Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 105, 2006
Ukrudtsstriglingens effekter på dyr, planter og ressourceforbrug
Indholdsfortegnelse
- 2.1 Ukrudtsstrigling som alternativ til herbicider
- 2.2 Hvorfor undersøge effekter på fugle, hvirvelløse dyr, den vilde flora, og landbrugets ressourceforbrug?
- 3.1 Oversigt over projektets delelementer
- 3.2 Pilotprojekt
- 3.3 De eksperimentelle undersøgelser
- 3.4 Modelleringsdel - sanglærke, løbebille, tæppespinder
- 3.5 Systemanalyse af landbrugets ressourceforbrug og driftsøkonomi
- 5.1 Eksperimentelle forsøg
- 5.2 Landbrugets ressourceforbrug
- 5.3 Fauna modellering på Landskabsniveau
- 6.1 Effekter på ukrudt og høstudbytte
- 6.2 Effekter på energiforbrug
- 6.3 effekter på fauna
- 6.4 Samlet vurdering
Bilag A: Beskrivelse af ALMaSS-modelsystemet
Bilag B: Vegetation growth simulation in ALMaSS 4.0
Bilag C: Validation procedures for ALMaSS, and other complex adaptive systems
Forord
Nærværende projekt, der blev udført på Kalø Gods i perioden 2003 – 2006, har været finansieret gennem Miljøstyrelsens pesticidforskningsmidler. Projektet er udført af Afdeling for Vildtbiologi og Biodiversitet og Afdeling for Terrestrisk Økologi (Danmarks Miljøundersøgelser), Institut for Jordbrugsvidenskab (Den Kgl. Veterinær- og Landbohøjskole) og Afdeling for Jordbrugsproduktion og Miljø (Forskningscenter Foulum, Danmarks JordbrugsForskning). Lektor Mogens Erlandsen (Institut for Biostatistik, Århus Universitet) har under projektforløbet været tilknyttet projektet som statistisk konsulent.
En tak skal rettes til medlemmerne af projektets følgegruppe: professor Peter Sandøe (Den Kgl. Veterinær- og Landbohøjskole), sektionsleder Poul Henning Petersen (Dansk Landbrugsrådgivning Landscentret), specialkonsulent Jørn Kirkegaard (Miljøstyrelsen, formand), cand. agro. Lene Gravesen (Miljøstyrelsen), seniorforsker Kristian Kristensen (Danmarks JordbrugsForskning, Forskningscenter Foulum), afdelingsleder Niels Lindemark (Dansk Planteværn), seniorforskere Pernille Kaltoft og Mette Jensen (Danmarks Miljøundersøgelser, Afd. for Systemanalyse), professor Peter Esbjerg og forskningsadjunkter Søren Navntoft og Anne-Mette M. Jensen (Den Kgl. Veterinær- og Landbohøjskole), forsker Jens Erik Ørum (Fødevareøkonomisk Institut, Den Kgl. Veterinær- og Landbohøjskole), biolog Bo Svenning Petersen (Orbicon), professor Jørgen Eivind Olesen, seniorforsker Bent J. Nielsen, seniorforsker Lise Nitrup Jørgensen og seniorforsker Egon Noe (alle Danmarks JordbrugsForskning, Forskningscenter Foulum). Der har i projektgruppen været mange gode faglige diskussioner, og gruppens kommentering af tidligere rapportudkast har været af stor værdi for det færdige resultat.
Under projektperioden har seniorforsker Marianne Bruus Pedersen afløst (marts 2004) forskningsprofessor Martin Holmstrup (begge afdeling for Terrestrisk Økologi, Danmarks Miljøundersøgelser, og lektor Sherow Raza har afløst (juni 2004) adjunkt Christian Bugge Henriksen (begge Den Kgl. Veterinær- og Landbohøjskole). Tak til Annie Laursen og Else-Marie Nielsen (Danmarks Miljøundersøgelser, Afd. for Vildtbiologi og Biodiversitet) som har foretaget hhv. opsætning og korrekturlæsning af rapporten samt Kathe Møgelvang og Tinna Christensen (Danmarks Miljøundersøgelser, Afd. for Ferskvandsøkologi), som har udført det grafiske arbejde i rapporten.
En særlig tak skal rettes til Peter Lange og Tore Kroier, som i foråret og sommeren 2004 og 2005 stod for arbejdet med at følge lærkernes reproduktion på de to forsøgsmarker på Kalø. Uden deres kompetente og utrættelige arbejde havde dette eksperimentelle arbejde ikke kunnet udføres. Ligeledes rettes en stor tak til Elin Jørgensen, Mette Thomson, Karsten Tværmose Andersen og Zdenek Gavor, som har gravet regnorme og mikroleddyr op af den stive lerjord og efterfølgende artsbestemt, talt og vejet dem, samt til landbrugsteknikker Jens B. Kjeldsen der i samarbejde med DJF’s afdeling for Mark- og Stalddrift har haft ansvaret for forsøgshøstning, opgørelse af kerne- og halmudbytte, samt analyserne på Foulums laboratorium.
Projektets eksperimentelle dele var færdige i sensommeren 2005, og den foreliggende afsluttende rapport blev endeligt godkendt af Miljøstyrelsen november 2006. Til slut en tak til vores koner, mænd og kærester, som har måttet bære over med, at vi i perioder mest var gift med vores arbejde!
Sammenfatning
Ukrudtsstriglingens omkostninger og utilsigtede effekter
I nærværende projekt undersøges ukrudtsstriglingens effekter på markfladens naturlige plante og dyreliv samt landbrugets ressourceforbrug. På baggrund af resultaterne fra den undersøgte fauna anbefales højst to striglinger i vår-kornafgrøder, ikke senere end ca. 30 dage efter såning. I vinter-korn bør efterårsstrigling undgås, og der bør kun foretages en forårsstrigling inden ca. midten af april. Striglinger kan udføres uden betydelige påvirkninger af energiforbrug og dækningsbidrag, men betragtes med nuværende teknik - og uden ændringer i sædskiftet - ikke at være et realistisk alternativ til herbicider på konventionelle bedrifter men alene et supplement.
Bichel-udvalgets arbejde behandlede mulighederne for en hel eller delvis afvikling af pesticidanvendelsen i Danmark og de ulemper og fordele der vil være forbundet hermed. Forsøgsarbejde siden da har vist, at ukrudtsstrigling kan indgå som alternativ til herbicider i særlige afgrøder, og i særlige områder hvor pesticidanvendelse måtte ønskes begrænset af hensyn til miljø og natur. Pesticidanvendelse har været en af de væsentlige årsager til den tilbagegang som har været observeret for flere agerlandsfugle i Vesteuropa inden for de seneste to årtier. Pesticidernes negative indvirkning har primært været indirekte gennem en reduktion af vilde planter og leddyr. Ukrudtsstriglinger anvendes stort set ikke i konventionel dyrkning, selvom der har været interesse for at kombinere strigling og herbicidanvendelse for at reducere pesticidforbruget. Effekterne på ukrudtsbiomasse og plantetæthed er velundersøgt, mens der kun foreligger få eksperimentelle undersøgelser, der belyser de direkte og indirekte effekter af mekanisk ukrudtsbehandling på fugle og hvirvelløse dyr i dyrkede marker.
Det er projektets formål gennem felteksperimenter og modellering at kvantificere direkte og indirekte effekter af ukrudtsstrigling på sanglærkens reproduktion, på en række leddyr, på ukrudtsfloraen samt på landbrugets ressourceforbrug. Et vigtigt element i undersøgelsen er for faunaens vedkommende at modellere ukrudtsstriglingens evt. langsigtede effekt på bestandene. Ressourceforbruget i landbrugsproduktionen opgøres for at sammenholde naturmæssige påvirkninger med energiforbruget ved ukrudtsstrigling. På baggrund af undersøgelsens resultater udarbejdes anbefalinger for den mest hensigtsmæssige og skånsomme udførelse af ukrudtsstriglinger, samt indflydelsen heraf på driftsmæssige forhold.
Projektet, der blev udført på Kalø Gods (2003 – 2006), er finansieret gennem Miljøstyrelsens pesticidforskningsmidler. Projektet er udført af afdelingerne Vildtbiologi og Biodiversitet og Terrestrisk økologi (DMU), Inst. for Jordbrugsvidenskab (KVL) og Forskningscenter Foulum (DJF). Lektor Mogens Erlandsen (Inst. for Biostatistik, Århus Universitet) har været statistisk konsulent. Projektet er resultatet af en sammenlægning mellem to selvstændige ansøgninger til Miljøstyrelsen program for Bekæmpelsesmiddel forskning 2003-2005: 1)”Gør mekanisk ukrudtsbekæmpelse en forskel? Effekter på naturkvalitet, indikeret ved jordlevende dyr, og økologisk bæredygtighed, indikeret ved det fossile energiforbrug i dyrkningssystemer med og uden brug af herbicider” og 2) ”Effekten af markstrigling/radrensning på sanglærke, hvirvelløse dyr og vilde planter”.
Projektets resultater kan samles i følgende:
- Mellem 50 og 100 % af de lærkereder, der udsættes for ukrudtsstriglinger, forventes ødelagt.
- Timingen for den (de) enkelte striglinger i den enkelte mark eller afgrøder afgør, hvor mange reder der potentielt udsættes for strigling.
- Under normale dyrkningsforhold er der ikke stor risiko for, at striglinger påvirker lærkernes reproduktion i økologisk dyrket vårhvede eller en konventionel dyrket vårbyg eller vinterhvede, hvor striglinger erstatter den kemiske ukrudtsbekæmpelse.
- Mht. til sanglærken bør strigling i vårbyg derfor undgås efter ca. 10. maj (svarende til ca. 30 dage efter såning), og i vinterhvede bør der ikke strigles om foråret efter ca. 10. april.
- Ukrudtsstriglinger udført under normale dyrkningsforhold påvirker de udvalgte og modellerede arter af leddyr, og den samlede effekt af tre eller fire striglinger medfører alvorlige negative påvirkninger af bestandene.
- Hovedkonklusionerne for de modellerede arters vedkommende er, at de negative konsekvenser ved ukrudtsstrigling i kornafgrøder kan reduceres ved:
- højst at udføre to striglinger i vår-korn ikke senere end ca. 30 dage efter såning
- ikke at udføre striglinger i vinter-korn om efteråret, og kun at udføre én forårsstrigling inden ca. midten af april
- Regnorme, springhaler og mider vil sandsynligvis ikke blive påvirket af striglinger i vårhvede på lerjord. Det er mere usikkert om disse resultater også gælder på lettere og mere sandede jordtyper.
- Ukrudtsstriglinger kan udføres uden betydelige påvirkninger af energiforbrug og dækningsbidrag.
- Strigling vurderes ikke at være et realistisk alternativ til herbicider på konventionelle bedrifter, men alene et supplement.
- I forsøgene havde kun den første strigling effekt på ukrudt og afgrøde. De efterfølgende striglinger var virkningsløse.
Feltforsøgene 2004 & 2005
Der blev i 2004 foretaget i alt 59 yngleforsøg på de to forsøgsmarker fordelt på 32 territorier. I 2005 blev der foretaget 33 yngleforsøg fordelt på 20 territorier. Der kunne ikke påvises forskel på fordelingen af antallet af reder pr. territorium som følge af strigling. I ingen af årene blev der registreret redeforsøg ved tidspunktet for første strigling. Samlet betragtet har den direkte effekt af strigling ikke haft nogen indflydelse på den samlede reproduktion. Der blev ikke fundet nogen signifikante forskelle mellem år eller mellem striglede/ustriglede arealer mht. antallet af æg, redeunger og ’udfløjne’ unger pr. rede eller pr. territorium.
I ingen af forsøgsårene blev der fundet signifikante effekter af strigling på antallet af regnorme, regnormearter eller regnormevægt. Effekten af striglingerne på antal mider og springhaler varierede meget. Der var ikke tale om nogen entydig effekt af forskellige antal striglinger, og når datamaterialet blev testet for effekt af strigling mod ingen strigling, var der en statistisk signifikant, men numerisk meget lille, negativ effekt på antal springhalearter oktober 2004 samt en signifikant positiv effekt på antallet af springhaler i maj 2005.
I gennemsnit reducerede striglingerne ukrudtsbiomassen 33 %, men der var ingen sikker forskel mellem 1, 2 og 3 striglinger. Den procentuelle effekt af strigling var således uafhængig af ukrudtsfloraens sammensætning. Høstudbyttet var negativt påvirket af strigling, men heller ikke her var der nogen forskel på antallet af striglinger. Strigling reducerede i gennemsnit udbytterne 6 %. Ved visuel bedømmelse af afgrøden efter sidste strigling, var det ikke muligt at skelne de ubehandlede parceller fra de behandlede, hvilket vidner om at striglingerne har været skånsomme.
I markforsøgene var de naturlige variationer i ukrudtet så stor, at det ikke var muligt at registrere nogen generel effekt af striglingerne på ukrudtet. En væsentlig årsag til variationen i ukrudtsmængden og artssammensætningen på markniveau skyldes meget væsentlige forskelle i jordbundsforholdene.
For et kvægsædskifte vil det gennemsnitlige dieselforbrug stige med godt 10 liter/ha, svarende til en gennemsnitlig stigning på ca. 10 %.
Overgang til ren mekanisk ukrudtsbekæmpelse vil betyde relativt mere for dieselforbruget på svine- og planteavlsbrug end på kvægbrug (se nedenstående Figur A), som således vil kunne overgå til mekanisk ukrudtsbekæmpelse med den mindste energiomkostning. Sammenlignes tilsvarende forøgelsen af det samlede direkte og indirekte energiforbrug ved overgang til mekanisk ukrudtsbekæmpelse med det samlede energiforbrug på tre brugstyper, viser der sig den samme tendens: på kvægbrugene øges det samlede energiforbrug med ca. 1-2 %, på svinebrugene med 2-3 %, og på planteavlsbrugene med 6-7 %. Disse effekter er opgjort for en intensiv mekanisk ukrudtsbekæmpelse, som skønnes nødvendig for på lang sigt og i et ’worst case’ eksempel at styre ukrudtsproblemerne.
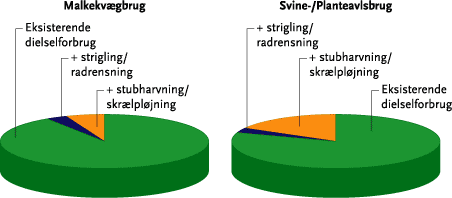
Figur A. Eksempel på beregnet ekstra dieselforbrug ved indførsel af intensiv mekanisk ukrudtsbekæmpelse (opdelt på strigling/radrensning og stubharvning/skrælpløjning) på malkekvægbrug og på svine-/planteavlsbrug, målt i forhold til dieselforbruget inden overgangen til mekanisk ukrudtsbekæmpelse. Figuren viser et ”worst case” eksempel på behov for mekanisk ukrudstbekæmpelse.
Modelsimuleringer på sanglærke, edderkop og løbebille
De udførte modelsimuleringer viste, at ingen af de udførte striglinger (hhv. 3 og 2 i vårbyg og vinterhvede) under standard dyrkningsbetingelser på type-kvægbedrifter havde nogen negative effekter på de målte variabler for sanglærkens vedkommende. Alle striglinger blev udført på tidspunkter der lå før lærkernes etablering af 1. yngleforsøg.
I monokulturer med vårbyg og vinterhvede (se afsnit 3.4) var der for vårbygs vedkommende ingen negative effekter af de fire udførte striglinger på den årlige totale bestand. Effekten af at ’udskyde’ 4. og sidste strigling med 10 dage til d. 20/5 var at reducere den totale årlige bestand alle år. Yderligere en udsættelse af 4. strigling til den 30/5 bevirkede, at dette mønster blev forstærket, da striglingen nu udføres på et tidspunkt, hvor både reder med æg og reder med unger bliver ødelagt af striglingen (se nedenstående Figur B).
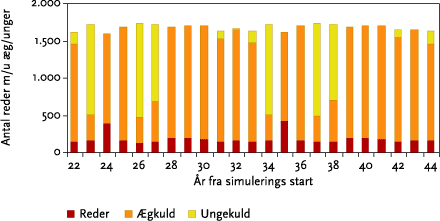
Figur B. Gennemsnitlige årlige tab af reder m/u æg og unger ved strigling i vårbyg foretaget d. 30. maj.
I et landskab, hvor der kun dyrkes vinterhvede, kan bestanden af sanglærker ikke overleve. Ved den senest udførte strigling d. 20/4 påvirkes det gennemsnitlige antal af flyvefærdige unger, og den gennemsnitlige totale bestandsstørrelse mindskes kraftigt.
For edderkoppernes vedkommende var den samlede effekt af striglingerne i gennemsnit en reduktion på 22 % af totalpopulationen i ’Basis-kvæg’ scenariet. For løbebillernes vedkommende var der en samlet reduktion på 19 %.
Den isolerede effekt på edderkoppen af 2., 3. og 4 strigling i vårbyg er af samme størrelsesorden på ca. 20 %. For løbebillens vedkommende er de isolerede effekter af strigling større end for edderkoppens vedkommende og størst efter 1. strigling både i vårbyg og i vinterhvede.
Summary
Mechanical weeding with a view to benefits for open-field fauna
This project investigated the impact of mechanical weeding on the wildlife contained in open fields together with the agricultural resource usage. Based on the results it is recommended that a maximum of two mechanical weeding operations are used in spring cereal crops, with the last not later than 30 days after sowing. Mechanical weeding can be used without significant impacts on energy budgets or gross margin, but at the current level of technology, and without changing rotations, it must be considered a supplement rather than a realistic alternative to herbicides on conventional farms.
The Bichel-committee considered the possibility for a reduction or cessation of pesticide use in Denmark and the associated benefits and problems. Subsequent research has demonstrated that mechanical weeding can be used as an alternative to herbicides in certain crops and in areas where environmental considerations require reduced pesticide use. Pesticide use has been an important factor in the decline of a range of European farmland bird species over the last 20 years, primarily via indirect effects on wild plants and arthropods. Mechanical weeding is rarely used in conventional farming, despite the interest to combine this with herbicide use to reduce pesticide usage. The effectiveness of mechanical weeding for weed control is well documented but there are few experimental investigations into its direct or indirect impact on birds and arthropods in fields.
The aim of the project was to quantify direct and indirect effects of mechanical weeding on skylark reproduction, a number of arthropod species, arable weeds, and agricultural resource usage through the use of field experiment and modelling approaches. An important element in the modelling was to consider the mechanical weeding’s long-term effect on wildlife. Agricultural resource usage is considered in order to compare overall wildlife impacts with energy usage for mechanical weeding. Experimental and modelling results were used to prepare recommendations for ‘wildlife friendly’ use of mechanical weeding together with its influence on agricultural management.
This project (2003-2006), was carried out on the Kalø Estate, by NERI departments of Wildlife Ecology and Biodiversity and Terrestrial Ecology, together with KVL Institute for Agricultural Sciences and DIAS, Foulum. Lektor Mogens Erlandsen (Institute for Biostatistics, Århus University) acted as a statistical consultant. The project is a result of the combination of two separate applications to the Danish EPA’s programme for pesticide research 2003-2005: 1) “Gør mekanisk ukrudtsbekæmpelse en forskel? Effekter på naturkvalitet, indikeret med jordlevende dyr, og økologisk bæredygtighed, indikeret ved det fossile energiforbrug i dyrkningssystemer med og uden brug af herbicider” and 2) ”Effekten af markstrigling/radrensning på sanglærke, hvirvelløse dyr og vilde planter”.
- Between 50 and 100 % of the lark nests exposed to mechanical weeding are expected to be destroyed;
- Timing of the individual weeding operations in the field determines the number of nests potentially exposed;
- Under normal agricultural practices where mechanical weeding replaces herbicides, the risk of weeding affecting skylark reproduction in organic spring wheat or a conventionally managed spring barley or winter wheat is low.
- W.r.t. skylark mechanical weeding in spring barley should not occur after approximately 10th May (or 30 days after sowing), and in winter wheat it should not be used after approximately 10th April.
- Mechanical weeding carried out under normal farming practice impacted the modelled arthropods with the combined effect of three or four consecutive operations resulting in serious negative consequences for the populations.
- Main conclusions from the results for the modelled species were that the negative consequences of mechanical weeding in cereal crops can be reduced by:
- Maximally using two mechanical weeding operations in spring cereal crops and not later than 30 days after sowing.
- Not to use weeding in winter cereal crops in autumn, and to only carry out one mechanical weeding operation in the spring before mid-April.
- Earthworms, springtails and mites will probably not be affected by mechanical weeding in spring barley on clay soils. It is uncertain whether these results will hold for lighter soils.
- Mechanical weeding can be used without significant negative impacts on energy budgets or gross margins.
- Mechanical weeding is not considered to be a realistic alternative to herbicides on conventional farms, but rather as a supplement.
- In the field experiment here, only the first weeding operation had an effect on the weeds and crop, the following applications were without effect.
Field Experiments 2004 & 2005
Fifty-nine breeding skylark attempts were made in 2004 on the two experimental fields, in 32 breeding territories. In 2005 there were 33 attempts in 20 territories (Table 5.1 and Figure 5.2 A).There were no significant differences between the number of ‘active’ nests (with eggs or nestlings) per territory as a result of mechanical weeding. There were no nesting attempts before the first mechanical weeding operation, and all in all weeding had no direct influence on the overall reproduction. In neither year was there a significant difference between mechanical weeded and un-treated areas in the number of eggs, nestlings, or fledged young per nest or per territory (Table 5.4 and Table 5.5).
None of the experiments showed significant effects of mechanical weeding on the number of earthworms, earthworm species, or earthworm weight (Figure 5.3). The effect on the numbers of springtails and mites was very varied (Table 5.6), and there was no clear effect of the different number of weeding operations (Figures 5.3-5.6). Tests for weeding vs no-weeding showed a statistically significant but very small negative impact on the number of springtails in October 2004, together with a positive effect on number of springtails in May 2005.
On average mechanical weeding reduced weed biomass by 33 %, but there was no clear difference between 1, 2, or 3 operations (Figure 5.7). The percentage reduction was independent of the weed community structure. Harvest was negatively affected by mechanical weeding, but again there was no difference between the numbers of operations used. Yield was reduced on average by 6 %. At a visual evaluation of the plots after the last weeding operation it was not possible to identify the treated from non-treated plots, which indicates that the weeding operation was relatively gentle.
In the field experiment, variation in weed biomass and species composition was so large that it was not possible to determine any general effect of mechanical weeding on weeds. A major cause of this variation was variation in soil type within the field.
For a cattle farm rotation the average diesel consumption will increase by at least 10 l/Ha, representing an average increase of 10 % (calculated on the basis of standard crop values from Figure 5.15).
Conversion to purely mechanical weed control will mean relatively more diesel consumption for pig and arable units compared to cattle farms (see Figure 5.16). Comparing the increase in combined direct and indirect energy consumption associated with conversion to mechanical weed control with the total energy consumption of the three farm types (Table 5.8) show the same tendency: cattle farms increase their overall energy consumption by 1-2%, whereas pig farmers increase by 2-3 % and arable farmers by 6-7 %. These effects are calculated for intensive mechanical weeding which is deemed necessary for a long-term and worst case example for weed control.
Model simulations of skylark, spider and beetle
The simulations showed that none of the standard mechanical weeding operations (i.e. 3 or 2 in spring barley and winter wheat) under the cattle farm simulation had any negative impact on the variables considered. In fact all operations were carried out before the period for the first skylark breeding attempt.
In monoculture spring barley (see section 3.4) there was no negative impact of the four weeding operations on the total population of skylarks (Figure 5.17 B). The effect of delaying the 4th operation by 10 days until the 20/5 was a reduction of the overall population in all years (Figure 5.17 B). Further delaying of the operation until the 30/5 resulted in a greater reduction since the operation was then carried out when nests with both eggs and young could be destroyed (Figure 5.19).
In a landscape where only winter wheat is grown, the model skylark population did not survive (Figure 5.20). However, the latest mechanical weeding operation (20/4) has a powerful effect of reducing the mean number of fledglings produced and the overall population size (Table 5.12).
The impact of mechanical weeding on the spider was a 22 % reduction of the total population of the baseline cattle scenario (Figure 5.18 A). For the beetle, the impact was a 19 % reduction (Figure 5.18 B).
The isolated effect on spiders of the 2nd to 4th operation in spring barley is of the same magnitude (ca. 20 %) (Figure 5.21B; Table 5.13). The impact on the beetle was greater then the spider and highest with the first operation both in spring barely and winter wheat (Figure 5.22 A and C; Table 5.14).
1 Formål
Forsøgsarbejde har vist, at ukrudtsstrigling kan indgå som alternativ til herbicider i særlige afgrøder og i særlige områder, hvor pesticidanvendelse måtte ønskes begrænset af hensyn til miljø og natur. Pesticidanvendelse har været en af de væsentlige årsager til den tilbagegang som har været observeret for flere agerlandfugle i Vesteuropa inden for de seneste to årtier. Pesticidernes negative indvirkning har primært været indirekte gennem en reduktion af vilde planter og leddyr. Ukrudtsstriglinger anvendes stort set ikke i konventionel dyrkning, selvom der har været interesse for at kombinere strigling og herbicidanvendelse for at reducere pesticidforbruget. Effekterne på ukrudtsbiomasse og plantetæthed er velundersøgt, mens der kun foreligger få eksperimentelle undersøgelser, der belyser de direkte og indirekte effekter af mekanisk ukrudtsbehandling på fugle og hvirvelløse dyr i dyrkede marker.
Med baggrund i den nuværende handlingsplan for nedbringelse af pesticidforbruget (Pesticidplan 2004-2009) har det været dette projekts primære formål at belyse de faunamæssige, dyrkningsmæssige og økonomiske konsekvenser, der er knyttet til mekanisk ukrudtsbekæmpelse. Det er projektets formål gennem felteksperimenter og modellering at kvantificere direkte og indirekte effekter af ukrudtsstrigling på sanglærkens reproduktion, på en række hvirvelløse dyr, på ukrudtsfloraen samt på landbrugets ressourceforbrug. Et vigtigt element i undersøgelsen er for faunaens vedkommende (sanglærke, løbebiller og edderkopper) at modellere ukrudtsstriglingens langsigtede effekt på arternes populationsudvikling. I dyrkningsscenarier opgøres ressourceforbruget i landbrugsproduktionen for derved at kunne sammenholde eventuelle naturmæssige påvirkninger med det konkrete energiforbrug ved ukrudtsstrigling. Effekter af mekanisk ukrudtsbehandling sammenholdes med effekter af kemisk ukrudtsbekæmpelse (herbicider). På baggrund af undersøgelsens resultater udarbejdes der anbefalinger for den mest hensigtsmæssige og skånsomme udførelse af ukrudtsstriglinger, samt indflydelsen heraf på driftsmæssige forhold.
2 Baggrund
- 2.1 Ukrudtsstrigling som alternativ til herbicider
- 2.2 Hvorfor undersøge effekter på fugle, hvirvelløse dyr, den vilde flora, og landbrugets ressourceforbrug?
2.1 Ukrudtsstrigling som alternativ til herbicider
Bichel-udvalgets arbejde behandlede mulighederne for en hel eller delvis afvikling af pesticidanvendelsen i Danmark og de ulemper og fordele der vil være forbundet hermed (Bichel-udvalget 1998). Siden da er der foregået en omfattede forskning og et sideløbende forsøgs- og udviklingsarbejde. Generelt er resultatet heraf, at der er mulighed for at ukrudtsstrigling kan indgå som alternativ til herbicider i særlige afgrøder (fx markært), og i særlige områder hvor pesticidanvendelse måtte ønskes begrænset af hensyn til miljø og natur. Ligeledes er effektiv ukrudtsstrigling en vigtig faktor for rentabel økologisk planteavl.
Igennem de seneste to årtier har der været udført en betydelig forskningsmæssig indsats med fokus på pesticidernes direkte og indirekte effekter på agerlandets vilde flora og fauna (Braae et al. 1988, Hald & Reddersen 1990). Det er veldokumenteret, at pesticidanvendelse har været en af de væsentligste grunde til flere agerlandfugles markante tilbagegang i de fleste vesteuropæiske lande (Fuller et al. 1995, Chamberlain et al. 2000). Pesticidernes negative indvirkning har primært været indirekte gennem en reduktion af vilde planter, insekter og andre hvirvelløse dyr (Potts 1986, Odderskær et al. 1997, Morris et al. 2005).
Blandt de mest udbredte metoder for ukrudtsbekæmpelse i økologisk dyrkede kornafgrøder er anvendelsen af ukrudtsstriglinger. Denne alternative form for ukrudtsbekæmpelse anvendes stort set ikke i konventionel dyrkning, selvom der har været en vis forsøgsmæssig interesse for at kombinere strigling og herbicidanvendelse i bestræbelse på at reducere pesticidforbruget. Effekterne på ukrudtsbiomasse og plantetæthed er variable og i en del tilfælde utilfredsstillende, og afhænger af blandt andet af ukrudtsflora, behandlingstidspunkter og vejrforhold (Rasmussen 1996, Rasmussen & Ascard 1995). Der foreligger kun få eksperimentelle undersøgelser, der belyser de direkte og indirekte effekter af mekanisk ukrudtsbehandling på fugle og hvirvelløse dyr i dyrkede marker (Thorbek & Bilde 2004; Esbjerg et al. 2006).
2.2 Hvorfor undersøge effekter på fugle, hvirvelløse dyr, den vilde flora, og landbrugets ressourceforbrug?
Effekten af mekanisk ukrudtbekæmpelse målt i forhold til anvendelse af herbicider kan måles på mange forskellige indikatorer. I dette projekt undersøges effekten af ukrudtsstrigling på forekomsten af vilde planter og udvalgte dyrearter/grupper, der muliggør undersøgelser af effekter på mange trofiske niveauer og funktionelle grupper af vilde dyr og planter. Undersøgelserne omfatter sanglærke, en udvalgt art af løbebiller (Bembidion lampros), en udvalgt art af edderkopper (tæppespinderen Erigone atra), mikroleddyr (springhaler og mider), regnorme og ukrudtsflora.
Effekter på sanglærkens reproduktionssucces er vigtige at opgøre fordi ukrudtsstrigling foregår i den første halvdel af fuglenes yngleperiode. Sanglærken er den fugleart i agerlandet, der er mest eksponeret for denne form for ukrudtsbekæmpelse, da reden etableres på jorden og den overvejende del af bestanden yngler i kornmarker.
Biller og edderkopper er vigtige prædatorer på forskellige mikroleddyr (herunder springhaler og mider) samt bladlus og andre skadedyr, og tjener derudover som vigtige byttedyr for andre dyr på højere trofiske niveauer (herunder sanglærken og andre agerlandsfugle), og bidrager dermed til at oprette en høj biodiversitet i landbrugssystemet. De udvalgte arter er hyppigt forekommende arter i dyrkede kornmarker, og er begge arter tilpasset et omskifteligt miljø.
Jordens mikroleddyr og regnorme har vigtige funktioner som fremmere af den biologiske omsætning af plantenæringsstoffer i jorden. De forøger jordens egnethed som dyrkningsmedium for afgrøder. De tjener som vigtige byttedyr for andre dyr på højere trofiske niveauer.
Den naturlige flora (ukrudt) undersøges fordi den er en vigtig dyrkningsmæssig faktor samt forventes at spille en rolle for de indirekte effekter knyttet til ukrudtsstriglinger. Høstudbyttet registreres i striglede og ustriglede markhalvdele samt i de udlagte blokforsøg med henblik på at kunne fastlægge den optimale striglingsintensitet under de givne forsøgsbetingelser.
I de følgende afsnit uddybes begrundelsen for undersøgelsen.
2.2.1 Effekter på højere dyr
Kun få marktilknyttede arter af pattedyr og fugle er i perioder af deres liv potentielt udsatte for en risiko gennem mekanisk ukrudtsbehandling af afgrøder (sanglærke, vibe, bomlærke, hare, agerhøne, fasan). Blandt disse, er sanglærken den potentielt mest udsatte, idet hovedparten af alle reder placeres i markerne, direkte på jorden. Der foreligger kun sparsomme oplysninger om effekter af mekanisk behandling på højere dyr. For sanglærken foreligger der igennem pilotundersøgelser oplysninger, der - selvom det foreliggende datamateriale er spinkelt - tyder på, at reder med æg der udsættes for strigling påføres en dødelighed på mellem 40-60 % (Odderskær, 2002). Derudover undersøges striglingens effekter på sanglærke og leddyr knyttet til jordoverfladen i et igangværende projekt (’Flora og fauna i marker: Betydningen af mekanisk ukrudtsbekæmpelse – og værdisætning af skånsom plantebeskyttelse’) under Miljøstyrelsens pesticidforskningsmidler. De negative følgevirkninger for sanglærken og andre jordrugende arter afhænger af tidspunktet for behandling(er), samt frekvens af og interval mellem disse, og størrelsen af evt. følgevirkninger vil således afhænge af de enkelte arters adfærd og biologi. For sanglærkens vedkommende er der behov for på et mere solidt grundlag at kvantificere den direkte effekt af mekanisk behandling på denne karakterarts samlede reproduktion.
2.2.2 Effekter på hvirvelløse dyr
Der foreligger kun få eksperimentelle undersøgelser, der belyser de direkte og indirekte effekter af mekanisk ukrudtsbehandling på hvirvelløse dyr i dyrkede marker. Nye danske undersøgelser har vist, at de direkte effekter af strigling i kornafgrøder på nogle af de hyppigst forekommende insektgrupper (løbebiller og rovbiller) og edderkopper (tæppespindere) i dyrkede marker kan svinge fra 25 – 60 % dødelighed for hhv. billernes og edderkoppernes vedkommende (Thorbek & Bilde, 2004). Effekten på den samlede biomasse af marklevende hvirvelløse dyr kendes øjensynligt ikke. Tæppespindere (edderkopper af familien Linyphiidae) og løbebiller er valgt som studiearter, fordi de både er mest talrige og mest effektive nyttedyr blandt leddyr i dyrkede marker. De er af økonomisk betydning, fordi store populationer af disse generalist-prædatorer kan forhindre udbrud af skadedyr. Derudover er disse dyr vigtige som føderessource for andre arter herunder fugle som sanglærken.
Jordens mikroleddyr (primært springhaler og mider) og regnorme er velegnede som indikatorer for jordøkosystemets struktur og funktion. Mikroleddyr optræder ofte i høje tætheder i landbrugssystemer (Petersen & Luxton 1982; Axelsen & Kristensen 2000) og er lette at kvantificere præcist. Artsdiversiteten af mikroleddyr er stor i landbrugssystemer, hvilket giver mulighed for at kvantificere forskelle i biodiversitet mellem forskellige dyrkningssystemer (Krogh 1991; Krogh et al. 1996). Mikroleddyr har en effekt på funktionen af jordøkosystemet idet de lever af bakterier og svampehyfer i jorden. Denne græsning er medvirkende til at holde disse primære nedbryderorganismer i en eksponentiel vækstfase og dermed maksimere mineraliseringen af dødt organisk stof og frigivelsen af plantenæringsstoffer. Mikroleddyr er sandsynligvis også medvirkende til spredning af svampesporer, således at tilført dødt plantemateriale hurtigere bliver koloniseret af svampe (Petersen 2002).
Mikroleddyr er vigtige fødeemner for generalist insektprædatorer i landbrugsøkosystemer (Toft & Wise 1999; Marcussen et al. 1999; Bilde et al. 2000). I den tidlige vækstsæson udgør springhaler hovedparten af føden for mange insektprædatorer, f.eks. edderkopper og løbebiller. Dette betyder, at populationen af insektprædatorer er høj på det tidspunkt hvor angreb af skadelige insekter som bladlus bliver af betydning senere i vækstsæsonen. En høj forekomst af springhaler er derfor med til at fremme bestanden af skadevolderes naturlige fjender, som potentielt kan være til gavn for landbrugsproduktionen. Det er derfor vigtigt at undersøge effekten af den mekaniske ukrudtsbekæmpelse på mikroleddyrene.
Regnormenes effekter på jordstruktur, udluftning og dræning er velkendt fra mange studier. Artsdiversiteten af regnorme er ikke stor i landbrugssystemer, men deres biomasse og graveaktivitet er så stor, at de har stor betydning for plantevækst (Edwards & Bohlen 1996). Desuden er regnorme en vigtig fødekilde for mange højerestående dyr. Anvendelsen af mikroleddyr og regnorme som indikatorer for naturindhold er derfor relateret til både struktur og funktion af jordøkosystemet.
Herbiciders effekt på jordlevende dyr er forholdsvis godt undersøgt. Almindelig brug af herbicider har sandsynligvis ingen eller kun ringe effekt på regnorme (Edwards & Bohlen 1992). Mange i Danmark brugte herbicider har ligeledes ringe effekt ved markdosis, hvis nogen, på jordlevende leddyr (Scott-Fordsmand et al. 2000; Cortet & Poinsot-Balaguer 2000).
