirker bærbærende hegnstræer. Hertil kommer en vurdering af, om en ændring af sprøjtemiddelanvendelsen vil resultere i øget fødemængde for bærspisende organismer i hegn, specielt fugle, under danske vilkår. Af samme grund kan der heller ikke på nuværende tidspunkt siges noget endegyldigt om i hvilken grad den fauna, der er knyttet til tjørnehegn og andre hegn, påvirkes, kun at det er sandsynligt at der er en påvirkning og at den kan være betydelig. Ved at fokusere på hvorledes variation af sprøjtemiddel, sprøjtedosering, dysetype, vejrtype og afstand fra markkant til hegn vil påvirke bærmængden, kan der dannes basis for at vurdere en eventuel gevinst ved administrativ regulering af sprøjtemiddelanvendelsen (se afsnit 5.3). 5.3 Perspektiver for reguleringen af sprøjtemidlerNye sprøjtemidler og sprøjtemidler under revurdering skal blandt andet vurderes for deres økotoksiske virkning overfor terrestriske organismer. Denne vurdering baseres normalt på laboratorietest med mikroorganismer, regnorme, bier, fugle, pattedyr og eventuelt leddyr. Leddyr omfatter primært nyttedyrsfaunaen i marken. I den europæiske lovgivning omfattes disse sammen med planter under "Other non-target organisms (flora and fauna) believed to be at risk". Der er ikke noget krav om, at der laves en vurdering af non-target plantearter. I oktober 2002 blev et udkast til et nyt "Guidance document on terrestrial Ecotoxicology " offentliggjort. Dette fungerer som et baggrundsdokument for direktiv 91/414/EEC. Heri foreslås, at planter testes enten som foreslået i den reviderede OECD plante test guideline eller ifølge OPPTS ("Office of Prevention, Pesticides and Toxic Substances") test guidelines, der er offentliggjort af den amerikanske miljøstyrelse. Resultaterne i nærværende rapport indikerer tydeligt, at effekter på ikke-måls planter (her forstået som de der findes uden for det dyrkede areal) må, ved den på etiketten angivne maksimal-dosis, forventes som et resultat af afdrift af ukrudtsmidler. De understøtter hermed også synspunktet, at plantetest er vigtige. De umiddelbare effekter af metsulfuron (første år) var størst for frugtsætningen. Det viser, at der findes ukrudtsmidler, der påvirker reproduktionen mere end de påvirker den vegetative vækst. De test, der er foreslået i det ovenfor nævnte "guidance document" vil ikke dokumentere sådanne effekter, idet alle test er "tidlig vækst" test, hvor testplanten ikke når til det reproduktive udviklingsstadie før testen afsluttes. Herudover udføres disse test på enårige planter. Sammenligning af vores data med data for metsulfurons virkning på "den mest følsomme enårige plante" præsenteret af McKelvey et al. (2002) understreger vigtigheden af at overveje implikationer af sprøjtemiddelafdrift på flerårige plantearter og på reproduktion (Figur 4.5), idet de refererede data i McKelvey et al. (2002) er vækstmålinger.
Figur 4.5 Sammenligning af følsomheden overfor metsulfuron hos "den mest følsomme enårige plante" som præsenteret af (McKelvey et al. 2002) og hos engriflet hvidtjørn Crataegus monogyna. Dette behov understreges yderligere af, at tydelige effekter kan observeres året efter den aktuelle eksponering har fundet sted. I forbindelse med Figur 4.5 er det vigtig at bemærke, at data for de enårige planter formodentlig refererer til den ønskede dosis, hvorimod data for tjørn er opgivet som den dosis, der er afsat på bladene. Dette betyder, at det må forventes, at de enårige planter har modtaget mindre sprøjtemiddel end angivet, og de to kurver vil nærme sig hinanden. Jagers op Akkerhuis et al. (1998) fandt at en afgrøde modtager op til 20% af den udsprøjtet dose på bladene Hvis dette også gælder for den enårige plante (agurk) vil dosis - respons kurven stadig ligge til højre for kurven for tjørn. Ved en effektvurdering vil det være væsentligt at inddrage afdriften fra flere sprøjtespor og depositionens fordeling over højden. LitteraturAl-Khatib K., Parker R. & Fuerst E. P. (1992) Sweet cherry (Prunus avium) response to simulated drift from selected herbicides. Weed Technology 6: 975- 979. Arvidsson T. (1997) Spray drift as influenced by meteorological and technical factors a methodological study. Swedish University of Agricultural Sciences Uppsala 1997, Doctors Dissertation in Agraria 71: 1-144. Babcock J. M., Brown J. J. & Tanigoshi L. K. (1990) Volume and coverage estimation of spray deposition using an amino nitrogen colorimetric reaction. Journal of Economic Entomology 83: 1633-1635. Bestman H. D., Devine M. D. & Vanden Born W. H. (1990) Herbicide Chlorsulfuron Decreases Assimilate Transport Out of Treated Leaves of Field Pennycress (Thlaspi arvense L.) Seedlings. Plant Physiol. 