Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 103, 2006
Fungiciders påvirkning af mykorrhizasvampes diversitet og funktion
Indholdsfortegnelse
2 Forsøgsmark og generelle metoder
- 2.1 Forsøgsareal og -jord
- 2.2 Fungicider
- 2.3 Forsøgsplanter og mykorrhizasvampe
- 2.4 Molekylære metoder
- 2.5 Kvantificering af mykorrhizasvampes vækst og fosfatoptagelse
- 2.6 Statistisk analyse
3 Diversitet og genotype fordeling af mykorrhizasvampe i økologisk og konventionelt dyrkede marker
4 Fungiciders indflydelse på mykorrhiza i parvise kombinationer mellem planter og mykorrhiza-svampe
- 4.1 Dosis-responsmålinger med forskellige arter af mykorrhizasvampe
- 4.2 Effekt af fungicider på forskellige genotyper (isolater) af samme mykorrhizasvampeart
5 Fungiciders indflydelse på funktion af mykorrhiza baseret på rekonstruerede svampe-samfund
6 Fungiciders indflydelse på forsøgsmarkernes naturlige mykorrhizasvampe-samfund
Forord
Denne rapport er resultatet af et samarbejde mellem Afdeling for Biosystemer, Forskningscenter Risø og Biologisk Institut, Københavns Universitet i perioden 2002-2005. Arbejdet er udført af Iver Jakobsen, Lisa Munkvold Murmann, Anette Olsen og Anne Olsen fra RISØ samt Søren Rosendahl, Randi Gregersen og Eva H. Stukenbrock fra KU. Lise Nistrup Jørgensen, Danmarks JordbrugsForskning, deltog som konsulent i forbindelse med udvælgelsen af fungicider til projektet.
Projektet bygger på resultaterne af projektet ’Pesticiders indflydelse på nedbrydnings- og opbygningsprocesser i rodzonen’, som blev gennemført i 1996-1999 under Den Tværministerielle Pesticidforskningsindsats. Projektet har derfor udnyttet det samme forsøgsområde som det foregående, et økologisk og et konventionelt dyrket sædskifte ved KVL’s forsøgsgård i Tåstrup.
Der har været nedsat en følgegruppe bestående af:
Bent K. Christensen, Dansk Juletræsdyrkerforening
Claus Rasmussen, Skov- og Naturstyrelsen
Hans M. Hedegaard, Dansk Skovforening
Michael Stoltze, Dansk Naturfredningsforening
Claus Hansen, Miljøstyrelsen
Carsten Suhr Jacobsen, GEUSWiborg, Dansk Landbrugsrådgivning
Sammenfatning
Denne undersøgelse anlægger en bred synsvinkel på fungiciders indflydelse på mykorrhizasvampe. Tidligere studier har især sammenlignet forskellige fungiciders effekt på enkelte svampeisolater og har kun i begrænset omfang inkluderet dosis-respons undersøgelser. Eksperimenterne beskrevet i denne rapport har undersøgt fungicideffekter på mykorrhizasvampe på individ-, populations- og samfundsniveau. Denne integrerede tilgang giver et mere realistisk billede af de mulige konsekvenser af landbrugets anvendelse af fungi-cider på betydningen af mykorrhiza som plantevækstfaktor.
På individniveau bekræftede undersøgelsen, at fungicider kan hæmme mykorrhizasvampene. NOAEL-grænseværdier (no observed adverse effect level) var langt lavere for carbendazim (0,1 – 0,5 x markdosis) end for mancozeb (5 – 25 x markdosis). Tilførsel af små doser (< NOAEL) af begge fungicider havde derimod en generelt stimulerende effekt på mykorrhizasvampene. Den fysiologiske baggrund for stimuleringen blev ikke klarlagt, men involverer formodentlig overkompensation i svampens reaktion på en svag stress påført af fungiciderne. Forsøgene viste også, at der kan være stor forskel på, hvordan forskellige arter og selv forskellige genotyper indenfor samme art reagerer på fungicider.