Effekten på jordlevende dyr af ukrudtsstrigling, som den anvendes i moderne landbrug, er ikke undersøgt. I og med at mekanisk bekæmpelse i mange tilfælde medfører en mere eller mindre kraftig forstyrrelse af jorden (f.eks. radrensning eller strigling), er det dog sandsynligt, at der vil være en vis negativ effekt på regnorme og mikroleddyr. Dette skyldes dels at dyrene i overfladen kan blive dræbt af jordbehandlingen, men jordløsningen kan også betyde en øget udtørring af jorden. Jordens fugtighedsforhold er en særdeles vigtig faktor for bestanden af jordlevende dyr (Rapoport & Tschapek 1967; Edwards & Bohlen 1996). Det er derfor sandsynligt, at mekanisk bekæmpelse vil have nogen effekt, måske endda en kraftig effekt på jordens organismer. Der mangler i høj grad videnskabelige undersøgelser til at belyse effekterne af ukrudtsstrigling på jordbundsfaunaens artsdiversitet og biomasse.
2.2.3 Effekter på ukrudt
Det er ikke projektets formål at dokumentere ukrudtsstriglingens potentiale som alternativ bekæmpelsesforanstaltning, da dette spørgsmål er belyst i talrige forsøg gennem de sidste 10-15 år (Rasmussen et al., 1997). I forsøg hvor ukrudtsstrigling indgår, er det imidlertid vigtigt at registrere ukrudtsstriglingens effekter på ukrudt og udbytte, for at få en meningsfyldt beskrivelse af striglingens aggressivitet. I modsætning til herbicidbehandlinger, hvor angivelse af herbicidnavn og dosering giver en meningsgivende beskrivelse af behandlingen, giver det kun ringe mening at angive striglingens aggressivitet gennem en beskrivelse af den anvendte strigle og dens aktuelle anvendelse. At der eksempelvis har været kørt 8 km/t med et givet striglefabrikat, siger intet om striglingens aggressivitet (Rasmussen, 1992). Det er først når effekterne på ukrudt og afgrøde er kendte, at det er muligt at vurdere om striglingens aggressivitet i forhold til almindelig landbrugsmæssig praksis (Rasmussen, 1992). For at kunne vurdere om aggressiviteten af de i forsøgene gennemførte striglinger er stor, middel eller lille i forhold til almindelig landbrugsmæssig praksis, er det således nødvendigt at gennemføre responsmålinger på ukrudt og afgrøde.
2.2.4 Effekter på landbrugets ressourceforbrug
Den direkte og indirekte effekt af ukrudtsstrigling på landbrugets ressourceforbrug og økologiske bæredygtighed kan med fordel opgøres ved en systemanalyse af energiforbrug og – produktion. Energibalancen integrerer en lang række af de ressourceforbrug i form af pesticider, næringsstoffer, maskiner etc., der påvirker den økologiske bæredygtighed af landbrugsproduktionssystemer (Fluck 1992; Dalgaard 2001). En anden fordel er, at for at opgøre energiforbruget og produktionen må hele produktionssystemet beskrives, og denne systembeskrivelse kan anvendes til beregning af produktions- og miljøøkonomiske forhold og sammenligning af de forskellige produktionssystemer med og uden anvendelse af herbicider. Desuden har energiforbruget relation til udledningen af drivhusgassen kuldioxid.
I nærværende projekt opgøres energiforbruget som summen af direkte energi forbrugt som olie, el, kul etc. og indirekte energi til fremstilling af gødning, pesticider, maskiner etc. I Danmark er der udviklet modeller til opgørelse af såvel det direkte som det indirekte energiforbrug i landbruget (Dalgaard et al. 2001), såvel som den afledte effekt på udledning af drivhusgasser (Olesen 2005, Dalgaard et al. 2002). Generelt gælder der, at ukrudtsstrigling påvirker energiforbruget, idet specielt dieselforbruget stiger (Dalgaard et al., 2003 a). Men sammenhængen mellem plantebeskyttelsesstrategi og den samlede energibalance samt drivhusgasudledning i landbruget er ikke simpel, idet såvel energiinput som -output påvirkes, og idet energiindholdet i indsatsfaktorer såsom gødning, pesticider og maskiner, der påvirkes af dyrkningssystemet, også bør medregnes (Dalgaard 1998).
2.2.5 Modellering på landskabsniveau
Det har tidligere i praksis været umuligt at kvantificere effekterne af forskellige dyrkningsmæssige tiltag på dyrepopulationer set på landskabsskala, dels på grund af problemer både med at definere landskaber med tilstrækkelig kompleksitet og inkorporering af rumlige og tidsmæssige interaktioner. Den seneste udvikling indenfor terrestrisk populationsmodellering har imidlertid givet mulighed for at løse disse problemer gennem konceptet landskabsøkotoxikologi og landskabsskala modelsimulering (Topping et al., 2003; Topping & Odderskær, 2004). I denne type af modellering er dyrenes individuelle adfærd og økologi opskaleret til et landskabsniveau. Integrationen af dyreøkologi og påvirkninger fra f.eks. pesticidanvendelse/ukrudtsstriglinger sker via en integration med en detaljeret dynamisk landskabssimulering af dyrkningspraksis og den konkrete anvendelse af pesticider/strigling. DMU har opbygget et modelsimuleringredskab, ALMaSS, som er i stand til i praksis at håndtere ovenstående problemstilling. ALMaSS modelsystemet består af individbaserede modeller for en række dyrearter og en landskabsmodel (en række konkrete landskaber er digitaliseret). Den enkelte bedriftsstyring foregår igennem en ”farm manager”, og derudover tager modelsystemet imod inputdata i form af f.eks. vejrlig, pesticidbrug, ukrudtsstrigling, behandlingshyppigheder, arealstørrelser for de enkelte behandlinger mm. (se endvidere Bilag A for en nærmere beskrivelse). Fordelen ved at anvende ALMaSS i nærværende projekt kan opsummeres som følger:
- Modelsystemet er allerede udviklet og er peer reviewed,
- Det er i stand til at integrere effekter af pesticidbrug og ukrudtsstrigling,
- Rumlig og tidsmæssig heterogenitet er indbygget,
- Der er udviklet dyremodeller for løbebille (typeart) (Bilde & Topping, 2004), edderkop (Thorbek & Topping 2003) (typearter af tæppespindere) og sanglærke (Topping & Odderskær, 2004),
- Der eksisterer en landskabsmodel som indeholder forsøgsarealer, der tidligere har været anvendt ved undersøgelser over arealudnyttelsens og driftspraksis’s betydning for sanglærken (Jepsen et al. 2005),
- Der er udviklet beskrivelser for driftspraksis for forskellige brugstyper,
- Der er udviklet dyrkningsplaner og vækstmodeller for de mest almindeligt forekommende afgrøder i Danmark (se eksempel i Topping & Odderskær 2004 samt Bilag B).
Anvendelsen af modelsystemet ALMaSS i nærværende projekt ses som det bedste middel for under samme landskabsmæssige betingelser at foretage sammenlignende undersøgelser af indvirkningen af mekanisk og kemisk ukrudtsbekæmpelse på udvalgte indikator arter af dyr.
3 Materiale og metoder
- 3.1 Oversigt over projektets delelementer
- 3.2 Pilotprojekt
- 3.3 De eksperimentelle undersøgelser
- 3.4 Modelleringsdel - sanglærke, løbebille, tæppespinder
- 3.5 Systemanalyse af landbrugets ressourceforbrug og driftsøkonomi
3.1 Oversigt over projektets delelementer
Projektet består af fire dele:
1) en pilotdel, 2) en eksperimentel del, 3) en modelleringsdel samt 4) en systemanalysedel
ad 1) Det undersøges under feltforhold om evt. effekter af ukrudtsstriglinger på en objektiv måde kan erkendes på udlagte kunstigt fremstillede reder med æg.
ad 2) Ukrudtsstriglingers effekter på a) lærkereder (naturlige, + kunstigt anlagte), b) jordlevende dyr (springhaler, mider, regnorme), c) ukrudtsbiomasse, d) plantebestand og e) høstudbytte undersøges eksperimentelt på to vårhvedemarker i to år på Kalø Gods.
ad 3) I modelleringsdelen simuleres effekterne af mekanisk ukrudtsbehandling og traditionel konventionel dyrkning med anvendelse af pesticider for sanglærke, løbebille (Bembidion lampros) og edderkop (tæppespindere, Erigone sp.) i et konkret (digitaliseret) landskab. Ud over data fra eksisterende litteratur anvendes der for sanglærkens vedkommende data for striglingseffekter fra den eksperimentelle undersøgelse.
ad 4) I systemanalysedelen opgøres det direkte og indirekte energiforbrug på typiske landbrugsbedrifter og det ekstra dieselforbrug forbundet med mekanisk ukrudtsbekæmpelse estimeres. Desuden opgøres maskin- og arbejdsomkostningerne samt det direkte energiforbrug i det dyrkningssystem, som markforsøgene på Kalø indgår i (forskellige vårhvede dyrkningsstrategier med og uden strigling), og dækningsbidraget i de to systemer sammenlignes.
Sammenhængen mellem de fire delelementer er vist i Figur 3.1, som viser hvorledes resultater fra den eksperimentelle del (2) indgår i modelleringsdelen (3) og systemanalysedelen (4). I modelleringsdelen beregnes en række scenarier (1-9) for effekter af ukrudtsstriglingen på sanglærke, løbebille og edderkop. I systemanalysedelen konsekvensvurderes ukrudtsstriglingerne i markforsøget mht. dækningsbidrag og energibalance (Scenario E), og de energimæssige konsekvenser af at indføre ukrudtsstrigling og anden mekanisk bekæmpelse beregnes for fire forskellige brugstyper (Scenario A-D).

Figur 3.1. Sammenhængen mellem projektets delelementer. For nærmere beskrivelse af scenarier se afsnit 3.4.
3.2 Pilotprojekt
3.2.1 Forsøgsstrigling med kunstige reder
Den 27. oktober 2003 udførtes to forsøgsstriglinger på en vinterbygmark på Kalø Gods. Striglingerne blev udført med det formål at undersøge anvendeligheden af kunstige reder i et forsøg på ad denne vej i tillæg til naturlige reder at kunne øge antallet af reder i de efterfølgende eksperimentelle forsøg. Afgrødehøjden var på striglingstidspunktet 6-7 cm med et (anslået) udviklingstrin på 32-37 (decimalskala). Der var forud for striglingerne udlagt og registreret data for 38 kunstige reder på forsøgsarealet. Rederne blev konstrueret ud fra data fra eksisterende litteratur samt fra erfaringer indhentet fra tidligere undersøgelser (Odderskær et al. 1997). Rederne blev lavet af vissent græs, som før udlægning i marken blev formet som en rede. Det tilstræbtes, at de færdige reder efter anbringelse i marken havde en øverste diameter på mellem 7-8 cm, og at den indvendige største dybde var mellem 6 og 7 cm. Dette svarer til, hvad der foreligger af oplysninger fra en tysk undersøgelse (Pätzhold, 1975) samt data indhentet fra den foreliggende undersøgelse (upublicerede data). Umiddelbart før 1. strigling blev der i de enkelte reder placeret hhv. 3 (32 reder) eller 4 (8 reder) vagtelæg med en gns. vægt på 12.8 g. Denne vægt er ca. fire gange større end den gns. vægt på lærkeæg. Der blev kun lagt æg ud i forbindelse med 1. strigling.
Striglingerne blev udført samme dag, som rederne var lagt ud. I tidsrummet mellem 1. og 2. strigling blev der ikke observeret mulige prædatorer (primært kragefugle). Efter hver strigling blev de enkelte reders skæbne (intakte, skadede eller ødelagte reder samt antallet af intakte, ødelagte eller ’fjernede’ æg) registreret.
Ved beregningen af striglingens påvirkning af reder/æg blev alle reder, der tydeligt var påført mekanisk skade af redematerialet og som samtidigt indeholdt et eller flere ødelagte æg henregnet til gruppen af ”ødelagte/tabte” reder. Hvis sådanne reder ikke var påført skader på æg, blev rederne betragtet som ”overlevende”, dvs. det blev antaget, at et en lærkehun ville have fortsat rugningen under naturlige forhold. Den resterende del af rederne var intakte (uden synlige påvirkninger fra striglingerne).
Ukrudtsstriglingen blev foretaget med en 12 m strigle af mærket CMN ved en hastighed på 10 km i timen og et striglingstryk på 40 bar, hvilket bevirkede en effektiv striglingsdybde på ca. 2-3 cm.
3.3 De eksperimentelle undersøgelser
3.3.1 Forsøgsarealer og forsøgsdesign
De eksperimentelle undersøgelser udførtes på to marker på Kalø Gods. Markerne, der har et samlet areal på ca. 60 ha, blev i 2004 og 2005 tilsået med økologisk dyrket vårhvede. Begge marker blev opdelt hver i to arealmæssigt lige store halvdele, hvoraf den ene halvdel blev striglet medens den anden halvdel forblev ustriglet (Figur 3.2). Første strigling udførtes umiddelbart efter kornets fremspiring. På begge marker blev der i 2005 skiftet mellem kontrol og ”behandlet” areal i forhold til 2004. I 2004 blev der udført tre striglinger hhv. d. 3/5, 14/5 og 21/5. I 2005 blev striglingerne udført hhv. d. 2/5, 12/5 20/5. Første strigling blev udført kort efter kornets fremspiring (ca. 5cm). Det var ikke muligt at fastlægge ’tændernes’ indstilling (vinkel) på striglingsudstyret, da denne var variabel (selvjusterende). Striglingerne blev udført med samme udstyr som beskrevet under pilotforsøget. Striglingstrykket var på 60-70 bar (variabelt pr. automatik) og kørehastigheden var mellem 12-15 km/t.
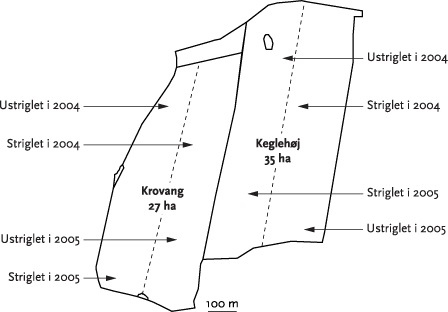
Figur 3.2. Skitse over de to forsøgsmarker med angivelse af de markafsnit der blev striglet i hhv. 2004 og 2005.
Effekter på ukrudtsbiomasse og jordlevende dyr blev undersøgt ved hjælp af et fuldstændig randomiseret blokdesign med 4 behandlinger (kontrol, 1 årlig strigling, 2 årlige striglinger og 3 årlige striglinger) og 4 gentagelser som blev lagt i de striglede markafsnit. Parcelstørrelsen var 12x12 m (netto). Hvert år blev der således etableret 2 forsøg med hver 16 forsøgsparceller (Figur 3.3). Prøvetagninger af ukrudtsbiomasse blev foretages i samtlige delparceller (2x16 hvert år), mens de jordlevende dyr kun blev indsamlet i det ene blokforsøg (Keglehøj).
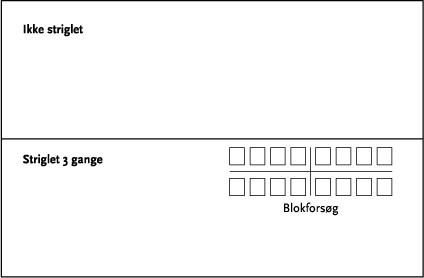
Figur 3.3. Skitse over forsøgsareal (én mark).
3.3.2 Sanglærke
Markforsøget (2 x 2 storparceller (= de striglede/ustriglede dele af hver mark):
I 2004 og 2005 blev der på Krovang og Keglehøj markerne udført mellem 20 og 30 moniteringer af territorier i perioden medio april til primo august. De hyppige moniteringer blev udført for at sikre et nøjagtigt kendskab til og placering af antallet af territorielle hanner, samt hvilke af disse der var udparrede. Derudover var disse registreringer med til at fastlægge, hvornår og i hvilke territorier de enkelte hunner havde påbegyndt redebygning/æglægning og rugning.
Hverken i 2004 eller i 2005 blev der konstateret redeforsøg på tidspunktet for 1. strigling, og der blev følgelig ikke udlagt kunstige reder i forbindelse med denne strigling. Ved både 2. og 3. strigling blev der på hver mark (2004) udlagt i alt 24 kunstige reder. Disse blev i 2004 udlagt (8 i hver) i tre forskellige typer af jordstruktur (visuelt bedømt) I) knoldet/leret, II) løs/muldrig og III) sandet. I 2005 blev der på Keglehøj udlagt 16 kunstige reder (kun 2 jordtyper kunne erkendes) og på Krovang blev der udlagt 32 reder (16 reder udlagt i type III). Umiddelbart før hver strigling blev der udlagt dværgvagtelæg i rederne med følgende fordeling af ægkuldstørrelser: 2 æg (1), 3 æg (6), 4 æg (15) og 5 æg (2). Denne fordeling afspejler data fra tidligere undersøgelser af sanglærken (Odderskær et al. 1997). Rederne blev udlagt på hver side af et kørespor med en indbyrdes afstand i køreretningen på 6 m. Derudover var rederne anlagt på hver side af køresporet med en afstand fra køresporet på hhv. 1.5, 2, 4 og 5 meter. Før og efter strigling blev der som dokumentation taget et foto af de enkelte reder, og redernes skæbne blev noteret som beskrevet under afsnit 3.2.1.
Umiddelbart før og efter hver strigling blev de igangværende naturlige reder undersøgt, og redeskæbnen blev noteret (levende/døde æg/unger, dødsårsag). Yngleresultater fra samtlige naturlige redeforsøg registreredes igennem hele yngleperioden for en vurdering af forskelle i det samlede antal af udfløjne unger i kontrol og behandlede arealer. Følgende variabler blev registreret, i den udstrækning det var muligt: start og sluttidspunkt for redebygning, æglægning, rugeperiode og ungeperiode samt antal æg, redeunger og udfløjne unger, ungevægte fra unger i alderen fra 5 dg. til udflyvning, dødelighed af æg/unger, dødsårsag (herunder strigling), territorie-ID samt antal af og kronologi for de enkelte ynglepars yngleforsøg.
3.3.3 Ukrudt- og afgrøderegistreringer
Blokforsøget (2 x 16 parceller):
For at bestemme striglingernes effekt på ukrudtet, blev ukrudtet i blokforsøgene høstet i juni måned i begge år. I hver af de 32 parceller blev der i begge år høstet 4 tilfældige 0,25 m² prøveflader. Ukrudtet blev opdelt i de 2-3 dominerende arter (de arter, som bidrager med mindst 20 % af den samlede ukrudtspopulations biomasse) og en rest bestående af de resterende arter. Efter tørring og vejning blev tørstoffet omregnet til ukrudtsbiomasse pr m². Kun enårige ukrudtsarter blev høstet. De flerårige arter blev kun visuelt bedømt, da de ikke påvirkes af strigling. I markforsøgene blev der foretaget en kvantitativ beskrivelse af ukrudtsforekomsten, og markerne blev systematisk fotograferet.
For at få et mål for striglingernes aggressivitet, blev der kort tid efter sidste ukrudtsstrigling foretaget en visuel bedømmelse af afgrøden (Rasmussen 1996).
På Keglehøj var der en stor men uensartet bestand af Agertidsel, som ikke blev registreret systematisk, da Agertidsel ikke påvirkes af ukrudtsstrigling. På Krovang var der spredte forekomster af alm. kvik, som heller ikke blev registreret systematisk, da denne art ikke påvirkes af ukrudtstrigling.
Markforsøget (2 x 2 storparceller):
I alle markhalvdele blev der foretaget visuelle vurderinger af ukrudt og afgrøde på samme tidspunkt som i blokforsøgene. Vurderingerne blev dokumenteret på grundlag af fotoserier, da dette gav den mest repræsentative dokumentation i forhold til de ressourcer der var afsat til registreringerne. Da blokforsøgene viste, at blokvariationen var betydeligt større end behandlingsvariationerne, og vurderingerne af markhalvdelene viste, at der var meget betydende variabilitet indenfor storparcellerne, blev det alene søgt vurderet, om det var muligt at fastslå en generel effekt af striglingerne uden en egentlig kvantificering.
3.3.4 Høstudbytte
Markforsøget (2 x 2 storparceller), samt blokforsøget (2 x 16 parceller):
Høstudbyttet blev registreret i blokforsøg og markforsøg. I blokforsøgene måltes høstudbyttet i de enkelte delparceller på basis af afhøstninger (2x12m) med forsøgsmejetærsker. Datoer for tilsåning, høst af marker og forsøgsparceller er angivet i Tabel 3.1.
Tabel 3.1. Dyrkningskronologi for de to forsøgsmarker (Ke = Keglehøj, Kr = Krovang) i 2004 og 2005. Angivelser for spiringsdatoer er estimerede. 1. – 3. Str = 1., 2. og 3. strigling.
| Mark | Såning | Spiring | 1. Str | 2. Str | 3. Str | Høst af mark | Høst af forsøg | |
| 2004 | Ke | 14+15/4 | 27/4 | 3/5 | 14/5 | 21/5 | 5+6/9 | 8/9 |
| Kr | 15+16/4 | 30/4 | 3/5 | 14/5 | 21/5 | 3+4/9 | 8/9 | |
| 2005 | Ke | 12+13/4 | 26/4 | 2/5 | 12/5 | 20/5 | 31/8 | 5/9 |
| Kr | 16+17/4 | 31/4 | 4/5 | 12/5 | 20/5 | 29+30/8 | 5/9 |
Der var ikke muligt i mejetærskeren inden høst at installere GPS-baseret udstyr til kontinuert høstudbyttemåling. Prøver fra forsøgsafhøstning (halm og kerneudbytte) blev analyseret på laboratorium i Foulum. For kerneprøverne blev opgjort værdier for hektolitervægt og indhold af protein, stivelse og gluten14. Desuden blev tørstofprocenten analyseret for både halm- og kerneprøverne. Grundet tekniske mangler ved den forsøgsmejetærsker, det oprindeligt var påtænkt at benytte, blev det i stedet valgt at benytte Foulums forsøgsmejetærsker for at kunne veje halm, hvilket er nødvendigt for at kunne opgøre energibalancen.
3.3.5 Mikroleddyr og regnorme
I maj 2004 og maj 2005 blev der indsamlet regnorme og leddyr (2004: regnorme d. 1. - 2. juni, mikroleddyr d. 27. og 28. maj; 2005: regnorme d. 26. – 27. maj, mikroleddyr d. 26. maj og d. 3. oktober). Der blev som planlagt indsamlet to regnormeprøver pr. prøvefelt i blokforsøget i den ene forsøgsmark, Keglehøj i 2004 og Krovang i 2005. Hver prøve var 50 cm bred, 50 cm lang og 30 cm dyb. Prøverne blev gravet op med spade og derefter lagt på et stort stykke plastic. De tilstedeværende orme blev fundet ved at håndsortere prøven, hvorefter jorden blev lagt tilbage i hullet. Ormene blev placeret på vådt filterpapir i 24 timer for at tømme tarmen, hvorefter dyrene blev bestemt til art (adulte) eller slægt (juvenile) og vejet individuelt.
Leddyrene blev i maj indsamlet i to dybder, 0-5 og 5-10 cm. I hvert prøvefelt skulle der være taget 2 x 6 prøver, men antallet af prøver i 5-10 cm dybde blev i maj 2004 reduceret til 4 pr. prøvefelt pga., at jorden i maj 2004 var meget hård, hvilket gjorde prøvetagningen både mere tidskrævende og mere fysisk belastende end beregnet. I maj 2005 blev der taget et fuldt prøvesæt i begge dybder. Dyrene blev efter uddrivning i varme-/tørkegradient sorteret til art (springhaler) eller gruppe (mider) og talt.
I oktober 2004 og oktober 2005 blev der kun indsamlet leddyr, og kun i 0-5 cm dybde. Prøvetagning og – behandling foregik i øvrigt som beskrevet ovenfor.
3.4 Modelleringsdel – sanglærke, løbebille, tæppespinder
For sanglærke er der udført modelsimuleringer for bestandsudvikling samt reproduktivt output i et 10 x10km stort modellandskab beliggende ved Bjerringbro. Følgende output-variabler har været anvendt for sanglærkens vedkommende (alle variabler udtrykkes med én værdi årligt):
- Total population: alle individer i live ved migration om efteråret
- Total antal juvenile: alle årsunger fra det enkelte år der overlever til migration
- Klækningssucces: antal klækkede æg i alt som andel af antal lagte æg i alt
- Udflyvningssucces: antal udfløjne unger i alt som andel af antal klækkede æg i alt
- Ynglesucces: antal udfløjne unger i alt som andel af lagte æg i alt
- Redemortalitet: antal reder (før æglægning) der er gået tabt som følge af strigling(er)
- Ægkuldmortalitet: antal reder med æg (rugeperioden) der er gået tabt som følge af strigling(er)
- Ungekuldmortalitet: antal reder med redeunger der er gået tabt som følge af strigling(er)
Striglingens direkte (mortalitet) og indirekte (migration) effekter på løbebille og edderkop er som ovenfor nævnt ikke undersøgt eksperimentelt i nærværende projekt. Som datagrundlag for denne parameterisering i de anvendte artsmodeller anvendes som udgangspunkt data fra en eksperimentel dansk undersøgelse af striglingers effekter på bl.a. løbebiller (effekt=27 %) og edderkopper (effekt=35 %) (Thorbek & Bilde, 2004). Som output-variabler for edderkop og løbebille er anvendt ’det totale antal individer’ i modelområdet (hvert enkelt individ repræsenterer hver især et stort antal individer; Scheffer et al. 1995) registreret sidste dag i de enkelte år.
Der er udført følgende ni scenarier:
’Ordinære’ scenarier (1-2):
1) Kvæg_basis: standard konventionelt dyrkede kvægbedrifter (herunder at alle ukrudtsbehandlinger udføres ved kemisk bekæmpelse på alle bedrifter i modelområde)
2) Kvæg_strigling: som under 1, men alle ukrudtsbekæmpelser foregår mekanisk ved strigling (3 i vårbyg, 2 i vinterhvede) på alle bedrifter i modelområdet
Tekniske scenarier (3-8):
3) Vårbyg_basis: på alle marker i modelområdet dyrkes der vårbyg efter
standard konventionel dyrkningspraksis men uden anvendelse af herbicider og strigling
4) Hvede_basis: på alle marker i modelområdet dyrkes der vinterhvede efter
standard konventionel dyrkningspraksis men uden anvendelse af herbicider og strigling
5) Vårbyg_antal: den kumulerede effekt af hhv. (1), 2, 3 og 4 striglinger udført i konventionelt dyrket monokultur af vårbyg, hvor alle ukrudtsbekæmpelser foregår ved mekanisk strigling på alle bedrifter i model området
6) Hvede_antal: den kumulerede effekt af hhv. (1), 2, 3 og 4 striglinger udført i konventionelt dyrket monokultur af vinterhvede, hvor alle ukrudtsbekæmpelser foregår ved mekanisk strigling på alle bedrifter i modelområdet
7a) Vårbyg_tidspunkt: den isolerede effekt (striglingstidspunkt) af hhv. 1., 2., 3. og 4. strigling udført i konventionelt dyrket monokultur af vårbyg, hvor alle ukrudtsbekæmpelser foregår ved mekanisk strigling på alle bedrifter i modelområdet
7b) Vårbyg_sent: som under 7a, men hvor datoen for udførelsen af 4. og sidste strigling d. 10. maj er udskudt med hhv. 10 og 20 dage
8) Hvede_tidspunkt: den isolerede effekt (striglingstidspunkt) af hhv. 1., 2., 3. og 4. strigling udført i konventionelt dyrket monokultur af vinterhvede, hvor alle ukrudtsbekæmpelser foregår ved mekanisk strigling på alle bedrifter i modelområdet
Scenario 1 og 2 kan betegnes som ’ordinære’ scenarier, hvor effekterne af ukrudtsbekæmpelse via benyttelse af hhv. kemisk eller mekanisk vej undersøges i et ’standard’ konventionelt (kvæg) sædskifte, mens scenarierne 3 – 8 i det følgende betegnes som ’tekniske’ scenarier (scenarier med monokulturer af vårbyg og vinterhvede), hvor de rene, isolerede (scenarierne 5 og 6) eller kumulerede (scenarierne 7-8) effekter opstået som følge af ukrudtsstriglinger undersøges i to dyrkningssystemer, hvor der kun dyrkes en vår- eller vinterafgrøde. I scenarierne 1 og 2 er der kun undersøgt brugstypen ’kvægbrug’, da indførelsen af mekanisk (kun) ukrudtsbekæmpelse i svine- og plantebrug ville medføre et ændret sædskifte, hvilket ville fjerne muligheden for kun at sammenligne effekter, der skyldes ukrudtsstrigling(er).
For løbebille og edderkop modelleres den årlige bestandsudvikling i scenarierne 1 – 7a og 8.
Ved arealfordelingen anvendes hele bedriftsarealer. Under scenario 1 og 2 anvendes der alene standard sædskifte for brugstypen kvægbrug (Tabel 3.2) udarbejdet efter det tværfaglige forskningsprojekt ARLAS (Dalgaard et al. 2003 b).
Tabel 3.2. Oversigt over sædskifteafgrøder på bedriftstypen kvægbrug.
| Afgrøde |
| Vårbyg m. udlæg |
| 1. års kløver græs til slet/afgræsning |
| 2. års kløver græs til slet/afgræsning |
| Vinterhvede |
| Vårbyg |
| Vårbyg m. udlæg |
| 1. års kløver græs til slet/afgræsning |
| Vårbyg m. udlæg |
| Vårbyg |
For alle benyttede afgrøder anvendes der standardiserede dyrkningsmodeller udarbejdet på baggrund af officielle dyrkningsvejledninger (Dansk Landbrugsrådgivning, Landscenter) og konsulentrådgivning fra Djurslands Landboforening – Landbocentret Følle samt Godsforvaltningen ved Fussingø Statsskovdistrikt. Vækstmodeller for afgrøderne er udarbejdet på grundlag af eksisterende modeller (se endvidere Topping & Odderskær, 2004 for yderligere beskrivelse).
Det aktuelle landskab der danner baggrund for model scenarierne er et allerede digitaliseret modellandskab (10x10km) beliggende ved Bjerringbro, som tidligere er anvendt i tidligere undersøgelser. Hvor herbicidbehandling er erstattet af mekanisk ukrudtsbekæmpelse, er alle tidspunkter og frekvenser for striglinger (med undtagelse af scenario 7b) i de konventionelle afgrøder taget fra de tilsvarende økologiske dyrkningsmodeller for de samme afgrøder. I de tekniske scenarier udføres de fire striglinger på følgende datoer - vårbyg: 10/4, 20/4, 30/4 og 10/5, samt i scenario 7b) 20/5 og 30/5; - vinterhvede: 1/10, 31/3, 10/4 og 20/4. De første tre og to datoer for hhv. vårbyg og vinterhvede er midtpunktdatoer for det tidsinterval, striglingerne udføres i under økologiske forhold. Striglinger på de efterfølgende datoer udføres ikke under normale dyrkningsforhold, og er kun medtaget i de tekniske scenarier. Datoerne er valgt ved yderligere at tilføje 10 dage. Alle bedrifter er de aktuelle i området. For en mere uddybende beskrivelse af ovenstående modeller se Bilag A.
3.5 Systemanalyse af landbrugets ressourceforbrug og driftsøkonomi
I 2004 blev der opstillet en model til beregning af fossilt energiforbrug i jordbrugssystemer med og uden mekanisk ukrudtsbekæmpelse. Denne model, som er en tilpasning af Dalgaard et al.’s (2001) generelle model til opgørelse af landbrugets energiforbrug, er dokumenteret i bilag D. Dette bilag indeholder en oversigt over litteraturen på området, og dokumenterer hvorledes det direkte og indirekte energiforbrug opgøres i projektet.
Modellen er afgrænset til opgørelser af energiinput og -output i den primære landbrugsdrift. Dvs. det energiinput, der medgår til forarbejdning og transport, efter produkterne har forladt landbrugsbedrifterne, medregnes ikke, mens den energi, der går til at fremstille input, som anvendes på bedriften, samt den energi der går til indengårds transport, medregnes.
I Tabel 3.3 er vist en oversigt over de forskellige former for direkte og indirekte energiforbrug i landbruget, som opgøres i modellen. Det direkte energiforbrug omfatter de kilder, som direkte kan omsættes til energi (fx brændstof, smøremidler og elektricitet). Det indirekte energiforbrug omfatter den energi, der behøves for at producere de input som anvendes i landbrugsproduktionen (fx bygninger, maskiner, kraftfoder, kunstgødning, kalk og sprøjtemidler). Energiforbruget opgøres i Joule (J), hvor 1.000.000 J= 1.000 MJ=1 GJ. Fx er energiindholdet i dieselolie 35,9 MJ/L, og der medgår yderligere 14 % til udvinding, raffinering og transport af olien (Boo 1993). Dvs. energiforbruget ved forbrug af 1L diesel svarer til 40,9 MJ/L. 1 kilowatt-time el udgør tilsvarende 9,5 MJ, hvis energitabet ved fremstilling og distributionen medregnes.
Tabel 3.3. Oversigt over de forskellige former for direkte og indirekte energiforbrug i landbruget (Dalgaard et al., 2004).
| Direkte energiforbrug | Indirekte energiforbrug | |
| Brændstof | Øvrig energi | |
|
|
|
Opgørelsen af energiforbruget til mekanisk ukrudtsbekæmpelse udgør således kun en lille del af det samlede energiforbrugsregnskab (Tabel 3.3). Især bør det bemærkes, at en ukrudtsstrigling koster væsentligt mindre energi end stubharvning/kvikharvning eller skrælpløjning, der her også er kategoriseret som mekanisk ukrudtsbekæmpelse og listet under kategorien ”Plantebeskyttelse”. I Tabel 3.4 er vist et eksempel på brændstofforbruget til udvalgte markoperationer i de forskellige kategorier, og i Tabel 3.5 er vist eksempler på tilknyttede, øvrige direkte og indirekte energiforbrugsposter.
Tabel 3.4. Oversigt over nøgletal/normer (D) for dieselforbrug ved enkeltoperationer i afgrødeproduktionen. De viste normer (D) er ikke korrigeret for jordtype.
| D | ||
| Enhed | norm | |
| Jordbearbejdning og såning | ||
| Pløjning (21 cm), forår | L/ha | 20,0 |
| Pløjning (21 cm), efterår | L/ha | 23,0 |
| Jordpakning | L/ha | 2,0 |
| Såbedsharvning, let | L/ha | 4,0 |
| Såbedsharvning, tung | L/ha | 6,0 |
| Tromling | L/ha | 2,0 |
| Såning | L/ha | 3,0 |
| Gødskning og kalkning | ||
| Spredning & læsning, fast gødning | L/t | 0,6 |
| Spredning, gylle/ajle | L/t | 0,3 |
| Spredning, handelsgødning | L/ha | 2,0 |
| Kalkning | L/ha/år | 1,5 |
| Plantebeskyttelse | ||
| Sprøjtning | L/ha | 1,5 |
| Ukrudtsharvning | L/ha | 2,0 |
| Stubharvning/skrælpløjning | L/ha | 7,0 |
| Radrensning | L/ha | 3,0 |
| Høst og presning | ||
| Mejetærskning | L/ha | 14,0 |
| Roeoptagning | L/ha | 17,0 |
| Roeaftopning | L/ha | 10,0 |
| Græsslåning | L/ha | 5,0 |
| Højtrykspresning + håndtering | L/t | 2,0 |
| Afpudsning | L/t | 0,5 |
| Stængelbrydning | L/t | +0,2 |
| Grønthøstning | L/t | 1,0 |
| Transport, læsning og håndtering | ||
| Maskintransport | L/km | 0,04 |
| Gødnings-/ fodertransport | L/t/km | 0,2 |
| Læsning | L/t | 0,3 |
| Læsning og håndtering | L/t | 0,5 |
| Udfodring | L/t | 0,3 |
| Øvrig håndtering | L/t | 0,5 |
| Håndtering (total gennemsnit) | L/t | 1,3 |
Tabel 3.5. Oversigt over nøgletal for andet energibrug end diesel i afgrødeproduktionen.
| Enhed | Norm | |
| Smøremidler | MJ/(L diesel) | 3,6 |
| Maskiner | MJ/(L diesel) | 12 |
| Markvanding | MJ/mm | 52 |
| Korntørring | MJ/t/(%-point) | 50 |
| Kvælstof handelsgødning | MJ/(kg N) | 50 |
| Fosfor handelsgødning | MJ/(kg P) | 12 |
| Kalium handelsgødning | MJ/(kg K) | 7,0 |
| Jordbrugskalk | MJ/t | 30 |
| Herbicider | MJ/kg | 40a |
| Insecticider | MJ/kg | 40a |
| Fungicider | MJ/kg | 40a |
a) per kg formuleret middel
Til analyserne blev der opstillet en række typeafgrøder og typebedrifter, som anvendes i projektets scenarieberegninger. Eksempler på opgørelser af energiforbruget for disse typeafgrøder og typebedrifter er publiceret i Jørgensen & Dalgaard (2004). Typebedrifterne omfatter konventionelle malkekvægbrug, svinebrug, planteavlsbrug og økologiske malkekvægbrug, hvor afgrødesammensætningen og indsatsfaktorforbruget kendes fra Fødevareøkonomisk Instituts Regnskabsstatistik 1999 (se Jørgensen & Dalgaard 2004). Ud fra disse informationer kan maskin- og arbejdsomkostningerne opgøres, og dækningsbidraget beregnes på afgrødeniveau, ifølge principperne i Gravsholt et al. (2005). Nærværende projekt opgør således simple dækningsbidragskalkuler for udvalgte dyrkningspraksis og produktionsmuligheder. Mere avancerede modeller er beskrevet i Ørum (2003).