93: 1441- 1448. Bhatti M. A., Al-Khatib K., Felsot A. S., Parker R. & Kadir S. (1995) Effects of simulated chlorsulfuron drift on fruit yield and quality of sweet cherries (Prunus avium L.). Environmental Toxicology and Chemistry 14: 537-544. Byass J. B. & Lake J. R. (1977) Spray drift from a tractor-powered field sprayer. Pestic. Sci. 8: 117-126. Christensen K. D., Jacobsen E. M. & Nøhr H. (1996) A comparative study of bird faunas in conventionally and organically farmed areas. Dansk Ornithologisk Forenings Tidskrift 90: 21-28. Croxton P. J. & Sparks T. H. (2002) A farm-scale evaluation of the influence of hedgerow cutting frequency on hawthorn (Crataegus monogyna) berry yields. Agriculture Ecosystems & Environment 93: 437-439. Davis B. N. K., Brown M. J., Frost A. J., Yates T. J. & Plant R. A. (1994) The effects of hedges on spray deposition and on the biological impact of pesticide spray drift. Ecotoxicology and Environmental Safety 27: 281-293. de Snoo G. R. & van der Poll R. J. (1999) Effect of herbicide drift on adjacent boundary vegetation. Agriculture Ecosystems & Environment 73: 1-6. Esbjerg P. & Petersen B. S. (2002) Effect of reduced pesticide use on flora and fauna in the agricultural fields. Danish Environmental Protection Agency. Fletcher J. S., Pfleeger T. G., Ratsch C. & Hayes R. (1996) Potential impact of low levels of chlorsulfuron and other herbicides on growth and yield of nontarget plants. Environ. Toxicol. Chem. 15: 1189-1196. Freemark K. & Boutin C. (1995) Impacts of agricultural herbicide use on terrestrial wildlife in temperate landscapes: A review with special reference to North America. Agriculture Ecosystems & Environment 52: 67-91. Gilbert A. J. & Bell G. J. (1988) Evaluation of the drift arising from spray application. Aspects of Applied Biology 17: 363376. Hald A. B., Reddersen J. & Elbek-Pedersen H. (1994) Sprøjtefri randzoner i sædskiftemarker - Plante- og insektliv samt udbytte. Landsforsøg 1987-92. Miljøstyrelsen, København. Hald A. B., Reddersen, J. (1990) Fugleføde i kornmarker - insekter og vilde planter. Undersøgelser på konventionelle og økologiske landbrug 1987-88 pp. 1-107. Miljøstyrelsen, Copenhagen. Jagers op Akkerhuis G. A. J. M., Axelsen J. A. & Kjær C. (1998) Towards predicting pesticide deposition from plant phenology; a study in spring barley. Pestic. Sci. 53. Kennedy C. E. J. & Southwood T. R. E. (1984) The number of species of insects associated with British trees: A reanalysis. Journal of Animal Ecology 53: 455-478. Kjær C., Strandberg M. & Elmegaard N. (2002) Effekter af sprøjtemiddelafdrift på bærbærende buske og træer som indikator for biodiversitetsforandringer. Bekæmpelsesmiddelforskning 60. Kleijn D. & Snoeijing G. I. J. (1997) Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34: 1413-1425. Longley M., Cilgi T., Jepson P. C. & Sotherton N. W. (1997) Measurements of pesticide spray drift deposition into field boundaries and hedgerows: 1: Summer application. Environmental Toxicology and Safety 16: 165-172. Longley M. & Sotherton N. W. (1997) Measurements of pesticide spray drift deposition into field boundaries and hedgerows: 2. Autumn applications. Environmental Toxicology and Safety 16: 173-178. Marrs R. H. & Frost A. J. (1997) A microcosm approach to the detection of the effect of herbicide spray drift in plant communities. Journal of Environmental Management 50: 369-388. Maybank J., Yoshida K. & Grover R. (1978) Spray drift from agricultural pesticide applications. Journal of the Air Pollution Control Association 28: 1009- 1014. McKelvey R. A., Wright J. P. & Honegger J. L. (2002) A comparison of crop and non-crop plants as sensitive indicator species for regulatory testing. Pest Management Science 58: 1161-1174. Nordbo E., Kristensen K. & Kirknel E. (1993) Effects of Wind Direction, Wind Speed and Travel Speed on Spray Deposition. Pesticide Science 38: 33- 41. Nordby A. & Skuterud R. (1975) The effects of boom height, working pressure and wind speed on spray drift. Weed Research 14: 385-395. Parkin C. S. & Merritt C. R. (1988) The measurement and prediction of spray drift. Aspects of Applied Biology 17: 351-361. Reddersen J., Elmholt S. & Holm S. (1998) Indirect Effects of Fungicide and Herbicides on Arthropods. Danish Environmental Protection Agency, Copenhagen. Snow B. & Snow D. (1988) Birds and berries - a study of an ecological interaction. T & A D Poyser, Calton. Sprague C. L. & Hager A. G. (2003) Brush control in Illinois. In: Illinois agricultural pest management handbook - 2003. University of Illinois Extension, College of Agricultural, Consumer and Environmental Sciences. Stephenson G. L., Feisthauer N. C., Koper N., McCann J. H. & Scroggins R. P. (2001) The Influence of Four Types of Water on Seedling Emergence and rowth of Barley and the Toxic Interaction with Copper Sulfate. Standard Technical Publication 1403: 134-145. Sutton R. D. (1984) The effect of host plant flowering on the distribution and growth of hawthorn psyllids (Homoptera:Psylloidea). Journal of Animal Ecology 53: 37-50. Topping C. J. & al. e. (in press) Monitering af de miljømæssige effekter på det terrestriske miljø ved reduktion af pesticidanvendelsen. Vanden Borne W. H., Bestman H. D. & Devine M. D. (1988) The inhibition of assimilate translocation by chlorsulfuron as a component of its mechanism of action. Proc. EWRS symp.: 69-74. Weisser P., Landfried M. & Koch H. (2002) Off-crop drift sediments on plant surfaces - exposure of non-target organisms. Aspects of Applied Biology 66: 225-230. Yates W. E., Akerson N. B. & Bayer D. E. (1978) Drift of glyphosate sprays applied with aerial and ground equipment. Weed Science 26: 597-604. Bilag A Forhold under forårssprøjtning
Bilag B Forhold under sommersprøjtning
Bilag C Datagrundlaget for eksponeringsåret
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||