På populationsniveau viste undersøgelsen, at der er et betydeligt antal genotyper af forskellige arter af mykorrhizasvampe til stede i marken. Den genotypiske variation og fordeling indenfor de enkelte marker kunne forklare det meste af variationen, når nabomarker med økologisk eller konventionelt dyrkningssystem blev sammenlignet. Forskellene mellem konventionel og økologisk dyrkning gav altså ikke anledning til en ændret genetisk diversitet indenfor arterne. Derfor skal det ikke forventes, at eventuelle effekter af forskelle i jordens indhold af pesticidrester slår igennem på populationsniveau.
Dyrkningshistorien (konventionel eller økologisk dyrkning) har derimod en væsentlig effekt på diversiteten på samfundsniveau, selvom diversiteten også varierer inden for marken. Disse mark- og stedbetingede forskelle var tydeligt større end små fungicid-inducerede effekter. Det blev dog også vist, at fungicidbehandling kunne ændre udfaldet af interaktionerne i de mulige kombinationer af tre svampe, og at dette udfald var svært at forudsige ud fra viden om de enkelte svampe. Fungicidbehandling kunne øge fosfatoptagelsen ved at hæmme en ineffektiv svamp i kombinationerne, men dette betyder ikke at fungicid behandling altid vil give denne effekt.
Det konkluderes, at den høje kompleksitet af svampesamfund i markjord bidrager til, at fungicidanvendelsen i dansk landbrug har en ringe effekt på samfundet af mykorrhizasvampe i jord og planterødder. Markdosis af mancozeb og små mængder af det i øvrigt stærkt hæmmende carbendazim viste sig at kunne stimulere udvikling og funktion af mykorrhiza. Denne hormesis-lignende effekt kan være en del af forklaringen på, at der kun sjældent er blevet observeret effekter af fungicidbehandling på forekomsten af mykorrhiza under markforhold. En høj kompleksitet af svampesamfund, og stor genetisk variation på populationsniveau, vil modvirke effekter af eksterne faktorer, som fx. fungicider, på samfundets funktionelle egenskaber.
Summary
This study describes studies on fungicide effects on arbuscular mycorrrhizal fungi. Previous studies have compared various fungicides effects on specific fungal isolates, and only few studies have included dose-response experiments. The experiments described in this report include fungicide effects on individual, population and community level. This integrative approach should provide a more realistic demonstration of the impact of agricultural fungicides on the importance of mycorrhiza as a plant growth promoting factor.
At the individual level fungicides were found to inhibit mycorrhizal fungi. The NOAEL-limit (no observed adverse effect level) was lower for carbendazim (0,1 – 0,5 x field dose) than for mancozeb (5-25 x field dose). Low doses (< NOAEL) of both fungicides had a stimulating effect on the fungi. The physiological background for this stimulation was not elucidated, but an over compensation in the reaction of the fungi to low stress levels could be involved. The experiments showed that different species and genotype may react differently to the fungicides.
Studies at the population level revealed a significant number of genotypes in the field. The genotypic variation and distribution within the fields explained most of the variation. Comparison of adjacent fields grown conventionally or organic, showed that most of the variation was attributable to variation within the field. Differences between conventionally and organic praxis did not influence the genetic diversity within the species. Consequently, fungicides can not be expected to influence the fungi at the population level.
A more pronounced effect of the soil management (conventionally or organic) was seen on the diversity at the community level, though some variation in diversity was seen within the fields. These field and site differences were more pronounced that the fungicide induced effects. It was also shown that the fungicide treatment could change the result of the interaction in the possible combination of the three fungi. The outcome of the interactions could not be concluded from the knowledge obtained from experiments with the fungi alone. The fungicide treatment could increase the phosphorus uptake by inhibition of an inefficient fungus in some of the combinations.