Arealanvendelsen på malkekvægbedrifterne er søgt repræsenteret ved det i afsnit 3.4 definerede modelsædskifte, som indgår i faunamodelleringen. Arealanvendelsen på kvægbrugene domineres således af græs- og kløvergræsmarker til slæt og afgræsning, samt marker med vårkorn og vinterkorn, hvoraf de fleste vårkornsmarker er med udlæg og/eller høstes som helsæd. Arealanvendelsen på plante- og svinebedrifterne omfatter desuden typiske salgsafgrøder såsom raps, frøgræs og ærter, samt en større andel af grønbrak end på kvægbedrifterne. I alt defineres følgende 8 typeafgrøder, som anvendes ved opgørelserne af de ressourcemæssige konsekvenser ved overgang til mekanisk ukrudtsbekæmpelse: vårkorn, vinterkorn, ærter, raps, frøgræs, helsæd, græs omdrift og grønbrak. Typeafgrøden ”Græs omdrift” samler kløvergræs- og græsmarker dyrket hhv. med henblik på slæt og afgræsning, idet det på baggrund af observationer på private studielandbrug antages, at 41 % af disse marker høstes til slæt, og den resterende del afgræsses. Denne fordeling er vigtig at kende, fordi der er stor forskel på energiforbruget ved afgræsning og ved slæt af græsmarker.
4 Statistiske analyser
4.1 Lærkeundersøgelse
4.1.1 Statistisk metode – kunstige reder
Hver kunstig rede er klassificeret som værende enten ødelagt eller intakt efter hver strigling, og data analyseres derfor vha. logistisk regression. På grund af de små antal er der valgt eksakt logistisk regression (som generaliserer Fishers eksakte test i 2 x 2 tabeller) med de 3 faktorer: mark, behandling og år. Der beregnes derfor kun p-værdier og ingen effekt-størrelser. Data analyseres efter 2. og efter 3. strigling, og efter 3. strigling vurderes endvidere om reder, der har overlevet 2. strigling, har samme chance for at overleve 3. strigling som de øvrige reder; denne faktor kaldes Status (S). Hvis ikke andet er anført, er statistiske analyser udført i SAS (SAS Institute, 2004).
4.1.2 Statistisk metode – naturlige reder
Studiet blev designet som et cross-over forsøg, hvor hver mark blev delt i 2 markhalvdele (plots), og den ene halvdel blev striglet det første år, men ikke det andet år og omvendt for den anden halvdel, jf. Tabel 4.1.
Tabel 4.1. Cross-over design.
| År | |||
| Mark | Markhalvdel | 2004 | 2005 |
| Keglehøj | Plot 1 | Ustriglet | Striglet |
| Plot 2 | Striglet | Ustriglet | |
| Krovang | Plot 3 | Ustriglet | Striglet |
| Plot 4 | Striglet | Ustriglet | |
Grundmodel
I dette design er forsøgsenheden plots. I princippet bør variationen mellem plots betragtes som ”tilfældig”; men der indgår kun 4 plots i alt og den behandles derfor som en systematisk faktor (P: 4 niveauer). Af samme årsag behandles variationen mellem de 2 år som en systematisk faktor (Y: 2 niveauer). Endvidere indgår behandling (striglet/ustriglet) som en systematisk faktor (B: 2 niveauer). Modellen for en vilkårlig responsvariabel (R) kan derfor symbolsk skrives som:
R = P + Y + B (1)
I et cross-over forsøg som dette kan man ikke udelukke en carry-over effekt, i dette tilfælde svarende til om foregående års behandling kan have indflydelse på responsen i år. En carry-over effekt svarer til tilstedeværelsen af en interaktion mellem Y og B, symbolsk skrevet Y*B. I det her valgte design er det dog ikke muligt at tilføje dette led til modeludtrykket (1) ovenfor, fordi effekterne P, Y, B og Y*B ikke længere vil være entydigt bestemt (som 2 ligninger med 3 ubekendte). Hvis man i stedet erstatter faktoren plots med mark (M: 2 niveauer) vil det være muligt at tilføje interaktionsleddet:
R = M + Y + B + Y*B (2)
Men i denne model antages implicit, at variationen mellem de 2 plots i samme mark er uden betydning for responsen. Vi har valgt at benytte model (1) fordi den bedst svarer til det valgte design med de etablerede plots, og vi antager derfor at carry-over effekten er forsvindende.
Analyse af ynglesucces
Som responsvariabel er benyttet "mindst 1 flyvefærdig unge pr rede". Det er antaget, at hvert ynglepar for hver af deres reder har samme sandsynlighed for at producere mindst en flyvefærdig unge. Med denne antagelse kan man for hvert ynglepar beskrive antallet af succesfulde yngleforsøg, dvs. forsøg med mindst 1 flyvefærdig unge, med en binomialfordeling med antalsparameter N = antal yngleforsøg og sandsynlighedsparameter = sandsynligheden for et succesfyldt yngleforsøg (for det givne par). Selve den statistiske analyse er gennemført som en standard generaliseret lineær model med binomialfordeling (= logistisk regression på aggregerede data).
Analyse af ungevægte
Ungevægte er analyseret som en ”mixed model”, symbolsk skrevet som:
W = P + Y + B + A + B*A + [U] + [R] + [R*A] + [E] (3)
De systematiske faktorer er: P, Y og B defineret som ovenfor (plot, år og behandling). Faktor A betyder alder (5, 7 eller 8 dage) og B*A er interaktion mellem behandling og alder. De tilfældige faktorer er: [U] svarende til gentagne målinger på den enkelte unge, [R] gentagne målinger på unger fra samme rede og [R*A] gentagne målinger på unger fra samme rede og med samme alder. [E] er residual-variation. En varianskomponent svarende til gentagne reder i samme territorium (samme ynglepar) var uden betydning.
Specielle problemer
I mange tilfælde kan yngleparrene nå at lave flere yngleforsøg i deres respektive territorier. Hvis yngleparrene ikke er lige succesrige (enten pga. yngleparrets egne evner eller det valgte territoriums egnethed) vil der kunne opstå en korrelation mellem responsen fra forskellige reder fra samme ynglepar. Dette problem er forsøgt løst via den såkaldte GEE-metode (GEE = Generalized Estimation Equation), som kort fortalt tillader korrelation mellem gentagne målinger på det enkelte territorium. I almindelighed påvirker det ikke estimaterne af de relevante parametre, men derimod øges deres standard errors og dermed vil p-værdierne blive større.
Responsvariablen CLUTCH er defineret som ´det maksimale antal lagte æg’. Pr. definition vil en aktiv rede indeholde mindst 1 æg (CLUTCH>0). Man kan derfor ikke umiddelbart anvende f.eks. en Poisson fordeling eller en negativ binomialfordeling til at beskrive antal æg pr. aktiv rede, fordi disse standardfordelinger vil tildele begivenheden ”0 æg” i en rede en positiv sandsynlighed. I stedet har vi valgt at bruge en såkaldt trunkeret Poisson fordeling (Grogger 1991), hvor der betinges med at CLUTCH skal være positiv. Det samme gælder for antal aktive reder pr. territorium.
Hvis reder med ”BROOD” (= maks. antal redeunger) eller ”FLEDGL” (= antal ’udfløjne’ unger) præderes, vil det oftest være hele reden der tømmes, dvs. det antal BROOD/FLEDGL, der registreres, vil blive 0. På den måde kan prædation give en overhyppighed af 0’er (og det understøttes faktisk af data), som ikke umiddelbart kan beskrives med f.eks. en Poisson fordeling. Fænomenet kaldes på engelsk ”zero-inflation” (Lambert 1992). For at løse dette problem har vi valgt en model, som kombinerer en Poisson model for BROOD/FLEDGL med en logistisk regressionsmodel for overhyppigheden af 0’er. Den logistiske regressionsmodel kan modelleres efter samme koncept som Poissonmodellen, men de relativt få observationer sætter grænser for hvor mange effekter der kan estimeres. I prioriteret rækkefølge har vi valgt at fokusere på: 1. forskel på de 2 år, 2. forskel på de 2 marker og 3. forskel på ustriglet/striglet. Analyserne med trunkeret og ”zero-inflated” Poisson-model med GEE er lavet ved hjælp af Stata 9 (StataCorp. 2005).
Tolkning af parameterestimater
Poisson-modellerne er såkaldte log-lineære modeller, dvs. den naturlige logaritme til middelværdien er lineær i de forklarende variable, jf. Model (1) ovenfor. Den statistiske analyse giver derfor estimater på den logaritmiske skala. Når estimaterne transformeres tilbage med exponentialfunktion fås estimater for de relative forskelle i middelværdierne. Betragt f.eks. behandlingen (B) i Model (1). Hvis vi vælger behandlingsniveauet ”Ustriglet” som reference, vil det tilbagetransformerede estimat for ”Striglet” kunne fortolkes som den relative forskel i forventet antal mellem ”Striglet” og ”Ustriglet”. Hvis det tilbagetransformerede estimat er 0.9, vil det forventede antal på den striglede markhalvdel være 90 % af antallet på den ustriglede markhalvdel etc.
Logistisk regression/binomialfordelings-modellerne (mindst 1 flyvefærdig unge pr. rede, zero-inflation) er log-lineære på odds, dvs på den naturlige logaritme til forholdet mellem sandsynlighederne for ”succes” og ”ikke succes”. Når de tilsvarende estimater tilbagetransformeres med exponentialfunktion fås estimater for odds ratio (OR), det vil sige forholdet mellem 2 odds. Som i Poissonmodellen vælges et referenceniveau, f.eks. ”År 2004”. Odds ratio for år 2005 vil da udtrykke forholdet mellem de 2 odds for år 2005 og år 2004. Hvis OR<1 vil odds for 2005 være mindre end odds for 2004, det betyder at sandsynligheden for succes vil være mindre i 2005 end i 2004. Hvis OR=1 er der ikke forskel på de 2 år, og hvis OR>1 vil sandsynligheden for succes være større i 2005 end i 2004. Bemærk, at ”succes” bruges i betydningen ”et redeforsøg med min. en udfløjen unge”.
4.1.3 Statistisk metode – modelleringer på landskabsniveau
Modelsimuleringer bliver udført (10 replikater) for de udvalgte arter for en periode på 50 år. Statistiske analyser af forskelle mellem de enkelte scenarier er meningsløse (Topping & Odderskær, 2004), der kan udføres så mange replikater der kræves for at opnå en given fastsat konfidensgrænse for resultaterne (se endvidere afsnit 5.3).
4.2 Mikroleddyr, regnorme og ukrudt/høstudbytte
Data for antal dyr, antal arter og vådvægt blev analyseret vha. variansanalyse for effekter af behandling (antal striglinger samt striglet/ustriglet), blok og behandling*blok med blok som tilfældig variabel (proc MIXED i SAS; SAS Institute, 1992). Data for springhaler og mider blev kvadratrodstransformeret, idet denne transformation i gennemsnit gav den bedste tilnærmelse til normalfordelingen. Test af utransformerede og logaritmetransformerede data viser imidlertid, at analyserne er temmelig ufølsomme over for valget af transformation. Da der i nogle tilfælde ikke kunne opnås varianshomogeneitet, blev der taget hensyn til dette ved at indføre et ”repeated”- statement i proc mixed.
Der blev anvendt variansanalyser til at undersøge om strigling påvirker ukrudtet og afgrøden. Sammenhængen mellem ukrudtsbiomasse og jordfaunadata blev undersøgt ved at teste Spearman korrelationskoefficient, som er ret ufølsom over manglende normalfordeling og ikke forudsætter lineær sammenhæng, i proc CORR i SAS.
4.3 Ukrudt og høstudbytte
Ukrudtsbiomasse og høstudbytte (kerne og halm) blev analyseret statistisk med en variansanalysemodel med behandling, mark og år som systematiske klassevariabler og blok var nestet inden for år og mark. Alle vekselvirkninger mellem behandling, mark og år ved inkluderet i modellen. Efter inspektion af residualerne blev det besluttet at log-transformere alle data for at skabe varianshomogenitet. Ved opsplitning af kvadratafvigelsessummen (SAK) blev det testet om der var forskelle mellem de behandlede parceller (1, 2 og 3 harvninger) eller forskelle mellem de behandlede i én gruppe (1, 2 og 3 harvninger) og de ubehandlede (0 harvninger) jævnfør Little (1991). Data blev analyseret med proceduren GLM i SAS.
5 Resultater
- 5.1 Eksperimentelle forsøg
- 5.2 Landbrugets ressourceforbrug
- 5.3 Fauna modellering på Landskabsniveau
5.1 Eksperimentelle forsøg
5.1.1 Sanglærke
Effekter på kunstige reder – pilotforsøg 2003
Lidt over halvdelen af rederne gik tabt ved første strigling. Af de ”overlevende” reder fra 1. strigling var der efter 2. strigling en mortalitet på 78 %. Kun ca. én procent af rederne overlevede de to på hinanden følgende striglinger.
Effekter på kunstige reder - eksperimentelle forsøg 2004 og 2005
Resultaterne af 2. og 3. strigling i 2004 og 2005 er vist i Figur 5.1 A og B. Tabet af reder ved 2. strigling var i gennemsnit (begge forsøgsmarker) ens i begge år (73 %), hvorimod det ved 3. strigling var lavere (65 %) i 2004 end i 2005 (79 %). Redemortaliteten set i relation til jordtype varierede mellem 44 % - 100 % i 2004 og mellem 50 – 100 % i 2005. Der var ingen signifikante forskelle på redeoverlevelsen i relation til år, mark og jordtyper efter 2. strigling (År: p=1, Mark: p=0.16, Jordtype: p=0.33). Ved 3. strigling var der en signifikant forskel på redemortaliteten mellem jordtyper (størst i den ’stenet/lerede’ jordtype mindst i den ’sandet/løse’). Der kunne ikke konstateres nogen signifikant forskel i overlevelsen mellem reder, der overlevede 2. strigling, og de nye reder der blev lagt ud ved 3. strigling. Det samlede gennemsnit for redetab (begge marker og år) var på 72,4 %. Dette gennemsnit blev benyttet som fast variabelværdi for redemortalitet i forbindelse med striglinger i de udførte modelsimuleringer. I begge forsøgsår var der 13 kunstige reder, der ’overlevede’ den 2. strigling, og ud af disse overlevede kun hhv. 5 (2004) og 6 (2005) den tredje og sidste strigling.
Ægmortaliteten (% ødelagte og bortkomne æg i alt jvf. afsnit 3.2.1) var relativt lav i begge år (< 20 %), højest efter 3. strigling (Figur 5.1 B).
Registrering af territorier
Der blev i 2004 i alt registreret 17 territorier på Krovang marken – hvoraf der blev foretaget yngleforsøg i de 16. Alle territorielle hanner blev udparrede. På Keglehøj blev der registreret 15 territorier, hvoraf der blev foretaget yngleforsøg i de 14. Ligesom på Krovang blev samtlige territorielle hanner udparrede. Territorietætheden i 2005 var betydeligt lavere med 13 territorier og yngleforsøg på Krovang marken, og 9 territorier med 7 yngleforsøg på Keglehøj. Fordelingen af aktive reder i relation til år/mark/behandling er vist i Tabel 5.1.
Territoriebesættelsen foregik begge år sent på begge marker. I 2004 foregik den ca. tre uger tidligere på Keglehøj (primo maj), mens territoriebesættelsen i 2005 tidsmæssigt var ens på begge marker. I begge år var territoriebesættelsen først tilendebragt i slutningen af maj.
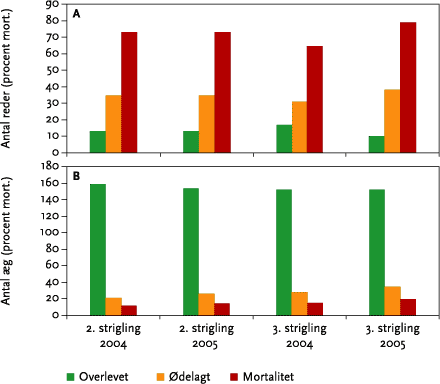
Figur 5.1. Den samlede dødelighed for kunstige reder (A) og æg (B) (begge forsøgsmarker i 2004 og 2005), efter 2. og 3. strigling i 2004 og 2005. Mortalitet = den samlede procentvise dødelighed for hhv. reder og æg.
Tabel 5.1. Antal aktive reder fordelt efter år, mark og behandling.
| 2004 | 2005 | ||
| Mark/Behandling | Reder i alt | Reder i alt | |
| Keglehøj | Striglet | 14 | 8 |
| Ustriglet | 11 | 8 | |
| Krovang | Striglet | 10 | 8 |
| Ustriglet | 17 | 9 | |
| Reder i alt | 52 | 33 | |
Yngleforsøg og ynglesucces
Ynglesæsonen var meget lang i både 2004 og 2005. Den sidste aktive rede (= rede med æg/unger) blev fulgt indtil d. 20. august, hvilket er ca. en måned længere end normalt for de fleste andre konventionelt drevne marker med veletablerede kornafgrøder.
Der blev i 2004 foretaget i alt 59 yngleforsøg på de to forsøgsmarker fordelt på 32 territorier (heraf 7, reder hvor der ikke blev påbegyndt æglægning). I 2005 blev der foretaget 33 yngleforsøg fordelt på 20 territorier (Tabel 5.1 og Figur 5.2 A). Der kunne ikke påvises forskel på fordelingen af antal aktive reder pr. territorium mellem de 2 behandlinger eller de to år (p=0.58, 0.56; zero truncated Poisson-model med plot (markhalvdel), år og behandling - se design skitse afsnit 4.1).
Fordelingen af lagte æg, redeunger og udfløjne unger i relation til år, mark og behandling (striglet/ustriglet) er angivet i Tabel 5.2.
Som den eneste variabel er ’ynglesuccesen’ også blevet testet med den statistiske ’Model 2’ beskrevet i afsnit 4.1. Der blev ikke fundet nogen interaktion mellem år og mark (Y*B>0.05), og der blev ikke fundet nogen signifikante forskelle på antallet af succesfulde reder (= reder med min en ’udfløjen’ unge) set i relation til mark og behandling (Tabel 5.3 og Figur 5.2), selvom der her var en tendens til flere succesfulde reder på Krovang marken. Antallet af succesfulde reder i 2005 var signifikant større end i 2004.
Tabel 5.2. Antal æg/unger/udfløjne unger i relation til år, mark og behandling.
| 2004 | 2005 | ||||
| Striglet | Ustriglet | Striglet | Ustriglet | ||
| Antal æg | Keglehøj | 47 | 41 | 30 | 29 |
| Krovang | 40 | 62 | 26 | 36 | |
| Klækkede æg | Keglehøj | 19 | 17 | 17 | 20 |
| Krovang | 26 | 35 | 15 | 29 | |
| Udfløjne unger | Keglehøj | 11 | 10 | 13 | 16 |
| Krovang | 16 | 21 | 15 | 25 | |
Tabel 5.3. Odds Ratio (OR) for ”ynglesucces” (=min. en udfløjen unge) vs ”ikke-ynglesucces”, når det viste niveau sammenlignes mod en valgt reference for hver af faktorerne: år (Y), Behandling (B) og Mark (M). jf. grundmodellerne (1) og (2). I model 2 er forinden testet for interaktion Y*B (”carry over” effekt), p=0.40.
| Model 1 | |||||
| 95% CI | |||||
| Effekt | OR | Nedre | Øvre | p | |
| År | 2005 | 3,75 | 1,42 | 9,95 | 0,008 |
| 2004 | 1 | (reference) | |||
| Behandling | Striglet | 0,55 | 0,21 | 1,46 | 0,23 |
| Ustriglet | 1 | (reference) | |||
| Model 2 | |||||
| 95% CI | |||||
| Effekt | OR | Nedre | Øvre | p | |
| År | 2005 | 3,74 | 1,45 | 9,66 | 0,006 |
| 2004 | 1 | reference | |||
| Behandling | Striglet | 0,65 | 0,26 | 1,64 | 0,36 |
| Ustriglet | 1 | reference | |||
| Mark | Keglehøj | 0,47 | 0,19 | 1,19 | 0,11 |
| Krovang | 1 | reference | |||
Effekter på æg og unger
I ingen af årene blev der blev registreret redeforsøg ved tidspunktet for første strigling. I 2004 blev to reder udsat for 2. strigling - heraf var den ene med et æg. Begge reder blev opgivet få dage efter strigling. I 2005 blev der ikke konstateret redeforsøg ved 2. strigling. Ved 3. strigling blev yderligere to reder udsat for strigling i 2004 – begge før æglægning. Den ene af disse blev opgivet dagen efter strigling. I 2005 blev en rede m. æg udsat for 3. strigling. Den ”udsatte” rede overlevede striglingen. Samlet betragtet har den direkte effekt af strigling - 5 ud af 92 redeforsøg - ikke haft nogen indflydelse på den samlede reproduktion i striglede og ustriglede markafsnit på de to marker.
Der blev ikke fundet nogen signifikante forskelle mellem år eller behandlingstyper i antallet af æg (ved start af rugning), redeunger (max antal) og ’udfløjne’ unger pr. rede (Tabel 5.4 ). Til sammenligning med de estimerede sandsynligheder for mislykkede (hovedsageligt præderede) reder (’p-zero’ i Tabel 5.4) var de tilsvarende observerede værdier for ’0 redeunger’ 44 og 19 % i hhv. 2004 og 2005, og værdierne for ’0 udfløjne unger’ var 63 og 33 % i hhv. 2004 og 2005.
Antallet af æg/redeunger/udfløjne unger pr. territorium var ikke forskelligt mellem år og mellem behandlingstyper (Tabel 5.5), men der var en tendens til højere værdier for alle tre variabler på Krovang marken (Tabel 5.5 og 5.2).
Tabel 5.4. Estimater af relative forskelle i forhold til valgt reference af antallet af æg/redeunger/udfløjne unger pr. rede som en funktion af år og behandling (Se afsnit 4.1.2). En sammenligning af Zero inflation (Se forklaring i afsnit 4.1.2) i de to forsøgsår 2004 og 2005 gav: redeunger p = 0,076; udfløjne unger: p = 0,004.
| 95% CI | ||||
| Æg | Estimat | Nedre | Øvre | P |
| 2005 | 0,99 | 0,91 | 1,08 | 0,797 |
| 2004 | 1 | (reference) | ||
| Striglet | 0,95 | 0,86 | 1,04 | 0,28 |
| Ustriglet | 1 | (reference) | ||
| Redeunger | Estimat | Nedre | Øvre | P |
| 2005 | 0,94 | 0,79 | 1,12 | 0,484 |
| 2004 | 1 | (reference) | ||
| Striglet | 0,93 | 0,77 | 1,11 | 0,484 |
| Ustriglet | 1 | (reference) | ||
| p(zero,2004) | 0,42 | |||
| p(zero,2005) | 0,14 | |||
| Udfløjne unger | Estimat | Nedre | Øvre | P |
| 2005 | 1,03 | 0,81 | 1,3 | 0,822 |
| 2004 | 1 | (reference) | ||
| Striglet | 1,1 | 0,88 | 1,38 | 0,41 |
| Ustriglet | 1 | (reference) | ||
| p(zero,2004) | 0,61 | |||
| p(zero,2005) | 0,29 | |||
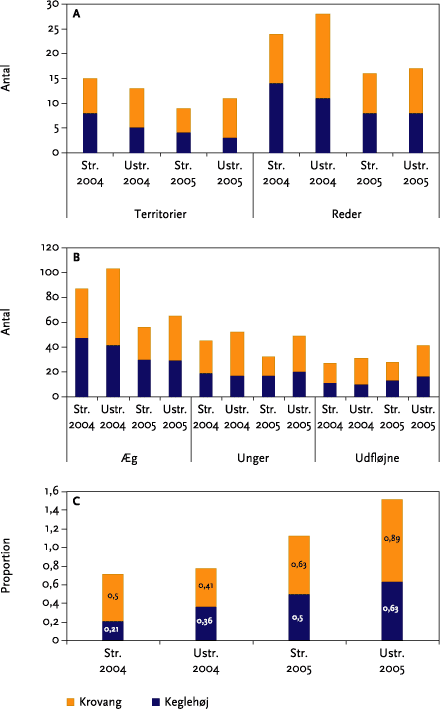
Figur 5.2. Antallet af Territorier, Aktive reder (m. æg/unger) (A), og (B) æg, redeunger, og udfløjne unger i relation til år, mark (Kr = Krovang, Ke = Keglehøj) og behandling (ST = striglet, UST = ustriglet). C: andelen af succesfulde reder (min. èn udfløjen unge) i relation til år, mark og behandling.
Antallet af reder/æg/redeunger/udfløjne unger pr. år, mark og behandlingstype gav den samme tendens som beskrevet for reproduktionen pr. territorium på Krovang, specielt på den ustriglede markhalvdel af Krovang marken (Figur 5.2 A og B). Denne tendens ses ikke i andelen af succesfulde reder i hverken 2004 eller 2005 (Figur 5.2 C).
Tabel 5.5. Estimater af relative forskelle i forhold til valgt reference af antallet af æg/redeunger/udfløjne unger pr. territorium som en funktion af år og behandling (se forklaring i afsnit 4.1.2). En sammenligning af Zero inflation (se afsnit 4.1.2) i de to forsøgsår 2004 og 2005 gav: redeunger: p = 0,006; udfløjne unger: p = 0,004. p (zero) = estimerede sandsynlighed for mislykkede reder.
| 95% CI | ||||
| Æg | Estimat | Nedre | Øvre | P |
| 2005 | 1,06 | 0,84 | 1,34 | 0,612 |
| 2004 | 1 | (reference) | ||
| Striglet | 0,90 | 0,71 | 1,14 | 0,392 |
| Ustriglet | 1 | (reference) | ||
| Redeunger | Estimat | Nedre | Øvre | P |
| 2005 | 1,02 | 0,75 | 1,38 | 0,89 |
| 2004 | 1 | (reference) | ||
| Striglet | 1 | 0,74 | 1,36 | 0,99 |
| Ustriglet | 1 | (reference) | ||
| P("zero",2004) | 0,30 | |||
| P("zero", 2005) | 0 | |||
| Udfløjne unger | Estimat | Nedre | Øvre | p |
| 2005 | 1,20 | 0,83 | 1,75 | 0,34 |
| 2004 | 1 | (reference) | ||
| Striglet | 1,08 | 0,74 | 1,57 | 0,691 |
| Ustriglet | 1 | (reference) | ||
| P(zero, 2004) | 0,45 | |||
| P(zero, 2005) | 0,13 | |||
Effekter på ungevægt
Der blev ikke fundet nogen signifikante forskelle mellem ungevægtene (beregnet på data for unger vejet både på 5., 7. og 8. dagen) set i relation til behandling (p=0,76) (behandling * alder: p = 0,62). Den gennemsnitlige forskel mellem ungevægtene på de striglede og de ustriglede markhalvdele (plots) 5., 7. og 8. dagen var hhv. -0,11g, 0,36g og 0,68g.
5.1.2 Mikroleddyr og regnorme
Regnorme 2004 og 2005
Der var ingen signifikante effekter af antal striglinger på antal regnorme, antal regnormearter eller regnormevådvægt i juni 2004 (p mellem 0,33 og 0,81). Bloknummeret bidrog med 3,7-40 % af den samlede tilfældige varians, interaktionen mellem bloknummer og behandling med 0-4,9 %. Heller ikke hvis man udelukkende skelner mellem striglet og ikke striglet, dvs. ikke tager hensyn til antal striglinger, var der signifikante effekter på regnormene (p>0,33). Middelværdier og standardfejl fremgår af Figur 5.3.
Heller ikke i maj 2005 var der signifikante effekter af antal striglinger på antal regnorme, antal regnormearter eller regnormevådvægt (p mellem 0,41 og 0,61). Bloknummeret bidrog med 0-36 % af den samlede tilfældige varians, interaktionen mellem bloknummer og behandling med 0-27 %. Heller ikke hvis man udelukkende skelner mellem striglet og ikke striglet, dvs. ikke tager hensyn til antal striglinger, var der signifikante effekter på regnormene (p>0,2). Middelværdier og standardfejl fremgår af Figur 5.3.
Mikroleddyr maj og oktober 2004 og 2005
For begge prøvetagninger gælder, at effekten af striglingerne på antal mider og springhaler varierede meget mellem blokkene, hvilket kan ses af det store interval for bloknummerets bidrag til den samlede, tilfældige variation (Tabel 5.6), idet det numeriske blokbidrag er fælles for alle blokke, mens residualvariationen varierer kraftigt, og blokbidraget i % derfor varierer med residualvariationen. Der var ikke tale om nogen entydig effekt af forskellige antal striglinger (Figurerne 5.3-5.6), idet der i maj 2004 for antal springhalearter i dybden 0-5 cm var en signifikant positiv effekt af antal striglinger, mens der i dybden 5-10 cm var en signifikant negativ effekt på antal springhaler. Maj 2005 var der en signifikant positiv effekt på såvel antal springhaler som antal springhalearter i dybden 5-10 cm. Når datamaterialet blev testet for effekt af strigling mod ingen strigling, var der en statistisk signifikant, men numerisk meget lille, negativ effekt på antal springhalearter oktober 2004 samt en signifikant positiv effekt på antallet af springhaler i maj 2005.
Tabel 5.6. P-værdier og bloknummerets bidrag til den samlede, tilfældige variation for variansanalyser af effekten af strigling/ingen strigling samt antal striglinger på springhaler og mider i maj og oktober 2004 og 2005. Majprøver er taget i to dybder (0-5 cm og 5-10 cm), mens oktoberprøver kun er taget i en dybde (0-5 cm).
| Striglet/ustriglet | Antal striglinger | ||||||||
| 0-5 cm | 5-10 cm | 0-5 cm | 5-10 cm | ||||||
| p | Blok-var, % | P | Blok-var, % | p | Blok-var, % | P | Blok-var, % | ||
| Maj 2004 | Springhaler | 0,17 | 0,02-1,7 | 0,51 | 1,7-17 | 0,47 | 0,1-18 | 0,0002 | 1,5-47 |
| Springhale- arter |
0,12 | 0 | 0,15 | 0 | 0,0007 | 0 | 0,45 | < 6·10-36 | |
| Mider | 0,37 | 7,5-45 | 0,49 | 0 | 0,90 | 5,2-21 | 0,089 | 0 | |
| Maj 2005 | Springhaler | 0,024 | 0 | 0,59 | 1,1-13 | 0,64 | 0 | 0,033 | 0,69-21 |
| Springhale- arter |
0,55 | 1,6-11 | 0,29 | 2,2-15 | 0,88 | 0 | 0,0025 | 4,7-48 | |
| Mider | 0,12 | 26-60 | 0,43 | 5,8-38 | 0,19 | 16-61 | 0,36 | 2,5-41 | |
| Okt. 2004 | Springhaler | 0,38 | 0 | 0,66 | 0 | ||||
| Springhale- arter |
0,016 | < 6,8·10-16 | 0,91 | 0 | |||||
| Mider | 0,48 | 0 | 0,84 | 0 | |||||
| Okt. 2005 | Springhaler | 0,88 | 4,1-20 | 0,057 | 2,2-11 | ||||
| Springhale- arter |
0,43 | 13-39 | 0,40 | 10-42 | |||||
| Mider | 0,54 | 8,8-15 | 0,066 | 7,5-29 | |||||
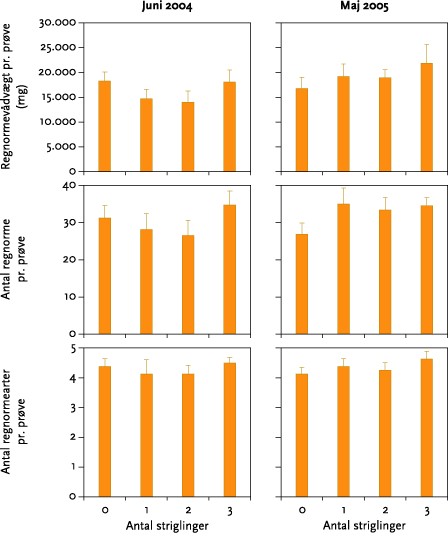
Figur 5.3. Regnormefriskvægt, antal regnorme og antal regnormearter pr. prøve (50 cm bred x 50 cm lang x 30 cm dyb) juni 2004 og maj 2005. Søjlerne repræsenterer middelværdier og standardfejl på disse.
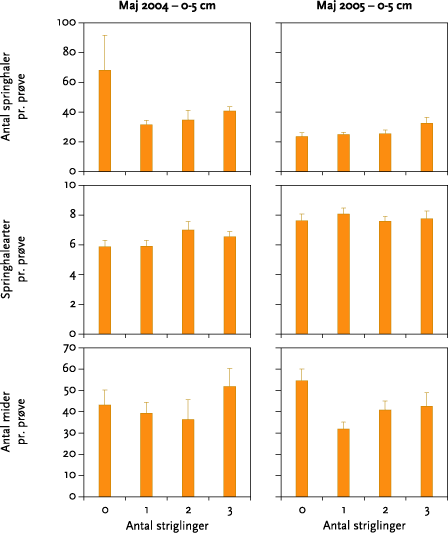
Figur 5.4. Antal springhaler, springhalearter og mider pr. prøve (0,0028 m²) i 0-5 cm dybde maj 2004 og maj 2005 som funktion af antal striglinger. Figuren viser middelværdier og SEM.
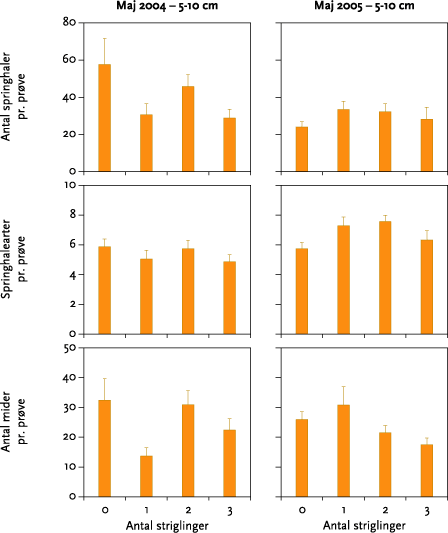
Figur 5.5. Antal springhaler, springhalearter og mider pr. prøve (0,0028 m²) i 5-10 cm dybde maj 2004 og maj 2005 som funktion af antal striglinger. Figuren viser middelværdier og SEM.
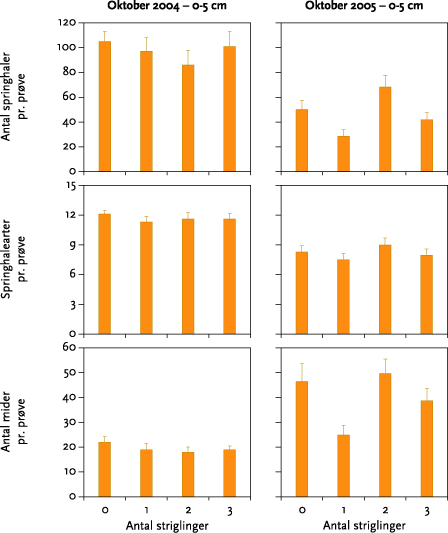
Figur 5.6. Antal springhaler, springhalearter og mider pr. prøve (0,0028 m²) i 0-5 cm dybde oktober 2004 og oktober 2005 som funktion af antal striglinger foretaget foråret 2004. Figuren viser middelværdier og SEM.
Korrelationer mellem ukrudt og jordfauna
Der var for 2004-data ingen signifikante sammenhænge mellem ukrudtbiomasse og hvirvelløse dyr (R<0,38, p>0,15). For 2005-data var der en signifikant positiv korrelation mellem regnormevådvægt og ukrudtbiomasse (R=0,55, p=0,029) og mellem antal mider i dybden 0-5 cm og ukrudtbiomassen (R=0,53, p=0,036). For de øvrige variable for hvirvelløse dyr var der ingen signifikant korrelation med ukrudtbiomassen (R<0,18, p>0,35).
5.1.3 Ukrudt og høstudbytte
Blokforsøg
Den statistiske analyse viste, at der med hensyn til ukrudtsbiomasse ikke var vekselvirkninger mellem strigling og år (P = 0,30), strigling og mark (P = 0,65) og strigling, år og mark (P = 0,57). Et lignende mønster blev fundet for kerneudbytter og halmudbytter, hvor der heller ikke var vekselvirkninger mellem strigling, år og mark. For kerneudbyttet var de tilsvarende P-værdier: 0,67, 0,59 og 0,24. Da striglingseffekterne udtrykt i procent således var konstante over år og mark, præsenteres der i det følgende kun gennemsnit over år og marker i form af tilbagetransformerede modelestimater.
