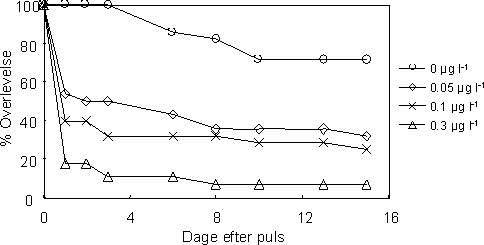
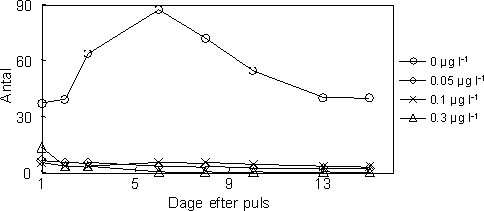
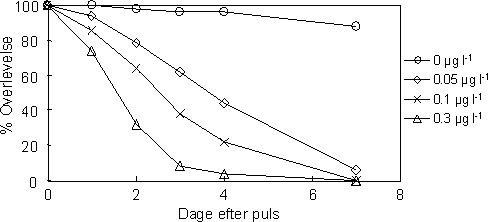
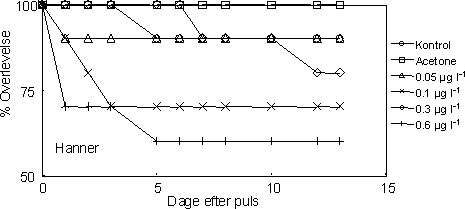
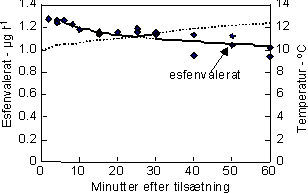
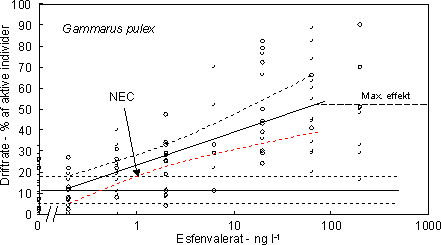
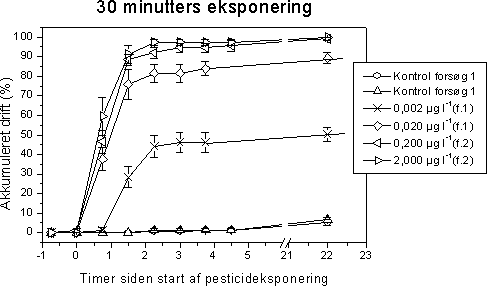
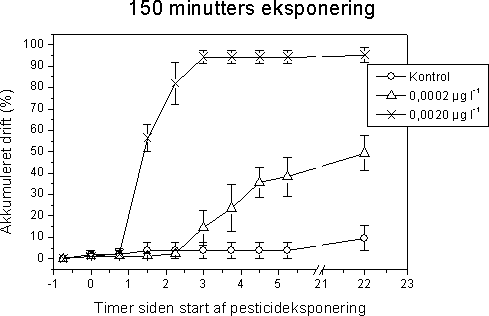
|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 82, 2004 Effekt af bekæmpelsesmidler på flora og fauna i vandløbIndholdsfortegnelse2 Vandløb og tilførsel af pesticider
3 Effekter af pesticider på alger i vandløb 4 Effekter af pesticider på invertebrater i vandløb
Bilag A: Bentiske mikroalger ForordDette projekt "Effekt af bekæmpelsesmidler på flora og fauna i vandløb" er udarbejdet for Miljøstyrelsen i perioden fra 1. marts 2001 til 1 juli 2003. Projektet blev udbudt af Miljøstyrelsen ved udbudsbekendtgørelse i sommeren 2000 og indeholder resultater af forsøg som er udført med naturligt forekommende alger og dyr fra danske vandløb. Projektet er udført af en projektgruppe bestående af: DHI – institut for vand & miljø:
Roskilde Universitets Center:
Danmarks Miljøundersøgelser:
Projektet er gennemført som et samarbejde mellem de tre institutioner. De forskellige delprojekters indhold og udførelse er løbende blevet diskuteret af parterne for at sikre, at projektets overordnede mål er fulgt, at relevante forsøgsresultater for projektgruppen er kommunikeret samt at foreslå ændringer i forsøg, hvis resultater gav anledning hertil. De enkelte institutioners primære ansvarsområder har været: DHI vand & miljø har forestået projektledelsen og haft det overordnede ansvar for projektets gennemførelse og rapportering. Endvidere har DHI udført undersøgelser af pesticiders effekt på bundlevende mikroalger, udført forsøg med invertebrater til belysning af eksponeringsvarighedens på dødeligheden samt udført adfærdsforsøg i recirkulerende strømrender. Roskilde Universitets Center har forestået undersøgelser af pesticiders effekt på forskellige livsstadier samt undersøgt effekten på en fuld livscyklus hos et insekt. Danmarks Miljøundersøgelser har udført undersøgelser af pesticiders effekt på krebsdyrs og insekters driftsmønster i store gennemstrømningssystemer under feltlignende forhold samt udført undersøgelser til belysning af pesticiders indirekte effekt på vandløbsdyrs nedbrydning af detritus. Projektet har været fulgt af en styregruppe med følgende medlemmer: Jørn Kirkegård, Miljøstyrelsen Styregruppen har fulgt arbejdet med projektet og deltaget intensivt i diskussioner af resultater og konklusioner. Vil især fremhæve konstruktive bidrag fra Peter Wiberg-Larsen, Niels-Martin Frost og Nina Cedergren Forchhammer. Generelt giver de opnåede resultater mulighed for andre fortolkninger af risikoen for effekter af pesticider i danske vandløb. Ansvaret for rapportens resultater og konklusioner er dog projektgruppens. Sammenfatning og konklusionerRapporten beskriver resultater fra 2½ års undersøgelser af en række forskellige pesticiders effekter på bundlevende mikroalger og et repræsentativt udvalg af invertebrater fra danske vandløb. Undersøgelserne er finansieret af Miljøstyrelsen og er gennemført i et samarbejde mellem DHI – institut for vand & miljø, Roskilde Universitets Center og Danmarks Miljøundersøgelser. Formålet med undersøgelserne var at belyse effekter af pesticider på naturligt forekommende alger og invertebrater. Langt hovedparten af vores nuværende viden om effekter baserer sig på undersøgelser gennemført under standardiserede betingelser så som konstante koncentrationer af pesticider og med dyr og planter fra stillestående vande. Resultater herfra kan ikke umiddelbart anvendes til at forudsige effekter i vandløb, fordi pesticider her vil optræde kortvarigt i høje koncentrationer og fordi standardorganismer så som dafnier ikke nødvendigvis er lige så følsomme som vandløbsorganismer. I undersøgelserne blev der fokuseret på effekter af realistiske eksponeringer med varigheder ned til ½ time og efterfølgende effekter på overlevelse, vækst, reproduktion og adfærd i op til 30 dage efter eksponeringen. De undersøgte pesticider (i alt 14) blev udvalgt efter deres kemiske egenskaber og forbruget i Danmark. Endvidere blev det tilstræbt at hovedparten af pesticidernes virkemekanismer var repræsenteret. Der blev undersøgt for effekter på ”naturlige” samfund af bundlevende mikroalger, på populationer af krebsdyr (2 arter), dansemyg, slørvinger (2 arter), vårfluer (2 arter), døgnfluer og snegle. Hos én art blev der undersøgt for effekter på forskellige stadier i livscyklus og hos én art gennemførtes en fuld livscyklus. Hovedparten af eksponeringerne er sket i vandet, mens 2 tests er gennemført hvor sedimentet eller føden blev kontamineret. Kombinationer mellem undersøgte pesticider og organismer blev besluttet udfra eksisterende viden om pesticidernes toksicitet og virkemekanismer. Således var der overvægt af herbicider i undersøgelserne med alger og en overvægt af insekticider i undersøgelserne med vandløbsdyr, mens fungicider blev undersøgt både på alger og dyr. Forsøgene med bundlevende mikroalger viste at herbiciderne isoproturon og metribuzin hæmmede fotosyntesen ved koncentrationer i intervallet 0,3 – 1 µg l-1. Dette ligger inden for det interval som er påvist i flere danske vandløb for isoproturon og for stofgruppen triaziner, f.eks. atrazin, terbuthylazin, simazin, der med hensyn til toksicitet ligner metribuzin. Det er derfor meget sandsynligt, at disse herbicider kan påvirke stofproduktionen i enkelte vandløb. Da effekterne var fuldt reversible og varigheden af forhøjede koncentrationer typisk er 2-10 timer i små vandløb vurderes den direkte effekt på mikroalgerne og den indirekte effekt på invertebraterne dog at være af mindre betydning. Herbicidet pendimethalin påvirkede algesamfundene ved relativt lave koncentrationer. Især grønalger var følsomme og effekten på deres biomasse forstærkedes selv efter overførsel til rent vand ved den lavest anvendte koncentration (10 µg l-1). Sammenlignet hermed udløstes der først mortalitet hos krebsdyret Gammarus pulex i mg l-1 området og kun efter kontinuerlig eksponering i flere dage. Pendimethalin er et af de mest anvendte herbicider i jordbruget, men fundprocenter og målte koncentrationer er relativt lave pga. lovbestemte sprøjtefrie zoner langs vandløbene og en lav mobilitet. Alligevel vurderer vi at pendimethalin kan medføre effekter på de bentiske alger, fordi det antageligt akkumuleres i vandløbsbunden hvor mikroalgerne sidder og eksponeres, fordi det er relativt persistent og fordi effekterne er længerevarende. Der er dog ingen målinger af pendimethalin i vandløbssedimenter som kan støtte denne vurdering. Fungicidet azoxystrobin var kun svagt toksisk overfor alger, mens effektgrænsen for mortalitet hos den mest følsomme invertebrat, Gammarus pulex var betydeligt lavere (LC50(4-14 dage) = 3-10 µg l-1). Kortvarige eksponeringer (0,5-2 timer) førte til forsinket mortalitet, men kun hvis eksponeringskoncentrationen havde været urealistisk høj (2,5 mg l-1). Derimod blev fødebearbejdningsaktiviteten hos G. pulex signifikant reduceret ved kortvarige (½ time) eksponeringer til 20 µg l-1. Til sammenligning var der ingen effekt på nedbrydningen af kontaminerede blade. Azoxystrobin er først indført i jordbruget i 1998 og følgelig er der ringe erfaring med forekomst i danske vandløb. Forekomst af azoxystrobin i koncentrationer omkring 20 µg l-1 i danske vandløb er ikke sandsynlige selv om mobiliteten er relativt høj og nedbrydningen meget langsom, men da azoxystrobin er nyt på markedet og endnu ikke har indgået i omfattende overvågningsprogrammer kendes de realistiske koncentrationer ikke. Fungicidet fenpropimorph var som Azoxystrobin kun svagt toksisk overfor mikroalger og der var fuldt recovery af både fotosynteseaktivitet og biomsser efter overførsel til rent vand. I NOVA programmet er fenpropimorph påvist i 2-4% af de undersøgte prøver, med en median-maximum koncentration på 0,07 µg l-1 (DMU 2001, DMU 2002). Med den baggrund anser vi effekter af fenpropimorph på algesamfundene for usandsynlige. Fungicidet prochloraz var moderat toksisk overfor Gammarus pulex med stærkt faldende effektkoncentration med øget eksponeringstid (LC50,24 timer = 22 mg l-1, LC50(14 dage) = 0,6 mg l-1. Ved en realistisk eksponeringsvarighed (< 24 timer) var der ingen akut mortalitet selv ved 10 mg l-1. Derimod blev der ”afsat” toksicitet ved kortvarige (½-1 time) eksponeringer til meget høje koncentrationer (>5 mg l-1). Prochloraz er ikke påvist i danske vandløb under NOVA programmet og sammen med de høje effektkoncentrationer anser vi det for usandsynligt at fungicidet kan påvirke invertebratfaunaen i danske vandløb. Toksiciteten af insekticidet dimethoat var lav overfor alle undersøgte invertebrater med LC50,96timer værdier på 5 mg l-1 eller højere. Hos krebsdyret Gammarus pulex øgedes toksiciteten med eksponeringsvarigheden, men selv efter 14 dage var LC50 værdien højere end 1 mg l-1. Der var ingen effekt på Gammarus' nedbrydning af elleblade selv efter eksponering til 2 mg l-1. Set i forhold til meget lave fundprocenter og koncentrationer i danske vandløb anser vi derfor, at effekter af dimethoat for lidet sandsynlige. Insekticidet pirimicarb var det mindst giftige af de undersøgte pesticider. Eksponering til så høje koncentrationer som 5 mg l-1 medførte ingen toksicitet hos 5 forskellige arter selv efter 14 dage. Sammenholdt med lave fundprocenter (4%) og meget lave koncentrationer (max 0,026 µg l-1) anser vi at effekter af pirimicab er meget usandsynlige i danske vandløb. Toksiciteten af insekticidet diflubenzuron blev undersøgt overfor 5 forskellige invertebrater. Der kunne ikke påvises nogen akut (96 timer) mortalitet ved de højeste anvendte koncentrationer (Gammarus: 0,2 mg l-1, øvrige arter: 3-5 mg l-1). Insekticider indenfor benzoylphenylurea gruppen er specifikt virkende på vækst/hudskifte hos insekter (og formentligt krebsdyr) og der kan derfor ikke forventes effekter ved kortvarige eksponeringer. I Danmark anvendes insekticider af benzoylphenylurea gruppen ikke i jordbruget og risikoen for transport til vandløb er meger ringe. Insekticidet esfenvalerat der tilhører gruppen af pyrethroider var langt det mest giftige af de undersøgte pesticider. Standardiserede akuttest viste LC50,96 timer værdier for Gammarus pulex på samme niveau som hos dafnier (ca 200 ng l-1) og de akut toksiske koncentrationer var markant lavere end hos de øvrige undersøgte invertebrater. Ved længerevarende eksponering (14 dage) reduceredes LC50 værdien til ca 30 ng l-1. Selv kortvarige (½-1 time) eksponeringer til lave koncentrationer (50-200 ng l-1) havde efterfølgende effekt på overlevelse, vækst/udvikling, reproduktion og fødebearbejdning. Undersøgelser af driften hos invertebrater efter eksponering til esfenvalerat viste effektkoncentrationer mellem 0,2 og 20 ng l-1 med Gammarus pulex som den suverænt mest følsomme af de undersøgte arter. Hos de to undersøgte vårfluearter havde esfenvalerat ingen effekt på driftmønstret. Der er ikke påvist esfenvalerat eller andre pyrethroider under NOVA programmet, men under flomhændelser i danske vandløb er der målt koncentrationer op til 660 ng l-1. Den forøgede drift ved en nominel koncentration på 0,2 ng l-1 er langt lavere end de højeste målte koncentrationer, men også meget lavere end detektionsgrænsen for kemiske analyser. Det er velkendt, at pyrethroider oa. pesticider med høj partikelaffinitet under naturlige forhold især forekommer som bundet til partikler. Og det er vist, at den partikulere fraktion er mindre giftig end den opløste. Det gør det vanskeligt på basis af kemiske analyser at forudsige om pyrethroider påvirker invertebratfaunaen i danske vandløb. Vi mener dog at med så lave effektkoncentrationer selv efter kortvarige eksponeringer og den brede påvirkning på vitale processer som overlevelse, reproduktion og adfærd vil pyrethroider blandt pesticiderne være den største trussel for faunaen i danske vandløb. Summary and conclusionsThis report presents results from a 2½ year study of the effects of a range of pesticides on benthic microalgal communities and on representative benthic invertebrates from Danish streams. The study was financed by the Danish Environmental Protection Agency and was conducted in collaboration among DHI Water and Environment, Roskilde University, and the Danish National Environmental Research Institute. The objective of the study was to elucidate the effects of pesticides on naturally occurring algae and invertebrates. The majority of our present understanding of pesticide effects is based on tests conducted under standardised conditions, including exposure to constant concentrations of pesticides and using animals and plants from stagnant water bodies. Such results cannot directly be used to predict effects in streams in which pesticides will tend to occur at high concentrations for short periods of time and because standard test organisms, such as daphnids, are not necessarily as sensitive to pesticides as stream-dwelling species. The present study focuses on measuring effects of realistic exposure durations, as short as 30 minutes, and examines subsequent (delayed) effects on survival, growth, reproduction and behaviour for up to 30 days post exposure. The pesticides examined in the study (a total of 14) were selected on the basis of their chemical characteristics and extent of use in Denmark. Furthermore, the aim was to ensure that most of the toxic mechanisms of action were represented. We investigated effects on 'natural' communities of benthic microalgae, on populations of crustaceans (2 species), midges, stoneflies (2 species), caddis flies (2 species), mayflies, and snails. We examined effects in different life stages within species, and for one of the species (the midge, Chironomus riparius) we conducted a full life-cycle study. Most of the exposures were carried out in water, however two tests were conducted in which either sediment or food was contaminated. Combinations of the selected pesticides and study species were chosen on the basis of existing knowledge about the pesticides' toxicity and mechanism of action. Therefore we tested primarily herbicides in tests with algae and primarily insecticides in tests with invertebrates, whereas fungicides were tested with both taxonomic groups. The experiments with benthic microalgae showed that the herbicides isoproturon and metribuzine inhibited photosynthesis at concentrations in the range 0.3 – 1 µg l-1. These concentrations are in the range shown to occur in several Danish streams for isoproturon and for pesticides belonging to the triazine group (i.e., atrazine, terbuthylazine, simazine) that have a toxicity similar to metribuzine. It is therefore very likely that these herbicides can influence primary production in a few streams. However since the effects were fully reversible, and since the duration of increased concentrations is typically 2 – 10 hours in small streams, we conclude that direct effects on microalgae and indirect effects on invertebrates are minimal. The herbicide pendimethalin affected algal communities at relatively low concentrations. Green algae were particularly sensitive, and effects on their biomass intensified even after transfer into clean water following exposure to the lowest test concentration (10 µg l-1). In comparison, mortality of the crustacean, Gammarus pulex, first occurred at concentrations in the mg l-1 range, and then only after continuous exposure for several days. Pendimethalin is one of the most widely used herbicides in agriculture, but measured concentrations and the percentage of samples in which it has been detected are relatively low due to mandatory buffer zones along streams and due to a low mobility. In any case, we estimate that pendimethalin may cause effects on benthic algae because it probably accumulates in stream bottoms where benthic microalgae are exposed, because it is relatively persistent, and because effects are long-lasting. There are, however, no measurements of pendimethalin in stream sediments that support this suggestion. The fungicide azoxystrobin was only weakly toxic to algae, whereas concentrations resulting in mortality of the most sensitive invertebrate, Gammarus pulex, were markedly lower (LC50(4-14 days) = 3-10 µg l-1). Brief exposures (0.5 – 2 hours) resulted in delayed mortality, but only if exposure concentrations were unrealistically high (2.5 mg l-1). On the other hand, feeding activity in G. pulex was significantly reduced even after short (0.5 hour) exposures to 20 µg l-1. In comparison, breakdown of contaminated leaves was not affected. Azoxystrobin was first introduced to agriculture in 1998, and consequently there is little information on its occurrence in Danish streams. The occurrence of azoxystrobin in concentrations around 20 µg l-1 in Danish streams is not likely despite the fact that its mobility is relatively high and its degradation very slow. However, since azoxystrobin is new on the market and has not been included in comprehensive monitoring programmes, realistic environmental concentrations are unknown. The fungicide fenpropimorph was, like azoxystrobin, only weakly toxic to microalgae, and there was full recovery of both photosynthesis and biomass following transfer to clean water. In the NOVA program fenpropimorph has been found in 2-4% of the samples taken, with a median-maximum concentration of 0.07 µg l-1 (DMU 2001, 2002). On this background we consider that effects of fenpropimorph on algal communities are unlikely. The fungicide prochloraz was moderately toxic to Gammarus pulex with effect concentrations falling sharply with increased exposure duration (LC50(24 hours) = 22 mg l-1, LC50,14 days = 0.6 mg l-1. There was no acute mortality at realistic exposure durations (< 24 h) up to concentrations of 10 mg l-1. On the other hand we observed delayed toxicity following brief exposures (0.5 - 1 h) to very high concentrations ( > 5 mg l-1). Prochloraz has not been detected in Danish streams by the NOVA program, and together with the high effect concentrations we consider it unlikely that this fungicide affects the invertebrate fauna in Danish streams. Toxicity of the insecticide dimethoate was low for all the invertebrates tested with LC50,96 h values of 5 mg l-1 or higher. In the crustacean Gammarus pulex, toxicity increased with increasing exposure duration, but even after 14 days the LC50 was greater than 1 mg l-1. There was no effect on Gammarus' breakdown of alder leaves even after exposure to 2 mg l-1. Considered in relation to the very low percent detection and measured concentrations in Danish streams, we consider therefore that the effects of dimethoate are not very likely. The insecticide pirimicarb was the least toxic of the pesticides tested. Exposure to concentrations as high as 5 mg l-1 resulted in no toxicity in five species even after 14 days. Compared with the low percent detection (4%) and very low measured concentrations (maximum 0.026 µg l-1), we consider that effects of pirimicarb are very unlikely in Danish streams. The toxicity of the insecticide diflubenzuron was investigated in five different invertebrates. We only found acute (96 h) mortality at the highest test concentration (Gammarus pulex: 0.2 mg l-1; other species: 3-5 mg l-1). Insecticides in the benzoylphenylurea group act specifically on growth/molting in insects (and presumably crustaceans), and effects of short-lived exposures would not be expected. In Denmark insecticides of the benzoylphenylurea group are not used in agriculture, and therefore the risk of transport to streams is very small. The pyrethroid insecticide esfenvalerate was by far the most toxic of the investigated pesticides. Standardised acute tests showed an LC50,96 h value for Gammarus pulex to occur at the same level as in daphnids (ca. 200 ng l-1), and the acutely toxic exposure concentrations were markedly lower than observed for the other invertebrates tested. During longer term exposures (14 days) the LC50 reduced to a value of approx. 30 ng l-1. Even brief (0.5 – 1 h) exposures to low concentrations (50-200 ng l-1) resulted in subsequent effects on survival, growth/development, reproduction and feeding activity. Investigations of invertebrate drift following exposure to esfenvalerate showed effect concentrations between 0.2 and 20 ng l-1 with Gammarus pulex as clearly the most sensitive of the tested species. Esfenvalerate had no effect on drift of the two caddis fly species tested. Neither esfenvalerate nor other pyrethroids have been detected in the NOVA program, however, under spate conditions in Danish streams concentrations up to 660 ng l-1 have been measured. The increased drift at a nominal concentration of 0.2 ng l-1 is much lower than the highest measured concentration, but also much lower than the detection limit for chemical analysis. It is well known that pyrethroids and other pesticides with a high particle affinity under natural conditions occur primarily bound to particles. It has also been shown that the particle-bound fraction is less toxic than the dissolved. This makes it difficult to predict effects of pyrethroids on the invertebrate fauna in Danish streams on the basis of chemical analyses. We believe, however, that with such low effect concentrations even after brief exposures and the widespread effects on vital processes such as survival, reproduction and behaviour, pyrethroids present the greatest pesticide threat to invertebrate fauna in Danish streams. 1 BaggrundSammenlignet med stillestående vande er vores viden om pesticiders effekter i vandløb yderst begrænset. Det skyldes, at man først indenfor det seneste 10-år har opbygget en nødvendig viden om forekomsten af pesticider i vandløb, men også vanskeligheder med at efterligne en realistisk pesticideksponering og holde typiske vandløbsorganismer under kontrollerede forhold i forsøg. Vores nuværende viden er således primært centreret omkring organismer repræsentative for stillestående vande. Denne viden kan ikke umiddelbart overføres til vandløb på grund af forskelle i pesticidernes eksponeringsforløb, organismernes fysiologi og livsstrategier samt forskelle i det fysiske miljø. Det primære grundlag for vurderinger af pesticiders effekter i det akvatiske miljø udgøres i dag af enkeltartstest med flere arter tilhørende forskellige trofiske niveauer og systematiske grupper. I disse tests har reproducerbarhed haft høj prioritet, hvilket delvist er gået ud over miljørealismen. Toksiciteten af kemiske stoffer, herunder pesticider, bestemmes overfor ”standard” organismer efter standardiserede vejledninger, hvor eksponeringskoncentrationerne søges holdt konstant, og hvor mortaliteten aflæses efter nøje fastlagte tidsintervaller. Ofte rapporteres den målte effekt kun som en effektparameter efter en standardiseret eksponeringsperiode, f.eks. LC50,96timer for fisk, EC50,48 timer for krebsdyr og EC50,72 timer for planktonalger. I miljøet udsættes organismerne derimod for koncentrationer, der varierer med tiden – mest udtalt i rindende vand. Her sker eksponeringen af den fastsiddende flora og fauna til pesticider i pulser i nær tilknytning til udsprøjtning og afstrømningshændelser. Da ”standard” organismerne uden undtagelse er repræsentanter for stillestående vande og de publicerede effektværdier primært er gældende for kontinuerlig eksponering er det vanskeligt at anvende eksisterende toksicitetsværdier til forudsigelse af effekter i vandløb. Nedenfor beskrives svagheder i det nuværende grundlag for organismer i vandløb:
Det er på den baggrund, at nærværende projekt blev igangsat og financieret af Miljøstyrelsens pesticidforskningsprogram. Der er derfor behov for at tilvejebringe et større datagrundlag omkring miljørealistiske effekter af pesticider på vandløbsorganismer, som både repræsenterer økologisk vigtige arter, grupper samt livsstadier og som kan udgøre fundamentet for realistiske effektvurderinger. 1.1 Pesticiderne i jordbrugetPesticider (plantebeskyttelsesmidler) er miljøfremmede stoffer, som udbringes på marker, skove, gartnerier, parker og haver for at slå svampe, insekter og ukrudt ihjel. Afhængig af målgruppe inddeles pesticiderne i ukrudtsmidler (herbicider), insektmidler (insecticider) og svampemidler (fungicider). Endvidere anvendes vækstregulerende midler (stråforkortere) i betydelig udstrækning i jordbruget. Pesticiderne indeholder et eller flere aktivstoffer rettet mod én eller flere skadegørere og normalt forekommer aktivstoffet sammen med hjælpestoffer, opløsningsmidler og vand, der sikrer effektiv dosering og optimal virkning på skadevolderne. Før 1980 havde reguleringen af pesticidanvendelsen sigte på at beskytte mennesker, husdyr og bier, men med forøget viden og fokusering på det omgivende miljø har reguleringen siden også fokuseret på at minimere effekterne i miljøet. Det er således dokumenteret, at der sker en påvirkning af flora og fauna i vandløb, søer og småbiotoper og at der udvaskes pesticider til grundvandet (Miljøstyrelsen 1999) Tabel 1. Forbrugsmønster af pesticider i dansk jordbrug i 1992 og 2001 (Miljøstyrelsen 1993, 2002)
I løbet af en 10-årig periode er pesticidanvendelsen i jordbruget (dvs. excl. gartnerier, haver o.l.) faldet ca. 34 % (fra 4653 t aktivstof til 3075 t), mens reduktionen i det sprøjtede areal “kun” udgør 15% (Tabel 1). Miljøgevinsten har antagelig været større ved reduktionen i antallet af aktivstoffer (85 68) som især har omfattet de mest persistente. Og for det akvatiske miljø har indførsel af sprøjtefrie zoner langs vandløb og vandhuller med stor sandsynlighed været en betydelig gevinst, da risikoen for tilførsel til vandet er blevet mindre. 2 Vandløb og tilførsel af pesticider2.1 Danske vandløbDanske vandløb og vandløbsoplande er generelt små. Halvfjers procent af det totale areal afstrømmes af vandløb med oplandsarealer mindre end 500 km² og kun to vandløb (Gudenå og Skjern Å) er længere end 100 km. Hovedparten af de danske vandløb (75%) er smallere end 2,5 m og er karakteriseret af en lav vandføring (Sand-Jensen & Friberg 2000). I den østlige del af Danmark er grundvandsdannelse mindre end i den vestlige del af landet (hovedparten af Jylland) pga. forskelle i jordtyper og nedbør. Dette bevirker at variationen i vandføring over året er størst i de østdanske vandløb, da de modtager hovedparten af deres vand fra overfladeafstrømning, mens vandløbene i den vestlige del er karakteriseret af mere stabile afstrømningsforhold. Sommervandføringen vil derfor være lavest i vandløbenei den østlige del af landet.