In conclusion the complexity in the fungal communities in agricultural fields, can explain why the use of fungicides in Danish agriculture have limited effects on the community of mycorrhizal fungi in soil and plant roots. Field doses of mancozeb and low doses of carbendazim could stimulate the development and the function of mycorrhiza. This hormesis effect can partly explain why fungicide effects on mycorrhiza under field conditions are rarely observed. The pronounced complexity in communities and the genetic variation at the population level will conceal effects of external factor as fungicides on the functional properties of the communities.
1 Indledning
Stort set alle planter lever i symbiose med svampe, som bidrager til planternes næringsstofoptagelse. Symbiosen kaldes mykorrhiza (svamperod) og arbuskulær mykorrhiza er den fremherskende type hos agerlandets kulturplanter. Mykorrhiza-svampenes mycelium vokser delvis i planteroden, delvis i den omgivende jord og grænsefladerne til plante og jord spiller en vigtig rolle for mykorrhizaens funktion. Svampen optager uorganiske næringsstoffer fra jorden, og en del af disse ender i planten, som til gengæld udgør svampens eneste forsyningskilde for kulstofforbindelser i form af simple sukkermolekyler. Mykorrhizasvampene spiller en vigtig rolle i landbrugsplanternes fosfor- (P) optagelse (Jakobsen & Rosendahl, 1993; Smith et al., 2003;Thingstrup et al., 1998; Schweiger & Jakobsen, 1999; Smith et al., 2003), og svampene har en række andre effekter på planterne fx nedsat modtagelighed for rodsygdomme (Rosendahl, 1985; Bodker et al., 1998) og tørkestress (Michelsen & Rosendahl, 1990). Hvor tidligere studier har fokuseret på at beskrive mykorrhizas effekt ved at sammmenligne planter med og uden mykorrhiza, har de senere års forskning fokuseret mere på betydningen af diversiteten af mykorrhizasvampene. Opretholdelsen af en høj diversitet af mykorrhizasvampe viste sig således at have afgørende betydning for planters vækst og planteøkosystemers diversitet (van der Heijden et al., 1998).
I PEASE projektet under Den Tværministerielle Pesticidforskningsindsats blev der udviklet metoder til at undersøge pesticiders virkning på mykorrhizasvampe. Metoderne tillader måling af svampenes P transport til planterne (Schweiger & Jakobsen, 1998; Schweiger et al., 1999; Schweiger et al., 2001) og af svampenes diversitet i marken (Kjoller & Rosendahl, 2000a; Kjoller & Rosendahl, 2001). Metoderne har sat os i stand til at bestemme diversiteten af svampene samt at kvantificere svampenes bidrag til planternes ernæring. Herved er det blevet muligt at måle både direkte og indirekte effekter af forskellige miljøpåvirkninger.
En række undersøgelser har søgt at klarlægge pesticiders effekt på jordens mikroflora, men ofte er det vanskeligt at anvende denne viden til at vurdere miljømæssige effekter på et højere niveau. Dette hænger bl.a. sammen med at vi kun har et begrænset indblik i betydningen af den mikrobielle diversitet for økosystemers funktion.
Pesticiders mulige indflydelse på sammensætningen af jordens mikrobielle samfund er målbar ved anvendelse af molekylære teknikker til at karakterisere de mikrobielle komponenter i samfundene. Ønsker man derudover at kvantificere den funktionelle betydning af sådanne ændringer i mikrobielle samfund, vil det ofte være forbundet med store metodiske vanskeligheder. Det gælder især, når funktionen af en specifik organisme er ukendt, f.eks. i forbindelse med nedbrydning af organisk stof. Derfor er det vanskeligt at vurdere konsekvensen af at eliminere en bestemt art fra det mikrobielle nedbrydersamfund.