I gennemsnit reducerede striglingerne ukrudtsbiomassen 33 % (P < 0.001), men der var ingen sikker forskel mellem 1, 2 og 3 striglinger (P = 0.91) (Figur 5.7). Den procentuelle effekt af strigling var således uafhængig af ukrudtsfloraens sammensætning, der på Krovang var domineret af fuglegræs, ærenpris-arter og ager sennep og på Keglehøj af hvidmelet gåsefod, fuglegræs og ærenpris-arter. I 2005 var ager sennep totalt dominerede på Krovang, og der var så stor en ukrudtsbiomasse (cirka 200 g/m²), at der var tale om et reelt og betydende ukrudtsproblem (Figur 5.8).
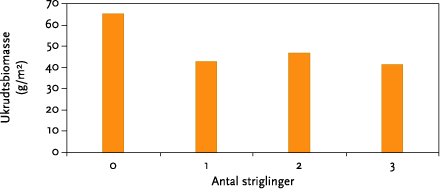
Figur 5.7. Ukrudtsbiomasse ved et stigende antal striglinger. Estimerede middelværdier over år og mark.
Høstudbyttet var negativt påvirket af strigling (P = 0,02), men de var ingen forskel mellem 1, 2 og 3 striglinger (P = 0.83) (Figur 5.9). Strigling reducerede i gennemsnit udbytterne 6 %. Udbytterne var generelt højere på Keglehøj (P < 0,001) og i 2005 var udbytteniveauet højere end i 2004 i begge marker (P < 0,001) (Figur 5.10). Ved visuel bedømmelse af afgrøden efter sidste strigling, var det ikke muligt at skelne de ubehandlede parceller fra det behandlede, hvilket vidner om at striglingerne har været skånsomme.
Halmudbyttet fulgte i høj grad udbytterne i kerne ved sammenhold af striglingernes effekt på kerne- og halmudbytte (Figur 5.9 og 5.11). Der var sikker effekt af striglingerne (P = 0,0014) men ingen forskel mellem 1, 2 og 3 striglinger (Figur 5.11). Halmudbytte var bemærkelsesværdigt stort på Krovang i 2005, hvilket ikke umiddelbart kan forklares (Figur 5.12).
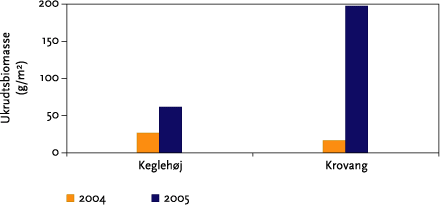
Figur 5.8. Gennemsnitlig ukrudtsbiomasse på de to marker i de to år. Estimerede middelværdier over 0, 1, 2 og 3 striglinger.
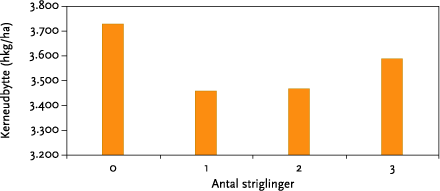
Figur 5.9. Kerneudbytte ved et stigende antal striglinger. Estimerede middelværdier over år og mark.
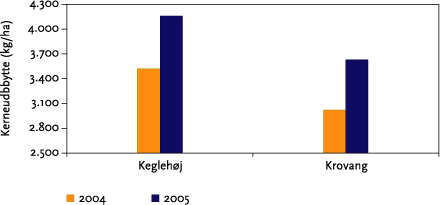
Figur 5.10. Gennemsnitlig kerneudbytte på de to marker i de to år. Estimerede middelværdier over 0, 1, 2 og 3 striglinger.
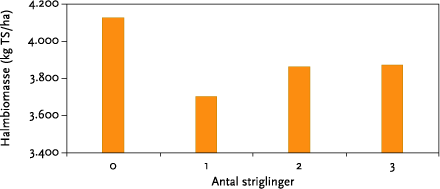
Figur 5.11. Halmudbytte ved et stigende antal striglinger. Estimerede middelværdier over mark og år.
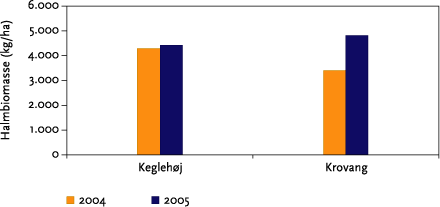
Figur 5.12. Gennemsnitlig halmudbytte på de to marker i de to år. Estimerede middelværdier over 0, 1, 2 og 3 striglinger.
Markforsøg
På Krovang var der en stor, men meget uensartet bestand af Agertidsel, som uden tvivl har konkurreret med afgrøden og på Keglehøj var der en meget uensartet bestand af Alm. Kvik, som også har konkurreret med afgrøden. Da flerårige arter ikke påvirkes af strigling, er disse arter ikke analyseret nærmere.
Ukrudtet i blokforsøgene var repræsentativt for markforsøgene med hensyn til artssammensætning. I markforsøgene var de naturlige variationer i ukrudtet imidlertid så stor, at det ikke var muligt at registrere nogen generel effekt af striglingerne på ukrudtet. En væsentlig årsag til variationen i ukrudtsmængden og artssammensætningen på markniveau skyldes meget væsentlige forskelle i jordbundsforholdene. I Figur 5.13 er vist et eksempel på dette. De fire billeder viser hvor stor variation der var på Krovang og Keglehøj markerne den 28. maj 2004.

Figur 5.13. Eksempler på variation i afgrødens vækst forskellige steder på de to forsøgsmarker Krovang (de to øverste billeder) og Keglehøj (de to nederste billeder) den 28. maj 2004.
Af figuren (nederste række) fremgår det, at der var en nøje positiv kobling mellem afgrødens og ukrudtets biomasseproduktion. Hvor afgrøden trivedes godt, trivedes ukrudtet også godt. De registrerede forskelle i udbytter mellem striglede og ikke striglede markhalvdele (Tabel 5.7), kan ikke betragtes som udtryk for striglingernes effekt, da de ikke er i overensstemmelse med blokforsøgene. Resultaterne må betragtes som udslag af tilfældig variation.
Tabel 5.7. Kerneudbytter (kkg) fra markforsøgene i 2004 og 2005 i relation til mark og behandling (ST = striglet; UST= ustriglet).
| Krovang | Keglehøj | |||
| ST | UST | ST | UST | |
| 2004 | 31,6 | 33,9 | 53,5 | 52,1 |
| 2005 | 46,7 | 55,5 | 65,3 | 60,3 |
5.2 Landbrugets ressourceforbrug
5.2.1 Energiforbruget på typiske landbrugsbedrifter (Scenario A-D)
Dieselforbruget er opgjort for hver af de 8 definerede afgrødetyper, dyrket ved brug af kemisk ukrudtsbekæmpelse. Derved kan dieselforbruget til planteværn i lighed med Bichel-udvalgets opgørelser (Miljøministeriet 1998a, 1998b) sammenlignes med det øvrige brændstofforbrug (Figur 5.14), og forskellen ved indførsel af mekanisk ukrudtsbekæmpelse kan vurderes (Figur 5.15). Som det ses, udgør dieselforbruget for afgrøder dyrket på lerjord, typisk mellem 15 og 125 liter per ha, idet dieselforbruget til plantebeskyttelse incl. stubharvning for alle afgrøder udgør under halvdelen af det samlede dieselforbrug. Det ses også, at der er stor forskel på det samlede dieselforbrug for de forskellige afgrøder. Ud over dieselforbruget til ukrudtsbekæmpelse (incl. stubharvning med henblik på bekæmpelse af kvik), er det især poster som pløjning (jordbehandling), udspredning og transport af husdyrgødning, samt høst og håndtering af grovfoderafgrøder, der vejer tungt.
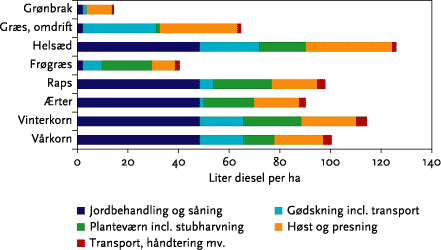
Figur 5.14. Eksempel på modelleret dieselforbrug for otte typeafgrøder, dyrket på lerjord og med kemisk ukrudtsbekæmpelse.
Det samlede direkte og indirekte energiforbrug er beregnet for typiske, konventionelle malkekvægbrug, planteavlsbrug, svinebrug, og for et typisk økologisk malkekvægbrug (Tabel 5.8). Det ses, at det indirekte energiforbrug til pesticider i alle tilfælde udgør under 1 % af det totale energiforbrug, og at forbruget af olie og smøremidler, hvorunder mekanisk ukrudtsbekæmpelse rubricerer, for alle bedriftstyper udgør en betydelig andel af det samlede energiforbrug (mellem 15 og 30 %). Dieselforbruget til stubharvning udgør en betydelig del af dieselforbruget til ukrudtsbekæmpelse i forhold til dieselforbruget til ukrudtsstrigling og sprøjtning (jf. bilag D).
Tabel 5.8. Beregnet fordeling af det direkte og indirekte energiforbrug på typiske, konventionelle malkekvægbrug (Scenario A), svinebrug (scenario B) og planteavlsbrug (scenario C). Desuden vises til sammenligning energiforbruget på et typisk økologisk malkekvægbrug scenario D).
| Energiforbrug (MJ/ha) | Malkekvæg (Scenario A) |
Svin (Scenario B) |
Planteavl (Scenario C) |
Øko-malkekvæg (Scenario D) |
| Olie og smøremidler | 5580 | 8315 | 5123 | 4123 |
| El | 4598 | 7095 | 2519 | 3864 |
| Handelsgødning og kalk | 3807 | 4184 | 4656 | 501 |
| Pesticider | 5 | 24 | 22 | 0 |
| Bygninger og maskiner | 5088 | 5309 | 2037 | 4141 |
| Foderindkøb | 17663 | 25333 | 2945 | 9589 |
| I alt | 36740 | 50260 | 17302 | 22218 |
Det vurderes, at afgrødesammensætningen og modelsædskiftet på kvægbedrifterne vil kunne fortsætte uændret efter overgang til mekanisk ukrudtsbekæmpelse. Derimod vurderes, at det vil være vanskelligt at bekæmpe ukrudtet tilfredsstillende med den eksisterende afgrødesammensætning på plante- og svinebrugene (se fx Forskningscenter for Økologisk Jordbrug 2006). Således vil der her ved en overgang til mekanisk ukrudtsbekæmpelse blive behov for at reducere kornafgrødernes andel i sædskiftet og indføre ukrudtssanerende afgrøder såsom frøgræs, grønbrak, plus om muligt afgræsningsmarker eller andre grovfoderafgrøder, som ikke høstes til modenhed. Desuden vil der antageligt være behov for at indføre radrensning (fx af rapsen), samt mini-sommerbrak hvor marken holdes sort efter en tidlig høst som grønafgrøde ultimo maj (Thomsen 2003).
Hvis den mekaniske bekæmpelse af rodukrudt i stedet kan udføres kemisk (fx i forb. med round-up sprøjtninger i efteråret og evt. hormonbehandlinger mod tidsler), og ukrudtsstriglinger indføres isoleret som erstatning for herbicidsprøjtninger først i vækstsæsonen, vil der derimod ikke være de samme behov for sædskiftemæssige tilpasninger.
For at illustrere effekten af en intensiv, mekanisk ukrudtsbekæmpelse på energiforbruget, opgøres effekten på typeafgrøderne i et eksempel på ”worst case” ukrudtsbekæmpelse (Figur 5.15). Det antages her at vårkorn, helsæd og ærter strigles 3 gange, at raps radrenses 3 gange, og at vinterkorn kun strigles 1 gang (Petersen 2006). Desuden antages en ekstra rodukrudt-bekæmpelse i korn, ærter og raps på 3 gange stubharvning/skrælpløjning årligt. For frøgræs antages samme rodukrudt-bekæmpelse hvert 2. år, og for græs i omdrift samt for grønbrak antages samme ukrudtsbekæmpelse hvert 3. år.
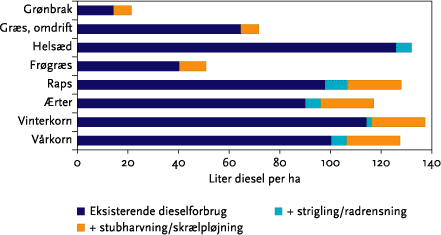
Figur 5.15. Eksempel på beregnet ekstra dieselforbrug til mekanisk ukrudtsbekæmpelse opdelt på strigling/radrensning og stubharvning/skrælpløjning i de 8 typeafgrøder, sammenlignet med det modellerede dieselforbrug på Figur 5.14. Figuren viser et eksempel på ’worst case’ behov for ukrudtsbekæmpelse.
På baggrund af det beregnede dieselforbrug til de opstillede typeafgrøder kan effekter på sædskifteniveau opgøres. For et kvægsædskifte, som indeholder 45 % helsæd, 10 % vårkorn, 10 % vinterkorn og 35 % græs omdrift, vil det gennemsnitlige dieselforbrug således stige med godt 10 liter/ha, svarende til en gennemsnitlig stigning på ca. 10 % (beregnet på baggrund af typeværdierne i Figur 5.15.). Som nævnt kræves der specielt på svine-/planteavlsbedrifter tilrettede sædskifte- afgrødedyrknings strategier, for at ukrudtet kan bekæmpes tilstrækkeligt med mekaniske midler. Hvis det antages at mekanisk ukrudtsbekæmpelse kan indføres i et sædskifte med 35 % vinterkorn, 10 % vårkorn, 10 % ært, 10 % raps, 25 % frøgræs og 10 % grønbrak, vil det samlede dieselforbrug stige med godt 25 % (Figur 5.15), idet det antages at ærtemarken høstes som grønært til helsæd med efterfølgende mini-sommerbrak. Som det ses, udgør dieselforbruget til stubharvning/skrælpløjning en væsentlig del af det ekstra energiforbrug ved overgang til mekanisk ukrudtsbekæmpelse. Isoleret set er det ekstra energiforbrug til ukrudtsstriglingen, som denne rapport fokuserer på, derfor væsentligt mindre. Imidlertid vurderes det, at supplerende stubharvning/skrælpløjning er nødvendig for at bekæmpe rodukrudtet i systemerne med mekanisk ukrudtsbekæmpelse, og det ekstra energiforbrug hertil bør derfor indregnes, hvis ikke der gives mulighed for supplerende kemisk bekæmpelse af rodukrudt.
Overgang til ren mekanisk ukrudtsbekæmpelse vil altså betyde relativt mere for dieselforbruget på svine- og planteavlsbrug end på kvægbrug (Figur 5.16), som således vil kunne overgå til mekanisk ukrudtsbekæmpelse med den mindste energiomkostning. Sammenlignes tilsvarende forøgelsen af det samlede direkte og indirekte energiforbrug ved overgang til mekanisk ukrudtsbekæmpelse (dvs. energiforbruget til både diesel, smøreolie, maskiner mv.) med det samlede energiforbrug på de tre brugstyper opgjort i Tabel 5.8., viser der sig den samme tendens: på kvægbrugene øges det samlede energiforbrug med ca. 1-2 %, på svinebrugene med 2-3 %, og på planteavlsbrugene med 6-7 %. Disse effekter er opgjort for en intensiv mekanisk ukrudtsbekæmpelse, som skønnes nødvendig for på lang sigt og i et ’worst case’ eksempel at styre ukrudtsproblemerne. Forsøg viser imidlertid, at der på kort sigt ikke er nogen signifikant effekt af en intensiveret mekanisk ukrudtsbekæmpelse, og at det enkelte års udbytte generelt ikke påvirkes signifikant af de forskellige former for mekanisk ukrudtsbekæmpelse (Miljøministeriet 1998a, 1998b; Pedersen 2002, Forskningscenter for Økologisk Jordbrug, 2006). Ligeledes vil ukrudtsstrigling i kombination med kemisk rodukrudtsbekæmpelse kunne indføres med færre energiomkostninger end ved ren mekanisk ukrudtsbekæmpelse, især på svine- og planteavlsbrugene.
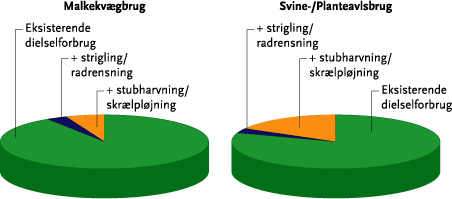
Figur 5.16. Eksempel på beregnet ekstra dieselforbrug ved indførsel af intensiv mekanisk ukrudtsbekæmpelse (opdelt på strigling/radrensning og stubharvning/skrælpløjning) på malkekvægbrug og på svine-/planteavlsbrug, målt i forhold til dieselforbruget inden overgangen til mekanisk ukrudtsbekæmpelse. Figuren viser et ”worst case” eksempel på behov for mekanisk ukrudstbekæmpelse.
5.2.2 Energibalance og dækningsbidrag for markforsøget på Kalø (Scenario E)
Ved opgørelse af energibalance og dækningsbidrag for markforsøgene, er maskin- og arbejdsomkostningerne opgjort hhv. for striglet og ustriglet dyrkning, idet markforsøget jo viste signifikant forskel på kerne og halm-udbyttet mellem disse to dyrkningspraksis (se afsnit 5.1.3). Derimod viste forsøget ikke nogen signifikant forskel mellem de forskellige intensiteter af strigling. Vi har derfor valgt at opgøre omkostninger og energiforbrug for en dyrkningspraksis med intensiv strigling, sammenlignet med ustriglet dyrkning (Tabel 5.9). Energiforbruget er opgjort som det samlede direkte og indirekte energiforbrug, og den fossile energibalance er opgjort som brændværdien i det høstede korn og halm (14,7 MJ/kg korn og 15,0 MJ/kg halm) minus det opgjorte energiforbrug.
Tabel 5.9. Gennemsnitlige maskin- og arbejdsomkostninger, energiforbrug og energibalancer for striglet og ustriglet, økologisk vårhvede på Kalø 2004-2005 Scenario E), opgjort hhv. i kroner og energienheder (MJ) per ha. Den økonomiske opgørelse er beregnet ifølge Gravsholt et al. (2005), og energiforbruget er opgjort ifølge Dalgaard og Dalgaard (2004) ifølge bilag D.
| Maskin- og arbejdsomkostninger | Striglet | Ustriglet | Striglet | Ustriglet |
| (kr/ha) | (kr/ha) | (MJ/ha) | (MJ/ha) | |
| Pløjning (1x) | 475 | 475 | 1435 | 1435 |
| Stubharvning (3x) | 420 | 420 | 1329 | 1329 |
| Spredning af husdyrgødn. (14 t) | 294 | 294 | 868 | 868 |
| Ukrudtsharvning (3x) | 345 | 0 | 365 | 0 |
| Komb. harvning & såning (1x) | 275 | 275 | 753 | 753 |
| Tromling (1x) | 125 | 125 | 133 | 133 |
| Mejetærskning (1x) | 809 | 809 | 798 | 798 |
| Hjemkørsel, korn (2 km) | 127 | 127 | 161 | 170 |
| Tørring (3 pct) | 479 | 479 | 540 | 570 |
| Halmpresning (1x) | 361 | 361 | 437 | 471 |
| Hjemkørsel, halm (2 km) | 92 | 92 | 171 | 185 |
| Maskin- og arb.omkostninger i alt | 3802 | 3457 | 6991 | 6712 |
| %-vis ændring fra striglet til ustriglet praksis | -9% | -4% | ||
| Energiudbytte kerne (brændværdi) | 52920 | 55860 | ||
| Energiudbytte halm (brændværdi) | 57000 | 61500 | ||
| Energibalance (netto overskud af fossil energi) | 102929 | 110648 | ||
Beregningerne viser, at de samlede maskin- og arbejdsomkostningerne var 10 % højere i de striglede end i de ustriglede parceller, mens energiforbruget var 4 % højere i de striglede end i de ustriglede parceller (Tabel 5.9).
Tilsvarende beregnes dækningsbidraget for striglet og ustriglet dyrkningspraksis. Da udbyttet var højest og omkostningerne lavest i de ustriglede forsøgsled, opnås her, som forventet, det bedste økonomiske resultat, svarende til at dækningsbidraget efter maskin- og arbejdsomkostninger her var 14 % (Tabel 5.10).
Andre markforsøg i vårhvede og en lang række andre afgrøder (Pedersen 2002, Forskningscenter for Økologisk Jordbrug, 2006) tyder på, at der ved en optimal strigling kan opnås et uændret eller forøget udbytte i forhold til en praksis, hvor der ikke foretages ukrudtsbekæmpelse ved strigling eller sprøjtning. Derfor gennemregnes også to scenarier for hhv. vellykket (Scenario E1) og meget vellykket (Scenario E2) ukrudtsstrigling. I de to scenarier antages 2 striglinger; hhv. en vellykket blindstrigling efter såning men før ukrudtet er kommet op + en enkelt opfølgende ukrudtsstrigling når ukrudtet har kimblade. Der er grund til at antage, at en sådan striglingsstrategi vil være mere effektiv end de 3 udførte striglinger i forsøget. Således antages i Scenario E1, at der opnås det samme udbytte som uden strigling i forsøget, og i Scenario E2 antages et 5 % højere udbytter end i de ustriglede forsøgsparceller.
Tabel 5.10. Gennemsnitligt dækningsbidrag for striglet og ustriglet, økologisk vårhvede på Kalø 2004-2005 beregnet ifølge Gravsholt et al. (2005) og med en forudsat pris på økologisk vårhvede til brødkorn på 120 kr/hkg (Pedersen 2005). Desuden vises resultatet af to scenarier, hvor ukrudtsstriglingen optimeres med en blindstrigling + en ukrudtsharvning, og hvor udbyttet er hhv. uændret (scenario E1), eller 5 % højere (scenario E2) end i de ustriglede parceller.
| Dækningsbidragskalkule (kr/ha) | Striglet | Ustriglet | Scenario E1 |
Scenario E2 |
| Udbytte | ||||
| Kerne | 4320 | 4560 | 4560 | 4788 |
| Halm | 1520 | 1640 | 1640 | 1722 |
| Hektar-støtte | 2451 | 2451 | 2451 | 2451 |
| Økologi-støtte | 870 | 870 | 870 | 870 |
| I alt | 9161 | 9521 | 9521 | 9831 |
| Stykomkostninger | ||||
| Udsæd | 788 | 788 | 788 | 788 |
| Næringsstofværdi i husdyrgødning | 504 | 504 | 504 | 504 |
| Bindegarn | 12 | 12 | 12 | 12 |
| I alt | 1304 | 1304 | 1304 | 1304 |
| Dækningsbidrag pr. ha | 7857 | 8217 | 8217 | 8527 |
| DB efter maskin- og arbejdsomkostninger | 4055 | 4760 | 4530 | 4840 |
5.3 Fauna modellering på Landskabsniveau
Variationen mellem de samme år i de gentagne modelsimuleringer (10 replikater) var meget små i den sammenlignede tidsperiode for såvel sanglærken (Figur 5.17 A), edderkop (eks. 95 % CL for ’Kvæg-basis’ scenariet: 45, gns.: 22.891; for ’Kvæg-strigling’ scenariet: 73, gns.: 18.556) og løbebille (eks. 95 % CL for ’Kvæg-basis’ scenariet: 75, gns.: 40.195; for ’Kvæg-strigling’ scenariet: 137, gns.: 32530).
Generelt var modelleringerne for alle tre arter meget afhængige af vejrforholdene de enkelte år (for løbebillens vedkommende varierer bestanden mellem 20.000-80.000), hvilket markeres ekstra af, at de anvendte vejrdata er fra en 11- års periode og således optræder cyklisk i alle afbildede modelleringer. Effekterne af striglingerne derimod var kun i mindre grad afhængige af vejrforholdene (eks. Figur 5.17 A og B), og udgjorde i sammenligning med disse kun 20 % i forhold til bestanden i ’Kvæg-basis’ scenariet (eks. løbebille og edderkop i Figur 5.18 A og B).
5.3.1 Kvægscenarier (basis – strigling)
Ingen af de udførte striglinger (hhv. 3 og 2 i vårbyg og vinterhvede) havde nogen negative effekter på de målte variabler for sanglærkens vedkommende. Alle striglinger blev udført på tidspunkter der lå før lærkernes etablering af 1. yngleforsøg.
For edderkoppernes vedkommende var den samlede effekt af striglingerne i gennemsnit 22 % af totalpopulationen i ’Basis-kvæg’ scenariet (Figur 5.18 A). For løbebillernes vedkommende var der en samlet effekt på 19 % (Figur 5.18 B).
5.3.2 Tekniske scenarier i monokulturer af vårbyg og vinterhvede
Sanglærke
For vårbygs vedkommende var der ingen negative effekter af de fire udførte striglinger på den årlige totale bestand (Figur 5.17 B). Dette resultat fremkommer til trods for, at et stort antal af reder (gns.: 1647) før æglægning blev ødelagt ved den 4. strigling (Figur 5.19 A og Tabel 5.11). Effekten af at ’udskyde’ 4. og sidste strigling med 10 dage til d. 20/5 var at reducere den totale årlige bestand i alle år (Figur 5.17 B). Årsagen hertil var en stor andel af reder med æg (gns.: 1484), der blev ødelagt af striglingen. Derimod blev kun få nyanlagte reder ramt, og ingen reder med unger (Figur 5.19 B og Tabel 5.11). De mange ødelagte reder med æg bevirkede desuden en reduktion i klæknings- og ynglesucces og en resulterende reduktion i antallet af unger, der overlevede til migration (Tabel 5.11). Yderligere en udsættelse af 4. strigling til den 30/5 bevirkede, at dette mønster blev forstærket som følge af, at striglingen nu udføres på et tidspunkt, hvor både reder med æg og reder med unger (gns.: hhv. 1157 og 341) bliver ødelagt af striglingen (Figur 5.19 C og Tabel 5.11).
I et landskab, hvor der kun dyrkes vinterhvede, kan bestanden af sanglærker ikke overleve (Figur 5.20). Den primære baggrund for dette er strukturelle forhold knyttet til afgrødens vækst (se endvidere afsnit 6.3.1). Overordnet har tredje strigling d. 10/4 ingen negative effekter på den totale bestandsstørrelse (Figur 5.20 A) eller nogen af de andre udvalgte bestandsvariabler (Tabel 5.12), selvom en del (gns.: 299) reder bliver ødelagt før æglægning (Figur 5.20 B). Ved den senest udførte strigling d. 20/4 påvirkes et tilstrækkeligt stort antal reder under rugefasen til at bevirke en reduktion i det gennemsnitlige antal af flyvefærdige unger med den følge, at den gennemsnitlige totale bestandsstørrelse mindskes kraftigt (Tabel 5.12). Den sene strigling bevirker en hurtigere uddøen af bestanden i modelområdet set i relation til basis scenariet (Figur 5.20 A).
Edderkop
Første strigling i vårbyg har som den eneste strigling en meget ringe effekt på bestandsstørrelsen af ’edderkopper’ (Figur 5.21 A; Tabel 5.13). Den isolerede effekt af 2., 3. og 4 strigling er af samme størrelsesorden på ca. 20 %. Allerede efter 2. strigling er den kumulerede effekt tæt på at halvere den gennemsnitlige bestandsstørrelse, og fra og med 3. strigling er bestanden mere end decimeret i forhold til basis (Figur 5.21 B; Tabel 5.13).
I vinterhvede er effekten af 1. strigling (d. 1/10) anderledes kraftig med en halvering af bestanden til følge (Figur 5.21 C; Tabel 5.13). Den
isolerede effekt af de tre efterfølgende striglinger er som for vårbyggens vedkommende af samme størrelsesorden, blot ca. 10 % større (Tabel 5.13). I vinterhveden medfører de kumulerede effekter en decimering af bestanden allerede efter 2. strigling, og efter 3. strigling uddør bestanden (Figur 5.21 D; Tabel 5.13). I vårbyg er bestanden på et eksistensminimum efter 3. strigling (Figur 5.21 C; Tabel 5.13), og efter 4. strigling uddør bestanden helt.
Løbebille
For løbebillens vedkommende er de isolerede effekter af strigling større end for edderkoppens vedkommende og størst efter 1. strigling både i vårbyg og i vinterhvede (størst efter efterårsstriglingen i vinterhvede; Figur 5.22 A og C; Tabel 5.14). De isolerede effekter af de efterfølgende tre striglinger er mindre end efter 1. strigling men stadig mellem 10 – 20 % større end for edderkoppens vedkommende (Figur 5.22 A og C; Tabel 5.14).
De kumulerede effekter for løbebillerne af de efterfølgende striglinger er sammenlignet med edderkopperne tilsvarende større. Allerede efter 2. strigling er bestanden i vinterhvede reduceret til et niveau, hvor bestanden overlever på et absolut minimum (Figur 5.22 D; Tabel 5.14). I vårbyg er den negative effekt også kraftig efter 2. strigling. I forhold til edderkopper er påvirkningen efter 2. strigling dobbelt så stor for løbebillernes vedkommende (Figur 5.22 D; Tabel 5.14). Efter 3. strigling er også bestanden i vårbyg reduceret meget kraftigt.
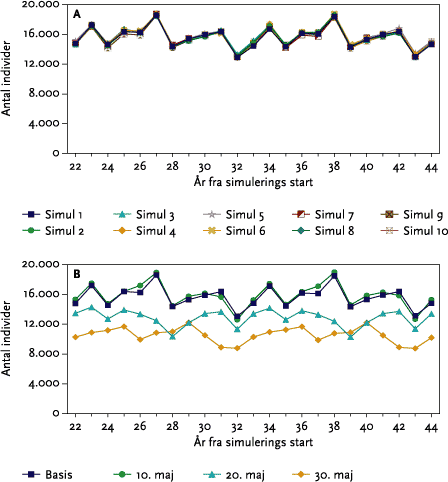
Figur 5.17. A: Den årlige totale bestand (gns. af 10 replikater) af sanglærker i ti modelsimuleringer (Simul 1-10) udført i vårbyg monokultur uden herbicidanvendelse og uden strigling = ’Basis’. B: Den årlige totale bestand af sanglærke i tekniske scenarier med vårbyg monokultur (fjerde og sidste strigling (d. 10/5) er forsøgsmæssigt udsat til hhv. d. 20/5 og d. 30/5). Basis = udgangsscenariet, hvor vårbyg er dyrket under ’standard’ konventionelle forhold som i A (se i øvrigt afsnit 3.4 for yderligere forklaring).
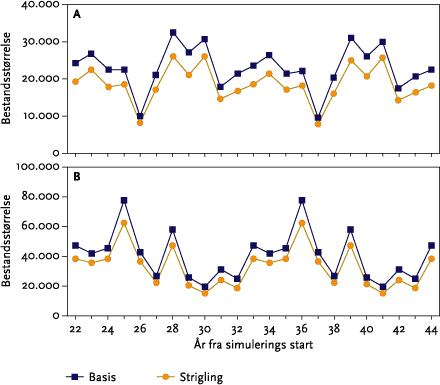
Figur 5.18. Den årlige totale bestand af hhv. edderkopper (A) (tæppespinderen Erigone sp.) og (B) løbebiller (Bembidion lampros) i kvægscenariet. Basis = konventionel dyrkningspraksis. Strigling = konventionel dyrkningspraksis men med benyttelse af strigling i stedet for kemisk ukrudtsbekæmpelse.
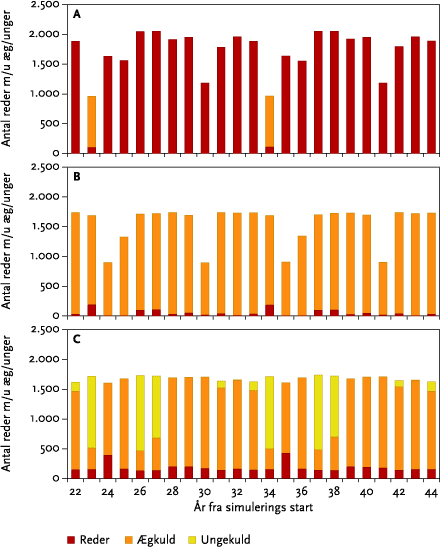
Figur 5.19. Det samlede årlige tab af lærkereder i vårbyg monokultur som følge af en enkelt strigling udført hhv. d. 10/5 (A), d. 20/5 (B) og d. 30/5 (C). ’Reder’ = reder før æglægning, ’Ægkuld’ = reder med æg, og ’Ungekuld’ = reder med unger (se i øvrigt Tabel 5.10 for yderligere forklaring).
Tabel 5.11. Modelresultater for sanglærke fra tekniske scenarier i monokultur af vårbyg (fjerde og sidste strigling (d. 10/5) er forsøgsmæssigt udsat til d. 20/5 og d. 30/5). Basis = som i Figur 5.20. Total-Bestand og Total-juvenil = hhv. det gennemsnitlige årlige antal (modelleret over 33år) individer og ’årsunger’ i alt der overlever til migration. Klæk. Success, Udflyv. Success og Ynglesuccess = hhv. den gennemsnitlige årlige klækningssuccess, udflyvningssuccess og ynglesuccess (for definition af disse se afsnit 3.4). ’Redemort.’, ’Ægmort.’ og ’ungemort.’ angiver den årlige gennemsnitlige dødelighed for hhv. ’reder’ (før æglægning), reder med æg (fra æglægning til klækning) og reder med unger (fra klækning til ’udflyvning’).
| Total- Bestand |
Total - Juvenil |
Klæk. Success |
Udflyv. Success |
Yngle- Success |
Rede- mort. |
Æg mort. |
Unge mort. |
|
| Basis | 15666 | 5934 | 0,74 | 0,58 | 0,43 | 0 | 0 | 0 |
| 10. maj | 15910 | 6020 | 0,75 | 0,59 | 0,44 | 1647 | 78 | 0 |
| 20. maj | 12789 | 4849 | 0,55 | 0,62 | 0,35 | 52 | 1484 | 0 |
| 30. maj | 10554 | 3998 | 0,52 | 0,54 | 0,26 | 184 | 1157 | 341 |
Tabel 5.12. Modelresultater for sanglærke fra tekniske scenarier i monokultur af vinterhvede (kun resultater fra ’Basis’ og 3. og 4. strigling hhv. d10/4 og 20/4 er vist). Basis = som i Figur 6.22. Det gennemsnitlige årlige antal (modelleret over 33år) individer og ’årsunger’ i alt (hhv. Total-Bestand og Total-juvenil) der overlever til migration. Klæk. Success, Udflyv. Success og Ynglesuccess = hhv. den gennemsnitlige årlige klækningssuccess, udflyvningssuccess og ynglesuccess (for definition af disse se afsnit 3.4). ’Redemort.’, ’Ægmort.’ og ’ungemort.’ angiver den årlige gennemsnitlige dødelighed for hhv. ’reder’ (før æglægning), reder med æg (fra æglægning til klækning) og reder med unger (fra klækning til ’udflyvning’).
| Total- Bestand |
Total - Juvenil |
Klæk. Success |
Udflyv. Success |
Yngle- Success |
Rede- mort. |
Æg mort. |
Unge mort. |
|
| Basis | 2565 | 857 | 0,54 | 0,32 | 0,18 | 0 | 0 | 0 |
| 10. april | 2560 | 862 | 0,54 | 0,33 | 0,18 | 299 | 0 | 0 |
| 20. april | 1028 | 240 | 0,44 | 0,27 | 0,12 | 55 | 229 | 0 |
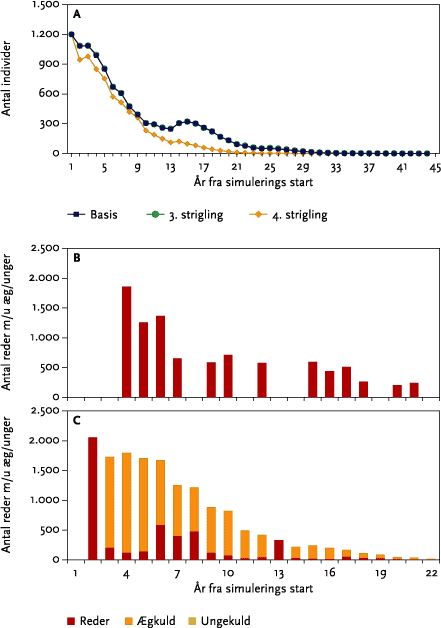
Figur 5.20. A: Den årlige totale bestand af sanglærke i tekniske scenarier med vinterhvede. Basis = vinterhvede monokultur uden herbicidanvendelse og uden strigling. Det samlede årlige tab af lærkereder i vinterhvede monokultur som følge af 3. (B) og 4. (C) strigling udført hhv. d. 10/4 og (C) d. 20/4. ’Reder’ = reder før æglægning, ’Ægkuld’ = reder med æg, og ’Ungekuld’ = reder med unger (se i øvrigt Tabel 5.11 for yderligere forklaring).