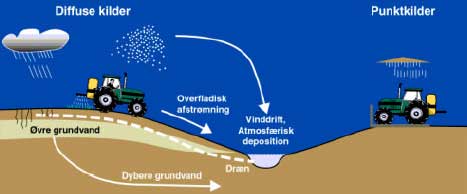
Figur 1. Hovedparten af danske vandløb er små. Da vandløbene generelt er små har de de en relativt stor kontaktflade til det terrestriske økosystem. Vandløbenes omgivelser bidrager med organisk stof (allochthont materiale) og forskellige kemiske forbindelser. Især i de mindre vandløb er denne tilførsel den primære kulstofkilde, og derfor af afgørende betydning for vandløbsøkosystemets struktur og funktion. Koblingen til omgivelserne er yderligere blevet forstærket pga. drænrør på lerede jorde, der har øget den hastighed hvormed vandet, samt opløste og partikelbundne stoffer, når vandløbet fra det terrestriske økosystem. Mere end 60% af arealanvendelsen i Danmark udgøres af landbrug. Det betyder at den overvejene andel af de danske vandløb ligger i et landbrugslandskab. De fleste af vandløbene er samtidig små og derfor påvirket af de aktiviteter, der forekommer i de vandløbsnære arealer. Det er i disse vandløb, der må forventes de største effekter af bekæmpelsesmidler på de økologiske forhold. Fortyndingen af de stoffer, der tilføres vandløbet enten via vinddrift, dræn eller overfladisk afstrømning vil være lille, samtidig med at bidraget potentielt kan være stort pga. den tætte kontakt med landbrugsarealerne. Det må yderligere forventes, at vandløb på lerede jorde med høj dræningsgrad og lav vandføring i sprøjtesæsonen er mest udsatte. 2.2 Biologiske komponenterVandløb er økosystemer der fysisk varierer på forudsigelig måde fra kildeudspring til udløb i havet. De øvre vandløb er karakteriseret af et stort fald, groft substrat og lav vanddybde. Desuden er vandføring ofte relativ konstant og temperaturen lav pga. en stor grundvandstilførsel (Sand-Jensen & Friberg 2000). I naturtilstanden vil det øvre vandløb være helt skygget af vegetationen i den ripariske zone og den helt overvejende kulstofkilde vil stamme fra det terrestriske økosystem. De biologiske samfund vil afspejle disse forhold. De få primærproducenter der forekommer er primært perifytiske mikroalger, især kiselalger, som vokser på sten. Er vegetationen langs vandløbet fjernet, vil primærproducenterne få en større betydning. Det vil stadig være mikroalger, men også trådalger som Cladophora, der dominerer. Det skyldes at det grove substrat og store fald medfører, at makrofyter har svært ved at etablere sig. Den lave primærproduktion bevirker at det øvre vandløb er heterotroft. Det kan lade sig gøre på grund den store tilførsel af allochthont materiale så som blade og grene fra træer langs bredden og en stor genluftning, der skaber et iltrigt miljø. Invertebratsamfundet i det øvre vandløb er domineret af detritivore, der enten lever af findele det grove organiske stof (CPOM: blade etc.) eller ved at samle fint organisk stof. Det fine organiske stof (FPOM) stammer dels fra omsætningen af blade i vandløbet og dels tilført FPOM fra det terrestriske økosystem. De inverterbrater der findeler CPOM til FPOM spiller en nøglerolle for økosystemets funktion ved at øge kvaliteten og tilgængeligheden af det øvre vandløbs primære føderessource, nemlig de nedfaldne blade. Disse invertebrater kaldes iturivere (shredders) og ferskvandstangloppen Gammarus pulex er en væsentlig ituriver i næsten alle danske vandløb. Mange af ituriverne, som f.eks. vårfluen Sericostoma personatum er desuden tilpasset til at leve i relativt kølige vandløb med lave sommertemperaturer og meget gode iltforhold. En del af det producerede FPOM vil indgå i fødenettet i det øvre vandløb, mens den resterende mængde vil transporteres nedstrøms . På de mellemste vandløbstrækninger har brinkvegetationen ikke samme skyggende virkning pga. af det bredere vandspejl. Derudover er faldet blevet mindre, substratet finere og dybden større, hvilket bevirker et skift i de autotrofe komponenter (Sand-Jensen & Friberg 2000). De højere vandplanter, makrofyterne, dominerer på de mellemste vandløbsstrækninger. I forårsperioden vil primærproduktionen være domineret af bundlevende mikroalger indtil de bliver skygget væk af makrofyterne i det sene forår. Makrofyterne vil opnå maksimal biomasse i sensommeren. Selvom vækstbetingelserne for mikroalger på bunden er dårlige sommeren igennem, udgør makrofyter et substrat i sig selv for epifytiske mikroalger. Pga. af planternes tredimensionelle struktur er det tilgængelige areal for mikroalger langt større end selve bunden af vandløbet og deres biomasse derfor større. Derfor er en meget hyppigt forekommende fødefunktionel gruppe græssende invertebrater, der lever af at skrabe biofilm (alger, heterotrofe mikroorganismer og diverse exudater) af især planterne. En meget hyppigt forekommende græsser på makrofyter er døgnfluer af slægten Baetis sp. Makrofyterne bliver kun i mindre omfang konsumeret levende. Hovedparten af makrofyterne vil først indgå i fødenettet efter de er døde. Pga. af mindre støttevæv, højere næringsstof-niveau og højere temperaturer i sommerperioden vil makrofyterne hurtigere end blade fra træer omsættes til FPOM i vandløbet (Sand-Jensen & Friberg 2000). Nedbrydningen vil i højere grad være mikrobiel, men også ituriverne har betydning i denne proces. Ituriverne spiller en mindre relativ rolle i det mellemste vandløb, men deres absolutte antal er ikke meget forskellig sammenlignet med det øvre vandløb. Også i det mellemste vandløb spiller Gammarus en meget væsentlig rolle sammen med f.eks. vårfluer tilhørende familien Limnephilidae. Mængderne af FPOM på de mellemste strækninger er større end i det øvre vandløb pga. af importen af FPOM fra opstrøms strækninger og pga. egen produktionen på strækningerne. Makrofyterne vil i sommerperioden bevirke at store mængder FPOM kan tilbageholdes, da de lokalt omkring grødeøerne sænker strømhastigheden og dermed øger aflejringen af det fine organiske stof. De invertebrater der lever af FPOM kaldes samlere og har stor betydning på de mellemste strækninger. Samlerne opdeles i filtratorer og sedimentædere. Typiske filtratorer i det mellemste vandløb er kvægmyg og netspindene vårfluer tilhørende familien Hydropsyche sp. Sedimentæderne lever i og på bunden, og de tilhører en lang række taxonomiske grupper. Gammarus der fødefunktionelt er meget plastisk, ernærer sig også som sedimentæder. Desuden vil forskellige dansemyg, børsteorme og døgnfluer fungere som sedimentædere. Det mellemste vandløb er mere produktivt og diversiten højere sammenlignet med de øvre strækninger. Dette skyldes at føderessourcerne er flere og af højere kvalitet, at der er flere habitater og et større oplandsareal. Den større energibase i fødenettet bevirker at der kan opretholdes en større biomasse af både invertebrat- og vertebratprædatorer. I det nedre vandløb vil vandspejlsfaldet være mindst, sedimentet finest og dybden størst. Strømhastigheden vil samtidig være høj, da bunden kun yder ringe modstand mod vandets flow. Makrofyter og deres påvækst af epifytiske mikrolager vil primært findes i kantzonen pga. dårlige lysforhold i midten af vandløbene. I store udenlandske vandløb vil der kunne forekomme egentlige fytoplanktonsamfund, men danske vandløb er for små, og opholdstiden for kort til at fytoplankton kan etablere sig med mindre der er en indskudt sø. I mange af de større danske vandløb kan der desuden forekomme makrofyter over hele vandløbets bredde. Invertebratsamfundet vil være det mest artsrige i vandløbskontinuumet pga. antallet af habitater og rekruttering af arter fra hele oplandet. Både mht. til fisk og invertebrater vil der forekomme arter der både lever i søer og i vandløb. Den kvantitativt mest dominerende føderessource vil være FPOM, der i stor udstrækning er importeret fra opstrøms strækninger. Antalsmæssigt vil samlerne dominere blandt invertebrater, men f.eks. en ituriver som Asellus aquaticus vil også forekomme, da den har sin naturlige forekomst blandt makrofyterne i kantzonen i større vandløb. Der vil være flere aktive filtratorer som muslinger, der ligger nedgravet i det fine sediment. Tilgengæld vil der relativt være færre af de passive filtratorer som Hydropsyche sp. da der vil være mangel på groft substrat, der er eksponeret for strømmen. Der vil ligeledes findes en række sedimentædere nedgravet i sedimentet. Imidlertid vil nedre vandløbsøkosystems struktur og funktion afhænge af lokale fysiske forhold og således kan man i de fleste af største danske vandløb finde partier med groft substrat og dermed andre invertebratsamfund end de teoretisk forudsagte. Vandløb er åbne systemer, der karakteriseret af den ensrettede vandbevægelse. I naturtilstanden er der derfor en god forbindelse i vandløbenes længderetning, fra kildeudspringet til udløbet i havet. Denne forbindelse er utrolig vigtig for vandløbets organismer og dermed økosystemets struktur og funktion. Spredning gennem vandløbet gør det muligt for organismerne at rekolonisere efter forstyrrelser, f.eks. store afstrømninger, der lokalt kan have reduceret eller fuldstændigt udryddet, populationer. Vandløbsøkosystemer er derfor generelt meget resiliente, dvs. at de efter en forstyrrelse vender hurtigt tilbage til den tilstand, der var før forstyrrelsen indtraf. Derudover har de akvatiske insekter mulighed for at sprede sig indenfor og mellem vandløb i deres flyvende voksenstadium. Dette er meget væsentligt for mulighederne for at rekolonisere områder hvor arter er gået tabt pga. forstyrrelser. Vandløb er naturligt forstyrrede systemer og derfor har et stort rekoloniseringspotentiale en stor adaptiv værdi. Desuden har invertebraternes spredningsevne også stor betydning for økosystemets mulighed for at komme sig over menneskeskabte forstyrrelser som f.eks. udledninger af miljøfremmede stoffer. 2.3 Tilførsel og koncentration af pesticiderPesticider tilføres utilsigtet vandløb fra diffuse kilder, som har ophav i udbringningen på marken eller fra punktkilder, især vaske- og spulepladser, hvor tanke og sprøjter tømmes for rester og rengøres (Fig. 2). Diffuse kilder omfatter vinddrift af aerosoler, overfladisk afstrømning under kraftige nedbørshændelser samt tilførsel via dræn og det dybere grundvand.
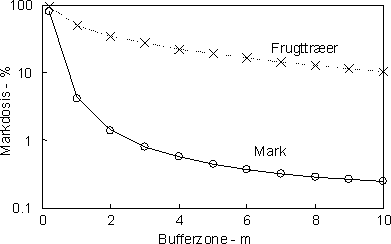
Figur 2. Kilder til pesticidforurening af vandløb. Vinddrift forekommer når der udsprøjtes tæt ved vandløbene og vindhastigheden er høj og i retning mod vandløbet. Størrelsen af vinddriften og hvor stor del som tilføres vandløbet afhænger udover vindhastigheden især af sprøjtedysernes højde over jorden samt af afstanden til vandløbet (bufferzone). I Figur 3 er vist den beregnede procentdel af markdosis, som når vandløbets overflade ved sprøjtning af frugttræer og afgrøder på marken ved anvendelse af forskellig bredde af sprøjtefri zone. Effekten af bufferzoner er dramatisk når der udsprøjtes ved jorden. Således kan der forventes en tilførsel på mindre end 1 % af markdosis, hvis den sprøjtefrie zone overstiger 4 m (og vindhastigheden er lavere ned 3-4 m s-1). Ved udsprøjtning i vækstsæsonen vil reduktionen være større pga. tilbageholdelse i afgrøden og i bufferzonens vegetation. Beregningerne viser, at for de mest giftige pesticider hvor der er lovpligtige sprøjtefrie zoner på 10-20 m, er risikoen for signifikant tilførsel med vinddrift lav, såfremt begrænsningerne overholdes.
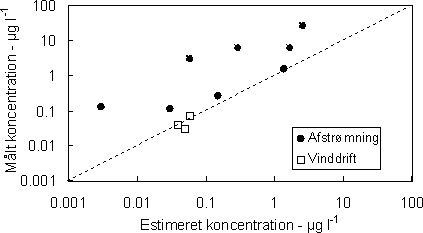
Figur 3. Markdosis (i %) der når vandløbets overflade som funktion af bufferzonens bredde. Beregninger er foretaget på baggrund af Ganzelmeiers formler for tidlig sprøjtning (dvs. inden løvspring) og ved lav vindhastighed (< 3-4 m s-1). (Ganzelmeier et al. 1995) Størrelsen af pesticidtilførslen ved afstrømning på overfladen eller via drænvand er afhængig af jordens karakter (porositet, organiske indhold) vandindholdet i jorden, hældningen mod vandløbet, plantedække i randzonen og på marken, pesticidernes egenskaber (partikelaffinitet, halveringstid) samt nedbørens størrelse og frekvens i forhold til udsprøjtningstidspunktet. De mange forhold som påvirker tilførslen gør det vanskeligt at generalisere, og kvalificerede bud på tilførsler forudsætter en dynamisk modellering som indeholder beskrivelser af de enkelte processer eller et omfattende undersøgelsesprogram. Tilførslen med overfladisk afstrømning kan ske som opløste eller partikelbundne pesticider, som igen bestemmes af pesticidernes egenskaber. Generelt vil forudsætning for betydelig overfladisk afstrømning være at jorden er vandmættet, har hældninger mod vandløbet større end flere procent og en stor nedbørsintensitet, f.eks. mere end 10 mm i løbet af 24 timer (Heidmann & Hansen 1995, Huber et al. 1998). Wauchope (1978) har på basis af litteraturstudier angivet hvor store dele af markjordens pesticidindhold der tabes ved erosion og som opløste pesticider (og evt. tilføres vandløb og vandhuller) under forskellige nedbørsforhold. Tabene angives af Wauchope til mellem 0,1 og 5%. Til sammenligning var tabene betydeligt lavere i danske undersøgelser (mellem 0,008 og 0,01%, Felding el al. 1997) og tyske (0,01-0,07%, Liess et al. 1999). Sammen med Ganzelmeiers (1995) undersøgelser har Wauchope's empiriske relationer udgjort grundlaget for flere simple eksponerings- og risikomodeller, f.eks. REXTOX (Møhlenberg et al. 2002), TOXWA, FSWRAT, DRIPS (Röpke et al. 2002). Ved sammenligning af beregnede og målte koncentrationer i vandløb er der en tendens til at afstrømningen underestimeres, mens modelforudsigelserne for vinddrift er i god overensstemmelse med målinger. I eksemplet (Fig. 4) kan afvigelserne for aftrømningen skyldes at den anvendte model (REXTOX) ikke inkluderer tilførsel med dræn.
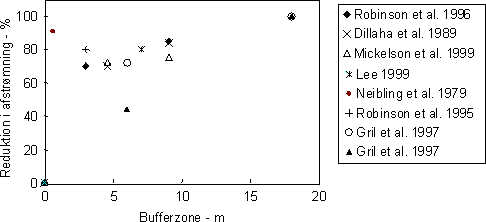
Figur 4. Beregnede og målte koncentrationer af pesticider i vandløb tilført ved vinddrift og overfladisk afstrømning. Tilførslerne er beregnet med REXTOX risikoindikator (omtegnet fra Møhlenberg et al. 2001) Som det er gældende for vinddrift er sprøjtefrie zoner langs vandløbene meget vigtige for tilførslen af pesticider med afstrømning. Således kan man forvente en reduktion på ca 70% gennem en bevokset bufferzone på 5 m (Figur 5).
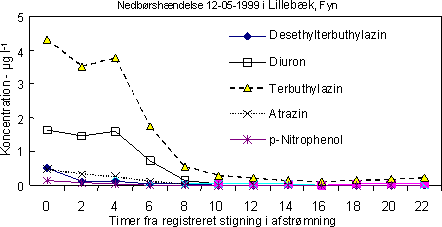
Figur 5. Reduktion af overfladisk afstrømning af pesticider gennem bufferzoner af forskellig bredde. Data repræsenterer tilbageholdelse af både opløste og partikelbundne pesticider gennem bufferzonen. I Danmark er markerne langs vandløbene normalt lavtliggende, med ringe hældning og det er estimeret at kun ca. 3% af det dyrkede areal er i risiko for erosion (Kronvang el al. 2000). Det betyder, at der kun undtagelsesvist (ved meget høje nedbørsintensiteter) vil forekomme overfladisk afstrømning. Det øvre grundvand og især dræn kan være en vigtig transportvej for pesticider til vandløb. Det er estimeret at mellem 0,1 og 5% af de tilførte pesticider kan tabes via dræn (Flury, 1996). I danske undersøgelser er der målt pesticider i dræn ved Højvads Rende (0 – 0,27 µg l-1; Spliid & Mogensen, 1995), ved Silstrup forsøgsstation (0 – 2,8 µg l-1), i dræn ved Lillebæk (0 - 1,6 µg l-1; Styczen, pers. com.), mens der under forsøgsforhold er fundet koncentrationer af prochloraz i dræn på 16 µg l-1 (Villholth 2000). Med det reviderede overvågningsprogram for vandmiljøet (NOVA) blev der sat fokus på miljøfremmede stoffer, bl.a. pesticider. I danske vandløb blev der påvist pesticider i hovedparten af de 263 analyserede prøver i 2001 med glyphosat og nedbrydningsproduktet AMPA tilstede i mere end 75% af de analyserede prøver (DMU 2002). Ud af 49 fundne pesticider var herbiciderne dominerende både i fundprocenter og antal pesticider (39), fulgt af fungicider (fundprocenter: 2 – 5%; antal: 3) og insekticider (fundprocenter: < 3%, antal insekticider: 6). Den samme fordeling mellem pesticiderne er også fundet i LOOP områderne, bl.a. Lillebæk på Fyn (Styczen, pers. com). Her har overvågningsprogrammet været mere intensivt og koncentreret omkring nedbørshændelser og i overensstemmelse hermed er der i korte perioder målt høje koncentrationer af især herbicider (Fig. 6).
Figur 6. Koncentration af 4 herbicider samt nedbrydningsproduktet p-nitrophenol (bl.a. fra parathion-Me) efter en nedbørshændelse. Undersøgelserne fra Lillebæk understreger, at forekomsten af høje koncentrationer er kortvarige i små vandløb og selv med ”intelligente” prøvetagningsprogrammer) vil det være vanskeligt af ”fange” toppene. Både i den landsdækkende overvågning og i LOOP områderne er der gentagne fund af pesticider som ikke længere er godkendte, f.eks. atrazin og nedbrydningsproduktet fra dichlorbenil BAM, som er det næsthyppigste stof i i vandløbene. 2.4 Undersøgte pesticider – deres anvendelse og repræsentativitet2.4.1 ForbrugPesticider til brug for forsøg er udvalgt efter en række kriterier så som en bred dækning af pesticidernes forskellige virkemekanismer, en passende repræsentation af insekticider, herbicider og fungicider, stoffernes anvendelse i Danmark, mulighed for at fremskaffe radioaktivt mærkede pesticider (for at gennemføre simple analyser af koncentrationer), samt ønsker fremsat af Miljøstyrelsen (jvf. Stofliste). De undersøgte insekticider dimethoat, esfenvalerat, deltamethrin og pririmicarb repræsenterer 3 vigtige stofgrupper (organofosfater, pyretroider, carbamater) og 2 virkemekanismer der påvirker transmissionen af impulser i nerverne (cholinesterasehæmmere, påvirkning af Na+/Ka+ pumpen) (se Tabel 2). Esfenvalerat anvendtes på 28% af arealet som behandles med insekticider og for pyrethroidgruppen som helhed anvendtes disse pesticider på 64% af arealet, der blev behandlet med insekticider i perioden 1999-2001. Målt i solgte mængder (Tabel 3) repræsenterer de 3 undersøgte insekticider ca. 78 % af det samlede insekticidsalg og ca 85% målt på den behandlede areal. De 6 undersøgte herbicider repræsenterer 5 vigtige stofgrupper (triaziner, phenylaurea, hydroxybenzonitril, sulfonylarea, dinitrioanilin), 4 forskellige virkemekanismer og 5 forskellige ”site of action” (Tabel 2). Målt på de solgte mængder udgjorde de 6 herbicider 18% af det samlede salg og på basis af det behandlede areal ca. 22% i 1999-2001. Det er dog afgørende, at de vigtigste virkemekanismer er repræsenteret med de 6 herbicider. Tabel 2. Oversigt over funktionelle stofgrupper og virkemekanismer af pesticider anvendt til forsøg.
De 3 undersøgte fungicider repræsenterer 3 forskellige stofgrupper (strombilurin, morpholin, imidazol) og 2 forskellige virkemekanismer (Tabel 2). Målt på de solgte mængder udgør de 3 fungicider 32% af det samlede salg, mens repræsentativiteten målt på det behandlede areal udgør 57% (Tabel 3). Tabel 3. Oversigt over forbrug, areal af behandlede arealer samt behandlingshyppighed i jordbruget af pesticiderne anvendt i forsøg. Værdierne repræsenteer gennemsnit af årene 1999-2001. For pesticider, der var forbudt i 2001 er der angivet gennemsnitsværdier for de år, hvor pesticiderne har været anvendt indenfor det seneste 10-år. Fra Miljøsryrelsen (2000, 2001, 2002).
2.4.2 Fysisk-kemiske egenskaberPesticiders risiko for transport fra marken til vandløb vil især afhænge af deres nedbrydelighed og graden af deres binding til jorden og afgrøder. For pesticider med kort halveringstid (dvs. hurtig nedbrydning) vil risikoen for transport udløst af stokastiske nedbørshændelser være mindre end for sværtnedbrydelige pesticider, der vil findes i jorden i relativt høje koncentrationer i længere tid efter udsprøjtning. Og pesticider med høj affinitet til partikler/organisk stof vil normalt kun transporteres ved erosion. Omvendt vil vandopløselige pesticider nemt transporteres med vandet f.eks. til dræn. I Tabel 4 er angivet de anvendte pesticiders halveringstid i jord samt octanol:vand fordelingskoefficienten. Til sammenligning er vist medianværdierne for samtlige insekticider, herbicider og fungicider der anvendes i Danmark. Det fremgår af Tabel 4 at variationen i de fysisk-kemiske egenskaber er stor indenfor de undersøgte pesticider, og endvidere at gennemsnittet indenfor hver pesticidgruppe placerer sig fornuftigt i forhold til medianværdierne for de anvendte pesticider i Danmark. Tabel 4. Fysisk-kemiske egenskaber af anvendte pesticider. Til sammenligning er angivet medianværdierne for samtlige insekticider, herbicider og fungicider, der anvendes i jordbruget (fed skrift). DT50 udtrykker halveringstiden af pesticiderne i jord; LogKow udtrykker logaritmen til fordeling af pesticidet i en blanding af octanol og vand. En høj LogKow udtrykker at pesticidet har en høj bindingsgrad til organisk stof (og til partikler) og vil bioakkumuleres i organismer.
3 Effekter af pesticider på alger i vandløbUndersøgelser af enkeltstoffers og komplekse blandingers (f.eks. spildevand) toksiske virkning over for vandlevende organismer udgør sammen med måling eller beregning af stoffernes koncentration i miljøet et af hovedelementerne i vurderingen af om et givet stof eller spildevand giver anledning til ikke-acceptable påvirkninger i miljøet. Øvrige forhold som indgår i den samlede vurdering omfatter stoffernes nedbrydelighed, bioakkumulerbarhed og eventuelle carcinogene effekter. Ved risikovurdering af pesticider (godkendelsesordningen) baseres vurdering af giftigheden på standardiserede undersøgelser udført på ”standardorganismer” holdt i kultur, typisk en planktonalge, en dafnie, en enkimbladet vandplante (oftest andemad) samt en fisk. Disse organismer repræsenterer arter der lever i stillestående vande. Desuden anvendes der fastlagte eksponeringsperioder til bestemmelse af toksiciteten. Således angives den akutte toksicitet typisk som EC/LC50 efter 48, 72 eller 96 timer for hhv. krebsdyr, alger og fisk, mens data for kroniske eksponeringer og effekter normalt kun foreligger for dafnier. Der er således en beskeden information om pesticiders effekt på andre endpoints så som vækst, reproduktion, og endnu mindre forekommer der undersøgelser til bestemmelse af indirekte effekter på økosystemprocesser. Det er velkendt at forskellige livsstadier af en art kan variere i følsomhed overfor et toksisk stof; oftest er juvenile individer mere følsomme end voksne individer, men om variationen i følsomhed mellem livsstadier indenfor én art er større eller mindre end forskellen i følsomheder mellem forskellige arter er uklar. I vandløb vil eksponeringskoncentrationen variere over tid og sted og have karakter af pulsudledninger afhængigt af de sæsonmæssige variationer i pesticidforbrug, behandlingshyppighed, nedbør samt vandløbets beskaffenhed. Det kan derfor være vanskeligt at anvende toksicitetsværdier opnået i standard tests til at forudsige eventuelle effekter af pesticider i vandløb. Kun undtagelsesvist eksisterer der toksicitetsdata fra eksponeringer på mindre end en dag – selvom denne type eksponeringer er hyppigt forekommende i mindre vandløb (se Fig. 6), og der er næsten ingen information om eventuelle eftervirkninger af kortvarige eksponeringer til pesticider på ferskvandsorganismer til trods for, at den tilgængelige litteratur påpeger mulige effekter længe efter eksponeringsophør (Abel & Garner 1986). Således var målet med nærværende projekt at tilvejebringe et mere realistisk billede af pesticiders mulige effekt på flora og fauna i vandløb. De mere specifikke formål med projektet var som følger:
3.1 Alger og højere planterI lysåbne vandløb er undervandsplanter og mikroalger vigtige komponenter i økosystemerne. Undervandsplanterne danner ofte tætte bestande og er vigtige for vandløbenes fysiske miljø, for habitater for invertebrater og fisk og som tilskud af organisk stof (Baattrup-Pedersen, 2000). Derimod ædes undervandsplanter kun i ringe grad men må gennem en mikrobiologisk nedbrydning før end det organiske stof kan udnyttes af vandløbsdyrene. I vandløbene er den største trussel for vandløbsplanterne den hyppige grødeskæring, som ændrer artssammensætningen mod robuste typer. Man må derfor forvente, at denne påvirkning langt vil overstige effekten af pesticider. 3.2 Effekt af pesticider på bentiske mikroalgerPå vandløbsbunden og på makrofytternes blade dominerer bentiske mikroalger. Deres variation er bestemt af lysintensiteten og deres betydning vil være stor, hvis undervandsplanterne beskæres hårdt på lysåbne vandløbsstrækninger og i skovvandløb inden løvspring (Friberg 1998). Mikroalgerne tjener som føde for en lang række vandløbsdyr (Lamberti 1996). De fleste algegrupper som kendes fra planktonet er repræsenteret blandt de bentiske mikroalger selv om kiselalger generelt dominerer. Der er en omfattende dokumentation for at herbicider i målte koncentrationer kan påvirke planktonalgesamfund i vandhuller og der er derfor nærliggende at også bentiske mikroalger i vandløb kan påvirkes af pesticider. Undersøgelser er dog relativt få (Jurgensen & Hoagland. 1990, Spawn et al. 1996, Krieger et al. 1988, Cradner & Hoagland 1998, Gruessener & Watzin 1995) og med en enkelt undtagelse (Kreutzweiser et al. 1996, 12 timers eksponering) har eksponeringerne været kontinuerlige over flere dage. Det er langt fra sikkert, at man kan overføre resultaterne fra forsøg med planktonalger, da eksponeringen i vandløb ofte vil være kortvarig med en højere koncentration end i vandhuller og søer. Da de mikrobentiske alger er et vigtigt fødegrundlag for vandløbsdyrene, er det vigtigt at kende effekter af pesticider på denne gruppe. Algerne har korte generationstider og rekoloniserer hurtigt. Men herbicider kan forårsage forskydninger i arts- og gruppesammensætningen, som kan ændre fødegrundlaget for vandløbenes invertebrater. De traditionelle toksikologiske enkeltartstest inkluderer ikke sådanne effekter, og endvidere medtager disse forsøg heller ikke den nedbrydning af pesticider, der vil være fra naturligt forekommende heterotrofe organismer. De mikrobentiske alger klæber til substrater ved hjælp af polysaccharidmatrixer, der i sig selv formentlig virker som barrierer for transporten af toksiske stoffer til algerne, og som pesticiderne i et uvist omfang vil klæbe sig til. Det er derfor vigtigt at inkludere sådanne faktorer, der kan påvirke omfanget af den toksiske effekt på perifytiske alger i forsøg. Yderligere fordele ved bentiske mikroalger (sammenlignet med makrofytter) er også at det praktisk er muligt at gennemføre statistisk ”korrekte” forsøg med flere replikater og at der eksisterer standardmetoder for kvantificering af funktionelle (primærproduktion) og strukturelle (f.eks. algepigmenter, mikroskopi) karakteristika (og end-points) i algesamfund. Tilsvarende forsøg med undervandsplanter på samfundsniveau er langt sværere at gennemføre. I dette projekt er en ny type test blevet designet, hvor intakte perifytonsamfund anvendes. I testet indgår der en 2-3 ugers koloniseringsperiode, hvor naturligt perifyton bevokser skiver af glas i et vandløb, og herefter anvendes de bevoksede skiver i laboratoriet for at undersøge herbiciders og fungiciders effekter på de algernes fotosynteseaktivitet og taksonomiske sammensætning på gruppeniveau. Da pesticider ofte forekommer i vandløb i pulser ved vinddrift eller efter regnskyl, er der i undersøgelsen lagt vægt på også at indrage korttidseksponering, for at kunne vurdere, om algernes opportunistiske leveform gør dem i stand til at klare sådanne pulseeksponeringer. Endvidere indgår der i forsøgene en ”recovery-fase”, for at kunne vurdere effekternes varighed, herunder om eksponeringen forårsager varige effekter, der således vil have indirekte effekter på invertebratfaunaen. Bentiske mikroalger i vandløbene varierer gennem sæsonen som følge af lystilgængelighed og strømhastighed. Under perioder med stærk strøm kan det finkornede sediment og perifytter skylles væk og i perioder og områder med stor dækningsgrad af undervandsplanter eller på vandløbsstrækninger med trædække kan perifytter blive udskygget. Derfor vil forårsperioden umiddelbart inden løvspring generelt være den optimale periode for indsamling. Det er også i denne periode hvor udbringning af herbicider normalt er størst. For at dække sæsonvariationen i artssammensætning blev nogle forsøg udført i sensommeren. 3.2.1 MetodePerifyton til forsøg kan indsamles fra vandløbsbunden, fra sten eller på kunstige substrater. De hyppigst anvendte kunstige substrater består af glasskiver (f.eks. Bahls 1993). De vigtigste fordele ved kunstige substrater er:
Den primære ulempe ved kunstige substrater er, at materialet til en vis grad vil favorisere visse algetyper (fastsiddende) fremfor andre (krybende former). Endvidere vil varigheden af eksponeringen i vandløbet påvirke sammensætningen af perifytsamfundet – hurtige kolonisatorer vil dominere efter kortvarig eksponering, mens længerevarende eksponering vil føre til større variation i samfundene på de enkelte glas. Indledende forsøg viste dog at der ved en eksponering (kolonisation) på 1-2 uger opnås et diverst samfund og en acceptabel lav variation mellem de enkelte glas. Der undersøgtes for effekter på en vital funktionel parameter i form af fotosynteseaktivitet og for effekter på biomasseudvikling og biomassens udvikling på gruppeniveau (kiselalger, grønalger, blågrønalger). Undersøgelsen gennemførtes som”hybrid design” med både replikater (3-4 for hver pesticideksponering og 6 kontroller) og 5 forskellige pesticidkoncentrationer. Dette giver mulighed for at estimere NEC (no effect concentration) ved regression og teste for forskelle mellem de forskellige eksponeringskoncentrationer ved ANOVA. ”No effect concentration” (NEC) og effekt koncentrationen (EC50) blev udregnet på basis af fotosyntesemålingerne ved log-lineær interpolation (Petersen & Gustavson 1998). I flere tilfælde kunne NEC ikke bestemmes. Istedet angives den lavest målte effektkoncentration (LOEC). Effekter af 5 herbicider og 2 fungicider undersøgtes på naturlige algesamfund fra Esrum Mølleå ca. 2 km nedstrøms udløbet fra Esrom Sø (Tabel 5). Denne vandløbsstrækning løber gennem et udyrket område og må formodes at være udsat for en meget lav pesticidpåvirkning. Tabel 5. Oversigt over anvendte pesticider og effektparametre
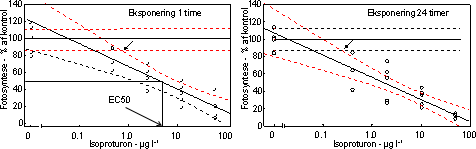
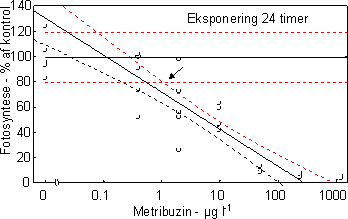
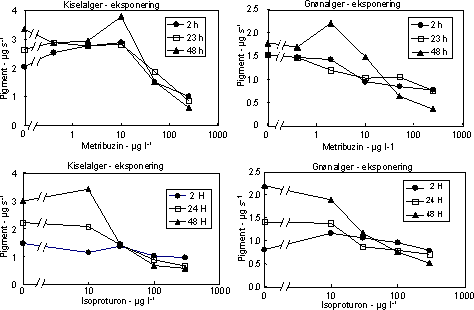
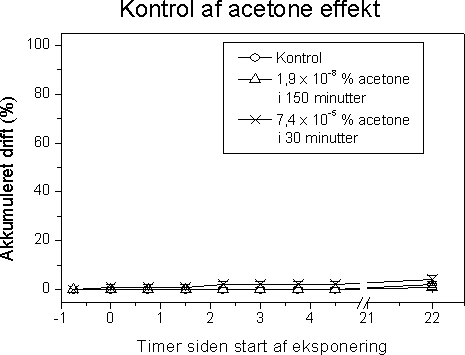
Ved en undersøgelses start udsattes 3 ramper hver indeholdene 170 glasskiver (1 cm i diameter) i vandløbet til etablering af påvækst. Efter indsamling og visuel kontrol for nær ens dækning med algevækst overførtes 400 glasskiver individuelt til 20 ml glasvials indeholdende filtreret åvand tilsat pesticid i 5 forskellige nominelle koncentrationer (bestemt efter range-finding test) + kontrol. De tungtopløselige pesticider Pendimethalin og Azoxystrobin blev inden tilsætning opløst i acetone og kontrolforsøgene gennemførtes med samme acetonekoncentration som pesticideksponeringerne Forsøgene gennemførtes med 3-4 replikater (6 replikater for kontrol). For at undgå næringsbegrænsning blev åvandet tilsat 100 µM NO3, 16 µM PO4 og 50 µM SiO2. Vials blev placeret på rystebord i termorum ved ”naturligt” lysmiljø (120 µE m-2 s-1; 14L:10M). Der blev udtaget prøver efter 1, 2, 24, 48 timer til bestemmelse for fotosynteseaktivitet og prøver efter 2, 24 og 48 timer til bestemmelse af koncentration og sammensætning af algegrupper ved HPLC (Schlüter et al. 1999). Ved hvert udtag overføres endvidere et identisk antal glas til rent åvand og inkuberes i reguleret lysmiljø (14 timers lys ved 250 µEm-2 s-1 og 10 timers mørke). Disse blev undersøgt for ændringer (genetablering) af algesammensætning efter yderlige 48 timer. I forsøget med pendimethalin kunne der ikke iagttages ”recovery” efter 48 timer i rent vand. På den baggrund gennemførtes et yderligere forsøg, hvor recovery fasen blev forlænget til 192 timer. Den fotosyntetiske aktivitet blev målt ved at tilsætte 1 µCi 14C til vials med bevoksede glasskiver ved slutningen af inkubationsperioden (1, 2, 24, 48 timer) samt efter 48 timer i rent vand. Glassene blev inkuberet ved in situ temperatur og lys i én time. Inkubationerne blev stoppet ved at tilsætte eddikesyre indtil pH var 2. Prøverne blev tørret ved 60 °C. Herefter blev 1 ml dimethylsulfoxid tilsat for at frigøre det inkorporerede 14C, og efter ½ time blev der tilsat scintillationsvæske og prøverne blev talt i scintillationstæller. Effekt på algernes biomasse og gruppesammensætning blev bestemt ved hjælp af pigmentanalyse på HPLC. Grundet en lavere følsomhed af pigmentindhold sammenlignet med fotosyntese gennemførtes pigmentforsøgene i flere tilfælde ved højere koncentrationer af pesticider. Indholdet på 3 glasskiver blev frosset i flydende kvælstof indtil analyse på HPLC. Forud for analysen blev prøverne tøet op, ekstraheret i 100% acetone i 24 timer, refiltreret gennem Teflon® sprøjtefiltre og analyseret på HPLC efter Schlüter et al. (2000). Algegrupper blev identificeret og kvantificeret ud fra diagnostiske pígmenter (Fucoxanthin = kiselalger, Klorofyl-b = grønalger, Zeaxanthin = blågrønalger). I modsætning til fotosyntesen bestemtes pigmentkoncentrationer i puljede prøver. For at få et indtryk af variationen mellem replikaterne blev der i 5 tilfælde analyseret på individuelle glasskiver. Disse analyser viste en variationskoefficient i gennemsnit på 26,0% (variationsbredde: 11-42%) for kisel- og grønalger og på 22,1% for summen af alger (klorofyl a). Til sammenligning var den gennemsnitlige variationskoefficient i fotosynteseprøverne 16,2%. Da vi ikke kender variationen indenfor den enkelte behandling kan der ikke gennemføres formelle statistiske test for forskelle mellem pesticideksponeringer og korrensponderende kontroller. Hvis vi antager at variansen er ens i eksponerede alger og kontrolforsøg, vil en gennemsnitlig variationskoefficient på 26% indenfor 3 prøver betyde, at der mindst skal være en forskel på 38% mellem eksponerede alger og kontrolalger før forskellen kan antages at være signifikant. 3.2.2 Effekt af fotosyntesehæmmereDe undersøgte fotosyntesehæmmere hexazinon, isoproturon og metribuzin havde alle en markant effekt på fotosynteseraten med entydige dosis-respons kurver for især isoproturon og metribuzin (Figur 7 & 8, Tabel 6). Hæmningen var nærmest momentan med tydelig reduktion allerede efter en times eksponering med laveste effektkoncentration mellem 0,24 µg l-1 (24 timer) og 1 µg l-1 (1 time). Effekten af hexazinon var mindre entydig: ved lave koncentrationer stimuleredes fotosyntesen ved kortvarig eksponering og ved 24 timers eksponering var effektkoncentrationen betydeligt højere sammenlignet med isoproturon og metribuzin (se Bilag A). Fotosyntesehæmmernes effekt på biomassen af kiselalger og grønalger afspejlede effekten på fotosyntesen, blot var effektkoncentrationerne højere og effekten forsinket sammenlignet med hæmningen i fotosyntesen (Fig. 9, Tabel 7).