Mykorrhizasvampes tætte samspil med planter gør det derimod muligt at inddrage funktionelle aspekter i effektstudier på denne gruppe af mikroorgansimer. Mykorrhizasvampene repræsenterer en unik gruppe af jordsvampe og udgør en væsentlig andel af jordens mikrobielle biomasse. Svampenes betydning for planters fosforoptagelse og vækst er velkendt, og svampenes funktionelle diversitet i denne fosforoptagelse er målbar. Det betyder, at pesticiders indflydelse kan måles, både som ændringer i disse svampesamfunds sammensætning og som ændringer i deres kvantitative bidrag til planters vækst. Mykorhizasvampene er derfor velegnet som en model for en fremtidig kvantificering af pesticideffekter på andre svampegrupper i de tilfælde, hvor der kan knyttes en målbar funktion til svampenes tilstedeværelse i jord-plante systemet.
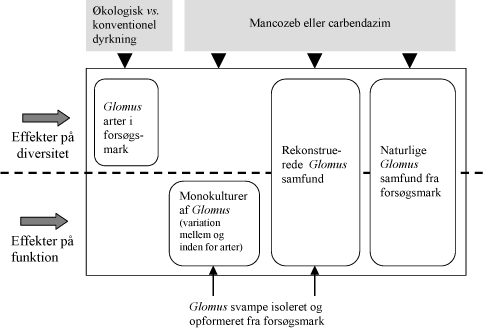
Figur 1.1. Skematisk fremstilling af de gennemførte undersøgelser i projektet.
Projektets indhold er illustreret i Figur 1.1. Der er gennemført fire hovedeksperimenter. Det første sammenligner konventionel og økologisk dyrkning med hensyn til effekt på diversiteten af populationer af mykorrhizasvampe i de to marker (Kapitel 3). Et andet hovedforsøg undersøger effekten af fungicider på mykorrhizaudvikling og funktion i symbioser mellem en planteart og monokulturer af mykorrhizasvampe isoleret fra forsøgsmarken. Et deleksperiment undersøger effekterne på artsniveau (Kap. 4.1), medens et andet undersøger effekterne på svampe tilhørende den samme art (Kap. 4.2). Effekter af fungicider på diversitet og funktion af rekonstruerede samfund af mykorrhizasvampe isoleret fra forsøgsmarken (Kap. 5) undersøges i et tredje hovedforsøg, og effekten på markjordens naturlige samfund af mykorrhizasvampe undersøges i fjerde hovedforsøg (Kap. 6).
1.1 Formål
Projektet havde som formål at undersøge fungiciders indflydelse på sammensætningen af samfund af mykorrhizasvampe i planterødder og på den afledte indflydelse på mykorrhizas betydning for planters næringsstofoptagelse og vækst. Udvikling af nye og optimering af eksisterende metoder fremstår som centrale virkemidler i projektet.
Projektarbejdet blev planlagt for at kunne teste følgende hovedhypoteser:
- Fungicidbehandling påvirker mykorrhizasvampes vækst og fosforoptagelse til værtplanten.
- Fungicidbehandling ændrer sammensætningen af mykorrhizasvampe-samfundet.
- Fungicidinducerede ændringer i svampesamfundets diversitet resulterer i et ændret fosforbidrag fra svamp til plante.
2 Forsøgsmark og generelle metoder
- 2.1 Forsøgsareal og -jord
- 2.2 Fungicider
- 2.3 Forsøgsplanter og mykorrhizasvampe
- 2.4 Molekylære metoder
- 2.5 Kvantificering af mykorrhizasvampes vækst og fosfatoptagelse
- 2.6 Statistisk analyse
2.1 Forsøgsareal og –jord
Projektet gennemføres med jord og svampe fra to marker ved Snubbekorsgård i Tåstrup. Markerne indgår i henholdsvis et økologisk (mark 29) og et konventionelt (mark 21) sædskifte (Figur 2.1). Pesticider har ikke været anvendt i det økologiske sædskifte i de seneste 15 år. De to marker blev dyrket med vinterhvede i 2002 og med markært i 2003. I hver af markerne blev der udlagt to transekter, og forsøgsjord og mykorrhizasvampe er indsamlet langs disse transekter.