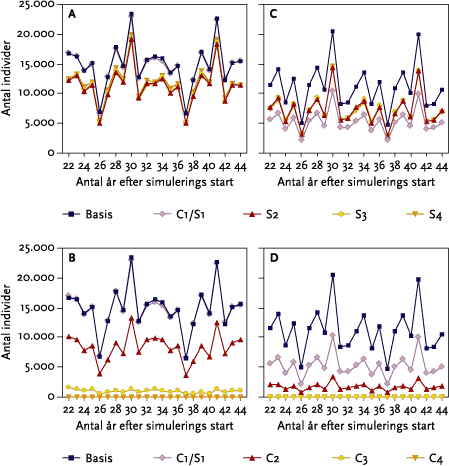
Figur 5.21. Den årlige totale bestand af edderkopper (tæppespinderen Erigone sp.) i tekniske scenarier med hhv. vårbyg (A og B) og vinterhvede (C og D). Effekten på total bestanden af kun én strigling udført på forskellige tidspunkter (S1-S4) i (A) vårbyg og (C) vinterhvede. C1-C4: den kumulerede effekt på total bestanden af én til fire striglinger i vårbyg (B) og vinterhvede (D).Basis = som i Figur 5.20.
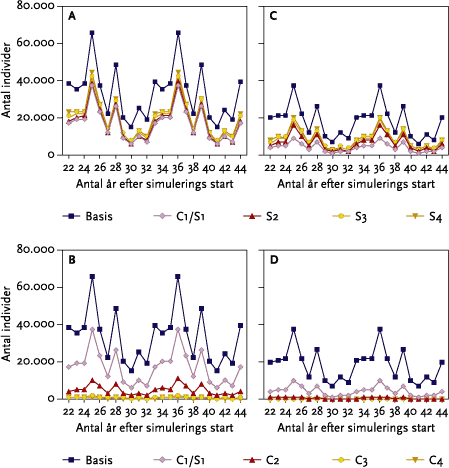
Figur 5.22. Den årlige totale bestand af løbebiller (Bembidion lampros) i tekniske scenarier med hhv. vårbyg (A og B) og vinterhvede (C og D). Effekten på total bestanden af kun én strigling udført på forskellige tidspunkter (S1-S4) i (A) vårbyg og (C) vinterhvede. C1-C4: den kumulerede effekt på total bestanden af én til fire striglinger i vårbyg (B) og vinterhvede (D). Basis = som i Figur 5.20.
Tabel 5.13. Gennemsnitlige effekter over 33 år (relative i forhold til basis scenario) af enkelt striglinger udført på fire forskellige tidspunkter (S1-S4) og kumulerede effekter af striglinger (C1-C4) på edderkop i hhv. vårbyg (VB) og vinterhvede (VH). Bestanden i VB-C4 og VH-C3,C4 uddør i alle modelsimuleringer.
| Edderkop | S1 | S2 | S3 | S4 | |
| Single | VB | 0,01 | 0,23 | 0,23 | 0,20 |
| VH | 0,53 | 0,35 | 0,35 | 0,36 | |
| C1(S1) | C2 | C3 | C4 | ||
| Kumuleret | VB | 0,01 | 0,44 | 0,93 | 0,99 |
| VH | 0,53 | 0,85 | 0,99 | 0,99 | |
Tabel 5.14. Gennemsnitlige effekter over 33 år (relative i forhold til basis scenario) af enkelt striglinger udført på fire forskellige tidspunkter (S1-S4) og kumulerede effekter af striglinger (C1-C4) på løbebille i hhv. vårbyg (VB) og vinterhvede (VH). Bestanden i VB-C4 og VH-C3,C4 uddør i alle modelsimuleringer.
| Løbebille | S1 | S2 | S3 | S4 | |
| Single | VB | 0,51 | 0,49 | 0,44 | 0,40 |
| VH | 0,77 | 0,63 | 0,58 | 0,53 | |
| C1(S1) | C2 | C3 | C4 | ||
| Kumuleret | VB | 0,51 | 0,86 | 0,97 | 0,99 |
| VH | 0,77 | 0,97 | 0,99 | 0,99 | |
6 Diskussion
- 6.1 Effekter på ukrudt og høstudbytte
- 6.2 Effekter på energiforbrug
- 6.3 effekter på fauna
- 6.4 Samlet vurdering
6.1 Effekter på ukrudt og høstudbytte
Ukrudtet udviklede sig så sparsomt i 2004, at det næppe har haft betydning for høstudbyttet. Det eneste tilfælde, hvor ukrudtet med sikkerhed har påvirket udbyttet, var på Krovang i 2005, hvor der var meget ukrudt i såvel de striglede som de ustriglede markhalvdele.
Ukrudtsstriglingerne havde ringe effekt på ukrudtet med en gennemsnitlig bekæmpelseseffekt på 33 %. Bemærkelsesværdigt nok var denne bekæmpelseseffekt upåvirket af år og mark, selvom der var store forskelle i den absolutte mængde ukrudt. Især i 2005 på Krovang, var ukrudtsfloraen påfaldende, idet agersennep totalt dominerede. Bekæmpelseseffekten var lav i forhold til, hvad der sædvanligvis opnås i vårsæd, hvor bekæmpelseseffekterne ofte ligger mellem 50-90 % (Rasmussen & Rasmussen 1994, 1995; Petersen 2000). Kun den tidligst udførte strigling havde effekt på ukrudt og udbytte, hvilket var forventeligt i forhold til agersennep, da denne aggressive og hurtigt voksende ukrudtsart kun kan bekæmpes med strigling i afgrødens tidligste vækststadier (Rasmussen & Rasmussen 1994). Det var imidlertid overraskende, at de øvrige arter ikke reagerede på de sene striglinger, da tidligere forsøg har vist, at disse striglinger godt kan forstærke effekten af de tidlige striglinger (Rasmussen & Rasmussen 1994, 1995). Årsagen kan være, at de senere striglinger ikke har været tilstrækkeligt kraftige, da det kræver ganske kraftig behandling at bekæmpe ukrudt i de sene vækststadier. Jo senere der behandles, jo kraftigere skal der strigles for at opnå en tilfredsstillende effekt, og på stiv lerjord, som der var tale om i forsøgene, kan det være svært at gennemføre en tilstrækkelig kraftig behandling.
På Landbohøjskolens økologiske forsøgsgård, Bakkegården, blev der i 2005 til sammenligning gennemført forsøg med 1, 2 og 3 striglinger i vårhvede, vårbyg, havre og vårtriticale (Gundersen 2006), og der blev opnået bekæmpelseseffekter på 53 %, 72 % og 77 %. Det stigende antal striglinger blev dog foretaget tidligere i kornets udvikling end på Kalø, og jorden var betydelig lettere at bearbejde. Selvom vårhvede havde lavere konkurrenceevne end både havre og vårbyg, var bekæmpelseseffekten udtrykt i procent uafhængig af kornart.
Der foreligger ikke nogen enkel forklaring på, at udbyttet blev reduceret ved strigling, men det må skyldes mekaniske skader på afgrøderne som følge af strigling. Det var ikke muligt at erkende de fysiske skader ved visuelle bedømmelser efter endt behandling. Normalt vil man kunne se skader efter behandling, der som her resulterer i 6 % udbyttetab, men dette var ikke været tilfældet i de pågældende forsøg. I markforsøgene blev der konstateret klare positive sammenhænge mellem afgrødens og ukrudtets vækst. Dette forhold kan have haft betydning i blokforsøgene, men er næppe hele forklaringen, da det så skulle være tilfældigt, at netop de ubehandlede parceller i begge år og på begge marker netop skulle have repræsenteret en generelt større jordfrugtbarhed end de øvrige parceller.
Resultaterne i dette projekt viser, at strigling kan have åbenlyse begrænsninger i form af lav bekæmpelseseffekt og negativ indflydelse på udbyttet. Dette betyder dog ikke, at strigling generelt kan afskrives som ineffektiv. I de tilfælde, hvor det er muligt at opnå mindst 70 % ukrudtsbekæmpelse uden skader på afgrøden, udgør strigling et økonomisk alternativ til herbiciderne. At sådanne effekter kan opnås, er tidligere dokumenteret (Rasmussen & Rasmussen 1995), og det er påvist, hvilke faktorer der skal være opfyldt for at opnå de positive resultater (Rasmussen 1991b). Her spiller selektivitetsforholdene en meget afgørende betydning (Rasmussen 1991a, 1991b), som blandt andet er betinget af behandlingens timing og ukrudtets sammensætning. En altafgørende forudsætning for at opnå et tilfredsstillende resultat er imidlertid, at jorden kan bearbejdes, så den ”falder godt for redskabet”. Denne forudsætning har i ringe grad været opfyldt i forsøgene på Kalø på grund af en stiv, knoldet og ubekvemme lerjord. I økologisk jordbrug og i visse afgrøder såsom markærter er ukrudtsstrigling et alternativ til pesticider, og hvis anvendelsen af pesticider begrænses eller forbydes i områder med særlige vand- eller naturinteresser, kan ukrudtsstrigling også her være et alternativ til pesticidanvendelse.
6.2 Effekter på energiforbrug
Opgørelserne i afsnit 5.2 viser, at mekanisk ukrudtsbekæmpelse kan udføres uden betydelige påvirkninger af energiforbrug og dækningsbidrag.
I forsøget med økologisk vårhvede forøgedes energiforbruget med 3-4 % ved intensiv strigling, og energibalancen blev reduceret fra 111 til 103 GJ/ha. Det vurderes, at det observerede fald i udbyttet ved striglingerne på Kalø vil kunne afværges ved en mere optimal ukrudtsstriglings-strategi, hvor der foretages èn blindstrigling og kun èn opfølgende ukrudtsstrigling. Derved vil dækningsbidraget ikke påvirkes nævneværdigt eller i hvert fald væsentligt mindre end de op til -15 %, der blev resultatet af markforsøget.
De energimæssige og økonomiske omkostningerne ved mekanisk ukrudtsbekæmpelse bør sammenlignes med de øvrige fordele og ulemper en pesticidfri dyrkning kan have. I denne rapport sammenlignes opgørelserne af energiforbrug og dækningsbidrag ved ukrudtsstrigling, således at disse effekter kan sammenholdes med udvalgte effekter på flora og fauna. Andre fordele ved ukrudtsstriglingen frem for anvendelse af pesticider vil fx være beskyttelsen af vandressourcerne mod pesticidforurening, hvilket bl.a. undersøges i et nyt pesticidforskningsprojekt om ”EU's landbrugsordninger og pesticidpolitikken” (2006-2008) (Schou, 2006). Pesticidfri dyrkning kan i den sammenhæng måske udløse forskellige støtteordninger i lighed med den økologistøtte og merpris for brødkornet, som opnås ved den økologiske produktion på Kalø (Tabel 5.10).
6.3 effekter på fauna
6.3.1 Sanglærke
En relativ sen fremspiring samt lav dækningsgrad (vårhvedens ’buskningstadie’ fremkom senere og i tydeligt mindre grad end eks. vårbyggen) bevirkede en langsom territorieopbygning på forsøgsmarkerne i begge år. I 2004 startede territoriebesættelsen 14 dage senere på Krovang marken, der igennem hele vækstsæsonen var præget af en mindre tæt afgrøde, hvilket tydeligt afspejledes i høstudbyttet på de to marker i 2004 (Tabel 5.7).
På trods af en sen territoriebesættelse på arealerne bevirkede vårhvedens åbne struktur under vækstforløbet, at tætheden af ynglepar (specielt i 2004) var høj sammenlignet med ’bestandstætheder’ i konventionelt dyrkede afgrøder under gode vækstforhold (Glutz & Bauer 1985). Den lysåbne afgrødevækst bevirkede tillige, at ynglesæsonen i begge forsøgsår blev lang, og at flere ynglepar havde to eller tre yngleforsøg. En medvirkende årsag til dette forhold var en relativ stor prædationsrate på rederne i primært første yngleforsøg, hvorved en stor del reder blev omlagt. Det samlede tab af reder (64 %) var størst i 2004, og det samlede tab af reder var størst i redeunge-stadiet. Tidligere undersøgelser (Odderskær et al. 1997) har vist en klar sammenhæng mellem et køligt og regnfuldt vejrlig og en høj dødelighed på sanglærkens æg og redeunger. Specielt i 2004 lå nedbørsmængden og antallet af regnvejrsdage i juni måned over gennemsnittet for denne måned. Det høje antal mislykkede reder på begge marker i 2004 (andel succesfulde reder i 2004: 0,2-0,5; i 2005: 0,5-0,9) var primært forårsaget af prædation (37 %), men også en del forladte reder og døde, underernærede unger tages som udtryk for, at de dårlige vejrforhold i en stor del af yngleperioden påvirkede reproduktionen negativt.
Antallet af ’naturlige’ reder, der direkte blev udsat for strigling var meget begrænset i begge år. Tilsvarende tages det lave antal ’naturlige’ reder (3), der blev ødelagt/opgivet som følge af striglingerne, som udtryk for, at den direkte effekt på det samlede yngleresultat på de striglede markhalvdele var ubetydelig. For striglinger udført inden for det samme tidsrum som denne undersøgelse, blev der opnået lignende resultater i en anden undersøgelse (Esbjerg et al. 2006), der bl.a. registrerede lærkens reproduktionsforhold i vårhvede. Til forskel fra nærværende projekt blev der i Esbjerg et al. (2006) også udført meget sene striglinger. Ved de sent udførte striglinger (ultimo maj – primo juni) var der, ud over kraftige effekter på andelen og antallet af påvirkede reder (se nedenfor), tillige indikationer for, at reder på de sent striglede marker efterfølgende var mere eksponerede for prædation end reder på marker, hvor der kun var foretaget tidlige striglinger. Ovenstående resultater tyder på, at sene striglinger – i hvert fald indenfor en periode – kan blotlægge reder, så de lettere opdages af primært kragefugle. En opgørelse over antallet af præderede og opgivne reder i nærværende undersøgelse viser ingen forskel i antallet af præderede eller opgivne reder i relation til behandlingstype, måske fordi alle striglinger blev udført inden lærkernes reproduktion var kommet rigtigt i gang.
Vurderet ud fra antallet af æg/redeunger, udfløjne unger pr. rede/territorium og den observerede ynglesucces (= min. én udfløjen unge/rede), var der ingen indikationer for, at de udførte striglinger skulle have haft nogen negativ indirekte indflydelse på fuglenes reproduktion eller nogen ’carry-over’- effekt mht. behandling det efterfølgende år. Heller ikke redeungernes vægt på 5., 7. eller 8. dagen indikerede nogen indirekte effekter relateret til behandling.
Data for begge forsøgsår viser imidlertid en tendens til et større antal succesfulde reder på Krovang marken. Denne tendens er mest udtalt i 2005, hvor antallet af mislykkede reder på begge marker generelt var mindre end året før. Årsagen til dette er usikker, men en medvirkende årsag kan have været et generelt lavere prædationstryk på denne mark.
Mens de direkte og indirekte effekter af mekaniske ukrudtsstriglinger på lærkebestandens ynglesucces i stor grad afhænger af tidspunktet for behandlingernes udførelse (se nedenfor), synes det derimod sikkert, at der ved striglinger med den her anvendte type af striglingsudstyr kan forventes effekter på den enkelte rede i en størrelsesorden på mellem 50 og 100 %. I størstedelen af tilfældene blev rederne enten dækket af jord/sten eller fysisk revet op af jorden. Den samlede gennemsnitlige redemortalitet på 72,4 % for kunstige reder ligger mellem de mortalitetsværdier, der er fundet for naturlige reder i to andre danske undersøgelser (Odderskær 2002, Esbjerg et al. 2006). Ud over nævnte undersøgelser, har ingen andre videnskabelige undersøgelser beskæftiget sig med samme problematik. Bedømt ud fra forsøgene med kunstige reder og æg var sandsynligheden for, at en rede overlevede to på hinanden følgende striglinger meget lille, da mortaliteten for den enkelte rede var af samme størrelsesorden ved begge striglinger.
Striglingens effekter afhænger til en vis grad af jordtype og vejrets påvirkning af disse. Ved den tredje og sidste strigling blev der fundet en signifikant forskel i redemortaliteten i relation til jordtype (størst tab v. stenet/leret jordtype, mindst v. sandet/løs jordtype), men om der kan uddrages noget konkret ud fra dette er tvivlsomt, da forsøg på at udtrykke jordstrukturernes ’fasthed’ var præget af meget stor variation indenfor de enkelte jordtyper.
Samlet set viser resultaterne vedrørende de direkte effekter på den enkelte rede altså en markant risiko for at reden går tabt, mens undersøgelser med ’naturlige’ reder viser, at der under normale dyrkningsforhold ikke er stor risiko for, at striglinger påvirker lærkernes reproduktion i vårhvede, fordi langt de fleste reder først anlægges efter at striglingerne er foretaget. Undersøgelsens modeldel underbygger, at dette under normal dyrkningspraksis heller ikke vil være tilfældet i en vårbyg eller vinterhvede, hvor striglinger erstatter den kemiske ukrudtsbekæmpelse. Striglingspraksis, som det er anvendt i modelscenarierne, afspejler en normal økologisk dyrkningspraksis for vårbyg og vinterhvede (Petersen 2006). I ’kvægscenariet’, hvor alle bedrifttyper i modellandskabet er kvægbrug med det samme sædskifte, når alle striglinger at blive udført før lærkernes ynglestart, og de tekniske scenarier viser, at der i monokultur af vårbyg først ses en markant reduktion i populationen ved striglinger, der udføres fra og med ca. d. 20. maj. I vinterhvede gælder dette for striglinger fra og med ca. d. 20. april. I vejrmæssigt normale år vil forårsstriglingen være overstået på dette tidspunkt (omkring midten af april). Scenariet med vinterhvede viser samtidigt, at populationen ikke kan overleve i et landskab, hvor der kun dyrkes vinterhvede, hvilket også er indikeret ud fra engelske undersøgelser (Wilson et al. 1997). Dette er i overensstemmelse med den viden, der foreligger vedr. lærkens reproduktionsforhold i tætte hurtigtvoksende vinterafgrøder (Schläpfer 1988, Jenny 1990, Daunicht 1998, Chamberlain et al. 1999). Det skal understreges, at størrelsen af de fundne effekter i forbindelse med sene striglinger vil afhænge af størrelsen af de striglede arealer, samt antal af og tidspunkt for de striglinger, der udføres.
’Timingen’ for den eller de sidste udførte striglinger er altså central for, i hvilken udstrækning rederne vil blive ’ramt’, og er som nævnt ovenfor afgrødespecifik og afhænger sekundært af flere andre faktorer herunder geografisk beliggenhed, jordbund, gødskning, lokale vejrforhold mm.
De udførte scenarier viser tillige, at der ikke har været nogen ’gavnlig’ effekt at spore ved at benytte mekanisk frem for kemisk ukrudtsbekæmpelse. En af de medvirkende årsager til dette er sandsynligvis, at den øvrige dyrkningspraksis (herunder anvendelsen af insekticider) forbliver uændret i forhold til det konventionelle udgangspunkt.
Resultaterne kan for lærkens vedkommende samles i følgende:
- i forbindelse med mekanisk ukrudtsbekæmpelse anbefales det:
- ikke at udføre striglinger i vår-kornafgrøder senere end ca. 30 dage efter såning
- ikke at udføre striglinger i vinter-kornafgrøder senere end ca. midt april (hvis vejrforholdene tillader det)
For sanglærken ville det i nogle områder, hvor der udføres flere sene striglinger kunne være et problem lokalt. Dette vil med den nuværende dyrkningspraksis kun være aktuelt på økologisk dyrkede bedrifter, da der ikke forekommer nogen nævneværdig ukrudtsstrigling i konventionelt dyrkede kornafgrøder.
Effekterne for lærken er ikke så kritiske som for viben, som på grund af en tidligere start på ynglesæsonen (starter æglægningen umiddelbart efter såning) og en længere rugetid (ca. 4 uger) risikerer at blive ramt af blindstriglinger såvel som efterfølgende striglinger, med en stor redemortalitet til følge (upublicerede data fra denne undersøgelse samt Esbjerg et al. (2006)). Data fra indeværende projekt vil indgå i sidstnævnte projekt i den endelige afrapportering for dette projekt. For vibens vedkommende er der yderligere det problem, at ingen eller kun meget få omlæggere ender med et succesfuldt yngleforløb, bl.a. fordi sent klækkede kyllinger har en dårlig overlevelse (Galbraith 1988).
6.3.2 Løbebille og tæppespinder
Bedømt ud fra resultaterne fra ’Kvægscenariet’ viser effekterne af vejrliget på begge de modellerede arter, at det – hvor det er muligt - er vigtigt at inkludere vejrmæssige betingelser i effektstudier på overfladelevende leddyr.
Bedømt ud fra de tekniske scenarier vil den samlede effekt af tre eller fire striglinger medføre alvorlige negative påvirkninger af bestanden af disse leddyr. Der er en række årsager til dette, og de skal alle findes i denne undersøgelses eksperimentelle set-up: scenarierne er ’worst-case’ scenarier og opnåede effekter er maksimale, idet alle dyrkningstiltag herunder striglingerne udføres synkront på samme datoer i kun én afgrøde. De tekniske scenarier er således designet til at både at kunne belyse effekterne af de enkelte (rene) striglinger og de kumulerede.
Striglingens effekter i et landskab, hvor der kun var ét sædskifte (kvægbedrift) var måske mindre markante end resultaterne fra tekniske scenarier kunne indikere. Selvom de påviste effekter i de tekniske scenarier altså er store allerede efter anden strigling, kommer de ikke til udtryk med samme styrke i kvægscenariet, fordi der er en sammensætning af forskellige afgrøder, hvor det for det første ikke er alle afgrøderne der strigles, og for det andet strigles vårbyggen og vinterhveden kun hhv. tre og to gange mod fire striglinger i de tekniske scenarier. Derudover er det ikke alle bygmarker, der strigles anden og tredje gang, og de enkelte striglinger udføres ikke synkront.
Selvom effekterne fra de enkelte striglinger altså er kraftigt negative for de udvalgte overfladelevende leddyrarter, bliver den samlede ’reelle’ effekt i det undersøgte – og forenklede – dyrkningslandskab betydeligt mindre, når de enkelte påvirkninger på denne måde ’spredes’ ud i tid og rum. De samlede effekter afhænger således af de enkelte former for dyrkningstyper og -praksis i et givet område (areal med, antal af og den geografiske spredning af bedrifts- og afgrødetyper), samt andelen af ikke dyrkede habitater i området. Det sidste er i denne undersøgelse specielt vigtigt for den modellerede løbebille, som overvintrer i og spredes fra netop sådanne seminaturlige ’kanthabitater’.
Resultaterne for edderkoppens og løbebillens vedkommende samles i følgende:
- i forbindelse med mekanisk ukrudtsbekæmpelse anbefales det mht. edderkoppen Erigone atra:
- ikke at udføre mere end to striglinger i vår-kornafgrøder, gerne udført så tidligt som muligt (blindstrigling + én efterfølgende strigling)
- Ikke at udføre mere end én strigling i vinter-kornafgrøder, og denne strigling har mindst skadelige effekt udført tidlig i foråret
- i forbindelse med mekanisk ukrudtsbekæmpelse anbefales det mht. løbebillen Bembidion lampros:
- ikke at udføre mere end én strigling i såvel vår- og vinter-kornafgrøder, og denne strigling har mindst skadelige effekt udført i foråret
6.3.3 Regnorme og mikroleddyr
Det er vanskeligt på baggrund af nærværende projekt at konkludere, om markstrigling har negative effekter på springhaler og mider i marken. Antal springhaler, mider og springhalearter responderer ikke entydigt på hverken antallet af striglinger eller strigling sammenlignet med fravær af strigling. For de fleste målinger var der således ingen signifikante effekter, og de effekter, der var, var ikke entydigt negative eller positive. I nogle, men ikke alle tilfælde var der store variationer mellem responset på strigling i de fire blokke, hvilket kan skyldes dels forskelle i forholdene inden for og mellem blokkene inden striglingerne, dels at striglingerne er faldet forskelligt ud i de forskellige blokke. Da forekomsten af jordlevende leddyr ikke generelt er korreleret med ukrudtbiomassen, kan forskellene i striglingseffekter mellem blokkene ikke forklares ad den vej.
For regnormenes vedkommende er billedet mere tydeligt, idet der konsekvent ingen signifikante effekter var af striglingerne hverken i 2004, da foråret var temmelig tørt, eller i 2005 da foråret var mere normalt. Vi kan derfor tillade os at konkludere, at striglinger foretaget som i dette projekt ikke er skadelige for regnormefaunaen på vårhvedemarker på lerjord. Sandsynligvis vil det også gælde for i hvert fald lignende afgrøder, mens det er mere usikkert, om forholdene vil være anderledes på lettere og mere sandede jordtyper.
6.4 Samlet vurdering
Nærværende undersøgelse har haft til formål at estimere effekterne af ukrudtsstriglinger på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug gennem inddragelse af flere trofiske niveauer og funktionelle grupper. De enkelte dyre- og plantearter har deres egen værdi i sig selv, men de udvalgte organismer i denne undersøgelse har også en funktion eller værdi, som kan have betydning af økonomisk og økologisk art.
Regnorme, springhaler og mider har en væsentlig indflydelse på omsætningen i jorden og dens frugtbarhed, og er desuden vigtige (springhalerne og miderne) fødeemner for den udvalgte løbebille og edderkop. Ud over selv at udgøre vigtige fødeemner for diverse fuglearter (Potts 1986, Odderskær et al. 1997), udgør disse som polyphage prædatorer et vigtigt økonomisk element gennem deres evne til at reducere mængden af skadevoldende insekter (primært bladlus) i landbrugsafgrøderne (Sunderland 1975, Edwards et al. 1979, Chiverton 1986, Östman et al 2003). Via deres tilstedeværelse i marken – eller mangel på samme - er de således indirekte med til også at påvirke behovet for anvendelse af insekticider, og har derigennem også potentielt en indirekte miljømæssig betydning.
Det er ikke muligt umiddelbart at overføre de fundne effekter på leddyr til andre af markfladens leddyr, men i kraft af de valgte arters forskellige levevis vil de fundne effekter med forsigtighed kunne overføres på andre arter af leddyr, hvilket tillige er indikeret ud fra andre undersøgelser (Navntoft & Esbjerg 2003; Bilde & Topping 2004).
Den lave og utilfredsstillende bekæmpelse af ukrudtet må antages at være en konsekvens af den svære og ubekvemme lerjord på Kalø, samt at de to sidste striglinger har været virkningsløse på grund at det sene tidspunkt, de blev udført på, hvilket gjorde det umuligt at bearbejde jorden tilstrækkeligt kraftigt til at opnå en signifikant effekt på ukrudtet. Antallet af og tidspunkter for ukrudtsstriglinger vil variere i relation til bl.a. afgrøde og ejendom, men under normale forhold bliver der på de tunge jorder på Kalø kun foretaget blindstrigling og én efterfølgende strigling efter at kornet er kommet op. Denne strategi er netop udviklet under hensyntagen til den tunge lerjord, som kendetegner forsøgsmarkerne. Hvis striglingerne havde været gennemført på en anden jordtype, hvor det havde været muligt at påvirke såvel ukrudt som afgrøde kraftigere, ville det muligvis også have medført større effekter på jordbundens leddyr og regnorme, men det kan nærværende forsøg ikke afgøre. Benyttelsen af modelsimuleringerne i modelsystemet ALMaSS har givet en mulighed for at udnytte de eksperimentelt baserede oplysninger fra markniveau til at give en vurdering af striglingens effekter på faunaen på landskabsniveau, hvorved det har været muligt også at inddrage variationer i tid og rum, og inkludere den variable påvirkning der altid vil påvirke levende organismer i samspil med de specifikke variabler, der som udgangspunkt er genstand for en undersøgelse. Derudover har modelsystemet givet mulighed for gennem specielle scenarier at kunne vurdere både isolerede og samlede effekter af ukrudtsstriglinger gennem modelsimuleringer udført i et heterogent dyrkningslandskab under dyrkningsmæssigt simplificerede forhold (kun én bedrifttype, samt rene monokulturlandskaber af vårbyg og vinterhvede).
Ud fra den erfaring der er indhentet fra brugen af ALMaSS-modelsystemet i andre sammenhænge (Jepsen et al. 2005; Thorbek & Topping 2003; Topping & Odderskær 2004), samt den specifikke validering, der er foretaget for de anvendte modelarter (eks. sanglærke se Bilag C) er det vurderet, at de fundne resultater giver pålidelige estimeringer af striglingens effekter på den udvalgte fauna. Når modelresultater og eksperimentelle resultater fra denne undersøgelse og ovennævnte undersøgelser fra Esbjerg et al. (2006) sammenholdes mht. den tidsperiode eller ’tidsrude’, hvori striglingerne vil have ingen eller kun meget begrænsede effekter, ses en stor grad af overensstemmelse. Denne overensstemmelse mellem eksperimentelle og modelrelaterede resultater tages som udtryk for, at de foretagne modelsimuleringer udtrykker en høj grad af realisme.
Der er ud fra resultaterne i denne undersøgelse flere ting der peger på, at det godt kan ’lade sig gøre’ - uden at påvirke den vilde fauna repræsenteret ved de i denne undersøgelse udvalgte arter/artsgrupper - at reducere herbicidforbruget ved anvendelse af mekanisk behandling. I praksis skal der dog tages hensyn til visse forudsætninger/begrænsninger. Dyrkningsmæssigt er den økonomiske omkostning ved en ’konvertering’ fra herbicidbenyttelse til mekanisk ukrudtsbekæmpelse mindst for kvægbrug, men også for svine- og plantebrug kan det med visse ændringer i sædskiftet lade sig gøre med et forholdsvis begrænset fald i dækningsbidraget. Med hensyn til faunaen er det samtidig en forudsætning, at der tages hensyn til striglingernes antal og tidsmæssige udførelse.
Samlet kan resultaterne for alle de modellerede arters vedkommende samles i følgende:
- i forbindelse med mekanisk ukrudtsbekæmpelse bør det anbefales:
- ikke at udføre mere end to striglinger i vår- kornafgrøder (blindstrigling + én efterfølgende strigling) ikke senere end ca. 30 dage efter såning
- ikke at udføre striglinger i vinter-kornafgrøder om efteråret, og kun at udføre én forårsstrigling inden ca. midt april, hvis vejrforholdene tillader det
Ukrudtsstrigling om efteråret kan under visse omstændigheder give en ganske effektiv ukrudtsbekæmpelse men den er forbundet med betydelig risici for alvorlige afgrødeskader (Rasmussen, 1998). I praksis vurderes det, at strigling på dette tidspunkt ikke anvendes i noget større omfang (pers. comm., Poul Henning Petersen, Dansk Landbrugsrådgivning Landscentret, Skejby). Med hensyn til forbruget af pesticider er der er i de udførte modelscenarier kun ændret praksis mht. til herbicidanvendelsen. Måske vil en udeladelse af herbicider nogle steder medføre, at der kommer lidt mere ukrudt til gavn for markfaunaen (her ses bort fra situationen i de tekniske scenarier), men miljømæssigt har der stadig været den samme belastning af andre kemiske hjælpemidler. De miljømæssige effekter (mindre risiko for nedsivning af kemiske stoffer) kunne være af betydning i denne forbindelse, men det er ikke undersøgt i nærværende projekt. De positive effekter kunne sandsynligvis have været større, hvis der samtidigt var sket en reduktion af andre pesticider (dosis/behandlingshyppighed af primært insekticider). Alt andet lige ville dette samtidig give plads til et potentiel større fødegrundlag i form af leddyr (biomasse, artsdiversitet). Hvorvidt dette rent dyrkningsmæssigt er en farbar vej vil afhænge meget af dyrkningstypen og en evt. reguleringsordning, der økonomisk tilgodeså forskellige former for ekstensiveret dyrkning. En vurdering af, om noget sådant er praktisk muligt/realistisk eller ønskeligt, ligger uden for rammerne af denne rapport.
7 Konklusioner
På baggrund af foregående resultat- og diskussionsafsnit gives i det følgende en kortfattet, samlet oversigt over denne undersøgelses hovedkonklusioner:
- Hvis lærkereder med æg eller unger rent fysisk udsættes for ukrudtsstriglinger med den her anvendte type af striglingsudstyr kan der for den enkelte rede forventes en risiko på mellem 50 og 100% for, at reden går til.
- De eksperimentelle dele i undersøgelsen viser imidlertid, at timingen for den (de) enkelte striglinger i den enkelte mark eller afgrøde er afgørende for, hvor mange reder der potentielt udsættes for fysiske påvirkninger.
- Under normal dyrkningspraksis er der ikke stor risiko for, at striglinger påvirker lærkernes reproduktion i økologisk dyrket vårhvede.
- Undersøgelsens modeldel underbygger, at dette under normal dyrkningspraksis heller ikke vil være tilfældet i en vårbyg eller vinterhvede, hvor striglinger erstatter den kemiske ukrudtsbekæmpelse.
- Mht. til sanglærken bør strigling i vårbyg derfor undgås efter ca. 10. maj (svarende til ca. 30 dage efter såning), og i vinterhvede bør der ikke strigles om foråret efter ca. 10. april.
- Striglinger udført før disse tidspunkter medfører ingen eller kun meget lille risiko for negative påvirkninger; striglinger udført senere end disse tidspunkter er potentielt forbundet med alvorlige negative effekter på sanglærkens reproduktion.
- Ukrudtsstriglinger udført under standard betingelser påvirker i højere grad end for sanglærkens vedkommende de udvalgte og modellerede arter af overfladelevende leddyr.
- Modelresultaterne viser, at alle ukrudtsstriglinger - om end i varierende grad afhængig af striglingstidspunkt og afgrøde - påvirker overlevelsen af edderkoppen Erigone atra og løbebillen Bembidion lampros negativt.
- Effekten på disse overfladelevende leddyr er mindst ved tidlige forårsstriglinger, men bedømt ud fra scenarier med monokulturer af vårbyg eller vinterhvede vil den samlede effekt af tre eller fire striglinger medføre alvorlige negative påvirkninger af bestanden af disse edderkopper og løbebiller. I det undersøgte ’kvægscenarie’ reducerer ukrudtsstriglingerne bestanden af begge arter med ca. 20 %.
- Overordnet kan resultaterne for de modellerede arters vedkommende samles i følgende:
- mht. at opnå færrest mulige negative effekter i forbindelse med ukrudtsstrigling i kornafgrøder anbefales det:
- ikke at udføre mere end to striglinger i vår- kornafgrøder (blindstrigling + èn efterfølgende strigling) ikke senere end ca. 30 dage efter såning
- ikke at udføre striglinger i vinter-kornafgrøder om efteråret, og kun at udføre èn forårsstrigling inden ca. midten af april, hvis vejrforholdene tillader det
- Regnormene blev ikke påvirket, og vil sandsynligvis ikke påvirkes af striglinger i vårhvede på lerjord.
- Springhaler og mider blev kun påvirket minimalt, og vil sandsynligvis kun i minimalt omfang påvirkes i vårhvede på lerjord.
- Sandsynligvis vil resultaterne vedr. regnorme og mikroleddyr også gælde for i hvert fald lignende afgrøder, mens det er mere usikkert, om forholdene vil være anderledes på lettere og mere sandede jordtyper.
- Ukrudtsstriglinger kan udføres uden betydelige påvirkninger af energiforbrug og dækningsbidrag.
- Med den nuværende teknik vurderes strigling ikke at være et realistisk alternativ til herbicider på konventionelle bedrifter, men alene et supplement, hvis der ikke foretages gennemgribende afgrøde- og sædskiftemæssige justeringer.
- En "ren" mekanisk ukrudtsbehandling (som fx kendes fra økologiske bedrifter, og som inkluderer kvikharvning, skrælpløjning og radrensning) kan resultere i en stigning i dieselforbruget på op til 10 %, og en forøgelse af det samlede direkte og indirekte energiforbrug på maksimalt 1-3 %; mindst på typiske kvægbrug og mest på svine- og planteavlsbrug.
- I forsøgene havde kun den første strigling (der blev ikke udført blinstrigling) effekt på ukrudt og afgrøde. De efterfølgende striglinger var virkningsløse.
- I forsøgene var bekæmpelseseffekten af ukrudt (33 %) lavere, end hvad der normalt kan opnås ved ukrudtsstriglinger. Dette vurderes at være forårsaget af jordtypen, som var svær og ubekvem lerjord.
- I begge år og på begge forsøgsmarker reducerede strigling udbyttet med 6 %. Dette vurderes at kunne afværges ved en mere optimal ukrudtsstriglings-strategi, hvor der foretages en blindstrigling og kun én opfølgende ukrudtsstrigling.
8 Perspektivering
8.1 Forskningsmæssige perspektiver
Direkte undersøgelser af striglingseffekter på leddyr og fugle er, så vidt det er forfatterne bekendt, stort set ikke eksisterende, og derfor udfylder dette projekt et forskningsmæssigt hul, bortset fra det igangværende pesticidforskningsprojekt ”Flora og fauna i marker: betydning af mekanisk ukrudtbekæmpelse og værdiberegning af mekanisk plantebeskyttelse”.
Umiddelbart peger resultaterne i nærværende projekt på, at striglingens effekt på energiforbrug og driftsøkonomi isoleret set er begrænset. Forsøgene med kunstige reder viste, at den direkte påvirkning på reder, der blev direkte udsat for strigling, var stor, hvorimod der blev ikke fundet effekter på springhaler eller regnorme under eksperimentelle betingelser. Resultaterne fra de udførte modelleringer indikerede en høj forventet negativ påvirkning for både edderkopper og biller, mens sanglærken under normal dyrkningspraksis kun vil blive direkte påvirket ved meget sent udførte striglinger.