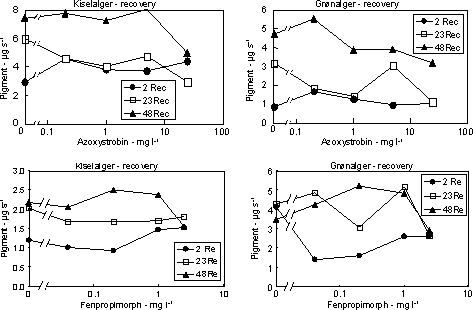
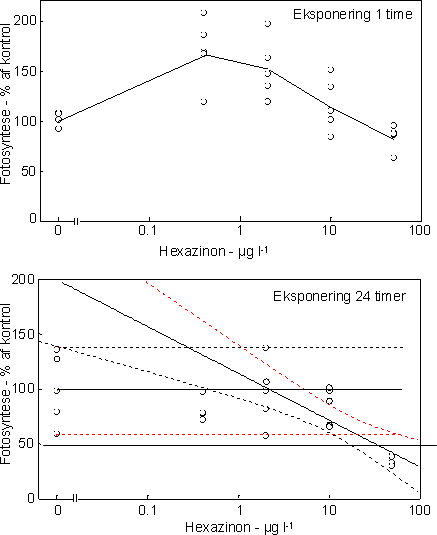
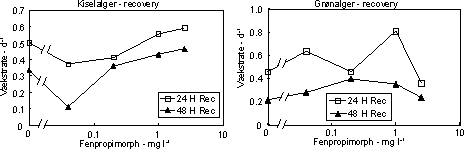
Figur 7. Dosis-respons relation for bentiske mikroalger eksponeret til isoproturon i 1 time (venstre) og 24 timer (højre). Skæringen (angivet med pil) mellem nedre konfidensgrænse for kontrolgruppen og den øvre konfidensgrænse for de eksponerede alger angiver den laveste signifikante effektkoncentration. Pilen nederst angiver den koncentration, som hæmmer fotosyntesen med 50% sammenlignet med kontrollen. Det var karakteristisk, at effekterne af fotosyntesehæmmerne var forbigående. Efter overførsel til rent vand var både fotosyntesen og biomassen af både kiselalger og grønalger næsten fuldt retableret i løbet af 48 timer (Fig. 10 & 11, Tabel 6 & 7). Blågrønalgernes koncentration var lav, men forløbet under eksponering og efterfølgende recoveryperiode var parallelt med de øvrige algegrupper (data ikke vist).
Figur 8. Dosis-respons relation for metribuzin. Skæringen (angivet med pil) mellem nedre konfidens-grænse for kontrolgruppen og den øvre konfidensgrænse for de eksponerede alger angiver den laveste effektkoncentration. En beregning af vækstraten udfra ændringerne i biomasserne under ophold i rent vand viste værdier mellem 0,05 og 0,6 d-1 med de højeste rater i forsøgene med de højeste pesticidkoncentrationer og hvor påvirkningen under eksponering havde været størst (Bilag A). Det kan således beregnes, at hvis perioden i rent vand havde været 3 dage i stedet for 2 ville der i alle forsøg havde været fuld recovery.
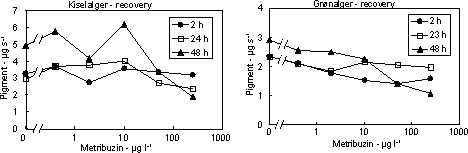
Figur 9. Effekt af fotosyntesehæmmere metribuzin (øverst) og isoproturon (nederst) på biomassen af kiselalger (venstre) og grønalger (højre) ved forskellig eksponeringsvarighed (2, 24 og 48 timer). Værdierne angiver et gennemsnit (µg diagnostisk pigment per glasskive) af 3 replikater. Bemærk at forsøgene med isoproturon ikke er de samme som vist i Figur 7.
Figur 10. Koncentration af kiselalger (venstre) og grønalger (højre) efter eksponering til metribuzin i 2, 24 og 48 timer og efterfølgende ophold i rent åvand i 48 timer. Værdierne angiver et gennemsnit af 3 replikater.
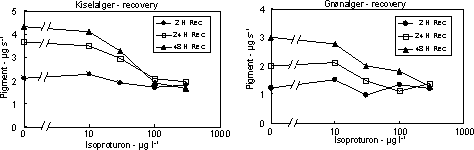
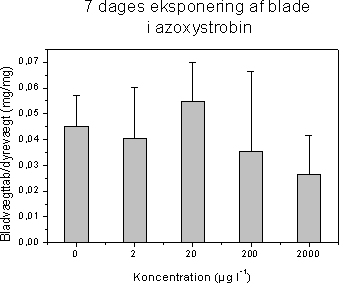
Figur 11. Koncentration af kiselalger (venstre) og grønalger (højre) efter eksponering til isoproturon i 2, 24 og 48 timer og efterfølgende ophold i rent åvand i 48 timer. Værdierne angiver et gennemsnit af 3 replikater. 3.2.3 Effekt af fungiciderBegge fungicider påvirkede fotosynteseraten hos mikroalgerne med reduktioner ved koncentrationer over og omkring 1 mg l-1 (Figur 12 & 13, Tabel 6). Efter overførsel til rent vand var fotosyntesen generelt reetableret efter 48 timer.
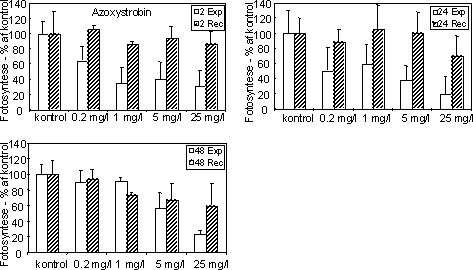
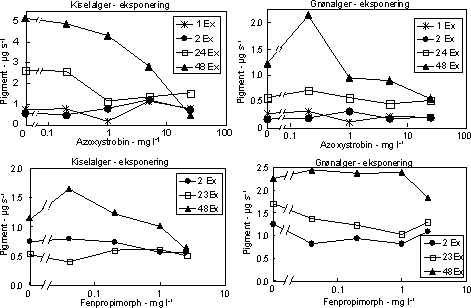
Figur 12. Fotosynteseaktivitet af bentiske mikroalger efter eksponering til azoxystrobin Søjlerne angiver den gennemsnitlige aktivitet (95 % konfidensgrænse) i procent af den korresponderende kontrol efter eksponering (hvide søjler) i 2 (øverst), 24 og 48 (nederst) timer samt efter 48 timers periode i rent vand (skraverede søjler).
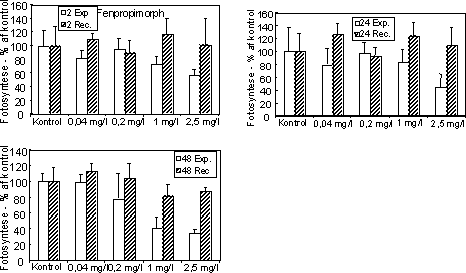
Figur 13. Fotosynteseaktivitet af bentiske mikroalger efter eksponering til azoxystrobin (venstre kolonne) og fenpropimorph (højre kolonne). Søjlerne angiver den gennemsnitlige aktivitet (95 % konfidensgrænse) i procent af den korresponderende kontrol efter eksponering (hvide søjler) i 2 (øverst), 24 og 48 (nederst) timer samt efter 48 timers periode i rent vand (skraverede søjler). Biomasseudviklingen af især kiselalger reduceredes ved høje koncentrationer af azoxystrobin og fenpropimorph (Fig. 14, Tabel 7). Effekten øgedes med eksponeringsvarigheden og var især tydelig efter 48 timers eksponering.
Figur 14. Effekt af azoxystrobin (øverst) og fenpropimorph (nederst) på biomassen af kiselalger (venstre) og grønalger (højre) ved forskellig eksponeringsvarighed (1, 2, 24 og 48 timer). Værdierne angiver et gennemsnit af 3 replikater. Efter overførsel til rent vand var biomasserne næsten fuldt reetableret i løbet af 48 timer (Fig. 15, Tabel 7). Det skyldes høje vækstrater under recoveryfasen hos de alger, som havde været eksponeret til de højeste koncentrationer i længst tid (se Bilag A).
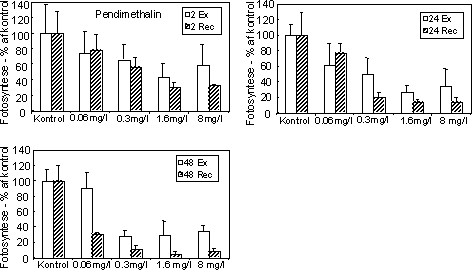
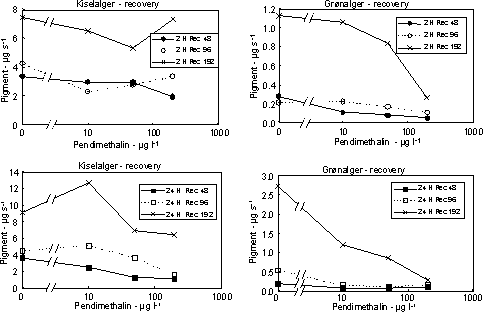
Figur 15. Koncentration af kiselalger (venstre) og grønalger (højre) efter eksponering til azoxystrobin (øverst) og fenpropimorph (nederst) i 2, 24 og 48 timer og efterfølgende ophold i rent åvand i 48 timer. Værdierne angiver et gennemsnit af 3 replikater. 3.2.4 Effekt af celledelingshæmmerDer blev gennemført 2 forsøgsserier med herbicidet pendimethalin, som virker ved hæmning af celledelingen i planter. Den første forsøgsrække viste en monoton reduktion i fotosynteseaktiviteten i koncentrationsintervallet 0,06 – 8 mg l-1 og et ufuldstændig recovery selv efter 2 timers eksponering (Fig. 16). Efter eksponering i 48 timer forstærkedes reduktionen i recoveryperioden ved alle undersøgte koncentrationer. En tilsvarende effekt og tidslig udvikling kunne også ses i biomassen af kiselalger, blågrønalger og især grønalger (se Bilag A).
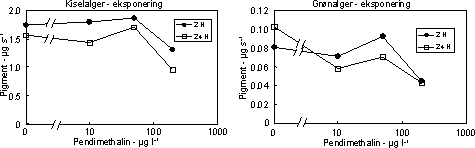
Figur 16. Fotosynteseaktivitet af bentiske mikroalger efter eksponering til pendimethalin Se tekst til Figur 12 for forklaring af symboler. På grund af mangel på recovery i første forsøg gennemførtes yderlige et forsøg ved lavere koncentrationer (10 – 200 µg l-1) og med en forlænget periode i rent vand (192 timer). For at undgå næringsbegrænsning blev der skiftet næringsberiget vand hver 2. dag under recoveryperioden. I eksponeringsperioden (2 – 24 timer) var effekterne på algebiomasserne moderate (Fig. 17), men effekten af pendimethalin på biomasseudviklingen hos grønalger blev tydelig under perioden i rent vand. Således var der ikke opnået fuld recovery af grønalger efter 192 timer i forsøget med 24 timers eksponering til 10 µg l-1 (Fig. 18, Tabel 7). At kiselalgerne er mindre følsomme end grønlalgerne kan skyldes, at kiselalgernes cellevæg (ydre skelet) udgør en mere effektiv barriere end grønalgernes ”nøgne” cellevæg. I terrestiske planter sker optagelsen af pendimethalin alene gennem skud og rodspidser, mens optagelsen gennem blade er negligibel.
Figur 17. Effekt af pendimethalin på biomassen af kiselalger (venstre) og grønalger (højre) efter eksponering i 2 og 24 timer. Værdierne angiver et gennemsnit af 3 replikater.
Figur 18. Koncentration af kiselalger (venstre) og grønalger (højre) efter eksponering til pendimethalin i 2 timer (øverst) og 24 timer (nederst) og efterfølgende ophold i rent åvand i 48, 96 og 192 timer. Værdierne angiver et gennemsnit af 3 replikater. Tabel 6. Effekt af pesticider på fotosynteseaktivitet hos perifytalger efter eksponering til 1 (2), 24 og 48 timer (Eksponering) og efterfølgende ophold i rent vand i 48 timer (Recovery). NEC: ”no effect concentration”; EC50 og 95 % konfidensinterval (i parantes) angivet. - = ikke forskellig fra kontrol, s = stimulering (signifikant højere aktivitet end i kontrol). Alle værdier i µg l-1.
Tabel 7. Effekt af pesticider på biomasseudvikling hos perifytalger efter eksponering til 1 (2), 24 og 48 timer (Eksponering) og efterfølgende ophold i rent vand i 48 timer (Recovery). LOEC = laveste effektkoncentration bestemt hvis afvigelse fra korresponderende kontrol var større end 38% (se tekst). (K = kiselalger; G = grønalger; B = blågrønalger). Alle værdier i µg l-1.
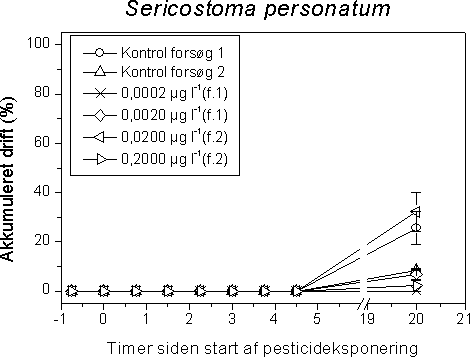
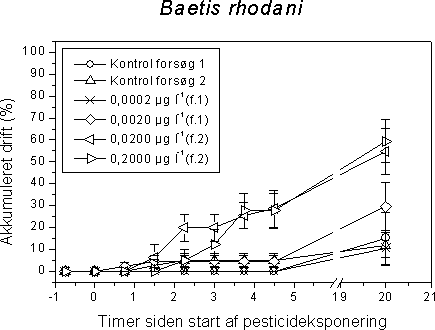
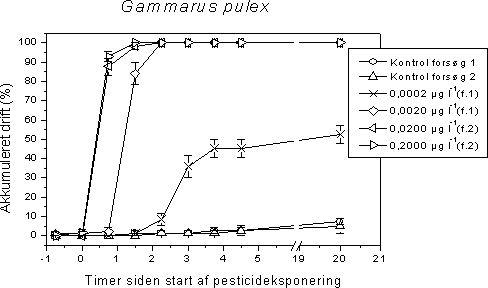
4 Effekter af pesticider på invertebrater i vandløbForsøgsdyrene blev udvalg således at de repræsenterede væsentlige elementer af den danske vandløbsinvertebratfauna. 70-80% af de taxa der findes i vandløb er insekter, og derfor er insekterne tilsvarende overrepræsenteret som forsøgsorganismer. De udsøgte organismer er desuden udvalgt så de varierer med hensyn til taksomisk gruppe, livscyklus, fødefunktionalitet, trofisk niveau og tolerance over for organisk forurening. Sidstnævnte kriterium betyder at de undersøgte organismer er indplaceret bredt i Dansk Vandløbsfaunaindeks (DVFI) (Miljøstyrelsen, 1998), der bruges som biologisk bedømmelsesmetode i det nationale overvågningsprogram (NOVA2003) på 1050 stationer samt flere steder i det regionale tilsyn. Derudover dækker de udvalgte forsøgsorganismer et bredt spektrum med hensyn til tolerance over pesticider. Wogram & Liess (2001) har fundet følgende rangorden med hensyn til de i vore undersøgelser anvendte invertebraters pesticidfølsomhed: Slørvinger > krebsdyr > døgnfluer > tovinger > vårfluer > snegle. Tabel 8 viser forekomst og densitet af hovedparten af de anvendte forsøgsorganismer i de 1050 NOVA 2003 overvågningsvandløb. Tabel 8. Fundfrekvens og tæthed på 1050 NOVA 2003 stationer i 2001 af hovedparten af de i projektet undersøgte invertebrater. Det taksonomiske niveau der anvendes i NOVA2003 er som minimum det i DVFI foreskrevne (Miljøstyrelsen, 1998), men enkelte amter udfører en fuld artsbestemmelse. Derfor indeholder tabellen både DVFI niveauet samt et højere taksonomisk niveau (slægt og art) hvor dette har forekommet. Hvad dette betyder for tolkningen af tabellen kan eksemplificeres for Gammarus pulex: Gammarus er identifikationsniveauet i DVFI, men en del amter har bestemt til artsniveau (G. pulex). Hovedparten af de Gammarus der er fundet på de 81% af stationerne vil formentlig være G. pulex da den anden art, G. lacustris, er langt mindre udbredt i vandløb. Imidlertid vides det kun med sikkerhed at der er G. pulex på 49% af stationerne.
4.1 Anvendte invertebraterGammarus pulex, ferskvandstangloppen, er et krebsdyr (Malacostraca; Amphipoda). Denne organisme blev primært valgt på grund af dens gode laboratoriegenskaber, samt dens store udbredelse i danske vandløb (Tabel 8). Ifølge undersøgelser foretaget af Fyns Amt (2000) udgør G. pulex ikke mindre end 25% af den samlede smådyrsfauna i vandløb og kan antage en tæthed på hen ved 8.000 individer pr. m². Dens hyppige forekomst betyder da også, at den udgør et meget vigtigt fødegrundlag for især fisk og fugle. Den udmærker sig desuden ved at være velbeskrevet i litteraturen, hvorfor sammenligningsgrundlaget for forsøg med G. pulex udsat for andre miljøfremmede stoffer er stort. G. pulex er et væsentligt element i DVFI, hvorfor den tillægges stor økologisk betydning i forbindelse med kvalitetsbedømmelse af danske vandløb. G. pulex vil findes både i rene og moderat forurenede vandløb. Den indgår i Nøglegruppe 3 og som positiv diversitetsgruppe i DVFI og er meget væsentlig for at kunne at skelne de let og moderat forurenede vandløb fra de stærkere forurenede vandløb. Fødefunktionelt er G. pulex meget plastisk men betegnes overvejende som en ituriver/samler, der primært ernærer sig ved nedfaldne blade og andet dødt organisk materiale. G. pulex opholder sig ved bunden eller brinkerne, hvor den søger føde. G. pulex er meget mobil og findes hyppigt i drift. Desuden udviser den meget udpræget opstrøms migration (positiv rheotaxi) for at kompensere for driften. G. pulex er derfor en god kolonisator, sålænge den ikke møder barrierer mod opstrømsmigration i form af f.eks. rørunderføringer ved veje. G. pulex udgør en nøglerolle i danske vandløb, da den kvantitativt er den mest dominerende invertebrat. Den udgør fødegrundlag for højere trofiske niveauer og har desuden en nøglerolle som bio-ingeniører i omsætningen af groft organisk stof til fint. Det fine organiske stof indgår som et væsentligt element i detritivorfødenettet, og har stigende betydning ned gennem vandløbet. Asellus aquaticus er som Gammarus et krebsdyr (Malacostraca; Isopoda). Hovedincitamentet for anvendelse af A. aquaticus er, at den ligesom G. pulex er vidt udbredt i de fleste vandløb. Asellus indtager en vigtig rolle i DVFI, blot med den forskel at dens forekomst anses at indikere en høj forureningsgrad. Fødefunktionelt betegnes den som ituriver/samler, hvorfor den i lighed med G. pulex udfylder en vigtig post som omsætter af organisk materiale. Naturligt forekommer A. aquaticus blandt makrofyter i kantzonen i større vandløb. Imidlertid vil A. aquaticus findes i flere vandløbstyper, når disse er påvirkede af organisk forurening. A. aquaticus findes på 63% af NOVA2003 stationerne (Tabel 8), hvilket bl.a. afspejler at halvdelen af disse vandløb ikke har en tilfredsstillende biologisk vandløbskvalitet (faunaklasse 4 eller lavere). A. aquaticus opholder sig primært på bunden af et vandløb, hvor den kan forekomme i meget store individantal ( 10.000 m²) i stærkt belastede vandløb. Isoperla er en slørvinge (Plecoptera) tilhørende familien Perlodidae. Isoperla tilhører gruppen Setipalpia, der er prædatorer. Isoperla er valgt fordi den befinder sig på et højere trofisk niveau i forhold til andre invertebrater der er indgået i forsøgsserierne, og fordi det er en meget rentvandskrævende organisme, der indgår i DVFI som Nøglegruppe 1 taxsa og positiv diversitetsgruppe. Isoperla grammatica findes i hele landet, mens den anden danske art, I. difformis, kun er fundet i Jylland. Isoperla er fundet i 7% af NOVA 2003 vandløbene (Tabel 8). De lever ikke i de mindste vandløb, men er ellers udbredt i hele vandløbskontinuumet sålænge vandkvaliteten tillader det. Isoperla har en 1-årig livscyklus og emergerer i maj-juni. Leuctra er slørvinger (Plecoptera; Leuctridae), der lever af at samle fint organisk stof (FPOM) og af at iturive blade og lign. (CPOM). Leuctra er primært knyttet til de mindre vandløb og de er forholdsvis almindelige (Tabel 8). Der er 4 arter i danske vandløb og de kræver alle gode iltforhold. Leuctra indgår således i Nøglegruppe 1 og tæller som positiv diversitetsgruppe i DVFI. De fleste Leuctra, men især Leuctra nigra, er imidlertid tolerante overfor lav pH i det omgivende vand. Alle Leuctra arterne har en 1-årig livscyklus med undtagelse af L. nigra, der har en 2-årig livscyklus. To arter emergerer i april-maj (hippopus og nigra) og to arter september-oktober (fusca og digitata). Der vil derfor være store Leuctra nymfer i vandløbene det meste af året såfremt vandkvaliteten tillader det. Baetis (Ephemeroptera; Baetidae) er en meget udbredt døgnflueslægt i danske vandløb (Tabel 8). Den vil forekomme fra de øvre kildebække til store vandløb. Baetis ernærer sig som skraber af alger og biofilm på sten og makrofyter. Desuden ernærer Baetis sig som samler af FPOM. Der er 10 arter i Danmark, hvoraf B. rhodani og B. vernus er de mest udbredte. B. rhodani er forholdsvis tolerant overfor forringede iltforhold sålænge vandet strømmer og kan derfor være hyppig i moderat forurenede vandløb. B. rhodani har to generationer om året, mens nogle af de øvrige Baetis-arter formentlig er 1-årige. Sericostoma personatum (Trichoptera; Sericostomatidae) er en typisk ituriver i skovvandløb. Den vil kunne forekomme i mere åbne vandløb sålænge temperaturen ikke bliver for høj. S. personatum lever i områder med lav strømhastighed, og er derfor særlig følsom for lavt iltindhold i det omgivende vand. Sericostomatidae indgår i Nøglegruppe 1 og tæller som en positiv diversitetsgruppe i DVFI. Sericostomatidae er af de undersøgte organismer den mest hyppigt forekommende Nøglegruppe 1 organisme i NOVA 2003 vandløbene (Tabel 8). S. personatum har en 2-årig livscyklus og emergerer om foråret. Agapetus fuscipes er en vårflue (Trichoptera; Glossosomatidae), der lever af at skrabe alger og andet biofilm af sten. Findes i Danmark primært i mindre vandløb men er forholdsvis sjælden (Tabel 8). A. fuscipes kræver gode iltforhold og familien Glossosomatidae indgår både i Nøglegruppe 1 og som positiv diversitetsgruppe i DVFI. A. fuscipes har en 1-årig livscyklus og kan emergere en stor del af året. Hydropsyche (Trichoptera; Hydropsychidae) har netspindende og fritlevende larver, der meget udbredte i danske vandløb (Tabel 8). Den ernærer sig ved at spinde et silkenet hvormed den fanger forskellige suspenderede partikler som tranporteres med vandet. Pga. denne fødestrategi er Hydropsyche afhængig af groft, stabilt substrat og en kontinuerlig vandstrøm. Desuden er tætheden af Hydropsyche afhængig af den mængde føde, der føres med vandløbet. Hydropsyche vil typisk findes på de mellemste vandløbsstrækninger, og kan findes i masseforekomst i søafløb, hvor de lever af de planktoniske organismer o. lign. der kommer fra søen. Hydropsyche er generelt tolerante overfor dårlig vandkvalitet og vil kunne findes i moderat forurenede vandløb. Der 7 arter af slægten Hydropsyche i Danmark, hvoraf en art (H. augustipennis) er udbredt og almindelig. Hydropsyche-arterne er formentlig alle 1-årige og flere har en lang flyveperiode, der strækker sig fra maj til august. Chironomus riparius er en dansemyg (Chironomidae) tilhørende underfamilien Chironomini. Som repræsentant for organismer med en fuldstændig forvandling og dermed også et terrestrisk livsstadie blev C. riparius valgt. Arten er naturligt forekommende i danske vandløb, men udmærker sig specielt ved at være egnet til at holde i laboratoriekultur, bl.a. fordi den har en meget kort generationstid. Sidstnævnte forhold betyder også at arten er særligt velegnet til gennemførsel af komplette livscyklus tests. Efter forgæves at have forsøgt at indsamle Chironomus fra felten blev vi i stedet nødt til at importere C. riparius fra en allerede eksisterende laboratoriekultur (se senere beskrivelse). Ifølge DVFI anses forekomst af Chironomus, i lighed med Asellus, at indikere en høj organisk forureningsgrad. Der er således tale om en meget forureningstolerant art, der tåler endog meget lave iltkoncentrationer. Vi fandt det imidlertid relevant at undersøge om denne tilsyneladende høje tolerancetærskel ligeledes gør sig gældende overfor forurening med pesticider. C. riparius er et udpræget bunddyr, der lever både i og af sediment, hvorfor den fødefunktionelt betegnes som en samler. Der er i visse tilfælde observeret forekomster på op til 50.000 individer pr. m². Imidlertid har de sidste årtiers spildevandsrensning bevirket, at Chironomus ikke længere er ret udbredt i danske vandløb og de blev således kun fundet på 6% af NOVA 2003 stationerne i 2001. Prodiamesa olivacea hører til underfamilien Prodiamesinae (familie: Chironomidae), der er en lillle underfamilie med få slægter og arter i Danmark. P. olivacea lever i sedimentet i vandløb. Den kræver forholdsvis gode iltforhold og sedimenter med stort indhold af organisk stof, som den ernærer sig af (sedimentæder). Den kan være meget talrig i moderat forurenede vandløb med store forekomster af fint organisk stof, men vil ikke findes i stærkt forurenede vandløb som f.eks. Chironomus. Bithynia tentaculata er en forgællesnegl tilhørende familien Hydrobiidae. Da vi ikke kunne få indsamlet et tilfredsstillende stort antal individer af huesneglen Ancylus fluviatilis blev denne almindeligt forekommende sumpsnegl anvendt som repræsentant for de græssende organismer. Den blev primært valgt udfra praktiske hensyn, idet den optrådte i stort antal på indsamlingslokaliteten. Den var således en meget hyppig bifangst ved indsamling af de øvrige organismer. B. tentaculata er ikke omfattet af DVFI, men det betyder ikke at der er tale om en økologisk ubetydelig art, idet den, med dens store individtæthed taget i betragtning, kan opfylde en vigtig rolle som græsser og skraber af diverse algetyper. B. tentaculata skraber biofilm af makrofyter og var i indsamlingsvandløbet især knyttet til Elodea (vandpest). Vi fandt det desuden relevant at vælge en art med en meget begrænset og langsom bevægelsesadfærd, der bevirker at den ikke umiddelbart kan forlade et forurenet område. 4.2 Standardtests til bestemmelse af akut toksicitet (LC50)Alle de arter som blev udvalgt til screeningtests er almindeligt forekommende vandløbsinvertebrater i Danmark, og de fleste spiller en vigtig rolle i DVFI til bedømmelse af et vandløbs forureningsgrad. Kriterierne for udvælgelsen af de respektive pesticider, samt ved hvilket koncentrationsniveau forsøgene skulle gennemføres er opsummeret i nedenstående:
Selvom de tilgængelige målinger over pesticidforekomster i danske vandløb sandsynligvis ikke repræsenterer den maksimale koncentration som kan forekomme i pågældende vandløb, er det værd at notere, at koncentrationer på over 2000 ng l-1 er yderst sjældne for noget pesticid (DMU 2002). Ikke desto mindre inkluderede alle screeningtests koncentrationer på minimum op til 2000 ng l-1, og i flere tilfælde endda en faktor 1000 højere. For mange af arterne var det imidlertid ikke muligt at bestemme en LC50-værdi på grund af lav eller slet ingen dødelighed ved den højeste koncentration. Hvis der ikke blev observeret dødelighed ved 2000 ng l-1 eller derover i løbet af en kontinuerlig eksponeringsperiode på 24 timer (eller længere) blev det antaget, at det pågældende pesticid ikke at havde en målbar effekt ved kortere pulseksponeringer med lignende koncentrationer. Derfor blev de pågældende art-pesticid kombinationer nedprioriteret med henblik på yderligere forsøg. Som det vil fremgå af resultaterne fra de udførte screeningstests viste Gammarus pulex sig konsekvent at være den mest sensitive af alle de testede arter. Af alle de testede pesticider var esfenvalerat klart det mest toksiske overfor ferskvandsinvertebrater efterfulgt af azoxystrobin. Ingen af de øvrige pesticider, der blev testet, havde målbare effekter på arter testet ved miljørealistiske koncentrationer. I bestræbelsen på at bestemme effekterne af miljørealistiske pulsekponeringer på andre endpoints end dødelighed, blev G. pulex herefter valgt som nøgleorganisme for videre studier til bestemmelse af følsomheden af forskellige livsstadier. Af de screenede arter var det udelukkende muligt (indenfor dette projekts begrænsede tidsramme) at udføre fulde livscyklusstudier med Chironomus riparius. En yderligere argumentation for at fokusere på C. riparius er, at det er den eneste sedimentlevende invertebrat som regelmæssigt (under kronisk eksponering) bliver anvendt til risikovurdering af pesticider. 4.2.1 MetodeDer blev udført screeningstests til bestemmelse af LC50-værdier med 5 forskellige arter udsat for 5 forskellige pesticider. De anvendte arter og pesticider fremgår af oversigtsskemaet under resultatafsnittet. Som nævnt indledningsvis viste den mest følsomme organisme sig at være G. pulex, hvorfor fokus under den følgende metode- og resultatgennemgang er lagt på denne art. Gammarus pulex Asellus aquaticus Bithynia tentaculata Chironomus riparius Prodiamesa olivacea Statistik 4.2.2 ResultaterSom det fremgår af oversigtstabellerne (Tabel 9 og 10) for alle de testede arter og pesticider var både voksne og juvenile individer af G. pulex mest følsomme, specielt overfor esfenvalerat, hvorfor der primært fokuseres på beskrivelse af data fra disse screeningstests. En lang række af de øvrige screeninger gav ikke anledning til forøget mortalitet hos de testede organismer, selv ikke ved urealistisk høje koncentrationsniveauer, hvorfor disse ikke kommenteres yderligere. Gammarus pulex eksponeret til esfenvalerat Overlevelsen af voksne var faldende med eksponeringstid mellem 24 og 48 timer, men ikke mellem 48 og 96 timer. LC50-værdier blev estimeret til ca. 0,34 µg l-1 efter 24 timers eksponering og 0,14 µg l-1 efter 48 og 0,13 µg l-1 efter 96 timers eksponering. Hunnerne så ud til at være mere sensitive end hanner og forskellen var specielt synlig efter 24 timers eksponering (Tabel 11). Tabel 9. Estimerede LC50-værdier (µg l-1) for voksne individer af krebsdyr og snegle. Testene med dansemyg blev gennemført med store individer (10-12 mm). Tallene i parenteserne angiver 95% konfidensintervaller.