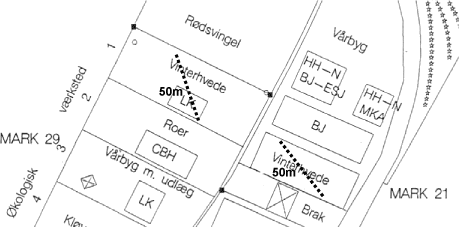
Figur 2.1. Svampe og jord til projektet er indsamlet langs to 50m transekter i økologiske (mark 29) og konventionelt (mark 21) dyrkede sædskifter ved Snubbekorsgaard, Tåstrup.
Jorden er en sandblandet lerjord (grovsand 23.7 %, finsand 26.2 %, silt 31.8 %, ler 16 %, organisk stof 2.3 %.) med pH 7.1 og et fosfortal på 2.7 og 4.0 i henholdsvis det økologiske og det konventionelle sædskifte.
2.2 Fungicider
Mancozeb er udvalgt som det centrale testfungicid i dette projekt, fordi mancozeb-forbruget tegnede sig for lidt over halvdelen af det samlede fungicidforbrug både i 2000 og 2004 (Anon., 2001; Anon., 2005). Mancozeb anvendes overvejende til kartofler, men også til bekæmpelse af ærtesyge. Mancozeb, det aktive stof i handelsvaren (fx. Dithane NT), er et polymer dithiocarbamate kontaktfungicid, som danner komplekser med metal-holdige enzymer og dermed hæmmer aktiviteten af bla. enzymer, der er involveret i produktion af adenosin triphosphate (ATP). Adsorptions koefficienten er høj (Koc ~ 6000) og mobiliteten i jordmatrix er derfor lav. Persistensen i jord er forholdsvis lav (T½ = 1-7 dage), men nedbrydningsproduktet ethylenethiourea (ETU) er mere persistent (T½ = 3-4 uger) (Jacobsen & Bossi, 1997). Halveringstider for mancozeb angives dog som 7-139 d af Christensen og Helweg (2003) med henvisning til The ARS Pesticide Properties Database, USDA.
Carbendazim har en kendt negativ virkning på mykorrhizasvampene (Kjoller & Rosendahl, 2000b; Schweiger & Jakobsen, 1998) og er derfor medtaget som referencestof, selv om stoffet ikke længere anvendes i Danmark. Carbendazim er et systemisk benzimidazole fungicide, som hæmmer polymeriseringen af tubulinmolekyler. Carbendazim adsorberes stærkt i jord (Koc ~ 3000) og er meget persistent (T½ = 6-12 måneder).
Når fungiciderne blev indblandet i jorden, blev den markækvivalente dosis beregnet på baggrund af en antagelse om, at hele markdosis opblandes i det øverste 5 cm jordlag. Dette estimat vil i betydeligt omfang afhænge af fungicidets adsorptions- og persistensegenskaber, samt af den mængde fungicid der tilbageholdes af afgrøden efter sprøjtning. De anvendte handelsvarer var Dithane DG NT (750 g mancozeb kg-1) og Derosal fl. (516 g carbendazim L-1) hvor anbefalet markdosis er hhv. 2 kg ha-1 og 0,7 L ha-1.
Indledende blev det vurderet i hvilket omfang effekten af de to fungicider på mykorrhizadannelse og –funktion ville afhænge af tilførselsmetode. I et potteforsøg med ært blev markdosis af hvert fungicid tilført enten direkte til jordoverfladen eller ved opblanding i jorden. Overfladetilførsel af carbendazim reducerede rodkolonisering fra 50 % til 36 %, medens opblanding resulterede i en stærk hæmning af mykorrhizaudviklingen til 2 %, og disse negative effekter kunne også aflæses i planternes fosfatindhold (2,0; 1,6 og 1,0 mg P pr. plante). Derimod havde mancozeb ikke signifikant effekt på hverken rodkolonisering eller P optagelse. Resultaterne viser, at tilførselsmetode kan have stor betydning for fungicideffekter på mykorrhiza. Det blev besluttet at anvende opblandingsmetoden i projektets planlagte eksperimenter for at sikre tilstedeværelsen af en kendt mængde fungicid i det jordvolumen, hvorfra mykorrhizasvampenes P optagelse skulle måles vha. radioaktivt P. Dette ville sikre den størst mulige reproducerbarhed i eksperimenterne.