I projektets eksperimentelle del har ukrudtsstrigling været anvendt i et økologisk dyrkningssystem, hvor betingelserne for en effektiv ukrudtsbekæmpelse ikke har været til stede. Dette skyldes primært de vanskelige jordbundsforhold, som har bevirket, at der kun er sket en mindre reduktion af ukrudtsmængden. Resultaterne fra forsøgene afspejler således først og fremmest de direkte skader på faunaen som følge af strigling og ikke de indirekte effekter som følge af, at ukrudtet fjernes. For at kunne foretage en bredere vurdering af striglingens utilsigtede effekter på faunaen, er der behov for forsøg, hvor strigling giver bekæmpelseseffekter, der er sammenlignelige med herbicidernes, således at både de direkte og indirekte effekter kan måles.
Modeltilgangen i dette projekt tillader at evaluere påvirkninger af striglinger på en landskabsmæssig skala under realistiske landbrugsmæssige betingelser - herunder en integrering af diverse dyrkningsmæssige operationer. Som en kontrast til disse resultater kan disse påvirkninger tillige sammenlignes med resultaterne fra ‘worst-case’ scenarier, hvor der er brugt samme tilgang (de tekniske scenarier). Denne kombination af at integrere mange variabler, og som en kontrast at sammenligne med en eksperimentel simplificering, er en ny og kraftfuld tilgang til vurdering af effekter gennem brug og evaluering af mange potentielle scenarier.
Modeltilgangen sammen med eksperimentelle, feltbaserede undersøgelser kan bruges til analyser af konsekvenser af forskellige kombinationer af ukrudtsstrigling med andre former for mekanisk og kemisk ukrudtsbekæmpelse, således at der skabes et bedre grundlag for at vælge ressourceøkonomiske og økologisk forsvarlige strategier for ukrudtsbekæmpelsen i Danmark.
8.2 Administrative perspektiver
Med baggrund i den nuværende handlingsplan for nedbringelse af pesticidforbruget (Pesticidplan 2004-2009) har det været dette projekts primære formål at belyse de faunamæssige, dyrkningsmæssige og økonomiske konsekvenser, der er knyttet til mekanisk ukrudtsbekæmpelse. Projektets resultater og anbefalinger mht. antal og ’timing’ af de enkelte mekaniske bekæmpelser af ukrudt vil kunne anvendes administrativt, hvis man ønsker at inddrage denne form for ukrudtsbekæmpelse i det mindste i dele af den konventionelle dyrkning for derigennem at bidrage til den generelle nedsættelse af pesticidforbruget. Hvis der var et politisk ønske om - måske som en del af en kommende handlingsplan? - at undersøge muligheden for at anvende en sådan form for ’ekstensiveret’ dyrkning, ville det sandsynligvis kræve en form for lovgivningsmæssige initiativer mht. evt. reguleringer i form af bestemte dyrkningsmæssige restriktioner og/eller anvendelse af dyrkningsmæssige, øremærkede tilskud og/eller afgifter evt. i forbindelse med udmøntningen af EU’s landbrugspolitik.
Den mest oplagte anvendelse af ukrudtsstrigling i konventionelt jordbrug er måske at bruge den som supplement til herbicidbehandling, således at herbicider ikke bliver anvendt hvert år og/eller kun bliver anvendt på steder i den enkelte mark, hvor der optræder en særlig stor ukrudtspulje, eller hvor der på anden vis har vist sig problemer med opformering af ’aggressive’ ukrudtsarter. Sådanne tiltag kunne eks. anvendes i områder, som man af forskellige årsager ønsker udlagt som miljøfølsomme områder (eks. vandindvindingsområder), men samtidig ønsker en rationel dyrkningsform der hindrer for kraftige opformeringer af ’problem’ ukrudtsarter.
Projektets anbefalinger for strigling vil desuden kunne anvendes, hvis man – enten i økologiske landbrugskredse eller administrativt – ønsker at sikre en praksis for markstrigling, som i videst mulige omfang tager hensyn til naturen på markfladen.
Dette projekt har endvidere genereret en ny viden, som sammen med de kommende resultater fra et sideløbende projekt under Miljøstyrelsens pesticidforskningsmidler (’Flora og fauna i marker: betydning af mekanisk ukrudtsbekæmpelse og værdisætning af mekanisk plantebeskyttelse’) kan medvirke til at vurdere ’omkostningerne’ i forbindelse med mekanisk ukrudtsbekæmpelse.
9 Litteraturliste
Axelsen, J. A. & K.T. Kristensen (2000). Collembola and mites in plots fertilised with different types of green manure. Pedobiologia 44, 556-566.
Bichel-udvalget (1998). Rapport fra Hovedudvalget. Miljøstyrelsen, København. ISBN 87-7909-296-9.
Bilde, T., Axelsen, J. A. & S. Toft (2000). The value of Collembola from agricultural soils as food for a generalist predator. Journal of Applied Ecology 37, 672-683.
Bilde, T. & C.J. Topping (2004). Life history traits interact with landscape composition to influence population dynamics of a terrestrial arthropod: a simulation study. - EcoScience 11(1): 64-73.
Boo, W. de (1993). Environmental and Energy Aspects of Liquid Biofuels. Centrum voor energisparing en shone technologie. Delft, Holland.
Braae, L., Nøhr, N. & B.S. Petersen (1988). Fuglefaunaen på konventionelle og økologiske brug. - Miljøprojekt nr.102, Miljøstyrelsen.
Chamberlain, D.E., Fuller, R.J., Bunce, R.G.H., Duckworth, J.C. & M. Shrubb (2000). Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. Journal of Applied Ecology, 37, 771-788.
Chamberlain, D.E., Wilson, A.M., Browne, S.J. & J.A. Vickery (1999). Effects of habitat type and management on the abundance of skylarks in the breeding season. Journal of Applied Ecology, 36: 856-870.
Chiverton, P.A. (1986). Predator density estimation and its effects on populations of Rophalosiphum padi (Hom.:Aphididae) in spring barley. Annals of Applied Biology 109: 49-60.
Cortet, J. & N. Poinsot-Balaguer (2000). Impact of phytopharmaceutical products on soil microarthropods in an irrigated maize field: the use of the litter bag method [French]. Canadian Journal of Soil Science 80, 237-249.
Dalgaard, T. (1998). Ændring i forbrug af fossil energi ved omlægning til pesticidfrit landbrug. Delrapport om Miljø- & Sundhedmæssige konsekvenser i scenario med 100 % pesticidfrit jordbrug i Danmark. Miljøstyrelsen, København. 16 p.
Dalgaard, T. (2001). Simulation and Generalisation of Agricultural Resource Use. PhD-thesis. The Royal Veterinary and Agricultural University, Copenhagen. 184 pp. ISBN 87-988287-3-8.
Dalgaard, T., Dalgaard, R. & A.H. Nielsen (2003). Energiforbrug på økologiske og konventionelle landbrug. Grøn Viden Markbrug nr. 260. Danmarks Jordbrugsforskning, Foulum. ISSN 1397-985X.
Dalgaard, T., Halberg, N. & J. Fenger (2002). Can organic farming help to reduce national energy consumption and emissions of greenhouse gasses in Denmark? In: EC van Lerland and AO Lansink (eds.) Economics of sustainable energy in agriculture. Economy and Environment vol. 24. p. 191-204. Kluwer Academic Publishers, Dordrecht, The Netherlands. ISBN 1-4020-0785-X.
Dalgaard ,T., Halberg, N. & J.R. Porter (2001). A model for fossil energy use in Danish agriculture used to compare organic and conventional farming. Agriculture, Ecosystems & Environment 87(1) 51-65.
Dalgaard, T., Kjeldsen, C., Rasmussen, B.M., Fredshavn, J.R., Munier, B., Schou, J.S., Dahl, M., Wiborg, I.A., Nørmark, P. & J.F. Hansen (2003)b. ARLAS' scenariesystem - Et grundlag for helhedsorienterede konsekvensvurderinger af ændringer i arealanvendelsen og landbrugsproduktionen, pp. 87-108, i Afslutningsseminar for projektet: Areasanvendelse og landskabsudvikling - Fremtidsperspektiver for natur, jordbrug, miljø og arealforvaltning. 3.- 4. december 2002, Golf Hotel, Viborg.
Daunicht, W.D. (1998). Zum ienfluss der Feinstruktur in der Vegatation auf die Habitatwahl, Habitatnutzung, Siedlungsdichte und populationsdynamik von Feldlerchen (Alauda arvensis) in grossparzelligem Ackerland. University of Bern, Germany. (Phd).
Edwards, C. & P.J. Bohlen (1992). The effects of toxic chemicals on earthworms. Rev. Env. Contam. Toxicol. 125, 23-99.
Edwards, C.A., Sunderland, K.D. & K.S. George (1979). Studies on polyphagous predators of cereal aphids. Journal of Applied Ecology 16: 811-823.
Edwards, C. & P. Bohlen (1996). Biology and Ecology of Earthworms. Chapman and Hall, London.
Esbjerg, P., Jensen, A., Johnsen, I., Kristensen, K., Navntoft, S., Petersen, B.S., Petersen, P.H. & J.E. Ørum (2006). Udkast til årsrapport 2005 til Miljøstyrelsen for forskningsprojektet ’Flora og fauna i marker: betydning af mekanisk ukrudtsbekæmpelse og værdisætning af mekanisk plantebeskyttelse.
Fluck, R.C. (1992). Energy in farm production. Energy in World Agriculture vol. 6. Elsevier, Amsterdam. 367 p. ISBN 0-444-88681-8.
Forskningscenter for Økologisk Jordbrug (2006). Temaside om ukrudtsbekæmpelse. www.foejo.dk/aktuelt/tema/pdyr/ukrudt.html. Forskningscenter for Økologisk Jordbrug, Foulum.
Fuller, R.J., Gregory, R.D., Gibbons, D.W., Marchant, J.H., Wilson, J.D., Baillie, S,R, & N. Carter (1995). Population declines and range contractions among lowland farmland birds. Conservation Biology, 9, 1425-1441.
Galbraith, H. (1988). Effects of agriculture on the breeding ecology of lapwings Vanellus vanellus. – J. Appl. Ecol. 25: 487-503.
Glutz von Blotzheim UN & K.M. Bauer (1985). Der Vogel Mitteleuropa. 10 (1), Passeriformes (1. Teil), 229-281.
Gravsholt H., Jørgensen K., Meinertsen P., Pedersen S.S., Jørgensen A., Udesen, F., Nørgaard, E., Fisker, C., Søeberg, H.P., Høy, J.J. & T.V. Jørgensen (2005). Økologikalkuler 2004. Landbrugsforlaget.
Grogger, J. T. & R. T. Carson (1991). Models for truncated counts. Journal of Applied Econometrics 6: 225- 238.
Gundersen, H. (2006). Afgrøders tolerance ved ukrudtsharvning. Institut for Jordbrugsvidenskab, Den Kongelige Veterinær- og Landbohøjskole. Speciale. 37 sider.
Hald, A.B. & J. Reddersen (1990). Fugleføde i kornmarker - insekter og vilde planter. Miljøprojekt nr. 125. Miljøministeriet, Miljøstyrelsen.
Jenny, M.(1990). Populationsdynamic der Feldlerche Alauda arvensis in einer intensiv genutzten Agrarlandschaft des Sweizerischen Mittellandes. Der Ornitilogischer Beobachter 87:153-163.
Jepsen, J.U., Topping, C.J., Odderskær, P. & Andersen, P.N. (2005). Evaluating consequences of land-use strategies on wildlife populations using multiple-species predictive scenarios. - Agriculture, Ecosystems & Environment 105(4): 581-594.
Jørgensen, U. & Dalgaard, T. (2004). Energi i Økologisk Jordbrug. Reduktion af fossilt energiforbrug og produktion af vedvarende energi. FØJO-rapport nr. 19. 164 s. www.foejo.dk.
Krogh, P. H. (1991). Perturbation of the soil microarthropod community with the pesticides benomyl and isofenphos. Pedobiologia 35, 71-88.
Krogh, P., Holmstrup, M., Jensen, J. & S. Petersen (1996). Økologisk vurdering af spildevandsslam i landbrugsjord. Arbejdsrapport fra Miljøstyrelsen, Nr. 43. Ministry of Environment and Energy, Denmark. 53 pp.
Lambert, D. (1992). Zero-inflated Poisson regression, with an application to defects in manufacturing. Technometrics 34: 1-14.
Little, L.M. (1981). Interpretation and presentations of results. HortScience 16, 637-40.
Marcussen, B. M., Axelsen, J. A. & S. Toft (1999). The value of two Collembola species as food for a linyphiid spider. Entomologia Experimentalis Et Applicata 92, 29-36.
Miljøministeriet (1998)a. Udvalget til vurdering af de samlede konsekvenser af en afvikling af Pesticidanvendelsen. Rapport fra hovedudvalget. http://miljoindflydelse.dk/default.asp?Sub=http://miljoindflydelse.dk/udgiv/Publikationer/2002/87-7972-210-5/html/helepubl.htm
Miljøministeriet (1998)b. Udvalget til vurdering af de samlede konsekvenser af en afvikling af Pesticidanvendelsen. Rapport fra den tværfaglige økologigruppe. http://miljoindflydelse.dk/default.asp?Sub=http://miljoindflydelse.dk/udgiv/Publikationer/2002/87-7972-210-5/html/helepubl.htm
Morris, J., Wilson, J.D., Whittingham, M.J. & R.B. Bradbury (2005). Indirect effects of pesticides on breeding yellowhammer (Emberiza citrinela). Agriculture, Ecosystemes & Environmenment, 106, 1-16.
Navntoft, S. & P. Esbjerg (2003). Arthropods. In: Flora and fauna changes during conversion from conventional to organic farming (S. Navntoft, ed.), Pesticides Research 74, 32-49. Danish Environmental Protection Agency. Danish Ministry of the Environment.
Odderskær, P. (2002). Konsekvenser for sanglærken ved omlægning til økologisk jordbrug, pp. 139-169, i "Omlægning til økologisk jordbrug i et lokalområde - Scenarier for natur, miljø og produktion. FØJO-rapport nr. 12, (Langer, V., Red.).
Odderskær P., Prang. A, Poulsen J.G., Elmegaard, N. & P.N. Andersen (1997). Skylark Reproduction in Pesticide Treated and Untreated Fields. Pesticide Research No 32. Ministy Of Environment and Energy, Denmark. Miljø- og Energiministeriet Miljøstyrelsen.
Olesen, J.E. (2005). Drivhusgasser fra jordbruget - reduktionsmuligheder. DJF rapport - Markbrug 113, 161 pp. http://web.agrsci.dk/djfpublikation/djfpdf/djfma113.pdf
Päzhold, R. (1975). Die Feldlerche Alauda arvensis. Die Neue Brehm-Bücherei. A Ziemsen Verlag - Wittenberg Lutherstadt.
Pedersen, C.A. (2000). Oversigt over Landsforsøgene. Dansk Landbrugsrådgivning, Skejby.
Pedersen, C.A. (2002). Oversigt over Landsforsøgene. Dansk Landbrugsrådgivning, Skejby.
Pedersen, C.A. (2005). Oversigt over Landsforsøgene. Dansk Landbrugsrådgivning, Skejby, 416s.
Petersen, H. (2002). General aspects of collembolan ecology at the turn of the millennium. Pedobiologia 46, 246-260.
Petersen, H. & M. Luxton (1982). A comparative analysis of soil fauna populations and their role in decomposition. Oikos 39, 287-388.
Petersen, P.H. (2006). 'Praksis vedr. ukrudtsharvning og radrensning i vintersæd'. Notat af 26/1 til styregruppen for pesticidforskningsprojekt "Ukrudtsstriglingens effekter på markfladens naturlige plante- og dyreliv samt landbrugets resourceforbrug".
Potts, G.R. (1986). The Partridge, Pesticides, Predation and Conservation. Collins, London.
Rapoport, E. & M. Tschapek (1967). Soil water and soil fauna. Rev. Écol. Biol. Sol 4, 1-58.
Rasmussen, J. (1991)a. A model for prediction of yield response in weed harrowing. Weed Research, 31: 401-408.
Rasmussen, J. (1991)b. Udbytteeffekter ved ukrudtsharvning - udvikling og anvendelse af en simuleringsmodel. 8. Danske Planteværnskonference/Ukrudt: 189-201.
Rasmussen, J. (1992).Testing harrows for mechanical control of weeds in agricultural crops. Weed Research 32: 267-274.
Rasmussen, J. (1996). Mechanical weed management. In: Second International Weed Control Congress, Copenhagen: 943-948.
Rasmussen, J. (1998). Ukrudtsharvning i vinterhvede. 15. Danske Planteværnskonference. DJF rapport nr 2. Markbrug: 179-189
Rasmussen, J. & J. Ascard (1995). Weed control in organic far*ming systems. In D.M. Glen, M.P. Greaves and H.M. Anderson (ed*s.), Ecology and Integrated Farming Systems, John Wiley & Sons Ltd, England, 49-67
Rasmussen, I.A, Melander, B., Rasmussen, K. & J. Rasmussen (1997). Regulering af ukrudt. In: Økologisk Planteproduktion. SP rapport nr. 15 (ed. E.S. Kristensen), 63-86. Forskningscenter for Økologisk Jordbrug, Foulum, Danmark.
Rasmussen, J. & K. Rasmussen (1994). Strategier for mekanisk ukrudtsbekæmpelse i vårsæd. 11.Danske Planteværns-konference/Ukrudt. SP rapport 6 (1994): 149-162
Rasmussen, J. & K. Rasmussen (1995). A strategy for mechanical weed control in spring barley. 9. EWRS (European Weed Research Society) Symposium Budapest 1995: Challenges for Weed Science in a Changing Europe: 557-564.
SAS Institute, Inc. (1992). SAS/LAB Software: User's Guide, Version 6, First Edition. SAS Institute Inc., Cary, NC. 291 pp.
SAS Institute Inc. (2004). SAS/STAT® 9.1 User’s Guide. Cary. NC: SAS Institute Inc.
Scheffer, M., Baveco, J.M., Deangelis, D.L., Rose, K.A & E.H. Vannes (1995). Super-individuals a sample solution for modelling large populations on an individual basis. Ecological Modelling, 80 (2-3): 161-170.
Schläpfer, A. (1988). Populationsökologie der Feldlerche Alauda arvensis in der intensiv genutzten Agrarlandschaft. Der Ornitologische Beobachter 85:309-371.
Schou, J. (2006). EU's landbrugsordninger og pesticidpolitikken. Revideret projektbeskrivelse. 21. marts. Miljøstyrelsen, København.
Scott-Fordsmand, J.J., Strandberg M. & M. Holmstrup (2000). Ecotoxicology of glyphosate. Report for the Danish Environmental Protection Agency. National Environmental Research Institute, Silkeborg, Denmark. 47 pp.
StataCorp. (2005). Stata Statistical Software: Release 9. College Station, TX: StataCorp LP.
Sunderland, K.D. (1975). The diet of some predatory arthropods in cereal crops. Journal of Applied Ecology 12: 507-515.
Thomsen, H. (2003). Kvikbekæmpelse med "minisommerbrak". Nyhedsbrev fra forskningscenter for Økologisk jordbrug. Nr. 4. www.foejo.dk.
Thorbek, P. & T. Bilde (2004). Reduced numbers of generalist arthropod predaters after crop managementJ. Appl. Ecol. 41: 526-538.
Thorbek; P. & C.J. Topping (2003). The influence of landscape diversity and heterogeneity on spatial dynamics of agrobiont linyphiid spiders: an individual based model. Ecol. Model.
Toft, S. & D.H. Wise (1999). Growth, development, and survival of a generalist predator fed single- and mixed.
Topping, C.J, Hansen, C.J., Jensen, T.S., Jepsen, J.U., Nikolajsen, F. & P. Odderskær (2003). ALMaSS, an agent-based model for animals in temperate European Landscapes. ECOMOD, 3318, pp 1-18, ELSEVIER.
Topping, C.J. & P. Odderskær (2004). Modelling the influence of temporal and spatial factors on the assessment of impacts of pesticides on skylarks. Environmental Toxicology and Chemistry.
Wilson , J.D., Evans, J., Brown, S.J. & J.R. King (1997). Territory distribution and breeding success of skylarks Alauda arvensis on organic and intensive farmland in southern England. Journal of Applied Ecology 34, 1462-1478.
Ørum, J.E. (2003). Driftsøkonomisk analyse af reduceret pesticidanvendelse i dansk landbrug – en opdatering af Bicheludvalgets analyser. Rapport nr. 163. Fødevareøkonomisk Institut, København.
Östman, Ö., Ekbom, B. & J. Bengtson (2003). Yield increase attributable to aphid predation by ground-living polyphagous natutal enemies in spring barley in Sweden. Ecological Economics 45, 149-158, ELSEVIER.
Bilag A
1 Beskrivelse af ALMaSS-modelsystemet
I det følgende gives en beskrivelse af ALMaSS-modelsystemet som anvendes i forbindelse med modelscenarierne i projekt ”Ukrudtsstriglingens effekter på dyr, planter og ressourceforbrug”.
1.1 ALMaSS-modelsystemet
Navnet ’ALMaSS’ er en forkortelse af ´Animal, Land and Man Simulation System´og refererer til et samlet dynamisk modelsystem, der er i stand til at tage højde for det dynamiske samspil af forskellige biologiske og abiotiske faktorer i tid og rum, der har afgørende betydning for, hvordan, og med hvilken effekt, en given samling af individer (lokalbestand) i et nærmere defineret landskab bliver påvirket af forskellige driftsbetingelser.
Modelsystemet består i princippet af to hovedkomponenter – en landskabsmodel og en (eller flere) individbaserede dyremodeller samt en række undermodeller (vejrmodel, dyrkningsmodeller, vækstmodeller for afgrøde og vegetation, sædskifter og ’driftsleder’; se nedenfor). Landskabsmodellen kan bestå af et konstrueret landskab eller en kopi (digitalisering)af et virkeligt landskab som i nærværende projekt. Landskaberne kan være simple eller meget detaljerede, og udgør det miljø eller den fysiske ramme de enkelte individer i dyremodellerne lever i.
De individbaserede modeller adskiller sig fra de fleste andre modeltyper (eks. matematiske) i, at de har det enkelte individ som udgangspunkt og ikke en samling eller population af individer, der opfører sig ens på alle udefra kommende stimuli. I ALMaSS har de enkelte individer deres egne agenda: på et hvilket som helst tidspunkt og sted at agere efter, hvad der er bedst mht. overlevelse og reproduktion (denne modelegenskab har giver navn til modeltypen ’agent-baserede modeller’ forkortet ABM) . Dyrene kan kommunikere og erkende tilstande og ændringer, i det miljø de lever. Samlet betyder dette, at det enkelte individ hele tiden reagerer på det dynamiske samspil af (eller interaktioner mellem) forskellige faktorer, der påvirker det. De enkelte dyremodeller er konstrueret således, at det løbende under modelsimuleringerne er muligt at indhente oplysninger om forskellige variabler vedr. reproduktion, population mm.
1.2 Integration af enkeltfaktorer gennem modellering
Bestandsudvikling af de enkelte arter er bestemt af en lang række faktorer. Ved ændret driftspraksis ændres mange af disse faktorer. Modellering af effekten af forskellige dyrkningsscenarier er en måde at integrere den foreliggende viden om samspillet mellem de modellerede arter og det landskab, de befinder sig i og dermed bidrage til at forstå enkeltfaktorernes betydning.
ALMaSS er derfor et godt redskab til at undersøge, hvilke konsekvenser eks. en bestemt ændring i dyrkningspraksis vil have for en given population - eller dele heraf – i et bestemt område. Da en given ændring i dyrkningspraksis oftest påvirker forskellige arter i forskellig retning, er det som i indeværende projekt en fordel at undersøge de samme ændringer for en række arter med meget forskellig levevis.
I tidligere undersøgelser har ALMaSS eks. været anvendt til at undersøge pesticiders effekter på sanglærken set i relation til markstørrelse, og afgrødetype (Topping & Odderskær 2004). Modelleringerne, der foretages i nærværende projekt, har til formål gennem ’tekniske scenarier’ kvantitativt at beskrive konsekvenserne af forskellige striglings intensitet og timingen for striglingernes udførelse for tre almindeligt forekommende agerlandsarter, som repræsenterer forskellige trofiske niveauer og levevis (sanglærke samt edderkop og løbebille). Effekter måles på forskellige variabler for reproduktion og bestandsforhold i et konkret landskab, hvor alle bedrift og dyrkningsforhold kan manipuleres (scenarier), men som udgangspunkt er baseret på konkrete forhold i ’værkstedsområdet’ (for nærmere beskrivelse af dette se: Dalgaard et al. 2004). I princippet vil antallet af outputvariabler være så godt som ubegrænset, men i praksis har hovedparten af outputvariabler været inden for årlig baseret populationstørrelse (inkl. mål for forsk. livsstadier) herunder vækst-, ekstensions- reproduktions- og mortalitetsrater, antal individer der påvirkes af en bestemt ændring (eks. fra strigling), og for en enkelt art (markmus) er det også muligt at få output i form af genetiske variabler. I nærværende projekt anvendes følgende modeloutput: alle tre arter: årlige total pop. pr 31/12, sanglærke: total antal årsunger, klægningssucces, udflyvningssucces, ynglesucces, redemortalitet, ægkuldmortalitet (for nærmere forklaring se afsnit 3.4 i rapport).
I det følgende beskrives de enkelte elementer i ALMaSS modelsystemet. I princippet kan alle modelparametre ændres efter behov, men nogle ændringer er lette at ændre medens en ændring af andre rent tidsmæssigt er forbundet med et stort arbejde. I det følgende beskrives disse forhold under de enkelte delmodeller. For beskrivelsen af de individbaserede modeller er der udvalgt eksempler for sanglærken, men der vil ikke i det følgende blive givet en detaljeret oversigt over parametriseringen af de enkelte artsmodeller, da dette ligger udenfor rammerne for nærværende notat. Detaljerede oversigter kan findes i følgende publikationer: -(sanglærke: Topping & Odderskær 2004; edderkop: Thorbek & Topping, 2005; løbebille: Bilde & Topping, 2004; samlet for hele modelsystemet: www.almass.dk).
1.3 Landskabsmodellen
Landskabsmodellen – eller værkstedsområdet, som er en kopi af et virkeligt område, udgør den ”verden” de modellerede dyr skal leve og overleve i. Den indeholder en detaljeret gengivelse af alle vigtige landskabselementer (læhegn, markskel, marker, veje og bygninger mm.). Med mindre det er ønsket at undersøge forskellige udgaver af et landskab, holdes størrelse og placering af disse elementer konstant både i ’før-scenariet’, som repræsenterer de nuværende forhold, og i alle ’efter- scenarier’ der simuleres, og kun markafgrøder (sædskifte, eller som i denne undersøgelse, dyrkningspraksis) ændres mellem scenarierne.
Det udvalgte værkstedsområde, hvori de enkelte modelscenarier er udført, er et 10 x 10 km stort område beliggende sydøst for Bjerringbro (Figur 1).
Figur 1. Udsnit (5x5 km, sydøst for Bjerringbro) af modellandskab, hvori modelkørsler foretages.
Området omfatter alle bedrifter i hele området. Dette valg er truffet ud fra modelmæssige hensyn, ud fra et ønske om, at det valgte landskab indeholder så stor andel dyrket land som muligt, hvilket for de pågældende modellerede arter er centralt. Ud fra flyfoto og markblok kort blev der foretaget en detaljeret digitalisering af området. Alle digitaliserede arealer (polygoner) blev defineret i forhold til en række udvalgte kategorier (marker, bygninger, haver, areelle småbiotoper, læhegn, skove, veje/vejrabatter, vandløb mm., Figur 2). Ved en efterfølgende inspektion af området blev alle arealinformationer vedrørende de forskellige polygontyper korrigeret for evt. fejl i forhold til placering, arealstørrelse og polygontype. For de individuelle læhegns vedkommende blev der desuden suppleret med oplysninger om højde, strukturel udformning mm. Alle oplysninger vedrørende bedriftstyper og ejerforhold (her: reelt oplysninger om arealer drevet af samme person) har været tilgængelige via GLR/CHR - register oplysninger.
Som udgangspunkt er de enkelte bedriftsenheder (i alt 123 bedrifter) og typer i modellandskabet defineret ud fra de konkrete forhold i 1998. Den enkelte bedriftsstørrelse og geografiske placering er defineret ud fra den person, der driver de pågældende marker (Figur 3). Alle ”beslutninger” om udførelsen af markoperationer på en given mark på et givet tidspunkt bliver foretaget i en ”Driftsleder” (se nedenfor), der samlet koordinerer oplysninger om bedriftstype, sædskifte, jordbunds- og vejrmæssige forhold.
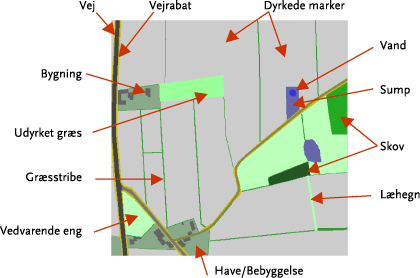
Figur 2. Udsnit af det digitaliserede landskabsmodel, med angivelse af selvstændigt definerede polygontyper (habitater).
Figur 3. Angivelse af bedriftsforhold: marker med samme farve er drevet af samme landmand.
Ændring af landskabsparametre
Arbejdsmæssigt er udarbejdelsen (digitalisering) af en detaljeret kopi af et virkeligt landskab som her anvendte (10x10 km) en omfattende proces, der tager ca. tre måneder at generere. Derefter er en ændring i f.eks. typen af eksisterende polygontyper let at foretage, mens en ændring i den fysiske placering eller ’mængden af’ en eller flere strukturer i landskabet (polygontyper som f.eks. læhegn eller skove) forbundet med et større arbejde – afhængigt af ændringernes omfang. Ligeledes vil eks. en bestemt – og individuel - ændring af eksisterende markers størrelse være forbundet med et større digitaliseringsarbejde, medens en ens opdeling af alle marker vil være forholdsvis let at foretage.
1.4 Vejrdata og vækstmodeller for afgrøder
De valgte modelarters muligheder for overlevelse og reproduktion er i høj grad afhængig af vækstforholdene og strukturen (bladareal, biomasse, højde) af de enkelte afgrøder indenfor yngle- eller ’home-range’ området. For på en realistisk måde at ”gengive” virkelige forhold, er der udviklet specifikke vækstmodeller for de afgrøder, der er anvendt i forskellige typer af sædskifter (se efterfølgende). Vækstmodellerne er udarbejdet på grundlag af eksisterende modeller (Olesen og Heidmann, 1990; Plauborg og Olesen, 1991) samt data indsamlet ved DJF og DMU. Modellerne er baseret på ”graddage”, således at både bladareal og højdeudvikling er relateret til temperatursummer med en basistemperatur på 0°C (for nærmere beskrivelse, se Bilag B). Vejrdata, der er anvendt i plantevækstmodeller, dyremodeller og ”Driftslederen” (se efterfølgende) er baseret på data fra DJF-Foulum fra perioden 1988-1999. I modelleringerne er der anvendt daglige gennemsnit for temperatur (°C), nedbør (mm) og vindstyrke (m/sek.). De samme vejrdata indgår således i det modellerede tidsrum på 55 år i alt i perioder af 11 år.
Ændring af vejrdata og vækstmodeller
En ændring af vejrdata er let at foretage. Det vil dog til dels afhænge af, i hvor stor grad det er muligt at fremskaffe over et specifikt lokalt område, og over hvor lang en (evt. bestemt) årrække, det er ønskeligt at anvende data fra. De enkelte vækstmodeller (afgrøder og urtevegetation generelt) er forbundet med et stort arbejde at foretage ændringer i.
1.5 ’Driftsleder’ og dyrkningsmodeller
Dyrkningsmodeller, der beskriver vækstforhold og dyrkningstiltag er baseret på dyrkningsvejledninger (Dansk Landbrugsrådgivning) og konsulentrådgivning fra Djursland Landboforening - Landbocentret Følle, Godsforvaltningen ved Fussingø Statsskovdistrikt og DMU, Afd. for Vildtbiologi og Biodiversitet. For hver enkelt afgrødemodel er der i videst mulig omfang medtaget de variationer i dyrkningsmæssige tiltag, der er bestemt af f.eks. brugstype, geografiske forhold, jordbund, vejrforhold mm. Den indbyggede variation bevirker, at vækstforhold og dyrkningstiltag indenfor de enkelte afgrøder skifter mellem marker og bedrifter i de enkelte modelår. I alt er der udarbejdet dyrkningsmodeller for ca. 50 forskellige afgrøder (inkl. forskellige dyrkningsvarianter af disse), herunder de afgrøder der benyttes under nærværende projekt (Tabel 1). Et eks. på en dyrkningsmodel for vårbyg er angivet i projektbeskrivelsen (Appendiks 2 i Odderskær et al. 2003) for nærværende projekt.
Tabel 1. Oversigt over modelafgrøder, der indgår i sædskiftet på bedriftstypen kvægbrug i nærværende projekt.
| Kvægbrug |
| Vårbyg m. udlæg |
| 1. års kløver græs til slet/afgræsning |
| 2. års kløver græs til slet/afgræsning |
| Vinterhvede |
| Vårbyg |
| Vårbyg m. udlæg (til ensilering) |
| 1. års kløver græs til slet/afgræsning |
| Vårbyg m. udlæg(til ensilering) |
| Vårbyg |
I dyrkningsmodellen for de enkelte afgrøder antages, at markoperationerne i Tabel 2 kun udføres under bestemte vejrforhold. De fleste markoperationer vil således ikke blive udført før 3 – 5 dage efter forudgående nedbør, høst foretages tidligst 5 dage efter nedbør og marksprøjtninger udføres kun i tørvejr og under gunstige vindforhold (vindstyrker < 4, Boufort skala).
Tabel 2. Samlet oversigt over de markoperationer der er anvendt i de eksisterende dyrkningsmodeller.
| Markoperationer | |
| Efterårspløjning | Fungicidbehandling |
| Efterårsharvning | Insekticidbehandling |
| Efterårstromling | Radrensning |
| Efterårssåning | Strigling |
| Forårspløjning | Blindstrigling |
| Forårsharvning | Kunstvanding |
| Forårstromling | Skårlægning |
| Forårssåning | Høst |
| Gødskning | Køer på græs |
| - NPKS | |
| - NPK | Græsslet til hø |
| - PK | Græsslet til ensilage |
| - Flydende ammoniak | Græsslet til hø/ensilage |
| - Gylle | Afpudsning af ukrudt |
| - Mangansulfat | Halmsnitning |
| - Fastgødning | Halm/Høvending |
| - Slam | Halm/Høpresning |
| - N-ammoniumsulfat | |
| Herbicidbehandling | Stubharvning |
| Vækstregulering | |
Som en vigtig del mht. sanglærken specielt er mængden af føde, der er tilgængelig fra en given habitat, afhængig af den tilstedeværende insektbiomasse (total for alle insekt fødeemner), som igen er proportionelt relateret til vegetationsstrukturen (højde og biomasse). Disse relationer blev for almindelige landbrugsafgrøder genereret fra feltdata indhentet i modelsimuleringsområdet ved Bjerringbro (Topping, unpublished). Ud fra disse data er der udarbejdet nogle generelle funktioner, der beskriver bestandsudviklingen af insekter i relation til den tidsmæssige udvikling (plantebiomasse) af de enkelte afgrøder og ukrudt (generelt). Insekttætheder omregnes herefter til en gennemsnitlig tørvægt per arealenhed for de vigtigste grupper af insektfødeemner efter eksisterende datamateriale (Odderskær et al. 1997).
Ændring af dyrkningsmodeller
En ændring i eksisterende dyrkningsmodeller er forholdsvis let (en til få dage) afhængig af, om det et er spørgsmål om f.eks. at ændre i den procentdel der udfører en bestemt markoperation, eller det f.eks. er en ændring i typen eller rækkefølgen af markoperationer). Derimod vil det arbejdsmæssigt tage fra to til tre uger i alt at lave og teste en ny afgrøde fra bunden afhængigt af, hvor kompliceret et dyrkningsmønster, der er tale om.
1.6 Effekter af markoperationer
Alle de tre modelarter i nærværende undersøgelse påvirkes af markoperationer i forskellig udstrækning. Som eks. på dette vises nedenstående - med udgangspunkt i sanglærken og insektfødeemner generelt (litteraturdata samt egne data) - en oversigt over de i lærkemodellen anvendte værdier for effekter af forskellige markoperationer (Tabel 3). Hvor data vedrørende de tilknyttede påvirkninger på æg/redeunger/insekter for en markoperation ikke har været tilgængelige i litteraturen, er der for æg og ungers vedkommende tilknyttet en dødelighed på 100 % (angivet med ** i Tabel 3) i de tilfælde, hvor den pågældende markoperation er af en umiddelbar radikal negativ karakter. I øvrige tilfælde er der som udgangspunkt tildelt en dødelighed på 0 %. De angivne dødeligheder på æg og unger som følge af markstrigling stammer fra de resultater (kunstige reder), der er fremkommet i nærværende undersøgelse (samlet gennemsnit for 2004 og 2005). Dødelighed på æg og unger som følge af pesticidsprøjtninger er ikke medtaget under de direkte følger af markoperationer, da disse indirekte påvirkninger (Odderskær et al. 1997) modelleres som påvirkninger gennem en fødereduktion.