Selvom antallet af unger der blev frigivet i løbet af den første time under eksponeringen ikke blev talt, blev disse observeret i live i grupperne op til 0,1 µg l-1, mens ungerne i grupperne 0,5, 1,0 og 2,0 µg l-1 alle var døde. I kontrollen var antallet af intakte par lavere efter 24 timer og resten af forsøgsperioden ud sammenlignet med 1 times eksponering. I modsætning hertil var der flere af de forstyrrede par i grupperne eksponeret til henholdsvis 0,01 og 0,05 µg l-1 der blev gendannet med tiden. Dette var imidlertid ikke tilfældet i de højere eksponeringsgrupper. Tabel 10. Estimerede LC50-værdier (µg l-1) for juvenile individer af krebsdyr og snegle. Testene med dansemyg blev gennemført med små individer (1-3 mm). Tallene i parenteserne angiver 95% konfidensintervaller.
Som det fremgår af resultatet af LC50-forsøg med eksponering af sammenhængende par viste den prækopulative fase sig at være yderst sensitiv overfor esfenvalerat. Derfor blev netop denne forsøgsparameter underkastet yderligere opmærksomhed i de efterfølgende undersøgelser til bestemmelse af følsomheder i forskellige livsstadier hos Gammarus pulex. Ligesom det var tilfældet for voksne individer blev der hos de juvenile observeret faldende overlevelse i tiden mellem 24 og 48 timer efter eksponeringen, men ikke mellem 48, og 72 eller 96 timer efter eksponeringen. De juvenile individer var mere sensitive end de voksne individer med estimerede LC50-værdier på 0,24 µg l-1 efter 24 timer, 0,14 µg l-1 efter 48 og 96 timer. Ingen af de overlevende individer i gruppen eksponeret til 0,5 µg l-1 var aktive efter 24 timers eksponering i modsætning til gruppen eksponeret til 0,1 µg l-1, hvor ca. 50 % var aktive og gruppen eksponeret til 0,05 µg l-1 hvor ca. 75 % af de overlevende var aktive. Tabel 11. Akut toksicitet af esfenvalerat på voksne Gammarus pulex. Der var 5 par voksne og 3 replikater per eksponeringsgruppe (n=30 pr gruppe).
Mortalitet i strømrender Tabel 12. Sammenligning af LC50,96 timer, (ng l-1) hos krebsbyr og insektlarver i strømakvarier. * Pirimicarb udløste ingen dødelighed ved den højest anvendte koncentration (1 mg l-1).
4.3 Effekt af eksponeringsvarighedFor at undersøge letale effekter af bekæmpelsesmidler i vandløb som funktion af eksponeringsvarigheden blev der udført laboratorieforsøg med invertebrater, primært Gammarus pulex, eksponeret for forskellige pesticider. Den eksperimentelle tilgang har inkluderet undersøgelser af sammenhængen mellem eksponeringsvarigheden og toksicitet samt undersøgelser af eventuelle forsinkede effekter af pesticider efter ophørt eksponering. Med stigende eksponeringsvarighed vil der ske et stigende optag af pesticid i organismen hvorfor effektkoncentrationen vil være faldende over tid. For let vandopløselige stoffer vil en ligevægt mellem optag og effekt indstille sig relativt hurtigt, og herefter vil effektkoncentrationen teoretisk set være stabil. Organismerne vil dog være udsat for et øget fysiologisk stress som funktion af eksponeringsvarigheden, hvorfor en forøget toksicitet kan forekomme efter at ligevægt mellem optag og effekt er indtruffet. Lipofile stoffer optages langsommere end vandopløselige stoffer, og en ligevægt vil først indtræffe efter længere tids eksponering. Lipofile stoffer vil have tendens til at bioakkumulere i organismernes fedtvæv, hvor koncentrationen vil overstige den i det omgivende miljø. Derfor kan toksiciteten af lipofile stoffer forventes at falde over en længere periode sammenlignet med vandopløselige stoffer. Ligeledes kan det forventes at bioakkumulerbare stoffer i højere grad end vandopløselige stoffer vil kunne medføre forsinkede effekter efter endt eksponering som følge af det akkumulerede pesticid. 4.3.1 MetodeDe gennemførte tests har i videst muligt omfang fulgt en standardiseret protokol for undersøgelse af akut toksicitet i det akvatiske miljø; OECD 202; Daphnia sp; Acute Immobilisation Test (OECD 1984). Der foreligger ikke egentlige guidelines til gennemførelse af test med de her anvendte arter, men principperne i OECD guideline nr. 202 er fulgt i overordnede træk. De gennemførte forsøg har på nogle områder afviget fra standardtesten; dels er relationen mellem toksicitet og eksponeringstid undersøgt over en længere serie af eksponeringstider, og dels er testorganismerne efter endt eksponering blevet overflyttet til rent vand og holdt under observation i op til 14 dage. Testorganismer blev indsamlet i vandløb af DMU og bragt til DHI, hvor de blev akklimatiseret under betingelser svarende til dem i forsøget i mindst 3 dage inden forsøgets start. Inden forsøget blev dyrene sorteret efter størrelse og det blev tilstræbt at anvende mindre individer af samme størrelse og dermed nogenlunde samme alder, idet optag og dermed effekt af kemiske stoffer bl.a. antages at afhænge af overflade-volumen forholdet samt udviklingsstadie af forsøgsorganismerne. Eksponeringskoncentrationerne i de enkelte forsøg blev fastlagt på baggrund af data for akut toksicitet for akvatiske organismer fundet i litteraturen og/eller databaser. Pesticiderne blev maksimalt testet i koncentrationer op til 5-10 mg l-1. Højere koncentrationer end dette niveau vurderedes ikke at være miljørealistisk. Eksponeringsforsøgene blev udført i åvand fra Brådebæk (Hørsholm). Testorganismerne blev eksponeret for en fortyndingsrække af det relevante pesticid i variable tidsforløb, hvorefter de blev overført til rent vand. Herefter er udviklingen af toksiciteten fulgt i en periode op til 14 døgn. Ca. 150 dyr blev eksponeret for hver testkoncentration i 20 L akvarier med svag gennembobling. Eksponeringstiderne var: ½; 1; 2; 4; 6; 24; 48; 72; 96; 168, 240 og 336 timer. Til hver eksponeringstid og –koncentration blev der overført 10 dyr til rent vand i 250 ml krystallisationsskåle forsynet med svag gennembobling gennem kanylespids. Forsøgene blev gennemført semi-statisk med to ugentlige vandskift i både eksponeringskar og krystallisationsskåle således at testkoncentrationerne i videst muligt omfang blev opretholdt i testperioden. Dødelighed samt synlige ændringer i adfærd blev bestemt, dels til de enkelte tider i løbet af eksponeringsperioden og dels hver 24. time efter at dyrene blev overført til rent vand. Testorganismerne blev fodret med Tetramin under eksponeringen såvel som efter overførsel til rent vand. Ilt og pH blev målt løbende både under eksponering og i rent vand. Eksponeringerne blev udført i klimarum ved ca. 15 °C, dog blev forsøg med vårfluelarver og juvenile slørvinger udført ved ca. 5 °C for at begrænse udviklingshastigheden og således sikre at dyrene ikke metamorfoserede i løbet af forsøget. Det blev tilstræbt at undersøge et bredt spektrum af pesticider for at afdække effekterne af pesticider med forskelligartede virkemekanismer inden for de forskellige grupper af insekticider, fungicider og herbicider. Der blev testet 7 pesticider i forsøg med Gammarus pulex: esfenvalerat, pendimethalin, azoxystrobin, pirimicarb, prochloraz, dimethoat og ioxynil. Esfenvalerat blev yderligere testet på Isoperla grammatica og Agapetus sp. I det omfang det har været muligt er eksperimenterne blevet udført med radioaktivt mærkede pesticider (esfenvalerat, pendimethalin og ioxynil). Ved brug af radioaktivt mærkede stoffer kan man dels følge den aktuelle eksponeringskoncentration og dels opnå informationer om akkumulering og koncentration af stoffet i testorganismerne. LC50 værdier for pesticidernes akutte letale effekt som funktion af eksponeringstid blev beregnet ved hjælp af probit analyse (Probit analyse, version 2.3, 1990). 4.3.2 ResultaterToksicitet under eksponering Eksponering af Gammarus pulex for de valgte pesticider viste som forventet en stigende mortalitet ved øget eksponeringsvarighed. For esfenvalerat og azoxystrobin, som var de mest toksiske pesticider, indtrådte en dødelig effekt i de højest testede koncentrationer allerede efter få timers eksponering (Fig. 19). For de øvrige pesticider sås letale effekter først senere i eksponeringsforløbet, hvorfor effektkoncentrationer (LC50) først kunne estimeres efter flere døgns eksponering. Idet eksponeringsforsøgene skulle tilvejebringe informationer om toksiciteten i et relativt langt tidsinterval (0-14 dage) var det vanskeligt at vælge koncentrationsserier som både medførte effekter på en meget kort tidsskala (minutter-timer) uden samtidig at slå hele testpopulationen ihjel inden for få døgn. Esfenvalerat havde en markant højere toksicitet overfor Gammarus pulex sammenlignet med de øvrige pesticider. Letale effekter af esfenvalerat gjorde sig gældende i koncentrationsområdet ng l-1 mens effekter af de øvrige pesticider sås i koncentrationsområdet mg l-1. Pirimicarb medførte ikke toksiske effekter overfor Gammarus pulex i de testede koncentrationer (0-5 mg l-1). LC50 blev bestemt som funktion af eksponeringstiden for de enkelte pesticider. I Figur 20 ses sammenhængen mellem eksponeringstid og LC50 for esfenvalerat, azoxystrobin, pendimethalin, prochloraz, dimethoat og ioxynil. I Bilag B er de eksakte værdier for både LC50 og LC10 koncentrationer angivet. Figur 20 illustrerer hvorledes pesticidernes toksicitet øges som funktion af eksponeringstiden. Toksicitetsforøgelsen sker typisk inden for de første 2 døgn efter begyndende toksicitet observeres, og efterfølgende flader toksicitetskurven ud. Dette skyldes bl.a. at der kræves en vis eksponeringsvarighed før der indtræder en ligevægt mellem pesticid koncentration i organismen og eksponerings-koncentration. Teoretisk set vil denne ligevægt indstille sig hurtigere for vandopløselige stoffer sammenlignet med lipofile stoffer. Lipofile stoffer kan akkumuleres over en længere periode, og effektkoncentrationen vil derfor falde over en længere tidsskala. De her undersøgte pesticider har vidt forskellige opløseligheder. Esfenvalerat, pendimethalin, prochloraz og ioxynil kan karakteriseres som lipofile stoffer (log Kow > 3, se Tabel 4) mens azoxystrobin og især dimethoat og pirimicarb er relativt vandopløselige. Effekten af eksponeringsvarighed synes dog ikke at variere for de hhv. opløselige og lipofile pesticider. LC50 stabiliseres ikke markant hurtigere for de vandopløselige pesticider azoxystrobin og dimethoat sammenlignet med de øvrige, mere lipofile pesticider. For alle 6 pesticider ses imidlertid en tendens til at toksiciteten vedbliver med at falde, hvilket kan skyldes det konstante fysiologiske stress dyrene er udsat for under eksponeringen. De anvendte pesticider udgøres af både insekticider, herbicider og fungicider. For at sammenligne den akutte toksicitet for de forskellige typer af pesticider er de beregnede LC50 værdier efter endt eksponering vist i Tabel 13. Således fås følgende rangordning af de 7 pesticiders toksicitet: esfenvalerat > azoxystrobin > pendimethalin > prochloras > demethoat > ioxynil > pirimicarb. Tabel 13. Sammenlignende toksicitet af de anvendte pesticider efter 14 dages eksponering
Det fremgår af Tabel 13 at de undersøgte insekticider ikke konsekvent var de mest toksiske af pesticiderne på trods af at insekticiderne i kraft af deres egenskaber kunne forventes havde en mere specifik virkning på invertebrater end herbicider og fungicider. For pendimethalin, prochloraz, dimethoat, ioxynil og pirimicarb indtraf der ikke akutte letale effekter inden for det første døgn i de testede koncentrationer. Da de testede koncentrationer generelt er meget høje i forhold til hvad der er målt i danske vandløb kan der ikke forventes umiddelbare toksiske effekter ved realistiske koncentrationer. Forsinket toksicitet Især to faktorer gør tolkningen af de forsinkede effekter hos Gammarus pulex vanskelige. I de glas hvor dyrene opholdt sig i rent vand blev der observeret en stigende grad af kannibalisme med tiden, trods regelmæssig fodring igennem hele forsøget. De individer der blev spist af deres artsfæller blev udeladt fra beregningerne. Dertil medfører forsøgsdesignet at de individer der overføres til rent vand kan have en anderledes tolerance overfor det anvendte pesticid i forhold til den samlede testpopulation. Ved koncentrationer der ikke medfører akutte toksiske effekter (lave koncentrationer, kort eksponeringsvarighed) må individer der udtages til rent vand formodes at have en tolerance der er repræsentativ for hele populationen. Ved koncentrationer hvor der observeres en høj akut mortalitet (høje koncentrationer, lang eksponeringsvarighed) må de individer der overføres til rent vand formodes at have en højere tolerance overfor det anvendte pesticid, idet de mest følsomme individer allerede er døde. I relation til miljørealistiske eksponeringsscenarier er det mest relevant at se på de kortvarige eksponeringer hvor koncentrationen af det givne pesticid ikke medfører synlige toksiske effekter, og hvor de individer der overføres til rent vand må formodes at have en følsomhed svarende til gennemsnittet for testpopulationen. Eksponeringer på op til 14 dage er væsentligt længere end den relevante tidsskala for eksponering i vandløb (se Fig. 6). For at undersøge om kortvarige eksponeringer kan medføre forsinkede effekter er det således valgt at fokusere på sub-letale eksponeringskoncentrationer, dvs. koncentrationer der ikke medførte mortalitet under eksponeringen. For de pesticider hvor der blev observeret en forsinket mortalitet i rent vand som følge af eksponering i op til 24 timer blev LT50rent vand beregnet. LT50rent vand angiver det ”mediane letale tidspunkt”, hvor 50% af individerne i rent vand forventes at dø som følge af en kortvarig eksponering. Forsinkede effekter af kortvarige, sub-letale eksponeringer gjorde sig gældende for esfenvalerat, azoxystrobin og prochloraz. For pendimethalin, dimethoat, ioxynil og pirimicarb blev der ikke observeret forsinkede effekter som følge af korttidseksponering (0-24 timer). Esfenvalerat medførte ingen akutte effekter i op til 4 timers eksponering (0-1 µg l-1), men i den efterfølgende periode i rent vand indtraf en stigende dødelighed hos dyr eksponerede for hhv. 0,25, 0,5 og 1 µg l-1. Figur 21 viser hvorledes mortaliteten øges hos individer der er overført til rent vand efter 0-4 timers eksponering for 1 µg l-1 esfenvalerat.
Figur 21. Forsinket mortalitet hos Gammarus pulex efter eksponering til 1 µg l-1 esfenvalerat i hhv. 0,5, 1, 2 og 4 timer For Gammarus pulex eksponeret for 1 µg l-1 esfenvalerat blev LT50rent vand beregnet til 16,4 dage efter 0,5 timers eksponering, 2,1 dage efter 1 times eksponering og 0,76 dage efter 4 timers eksponering (Tabel 13). Således falder LT50rent vand med stigende eksponeringsvarighed. Endelig kan det ses at den samme dødelighed (ca. 100%) opnås tidsforskudt som følge af både 1, 2 og 4 timers eksponering til esfenvalerat. Azoxystrobin (0-6,25 mg l-1) medførte akut mortalitet i de testede koncentrationer allerede efter 2 timers eksponering, hvorfor der kun er begrænsede data for sub-letale eksponeringer. Ved 0,5-1 times eksponering for 2,5 mg l-1 sås en forsinket mortalitet i rent vand med en LT50rent vand på 2,8-5,4 dage (Tabel 14). Ved 0,5-1 times eksponering for 6,25 mg l-1 sås en forsinket mortalitet i rent vand med en LT50rent vand på 1,4-1,9 dage. Prochloraz medførte ikke akutte effekter inden for de første 6 timers eksponering (0-10 mg l-1), men i den efterfølgende periode i rent vand indtraf en stigende dødelighed hos dyr eksponeret for hhv. 2, 5 og 10 mg l-1. Figur 22 viser hvorledes mortaliteten øges hos individer der er overført til rent vand efter 0,5-6 timers eksponering for 10 mg l-1 prochloraz.
Figur 22. Forsinket mortalitet hos Gammarus pulex som følge af eksponering til 10 mg/l prochloraz i hhv. 0,5, 1, 2, 4 og 6 timer For Gammarus pulex eksponeret til 10 mg l-1 prochloraz blev LT50rent vand beregnet til 5,7 dage efter 0,5 timers eksponering, 4,3 dage efter 1 times eksponering, 3,8 dage efter 4 timers eksponering og 2,8 dage efter 6 timers eksponering (Tabel 14). Tabel 14. Median letale tid (LT50rent vand), hvor 50% af individerne dør efter en kortvarig eksponering.
For de mest toksiske pesticider ses således en tendens til at selv ganske kortvarige, ikke letale eksponeringer helt ned til 0,5 timer udløser et forsinket toksisk respons i form af en forøget dødelighed efter eksponeringens ophør. Dette kan skyldes en tidsforskydning mellem eksponering og effekt ved korte pulseksponeringer, da pesticidet ikke når dets ”site of action” under eksponeringen. I sådanne tilfælde kan tidsrummet mellem eksponering og effekt forventes at afhænge af pesticidernes virkemekanismer samt optagelses- og fordelingskinetik, idet transporthastigheden til de forskellige ”sites of action” vil variere. Eksemplerne ovenfor illustrerer således at korte pulseksponeringer, der ikke medfører synlige akutte effekter, kan underestimere de toksiske effekter. Det ses også at eksponeringstiden influerer på hvor hurtigt det efterfølgende toksiske respons indtræffer, idet LT50rent vand bliver fremskyndet med stigende eksponeringsvarighed. Den forsinkede mortalitet, der blev observeret i forsøg med esfenvalerat, azoxystrobin og prochloraz er betinget af relativt høje eksponerings-koncentrationer. I vandløb vil koncentrationerne typisk være langt lavere, og der kan ikke forventes forsinket mortalitet efter kortvarig eksponering. Forsinkede effekter kan dog også komme til udtryk i form af sub-letale effekter som f.eks. hæmmet udvikling, vækst eller reproduktion, som på længere sigt kan påvirke populationerne (se 4.5). De udførte forsøg illustrerer dog vigtigheden af at vælge eksponeringsscenarier der tager højde for eksponeringsvarighed samt semi-kroniske effekter, der indtræffer med forsinket virkning. Resultaterne indikerer således, at de almindeligt anvendte parametre til bestemmelse af akut toksicitet (48-96 timers LC50) i flere tilfælde ikke kan anvendes til at forudsige korttidstoksiciteten af pesticider i vandløb, da man ved ekstrapolering til effektkoncentrationer ved korte eksponeringstider ikke tager højde for eventuelle forsinkede effekter. For at kortlægge effekterne af pulseksponeringer for pesticider i vandløb er det således relevant at inddrage en ”post-eksponerings fase” der også omfatter eventuelle forsinkede effekter. 4.4 Bestemmelse af interne effekt koncentrationerDet normalt anvendte endpoint i toksicitets test, EC/LC50, angiver den testkoncentration i det omgivende miljø der medfører en effekt på 50% af testpopulationen. EC/LC50 kan derfor betragtes som den eksterne effekt koncentration. Denne varierer som følge af almindelig biologisk variation testorganismer imellem, men også som følge af betingelserne i det omgivende miljø. F.eks. vil biotilgængeligheden af lipofile stoffer være påvirket af mediets sammensætning, indhold af partikler og organisk materiale samt typen af testbeholdere, da disse parametre influerer på stoffets sorption. Som supplement til eksterne effekt koncentrationer kan man undersøge kemiske stoffers interne effektkoncentration ved at relatere en given effekt (f.eks mortalitet) til den molære koncentration af stoffet i organismen (mmol/kg vådvægt). Den interne effektkoncentration for et kemisk stof er teoretisk set konstant for forskellige organismer, idet det antages at en toksisk effekt udløses når vævskoncentrationen når en effekt-specifik tærskelværdi. Det er dertil antaget at stoffer med forskellige virkemekanismer vil resultere i forskellige interne effektdoser. 4.4.1 MetodeI denne undersøgelse blev den interne letale koncentration (lethal body burden, LBB) bestemt i forsøg med de radioaktivt mærkede pesticider esfenvalerat, pendimethalin samt ioxynil eksponeret til Gammarus. LBB blev kvantificeret ved at bestemme den akkumulerede radioaktivitet i nyligt døde individer indsamlet løbende fra forsøgene med varierende eksponeringsvarighed. Således blev der beregnet LBB værdier til forskellige eksponeringstider og eksponeringskoncentrationer. 4.4.2 ResultaterDe beregnede LBB værdier syntes at være uafhængige af eksponeringstiden mens der var en tendens til LBB steg med eksponeringskoncentrationen (data ikke vist). Spektret af de beregnede LBB værdier er vist i Figur 23 (bemærk logaritmisk skala).
Figur 23. Lethal body burdens for esfenvalerat, pendimethalin og ioxynil. Gennemsnittet af LBB værdierne for de tre pesticider er vist i Tabel 15. Ligeledes er LC50 efter endt eksponering (14 dage) angivet for at illustrere den relative toksicitet af pesticiderne. Tabel 15. Lethal body burdens for esfenvalerat, pendimethalin og ioxynil
Trods spredningen af LBB værdierne for de enkelte pesticider fremgår det at der er en simpel sammenhæng mellem LBB og pesticidernes akutte toksicitet, idet LBB falder med stigende toksicitet. Således er den interne kritiske koncentration lavest for esfenvalerat som er det mest toksiske af de tre pesticider, mens den indre kritiske koncentration er væsentligt højere for ioxynil, som har en relativt lav akut toksicitet. Med andre ord er den akutte toksicitet negativt korreleret med den interne kritiske koncentration: jo højere toksicitet, jo lavere er den indre kritiske koncentration der udløser det toksiske respons. Dette kan understøtte teorien om, at forskellige virkemekanismer er associeret med forskellige spektre af body burdens. Det var i denne undersøgelse ikke muligt at skaffe flere radioaktivt mærkede pesticider, hvorfor LBB ikke kunne beregnes for et bredt udsnit af pesticider med forskellige virkemekanismer. Både esfenvalerat, pendimethalin og ioxynil er relativt lipofile stoffer hvorfor en akkumulering i organismerne over tid kan forventes. Det havde været relevant at undersøge om forsinkede effekter generelt var betinget af et højt akkumuleringspotentiale, idet den akkumulerede body burden forventes at være højere for lipofile stoffer sammenlignet med vandopløselige stoffer. Dette var dog ikke muligt at gennemføre i nærværende projekt. 4.5 Livscykluseffekter hos Gammarus pulexDet var det primære mål med denne delundersøgelse at kvantificere den relative følsomhed hos forskellige livsstadier hos krebsdyret Gammarus pulex. Effekter på livscyklus hos insektet Chironomus riparius er omtalt i afsnit 4.6. Sammenholdt giver disse forsøg mulighed for at vurdere forskelle i følsomhed mellem arter, forskellige stadier og effektparametre. I projektets sene fase blev der forsøgt at udføre livscyklusundersøgelser med dansemyggen Prodiamesa olivacea, der i modsætning til C. riparius lever udbredt i vandløb og med betydeligt højere krav til iltindhold og formodes at være mere følsom end C. riparius. Der var dog problemer med kultiveringen af P. olivacea og forsøgene kunne ikke gennemføres. I Bilag F er beskrevet forsøgene på kultivering. 4.5.1 Indsamling og forsøgsoversigtIndivider af G. pulex blev indsamlet fra Værebro Å, en sedimentrig og langsomtløbende å nær Roskilde Amts prøvestation nr. 1213. Indsamlingslokaliteten, der på den ene side var omgivet af brakmarker og på den anden side af buske og træer, klassificeres af Roskilde Amt (1998) som tilhørende forureningsgrad II/III eller faunaklasse 4. Indsamlingen af organismer foregik ved brug af ketcher og sparkeprøver. I laboratoriet blev organismerne forsigtigt sorteret i vandfyldte plastkar og kun ubeskadigede og ikke-parasitinficerede (Echinorrhyncus sp.) individer blev udvalgt til senere eksperimentelt brug. Opbevaring og kultivering af dyrene fulgte generelt McCahon & Pascoe's (1988a; 1988b) anvisninger herfor. Minimum 14 dage før start af et eksperiment blev dyrene opbevaret i 10 liters plastakvarier med en tæthed på ca. 200-300 dyr pr. akvarium. Dyrene blev holdt i luftgennemboblet vand, der bestod af en blanding (50:50) af vand indsamlet på feltlokaliteten og kunstigt ferskvand. Det kunstige ferskvand blev fremstillet efter OECD Guideline 202. Både under kultivering og eksperimenter blev dyrene holdt i klimarum ved 13°C og kunstige lysforhold (12 timer lys:12 timer mørke). Foderet bestod af delvist opløste elleblade (Alnus sp.) indsamlet på feltlokaliteten. Bladene, der alle var på samme forrådnelsesniveau, blev konditioneret i vand indsamlet på feltlokaliteten i en periode på minimum 14 dage inden brug. Afhængig af eksperimentet varierede testkamrene i størrelse og volumen, men i de fleste tilfælde blev der anvendt plastbeholdere med et volumen på 200 ml vand. For at minimere turbulens i testkamrene og stress af testorganismerne, blev der i visse eksperimenter tilført luft gennem kanylespidser som beskrevet af Welton & Clark (1980). I enkelte eksperimenter bestod testkamrene ligeledes af en sedimentfase. I tilfælde af en sedimentfase blev der anvendt vasket akvariegrus med en partikelstørrelse på 2-3 mm. Der blev udført 6 forskellige forsøg med Gammarus pulex varierende fra ”worst-case” eksponering af voksne individer til detaljerede forsøg med esfenvalerats effekt på parringsadfærd og effekter på afkom. Detaljer om dosering and analyse af esfenvalerat samt anvendt statestik fremgår af Bilag C. 4.5.2 Worst-case eksponering af G. pulexDette forsøg blev udført med henblik på bestemmelse af effekten af et ”worst-case scenarium”, hvor G. pulex blev eksponeret til en 1-times puls af esfenvalerat ved koncentrationerne: 0 og 4,65 µl-1, hvor den høje koncentration svarer til den højeste mulige eksponering ved direkte oversprøjtning på en 2 meter dybt vandområde, beregnet udfra anvendelse af EU's maksimale dosis på 93 g a.i./ha. Herudover blev der desuden anvendt en acetonekontrol. I både kontrolgruppen og acetonekontrolgruppen blev der anvendt 10 voksne (kropslængde = 10-14 mm) individer, mens der i eksponeringsgruppen blev anvendt 20 voksne individer. Efter pulseksponering, der foregik i bægerglas, blev dyrene skyllet i rent vand og overført til testkamre indeholdende rent vand. Overlevelse og immobilitet blev herefter observeret over en periode på 144 timer. Undervejs blev dyrene fodret med bladskiver (diameter = 18mm). Figur 24 illustrerer effekter på overlevelsen af voksne G. pulex efter, at de har været udsat for et “worst-case pulseksponerings scenarium”. Allerede 1 time efter pulseksponeringen er kun 60% af de eksponerede individer i live, mens der blev observeret 100 % dødelighed 144 timer efter eksponeringen. Til sammenligning er der henholdsvis 10% og 90% overlevelse i kontrol- og actonegruppen.