2.3 Forsøgsplanter og mykorrhizasvampe
Ært cv Solare blev anvendt i alle eksperimenter bortset fra sammenligningen af svampeisolater inden for samme art (kapitel 4.2), hvor modelplanten Medicago truncatula blev anvendt i stedet. Ært blev valgt som primær forsøgsplante, fordi en række af de metoder, der skulle anvendes i nærværende projekt, delvis var udviklet i PEASE-projektet under Den Tværvidenskabelige Pesticidforskningsindsats. Valget af M. truncatula er begrundet i afsnit 4.2. Mykorrhizasvampe, som er anvendt i dyrkningsforsøgene, blev overvejende isoleret fra det økologiske sædskifte, mark 29, ved Snubbekorsgaard. Svampene blev i alle tilfælde opformeret i potter med jordkløver dyrket i 2-3 måneder en P-fattig bestrålet jord-sand blanding. Mykorrhizainokulum består af lufttør dyrkningssubstrat fra disse pottekulturer, med indehold af koloniserede rodstykker og sporer. Alt inokulum anvendt i projektet gennemløb en kvalitetstest i form af et bioassay, hvor koloniseringen af jordkløverplanter, dyrket i en 9:1 jord-inokulum blanding, blev målt efter tre ugers vækst. Der blev ikke anvendt inokulum, som resulterede i < 30 % kolonisering af testplanterne.
2.4 Molekylære metoder
2.4.1 PCR detektion af arbuskulær mykorrhizasvampe i planterødder
Sammenlignende undersøgelser af arbuskulære mykorrhizasvampe vanskeliggøres af, at svampene ikke kan dyrkes på kunstige medier. Samtidig er det ikke umiddelbart muligt at adskille svampearter i rødder på baggrund af forskelle i deres udseende. Hvis man ønsker at bestemme hvilke svampe der er til stede i en given plante, er det derfor nødvendigt at benytte en molekylærbiologisk strategi.
Anvendeligheden af molekylærbiologisk baserede detektionsmetoder afhænger af kendskabet til de organismer, man ønsker at detektere. Jo bedre kendskab man har til organismernes diversitet, jo bedre er mulighederne for at finde karakterer, der er specifikke for den givne organisme. Hvis man anvender en specifik DNA sekvens som mål (target) for sin detektion, er det derfor nødvendigt at have adgang til en database med nukleotidsekvenser fra beslægtede organismer, således at man kan vurdere sekvensens specificitet. I mange tilfælde kan man anvende sekvenser fra GenBank, som er tilgængelig via internettet (http://www.ncbi.nlm.nih.gov/).
Ved hjælp af PCR metoden mangfoldiggør man et specifikt stykke DNA til millioner af kopier. Metoden kræver at man kender lidt af den sekvens, man er interesseret i for at kunne fastgøre nogle små stykker syntetisk fremstillet DNA (ca 20 bp lange), de såkaldte primere. Primerne binder sig til de komplementerende sekvenser på DNA-prøven og definerer dermed, hvor der bliver amplificeret. PCR reaktionsbufferen indeholder foruden primere og DNA-prøve, også nukleotider til opbygning af de nye kopier og en varmestabil DNA polymerase, der indbygger nukleotiderne på de rigtige steder. Nukleotidsekvensen i det mangfoldiggjorte DNA kan herefter bestemmes og sammenlignes med kendte organismers nukleotidsekvens.