Tabel 3. Estimerede effekter (angivet i % dødelighed) af forskellige markoperationer på sanglærkeæg og –unger anvendt i modellen (kilder: Busche 1989, Jenny 1990, Odderskær et al. 1997 b, Daunicht 1998, Odderskær 1998, Poulsen et al. 1998, Wakeham-Dawson et al. 1998, Odderskær et al. 2001), samt insekter (kilder: Sunderland & Topping 1993, Poulsen et al. 1998, Topping & Sunderland 1998, Wakeham-Dawson et al. 1998). For æg og unger angiver de opførte procenter den individuelle dødelighed i det tilfælde de udsættes for den pågældende markoperation. De anførte græsningseffekter på æg/unger er en daglig (tilfældig) dødelighed pr. rede m. æg/unger. For insekter er den angivne dødelighed en gennemsnitlig dødelighed (reduktion) for populationen af insekter i den pågældende mark, der udsættes for den givne markoperation. * : indirekte effekt modelleret gennem en reduktion af insekter (for æg/unger) og plantebiomasse (for insekter). **: angiver, at den pågældende begivenhed er skønnet til at medføre en dødelighed på 100 %, men at den pågældende begivenhed i øvrigt kun i sjældne tilfælde vil finde sted.
| Markoperation | Voksen | Æg | Redeunger | Udfløjne unger | Insekter (generelt) |
| Efterårspløjning | 0 | ** | ** | 0 | 90 |
| Efterårsharvning | 0 | ** | ** | 0 | 70 |
| Efterårstromling | 0 | ** | ** | 0 | 0 |
| Efterårssåning | 0 | ** | ** | 0 | 0 |
| Forårspløjning | 0 | 100 | 100 | 0 | 90 |
| Forårsharvning | 0 | ** | ** | 0 | 70 |
| Forårstromling | 0 | ** | ** | 0 | 0 |
| Forårssåning | 0 | ** | ** | 0 | 0 |
| NPKS | 0 | 0 | 0 | 0 | 0 |
| NPK | 0 | 0 | 0 | 0 | 0 |
| PK | 0 | 0 | 0 | 0 | 0 |
| Flydende ammoniak | 0 | 100 | 100 | 0 | 0 |
| Gylle | 0 | 2 | 2 | 0 | 0 |
| Mangansulfat | 0 | 0 | 0 | 0 | 0 |
| Fastgødning | 0 | 0 | 0 | 0 | 0 |
| Slam | 0 | 0 | 0 | 0 | 0 |
| N-ammoniumsulfat | 0 | 0 | 0 | 0 | 0 |
| Herbicid | 0 | 0 | * | * | * |
| Vækstregulator | 0 | 0 | 0 | 0 | 0 |
| Fungicid | 0 | 0 | 0 | 0 | 0 |
| Insekticid | 0 | 0 | * | * | 80 |
| Radrensning | 0 | 100 | 100 | 0 | 75 |
| Strigling, | 0 | 72 | 72 | 0 | 30 |
| Vanding | 0 | 0 | 0 | 0 | 0 |
| Skårlægning | 0 | 2 | 2 | 0 | 50 |
| Høst | 0 | ** | ** | 0 | 60 |
| Kvæg afgræsning | 0 | 2 | 2 | 0 | * |
| Græsslet til hø | 0 | 100 | 100 | 0 | 60 |
| Græsslet til ensilage | 0 | 100 | 100 | 0 | 60 |
| Halmsnitning | 0 | ** | ** | 0 | 60 |
| Høvending | 0 | 100 | 100 | 0 | 0 |
| Halmpresning | 0 | 0 | 0 | 0 | 0 |
| Afbrænding af stub | 0 | ** | ** | 0 | 60 |
| Stubharvning | 0 | ** | ** | 0 | 75 |
De fleste typer jordbehandling (pløjning, harvning, tromling, såning, stubharvning) er af en så radikal karakter, at de må antages at være forbundet med 100 % dødelighed for æg og redeunger, der bliver udsat for sådanne begivenheder. Mange af disse markoperationer udføres imidlertid på tidspunkter i lærkens yngleforløb, som bevirker, at ingen eller kun meget få reder bliver berørt. Derimod foregår mekanisk ukrudtsbekæmpelse ofte i starten af lærkernes yngleperiode. Af særlig interesse er ukrudtsstriglinger, hvis effekter på højere dyr ikke er beskrevet i litteraturen. Bedømt ud fra resultaterne fra forsøgene med kunstige reder i nærværende undersøgelse er der en stor risiko for, at en rede med æg eller unger vil gå tabt, hvis den fysisk bliver udsat for en eller flere striglinger.
Den samlede effekt af strigling i en markafgrøde vil afhænge af antallet af striglinger, der udføres, deres indbyrdes tidsmæssige placering og af antallet af reder m. æg/unger, der er i den pågældende afgrøde, på de tidspunkter striglingerne udføres. Striglingseffekten på æg og unger stiger med antallet af striglinger udført med tidsmæssige korte intervaller. For et succesfuldt yngleforsøg behøver sanglærken en tidsmæssig periode på mellem 25-28 dage fra det første æg er lagt, til ungerne har forladt reden. Bliver reden ødelagt i første eller andet yngleforsøg, vil en ny rede ofte blive påbegyndt indenfor 4-7 dage. Hvis der i en afgrøde udføres to eller flere striglinger med en indbyrdes tidsmæssig afstand på 8-10 dage, vil chancen for at et ynglepar får ødelagt to på hinanden følgende yngleforsøg indenfor en måned være markant. Hvis et yngleterritorium kun indeholder den ene (striglede) afgrøde, vil resultatet, som ovenfor nævnt ofte være, at et yngleterritorium opgives, da væksten af afgrøden i den mellemliggende periode gør den uegnet som ynglehabitat. Den samlede effekt på bedrifts- eller landskabsniveau vil derudover igen afhænge af yderligere en lang række biotiske og abiotiske faktorer.
Ændring i effektvariabler
Hvis der forligger data for effekten af en bestemt markoperation på f.eks. æg/unger/voksne af en modelart, er det meget let efterfølgende at ændre i disse, hvis det skulle være påkrævet. At fremskaffe sådanne data er som regel forbundet med et stort arbejde at fremskaffe, og i mange tilfælde eksisterer data ikke, og man må efter en ’rundspørge’ blandt specialister anvende kvalificerede gæt. I sådanne tilfælde kan det være nødvendigt at foretage en følsomhedsnalyse af en eller flere sådanne variabler.
1.7 Sædskifter og afgrøder
I modellen gennemløber hver enkelt mark det sædskifte, der er beskrevet og begrundet for forskellige bedriftstyper. I nærværende projekt er der kun opstillet ét konventionelt sædskifte (kvægbrug) baseret på de beslutninger, der blev taget og beskrevet i et tidligere notat til styregruppen (FØL.JORD 12/3/4). Udgangspunktet for det anvendte sædskifte er et sædskifte baseret på de faktiske forhold i området beskrevet under det tværfaglige forskningsprojekt ARLAS (Dalgaard et al. 2004). De modifikationer, der er foretaget i forhold til dette, er gjort for at kunne erstatte herbicidsprøjtningerne i de konventionelle afgrøder med striglinger (tidspunkter for striglingerne taget fra økologisk dyrkede ’søster afgrøder’). De enkelte marker på en bedrift har forskellige afgrøder som udgangspunkt i opstartsåret for modelleringerne. For at kunne simulere forekomsten af afgrøder i forhold til deres arealmæssige forekomst, er det muligt at opstille mangeårige model-sædskifter for den enkelte mark.
Vedvarende græsarealer udgør i scenarierne for kvægbrug 10 %. Dette svarer godt til den procentuelle andel i værkstedsområdet (Dalgaard et al. 2004).
Ændring i sædskiftemodeller
Ændringer sædskiftemodeller kan foretages hurtigt, hvis der ikke er bestemte ønsker om, at disse sædskifter skal være baseret på statistikker for specifikke bedriftstyper i et område, eller at der skal foreligge et grundlag baseret på de enkelte afgrøders specifikke arealandele. Hvis sådanne ønsker derimod foreligger, indebærer det en arbejdsindsats på flere uger afhængig af, om sædskifterne også arealmæssigt skal forbindes til den person der driver de enkelte marker.
Som i foregående afsnit er det for anskuelighedens skyld valgt i de følgende to afsnit at anvende eksempler fra sanglærkemodellen.
1.8 De individbaserede modeller
Udviklingen af individbaserede modeller har givet nye muligheder for realistisk at simulere og modellere dyre- og plantearters (Huston et al. 1988; Hogeweg & Hesper 1990; Grimm 1999; McGlade 1999) økologiske respons på ændrede livsbetingelser i et modellandskab, der (som det er tilfældet i nærværende undersøgelse) kan være en detaljeret kopi af et eksisterende landskab. De enkelte dyrearters biologi er bestemmende for kravene mht. arealskala og detaljeringsgrad og type af arealinformationer der er behov for. Arealskalaen for modelleringen af et større rovdyr vil eksempelvis betyde, at der arbejdes på et landskabsniveau, men ikke nødvendigvis at der er behov for den samme detaljeringsgrad af den arealinformation, der for eksempel ville være nødvendig ved modelleringen af en insektart, hvor arealskalaen typisk vil være på mark- eller bedriftsniveau.
Udgangspunktet for individbaserede modeller er, som navnet antyder, det enkelte individ, for hvilket der er defineret et artsspecifikt sæt af regler, der er bestemmende for dyrets adfærd (eks. sociale interaktioner, energetik, reproduktion, bevægelse og spredning) under tidsmæssigt og rumligt skiftende forhold. Overgangen fra ét sæt af adfærdsregler, der beskriver og styrer begivenhederne i en bestemt periode (”stadie”) af et individs livsforløb, til et nyt stadie er beskrevet af et andet regelsæt (”overgangsstadie”), som er karakteristisk for den givne art (Figur 4). En del af et sådant stadie – overgangs diagram for en hunsanglærke er illustreret i Figur 5. Et komplet sæt af indbyrdes afhængige stadier og overgange mellem disse beskriver den specifikke individ-baserede biologiske model for en pågælden art.
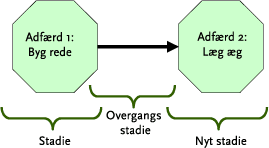
Figur 4. To stadier i en hunsanglærkes liv: hvert stadium og overgangen mellem disse er beskrevet af et bestemt sæt adfærdsregler, der bestemmer hunlærkens beslutninger under de givne omstændigheder.
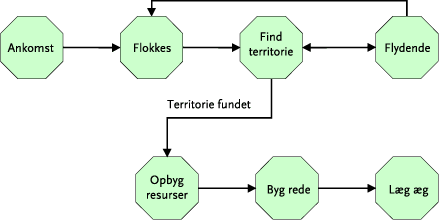
Figur 5. Brudstykke af et ”State/Transition-diagram” for hunsanglærken, der beskriver stadierne fra ankomst til ynglepladsen til påbegyndelse af æglægning. Pilene angiver overgangsstadier.
Modeltypen er karakteristisk ved, at alle individer til et givet tidspunkt i en given lokalitet på samme tid kan få alle relevante informationer om forskellige biotiske (f.eks. afgrødetype, -højde og dækningsgrad, tæthed af fødeemner, tilstedeværelsen af andre individer (f.eks. fra naboterritorier)) og abiotiske forhold (f.eks. pløjning, harvning, sprøjtning, temperatur og nedbør), der er betydende for de enkelte individers reaktioner og handlinger (Figur 6). På denne måde kan et antal individer modtage informationer om, og udnytte eksempelvis en lokal føderesource på en og samme tid. Den lokale population der modelleres, består således af indbyrdes ”kommunikerende” individer, der i tid og rum kan opfatte og reagere på forhold i det lokale miljø. For sanglærkens vedkommende afhænger eks. territoriestørrelsen dels af territoriets kvalitet, dels af den enkelte fugls styrkeforhold i forhold til andre territoriehævdende nabofugle, hvilket medfører en tæthedsafhængig modellering af populationen. De biologiske modeller består af en række undermodeller, der hver især beskriver de regelsæt, der karakteriserer de specielle forhold, der gør sig gældende for eks. hanlærker, hunlærker, ægudvikling, udvikling af redeunger og udvikling af ikke flyvefærdige unger.
En samlet oversigt over den komplekse struktur og sammenhæng mellem den overordnede models delelementer er vist for sanglærken på Figur 6.
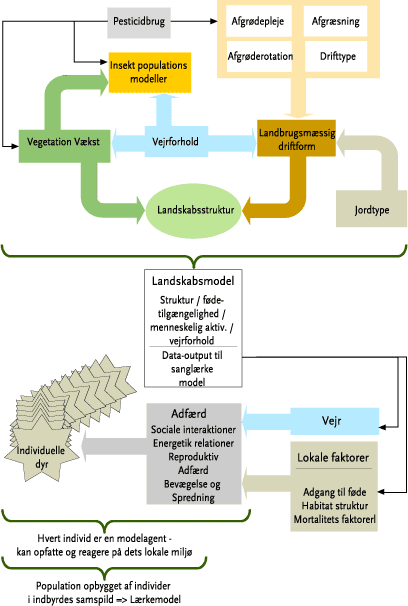
Figur 6. Diagrammatisk fremstilling af de faktorer, der alle påvirker de individuelle dyr, der modelleres i sanglærkemodellen.
1.9 Parameterisering
For at kunne vurdere modellens evne til at ”forudsige” lokale bestandstætheder under givne dyrkningsforhold, blev der for sanglærkens vedkommende i 16 delområder (af ca. 40 ha) indenfor et 10 x 10 km stort område ved Bjerringbro udført moniteringer af lærketætheder (lokalisering af territorier, udparrede hanner mm.) i 1998, 1999 og 2000 (Topping & Odderskær 2000). Efterfølgende er der ved at sammenholde modellens ”forudsigelser” af antal og placering af lærketerritorier og reder i de 16 delområder, hvori der gennem moniteringer forelå eksakte oplysninger om lokale bestandstætheder (herunder territorieplaceringer) i forhold til fysiske parametre (f.eks. tilstedeværelsen af læhegn, markskel, bygninger mm. – Figur 7), foretaget en kontrol og efterfølgende ”justering” af centrale biologiske modelparametre, indtil der blev opnået en generel god overensstemmelse mellem modelforudsigelser og reelle forhold. Modellens ’forudsigelser’ og de faktisk registrerede lærketerritorier i de 16 delområder viste en god overensstemmelse (Odderskær 2005).
Efter den foretagne justering af modeldelen der vedrører rugetid (notat fremsendt til styregruppemødet for nærværende projekt d. 14/11 2005: ’Notat vedrørende justering af modeldel for sanglærkens udrugningstid’) er der gennem de foretagne modelvalideringer af modelsystemet, konstateret en god overensstemmelse mellem faktiske og modellerede forhold.
Ændring i variabler, der indgår i dyremodeller
Genereringen af nye artsmodeller er den selvstændige del, der tager længst tid at udføre. I gennemsnit skal der regnes med ca. 1 års arbejde før en ny model er ’køreklar’. Efter en ny artsmodel er taget i anvendelse, vil der som regel med tiden blive behov for at justere på delelementer af modellerne, alene i kraft af at der med tiden dukker ny viden op på områder, hvor der tidligere kun har foreligget sparsomme eller ingen data. På dette punkt er forholdene ens med de betingelser, der eksistere for andre modelsystemer – der vil med tiden være behov for en løbende opdatering.
Der kan ikke siges noget konkret om, hvor vanskeligt det er at udføre specifikke ændringer i de enkelte modeller. Dette afhænger meget af, hvilken art (simpel/kompliceret model) der er tale om, og hvilke ændringer der er tale om, men det er erfaringen, at der fra det øjeblik det vides, hvordan en konkret ændring skal foretages, vil være behov for mellem en til to ugers arbejde, da der under alle omstændigheder efterfølgende skal foretages en række ’testsimuleringer’ for at sikre, at øvrige centrale variabler i modellen ikke har ændret sig.
Figur 7. Eksempel på modellens indplacering af lærkepar og reder (blå og røde prikker) i et udsnit af værkstedsområdet. Forskellige tætheder i udvalgte delområder er angivet med pile.
2 Litteratturliste
Bilde, T. & C.J. Topping (2004). Life-history traits interact with landscape composition to influence population dynamics of a terrestrial arthropod: a simulation study. EcoScience 11(1): 64-73.
Busche, G. (1989). Drastische Bestandseinbußen der Feldlerche Alauda arvensis auf Grünlandflächen in Schlewig-Holstein. Die Vogelwelt, 2: 51-58.
Dalgaard, T., Kjeldsen, C., Rasmussen, B.M., Fredshavn, J.R., Munier, B., Schou, J.S., Dahl, M., Wiborg, I.A., Nørmark, P. & J.F. Hansen (2004). ARLAS’ scenariesystem – Et grundlag for helhedsorienterede konsekvensvurderinger af ændringer i arealanvendelsen og landbrugsproduktionen, pp. 97-119, i DJF rapport Markbrug nr. 110.
Daunicht, W.D. (1998). Zum einfluss der Feinstruktur in der Vegatation suf die Habitatwahl, Habitatnutzung, Siedlungsdichte und populationsdynamik von Feldlerchen (Alauda arvensis) in grossparzelligem Ackerland. PhD thesis, University of Bern.
Grimm, V. (1999) Ten years of individual based modelling in ecology: what have we learned and what could we learn in the future. Ecological Modelling 115, 129-148. Elsevier.
Hogeweg, P. & B. Hesper (1990). Individual-oriented Modelling in Ecology, 13 (6): 83-90.
Huston, M., DeAngellis, D, & W. Post(1988). New Computer models Unify Ecological Theory. BioScience, 38 (10).
Jenny, M. (1990). Territorialität und Brutbiologie der Feldlerche Aluda arvensis in einer intensiv genutzten Agrarlandschaft. Journal für Ornithologie, 131: 241-265.
McGlade, J.M. (1999). Induvidual-based models in ecology. In: Advanced Ecological Theory, Principles and Applications (ed. Jacqueline McGlade). Blackwell Science.
Olesen, J.E. & T. Heidmann (1990). EVACROP. Et program til tilberedning af aktuel fordampning og afstrømning fra rodzonen. Version 1.00. AJMET Arbejdsnotat nr. 9.
Odderskær P., Prang A, Poulsen J.G., Elmegaard, N. & P.N. Andersen (1997). Skylark Reproduction in Pesticide Treated and Untreated Fields. Pesticide Research No 32. Ministy Of Environment and Energy, Denmark. Miljø- og Energiministeriet Miljøstyrelsen.
Odderskær, P., Lilleør, O. & Dietz, H.D. (1997). Den vilde faunas forekomst i forbindelse med et udendørs svinehold. Slutrapport for delprojekt ”Den vilde fauna” under ”Nærmiljø og produktionssystemers indflydelse på sundhed i udendørs svineproduktion”, Strukturdirektoratet, Ministeriet for Fødevarer, Landbrug og Fiskeri.
Odderskær, P. (1998). Undersøgelser af sanglærkers levevilkår i agerlandet. Statusrapport for delprojekt Samfunds- og miljømæssige konsekvenser af forskellige strategier for udvikling og udbredelse af økologiske jordbrugssystemer under Det Strategiske Miljøforskningsprogram, Strukturdirektoratet.
Odderskær, P., Cracknell, G. & T. Kroier (2001). Sanglærker og økologisk jordbrug – undersøgelser på Ny Ryomgård 199-1998. Posters til besøgslandbrug Ny Ryomgård.
Odderskær, P. (2005). Notat vedrørende justering af modeldel for sanglærkens udrugningstid – ’Ukrudtsstriglingens effekter på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug’. Notat til styregruppen for ’Ukrudtsstriglingens effekter på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug’ – 31/10/2005.
Plauborg, F. & J.E. Olesen (1991). Udvikling og validering af modellen MarkVand til vandingsstyring i landbruget. Tidsskrift for Plantealvs Specialserie, S 2113.
Poulsen, J.G., Sotherton, N.W. & N.J. Aebischer (1998). Comparative nesting and feeding ecology of skylarks Alauda arvensis on arable farmland in southern England with special reference to set-aside. Journal of Applied Ecology, 35: 131-147.
Sunderland, K.D. & C.J. Topping (1993). The spatial dynamicks of linyphiid spiders in winterwheat. Memoires of the Queensland Museum, 33: 639-644.
Thorbek, P. & Topping, C.J. (2005). The influence of landscape diversity and heterogeneity on spatial dynamics of agrobiont linyphiid spiders: an individual-based model. BioControl 50: 1-33.
Topping, C.J. & K.D. Sunderland (1998). Population dynamics and dispersal of Lepthyphantes tenuis in an ephemeral habitat. Entomologia Experimentalis et Applicata, 87: 29-41.
Topping, C. & P. Odderskær (2000) Udvikling af faunamodeller og landskabsmodel. Statusrapport for delprojekt Arealanvendelse og landskabsudvikling belyst ved scenariestudier – vekselvirkninger mellem natur, jordbrug, miljø og arealforvaltning (ARLAS) under forskningsprogrammet Arealanvendelse – jordbrugeren som lansskabsforvalter, Direktoratet for FødevareErhverv – Forskningskontoret.
Topping C.J. & Odderskær, P. (2004). Modelling the influence of temporal and spatial factors on the assessment of impacts of pesticides on skylarks. Environmental Toxicology & Chemistry 23, 509-520.
Wakeham-Dawson, A., Scoszkiewitz K., Stern K. & N.J. Aebischer (1998). Breeding Skylarks Alauda arvensis on Environmentally Sensitive Area arable reversion grass in southern England: survey-based and experimental determination of density. Journal of Applied Ecology, 35, 635-648.
Bilag B
1 Vegetation growth simulation in ALMaSS 4.0
Chris Topping¹ & Jørgen E. Olesen²
¹ Department of Wildlife Ecology and Biodiversity, National Environmental Research Institute, Grenåvej 12, DK-8410 Rønde, Denmark.
² Danish Institute of Agricultural Sciences, Dept. of Agroecology, Research Centre Foulum, P.O. Box 50,DK-8830 Tjele, Denmark.
This document describes the implementation of vegetation growth models in ALMaSS. These models have the aim of providing information on vegetation biomass and height for use by the animal model for which the system is designed.
Vegetation biomass and height are updated on a daily basis and independently for each vegetated element in the ALMaSS landscape. Biological systems are based primarily on photosynthesis, and are thus dependent on incoming radiation. However, the potential for production set by the radiation is greatly modified by temperature. Hence both incoming radiation and temperature were chosen as the driving factors for the vegetation models. Although also important, the current version of ALMaSS does not incorporate rainfall as a driving factor for vegetation growth, but assumes sufficient water availability. The main effect of temperature on plant growth is to control the duration of the period when growth is possible in each year (Olesen and Bindi, 2002), but other processes linked with the accumulation of dry matter (leaf area expansion, photosynthesis, respiration etc.) are also directly affected by temperature.
These effects are summarised in the following equation:
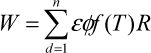
where W is accumulated dry matter (g/m²), ε is radiation use efficiency (g/MJ), φ is fraction of incoming light intercepted by crop canopy, ƒ(T) is effect of temperature T (°C) on dry matter growth, and R is incoming global radiation (MJ/m²).
The maximum radiation use efficiency (ε) depends on the species. However, a general distinction may be made only between C3 and C4 species (Figure 1). For C3 species (most Danish species) ε can be generally estimated to be 1.4 g/MJ and for C4 species (in our case maize) 1.7 g/MJ should be used (Sinclair and Muchow, 2000). The temperature response ƒ(T) on DM growth is also different for the different species as illustrated in Figure 1.
Figure 1. The effect of temperature on radiation use efficiencies in c3 & c4 crops (Preening de Vries eta al, 1989; Sinclair & Muchow, 2000).
However, the use of ε of 1.4 for C3 species is too great a simplification. Olesen et al. (2004) used crop specific constants for light saturated photosynthesis per unit leaf area depending on leaf thickness (specific leaf area). These specific leaf areas increased in the order: grass < barley < wheat < rape. Assuming that these figures relate to light use efficiencies (RUE), the following RUE values would be applied to the C3 crops: 1.1 g/MJ for grass, 1.2 g/MJ for barley, 1.4 g/MJ for wheat, and 1.5 g/MJ for rape.
| Vegetation Type | c |
| Natural Grass | A |
| Set-aside | A |
| Young Forest | A |
| Carrots | A |
| Grass /Clover | A |
| Permanent Grass | A |
| Permanent Set-aside | A |
| Grass for seed | A |
| Field Peas | B |
| Beets (fodder) | B |
| Oats | B |
| Potatoes | B |
| Spring Barley | B |
| Winter Barley | B |
| Winter Rye | B |
| Spring Wheat | C |
| Triticale | C |
| Winter Wheat | C |
| Oil Seed Rape | D |
Table 1. The basic crop types currently used in ALMaSS and the constant applied to modify their light use efficiencies (see text for constant values)
In the model implementation this is achieved by using the standard C3 values, but multiplying by a constant ( c ). c may have one of four values, which scale the required RUE relative to 1.4:
A = 0.786; B = 0.857; C = 1.000; D = 1.071.
Crops other than the four crops above were allocated values based on the closest crop type (Table 1).
The value for ε is modified by ƒ(T) for C3 and C4 species following Fig. 1, using the mean daily temperature as the value T. The resultant temperature related value for RUE is then effectively scaled by multiplication by c.
The fraction of intercepted light can be estimated from the green leaf area index (L) as:
φ = 1–exp(–kL)
where k is the extinction coefficient, which in general may be set to 0.4. L was calculated for 40 different crop or crop variants as being dependent upon day degrees from either a certain time of year, or a certain farming event. The relationships between leaf-area index for green and total leaf area (LAI-green & LAI-total) and height with day-degrees are listed in CropCurves.pdf, and are based on Olesen and Heidmann (1990) and Plauborg and Olesen (1991) together with unpublished data. Critical times of year were considered to be 1st January and 1st March. These were chosen to represent the start of a new year, and the time before which crop growth would not occur. Critical events were sowing, and harvest events, both of which will obviously alter the growth pattern of the crop. Hence following a critical event or date, the day degrees sum is reset to zero, and often the values for LAI-total, LAI-green and height are reset to some pre-determined value (e.g. 10 cm height after harvest). This is denoted by a ‘-1’ in the day-degrees sum in CropCurves.pdf. The curves for LAI-total, -green, and height are presented as a set of linearized curves and can therefore be described as a set of fixed points joined by straight lines with a particular slope denoting the rate of change in leaf area and height with day degrees. Implementation in ALMaSS is by determining the rate of change in response variable (leaf area or height) with day-degrees and using this to control the leaf-area and height on a daily basis as the crop grows within the model. As day-degrees are summed the rate of change is read from the curve until a fixed point is passed, when a new rate of change will be used. Thus, as growth proceeds, the day degrees are summed following each critical point, and the correct rate of change selected depending on last fixed point that is met. If the current day’s addition to the day-degrees sum overshoots the next fixed point, then the model calculates growth up to the next fixed point, then using the remaining day-degrees, calculates growth from that point. The result of this procedure is for height and leaf-area for all crops in ALMaSS to quite accurately match the input curves, but be dependent upon the temperature information in the weather data used to determine at what date a certain point will be reached. As a result, if a crop is sown too late or if a weather set is used that results is very slow growth, then it likely that, as in reality, the crop will not reach its mature height or leaf area.
To simulate the effect of altered fertiliser regimes on the gain in biomass another constant is used ( p ). This constant takes four values: 1.00, the default value; 0.8 for low fertilizer levels (all non-grass organic crops); 0.67 for ex-arable land (set-aside and young forests); and 0.5 for non-farm habitats (natural grass).
The radiation R can also be taken as the measured daily global radiation, but since this information is rarely available, an alternative is to use the normal daily global radiation, which can be approximated by a Fourier series (Olesen, 1991):
Rd = b0 + b1cos(ωd) + b2sin(ωd) + b3cos(2ωd) + b4sin(2ωd)
where Rd is global radiation on day number d in the year, ω = 2π/365, and b0 to b4 are constants. For the climate station Ødum, from where the weather data normally used for ALMaSS simulations for Denmark comes, these constants have been estimated for the period 1961 to 1990 as b0 = 9.478, b1 = -9.221, b2 = 1.458, b3 = 0.653 and b4 = -0.421.
1.1 Determination of model curves for natural grass vegetation
Growth of non-crop grassland was based on data from Al-Mufti et al (1977). This work provides seasonal variation in dry weight of live shoot material and dead litter from three grassland sites in Yorkshire, UK. These data were pooled and mean figure estimated for each month (Figure 2).
To recreate this curve it was necessary to estimate the LAI total and LAI green required to generate the seasonal pattern required. For LAI total, this was achieved by assuming a maximum LAI total of 5.5, corresponding to 550 g/m², and scaling this so that at minimum dead biomass the LAI-total was 3.85 corresponding to 385 g/m² ( Figure 2 ). LAI-green was then fitted by trial and error until the resulting pattern of seasonal biomass changes matched Figure 2. Fitting was carried out using an 11-year weather data series from Ødum in Denmark that spanned 1989-1999 inclusive. The mean biomass curve over the 11 years that resulted fitted the target value for all months within 5%. The resulting LAI-total and LAI-green values are listed in Table 2.
Figure 2. Seasonal changes in live and dead biomass as a mean response from three UK grasslands (estimated from Al-Mufti et al, 1977).
| TSum | LAI-Total | LAI-Green | Height | |
| From Jan 1st | 0 | 4.83 | 0 | 70 |
| 105 | 3.85 | 0 | 56 | |
| From Mar 1st | 0 | 3.85 | 0 | 56 |
| 289 | 4.5 | 1 | 65 | |
| 695 | 5 | 1.25 | 73 | |
| 1017 | 5.5 | 0 | 80 | |
| 2619 | 5.5 | 0 | 80 | |
| 2787 | 4.83 | 0 | 70 | |
| 99999 | 4.83 | 0 | 70 | |
| After Cutting | 0 | 0.3 | 0 | 10 |
| 289 | 4.5 | 1 | 65 | |
| 695 | 5 | 1.25 | 73 | |
| 1017 | 5.5 | 0 | 80 | |
| 2619 | 5.5 | 0 | 80 | |
| 2787 | 4.83 | 0 | 70 | |
| 99999 | 4.83 | 0 | 70 |
Table 2. The values used in ALMaSS to predict vegetation growth for non-crop grasslands. Tsum is the number of day degrees experienced. LAI-Total is a measure of the total leaf-area index, and LAI green is a measure of the available photosynthetic surface. Height indicates the height in cm achieved for the day-degrees experienced.
2 References
Al-Mufti, M.M., Sydes, C.L., Furness, S.B., Grime, J.P. & , S.R. Band (1977). A quantitative analysis of shoot phenology and dominance in herbaceous vegetation. J. Ecol. 65: 759-791.
Olesen, J.E. (1991). Jordbrugsmeteorologisk årsoversigt 1990. Statens Planteavlsforsøg, beretning nr. S2130.
Olesen, J.E. & M. Bindi (2002). Consequences of climate change for European agricultural productivity, land use and policy. European Journal of Agronomy 16, 239-262.
Olesen, J.E. & T. Heidmann (1990). EVACROP. Et program til beregning af aktuel fordampning og afstrømning fra rodzonen. Version 1.00. AJMET Arbejdsnotat nr. 9.
Olesen, J.E., Rubæk, G., Heidmann, T., Hansen, S. & C.D. Børgesen (2004). Effect of climate change on greenhouse gas emission from arable crop rotations. Nutrient Cycling in Agroecosystems 70, 147-160.
Penning de Vries, F.W.T., Jansen, D.M., ten Berge, H.F.M. & A. Bakema (1989). Simulation of ecophysiological processes of growth in several annual crops. Simulation Monographs. Pudoc.
Plauborg, F. & J.E. Olesen (1991). Udvikling og validering af modellen MarkVand til vandingsstyring i landbruget. Tidsskrift for Planteavls Specialserie S2113.
Sinclair, T.R. & R.C. Muchow (2000). Radiation use efficiency. Advances in Agronomy 65, 215-265.
Bilag C
1 Validation procedures for ALMaSS, and other complex adaptive systems
ALMaSS is an agent-based model (ABM), or complex adaptive system (CAS). It is an entirely different class of model to traditional population models which are usually based upon simple equations and relationships. Agent-based complex systems such as ALMaSS are dynamic networks of many interacting entities mimicking the processes and interactions we see in the real world. Whilst unlike statistical modelling there is no general framework for designing, testing, and analyzing bottom-up models as yet established, recent advances in ecological modelling have come together in a general strategy called pattern-oriented modelling.
What is pattern-oriented modelling? The patterns refer to system properties that we see around us, e.g. population growth curves, spatial distribution of organisms, individual developmental rates, or any other measurable characteristic of a system resulting from the interaction of agents. Patterns are therefore the defining characteristics of the modelled system.
The test of whether a CAS model is sufficient to be able to explain the cause of dynamics of the system is whether it can generate enough sufficiently accurate patterns. If it can do this then the mechanisms that are used to build up the model are considered to be sufficient to generate system behaviour akin to that seen in the real world system. In other words if enough of the basic mechanisms are incorporated so that the model system responds to changes like the real world system, then sufficient confidence can be generated in the model to consider its use as a predictive tool.
There are two kinds of pattern that must be evaluated, basic patterns and complex patterns. The complex patterns are what are termed ‘emergent properties’ and are the product of interactions between agents in the model. The basic patterns are features of the behaviour of the model that are directly programmed in. Hence checking these requires a range of trivial although laborious tests.
Checking the basic patterns are more or less checks on the system to see that all is functioning as intended. In ALMaSS this includes checks on weather input, landscape structure, vegetation growth, crop allocation, and not least crop or other habitat management. All these basic model parts need to be checked to see that they are producing the correct patterns. The same process is also carried out for the animal models. All kinds of individual behaviours need to be checked to see if they function correctly. These kinds of behaviours are those that are part of the processes directly programmed into the model and are not emergent properties. These basic checks can be thought of as a complex debugging process, i.e. afterwards the individual sub-components of the model should behave according to expectations (e.g. a farmer should manage his crop following the plan provided).
Once all the basic patterns are correct the interactions between these results in emergent properties, i.e. complex patterns that are not directly programmed responses. Relatively few complex patterns are required for successful analysis since they are a function of a great many model components or mechanisms. Probably the simplest and first of these is the plausibility criteria. In ALMaSS where we are simulating animal species this usually consists of an ecologist’s evaluation of the behaviour of the model animals, i.e. do they behave like the real thing? Subsequently numerical comparisons with observed patterns are utilized. These patterns, such as changes in animal population numbers with time, are a result of integrating all factors affecting growth, mortality, reproduction and dispersal across spatially and temporally heterogeneous landscapes affected by agricultural management.
If the model can predict a number of emergent patterns simultaneously from the same set of inputs (e.g. spatial distribution of animals at the same time as their developmental rates), then confidence in the model grows. As we increase the numbers of patterns that are used to compare model outputs to, we reduce the set of potential input parameter values (parameter space) that can be used. Why is this? The reason is that each new pattern that is added requires a certain specific set of parameter values to achieve a good match, as new patterns are used these new values much be selected from the set of values used to match the preceding patterns. The result is therefore an ever-dwindling range of possible parameter values that can achieve a match to all patterns; hence adding further patterns adds further limitations to the inputs. If sufficient real world data were available to add sufficient new patterns, then the continual reduction in input parameter space will eventually lead to a situation where deviation from a distinct set of parameters leads to failure to simulate all patterns. At this point the model cannot be improved and not only is the model structure (interactions between agents) well tested, but the inputs are also narrowed down, reducing uncertainty and providing a powerful predictive capability.
Bilag D
1 Model til beregning af fossilt energiforbrug i jordbrugssystemer med og uden mekanisk ukrudtsbekæmpelse
Tommy Dalgaard og Randi Dalgaard
Danmarks JordbrugsForskning, Forskningscenter Foulum
1.1 Dokumentation af energiforbrugsmodellen
I dette bilag beskrives energiforbrugsmodellen som anvendes i forskningsprojektet ”Ukrudtsstriglingens effekt på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug”, under Miljøstyrelsens Pesticidforskningsprogram 2003-2006. Formålet er at: 1) Dokumentere energiforbrugsmodellen og dens parameterisering, 2) Definere typeafgrøder og typebedrifter, som kan anvendes ved senere konsekvensanalyser af scenarier for ændret energiforbrug ved mekanisk ukrudtsbekæmpelse, og 3) Diskutere de mulige konsekvensanalyser af ændret energiforbrug ved mekanisk ukrudtsbekæmpelse, samt forudsætningerne for modelberegningerne ved en gennemgang af litteraturen.
Modellen er afgrænset til opgørelser af energiinput og -output i den primære landbrugsdrift. Dvs. det energiinput, der medgår til forarbejdning og transport, efter produkterne har forladt landbrugsbedrifterne, medregnes ikke, mens den energi, der går til at fremstille input, som anvendes på bedriften, samt den energi der går til indengårds transport, medregnes.