Figur 24. Overlevelse efter worst-case (4.56µg l-1) pulseksponering af G. pulex. 4.5.3 Effekt på juvenile og ikke-reproducerende voksneDette forsøg undersøgte overlevelsen af både juvenile (kropslængde = 3-7 mm) og ikke-reproducerende voksne (kropslængde = 9-15 mm) eksponeret til en 1-times puls af esfenvalerat. Foruden en kontrol og en acetone-kontrolgruppe blev der anvendt testkoncentrationer på 0,1 og 0,3 µl-1. I forsøget blev testkamrene foruden vand tilført sediment (1 del sediment: 5 dele vand, på volumen basis). I halvdelen af eksponeringsgrupperne blev testorganismerne efter pulseksponeringen overført til testkamre med rent vand og rent sediment, mens den anden halvdel af testorganismerne blev overført til testkamre hvor kun vandet blev skiftet. Formålet med dette forsøgsdesign var at bestemme i hvor høj grad esfenvalerat bandt sig til sedimentpartiklerne under pulseksponeringen, og om det således efterfølgende kunne virke som en kontinuerlig eksponeringskilde. Der blev anvendt 10 replikater af hver 10 individer pr koncentration for voksne og 10 replikater af 5 individer pr koncentration for juvenile. Umiddelbart efter eksponering blev dyrene overført til deres respektive testkammer, hvor overlevelsen blev observeret hver anden dag over en periode på to uger. Alle voksne individer overlevede den 1 times pulseksponering med esfenvalerat koncentrationer på op til 0,3 µg l-1. Overlevelsen var faldende i perioden efter eksponeringen, men variationer i median overlevelsestid testgrupperne imellem blev først detekteret syv dage efter pulsen (Kruskal Wallis p-værdier var som følger: Dag 1: 0,746; Dag 2: 0,746; Dag 3: 0,141; Dag 4: 0,179; Dag 7: 0,033; Dag 9: 0,005; Dag 11: 0,046; Dag 14: 0,038). Gennemsnitlig og median overlevelsestid for hver gruppe, estimeret udfra overlevelseskurverne, fremgår af Tabel 16. Table 16. Gennemsnitlig og median overlevelsestid (dage) hos juvenile og voksne efter 1-times pulseksponering til esfenvalerat. Værdierne i den sidste kolonne er fra sammenligninger i overlevelseskurver (Tarone-Ware Log-rank test) mellem juvenile og voksne indenfor hver eksponeringsgruppe.
Betragtes overlevelseskurverne ses en tendens til at overlevelsen i de 14 dage efter pulseksponeringen faldt hurtigere i de eksponerede grupper end i kontrollerne og at overlevelsen i grupperne eksponeret til 0,3 µg l-1 faldt hurtigere end grupperne eksponeret til 0,1 µg l-1 (Figur 25). Selvom figuren indikerer en lille forskel i overlevelse mellem grupperne der modtog den samme pulskoncentration - men som blev overført til testkamre henholdsvis med eller uden kontamineret sediment - var forskellen mellem kurverne ikke signifikant (Tarone-Ware; 0,3 µg l-1: p=0.808; 0,1 µg l-1: p=0,601). Overlevelsen var generelt høj i alle grupper og oversteg 80 % i alle grupper (Figur 25). Alle juvenile individer overlevede en times pulseksponering til esfenvalerat koncentrationer på op til 0,3 µg l-1. Overlevelsen faldt over tid i perioden efter pulsen, og i modsætning til de voksne individer kunne der detekteres variationer i median overlevelsestid grupperne imellem allerede et døgn efter pulsen (Kruskal Wallis p-værdi 0,001 for alle dage). Overlevelseskurverne indikerer at overlevelsen faldt hurtigere efter pulsen i de eksponerede grupper end i kontrol grupperne og at overlevelsen i grupperne eksponeret til 0,3 µg l-1 faldt hurtigere end i grupperne eksponeret til 0,1 µg l-1 (Figur 26). Selvom figuren indikerer en lille forskel i overlevelse mellem grupperne der modtog den samme pulskoncentration - men som blev transporteret til testkamre henholdsvis med eller uden kontamineret sediment - var forskellen mellem kurverne ikke signifikant (Tarone-Ware; 0,3 µg l-1: p=0,638; 0,1 µg l-1: p=0,184). Gennemsnitlig og median overlevelsestid for hver gruppe, estimeret udfra overlevelseskurverne, kan ses i Tabel 16. Ved slutningen af forsøget var overlevelsen i kontrollen og acetonekontrollen på henholdsvis 74 og 76 %, mens overlevelsen i grupperne eksponeret til 0,1 µg l-1 var 24% (rent sediment) og 14 % (kontamineret sediment). I grupperne eksponeret til 0,3 µg l-1 var overlevelsen på henholdsvis 0 % (kontamineret sediment) og 2% (rent sediment). Overlevelsen faldt hurtigere i alle eksponeringsgrupper hos de juvenile end hos de voksne individer, og trods små forskelle var der også signifikante forskelle i overlevelsen mellem juveline og voksne i kontrol- og acetonekontrolgrupperne (Tabel 16).
Figur 25. Overlevelse af voksne individer af G. pulex udsat for en 1-times puls af esfenvalerat.
Figur 26. Overlevelse af juvenile individer af G. pulex udsat for en 1-times puls af esfenvalerat. 4.5.4 Effekt på reproducerende par og deres afkomI dette forsøg undersøgtes effekten af en kort puls af esfenvalerat på reproduktionsadfærden hos voksne G. pulex. Fjorten sammenhængende par af G. pulex, alle med en ægbærende hun, blev eksponeret til hver en 1-times puls af følgende testkoncentrationer: 0, 0,05, 0,1 og 0,3 µl-1. Eksponeringsproceduren var den samme som beskrevet under tidligere eksperimenter. Efter eksponering blev hvert par placeret individuelt i testkamre og overlevelsen af de voksne og deres afkom observeret over en periode på to uger. Juvenile der blev frigivet fra de gravide hunner under eksponeringen og den efterfølgende observationsperiode blev talt og opbevaret separat. Afkommet blev isoleret for at undgå prædation fra de voksne under forsøget. Under den 1-times pulseksponering frigav hunnerne et stort antal befrugtet afkom (Tabel 17). Tabel 17. Oversigt over antal unger frigivet under eksponering, procent unger observeret ved forsøgets slutning, antal overlevende voksne (hunner + hanner), samt antal gendannede par af 14 mulige.
Den procentvise overlevelse hos de unger der blev frigivet under pulseksponeringen var faldende med stigende koncentration (Figur 27). Overlevelsen af afkommet fortsatte med at falde over tid under den to uger lange observationsperiode i rent vand og var afhængig af pulskoncentrationen. Parrede voksne var relativt sensitive overfor esfenvalerat (sammenlignet med tidligere studier af ikke-parrede voksne) og overlevelsen ved den højeste koncentration (0,3 µg l-1) var meget lav for både hanner og hunner (Figur 28). Hertil kommer, at overlevelsen hos hunnerne var faldende over tid og ca. halvt så stor som hos hannerne ved slutningen af den to uger lange observationsperiode, og dette forhold gjorde sig ligeledes gældende i kontrollen (Tabel 17).
Figur 27. Overlevelse af unger frigivet efter en 1-times pulseksponering af forældrene ved følgende koncentrationsniveauer af esfenvalerat: 0, 0,05, 0,1 og 0,3 µg l-1. Ingen par i de eksponerede grupper blev gendannet under den to ugers observationsperiode sammenlignet med kontrollen, hvor 11 ud af 14 par blev gendannet (Tabel 16). Ligeledes var overlevelsen blandt de juvenile, der blev produceret under de to uger efter eksponeringen betydeligt lavere i forsøgsgrupperne end i kontrollen (Figur 29).
Figur 28. Overlevelse af reproducerende voksne G. pulex efter en 1-times pulseksponering ved følgende koncentrationsniveauer af esfenvalerat: 0, 0,05, 0,1 og 0,3 µg l-1.
Figur 29. Antal overlevende unger produceret af voksne individer af G. pulex over en periode på to uger efter en 1-times pulseksponering til esfenvalerat ved følgende koncentrationsniveauer: 0, 0,05, 0,1 og 0,3 µg l-1. 4.5.5 Effekt på nyfødteDa det forgående forsøgt afslørede en relativ høj dødelighed blandt de unger, der var blevet frigivet af den tidligere eksponerede moder, blev dette forsøg udført for at undersøge nyfødte individers sensitivitet overfor en kortvarig puls af esfenvalerat. Halvtreds nyligt frigivede juvenile (kropslængde = 1-2 mm) pr. testgruppe blev eksponeret til en 1-times puls af hver af følgende koncentrationer: 0, 0,05, 0,1 og 0,3 µl-1 og overlevelsen observeret over en periode på en uge. Der blev ikke anvendt replikater, idet alle individer fra hver koncentration blev holdt i et enkelt testkammer. Under observationsperioden blev de juvenile fodret med fækalier produceret af voksne fra kulturakvarierne. Overlevelsen blandt nyfødte eksponeret for en 1-times puls af esfenvalerat faldt med stigende koncentration i ugen efter eksponering (Tarone-Ware Log-rank Test, 2=138,2, P<0,00001; Figur 30). Den gennemsnitlige overlevelsestid var: 6,8 dage (kontrol), 5,1 dage (0,05g l-1 gruppe), 3,5 dage (0,1 µg l-1 gruppe) og 2,3 dage (0,3 µg l-1 gruppe).
Figur 30. Overlevelse af nyligt producerede juvenile G pulex efter en 1-times pulseksponering til esfenvalerat ved følgende koncentrationsniveauer: 0, 0,05, 0,1 og 0,3 µg l-1. 4.5.6 Effekt på parringsadfærd og efterfølgende reproduktionI modsætning til tidligere beskrevne eksperimenter relateret til G. pulex's reproduktive adfærd, blev dette forsøg udført med par af G. pulex hvor hunnerne endnu ikke var blevet befrugtet og derfor endnu ikke bar juvenile på maven. Ti prækopulative par per testgruppe blev forsigtigt adskilt efter metoden beskrevet af Malbouisson (1995), idet det sammenhængende par blev lagt på et filterpapir indtil hannen gav slip på hunnen. Hanner og hunner blev herefter individuelt eksponeret til en 1-times puls af hver af følgende koncentrationer: 0, 0,05, 0,1, 0,3 og 0,6 µl-1. Kropslængden hos hannerne varierede mellem 10-15 mm mens kropslængden hos hunnerne varierede mellem 9-12 mm. Efter eksponering blev dyrene skyllet i rent vand og placeret i testkamre med den samme partner som før adskillelsen. Overlevelse, tid til evt. gendannelse af par, samt den procentvise gendannelse af par blev herefter observeret over en periode på to uger. Antallet af juvenile produceret per reproducerende hun blev ligeledes noteret. Juvenile og døde individer blev straks fjernet fra testkamrene. Par hvor en af parterne døde i løbet af observationsperioden blev ikke anvendt i den endelige dataanalyse. Under den 1-times eksponeringspuls blev de fleste af individerne i de tre højeste koncentrationsgrupper immobile og forblev liggende på bunden af eksponeringskamrene. Denne effekt fortsatte et par dage efter eksponeringen. Til trods for den relativt store størrelsesforskel kønnene imellem, blev der ikke observeret nogen statistisk signifikans i overlevelsen mellem hanner og hunner (Tarone-Ware Log-rank test χ2≤0,07 og P≥0,78 for alle koncentrationer). Tretten dage efter pulseksponeringen var overlevelsen på 95 og 100% i henholdsvis kontrollen og acetonekontrollen, hvorimod overlevelsen kun var på 60% i den højeste eksponeringsgruppe (0,6 µg l-1). For begge køn viste 0,1 µg l-1 gruppen en lavere overlevelse end 0,3 µg l-1gruppen. Den gennemsnitlige overlevelsestid var: 17,4 dage (kontrol), 13,0 dage (acetone), 11,8 dage (0,05 µg l-1), 12,7 dage (0,1 µg l-1), 12,2 dage (0,3 µg l-1), og 10,9 dage (0,6 µg l-1) (Figur 31).
Figur 31. Overlevelse af reproducerende voksne G. pulex forstyrret fra præcopula ved separation og pulseksponering (1 time) til esfenvalerat ved følgende koncentrationer: 0, 0,05, 0,1, 0,3 og 0,6 g l-1. Der blev desuden anvendt en acetonekontrol. Øverst: hunner; Nederst: hanner. Efter to dage var 60% af de overlevende par i både kontrollen og acetonekontrollen gendannet. Til sammenligning tog det ca. seks gange så lang tid (12 dage) før de forstyrrede par, der havde været eksponeret til esfenvalerat, blev gendannet. Eksponeringskoncentrationen havde en signifikant effekt på den tid det tog for parrene at blive gendannet (Tarone-Ware Log-rank test χ2=16,29 og P=0,006). Det bør dog noteres, at tiden til gendannelse af par i de højeste eksponeringsgrupper er baseret på færre individer, idet overlevelsen var faldende med stigende koncentrationer (Tabel 18). Tabel 18. Varighed for genetablering af præcopula, procent hunner som reproducerede og gennemsnitligt antal afkom er hun efter pulseksponering til esfenvalerat.
Eksponering til en kortvarig puls af esfenvalerat havde en signifikant effekt på antallet af juvenile, der blev produceret i perioden efter pulsen (Figur 32). Hos alle hunnerne havde esfenvalerat en signifikant effekt på antallet af juvenile produceret efter eksponering (Kruskal-Wallis Test, P=0,002). Kontrolgruppen producerede signifikant flere juvenile end 0,1 µg l-1 gruppen og 0,6 µg l-1 gruppen, men ikke 0,3 µg l-1 gruppen. Betragter man udelukkende de ynglende hunner vil esfenvalerats effekt på antallet af producerede juvenile derimod være marginal signifikant (Kruskal-Wallis Test, P=0,09). Dette indikerer at effekten på reproduktionen formentlig kan tilskrives det lavere antal reproducerende hunner.
Figur 32. Gennemsnitlige antal unger produceret af gendannede par i de forskellige eksponeringsgrupper. Det største antal juvenile (total = 133 individer produceret af 8 hunner) blev fundet i kontrolgruppen, hvilket kan forklares med en høj overlevelse samt hurtig genparring blandt de voksne. Til sammenligning blev der fundet færrest juvenile i 0,6 µg l-1 gruppen (total = 17 individer produceret af 2 hunner). Antallet af juvenile produceret pr. ynglende hun varierede fra 4 (0,1 µg l-1 gruppen) til 25 (acetone gruppen). Det bør noteres, at antallet af juvenile fundet i hver gruppe kan være en smule underestimeret grundet prædation fra de voksne. 4.5.7 Effekt af ugentlige eksponeringer til korte pulserDette forsøg blev udført for at undersøge effekten af gentagende pulser af 1 times varighed ved koncentrationerne: 0 og 0,1 µl-1 på parringsadfærden hos G. pulex. Der blev anvendt tre replikater af fem par voksne (kropslængde = 10-14 mm) individer af G. pulex per testgruppe. Testgruppe A blev udsat for tre pulseksponeringer (1 hver uge i tre uger), testgruppe B blev udsat for 2 pulseksponeringer (1 hver uge i de to første uger) mens testgruppe C blev udsat for en enkelt pulseksponering (1 i den første uge). Kontrolgruppen fik maksimal stressbehandling som testgruppe A, dvs. tre eksponeringer, blot med den forskel at eksponeringen foregik i rent vand. Eksponeringsproceduren var som tidligere beskrevet, idet parrene blev eksponeret i glasbeholdere og efterfølgende skyllet og overført til testkamre med rent vand. Dyrene var aldrig fødebegrænsede, idet de blev foderet regelmæssigt med bladskiver. Under den tre ugers forsøgsperiode blev overlevelse og gendannelse af parrene noteret. Den højeste overlevelse efter tre uger blev observeret i kontrollen (77 %), mens den laveste overlevelse blev observeret i testgruppe A (60 %), der havde været udsat for tre pulseksponeringer (Figur 33). I testgruppe B, der havde været udsat for to pulser, var overlevelsen 70 %, mens testgruppe C, der havde været udsat for en enkelt puls, sluttede med en overlevelse på 73 %. Gendannelsen af par var generelt lav i alle testgrupper efter første eksponering, hvilket antyder, at de fleste par allerede havde reproduceret og derfor ikke behøvede at gendannes.
Figur 33. Overlevelse af G. pulex par udsat for en ugentlig 1-times pulseksponering af esfenvalerat (0,1µg l-1). Pilene angiver eksponeringstidspunkterne. A blev udsat for tre pulseksponeringer (1 hver uge i tre uger), B blev udsat for 2 pulseksponeringer (1 hver uge i de to første uger) mens C blev udsat for en enkelt pulseksponering (1 i den første uge). 4.6 Livscykluseffekter hos Chironomus riparius400 larver fra henholdsvis en finsk og en engelsk kultur af Chironomus riparius blev anvendt til start af egen kultur. Kultiveringsteknikken fulgte generelt OECD Guidelines for the Testing of Chemicals (Proposal for a New Guideline 219). Således blev larverne holdt i kulturbure med følgende dimensioner: højde = 40 cm, længde = 34 cm, bredde = 20 cm, hvor bunden blev forsynet med et ca. 3 cm tykt lag kunstigt sediment fremstillet efter OECD's anvisning herfor. Vandet bestod af afboblet hanevand i en dybde på ca. 6 cm og der blev tilført luft og omrøring i vandsøjlen ved hjælp af en svag gennemboblinge. Toppen af kulturburene bestod af en aftagelig sværmekasse fremstillet af myggenet med en maskestørrelse på 1mm. Dyrene blev holdt ved 20C og kunstige lysforhold (12 timers lys: 12 timers mørke) med en lysintensitet på ca. 1000 lux. Foderet bestod almindeligt fiskefoder (TetraMin) knust og opløst i vand ved forholdet: 1 gram fiskefoder/20 ml vand. I henhold til OECD's guideline anbefales små larver en fodermængde på ca. 0,25-0,5mg/dag mens store larver kræver ca. 0,5-1 mg/dag for at opnå optimale vækstbetingelser. Producerede ægsække blev høstet dagligt og den eksakte læggedato noteret, idet de herefter blev overført til petriskåle. Anvendt statistik er anført i Bilag C. 4.6.1 Eksponering til en 1-times puls (uden sediment)Forsøget havde til formål at undersøge effekten af en kortvarig puls af esfenvalerat ved et koncentrationsniveau på 0,2µg l-1. Seks ægsække med samme alder ( 1 døgn) fra egen laboratoriekultur blev isoleret i en petriskål. En uge senere blev det ønskede antal larver udtaget og eksponeret i glasbeholdere med et vandvolumen på 0,5 liter. Der blev anvendt 4 replikater af 30 dyr pr. testkammer fordelt på tre forsøgsgruppe. Forsøgsgrupperne var omfattede: en kontrolgruppe; en testgruppe eksponeret til en 1-times puls på 0,2µg l-1 og overført til testkamre med rent sediment; samt en testgruppe eksponeret til en 1-times puls på 0,2µg l-1 og overført til testkamre med sediment der ligeledes havde været kontamineret til en 1-times puls på 0,2µg l-1. Efter endt eksponering blev dyrene med automatpipette overført til et større kar med en stort vandvolumen, for at undgå overførsel af pesticidrester. Herfra blev dyrene atter ved hjælp af automatpipette overført til de respektive forsøgskamre. Testkamrene målte 7 cm i diameter og 4 cm i højden og blev tilført 20 ml kunstigt sediment og 100 ml gennemboblet hanevand. 30 individer i hvert forsøgskammer gav en tæthed på: 1 individ/ 1,3 cm². Hvert testkammer blev forsynet med et aftageligt låg, hvis top bestod af myggenet, til opsamling af klækkede individer. Der blev fodret svarende til et niveau på 0,5 mg/larve/dag. Efterfølgende blev forsøgskamrene observeret i en periode på 39 dage med henblik på bestemmelse af følgende effektparametre:
Antal og køn af klækkede individer i hvert testkammer blev dagligt noteret og de færdigudviklede individer blev straks overført til sværmebure med følgende dimensioner: højde = 20 cm, længde = 34 cm, bredde = 20 cm. Sværmeburene indeholdt ligeledes en vandfyldt petriskål til opsamling af ægsække. Alle færdigudviklede individer tilhørende den samme forsøgsgruppe blev, uafhængigt af replikat, overført til det samme sværmebur. Dette var for at sikre et tilstrækkeligt stort antal individer pr. sværmebur til at reproduktion kunne foregå. Manglen på replikater som følge af denne forsøgsopstilling betød, at det ikke var muligt at udføre en statistisk analyse på de opnåede reproduktionsdata. Ved forsøgets afslutning blev sedimentet i samtlige testkamre sigtet for at bestemme antallet af evt. uklækkede larver og pupper. Figur 34 viser den procentvise kumulative andel af alle klækkede individer i forsøgsperioden, idet hver kurve repræsenter både hanner og hunner. Den højeste andel af klækkede individer over tid blev observeret i kontrolgruppen (ca. 50%), mens den laveste andel blev fundet i gruppen med kontamineret sediment (ca. 12%). Eksponering til esfenvalerat reducerede tiden til klækning i både gruppen med rent og kontamineret sediment (Tabel 19). Generelt var hunnerne længere tid om at klække end hannerne. Selvom der ikke var signifikans i interaktion mellem esfenvalerat og køn, indikerede separate parvise sammenligninger af hver gruppe, at det kun var i kontrolgruppen at klækningstiden hos hunner var signifikant længere end hanner (ANOVA; kontrol: P=0,001; rent sediment: P=0,88; esfenvalerat sediment: P=0,22). Median klækningstid er vist i Tabel 20. Klækningstiden blev reduceret for både hanner og hunner ved pulseksponering til esfenvalerat, men effekten var større på hunner end på hanner. Eksponering til esfenvalerat havde også en markant effekt på det totale antal af voksne der klækkede (Tabel 20; Pearson Chi-square = 49,44, df=2, P << 0,001). Dette forhold var gældende for både gruppen holdt i rent og kontamineret sediment efter pulseksponeringen.
Figur 34. Procentvise kumulative andel af alle klækkede individer over en forsøgsperiode på 39 dage. Tabel 19. Effekt af esfenvalerat og køn på tid til klækning hos Chironomus riparius. Interaktionen mellem esfenvalerat og køn var ikke signifikant (P=0,50) og en reduceret model, uden interaktion, blev anvendt.
Det totale antal æg produceret af kontrolgruppen var 1180, 164 æg blev produceret af gruppen holdt i rent sediment efter eksponeringen, mens der blev produceret 472 æg af gruppen holdt i kontamineret sediment efter eksponeringen. Af disse klækkede 835 succesfuldt i kontrolgruppen (70,1%), 122 i gruppen med rent sediment (74,4%) og 319 i gruppen med kontamineret sediment (67,6%). Tabel 20. Median tid til klækning og total antal klækkede voksne individer af Chironomus riparius.
4.6.2 Eksponering til en 1-times puls (med sediment)Da den mest realistiske eksponeringsvej for den sedimentlevende organisme C. riparius formodes at foregå gennem sedimentet blev testorganismerne i dette forsøg pulseksponeret mens de befandt sig i sedimentet, modsat i ovenstående forsøg, hvor testorganismerne blev pulseksponeret udenfor sedimentet. Foruden bestemmelse af effekten af en kortvarig puls til esfenvalerat ved et koncentrationsniveau på 0,2 µg l-1 havde dette forsøg til formål at undersøge betydningen af populationstætheden i testkamrene. Fra egen laboratoriekultur blev udtaget 6 ægsække med samme alder (1 døgn). Disse blev isoleret i en petriskål og en uge senere blev 600 larver udtaget og fordelt i følgende forsøgsgrupper af hver 10 replikater: en kontrolgruppe med en tæthed på 1 larve pr. 5 cm², en testgruppe med en tæthed på 1 larve pr. 5 cm² eksponeret til en 1-times puls esfenvalerat (0,2 µg l-1); en kontrolgruppe med en tæthed på 1 larve pr. 2,5 cm², en testgruppe med en tæthed på 1 larve pr. 2,5 cm² eksponeret til en 1-times puls esfenvalerat (0,2 µg l-1). Pulseksponeringen af testorganismerne foregik først et døgn efter, at larverne var blevet introduceret til de respektive testkamre indeholdende kunstigt sediment og gennemboblet hanevand i volumenforholdet (1:5). Herved havde larverne fået lejlighed til at grave sig ned i sedimentet inden selve eksponeringen. Testkamrene målte 8 cm i diameter og 4 cm i højden. Ligesom i det foregående forsøg blev hvert testkammer forsynet med et aftageligt låg til opsamling af klækkede individer. Der blev foderet svarende til et niveau på 0,25 mg/larve/dag. Herefter fulgte forsøget den samme procedure som beskrevet under forsøget med eksponering udenfor sedimentet, idet der blev testet for de samme endpoints, samt betydningen af individtætheden i testkamrene. Dette forsøg blev kørt over en periode på 44 dage. Figur 35 viser den procentvise kumulative andel af alle klækkede individer i forsøgsperioden, idet hver kurve repræsenter både hanner og hunner. Den højeste andel af klækkede C. riparius blev observeret i kontrolgruppen med lav individtæthed, hvor hele 93% af de tilførte testorganismer var klækket ved forsøgets afslutning efter 44 dage. Den laveste andel (57%) af klækkede C. riparius blev fundet i gruppen med høj individtæthed eksponeret til 0,2 µg l-1.
Figur 35. Procentvise kumulative andel af alle klækkede individer over en forsøgsperioden på 44 dage. Hos hanner havde esfenvalerat eksponering en reducerende effekt på tid til klækning, mens stigende individtæthed havde en øgende effekt på tid til klækning (Tabel 21). Hos hunner var der ingen effekt af esfenvalerateksponering på tid til klækning, mens stigende individtæthed havde en signifikant øgende effekt på tid til klækning. Både i kontrol og esfenvalerat eksponerede grupper, og ved både lave og høje individtætheder, blev der hos hunnerne observeret en signifikant længere tid til klækning end hos hannerne (p0,001 for alle tests). Table 21. Effekt af esfenvalerat og tæthed på tid til klækning hos Chironomus riparius. Hverken for hanner eller hunner var interaktionen mellem esfenvalerat og tætheden signifikant (Hanner; P=0,685, Hunner; P=0,241).
Tiden til 50% af individerne klækkede og det totale antal af klækkede individer i forsøgsperioden på 44 dage er vist i Tabel 22. Individtætheden havde en signifikant og negativ effekt på klækningssuccesen af larver i både kontrollen (Pearson Chi-square=24,02; P < 0,001) og gruppen eksponeret til esfenvalerat (Pearson Chi-square=24,10; P < 0,001). Derimod havde esfenvalerat ingen signifikant effekt på den procentvise andel af larver med klækningssucces i gruppen med lav individtæthed (Pearson Chi-square=1,45, P=0,23) eller i gruppen med høj individtæthed (Pearson Chi-square=1,22; P=0,27). I Tabel 23 er vist det samlede reproduktive output i de forskellige grupper over den 44 dage lange forsøgsperiode. Hverken eksponering til esfenvalerat eller individtæthed havde en signifikant effekt på ægproduktionen (Esfenvalerat: df=1,4; F=0,31; P=0,61; Tæthed: df=1,4; F=1,86; P=0,24; esfenvalerat x Tæthed: df=1,4; F=0,99; P=0,38). Ligeledes blev der heller ikke observeret nogen signifikant effekt på antallet af unger der klækkede (esfenvalerat: df=1,4; F=0,03; P=0,87; Tæthed: df=1,4; F=0,42; P=0,55; esfenvalerat x Tæthed: df=1,4; F=0,96; P=0,38). Den gennemsnitlige klækningssucces af æggene var for alle grupper 77% (±11,25=SD). Tabel 22. Median klækningstid og antal klækkede voksne individer af Chironomus riparius ved 1 larve pr. 5 cm² og 1 larve pr. 2.5 cm².
Tabel 23. Reproduktive output fra Chironomus riparius ved 1 larve pr. 5 cm² og 1 larve pr. 2.5 cm². Klækkede individer fra replikat 1-5 og replikat 6-10 blev puljet for at sikre tilstrækkeligt antal voksne individer til reproduktion.
4.7 Drift hos invertebrater efter pesticideksponeringEn sammenligning af følsomheden af forskellige effektparametre i test med pesticider i kunstige vandløb har vist, at forøget drift (korttidsrespons) af insektlarver og gammarider generelt er mere følsom end reduktion i abundans (langtidsresponse) (Møhlenberg et al. 2001). I vandløb er drift en naturlig adfærd hos bundlevende krebsdyr og insektlarver, der knytter sig til spredning, fødesøgning og forplantning. Ved ugunstige forhold f.eks. som følge af tørke og forekomst af toksiske stoffer så som pesticider kan der udløses katastrofedrift i invertebratbestandene. Denne drift kan dels være udtryk for en aktiv response for ugunstige forhold eller den kan skyldes en narkotisk virkning af et pesticid og en følgende passiv transport med strømmen. Under normale forhold kan 1-5 % af en population være i drift på et givet tidspunkt. Hvis bunddyrene udsættes for insekticider som lindan, permetrin oa. øges driftraten, i enkelte tilfælde med op til 10000 % (Møhlenberg et al. 2001). Generelt kan drift og ændring i abundans opfattes som reciprokke processer, idet øget drift fra et område vil føre til reduktion i abundans. I projektet er der undersøgt for pesticiders effekt på drift hos krebsdyr og insektlarver, dels under laboratorieforhold i små systemer (recirkulerende strømrender) dels under under ”semi-naturlige” forhold i kunstige vandløb uden recirkulering 4.7.1 Drift i recirkulerende strømrenderI eksperimenterne anvendtes 16 strømakvarier placeret i termorum (Fig. 36). Akvarierne er fremstillet i en blanding af transparent, lys og mørk grå, 5mm tyk akryl, bukket i form under opvarmning og samlet ved svejsning med akryltråd. Akvarierne er 1m lange, 20cm brede og adskilt i to parallelle render med en 80cm lang midtervæg. Vandstrømmen genereres af en cylinder i PVC med en diameter på 11cm. Ydervæggen er 7,5cm høj og under forsøgene er vanddybden 5cm ved 10 L vand, hvorved cylinderen stikker 2,5-3 cm ned i vandet. Strømhastigheden i systemerne bestemmes af omdrejningshastigeheden af cylinder, der styres af 2 frekvensregulerede motorer, som hver er koblet til 8 strømrender
Figur 36. To af de 16 strømakvarier. Under forsøg tilsættes kvartsgrus og konditionerede blade som substrat for invertebraterne i renden modsat den roterende cylinder I forsøgene anvendtes minimum 10 individer per art per strømakvarium. Et døgn inden et forsøg blev strømakvarierne fyldt med filtreret åvand, der placeredes 200 g kvartsgrus (5 mm) i renden modsat det roterende hjul over en strækning på 45 cm. Der placeredes yderlige 10 ”konditionerede” blade (elm, el, eg) fra en beholder med åvand. Bladene blev fastholdt med sten. Dyrene blev tilsat og vandstrømmen indstillet til 9-10 cm s-1 (fri hastighed ved overfladen). Efter 24 timer vil hovedparten af dyrene sidde i eller på blade/grus. Dyrenes placering (indenfor og udenfor grus) og aktivitet noteredes kontinuerligt indtil 40 min. Herefter tilsattes pesticidet (opløst i acetone for tungtopløselige pesticider) i koncentrationer fastlagt ved indledende forsøg og dyrenes placering og aktivitet noteres igen kontinuerligt indtil 90 min og derefter over en 15 min periode hver 24. time indtil 96 timer. Ved hver registrering (ca. 4-5 gange i minuttet) noteres dyrenes placering (på strækning udenfor sten). Aktiviteten udenfor afsnittet med sten inddeles i:
Intensiteten i optællingen varierede mellem 2-6 gange per 5 min periode afhængig af antal strømakvarier i et forsøg og især af dyrenes art og størrelse. For at undgå eventuelle bias blev observationerne midlet over 5 min intervaller. Hver forsøgsserie gennemførtes med minimum 2 kontrolrender og et varierende antal eksponeringsrender (6-14). I de tilfælde hvor det var nødvendigt at opløse pesticidet i acetone (esfenvalerat, deltamethrin, pendimethalin, azoxystrobin) blev der tilsat acetone i samme koncentration i alle strømrender. Temperaturen under forsøget blev afpasset til årstiden og in situ temperaturen (6-15 °C). Der anvendtes temperaturer indenfor intervallet 7 – 12 °C. Driftraten blev udtrykt i procent af aktive (og observerbare) individer:
hvor #D = antal levende individer i drift, #OP = antal individer under optrøms migration, #AB = antal individer der viste andre aktivitetsmønstre eller kunne observeres som levende. I stedet for at basere beregningerne på det totale antal tilsatte dyr i strømrenderne sikrede denne beregning, at døde og inaktive (evt. paralyserede) individer ikke indgik. Ulempen ved fremgangsmåden var dog en større relativ variation fordi ”nævneren” (summen af #D, #OP og #AB) var variabel og ofte væsentlig lavere end det tilsatte antal dyr (10). Bindingen af pesticider til overflader i strømakvarier kan mindske eksponeringen. Adsorptionen af esfenvalerat (som har den højeste Kd af de undersøgte pesticider) blev undersøgt ved hjælp af radioaktivt mærket esfenvalerat. Tabet blev bestemt til at være mellem 20-30% over en time, som var den normale forsøgsvarighed (Figur 37). Vi antog derfor, at de nominelle koncentrationer gav et dækkende udtryk for de reelle koncentrationer under forsøgene.