2.4.2 T-RFLP
Terminal Restriction Fragment Length Polymorphism (T-RFLP) anvendes ofte til at undersøge diversiteten af mikroorganismer i en prøve. Et bestemt gen fra prøven bliver mangfoldiggjort vha. universelle primere. Sekvensforskelle mellem de forstærkede gener visualiseres ved at skære generne med et eller to enzymer, der genkender specifikke områder i generne, hvor der er kendte forskelle. Fragmenterne kan derefter adskilles elektroforetisk efter deres længde, fx på en sekvenator. I princippet vil hvert fragment repræsentere en genetisk enhed fx en art eller en type. Teknikken kan på den måde anvendes til at estimere antallet af arter i en given prøve.
Mens teknikken er god til at lave relative sammenligninger af diversiteten i forskellige prøver, kan det være svært at verificere hvilke organismer, der er tale om. Generelt er der to måde at verificere eller identificere organismerne. En metode er at fremstille et klon-bibliotek på baggrund af sekvensering af klonede PCR prøver. Længden af sekvenserne i klon-biblioteket kan derefter sammenlignes med de fundne fragmenter. Alternativt kan sammenligne de fundne fragmenter med kendte sekvenser fra en database. Denne metode anvendes på organismegrupper, hvor genomet i forvejen er velkendt.
2.5 Kvantificering af mykorrhizasvampes vækst og fosfatoptagelse
Mykorrhizasvampes kolonisering af rødder bestemmes ved mikroskopi efter en klarings- og farvningsprocedure. Observation af mykorrhizastatus i skæringspunkterne mellem et antal tilfældigt placerede liniestykker og rødderne i tælleskålen gør det muligt at kvantificere både den koloniserede og den totale rodlængde (Newman, 1966). Svampenes hyfelængde i jorden bestemmes efter et lignende princip, hvor hyferne fra en jordprøve opsamles på et membranfilter, og hvor okularet indeholder et kvadratnet i stedet for et liniestykke (Jakobsen et al., 1992).
Mykorrhizasvampes P optagelse kan måles direkte ved anvendelse af dyrkningssystemer, der indeholder et rodfrit jordvolumen, som er mærket med radioaktivt P, 33P. Den radioaktive jord i det såkaldte hyfekammer er adskilt fra jord med rødder vha. et fintmasket net, der let passeres af de tynde hyfer, men ikke af rødder, og svampenes P optagelse kan måles som indholdet af 33P i planten (Schweiger et al., 1999). Et plastbæger fyldt med jord og lukket med et 25µm nylonnet havde i tidligere eksperimenter vist sig at være velegnet som hyfekammer (Smith et al., 2003), og det blev undersøgt, om hyfekammerets udformning ville påvirke mykorrhizasvampenes P optagelse til værtplanten (Figur 2.2).
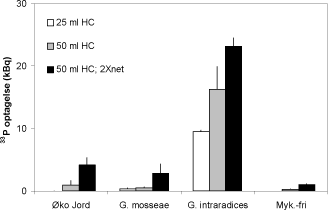
Figur 2.2. Mykorrhizasvampes optagelse af 33P fra jord placeret i nedgravet 25 ml eller 50 ml plastcylinder, hvor én åbning var forseglet med med nylonnet. Den sorte søjle angiver resultaterne med en 50 ml cylinder, hvor begge åbninger var forseglet med net. Ærteplanter blev dyrket i jord med det naturlige svampesamfund (øko-jord), i bestrålet jord tilsat Glomus mosseae eller G. intraradices eller i bestrålet jord uden tilsætning af mykorrhizasvampe. Middelværdier og standardfejl er angivet.
Glomus intraradices optog mest 33P, og optagelsen blev forøget både et større volumen af 33P mærket jord (50 ml HC) og et større areal for indvækst af mycelium (2 x net; Figur 2.2) Betydningen af det større areal for mycelieindvækst var også tydelig for de øvrige mykorrhiza-behandlinger og anvendelse af en 50 ml cylinder med net over begge åbninger vil derfor maksimere svampenes optagelse af 33P i testsystemet. Måling på forskellige tidspunkter bekræftede at 33P optagelse forøges med længden af vækstperioden.