I lighed med metoderne i Refsgaard et al. (1998) og Dalgaard et al. (2001) opgøres energiforbruget ved hjælp af nøgletal for energiforbrug ved forskellige operationer og ressourceforbrug.
I Tabel 2 er vist nøgletal for brændstofforbrug ifølge litteraturen. Der er opdelt i fem kategorier af operationer, hvorunder normer (D) for enkelt-operationer er valgt ud fra litteraturen suppleret med egne målinger af brændstofforbrug på private gårde, i et tidligere forskningsprojekt. Normerne for harvning, jordbearbejdning og såning kan korrigeres for jordtypen ved på sandede jorde at multiplicere med en faktor 0,9 og på lerede jorde ved at multiplicere med en faktor 1,1 (Dalgaard et al. 2001).
Tabel 3 og Tabel 4 viser nøgletal for øvrige energiforbrugsposter i henholdsvis afgrøde- og husdyrproduktionen. Det bør bemærkes, at specielt nøgletallene for indirekte energiforbrug er behæftet med stor usikkerhed.
Tabel 2. Oversigt over nøgletal/normer (D) for dieselforbrug ved enkeltoperationer i afgrødeproduktionen. Lav-høj er de laveste og højeste værdier for dieselforbrug ved den enkelte operation fundet hhv. i litteraturen (se referencer i tabellen), eller målt på private kvæggårde ifølge Dalgaard 1996 (N er antal målinger bag hver af de viste gennemsnit, Gns.). De viste normer (D) er ikke korrigeret for jordtype.
| D | Litteraturen | Målt dieselforbrug | |||||
| Enhed | norm | Lav-høj | Referencea | Gns. | Lav-høj | N | |
| Jordbearbejdning og såning | |||||||
| Pløjning (21 cm), forår | L/ha | 20.0 | 8.4-32.7 | 1,2,4 | 17 | 12.0-22.0 | 9 |
| Pløjning (21 cm), efterår | L/ha | 23.0 | 8.4-32.7 | 1,2,3,4,5,6 | 22 | 15.0-27.0 | 16 |
| Jordpakning | L/ha | 2.0 | 1.8 | 1,2 | |||
| Såbedsharvning, let | L/ha | 4.0 | 2.2-4.7 | 1,2,4,5 | |||
| Såbedsharvning, tung | L/ha | 6.0 | 4.9-16.8 | 1,2,4,5 | 6.2 | 4.9-7.1 | 3 |
| Tromling | L/ha | 2.0 | 1.8 | 1 | 1.6 | 1 | |
| Såning | L/ha | 3.0 | 0.9-21.6 | 2, 4 | 3.2 | 3.0-3.4 | 2 |
| Stubharvning | L/ha | 7.0 | 2.8-30.9 | 3,4 | 7.3 | 4.0-18.0 | 7 |
| Gødskning og kalkning | |||||||
| Spredning & læsning, fast gødning | L/t | 0.6 | 0.4-1.8 | 5,8 | 0.6 | 0.5-0.7 | 4 |
| Spredning, gylle/ajle | L/t | 0.3 | 0.2-1.1 | 3,5 | 0.5 | 0.3-3.7 | 7 |
| Spredning, handelsgødning | L/ha | 2.0 | 0.9-4.7 | 3,4,5,8 | 1.9 | 1 | |
| Kalkning | L/ha/år | 1.5 | |||||
| Plantebeskyttelse | |||||||
| Sprøjtning | L/ha | 1.5 | 0.8-1.7 | 3,4,5 | 1.2 | 1.1-1.4 | 2 |
| Ukrudtsharvning | L/ha | 2.0 | 1.5-2.4 | 3 | |||
| Kvikharvning (se stubharvning) | |||||||
| Radrensning | L/ha | 3.0 | 3.0-4.9 | 3,5 | |||
| Høst og presning | |||||||
| Mejetærskning | L/ha | 14.0 | 7.0-19.0 | 3,4,5,8 | 14 | 11.0-19.0 | 20 |
| Roeoptagning | L/ha | 17.0 | 8.4-22.0 | 4,5,10-13 | 13 | 1 | |
| Roeaftopning | L/ha | 10.0 | 7.8-21.0 | 4,8,11,12,13 | 10 | 7.4-13.0 | 4 |
| Græsslåning | L/ha | 5.0 | 5.3-10.4 | 5,11 | 8.0 | 5.0-27.0 | 2 |
| Højtrykspresning + håndtering | L/t | 1.5+0.5 | 1.3-1.7 | 2,5,8 | 1.6 | ||
| Afpudsning | L/t | 0.5 | 0.3-0.9 | 5,14,15 | |||
| Stængelbrydning | L/t | +0.2 | 10,11 | ||||
| Grønthøstning | L/t | 1.0 | 0.7-2.1 | 14,15,16 | 1.7 | 1.2-3.3 | 4 |
| Transport | |||||||
| Maskin-transport | L/km | 0.04 | 0.3-0.4b | 14 | 1.2b | 0.2-2.3b | 3 |
| Gødnings-/ foder-transport | L/t/km | 0.2 | 0.1-0.5 | 8,14 | 0.4b | 0.3-0.5b | 3 |
| Læsning og håndtering | |||||||
| Læsning | L/t | 0.3 | 0.1-0.5 | 8,14 | 0.3 | 0.2-0.3 | 5 |
| Læsning og håndtering | L/t | 0.5 | 0.3-1.1 | 14 | |||
| Udfodring | L/t | 0.3 | 0.1-0.4 | 14 | |||
| Øvrig håndtering | L/t | 0.5 | |||||
| Håndtering (total gennemsnit) | L/t | 1.3 | 0.3-3.8 | 14 | 1.6 | 1.1-2.1 | 41 |
a) 1: Nielsen 1987, 2: Nielsen 1989, 3: Nielsen and Sørensen 1994, 4: Stout et al. 1982, 5: McFate 1983, 6: Vitlox and Pletinckx 1989, 7: Brown1988, 8: Pick and Netik 1989, 10: White 1974, 11: Frith and Pomersberger 1974, 12: Pick 1984, 13: Nielsen 1985, 14: Nielsen 1991, 15: Cunney 1982. 16: Norén 1984.
b) Inklusiv læsning
Tabel 3. Oversigt over nøgletal for andet energibrug end diesel i afgrødeproduktionen. Lav-høj er det laveste og højeste normtal fundet i den refererede litteratur.
| Enhed | Norm | lav-høj | Referencea | |
| Smøremidler | MJ/(L diesel) | 3.6 | 3.6-5.7 | 1,2 |
| Maskiner | MJ/(L diesel) | 12 | 7,8,9 | |
| Markvanding | MJ/mm | 52 | 1,3 | |
| Korntørring | MJ/t/(%-point) | 50 | 4,5,6 | |
| Kvælstof handelsgødning | MJ/(kg N) | 50 | 43-75 | 3,4,8,11,12 |
| Fosfor handelsgødning | MJ/(kg P) | 12 | 12-17 | 4,8,11,12,14 |
| Kalium handelsgødning | MJ/(kg K) | 7.0 | 6.0-7.0 | 4,8,11,12,14 |
| Jordbrugskalk | MJ/t | 30 | 15 | |
| Herbicider | MJ/kg | 40b | 80-460c | 8,16,17,18 |
| Insecticider | MJ/kg | 40b | 58-580c | 8,16,17,18 |
| Fungicider | MJ/kg | 40b | 61-397c | 8,16,17,18 |
a) 1: Refsgaard 1992, 2: Dalgaard 1996, 3: Pimentel 1980, 4: Leach 1976, 5: Andersen et al. (1995), 6: Cunney 1982, 7: Sonesson 1993, 8: Kristensen 1995, 9: Bowers 1992, 11: Hjortshøj and Rasmussen 1977, 12: Bøckman et al 1991, 14: Åbyholm 1995, 15: Refsgaard 1992, 16: Stout et al. 1982, 17: Green 1987, 18: Fluck 1992.
b) per kg formuleret middel.
c) per kg aktivstof.
Tabel 4. Oversigt over nøgletal for energiforbrug i hhv. konventionel og økologisk kvæg- og svineproduktion. (1 DE= 1 Dyreenhed svarer 1 malkeko af stor race I 1 år eller 30 producerede slagtesvin. 1 FE= 1 foderenhed= 12 MJ metaboliserbar energi).
| Enhed | Norm | lav-høj | Referencea | |
| Operationer i stalden: | ||||
| for malkekvæg | GJ/DE | 8.0 | - | 2,7 |
| for andet kvæg | GJ/DE | 1.7 | - | 2,7 |
| for konventionelle søer | GJ/DE | 6.1 | - | 2,7 |
| for økologiske søer | GJ/DE | 3.2 | - | 2,7 |
| for konv. Slagtesvin | GJ/DE | 0.9 | - | 2,7 |
| for økologiske slagtesvin | GJ/DE | 0.4 | - | 2,7 |
| Opvarmning af stalde: | ||||
| for konventionelle søer | GJ/DE | 3.1b | - | 3,7 |
| for konv. Slagtesvin | GJ/DE | 0.6b | - | 3,7 |
| Bygninger, inventar etc. | GJ/DE | 2.5 | - | 5,6,7 |
| Importeret foder | MJ/FE | 5.7 | 1.4-7.7 | 1,4 |
| Egen-produceret foderc | MJ/FE | c | 0.2-3.7 | 7,8 |
a) 1: Refsgard et al. (1998), 2: The Danish Producers of Electricity (1994), 3: Søgaard (1999), 4: Cederberg (1998), 5: Kristensen (1995), 6: Refsgaard (1992), 7: Dalgaard et al. (1999), 8: Dalgaard et al (2001).
b) 80% af standard normen fordi ca. 20% af den konventionelle svineproduktion er uopvarmet. Økologisk svineproduktion antages 100% uopvarmet.
c) Beregnet for en række specifikke fodertyper i Dalgaard et al. 2001.
1.2 Definition af typebedrifter og typeafgrøder
De energimæssige konsekvenser af mekanisk ukrudtsbekæmpelse afhænger i høj grad bedriftstypen og af de ændringer i afgrødevalg og produktionsstrategi der følger med overgang til en ændret mekanisk ukrudtsbekæmpelsesstrategi. Det giver således ikke mening at opgøre energimæssige konsekvenser af mekanisk ukrudtsbekæmpelse isoleret set for den enkelte afgrøde eller på den enkelte mark, hvor den mekaniske ukrudtsbekæmpelse finder sted.
Derfor defineres en række typebedrifter, hvorpå konsekvenserne af mekanisk ukrudtsbekæmpelse kan illustreres. I tabel 5 er vist eksempler på en sådan bedriftstypologisering foretaget på baggrund af Fødevareøkonomisk Instituts Regnskabsstatistik (www.foi.dk).
Tabel 5. Eksempler på bedriftstyper, hvorpå energimæssige konsekvenser af mekanisk ukrudtsbekæmpelse kan opgøres.
| Brugstype | Jordtype | Husdyrtæthed (DE/ha) |
| Konventionelt malkekvægbrug, < 1,4 DE/ha | Sandjord | 1,0 |
| Konventionelt malkekvægbrug, 1,4-2,3 DE/ha | Sandjord | 1,7 |
| Konventionelt malkekvægbrug, > 2,3 DE/ha | Sandjord | 2,6 |
| Konventionelt malkekvægbrug, < 1,4 DE/ha | Lerjord | 0,9 |
| Konventionelt malkekvægbrug, 1,4-2,3 DE/ha | Lerjord | 1,2 |
| Konventionelt malkekvægbrug, > 2,3 DE/ha | Lerjord | 2,8 |
| Konventionelt svinebrug, <1,4 DE/ha | Sandjord | 0,8 |
| Konventionelt svinebrug, 1,4-1,7 DE/ha | Sandjord | 1,4 |
| Konventionelt svinebrug, >1,7 DE/ha | Sandjord | 2,5 |
| Konventionelt svinebrug, <1,4 DE/ha | Lerjord | 0,7 |
| Konventionelt svinebrug, 1,4-1,7 DE/ha | Lerjord | 1,4 |
| Konventionelt svinebrug, >1,7 DE/ha | Lerjord | 2,5 |
| Økologisk malkekvægbrug | Sandjord | 1,3 |
| Økologisk malkekvægbrug | Lerjord | 1,1 |
| Øvrige økologiske brug (ikke mælkebrug) | Sandjord | 0,3 |
| Øvrige økologiske brug (ikke mælkebrug) | Lerjord | 0,5 |
For hver af disse typebedrifter kendes afgrødersammensætningen, høstudbyttet af de enkelte afgrødetyper, husdyrproduktionen, handels- og husdyrgødningsforbruget mv.
På baggrund af disse oplysninger kan opstilles en model for energiforbruget. I tabel 6 er således vist en tilpasset model for norm-dieselforbruget ved forskellige markoperationer på lerjord og sandjord. Der skelnes mellem ”faste normer”, der er defineret ved antallet af markoperationer for en række typeafgrøder (Tabel 7 og Tabel 8), og ”variable normer”, der afhænger af den høstede mængde, eller mængden og typen af husdyrgødning, der ifølge ovenstående bedriftstypeoplysninger er udbragt på den aktuelle afgrødetype på den aktuelle bedriftstype.
Tabel 6. Norm dieselforbrug for markoperationer på lerjord og sandjord, og indikation af hvilke normer der er hhv. faste (jf. Tabel 7 og Tabel 8) eller variable for den enkelte afgrødetype og bedriftstype.
| Norm dieselforbrug | ||||
| Markoperation | Fast/variabel | Enhed | sandjord | lerjord |
| Pløjning (efterår) | F | L/ha | 20,7 | 25,3 |
| Pløjning (vår/vinter) | F | L/ha | 18 | 22 |
| Jordpakning | F | L/ha | 1,8 | 2,2 |
| Såbedsharvning | F | L/ha | 3,6 | 4,4 |
| Agerslæbning/tung såbedsharve | F | L/ha | 5,4 | 6,6 |
| Tromling | F | L/ha | 1,8 | 2,2 |
| Rotorsæt-såning | F | L/ha | 10,8 | 13,2 |
| Såning (radsåmaskine) | F | L/ha | 2,7 | 3,3 |
| Sætning/stenstrenglægning | F | L/ha | 8,1 | 9,9 |
| Rækkeafgrødeplantning | F | L/ha | 4,5 | 5,5 |
| Stubkultivering/tallerkenharve/fræsning | F | L/ha | 6,3 | 7,7 |
| Grubning | F | L/ha | 13,5 | 16,5 |
| Ton møg | V | L/ton | 0,55 | 0,55 |
| Ton gylle/ajle | V | L/ton | 0,3 | 0,3 |
| Bredspredning, kunstgødning | F | L/ha | 2 | 2 |
| Placering, kunstgødning | F | L/ha | 0,5 | 0,5 |
| Ammoniaknedfældning | F | L/ha | 7 | 7 |
| Kalkning | F | L/ha/år | 1,5 | 1,5 |
| Sprøjtning | F | L/ha | 1,5 | 1,5 |
| Ukrudtsharvning/strigling | F | L/ha | 2 | 2 |
| Radrensning | F | L/ha | 3 | 3 |
| Hypning | F | L/ha | 4 | 4 |
| Gasbrænding (incl. E til gas) | F | L/ha | 0 | 0 |
| Mejetærskning | F | L/ha | 14 | 14 |
| Halmsnitning | F | L/ha | 4 | 4 |
| Optagning (kartofler/roer) | F | L/ha | 17 | 17 |
| Aftopning af roer | F | L/ha | 10 | 10 |
| Sammenrivning | F | L/ha | 2 | 2 |
| Grønthøstning/afpudsning | F | L/ha | 5 | 5 |
| Højtrykspresning+læsning | V | L/Hkg | 0,2 | 0,2 |
| Wrapning | V | L/Hkg | 0 | 0 |
| Skårlægning | V | L/Hkg | 0,05 | 0,05 |
| Stængelbryder | V | L/Hkg | 0,02 | 0,02 |
| Finsnitning+friskfoderhøst | V | L/Hkg | 0,1 | 0,1 |
| Maskintransport | F | L/km | 0,04 | 0,04 |
| Gødningstransport | V | L/Hkg/km | 0,02 | 0,02 |
| Salgsafgrødetransport (korn+rod) | V | L/Hkg/km | 0,02 | 0,02 |
| Halmtransport | V | L/Hkg/km | 0,02 | 0,02 |
| Friskfodertransport | V | L/Hkg/km | 0,02 | 0,02 |
| Øvrig grovfodertransport | V | L/Hkg/km | 0,02 | 0,02 |
| Indlægning/læsning | V | L/hkg | 0,05 | 0,05 |
| Udfodring | V | L/hkg | 0,03 | 0,03 |
| Diverse kørsel | V | L/hkg | 0,05 | 0,05 |
Tabel 7. Oversigt over de antagelser der ligger bag de „faste“ normer for dieselforbrug for de enkelte afgrødetyper på konventionelle bedriftstyper i Tabel 5.
| Konventionelt dyrket | Vår- korn |
Vin- ter- korn |
Ær- ter |
Raps | Frø- græs |
Kar- tof- ler |
Ro- er |
Hel- sæd |
Grønt- afgrø- der |
Af- græs- nings- græs |
| Pløjning (efterår) | 1 | 1 | ||||||||
| Pløjning (vår/vinter) | 1 | 1 | 1 | 1 | 1 | |||||
| Jordpakning | 1 | 1 | 1 | 1 | 1 | |||||
| Såbedsharvning | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |||
| Agerslæbning/tung såbedsharve | 1 | 1 | 1 | 1 | 2 | 2 | 1 | |||
| Tromling | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Rotorsæt-såning | ||||||||||
| Såning (radsåmaskine) | 2 | 1 | 2 | 1 | 1 | 2 | ||||
| Sætning/stenstrenglægning | 2 | |||||||||
| Rækkeafgrødeplantning | ||||||||||
| Stubkultivering | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| Grubning | ||||||||||
| Bredspredning, kunstgødning | 2 | 2 | 2 | 3 | 3 | 2 | 2 | 3 | 5 | |
| Placering, kunstgødning | ||||||||||
| Ammoniaknedfældning | ||||||||||
| Kalkning | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Sprøjtning | 3 | 5 | 3 | 5 | 3 | 7 | 5 | 2 | 1 | 1 |
| Ukrudtsharvning/strigling | ||||||||||
| Radrensning | 2 | 1 | ||||||||
| Hypning | ||||||||||
| Gasbrænding (incl. E til gas) | ||||||||||
| Mejetærskning | 1 | 1 | 1 | 1 | ||||||
| Halmsnitning | 1 | 1 | 1 | |||||||
| Optagning (kartofler/roer) | 1 | 1 | ||||||||
| Aftopning af roer | 1 | 1 | ||||||||
| Sammenrivning | ||||||||||
| Grønthøstning/afpudsning | 1 | 2 | ||||||||
| Maskintransport (kilometer) | 42 | 46 | 42 | 48 | 34 | 58 | 48 | 40 | 24 | 30 |
Tabel 8. Oversigt over de antagelser der ligger bag de „faste“ normer for dieselforbrug for de enkelte afgrødetyper på de økologiske bedriftstyper i Tabel 5.
| Økologisk dyrket | Vår- korn |
Vin- ter- korn |
Ær- ter |
Raps | Frø- græs |
Kar- tof- ler |
Ro- er |
Hel- sæd |
Grønt- afgrø- der |
Af- græs- nings- græs |
| Pløjning (efterår) | 1 | 1 | ||||||||
| Pløjning (vår/vinter) | 1 | 1 | 1 | 1 | 1 | |||||
| Jordpakning | 1 | 1 | 1 | 1 | 1 | |||||
| Såbedsharvning | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |||
| Agerslæbning/tung såbedsharve | 1 | 1 | 1 | 1 | 2 | 2 | 1 | |||
| Tromling | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| Rotorsæt-såning | ||||||||||
| Såning (radsåmaskine) | 2 | 1 | 1 | 1 | 1 | 1 | ||||
| Sætning/stenstrenglægning | 2 | |||||||||
| Rækkeafgrødeplantning | ||||||||||
| Stubkultivering | 3 | 3 | 2 | 2 | 3 | 3 | ||||
| Grubning | ||||||||||
| Bredspredning, kunstgødning | ||||||||||
| Placering, kunstgødning | ||||||||||
| Ammoniaknedfældning | ||||||||||
| Kalkning | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Sprøjtning | ||||||||||
| Ukrudtsharvning/strigling | 1 | 3 | 4 | 3 | 2 | 2 | 3 | |||
| Radrensning | 3 | 3 | ||||||||
| Hypning | ||||||||||
| Gasbrænding (incl. E til gas) | ||||||||||
| Mejetærskning | 1 | 1 | 1 | 1 | ||||||
| Halmsnitning | 1 | |||||||||
| Optagning (kartofler/roer) | 1 | 1 | ||||||||
| Aftopning af roer | 1 | 1 | ||||||||
| Sammenrivning | ||||||||||
| Grønthøstning/afpudsning | 2 | 2 | ||||||||
| Maskintransport (kilometer) | 32 | 40 | 38 | 40 | 18 | 44 | 42 | 38 | 18 | 18 |
1.3 Diskussion
Det er hensigten at anvende ovenstående model til konsekvensanalyser af ændret energiforbrug ved mekanisk ukrudtsbekæmpelse.
I markforsøget sammenlignes det fossile energiforbrug ved produktion af vårhvede med ingen eller flere ukrudtsstriglinger med afgrødeudbyttet i form af halm og kerne. Derved kan bl.a. opstilles en energibalance hvor de marginale ændringer i det fossile energiforbrug ved ekstra striglinger sammenlignes med de marginale ændringer i energiproduktionen - dels i form af metaboliserbar energi (foderenheder), og dels i form af brændværdien i den marginale udbytteændring i kerne og halm (Pykh and Pykh 2002).
Desuden vurderes de energimæssige konsekvenser af mekanisk ukrudtsbekæmpelse for de forskellige afgrødetyper og bedriftstyper.
Opgørelserne kan sammenlignes med lignende opgørelser i litteraturen (fx LandboCentrum 2003), og DJF’s nyeste målinger af energiforbruget ved mekanisk ukrudtsbekæmpelse mv. (fx Nielsen et al., 2004).
2 Litteraturliste
Andersen, B., Birkkjær, K.O., Jørgensen, K., Laursen S.W., Mogensen, J., Nielsen, A., Nørgaard, E., Sørensen, P., 1995. Miljø- og ressourcestyring på landbrugsbedrifter -en håndbog (Environmental and ressource management on farms). Miljøgruppen på Landbrugets Rådgivningscenter. Landbrugets Informationskontor. Aarhus, Denmark . ISBN 87-7470-601-2. 69 s.
Boo, W. de, 1993. Environmental and Energy Aspects of Liquid Biofuels. Centrum voor energisparing en shone technologie. Delft, Holland.
Bowers, W., 1992. Agricultural Field Equipment. In: Fluck R.C. (ed). Energy in Farm Production. Elsevier, Amsterdam. ISBN: 0-444-88681-8. 10, 117-29.
Brown, R.H., 1988. Handbook of Engineering in Agriculture. Volume I: Crop Production Engineering. CRC Press. Boca Raton, Florida: ISBN 0-8493-3861-1. 261 pp.
Bøckman, O.C., Kaarstad, O., Lie, O.H., 1991. Landbruk og gjødsling. Mineralgjødsling i perspektiv (Agriculture and fertilisation. Mineral fertilisation in perspective). Norsk Hydro. Oslo. ISBN:82-90861-01-x. 248 pp.
Cederberg, C., 1998. Life Cycle Assessment of Milk Production. The Swedish Institute for Food and Biotechnology, Gothenburg. ISBN 91-7290-189-6. rep. 643. 86 pp.
Cleveland, C.J., 1995. The direct and indirect use of fossil fuels and electricity in USA agriculture, 1910-1990. Agric. Ecosys. and Environ. 55, 111-21.
Cunney, M.B., 1982. Energy aspects of harvesting and drying. In: Robinson, D.W., Mollan, R.C. (eds). Energy Management in Agriculture. Royal Dublin Society, Dublin. 11: p.143-154.
Dalgaard, T., 1996. Ressource Management of Fossil Energy in Crop Production. M.Sc. (Agric) Thesis. Royal Veterinary and Agricultural University, Copenhagen. 116 pp.
Dalgaard, T., Halberg, N. & Jørgensen, M.H., 2004. Status for energiinput og -output i økologisk jordbrug samt muligheder for energibesparelser. I: Jørgensen, U. & Dalgaard, T. (red.). Energi i økologisk jordbrug. Reduktion af fossilt energiforbrug og produktion af vedvarende energi. FØJO-rapport 19, 25-45.
Dalgaard T, Halberg N & Porter JR. 2001. A model for fossil energy use in Danish agriculture used to compare organic and conventional farming. Agriculture, Ecosystems & Environment 87(1) 51-65.
Dalgaard, T., Halberg, N., Fenger, J., 1999. Fossil-based energy use and emission of greenhouse gasses in three scenarios for conversion to 100% organic farming in Denmark. Danish Research Centre for Organic Farming (FØJO), Foulum. Report. 57 pp.
Danish Farmers Unions (“De Danske Landboforeninger”), 1998. Landøkonomisk Oversigt. (Agricultural Statistics). De Danske Landboforeninger, Copenhagen. ISSN 0107-7163. 127 pp.
Edwards, C.A., Grove, T.L., Harwood, R.R., Colfer, C.J.P., 1993. The role of agroecology and integrated farming systems in agricultural sustainability. Agric. Ecosyst. Environ. 46, 99-121.
Fluck, R.C., 1992. Energy in Farm Production. Elsevier. Amsterdam: ISBN 0-444-88681-8. 367pp.
Frith, R.R., Promersberger, W.J., 1974. Estimates of fuel consumption for farming and ranching operations under typical North Dacota conditions In: Nielsen, V. 1985. Energiforbruget ved håndtering og behandling af grovfoder (udredningsprojekt). Statens Jordbrugstekniske Forsøg. Horsens, Denmark. rep. 40. 35 pp.
Green, M., 1987. Energy in Pesticide Manufacture, Distribution and Use. In: Helsel, Z.R. (ed). Energy in Plant Nutrition and Pest Control. Elsevier, Amsterdam. ISBN 0-444-42753-8. 7: p.165-177.
Halberg, N. 1999. Indicators of ressource use and environmental impact for use in a decision aid for Danish livestock farmers. Agric. Ecosys. Environ. 76, 17-30.
Hitzhusen, F.J., 1993. Land degradation and sustainability of agricultural growth: some economic concepts and evidence from selected developing countries. Agric. Ecosys. Environ. 46, 69-79.
Hjortshøj, A., Rasmussen, S., 1977. En kortlægning af den primære jordbrugssektors energiforbrug (Mapping the energy use of the primary sector). Økonomisk Institut, Den Kongelige Veterinær- og Landbohøjskole. Copenhagen. rep.7. 119 pp.
Hulscher, W.S., 1991. Basic Energy Concepts. In: Energy for sustainable rural development projects. FAO, Rome. vol.1. p.5-26.
International Federation of Institutes for Advanced Study (IFIAS). 1974. Energy Analysis. Slesser, M. (eds.). International Federation of Institutes for Advanced Study. Stockholm. rap.nr.6. 89 pp.
Intergovernmental Panel on Climate Change (IPCC), 1997. Greenhouse Gas Inventory Reference Manual. Houghton J.T, et al. (eds.). Vol. 3. IPCC Technical Support Unit, London.
Kristensen, E.S.,1995. Forskning i økologisk landbrug ved Statens Husdyrbrugsforsøg (Research on organic farming at The Danish Institute of Animal Sciences). Statens Husdyrbrugsforsøg, Foulum, Denmark. Internal Report 42.105-112.
Kuemmel, B., Langer, V., Magid, J., de Neergaard, A., and Porter, J.R., 1998. Energetic, economic and ecological balances of a combined food and energy system. Biomass and Bioenergy 15 (4/5) 407-416.
LandboCentrum, 2003. Rapport over pesticidfri dyrkning 1998-2002.
Leach, G., 1976. Energy and Food Production. IPC Science and Technology Press Limited. Guilford, Surrey: 151 pp.
Madsen I., Petersen D. 1981. Hjemkundskab Grundbog (Cooking – a primer). 9.udg. Aschehoug Forlag, Copenhagen. ISBN 87-11-02770-3. 181 pp.
McFate, K.L., 1983. Farm Energy Analysis. A Programe aimed at fostering a team approach for implementing wise energy management and efficient energy use practices on American Farms. The National Food and Energy Council. Columbia, Missouri.
Ministry of Environment and Energy, 1995. Danmarks Energifremtider (Danish energy futures). The Danish Energy Agency, Copenhagen. 214 pp.
Mörschner, J., Bärbel, G., 1999. Direct and indirect energy use in arable farming - an example on winter wheat in Northern Germany. In: Weidema, B., Meeusen, M. (eds). Agricultural data for life cycle assessments. Proceedings of The Second European Invitational Expert Seminar on Life Cycle Assessments of Food Products, Den Haag 25th to 26th January 1999. Institute for Product Development, Lyngby, Denmark.
Nielsen, V., 1985. Energiforbruget ved håndtering og behandling af grovfoder (Energy use for management and treatment of roughage). Statens Jordbrugstekniske Forsøg. Horsens, Denmark. rep. 40. 35 pp.
Nielsen, V., 1987. Energiforbrug og arbejdsbehov ved direkte såning og traditionel jordbehandling (The need for energy and labour for direct sowing and conventional soil treatments). Statens Jordbrugstekniske Forsøg. Horsens, Denmark. rep.77. 35 pp.
Nielsen, V., 1989. Specific Fuel Consumption in European Countries. Denmark. In: Pick, E., Noren, O., Nielsen, V. (eds). Energy Consumption and Input-Output Relations of Field Operations. REUR Tecnical Series. Food and Agriculture Organization of the United Nations. Rome. rep.10. p.36-52.
Nielsen, V. 1991. Energiforbrug ved håndtering af græs (Energy use for management of grass). Statens Jordbrugstekniske Forsøg. Horsens, Denamrk. rep. 47. 86 pp.
Nielsen, V., Mortensen H., og Sørensen K., 2004. Brændstofforbrug, arbejdsforbrug og kapacitet ved reduceret jordbearbejdning og direkte såning. DJF rapport markbrug nr. 105.
Nielsen, V., Sørensen, C.G., 1994. "Drift". Et program til beregning af arbejdsbehov, arbejdskapacitet, arbejdsbudget og arbejdsprofil (A programme to calculate the need for labour). Statens Jordbrugstekninske Forsøg. Horsens, Denmark. rep. 53.124 pp.
Norén, O., 1984. Different methods of forage harvesting from an energy point of view. FAO-meeting, Bruxelles, February 1984. In: Nielsen, V. 1985. Energiforbruget ved håndtering og behandling af grovfoder (udredningsprojekt). Statens Jordbrugstekniske Forsøg. Horsens, Denmark. rep. 40. 35 pp.
Pick, E. (1984): Rationalisation of energy demands in field operations. FAO-meeting, Bruxelles, February 1984. In: Nielsen, V. 1985. Energiforbruget ved håndtering og behandling af grovfoder (udredningsprojekt). Statens Jordbrugstekniske Forsøg. Horsens, Denmark. rep. 40. 35 pp.
Pick, E. and Netik, O. (1989): Specific fuel consumption in European Countries. Czechoslovakia. In: Pick, E., Noren, O., Nielsen, V. (eds). Energy Consumption and Input-Output Relations of Field Operations. REUR Tecnical Series. Food and Agriculture Organization of the United Nations. Rome. rep.10.
Pimentel, D. 1980. Handbook of Energy Utilization in Agriculture. CRC Press. Boca Roton, Florida: ISBN:0-8493-2661-3. 475 pp.
Pimentel D., Pimentel M., 1996. Food, Energy and Society. ISBN 08-70-81386-2
Pimentel, D., Beradi, G., Fast, S., 1983. Energy efficiency of farming systems: organic and conventional agriculture. Agric. Ecosyst. Environ. 9, 359-372.
Pimentel, D., Brown, N., Vecchio, F., La Capra, V., Hausman, S., Lee, O., Diaz, A., Williams, J., Cooper, S., Newburger, E., 1992. Journ. Agric. Environ. Ethics 5(2), 113-146.
Pimentel, D., Hurd, L.E., Bellotti, A.C., Forster, M.J., Oka, I.N., Sholes, O.D., Whitman, R.J., 1973. Food Production and the Energy Crisis. Science. 182(4111), 443-449.
Plentinger, M.C., Penning de Vries, F.W.T., 1996. CAMASE. Register of Agro-ecosystems Models. Camase (Concerned Action for the development and testing of quantitative Methods for research on Agricultural Systems and the Environment). rep.II. Wageningen, Holland. 411 pp.
Pykh IG and Pykh YA., 2002. Sustainable Energy: Resources, Technology and Planning. WIT press. ISBN 1-85312-939-9. 351 p.
Refsgaard, K. 1992. Grundlag for beregning af energiindsatsen ved fremstilling af hjælpestoffer på primære landbrugsbedrifter (Tables for calculation of energy use on primary farms). Working paper, Danish Institute of Animal Sciences.
Refsgaard, K., Halberg N., Kristensen, E.S., 1998. Energy Utilization in Crop and Dairy Production in Organic and Conventional Livestock Production Systems. Agric. Systems 57(4) 599-630
Sensi, A., 1999. Agriculture and climate change. In: Agriculture, Environment, Rural development: Facts and Figures - A Challenge for Agriculture. EU Comission, DG VI, DG XI, and EUROSTAT. http://europa.eu.int/comm/dg06/envir/report/en/index.htm
Slesser, M.,1977. Energy Analysis. Science 196, 259-61.
Sonesson, U., 1993. Energianalyser av biobränslen från Höstvete, Raps och Salix (Energy analysis of biofuels from Winter wheat, Rape seed and Salix). Sveriges Lantbruksuniversitet. Uppsala, Sweden. rep. 174. 54 pp.
Soddy F. 1933. Welth, virtual welth and dept. Allen and Unwin, London. 57 pp.
Statistics Denmark (1996) Danish Agricultural Statistics. ISBN 87-501-1034-9. 310 pp.
Stout, B.A., Myers, C., Schwab, G., et al., (1982): Energy Management in US Production Agriculture. In: Robinson, D.W., Mollan, R.C. (eds). Energy Management and Agriculture. Royal Dublin Society, Dublin. 5: p.67-84.
Strudsholm, F., Nielsen, E.S., Flye, J.C. (1997) Fodermiddeltabel 1997 (fodder stuff table 1997). Danish Extension Service, Skejby. 69: 1-53.
Søgaard, J., 1999. Energiforbrug i landbruget (Agricultural energy use). Bygnings- og Maskinkontoret, Viborg, Denmark. Report, chapter 1, 7 pp.
The Danish Energy Agency. (1997) Energistatistik 1996. (Energy Statistics 1996). ISBN 87-7844-065-3. 36 pp.
The Danish Producers of Electricity (“De Danske Elværker”). 1994. Energirådgivning – Landbrug (Energy advise – agriculture). De Danske Elværkers Forening, Frederiksberg, Copenhagen. ISBN 87-87071-47-9. 225 pp.
The European Commission. 1997. Harmonisation of environmental life cycle assessment for agriculture. Audsley, E (eds.) Final Report Concerted Action AIR3-CT94-2028. European Commission DG VI Agriculture, Brussels.
Tsatsarelis, C.A. 1993. Energy inputs and outputs for soft winter wheat production in Greece. Agric. Ecosyst. Environ. 43, 109-118.
Tuthill Corporation. 1994. Fill-Rite Fluid Measurement Technology. Parts and Technical Service Guide for models 806B, 807B. Tuthill Corporation. Fort Wayne, Indiana, USA.
Uhlin, H. 1998. Why Energy Productivity is Increasing: An I-O Analysis of Swedish Agriculture. Agric. Ecosys. Environ. 56(4) 443-465.
Vester, J., 1995. Energibalancer (Energy balances). In: Olesen, J.E., Vester, J. (eds). Næringsstofbalancer og energiforbrug i økologisk jordbrug- fokus på kvægbedrifter og planteavl. Statens Planteavlsforsøg, Lyngby. rep. 8: p.106-116.
Vitlox, M., Pletinckx, 1989. Specific fuel consumption in European Countries: Belgium. In: Pick, E., Noren, O., Nielsen, V. (eds). Energy Consumption and Input-Output Relations of Field Operations. REUR Tecnical Series. Food and Agriculture Organization of the United Nations. Rome. rep.10. p.12-17.
Weidema, B., Meeusen, M. 1999. Agricultural data for life cycle assessments. Proceedings of The Second European Invitational Expert Seminar on Life Cycle Assessments of Food Products, Den Haag 25th to 26th January 1999. Institute for Product Development, Lyngby, Denmark
White, R.G., 1974. Fuel requirements for selected Farming Operations. Bulletin E-780. Michigan State University.
Åbyholm, P.I., 1995. Important Questions on Fertilizers and the Environment. Hydro Agri Europe. Bruxelles. 54 pp.
Version 1.0 Januar 2007 • © Miljøstyrelsen.