Figur 37. Tidslig variation i koncentrationen af radioaktivt esfenvalerat gennem 60 min i strømrende. Driftrespons blev undersøgt hos Gammarus sp., Asellus, Baetis, Leuctra, Hydropsyche, efter eksponering til pesticiderne pendimethalin, pirimicarb, azoxystrobin, deltamethrin samt esfenvalerate (Tabel 24). Sidstnævnte blev undersøgt som rent kemisk stof og i handelsproduktet Sumi-alpha, som indeholder detergenter og hjælpestoffer i ikke offentliggjort koncentration og art. Tabel 24. Oversigt over gennemførte forsøg med driftrespons (og toksicitet) i recirkulerende vandsløbsrender.
Undersøgelserne viste, at kun pyrethroiderne (esfenvalerat og deltamethrin) udløste drift hos krebsdyrene og insektlarverne (Fig. 38 - 40), mens ingen af herbiciderne, fungicidet azoxystrobin eller insekticidet pirimicarb udløste flugtrespons (Bilag D). For esfenvalerat var der forøget drift hos Gammarus ved 0,6 ng l-1 45-90 min efter tilsætning, mens følsomheden hos Asellus, Baetis og Leuctra var ringere (typisk 6 ng l-1). Undersøgelserne viste ingen drift hos Hydropsyche uanset koncentration eller art pesticid. Undersøgelser med anvendelse af esfenvalerat i det formulerede produkt Sumi-alfa viste, at driften hos alle arter udløstes tidligere og ved lavere koncentrationer end det rene stof (Fig. 40 & 41). ”No effect concentrationer” (NEC) blev bestemt ved log-lineær regression (se også Fig. 7) som skæringen mellem den øverste konfidensgrænse for kontrollen og den nederste konfidensgrænse for pesticideksponeringerne (Fig. 42). Ved høje koncentrationer af pyrethroider skete der ingen yderligere stigning i driftraten og ved længerevarende eksponering skete et fald i driftraten ved stigende koncentration fordi dyrene blev inaktive (se Fig. 40). Disse værdier indgik ikke i dosis-respons regressionen. EC50 koncentrationen blev beregnet udfra dosis-respons regressionen, hvis nedre grænse defineredes af driftraten i kontrolforsøgene og den øvre grænse af den maksimale driftrate i det enkelte forsøg (se Fig. 42). I eksemplet med esfenvalerat var kontroldriften 11% og den maksimale driftrate 53% ved 63,5 ng l-1.
Figur 42. Dosis-respons relation for Gammarus pulex eksponeret til esfenvalerat. ”No effect concentration” (NEC) bestemmes ved skæringen af konfidensgrænserne for kontrolforsøg og eksponeringsforsøg (markeret med pil). Tabel 25. Effekt af pyrethroider på drift hos Gammarus, Asellus, Baetis og Leuctra. ”No effect concentration” (NEC) og EC50 angivet i ng l-1 0-40 min og 45-90 min efter eksponeringsstart. - = kan ikke beregnes; nd = ingen data.
De højeste koncentrationer af pyrethroider som ikke udløste forøget drift (NEC) og EC50 koncentrationerne varierede mellem arter og pesticider og reduceredes med varighed af eksponering. Gammarus pulex var mest følsom med en NEC-værdi for esfenvalerat så lav som 0,55 ng l-1 (Tabel 25). Sammenlignet hermed var insektnymferne Baetis og Leuctra samt Asellus mindre følsomme med 10-20 gange højere NEC- og EC50-værdier. Testene med det formulerede produkt af esfenvalerat (Sumi-alfa) viste markant lavere effektværdier hos alle arter sammenlignet med det rene stof. Med undtagelse af EC50-værdien hos Asellus var øgningen i følsomhed over tid begrænset, hvilket peger på en hurtigere effekt og optagelse end i det ikke-formulerede stof. 4.7.2 Drift under in-situ forholdDriftrespons hos vandløbsinvertebrater er undersøgt før, under og efter pulseksponering med pyrethroidet esfenvalerat i semi-naturlige vandløbsrender. Undersøgelserne fokuserede hovedsageligt på krebsdyret Gammarus pulex men også døgnfluen Baetis rhodani og vårfluen Sericostoma personatum indgik i undersøgelserne. Under forsøgene blev såvel koncentration som varighed varieret for at få et indblik i driftresponset ved en række realistiske eksponeringsscenarier. Forsøgene blev først gennemført som enkeltarts test med G. pulex og derefter med samfund bestående af alle ovenstående arter. Metode
Fig. 43. Foto af forsøgsopstillingen med de 12 vandløbsrender. Driftnettene er placeret i de lodrette nedløbsrør forrest i billedet. Tabel 26. Fysiske og kemiske parametrer for vandet i renderne
Såvel vandløbsrenderne som sedimentet i vandløbsrenderne blev udskiftet mellem alle forsøgsserier således at hver forsøgsserie indledtes med at etablere sedimentet. Først blev hver vandløbsrende tilført 4,5 kg grus (1-3 cm) fordelt jævnt på en 3 meter lang strækning. Gruset var forinden konditioneret 7 dage i vandløbsvand for at skabe en naturlig biofilm på substratet. Efter fordelingen af gruset blev der udlagt knytnæve store sten for hver 30. centimeter. Disse sten blev indsamlet samme dag som etableringen af vandløbsrenderne i Hulbæk, et lille vandløb 5 km øst for Silkeborg. Inden stenene blev udlagt i vandløbsrenderne blev de renset for dyr og skidt. Efter udlægningen af sten blev der i strømlæet bag hver sten placeret 2 elleblade (Alnus glutinósa (L)) som forinden var konditioneret 5 dage i kildevand ved 10°C. Efter udlægningen af elleblade blev en Stowaway Tidbit temperatur logger placeret i nedstrømsenden af substratsstrækningerne. Efter etableringen af substratet blev et kendt antal invertebrater introduceret umiddelbart opstrøms for den tre meter lange substratsstrækning i alle render. 1,5 time efter introduktionen af invertebraterne blev driftnettene udskiftet og de opfangede invertebrater blev reintroducret opstrøms for substratsstrækningen.
Figur 44. Foto af de 3 meter lange substrat-strækninger bestående af grus, sten og elleblade. Næste formiddag, ca. 20 timer efter introduktionen af invertebraterne, blev driftnettene skiftet igen og de fangede invertebrater kasseret og indgik dermed ikke i selve forsøget. Herefter blev driftnettene udskiftet med 45 minutters intervaller resten af dagen og igen næste formiddag, ca. 20 timer efter pesticidtilførslen. Under hver udskiftning blev de opfangede dyr skyllet ud i en hvid fotobakke, talt og konserveret i ethanol. Forsøgene blev afsluttet efter tømningen af driftnettene 20 timer efter pesticidtilførslen. Ved forsøgenes afslutning blev substratet skyllet ud af renderne og tilbageværende invertebrater blev frafiltreret og talt. De ethanolkonserverede invertebrater blev i laboratoriet efterfølgende målt, således at deres tørvægt kunne bestemmes på baggrund af tidligere beskrevne relationer mellem længden af en given morfologisk enhed (f.eks. hovedkapsel bredden) og dyrenes tørvægt (Zelinka & Marvan, 1976, Iversen & Jessen, 1977, Friberg et al. 2002). Middel tørvægten af G. pulex og B. rhodani blev således bestemt til hhv. 3,85 ± 0,09 mg og 7,24 ± 0,63 mg mens den askefri tørvægt af S. personatum blev bestemt til 5,32 ± 0,17 mg. Esfenvaleratopløsningerne blev fremstillet ved at opløse en given mængde stof i 10 ml acetone og derefter lave en fortyndingsserie med demineraliseret vand til de ønskede koncentrationer. Koncentrationerne blev fremstillet således at tilsætning af 6 ml opløsning per minut gav den ønskede nominelle koncentration. Fremstillingsmetoden betød at acetonekoncentrationen i vandløbsrenderne varierede mellem 1,9 x 10-8 – 7,4 x 10-5 %. Efter 1,5 time, og dermed to driftnettømninger, blev pulstilførslen af pesticid indledt. Pesticidpulsen blev tilført fra 1,75 liters lavementposer forbundet med et infusionssæt indstillet til at afgive 6 ml pesticidopløsning per minut til opstrømsenden af vandløbsrenderne (Fig. 45). Undersøgelserne omfattede scenarier med pulsvarighed på 30 og 150 minutter og koncentrationen varierede mellem 0,2 ng l-1 og 2 µg l-1 (Tabel 27). Undersøgelserne blev gennemført i seks serier, hvor der i hver serie blev gennemført forsøg med to pesticidkoncentrationer. Under serierne blev hver af de to pesticidkoncentrationer tilført fire vandløbsrender mens de fire resterende vandløbsrender ikke blev tilsat pesticid og fungerede som kontrol. Under enkeltartsforsøgene med G. pulex blev der udført en forsøgsserie, hvor 4 vandløbsrender blev tilsat acetone svarende til koncentrationen under eksponeringen med 2 µg l-1 i 30 minutter og 4 vandløbsrender med acetone koncentrationer som under eksponeringen for 0,0002 µg l-1 i 150 minutter. I modsætning hertil blev der under samfundsforsøgene tilsat acetone til kontrolrenderne under selve forsøget i en koncentration, der svarede til højeste koncentration anvendt i eksperimentelrender.
Figur 45. Dosering af pesticidopløsning til vandløbsrender Tabel 27 Undersøgte eksponeringsscenarier
Statistisk metode Enkeltartsforsøg med G. pulex
Figur 46. Akkumuleret drift af G. pulex under enkelt artsforsøgene med 30 minutters esfenvalerat eksponering. Den Akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten. Tabel 28. Test af forskelle mellem de undersøgte koncentrationer ved 30 minutters eksponering. Værdierne angiver ?2(1) og de korresponderende P-værdier. n angiver antallet af individer tilstede til tiden –0,75 timer.
Under forsøget med 150 minutters esfenvalerat eksponering driftede 50% af individerne ved eksponering for 0,0002 µg l-1 (0,2 ng l-1) og 95% af G. pulex driftede ud af vandløbsrenderne eksponeret for 0,002 µg l-1 (Fig. 47). Sammenholder man resultaterne fra forsøgene med 30 og 150 minutters eksponering ved 0,002 µg l-1 ses det, at driften øges fra 50 til 95% ved den lange eksponering. Desuden svarede driftresponset, målt som akkumuleret drift ved forsøgets afslutning, under 150 minutters eksponeringen cirka til responset under 30 minutters eksponeringen efter eksponering for ti gange så høje koncentrationer. Driftresponset er altså ikke kun afhængig af koncentration men også varighed af eksponeringen.
Figur 47 Akkumuleret drift af G. pulex under enkelt artsforsøget med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten. Tabel 29, Test af forskelle mellem de undersøgte koncentrationer ved 150 minutters eksponering. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
Acetoneeksponering af G. pulex påvirkede ikke driftraten selv ved den højeste acetonekoncentration benyttet under esfenvaleratforsøgene (Figur 48; Tabel 30). Den forhøjede drift under eksponeringen for esfenvaleratopløsninger kan altså udelukkende tilskrives effekt af pesticidet. Dog kan det ikke udelukkes at der findes synergistiske effekter af esfenvalerat og acetone, selvom litteraturen ikke beskriver sådanne effekter.
Figur 48. Akkumuleret drift af G. pulex under kontrol forsøgene med acetone eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af acetone eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten. Tabel 30. Test af effekten af acetone eksponering på driften blandt G. pulex. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
Samfundsforsøg Driftraten hos vårfluen S. personatum blev hverken påvirket under eller efter nogen af pesticideksponeringerne (Figur 49). Natten var det eneste tidsrum, hvor der blev fanget S. personatum i driftnettene men denne drift var ikke større efter pesticid eksponering. Den udprægede natdrift kan tilskrives at S. personatum primært søger føde om natten.
Figur 49 Akkumuleret drift af S. personatum under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten. Døgnfluen B. rhodani responderede på esfenvalerat eksponeringen med forøget drift (Figur 50A). Dog var der intet driftrespons ved eksponering for 0,0002 µg l-1 og driften var først signifikant forhøjet ved eksponering for 0,02 µg l-1 (Tabel 31). Driften var signifikant forhøjet efter eksponeringen for 0,02 og 0,2 µg l-1 men ingen af koncentrationerne medførte en drift på mere end 60% af individerne mod cirka 10% i kontrolrenderne. Tabel 31 Test af forskelle driften af B. rhodani ved de undersøgte koncentrationer. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
Figur 50A Akkumuleret drift af B. rhodani under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten. Driftresponset observeret blandt G. pulex under samfundsforsøgene var i god overensstemmelse med mønstret observeret under enkeltartsforsøgene (Figur 50B, Tabel 32). Eksponering for 0,2 ng l-1 medførte også under samfundsforsøgene, at ca. 50 % af individerne gik i drift og eksponering for koncentrationer på 0,002, 0,02 og 0,2 µg l-1 forårsagede 100 % drift blandt G. pulex. Ligeledes blev der i samfundsforsøgene observeret en klar negativ korrelation mellem eksponeringskoncentration og start på drift respons.
Figur 50B Akkumuleret drift af G. pulex under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Tabel 32 Test af forskelle i driften af G. pulex ved de undersøgte koncentrationer efter 150 min eksponering. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
Sammenligner man arternes driftrespons ved de enkelte koncentrationer fremkommer et klart billede med G. pulex som responderer med en signifikant større andel individer i drift end både B. rhodani og S. personatum ved alle de testede koncentrationer (Tabel 33). Andelen af B. rhodani i drift var dog signifikant større end andelen af S. personatum ved samtlige esfenvalerateksponeringer (Tabel 33). Tabel 33. Test af forskelle i driften hos arterne ved de enkelte koncentrationer fra start af eksponering og resten af forsøget (0 – 20 timer). Værdierne angiver ?2(1) og de korresponderende P-værdier.
Sammenligner man driftresponset hos G. pulex og B. rhodani fra starten af pesticideksponeringen og de følgende 135 minutter fremgår det klart at G. pulex responderer langt hurtigere end B. rhodani på esfenvalerat (Tabel 34). Tabel 34. Test af forskelle i drift hos G. pulex og B. rhodani under de første 135 minutter efter start af pesticideksponering. Værdierne angiver ?2(1) og de korresponderende P-værdier.
4.8 Nedbrydning af detritusVandløbsinvertebraters nedbrydning af detritus under påvirkning af pesticider blev undersøgt under laboratorieforhold i klimarum. Forsøgene fokuserede på hastigheden hvormed krebsdyret Gammarus pulex (L) nedbrød bladplader af rødel (Alnus glutinósa) efter eksponering for forskellige pesticider. Undersøgelserne omfattede studier, hvor G. pulex blev direkte eksponeret til insekticiderne esfenvalerat og dimethoat samt fungicidet azoxystrobin i pulse af 30 og 150 minutters varighed inden de blev overført til forsøgsopstillingen. Desuden blev der gennemført undersøgelser af effekten af indirekte eksponering af G. pulex for esfenvalerat og azoxystrobin gennem elleblade eksponeret for pesticiderne. 4.8.1 MetodeForsøgsopstilling
Figur 51 Forsøgsopstillingen benyttet under forsøgene med nedbrydning af detritus. Hvert 100 ml bæger glas indeholder 80 ml vand, som bliver genemluftet af iltsten, 2 ellebladplader og en G. pulex. Blade Forsøgsdyr Forsøgene blev gennemført med to insekticider: dimethoat og esfenvalerat samt et fungicid: azoxystrobin. Pesticidopløsningerne til pulseksponering af dyrene blev fremstillet med demineraliseret vand. Dog blev det aktive stof opløst i 10 ml acetone under fremstillingen af pesticidopløsningerne med de tungt opløselige stoffer esfenvalerat og azoxystrobin. Efterfølgende blev acetoneopløsningen fortyndet i demineraliseret vand så der maksimalt var 0,1 µg acetone i opløsningen dyrene blev udsat for. Samtidig blev kontroldyrene/bladene uden pesticid også tilsat 0,1 µg acetone under eksponeringen. Direkte eksponering Indirekte eksponering Tabel 35. Oversigt over de undersøgte eksponeringsscenarier
Hvert kontamineringsscenarium blev gennemført med 15 replikater. Under forsøgene blev dyrenes tilstand observet dagligt. Forsøget blev afsluttet efter syv dage, ved afslutningen blev bladresterne fra hvert bægerglas overført til forvejede foliedigler og tørret ved 60°C til konstant vægt. Længden af første thorakalled af G. pulex blev målt og tørvægt blev fundet ud fra en kendt relation mellem længde og tørvægt (Iversen & Jessen, 1977). Sideløbende med forsøgene blev 50 bladpladepar konditioneret og vejet som under de øvrige forsøg. Bægerglassene de blev overført til indeholdt dog ingen dyr, hvorfor forholdet mellem vådvægt før og tørvægt efter forsøg uden påvirkning af dyr kunne etableres. 4.8.2 ResultaterHastigheden hvormed krebsdyret G. pulex nedbrød groft detritus (CPOM) til fint detritus (FPOM) efter pulseksponering for pesticider var generelt nedsat. Nedbrydningshastigheden efter eksponering for insekticidet esfenvalerat faldt med stigende eksponerings koncentration (Figur 52). Nedbrydningshastigheden faldt efter 30 minutters eksponering for 0,02 µg l-1 esfenvalerat og faldet var signifikant efter eksponering for 0,2 µg l-1 (Tabel 36). Det meget kraftige fald i nedbrydningshastighed efter eksponering for 2 µg l-1 skyldes primært, at 14 af 15 individer døde efter overførsel fra esfenvaleratopløsningen til forsøgsopstillingen. At bladvægttabet kan være negativt skyldes primært to ting: 1) Ikke to bladpar er ens og mængden af nervation i bladpladerne vil have betydning for det forhold vi etablerede mellem vådvægt før og tørvægt efter, 2) Mængden af mikroorganismer på bladene vil øges med tid og biomassen af disse mikroorganismer påvirkes af tilstedeværelsen af G. pulex. Efter 150 minutters eksponering med esfenvalerat blev der også observeret et fald i nedbrydningshastigheden med stigende koncentration. Faldet var dog ikke signifikant på 5 % niveau efter eksponering for 0,2 µg l-1 (Tabel 36). Faldet i nedbrydningshastighed var ikke entydigt blandt de testede insekticider da stoffet dimethoat ikke påvirkede nedbrydningshastigheden selv efter eksponering for 2000 µg l-1 (Bilag E). Tabel 36. Test af forskel fra kontrol under forsøg med esfenvalerat samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de korresponderende P-værdier.
Pulseksponering med fungicidet azoxystrobin forårsagede et fald i nedbrydningshastigheden hos G. pulex (Figur 53). Ved 30 minutters eksponering var falder signifikant efter eksponering for 20 µg l-1 (Tabel 37). Det kraftige fald i nedbrydningshastigheden efter eksponering for 2000 µg l-1 skyldes primært at 12 af 15 individer døde efter overførsel fra pesticidopløsningen til forsøgsopstillingen (Tabel 37). Nedbrydningshastigheden efter 150 minutters eksponering var også negativt korreleret med azoxystrobin koncentrationen dog uden at nogen af koncentrationerne medførte et signifikant fald nedbrydningshastigheden på 5 % niveau (Tabel 37). Tabel 37. Test af forskel fra kontrol under forsøg med azoxystrobin eksponeret til G. pulex samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de korresponderende P-værdier.
Under forsøgene med indirekte eksponering var G. pulex ikke eksponeret for pesticiderne i vandfasen men derimod gennem deres fødeemne - elleblade. Denne form for eksponering medførte nedsat nedbrydningshastighed under forsøgene med blade eksponeret for esfenvalerat (Figur 54). Nedbrydningshastigheden var signifikant nedsat ved eksponering af bladene for 0,002 µg l-1, hvorimod nedbrydningshastigheden ved blade eksponeret for 0,02 µg l-1 ikke var signifikant mindre end kontrolforsøgene (Tabel 38). Nedbrydningshastigheden var yderligere kraftigt reduceret under forsøgene med blade eksponeret for 0,2 og 2 µg l-1 (Figur 54). I ingen af forsøgene døde mere end et individ per eksponeringskoncentration og faldet i nedbrydningshastighed er altså ikke et artefakt af dødelighed blandt G. pulex som følge af pesticideksponering (Tabel 38). Forsøgene med indirekte eksponering af G. pulex for azoxystrobin gennem føden medførte ingen signifikant ændring af nedbrydningshastigheden (Bilag E).
Figur 54 Nedbrydningen af detritus efter syv dages eksponering af elleblade i esfenvalerat er afbilledet som bladpladernes vægttab per mg dyr mod eksponeringskoncentration. Søjlediagrammerne angiver gennemsnittet af de 15 replikater med standard error. * indikerer at bladvægttabet er signifikant forskelligt fra kontrollerne (F-test, p<0,05) Tabel 38 Test af forskel fra kontrol under forsøg med esfenvalerat eksponering af blade (7 dage) samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de korresponderende P-værdier.
5 DiskussionProjektets overordende mål var at undersøge og kvantificere effekter af almindeligt anvendte eller fundne pesticider under forsøgsforhold der var sammenlignelige med eller som gør det muligt at overføre resultaterne til mulige effekter i mindre vandløb. Det var endvidere et mål at sammenligne følsomheden af forskellige arter og effektparametre. Det har været tilstræbt at belyse mange forskellige problemstillinger omkring pesticiders effekt i vandløb og hvis forsøgsresultater har peget på særlige forhold, er der gået i dybden med disse. De undersøgte pesticider (ialt 14) udgør et repræsentativt udvalg af de pesticider som anvendes i jordbruget og/eller som er påvist i danske vandløb, og hovedparten af de aktive stofgrupper og virkemekanismer er repræsenteret. De biologiske effektundersøgelser har spændt vidt, både hvad angår eksponeringsmåde, -varighed og anvendte organismer. Der er undersøgt for effekter af kortvarige pulseksponeringer, der udfra målinger i danske vandløb er de mest realistiske, samt effekter under kontinuert eksponering til pesticider. Der er undersøgt effekter på “naturlige” mikroalgesamfund som repræsentant for primærproducenterne i vandløb. Disse undersøgelser har omfattet effekter på funktionelle (primærproduktion) og struktuelle (forskydninger mellem forskellige grupper af alger) parametre. Indenfor vandløbsdyrene er der undersøgt for direkte effekter af pesticider på overlevelse, adfærd (drift), samt indirekte effekter i form af nedbrydning af detritus. Forsøgsorganismerne har bestået af en række arter (ialt 11 arter) repræsenterende forskellige taksonomiske (krebsdyr, insekter, snegle) og fødebiologiske grupper. Der er endvidere undersøgt for effekter på forskellige livsstadier og der er gennemført undersøgelse af effekt på fuld livscyklus hos et insekt. I det følgende sammenlignes resultaterne med andre undersøgelser af samme eller beslægte pesticider og sandsynligheden for effekter under danske forhold vurderes i lyset af forbrug og fund af de undersøgte pesticider i danske vandløb. Yderligere sammenlignes følsomheden af de forskellige forsøgsmetoder og organismer og ved sammenligning med resultater fra standardtest diskuteres om der er behov for at udvide batteriet af standardtest med test som er målrettet mod at forbedre risokovurderingen af pesticider i vandløb. 5.1 Effekt af herbicider og fungicider på bundlevende mikroalgerI forsøgene er anvendt 2 effektparametre, fotosynteseaktivitet samt algebiomasse/sammensætning, der er vitale parametre for funktionen og delvist den biologiske struktur i vandløb. De undersøgte fotosyntesehæmmere, isoproturon og metribuzin havde ikke overraskende en entydig effekt på fotosynteseaktiviteten, Det var dog overraskende at effektkoncentrationerne var så lave som 0,3 - 1 µg l-1. Sammenlignet med fotosyntesehæmningen optrådte målbare effekter på biomasseudvikling ved væsentligt højere koncentrationer (isoproturon: 30 µg l-1, metribuzin: 50 µg l-1). Disse værdier er i god overensstemmelse med undersøgelser af væksthæmning hos planktonalger (Chlorella pyrenoidosa, Scenedesmus subspicatus, Chlamydomonas reinhardtii) med EC50,96timer mellem 12 og 40 µg l-1 (Traunspurger et al. 1996, Anton et al. 1993), mens koncentrationen og diversiteten af mikrobentiske kiselalger var reduceret ved 5 µg l-1, som var den laveste testkoncentration i forsøg udført af Pérès et al. (1996). I sidstnævnte forsøg var eksponeringsvarigheden dog væsentligt større end i nærværende undersøgelse. I NOVA programmet er isoproturon påvist i mere end 25 % af de analyserede prøver med en maksimal værdi på 0,63 µg l-1 og 3 % af prøverne havde indhold over 0,3 µg l-1 (DMU 2002). Metribuzin forekom i mindre end 5% af de undersøgte prøver, mens triaziner med samme giftighed som metribuzin (atrazin, terbuthylazin, simazin) forekom i mere end 30% af prøverne, med en maksimal koncentration på 2,5 µg l-1 . Det er derfor meget sandsynligt, at fotosyntesehæmmere kan påvirke stofproduktionen i enkelte vandløb. Da varigheden af forhøjede koncentrationer typisk er 2-10 timer i små vandløb (Styczen et al. 2003) og effekterne af fotosyntesehæmmerne var fuldt reversible efter 2-3 dage i rent vand vurderes den direkte effekt på mikroalgerne af denne stofgruppe dog at være af mindre betydning. Effektkoncentrationer af fungiciderne azoxystrobin og fenpropimorph var høje, omk. 1 mg l-1. EC50 for azoxystrobin i væksthæmningstest med planktonalger angives til mellem 100 og 360 µg l-1 med grønalger som de mest følsomme (EU 1998, USEPA 1997). For fenpropimorph varierer EC50 fra 170 µg l-1 til mere end 2 mg l-1. Set i forhold til en lav mobilitet på grund af lave opløseligheder og høj partikelaffinitet er det lidet sandsynligt at disse fungicider forårsager effekter på mikroalger i vandløbene. I NOVA programmet er fenpropimorph påvist i 2-4% af de undersøgte prøver, med en median-maximum koncentration på 0,07 µg l-1 (DMU 2001, DMU 2002). Med den baggrund anser vi effekter af fenpropimorph på algesamfundene for usandsynlige. Azoxystrobin indgår ikke i NOVA overvågningen (DMU 2001) og forekomst i danske vandløb er derfor dårligt belyst. Som det var tilfældet for fotosyntese-hæmmerne var effekterne (reduceret biomasse og fotosyntese) også fuldt reversible i løbet af 2-3 dage. Metsulfuron-methyl påvirkede fotosyntesen hos bentiske mikroalger ved koncentrationer over 6-7 µg l-1 og kun efter længerevarende eksponering (48 timer), mens der der kun kunne påvises effekter på biomassen ved koncentrationer over 1 mg l-1. Som det var gældende for isoproturon og metribuzin var var både fotosynteseraten og biomassen genetableret efter 48 timer ved alle undersøgte koncentrationer. Effektkoncentrationer i væksttest med planktonalger viser meget stor variation fra 2,3 og 1250 µg l-1 (Blanck & Björksäter 1998, Fahl et al. 1995). Den lave værdi blev fundet i en enkelt art blandt 40 eksponeret til midlet Ally og må vurderes som utypisk, da den gennemsnitlige EC50 af alle arter var ca. 100 gange højere. Indenfor gruppen af stoffer med samme virkemekanisme (hæmmer af aminosyresyntese, Acetolactat synthase, ALS) er effekten overfor planktonalger meget variabel, mens effektkoncentrationerne for højere planter (andemad) typisk er 100 gange lavere (Battaglin & Fairchild 2002). Da forbruget af metsulfuron og andre herbicider med samme virkemekanisme er meget begrænset og de ikke er fundet i danske vandløb under NOVA programmet (DMU 2002) er det ikke sandsynligt, at denne gruppe af herbicider vil have effekter på mikroalger i danske vandløb. Pendimethalin var giftig overfor bundlevende alger og i modsætning til fotosyntesehæmmerne og fungiciderne var reetablering af samfundene efter endt eksponering meget langsom. Især grønalger var følsomme og effekten på deres biomasse fortsatte selv efter overførsel til rent vand ved den lavest anvendte koncentration (10 µg l-1). I litteraturen angives effektkoncentrationer (EC50) overfor planktonalger til mellem 5 µg l-1 og 72 mg l-1 (f.eks. Clausen 1998, Møhlenberg et al. 2002). I flere lande udnytter man den store giftighed og bekæmper aktivt alger i det terrestiske miljø med pendimethalin. I denne undersøgelse var biomassen af grønalger reduceret med 50% efter 24 timers eksponering til 10 µg l-1 og en ”recovery periode” på 192 timer. Pendimethalin bindes stærkt til partikler og der er forbud mod sprøjtning i en 10 m zone langs vandløb, hvilket taler imod en signifikant transport til vandløbene. Både forsøg og overvågning i vandløb viser dog, at pendimethalin kan transporteres til vandløbene. I forsøg med simulerede nedbørshændelser (70-110 mm time-1) 24 timer efter udsprøjtning blev der tabt ca. 0.1% af den udsprøjtede mængde pendimethalin ved afstrømning, heraf blev 35% transporteret i partikelbundet form (Müller et al. 2002). Sammenlignet med mere opløselige herbicider var koncentrationerne i det afstrømmende vand lave (12-20 µg l-1) og tabsprocenten 3-10 gange lavere end de øvrige undersøgte pesticider. Pendimethalin optræder i ca. 10% af undersøgte prøver fra danske vandløb (DMU2002) men koncentrationerne er normalt lave. Nedbrydningen af pendimethalin sker langsomt i jord og sammen med det store forbrug er det antagelig årsagen til de høje fundprocenter i vandløbene. Grundet den store partikelaffinitet må det formodes, at sedimentet i vandløbenes aflejringsområder i perioder er beriget med pendimethalin. Vi er dog ikke bekendt med sådanne målinger og det er uvist om sedimentbundet pendimethalin vil være tilgængeligt for algerne. Undersøgelser har dog vist, at toksiciteten af herbicider med stor partikelaffinitet reduceres op til 900 gange, hvis der er sediment tilstede i forsøgene (Shillabeer et al. 2000) 5.2 Effekt af pesticider på vandløbsdyr5.2.1 MortalitetMed undtagelse af ioxynil har forsøgene med vandløbsdyr været centreret om undersøgelse af effekter af fungicider og insekticider. Gammarus pulex har været den langt mest benyttede organisme og forsøgene med denne art dominerer dette afsnit. Ioxynil var kun svagt toksisk overfor Gammarus pulex (EC50,14 dage= 5,7 mg l-1) og effekter indtrådte først efter 4 dages eksponering. Til sammenligning påvirkes dafnier tidligere, udtrykt ved en sammenlignelig EC50 værdi mellem 3,9-5,6 mg l-1 , der dog blev bestemt efter 48 timer (f.eks. Clausen 1998, Møhlenberg 2002). I NOVA programmet er fundprocenterne lave (6%) og den maksimalt målte koncentration er 0,033 µg l-1 (DMU 2001). Der vil derfor være minimal risiko for at ioxynil kan have effekter på invertebratfaunaen i danske vandløb. Azoxystrobin var toksisk overfor Gammarus pulex med stærkt faldende effektkoncentration ved øget varighed af eksponering, (LC50,2timer = 15 mg l-1, LC50,24timer = 120 µg l-1, LC50,4dage = 20 µg l-1, LC50,10dage = 10 µg l-1. Disse effektkoncentrationer er lavere end i andre sammenlignelige test med G. pulex (LC50,4dage = 350 µg l-1) og i test med Daphnia magma (110-280 µg l-1, Møhlenberg 2002). Kortvarige eksponeringer (½-2 timer) førte til ”forsinket” mortalitet i en efterfølgende periode med rent vand, men kun hvis eksponeringskoncentrationen havde været urealistisk høj (2,5 mg l-1). Derimod blev fødebearbejdningsaktiviteten hos G. pulex (målt som nedbrydning af elleblade) signifikant reduceret ved selv kortvarige (½ time) eksponeringer til 20 µg l-1 og højere koncentrationer. Sammenlignet hermed var der ingen effekt på nedbrydningen af kontaminerede blade. Koncentrationen af azoxystrobin i bladene var dog ukendt. Azoxystrobin er først indført i jordbruget i 1998 og følgelig er der ringe erfaring med forekomst i danske vandløb. Azoxystrobin indgår ikke NOVA programmet. Forekomst af azoxystrobin i koncentrationer omkring 20 µg l-1 i danske vandløb er ikke sandsynlige selv om mobiliteten er relativt høj og nedbrydningen meget langsom, men da azoxystrobin er nyt på markedet og endnu ikke har indgået i omfattende overvågningsprogrammer kendes de realistiske koncentrationer ikke. Azoxystrobin påvirkede ikke driften hos G. pulex, hvilket er i overensstemmelse med undersøgelser af bevægelsesadfærden under eksponering til azoxystrobin (Nørum & Bjerregaard 2003). Prochloraz var moderat toksisk overfor Gammarus pulex med stærkt faldende effektkoncentration med øget eksponeringstid (LC50,24 timer = 22 mg l-1, LC50,48 timer = 7 mg l-1, LC50,14 dage = 0,6 mg l-1). Under sammenlignelige forsøgsforhold var effektkoncentrationen hos Daphnia magma lavere (LC50,48 timer = 2,6 mg l-1, NOEC21dage = 22 µg l-1, Clausen 1998, Møhlenberg et al. 2002) end hos G. pulex. Ved en eksponeringsvarighed som er realistisk for mindre vandløb (< 24 timer) var der ingen akut mortalitet selv ved 10 mg l-1. Derimod blev der ”afsat” toksicitet ved kortvarige (½-1 time) eksponeringer til meget høje koncentrationer (>5 mg l-1) og disse kom til udtryk ved forøget mortalitet i de efterfølgende 5-10 dage i rent vand. Prochloraz er ikke påvist i danske vandløb under NOVA programmet og sammen med de høje effektkoncentrationer anser vi det for usandsynligt at fungicidet kan påvirke invertebratfaunaen i danske vandløb. Toksiciteten af insekticidet dimethoat var lav overfor alle undersøgte invertebrater med LC50,96 timer værdier på 5 mg l-1 eller højere. Hos Gammarus pulex øgedes toksiciteten med eksponeringsvarigheden, men selv efter 14 dages eksponering var LC50 værdien højere end 1 mg l-1. I litteraturen er der stor uenighed om den akutte toksicitet (LC50) af dimethoat overfor gammarider, fra 180 µg l-1 (Bækken & Aanes 1991), 200 µg l-1 (Johnson & Finley 1980), 2,4-3,5 mg l-1 (Nørum & Bjerregaard 2003) til 4,1 mg l-1 (Pantani et al. 1997). Den lavest angivne LC50,96 timer for vandløbsdyr (Baetis rhodani) er 7 µg l-1 doseret som aktivstof (Bækken & Aanes 1991). Effektkoncentrationer af dimethoat er generelt lidt lavere hos dafnier (gennemsnit LC50,48 timer = 460 µg l-1, Clausen 1998, Møhlenberg 2002) selv om variationen er meget stor,). Der var ingen effekt på Gammarus' nedbrydning af elleblade selv efter eksponering til 2 mg l-1. Set i forhold til meget lave fundprocenter og koncentrationer i danske vandløb (DMU 2002) anser vi derfor at effekter af dimethoat på vandløbsdyr for lidet sandsynlige. Pirimicarb havde den laveste toksicitet af de undersøgte pesticider overfor Gammarus pulex. Eksponering til så høje koncentrationer som 5 mg l-1 medførte ingen toksicitet selv efter 14 dage. Hos larver af stikmyg (Culex quinquefasciatus) angives LC50,24 timer til 8,5 mg l-1 (Magnin et al. 1988). Til sammenligning er toksiciteten af pirimicarb langt højere overfor dafnier (LC50,48 timer = 14-22 µg l-1, NOEC21dage = 1-2 µg l-1, Kusk 1996). Årsagen til denne usædvanligt store forskel i følsomhed er ikke kendt. Hvis de høje effektkoncentrationer overfor G. pulex er repræsentative for vandløbsdyrene generelt (se nedenfor) anser vi ikke at effekter af pirimicab er sandsynlige i danske vandløb, set i lyset af lave fundprocenter (4%) og meget lave koncentrationer, der er påvist (max 0,026 µg l-1). Toksiciteten af insekticidet diflubenzuron blev undersøgt overfor 5 forskellige invertebrater i akuttest. Der kunne ikke påvises nogen akut (96 timer) mortalitet ved de højeste anvendte koncentrationer (Gammarus: 0,2 mg l-1, øvrige arter: 3-5 mg l-1). Insekticider indenfor benzoylphenylurea gruppen er specifikt virkende på vækst/hudskifte hos insekter (og formentligt krebsdyr) og der kan derfor ikke forventes effekter ved kortvarige eksponeringer. I feltundersøgelser er er diflubenzuron vist at påvirke populationer af dafnier ved 5 µg l-1 (WHO 1995). I Danmark anvendes insekticider af benzoylphenylurea gruppen ikke i jordbruget og risikoen for transport til vandløb er meger ringe. Esfenvalerat var langt det mest giftige af de undersøgte pesticider. Standardiserede akuttest viste LC50,48 timer værdier for Gammarus pulex på 120 ng l-1, som er på niveau med de laveste værdier hos dafnier (LC50,48 timer = 100-1000 ng l-1, Clausen 1998, Møhlenberg 2002). Med øget eksponeringsvarighed (14 dage) reduceredes LC50 værdien til ca 30 ng l-1. Set i lyset af de korte varigheder af forhøjede koncentrationer i vandløb var det vigtigt, at selv kortvarige (½-1 time) eksponeringer til lave koncentrationer efterfølgende havde effekt på overlevelse, vækst/udvikling, reproduktion, fødebearbejdning samt adfærd i form af drift. En 1-times pulseksponering til esfenvalerat ved koncentrationer på mellem 100 – 600 ng l-1 havde effekter på overlevelsen, parringsadfærd bestemt som gendannelse af par, og det reproduktive output hos Gammarus pulex, der stadig kunne detekteres mindst to uger efter pulseksponeringen. Trods 100 % overlevelse under pulseksponering op til 2 g l-1 steg dødeligheden i visse tilfælde markant efter overførselen til rent vand. Lignende observationer hos G. pulex udsat for stoffer såsom cadmium, cyanid og pyrethoridet permethrin er gjort af Abel & Garner (1986). I denne undersøgelse var eksponeringskoncentrationerne, der medførte effekter efter kun en 1-times puls, ikke meget højere end de estimerede LC50-værdier (96 timer). Det betyder, at den toksiske dosis afsættes hurtigt i dyrene, men at effekterne i form af død først bliver synlige senere. Det betyder videre, at brugen af endpoints fra standard akuttoksicitetstests (f.eks. eksponering i 48 timer) ikke nævneværdigt overestimerer effekterne selv ved meget korte eksponeringer. Reproduktionsadfærden (prækopula) hos G. pulex viste sig at være yderst sensitiv overfor eksponeringer med esfenvalerat og en eksponeringskoncentration på 50 ng l-1 førte til øjeblikkelig forstyrrelse og separation af parrene, hvor de gravide hunner straks smed deres afkom. Denne forstyrrelse af parrene kan således virke forsinkende på reproduktionen, idet mange af de forstyrrede par aldrig eller først sent gendannedes efter overførsel til rent vand. Maltby & Naylor (1990) fandt lignende effekter hos G. pulex hunner eksponeret til zink over en periode på tre til fire uger, idet de observerede en stigning i antallet af aborterende hunner og dermed stigende variation i antallet af unger. De forklarede disse observationer ved en reduktion af den tilgængelig energimængde til vækst og reproduktion, forårsaget af nedsat fødeoptag hos de eksponerede hunner. Effekten af esfenvalerat på reproduktionadfærden hos G. pulex var dog momentan og skyldes nok den direkte effekt på impulsledningen i nerverne. I forsøgene med Chironomus riparius eksponeret til miljørealistiske koncentrationer (200 ng l-1) af esfenvalerat blev det vist, at eksponeringsvejen er af stor betydning for overlevelsen. Således blev der observeret en meget lavere klækning af individer i det forsøg, hvor dyrene var blevet pulseksponeret udenfor sedimentet, modsat forsøget hvor dyrene blev pulseksponeret, mens de befandt sig i nede i sedimentet. Da eksponeringsscenariet i andet forsøg formodes at repræsentere den mest realistiske eksponeringsvej er der på baggrund af de opnåede resultater ikke belæg for at tro, at kortvarige miljørealistisk pulser af esfenvalerat udgør en trussel for populationsdynamikken hos C. riparius. Det var imidlertid lidt overraskende, at overlevelsen af den anden undersøgte chironomid-art Prodiamesa olivacea, der ikke er nær så tolerant overfor lave iltkoncentrationer som C. riparius, ikke blev påvirket af esfenvalerat selv ved koncentrationer på op til 2 g l-1. Både i forsøg med og uden sediment under eksponeringen var klækningstiden hos hunner større end hos hanner. Dette er i overensstemmelse med tidligere studier, hvor der ikke er eksponeret med pesticid, hvorfor denne observation ikke kan relateres til eksponeringen med esfenvalerat. 5.2.2 DriftEffekt af pesticider på driften hos vandløbsdyr blev undersøgt ved to forskellige metoder, dels i recirkulerende render i laboratoriet, dels under in situ lignende forhold i langt større gennemstrømssystemer, hvor vand (og pesticidopløsning) kontinuerligt blev fornyet. I de recirkulerende render blev driften kvantificeret ved andelen af organismer som var i drift på et givet tidspunkt, mens driften i gennemstrømsrenderne blev kvantificeret som andelen der var driftet ud af renderne over en længere periode. Teoretisk vil gennemstrømsprincippet og opsamling af driftede individer være mest følsom, fordi drifteffekten akkumuleres over en periode på flere timer. Alligevel var effektgrænserne sammenlignelige i de to undersøgelser. Fordelene ved de recirkulerende render er en mere simpel logistik og forsøgsgang samt at resultaterne på grund af en større tidsopløsning umiddelbart kan anvendes i koblede skæbne-effekt modeller. I enkeltarts- og samfundsforsøgene gennemført under ”in situ” forhold var der forøget drift hos Gammarus pulex ved den lavest testede koncentration af esfenvalerat (0,2 ng l-1) og bedømt på responset er det sandsynligt at den laveste effektkoncentration er væsentligt under denne værdi (se f.eks. Fig. 47 & 50). Hos G. pulex var der en klar positiv korrelation mellem driftaktiviteten og esfenvaleratkoncentrationen under eksponeringen. I de recirkulerede render gennemførtes forsøgene ved 6-8 forskellige koncentrationer og det var muligt at beregne en ”no effect concentration” af esfenvalerat på 0,55 ng l-1 ved regression. Tiden fra start af esfenvalerateksponering og til start af driftrespons var negativt korreleret hos Gammarus pulex, driftresponset blev således initieret tidligere jo højere koncentration de blev eksponeret for. Hos de øvrige undersøgte arter var denne sammenhæng ikke så udpræget og den forøgede drift udløstes senere. Det hurtige respons hos G. pulex tyder på at de sanser pesticidet og flygter for at søge et refugium. Denne teori understøttes af et bioassay udført af Schulz og Liess (1999) hvor de indirekte påviser at G. pulex søger fra et hovedløb kontamineret med fenvalerat (pyrethroid) og parathion-ethyl op i et pesticidfrit tilløb. Yderligere støtte til teorien findes i studier af driftadfærden hos G. pulex under pyrethroid eksponering, hvorunder det blev observeret at hovedparten af G. pulex i drift var aktivt svømmende (Liess 1994) og i undersøgelser af bevægelsesadfærden kvantificeret ved videosporing (Nørum & Bjerregaard 2003). Det noget langsommere driftrespons hos de øvrige arter med driftrespons skyldes formentligt, at de først initierer drift, når de er fysiologisk påvirket af pyrethroidet. Denne teori understøttes af tidligere driftundersøgelser udført med det nært beslægtede pyrethroid lambda-cyhalothrin, hvor op mod 80 % af de driftende Baetis rhodani udviste påvirket adfærd, mod under 10 % af G. pulex (Lauridsen 2002). Sammenholdes resultaterne fra 30 og 150 minutters eksponering under enkeltartsforsøgene med G. pulex i gennemstrømsrenderne, samt resultaterne i 0-40 min og 45-90 min perioderne i forsøgene i recirkulerede render fremgår det, at ikke kun eksponeringskoncentrationen men også eksponeringstiden har betydning for driftresponset. Eksempelvis driftede 50 % af individerne under forsøgene med 30 minutters eksponering ved 2 ng l-1 mod 95 % under forsøgene med 150 minutters eksponering for samme koncentration. For at give et bud på responset på en given eksponering er det altså ikke nok at kende koncentrationen - man må også have kendskab til eksponeringens varighed. Driftforsøg i de recirkulerende render viste at esfenvalerat, når det forekom som formuleret produkt (Sumi-alfa) udløste forøget drift ved koncentrationer der var 3-20 gange lavere end det rene stof. Handelsprodukter er ofte tilsat overfladeaktive stoffer for at øge pesticidernes optag i og effekten på målorganismer. En sådan forøgelse af ”effektiviteten” gjorde sig også gældende i form af forøget driftrespons, når det formulerede produkt anvendtes i forsøgene. Driftrespons efter eksponering med esfenvalerat og andre pyrethroider er ofte rapporteret i litteraturen (Kreutzweiser & Sibley 1991, Breneman & Pontash 1994, Liess 1994, Jensen 2001). Imidlertid er der aldrig tidligere rapporteret forhøjet drift blandt invertebraterne ved en koncentration så lav som 0,2 ng l-1. Den lave effektkoncentration er dog i overensstemmelse med undersøgelser af bevægelsesadfærden hos Gammarus pulex, hvor den tilbagelagte afstand blev øget ved 0,3 ng l-1 og højere koncentrationer (Nørum & Bjerregaard 2003). Det observerede driftrespons ved en nominel koncentration på 0,2 ng l-1 er langt lavere end de højeste koncentrationer af esfenvalerat målt under flomhændelser i danske vandløb på 0,66 µg l-1 (Wiggers, 1999). Umiddelbart vil man derfor forvente, at brug af esfenvalerat fører til driftrespons blandt vandløbenes invertebrater. Imidlertid binder esfenvalerat stærkt til organisk materiale og esfenvalerat målt i vandløb vil derfor primært forekomme som bundet til suspenderet stof. Undersøgelser har dog vist, at også partikelbundet pyrethroid har effekt på driften blandt invertebrater selv om effektkoncentrationerne er højere (Schulz & Liess 2001). Effektkoncentrationer under 1 ng l-1 leder desuden til en ny problemstilling, da det ikke muligt at måle så lave koncentrationer med de tilgængelige metoder. Situationen er altså den, at vi observerer effekt af koncentrationer som ikke kan detekteres ved kemisk analyse. De økologiske effekter i naturlige vandløbssystemer af en øget drift vil afhængige af de tidslige og rumlige forhold. En kortvarig pulseksponering af en vandløbsstrækning med esfenvalerat vil pga. af økosystemets evne til at rette sig selv op ikke have nogen permanent effekt. Imidlertid må det forventes at såfremt koloniseringsmulighederne ikke er optimale, og såfremt vandløbet påvirkes gentagne gange, vil der kunne ske både strukturelle og funktionelle ændringer i økosystemet. Ligeledes vil påvirkninger af lange vandløbsstrækninger, evt. helt fra udspringet af mindre vandløb, kunne have store effekter på invertebratsamfundet. På baggrund af de gennemførte forsøg er det desuden svært at forudsige ændringer på samfundsniveau, da vi kun undersøgte enkeltarter og meget simple samfund bestående af tre arter. På længere sigt kan selv små påvirkninger forskyde konkurrence og prædationsforhold, og således ændre samfundets struktur. Lampert et al. (1989) har således for planktoniske systemer vist at samfundsniveauet er langt mere følsomt for herbicider end de enkelte arter som samfundet er opbygget af. 5.2.3 FødebearbejdningEksponeringen til esfenvalerat nedsatte generelt hastigheden hvormed Gammarus pulex nedbrød detritus i form af elleblade med en laveste effektkoncentration på 200 ng l-1. Lignende effekter af andre toksiske stoffer som tungmetaller og pesticider på vækst og ædelyst hos G. pulex er tidligere rapporteret (Naylor et al.,1989, Blockwell et al., 1998). Effekten på fødebearbejdningen var ikke større ved 150 minutters eksponering sammenlignet med 30 minutters eksponering. Der var tværtimod en tendens til at effekterne var mindre, og overvejende ikke-signifikante, ved 150 min eksponering. Årsagen til at de signifikante sammenhænge forsvandt ved 150 min eksponering er formentlig forhold relateret til eksperimentet, og er derfor ikke udtryk for en reel forskel mellem de to eksponeringsscenarier. Det virker sandsynligt, at der ikke skulle være nogen reel forskel på om G. pulex blev eksponeret i 30 eller 150 min med esfenvalerat og azoxystrobin. Indirekte eksponering af Gammarus pulex til esfenvalerat gennem eksponerede elleblade påvirkede også nedbrydningshastigheden i negativ retning med en laveste effektkoncentration på 2 ng l-1 under bladeksponering. Effekt på hastigheden, hvormed G. pulex nedbryder elleblade eksponeret for pyrethroider er tidligere observeret (Lauridsen, 2002). Eksponering af elleblade i azoxystrobin havde derimod ingen effekt på G. pulex. Både esfenvalerat og azoxystrobin havde effekt under den direkte eksponering men kun esfenvalerat påvirkede nedbrydningshastigheden ved indirekte eksponering. Dette kan skyldes forskellig virkemåde men kan også skyldes esfenvalerats meget høje affinitet for organisk materiale som medfører kraftig opkoncentrering af esfenvalerat til ellebladene under eksponeringen. Den kraftige binding af esfenvalerat og andre pyrethroider til organisk materiale kan medføre tilbageholdelse i vandløbet og kan som bundet til sedimentet formindske findelingen af organisk materiale i vandløbet. Hvad enten formindsket nedbrydningshastighed skyldes direkte eller indirekte eksponering for pesticiderne vil det have betydning for vandløbets økosystem. Iturivernes findeling af blade til fint organisk materiale er nemlig essentiel for andre led i fødenettet som filtratorer og samlerer, der lever af det fine organiske materiale. Formindskes disse organismers fødemængde vil det også påvirke højere trofiske niveauer. Forsøget viser også, at vandløbsinvertebrater påvirkes af pesticider, der er bundet til deres føde. Derfor må det forventes at de partikelbundne pyrethroider, der tilføres vandløbene fra dræn og indgår i vandløbets pulje af organisk stof, ligeledes vil påvirke de invertebrater som lever af denne føderessource. 5.3 Sammenligning af følsomhed mellem arter og effektparametreI forsøgene er der undersøgt for effekter på flere arter eksponeret til den samme pesticid og der er undersøgt for effekter på forskellige parametre indenfor samme art eller funktionelle gruppe. Dette afsnit vil forsøge at sammenholde disse informationer, der er vigtige for vurdering af mulige effekter i vandløb, hvis vi kun kender effektkoncentrationer for enkelte arter eller enkelte effektparametre. Ligeledes vil vi sammenligne effektværdierne for vandløbsorganismer med tilsvarende effektværdier for standardorganismer så som planktonalger og dafnier. Dette giver mulighed for at vurdere om resultater fra standardforsøg kan anvendes i risikovurderinger under vandløbsforhold. Figur 55 viser effektværdierne af 6 forskellige pesticider på bentiske mikroalger (fra denne undersøgelse) og litteraturværdier af EC50,96 timer fra standardiserede væksttest med planktonalger. Det skal understreges, at varigheden af eksponering er forskellig i forsøgene: i standardtest er effekten bestemt efter 96 timer, mens i denne undersøgelse har den maksimale eksponeringstid været 48 timer. Effektparametrene er ikke ens: værdierne for fotosyntese og væksthæmning hos planktonalger er repræsenteret ved EC50-værdier, og ”biomasse” ved den laveste koncentration, hvor enten den samlede biomasse eller biomassen af én algegruppe afveg fra kontrollen. Ligeledes er der stor forskel på effektusikkernederne angivet i figuren. Usikkerhederne på fotosynteseværdierne angiver 95% konfidensinterval beregnet udfra dosis-responskurverne, usikkerheden på biomasseværdierne er angivet som en ”fast” fraktion (0.22) af middelværdien (se afsnit 3.2.2), og usikkerhedsangivelsen på planktonalgerne repræsenterer standardafvigelsen i EC50 værdierne mellem forskellige arter. Det er dog vigtigt at bemærke, at ”biomassen” hos de bentiske alger inkluderer adskellige arter og grupper, som må formodes at variere i følsomhed overfor de forskellige pesticider. Nettoresultatet kan så formelt sammenlignes med middelværdien af effektkoncentrationer fra forskellige planktonarter og forsøg. Indenfor de bentiske mikroalger var fotosynteseraten generelt mere følsom (lavere effektkoncentration) end ændringer i biomassen, mest udpræget hos fotosyntesehæmmerne isoproturon og metribuzin. En klar undtagelse var dog forsøget med pendimethalin, hvor reduktionen i grønalger skete ved langt lavere koncentration end hæmning af fotosyntesen. Det skyldes, at grønalgerne delvist blev erstattet med mindre følsomme algegrupper, f.eks. kiselalger og blågrønalger, som kunne opretholde en høj fotosyntese. Ved sammenligning af effektværdierne for biomasseudvikling hos bentiske alger, og hæmning af vækst hos planktonalger er der ikke et entydigt mønster, som dels skyldes en stor variation i EC50- værdier hos planktonalgerne, f.eks. i forsøg med metsulfuron og især pendimethalin er der få meget høje værdier, som ”trækker” middelværdien op. Det er dog klart, at azoxystrobin er væsentligt mere toksisk overfor planktonalger end overfor bentiske alger. Med forbehold for de omtalte usikkerheder og de forskellige effektparametre er der intet, som tyder på, at bentiske mikroalger konsekvent er hverken mere end mindre følsomme end planktonalger. Dødelighed og drift under og efter eksponering til pesticider er i projektet undersøgt over en række arter og pesticider. Dette gør det muligt at sammenligne de forskellige arters følsomhed. Gammarus pulex var i alle sammenligninger baseret på EC50,96 timer det mest følsomme dyr blandt de undersøgte vandløbsdyr, mens rækkefølgen blandt de øvrige arter varierede afhængig af effektparametren og pesticidets art (Tabel 39). Med undtagelse af pirimicarb var effektkoncentrationerne hos Gammarus på samme niveau eller lidt lavere end hos dafnier. Invertebraternes følsomhed over for esfenvalerateksponering var vidt forskellige i driftundersøgelserne, hvor krebsdyret G. pulex responderede med forøget drift ved esfenvaleratkoncentrationer, som var 3-30 gange lavere sammenlignet med de øvrige arter (Tabel 39). Det noget svagere driftrespons blandt Baetis rhodani og Asellus skyldes formentligt, at disse arter først initierer drift, når de er fysiologisk påvirket af esfenvaleraten. Denne teori understøttes af tidligere driftundersøgelser af det nært beslægtede pyrethroid lambda-cyhalothrin, hvor op mod 80% af de driftende B. rhodani udviste påvirket adfærd, mod under 10% af G. pulex (Lauridsen 2002). Vårfluerne Sericostoma personatum og Hydropsyche udgjorde en undtagelse idet der ikke kunne påvises forøget drift. Dette skal nok tilskrives at disse arters tilbøjelighed til at gå i drift er generel lav (f.eks. Iversen 1980). Det skyldes formentlig flere årsager, bl.a. at S. personatum på grund af det tunge hus af sand og deres normale habitatvalg i langsomt strømmende partier i vandløbet ikke normalt går i drift selv når de er aktivt fødesøgende. Desuden vil store organismer som S. personatum være meget udsatte for størrelsesselektiv prædation fra f.eks. ørreder, såfremt de forekom i drift om dagen (f.eks. Brittain & Eikeland 1988). Imidlertid viste vore forsøg en tydelig natdrift i alle render hvilket formentlig afspejlede øget fødesøgningsaktivitet og det viser, at S. personatum var i stand til at gå drift under de anvendte forsøgsbetingelser. Årsagen til at S. personatum ikke går i drift er derfor muligvis, at den ikke er følsom overfor pesticideksponeringen med de anvendte koncentrationer eller mere sandsynligt, at den sænker aktiviteten ved pesticideksponering. Dette understøttes af, at tidligere observationer af S. personatum under pyrethroideksponering har vist at deres primære respons er at trække sig så langt ind i huset som muligt (Lauridsen 2002). Tabel 39. Sammenligning af følsomheder mellem vandløbsdyr og dafnier eksponeret til pesticider. Værdierne angiver den relative følsomhed i forhold til Gammarus pulex. Værdierne er baseret på LC/EC5048-96 timer værdier
* bestemt udfra adfærd. Den store følsomhed hos Gammarus sammenlignet med de øvrige vandløbsdyr er et meget væsentligt resultat, fordi arten er vidt udbredt i danske vandløb, og fordi den udgør en nøglerolle i vandløbene som ituriver og som føde for fisk. Sammenholdt med dens gode laboratorieegenskaber gør det arten meget velegnet som testorganisme over for potentielt giftige stoffer, f.eks. i forbindelse med godkendelse af nye pesticider. I hvilken udstrækning, at arten giver yderligere information sammenlignet med standardorganismen Daphnia kan diskuteres. Overfor eksponeringer til esfenvalerat og azoxystrobin var G. pulex 3-8 gange mere følsom, mens toksiciteten af andre pesticider, f.eks. pirimicarb er langt større overfor dafnier. Hvis man ønsker at supplere standardundersøgelserne i forbindelse med risikovurderinger målrettet mod vandløb vil G. pulex dog være et oplagt valg. Variationen i følsomheden af de forskellige effektparametre var meget stor. Hos Gammarus pulex eksponeret til esfenvalerat i én time øgedes driftraten med ca. 60% ved en koncentration på 0,2 ng l-1, mens reproduktionen og overlevelsen af juvenile blev reduceret med 90 - 95% ved 50 ng l-1 der dog var den laveste undersøgte koncentration (Fig. 56). Til sammenligning var fødebearbejdningen mindre følsom med ca. 50% reduktion ved 200 ng l-1. Overordnet må mortalitet opfattes som den mest konservative effektparameter ved ekstraponeringer til effekter i felten, mens drift på trods af en meget stor følsomhed kun kunne påvises overfor pyrethrioder. Endelig vil effekten af forøget drift hos Gammarus antagelig være begrænset, fordi dyrene vil kunne genindvandre fra refugierne (ikke påvirkede dele af vandløbssystemet) 5.4 Miljørelevans af undersøgelserMed få undtagelser er undersøgelserne udført på individer og populationer under laboratorieforhold, hvilket gør det vanskeligt at forudsige ændringer på samfundsniveau. På længere sigt kan selv små påvirkninger forskyde konkurrence- og prædationsforhold, og dermed ændre samfundets struktur. Således viser undersøgelser gennemført i store komplekse systemer (mesokosmos) eller i naturlige søer og vandløb, at effekten af pesticider typisk kommer til udtryk ved koncentrationer som er 10-flere gange lavere end de koncentrationer som forårsager mortalitet overfor “standardorganismer” (Møhlenberg et al. 2001). I dette projekt er der dog også undersøgt for ikke-lethale effekter, ligesom hovedparten af testene er gennemført med organismer, f.eks. Gammarus pulex, som er vist at være betydeligt mere følsomme end “standardorganismen” Daphnia i længerevarende test så som mesokosmosundersøgelser (Møhlenberg et al. 2001), og af de 11 bundlevende arter i denne undersøgelse var G. pulex konsekvent den mest følsomme. Det er derfor sandsynligt, at forskellen mellem effektgrænserne fundet i denne undersøgelse og de reelle effektgrænser i vandløbene er væsentlig lavere end en 10-faktor. I den anden retning trækker, at effektundersøgelserne i dette projekt er gennemført med pesticider tilsat i opløst form, hvor pesticider med høj partikelaffinitet så som esfenvalerat generelt vil forekomme bundet til overflader, f.eks. partikler i vandet, sediment eller på undervandsplanter. Tidligere undersøgelser har vist, at den opløste fraktion af pesticider med stor affinitet til partikler så som pyrethroider, er væsentlig mere toksisk end den partikel- eller sedimentbundne del (Clark et al. 1989, Chandler 1990, Chandler & Scott 1991, Green et al. 1996, Hill et al. 1988) uden at det dog er muligt at uddrage mere eksakte tal af disse undersøgelser. Baseret på nyere undersøgelser beregnede Boxall et al. (2001), at toksiciteten af pesticider reduceredes 3-1000 gange hvis der i forsøgene var sediment tilstede. Effekten var størst for pesticiderne med højest affinitet til partikler. I en række forsøg viste Schulz & Liess (2001a,b,c) at effektgrænsen efter 1 times pulseksponering af opløst fenvalerat til vårfluelarver var 10-100 gange lavere end hvis eksponeringen skete som sedimentbundet pesticid. Forskellen var størst for effektparametren dødelighed (faktor 100), og mindst for sublethale effekter som tidspunkt for emergens og biomasse. Der er således argumenter for at effektgrænserne beregnet på basis af projektets undersøgelser vil være højere end i naturlige vandløb – fordi der ikke er undersøgt på alle organismer og på samfundsniveau - men der er også argumenter for at de toksiske effekter er overvurderede pga. partikkelbinding i naturlige vandløb. Uden at under- eller overvurderinger nødvendigvis balancerer er det dog vores opfattelse, at effektgrænserne angivet i projektet er de bedst opnåelige (indenfor projektets rammer) og rimeligt realistiske. 6 LitteraturAbel PD & SM Garner. 1986. Comparisons of median survival times and median lethal exposure times for Gammarus pulex exposed to cadmium, permethrin and cyanide. Water Research 20(5): 579-582. Anton FA, Ariz M, & M Alia. 1993. Ecotoxic effects of four herbicides (Glyphosate, Alachlor, Chlortoluron and Isoproturon) on the Algae Chlorella pyrenoidosa Chick. Sci Total Environ (Suppl.) 845-851 Battaglin W & J Fairchild. 2002. Potential toxicity of pesticides measured in midwestern streams to aquatic organisms. Water Sci & Tech 45(9): 95-103. Baattrup-Pedersen, A. 2000. Planter i vandløb. DMU Temarapport 34, 38s. Blanck H & B Björksäter. 1998. Inhibition of growth of marine and freshwater microalgae by the sulfonylurea herbicides GLEAN and ALLY in the algal microtest battery. Final report to Nordisk Alkali Biokemi A/S. Blockwell S J, E J Taylor, et al. 1998. “The influence of fresh water pollutants and interaction with Asellus aquaticus (L.) on the feeding activity of Gammarus pulex (L.).” Archives of Environmental Contamination and Toxicology 34(1): 41-47. Boxall A, Brown C & K Barrett. 2001. Higher tier laboratory aquatic toxicity testing. Cranfield Centre for EcoChemistry research report No. JF 4317E for DETR, 70p. Breneman D.H & K W Pontasch. 1994. Stream microcosm toxicity tests - Predicting the effects of Fenvalerate on riffle insect communities. Environmental Toxicology and Chemistry 13(3): 381-387. Bækken T & KJ Aanes. 1991. Pesticides in Norwegian agriculture. Their effects on benthic fauna in lotic environments. Preliminary results Int. Assoc. Theor. Appl. Limnol. Proc./Int.Ver.Theor.Angew.Limnol.Verh. 24(4):2277-2281 Cambpell M J & D Machin 1990. Medical Statistics: a commonsense approach. Chichester, England, Wiley & Sons. Chandler GT. 1990. Effects of sediment-bound residues of the pyrethroid insecticide fenvalerate on survival and reproduction of meiobenthic copepods. Mar Environ Res 29: 65-76. Chandler GT & GI Scott 1991. Effects of sediment-bound endosulfan on survival, reproduction and larval settlement of meiobenthic polychaetes and copepods. Environ Toxicol Chem 10: 375-382. Clark JR, Goodman LR, Bothwick PW, Patrick JM Jr, Cripe GM, Moody PM, Moore JC & EM Lores. 1989. Toxicity of pyrethroids to marine invertebrates and fish: a literature review and test results with sediment-sorbed chemicals. Environ Toxicol Chem 8: 393-401. Clausen H 1998. Ændringer i bekæmpelsesmidlers egenskaber fra 1981-1985 frem til 1996. Faglig rapport fra DMU, nr. 223. Cradner JP & KD Hoagland. 1998. Combined effects of alachlor and atrazine on benthic algal communities in artificial streams. Envir. Toxicol. Chem. 17: 1415-1420. Dillaha TA, Reneau RB, Mostaghimi S & D Lee. 1989. Vegetative filter strips for agricultural nonpoint source pollution control. Trans of the ASAE 32: 513-519. DMU. 2001. Vandløb og kilder 2000. NOVA 2003. Bøgestrand J. (red.) Faglig rapport fra Danmarks Mljøundersøgelser nr. 378. 120 s. DMU. 2002. Vandløb 2001. NOVA 2003. Bøgestrand J (red). Faglig rapport fra DMU nr. 422. Danmarks Miljøundersøgelser. 40 s. EU 1998. Review report for the acitive substance azoxystrobin. 7581/VI/97-Rev.5. Fahl GM, Kreft L,. Altenburger R, Faust M, Boedeker W & LH Grimme. 1995. pH-dependent sorption, bioconcentration and algal toxicity of sulfonylurea herbicides Aquat.Toxicol. 31(2):175-187 Fairchild JF, La Point TW, Zajicek JL, Nelsen MK, Dwyer FJ, Lovely PA. 1992. Population-, community- and ecosystem-level responses of aquatic mesocosms to pulsed doses of a pyrethroid insecticide. Environ. Toxicol. Chem. 11, 115-129. Felding G, Mogensen BB, Sørensen JB & AC Hansen. 1997. Surface run-off of pesticides from farmland to streams and lakes. Bekæmpelsesmiddelforskning fra Miljøstyrelsen Nr. 29, 76 s. Flury M. 1996. Experimental evidence of transport of pesticides through field soils - A review. J Environ Qual 25: 25-45. Friberg N. 1998. Skove og skovvandløb. TEMA-rapport fra DMU 21/1998. Friberg N & D Jacobsen 1999. “Variation in growth of the detritivore-shredder Sericostoma personatum (Trichoptera).” Freshwater Biology 42(4): 625-635. Friberg N, AD Larsen, et al. 2002. “Shredder guilds in three Danish forest streams contrasting in forest type.” Archiv Fur Hydrobiologie 153(2): 197-215. Fyns Amt. 1999. Notat: Pesticidundersøgelser i vandløb, kildevæld og dræn 1994-1997. Notat, Fyns Amt. Natur- og vandmiljøafdelingen. Ganzelmeier H, Rautmann D, Spangenberg R, Streloke M, Herrmann M, Wenzelburger H-J. & H-F Walter. 1995. Studies on the spray drift of plant protection products. Mitteilungen aus der Biologischen Bundesanstalt für Land-und Forstwirtschaft. Heft 305. Berlin 1995. Green AS, Chandler GT & WW Piegorsch. 1996. Life-stage-specific toxicity of sediment-associated chlorpyrifos to a marine, infaunal copepod. Environ Toxicol Chem 15: 1182-1188. Gruessener B & MC Watzin. 1995. Response of aquatic communities from a Vermont stream to environmentally realistic atrazine exposure in laboratory microcosms. Envir. Toxicol. Chem. 15: 410-419. Heidmann T & AC Hansen. 1995. Precipitation and temperature in relation to runoff. I: Surface runoff, erosion and loss of phosphorus at two agricultural soils in Denmark – plot studies, 1989-92. SP-report No 14, June 1995, Vol 3. pp 47-76. Hill I, Hadfield ST, Kennedy JH & P Ekoniak. 1988. Assessment of the impact of PP321 on aquatic ecosystems using tenth-acre experimental ponds. Proceedings of the British Crop Protection Conference-Pests and Diseases. 1988. v. 1 p. 309. Huber A, Bach M & H-G Frede. 1998. Modeling pesticide loss with surface runoff in Germany. Sci Tot Envir: 177-191. IversenTM. 1980. Densities and energetics of two streamliving larval populations of Sericostoma personatum (Trichoptera). Holartic Ecology 3: 65-73. Iversen T M & J Jessen 1977. Life-cycle, drift and production of Gammarus-pulex-L (Amphipoda) in a Danish spring. Freshwater Biology 7(3): 287-296. Jensen K 2001. Esfenvalerat pulseffekter på makroinvertebrater i vandløb. Specialerapport Københavns Universitet. Johnson WW & MT Finley. 1980. Handbook of Acute Toxicity of Chemicals to Fish and Aquatic Invertebrates. Resour.Publ.137, Fish Wildl.Serv., U.S.D.I., Washington, D.C 98 p. Jurgensen TA & KD Hoagland. 1990. Effects of short-term pulses of atrazine on attached algal communities in a small stream. Arch Environ Contam Toxicol 19: 617-623. Kreutzweiser DP, Capell SC & BC Sousa. 1996. Hexazinone effects on stream periphyton and invertebrate communities. Envir. Toxicol. Chem. 14: 1521-1527 Kreutzweiser DP & P K Sibley. 1991. Invertebrate drift in a headwater stream treated with Permethrin. Archives of Environmental Contamination and Toxicology 20(3): 330-336. Krieger KA, Barker DB & JW Kramer. 1988. Effects of herbicides on stream Aufwuchs productivity and nutrient uptake. Arch. Environ. Contam. Toxicol. 17: 299-306 Kronvang B, Hansen B, Hald AB, Laubel AR & P Olsen. 2000. Jorderosion og bræmmer i landskabet. Vand & Jord 7(1): 20-27. Kusk KO. 1996. Bioavailability and effect of pirimicarb on Daphnia magna in a laboratory freshwater/sediment system. Arch Environ Contam and Toxicol 31: 252–255. Lamberti GA. 1996. The role of periphyton in benthic food webs, pp. 533-572 In: R.J. Stevenson, M.L. Bothwell, and R.L. Lowe (eds.). Algal Ecology: Freshwater Benthic Ecosystems. Academic Press, San Diego, CA. Lampert W, W Fleckner, et al. 1989. Herbicide effects on planktonic systems of different complexity. Hydrobiologia 188/189: 415-424. Lauridsen RB. 2002. Pesticideffekter på makroinvertebrater i vandløb - med særlig fokus på pyrethroidet lambda-cyhalothrin. Specialerapport Århus Universitet. Liess M. 1994. Pesticide impact on macroinvertebrate communities of running waters in agricultural ecosystems. Verh. Internat. Verein. Limnol. 25: 2060-2062. Liess M & R Schulz. 1999. Linking insecticide contamination and population response in an agricultural stream. Environ. Toxicol. Chem. 18: 1948-1955. Liess M, Schulz R, Liess MH-D, Rother B & R Kreuzig. 1999. Quantification of insecticide contamination in agricultural headwater streams. Water Res 33: 39-247. Magnin M, Marboutin E & N Pasteur. 1988. Insecticide resistance in Culex quinquefasciatus (Diptera: Culicidae) in West Africa. J Med Entomol. 25(2):99-104 Malbouisson JFC, TWK Young, et al. 1995. Use of feeding rate and re-pairing of precopulatory Gammarus pulex to assess toxicity of gamma-hexachlorocyclohexane (Lindane). Chemosphere 30(8): 1573-1583. Maltby L & C Naylor 1990. Preliminary observations on the ecological relevance of the Gammarus `Scope for Growth' assay: effect of zinc on reproduction. Funct. Ecol. 4: 393-397. McCahon C P & D Pascoe. 1988a. Culture techniques for three freshwater macroinvertebrate species and their use in toxicity tests. Chemosphere 17(12): 2471-2480. McCahon CP & D Pascoe. 1988b. “Use of Gammarus pulex (L.) in safety evaluation tests: culture and selection of a sensitive life stage.” Ecotoxicology and Environmental Safety 15: 245-252. Mickelson SK, Baker JL, Melvin SW, Fawcett RS, Tierney DP & CJ Peter. 1998. Effects of soil incorporation and setbacks on herbicide runoff from a tile-outlet terraced field. J Soil Water Conserv 531:18-25. Miljøstyrelsen. 1993. Bekæmpelsesmiddelstatistik 1992. Salg og hehandlingshyppighed 1992. Orientering fra Miljøstyrelsen. Miljøstyrelsen. 1998. Biologisk bedømmelse af vandløbskvalitet. Vejledning fra Miljøstyrelsen nr. 5 (1998). Miljøstyrelsen. 1999. Rapport fra underudvalget for Miljø og Sundhed Udvalget til vurdering af de samlede konsekvenser af en afvikling af pesticidanvendelsen (Bicheludvalget). Miljøstyrelsen. 2000. Bekæmpelsesmiddelstatistik 1999. Salg 1997, 1998 og 1999: Behandlingshyppighed 1999. Orientering fra Miljøstyrelsen, 11/2000. Miljøstyrelsen. 2001. Bekæmpelsesmiddelstatistik 2000. Salg 1998, 1999 og 2000: Behandlingshyppighed 2000. Orientering fra Miljøstyrelsen, 10/2001. Miljøstyrelsen. 2002. Bekæmpelsesmiddelstatistik 2002. Salg 1999, 2000 og 2001: Behandlingshyppighed 2001. Orientering fra Miljøstyrelsen, 5/2002. Müller K, Trolove M, James TK & A Rahman. 2002. Herbicide runoff studies in an arable soil under simulated rainfall. New Zealand Plant Prot 55: 172-176. Møhlenberg F, Friberg N, Gustavsson K, Lauritsen T & S Pedersen. 2001. Mesocosm experiments in the approval procedure for pesticides. Persticide Research No. 56. Møhlenberg F, Gustavson K & PB Sørensen. 2002. Pesticide Aquatic Risk Indicators -an examination of the OECD indicators REXTOX, ADSCOR and the Danish indicators FA and LI based on Danish sales data from 1992-2000, 73p. http://www.oecd.org/dataoecd/5/37/2752836.pdf Naylor C, L Maltby et al. 1989. Scope for growth in Gammarus pulex, a fresh-water benthic detritivore. Hydrobiologia 188: 517-523. OECD Guideline for testing of chemicals No. 202: Daphnia sp., Acute Immobilization Test. 1984.04.04. Pantani C, Pannunzio G, De Cristofaro M, Novelli AA & M Salvatori. 1997. Comparative acute toxicity of some pesticides, metals, and surfactants to Gammarus italicus Goedm. and Echinogammarus tibaldii Pink. Bull.Environ.Contam.Toxicol. 59(6):963-967 Pérès F, Florin D, Grollier T, Feurtet-Mazel A, Coste M, Ribeyre F, Ricard M and Boudou A. 1996. Effects of the phenylurea herbicide isoproturon on periphytic diatom communities in freshwater indoor microcosm. Environ Pollut 94: 141-152. Petersen S & K Gustavson. 1998. Toxic effects of tri-butyl-tin (TBT) on autotrophic pico-, nano- microplankton assessed by a size fractionated pollution-induced community tolerance (SF-PICT) concept. Aqua Toxicol 40:253-264. Probit analyse. Version 2.3, 1990.03.20. Statens Naturvårdsverk (Nat.ional Swedish Environmental Protection Board. The data section). Robinson JL, Brinsfield RB, Staver KW, Lucas WC & AH Todd. 1995. Water quality functions of riparian forest buffer systems in the Chesapeake Bay Watersheds. U.S. Environmental Protection Agency. Robinson CA, Ghaffarzadeh M & RM Cruse. 1996. Vegetative filter strip effects on sediment concentration in cropland runoff. J Soil Water Cons. 50: 227-230. Röpke B, Bach M, H-G Frede (i trykken). DRIPS - A DSS estimating the input quantity of pesticides for German river basins. I: Integrated Assessment and Decision Support 2002, Lugano, Switzerland: 150-155. Sand-Jensen K & N Friberg. 2000. (red.). De strømmende vande. Gads Forlag Schulz R & M Liess 1999. Validity and ecological relevance of an active in situ bioassay using Gammarus pulex and Limnephilus lunatus. Environ Toxicol and Chem 18(10): 2243-2250. Schulz R & M Liess 2001a. Runoff simulation with particle-bound fenvalerate in multispecies stream microcosms: Importance of biological interactions. Environ Toxicol and Chem 20(4): 757-762. Schulz R & M Liess. 2001b. Toxicity of aquarous.phase and suspended-particle-associated fenvalerate: chronic effects following pulse-dosed exposure of Limnephilus lunatus (Trichoptera). Environ Toxicol Chem 20: 185-191. Schulz R & M Liess. 2001c. Acute and chronic effects of particle-associated fenvalerate on stream macroinvertebrates: a run-off simulation study using outdoor microcosms. Arch Environ Contam Toxicol 40: 481-488. Shillabeer N, Smyth DV & L Tattersfield. 2000. Higher tier risk assessment of agrochemicals, incorporating sediment into algal test systems. Proc. of the BCPC Conference – Pests and Diseases, Brighton 2000, pp. 359-364. Spawn RL, Hoagland, KD & BD Siegfried. 1997. Effects of alachlor on an algal community from a midwestern agricultural stream. . Envir. Toxicol. Chem. 16: 785-793. Spliid NH & Mogensen BB. 1995. Udvaskning af pesticider fra Landbrugsjord. Bekæmpelsesmiddelforskning fra Miljøstyrelsen, nr. 11. 105 s. Styczen M, Wiberg-Larsen P & A Aagaard. 2003. Tag pulsen på pesticiderne i vandmiljøet. Vand & Jord 10: 84-87 Traunspurger W, Schafer H & A Remde. 1996. Comparative investigation on the effect of a herbicide on aquatic organisms in single species tests and aquatic microcosms. Chemosphere 33(6):1129-1141. USEPA. 1997. Pesticide fact sheet. Azoxystrobin. Villholth KG, Jarvis NJ, Jacobsen OH & H de Jonge. 2000. Field investigations and modeling of particle-facilitated transport in macroporous soil. Jour. Environ. Qual. 29: 1298-1309. Wauchope RD. 1978. The Pesticide Content of Surface Water Draining from Agricultural Fields – A Review. J. Environ. Qual. 7: 459-472. Welton JS & RT Clarke. 1980. Labratory studies on the reproduction and growth of the amphipod, Gammarus pulex (L.). Journal of Animal Ecology 49: 581-592. WHO 1995. Diflubenzuron Health and Safety Guide No. 99. Geneva 1995. Wiggers L. 1999. Pesticider i vandløb, kilder og søer i Århus Amt. Teknisk Rapport, Århus Amt, Natur og Miljø. Wogram J & M Liess. 2001. Rank ordering of macroinvertebrate species sensitivity to toxic compounds by comparison with that of Daphnia magna. Bulletin of Environmental Contamination and Toxicology 67(3): 360-367. Zelinka M & P Marvan. 1976. Notes to estimating production of zoobenthos. Folia Fac. Sci. Nat. Univ. Purkynianae Brunensis 17, Biol. 58(10): 1-53. Bilag A Bentiske mikroalger
Dosis-respons relation for bentiske mikroalger eksponeret til hexazinon i 1 time (øverst) og 24 timer (nederst).
Beregnede vækstrater af kiselalger (venstre) og grønalger (højre) efter eksponering til metribuzin (øverst) og isoproturon (nederst) i 2, 24 og 48 timer og efterfølgende ophold i rent åvand i 48 timer. Vækstraterne er beregnet som: ln(CR/C0)/t, hvor CR er algernes koncentration efter 48 timer i rent vand og C0 er koncentrationen efter eksponeringsperioden; t = recoveryperiodens længde (2 dage).
Effekt af pendimethalin på biomassen af kiselalger (øverst), grønalger (midt) og blågrønalger (nederst). Venstre kolonne: biomasse efter eksponering (1, 2 24 og 48 timer); højre kolonne: biomasse efter de eksponerede alger har været i rent vand i 48 timer. Forsøgene er udført om foråret i 2001.
Beregnede vækstrater af kiselalger (venstre) og grønalger (højre) efter eksponering til fenpropimorph i 24 og 48 timer og efterfølgende ophold i rent åvand i 48 timer. Vækstraterne er beregnet som: ln(CR/C0)/t, hvor CR er algernes koncentration efter 48 timer i rent vand og C0 er koncentrationen efter eksponeringsperioden; t = recoveryperiodens længde (2 dage). Bilag B Effekt af eksponeringsvarighedValg af testorganismerI henhold til projektoplægget var strategien at teste relevante pesticider på 8-10 forskellige arter/slægter, som repræsenterede forskellige hovedgrupper samt ernæringstyper. Det viste sig problematisk at gennemføre denne forsøgs strategi af en række årsager. De fleste af arterne var kun tilgængelige i en ganske kort periode (1-2 mdr.) og var vanskelige at arbejde med i laboratoriet da f.eks slørvinger og vårfluer nåede et flyvefærdigt stadie kort tid efter indsamlingen. Samtidig var det problematisk at indsamle et tilstrækkeligt antal testorganismer af de forskellige arter. I løbet af de første forsøgs serier der udført i 2001 blev det klart at eksponeringsscenariet ikke var velegnet til de mindre robuste vandløbs invertebrater som vårfluer og slørvinger. Længerevarende eksponerings forsøg med sådanne vandløbs invertebrater burde formentlig have inkluderet strømakvarier med bundsubstrat samt en lavere organisme tæthed. Et sådant testdesign var ikke praktisk muligt inden for projektets rammer. Det blev derfor valgt at fokusere på test med Gammarus pulex som kunne holdes i laboratoriet og som dertil viste sig at være den mest følsomme af de anvendte arter uanset om effekten var på mortalitet, drift eller reproduktion. Kvantificering af eksponeringskoncentrationerI forsøg med radioaktivt mærkede pesticider blev den aktuelle eksponeringskoncentration fulgt ved at udtage vandprøver før og efter hvert vandskift. Vandprøverne (10 ml) blev tilsat 10 ml scintillationsvæske (InstaGel II Plus, Packard, The Netherlands) og sat i køleksab mindst 24 timer før kvantificering af radioaktiviteten ved ”Liquid Scintillation Counting” (LSC). I forsøg med umærkede pesticider er LC50 udregnet på baggrund af nominelle koncentrationer. Kvantificering af interne effektdoserDen interne effektkoncentration (CBR) blev bestemt for de radioaktivt mærkede pesticider esfenvalerate, pendimethalin og ioxynil. Døde testorganismer blev opsamlet i vials og tilsat ca 1 ml vævsopløser (Soluene®-350) pr. vial (max 5 dyr pr. vial). Prøverne henstod i ca. 24 timer for at sikre total opløsning af vævet. Derefter blev der tilsat 15 ml scintillationsvæske (Ultima Gold, Packard, The Netherlands), prøverne blev grundigt rystet og blev stillet på køl i mindst 24 timer. Radioaktiviteten målt ved LSC. ResultaterI nedenstående tabeller angives for hvert af de undersøgte pesticider
Esfenvalerat Kumuleret dødelighed i %
LC10 og LC50 værdier
** Konfidensinterval kan ikke beregnes Azoxystrobin. Kumuleret dødelighed i %
* Dødelighed i kontrol overstiger de 10% der normalt accepteres i standardtest LC10 og LC50 værdier
Pendimethalin Kumuleret dødelighed i %
LC10 og LC50 værdier
** Konfidensinterval kan ikke beregnes Prochloraz Kumuleret dødelighed i %
LC10 og LC50 værdier
** Konfidensinterval kan ikke beregnes Dimethoat Kumuleret dødelighed i %
LC10 og LC50 værdier
** Konfidensinterval kan ikke beregnes Ioxynil Kumuleret dødelighed i %
LC10 og LC50 værdier
** Konfidensinterval kan ikke beregnes Bilag C LivcykluseffekterProcedure for pulseksponeringEsfenvalerat (kemisk navn = (S)-α-cyano-3-phenoxybenszl(S)-2-(4-chlorophenyl)-3-methylbutyrat, technical grade (HPLC) = 99,9%) blev indkøbt i pulverform fra Sigma-Aldrich, Danmark. Stamopløsninger blev fremstillet ved at opløse en kendt vægt af esfenvalerat i acetone (technical grade (GC) = 99,8). Koncentrationen af stamopløsningerne blev testet ved hjælp af GC-MS. Denne analysemetode blev anvendt, idet de laveste eksponeringskoncentrationer lå under den analytiske detektionsgrænse. I løbet af projektforløbet fik vi udviklet en analytisk metode ved hjælp af GC-ECD (Wiener, upubliceret) med em detektionsgrænse på 0,01 µl/l. For detaljeret beskrivelse af ekstraktionsprocedure henvises til bilag. Eksponeringskoncentrationerne blev opnået ved at tilføre en kendt volumen af stamopløsning til vandfasen ved hjælp af en mikrosprøjte (SGE – Syringe Perfection) efterfulgt af en grundig omrøring. Den maksimale koncentration af opløsningsmidlet acetone var 300µl/l. Mulige effekter af solventet blev testet ved anvendelse af en actetone-kontrolgruppe. Alle koncentrationer er angivet som nominelle værdier. Anvendt statistikOverlevelseskurverne for forsøgene med Gammarus blev analyseret ved hjælp af Kaplan-Meier estimator i SYSTAT (SYSTAT ver. 10, SPSS Inc. Chicago). Denne nonparametriske estimator tillader stratifikation (f.eks. af eksponeringsgrupper, køn eller livsstadier) og kan håndtere ”censurerede” data (f.eks. individer der stadig er i live ved slutningen af forsøget). Overlevelseskurver blev sammenlignet ved hjælp af en Tarone-Ware log-rank test der vægter hver død ved kvadratroden af det totale antal individer udsat for risiko. Der lægges således mindre vægt på de dødsfald der forekommer senere i eksperimentet. Median overlevelsestid blev estimeret udfra den beregnede ”quantiles”. I forsøg 2 og 3 blev der anvendt en nonparametrisk Kruskal-Wallis test for at sammenligne median antal overlevende individer testgrupperne imellem til forskellige tider efter pulseksponering. Kaplan-Meier teknikken blev brugt til at analysere paringsadfærden hos voksne efter pulseksponering (eksperiment 5). Den første dag hvor et par blev observeret gendannet blev noteret for hvert par, og den kumulative andel af gendannede par blev beregnet for hver eksponeringsgruppe. Det reproduktive output efter eksponering blev analyseret ved brug af Kruskal-Wallis nonparametriske ANOVA-test (SYSTAT ver. 10), idet data ikke var normalfordelt. Parvis sammenligning blev udført ved hjælp af en nonparametrisk version af Turkey-test (Nemenyi Test, Zar 1996). Effekten af pulseksponering til esfenvalerat og køn på tid til klækning af C. riparius blev i Forsøg 1 analyseret ved hjælp af en 2-vejs ANOVA. I forsøg 2 blev pulseksponering til esfenvalerat og populationstæthedens effekt på tid til klækning af C. riparius analyseret ved hjælp af en 2-vejs ANOVA, henholdsvis for hanner og hunner. Variationer i tid til klækning kønnene imellem blev i forsøg 2 analyseret ved hjælp af en 1-vejs ANOVA, henholdsvis for hver af de repræsenterede eksponeringsgrupper og tæthedsgrupper. Effekten af esfenvalerateksponering (Forsøg 1 og 2) og individtæthed (Forsøg 2) på den procentvise andel af larver med klækningssucces blev testet med en ”contingency table analyse” ved hjælp af Pearson's Chi-square test statistik. I Forsøg 1 blev alle færdigudviklede individer tilhørende den samme forsøgsgruppe overført til det samme sværmebur ufafhængigt af replikat. Dette var nødvendigt for at sikre et tilstrækkeligt stort antal individer af hvert køn pr. sværmebur, så der kunne foregå reproduktion. I bestræbelsen på at have voksne individer nok til formering i forsøg 2 blev klækkede voksne individer fra de 10 originale relikater pr gruppe delt i to lige grupper, således at replikat 1-5 udgjorde en gruppe og replikat 6-10 en anden. Herefter blev eksponering med esfenvalerat og populationstæthedens effekter på ægproduktionen samt klækningssuccesen af unger undersøgt ved hjælp af en 2-vejs ANOVA med n=2. Alle statistiske analyser blev udført ved hjælp af SYSTAT ver. 10 (SPSS, Chicago, IL). Bilag D Drift i recirkulerende vandløbsrenderKlik her for at se Figur til Bilag D. Bilag E Indirekte effekterKlik her for at se Figur til Bilag E. Test af forskel fra kontrol under forsøg med dimethoat samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset.
Nedbrydningen af detritus efter syv dages eksponering af elleblade i azoxystrobin er afbilledet som bladpladernes vægttab per mg dyr mod eksponeringskoncentration. Søjlediagrammerne angiver gennemsnittet af de 15 replikater med standard error. * indikerer at bladvægttabet er signifikant forskelligt fra kontrollerne (F-test, p<0,05) Test af forskel fra kontrol under forsøg med azoxystrobin samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset.
Bilag F Kultivering af Prodiamesa olivaceaDen 13/11-02 blev ca. 800-1000 individer af Prodiamesa olivacea indsamlet fra Gadevang Renden, umiddelbart efter udløbet fra Gadevang Rensningsanlæg. Renden er ca. 1-2 meter bred, 5-400 cm dyb og har relativt langsomt rindende vand. Bunden udgøres primært af sand og løst blødt sediment bestående af henrådnende plantedele og blade fra den omgivende vegetation, der består af høje bøgetræer. Renden løber således udelukkende gennem skov, hvorfor der må formodes at være en ringe lyspåvirkning i vandløbet ved løvspring. Ved indsamlingslokaliteten blev vandtemperaturen målt til 8°C mens lufttemperaturen blot blev målt til 2°C. Den relativt høje vandtemperatur i forhold til lufttemperatur forventes at være forårsaget af lokalitetens nærhed til rensningsanlægget. IndsamlingsprocedureIndsamlingen foregik ved hjælp af en håndketcher (køkkensigte) med en maskevidde på 1 mm. Denne blev ført gennem det øverste sedimentlag (5 cm dybde), hvorefter dette blev sigtet i vandoverfladen. De nu tydeligt synlige myggelarver blev herefter nænsomt pillet fra ved hjælp af en blød pincet og overført til små plastbeholdere indeholdende vand fra indsamlingslokaliteten. Akklimatisering og kultiveringVed hjemkomsten blev alle individer anbragt i klimarum ved 12°C og fodret med en opslemmening af fiskefoder (TetraMin) opløst i hanevand. Dagen efter indsamlingen blev individerne overført til sværmebure med mellem 200-300 individer i hvert. Sværmeburene, der havde dimensionerne 31(H)×17(B)×42(L) cm var på forhånd forsynet med et ca. 1,5 cm tykt lag sediment (kunstigt fremstillet efter OECD's guideline) og tilført afboblet hanevand til en dybde på ca. 8-10 cm. Vandsøjlen blev tilført luft via en almindelig akvariepumpe monteret med luftslanger og tilsat to pimpsten. Efter ca. en uge ved 12°C blev sværmeburene akklimatiseret til et rum med stuetemperatur (20°C ± 1). Larverne blev fodret en gang om ugen, idet kulturakvarierne blev tilsat et par dråber af omtalte foderblanding (konc.= 1g fiskefoder/20ml hanevand). Halvdelen af vandet i kulturburene blev skiftet hver 14 dag. De første sværmende myg blev observeret den 25/11-02 og den første ægmasse (lange strimler) blev observeret den 19/12-02. Der kom dog først rigtig gang i klækningen den 19/12-02, da sværmeburene blev tilsat en direkte lyskilde (800-1000 lux) med 12 timers lys og 12 timers mørke, hvilket resulterede i en øjeblikkelig forøgelse i antallet af sværmende individer. Trods det stigende antal klæggede individer udeblev reproduktionen, idet kun ganske få ægmasser blev observeret. De enkelte ægmasser blev straks overført til petriskåle og den eksakte læggedato noteret. Desværre gav ingen af de i alt 6 observerede ægmasser levende afkom, hvorfor de resterende larver blev anvendt til LC50-forsøg.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||