2.6 Statistisk analyse
Styrkeberegninger blev gennemført for en række centrale variable og resultaterne viste, at anvendelse af 5 gentagelser af en given forsøgsbehandling gav en restvarians, som var tilstrækkelig lav til, at de forventede behandlingseffekter kunne måles. Begge hovedeksperimenter anvendte derfor 5 gentagelser (Kapitel 5 og 6). Behandlingseffekter blev analyseret vha. variansanalyse (S-Plus og SAS).
3 Diversitet og genotype fordeling af mykorrhizasvampe i økologisk og konventionelt dyrkede marker
3.1 Baggrund
De økologisk og konventionelt dyrkede sædskifter ved Snubbekorsgaard adskiller sig bl.a. i anvendelsen af sprøjtemidler siden 1987. En mulig påvirkning af sammensætningen af markens samfund af mykorrhizasvampe er undersøgt i afsnit 6, men det er endnu ikke afdækket, om dyrkningspraksis har en betydning for den genetiske diversitet inden for arterne. Variation er et særdeles relativt begreb, og variationen mellem marker med forskellig dyrkningspraksis bør derfor analyseres i forhold til den variation, der findes inden for markerne. Ved at sammenligne den genetiske diversitet på forskellige hierarkiske niveauer, er det muligt at afdække, hvad der bedst kan forklare variationen. Den hierarkiske indsamling blev foretaget ved at indsamle inden for markerne langs en transekt i begge marker. Langs transekter blev der indsamlet inden for 4 plots med 10m afstand.
3.2 Formål
Det var formålet at bestemme om den genetiske diversitet og den genetiske struktur inden for arter af mykorrhizasvampe er forskellig i økologisk og konventionelt dyrkede marker. Anvendelse af fungicider er én af de dyrkningsfaktorer, der adskiller de to dyrkningssystemer. Forskelle mellem marker blev sammenlignet med forskelle inden for marker.
3.3 Eksperimentelt
Der blev indsamlet sporer af tre arter af mykorrhizasvampe: Glomus mosseae, G. caledonium og G. geosporum i de to marker i Taastrup. Sporerne blev indsamlet i fire plots langs en 50 m transekt i begge marker. I hvert plot blev der indsamlet 4 prøver og der blev isoleret sporer fra jordprøverne. Fra den konventionelle mark blev der samlet 27 G. mosseae, 60 G. caledonium, og 13 G. geosporum sporer og fra den økologisk dyrkede blev der samlet 32 G. mosseae, 16 G. caledonium og 12 G. geosporum. Sporerne blev bestemt til genotype ved anvendelse af nested multiplex PCR, hvor der blev karakteriseret 3 uafhængige DNA markører fra hver spore (Stukenbrock & Rosendahl, 2005b). En genotype blev defineret som en unik kombination af de tre markører. Frekvensen af genotyper for de tre arter blev bestemt i de to marker. For at undersøge hvad der gav anledning til den opnåede genetiske variation, blev den genetiske variation opdelt i variation (i) inden for marken, (ii) mellem markerne, (iii) inden for plots og (iv) mellem plots. Såfremt dyrkningspraksis betyder noget for genotypefordelingen, vil det forventes at den største del af variationen stammer fra variation mellem marker (Gordon et al., 1992).
Dette kan beskrives ved at opfatte den totale variation (heterogenitet) betegnet Ht, hvor Ht sammensat af heterogenitet mellem marker og mellem plots samt inden for plots: Ht = Hs + Dst, hvor Ht er den totale heterogenitet, Hs er variationen inden for de enkelte marker og Dst er variationen mellem markerne. Udfra dette kan Gst = Dst/Ht bestemmes. Gst vil dermed beskrive, hvor meget af den totale heterogenitet, der kan forklares som variation mellem markerne; resten vil stamme fra variation inden for marker. Hs kan yderligere opdeles i variation inden for og mellem de enkelte plots: Hs = Hc + Dcs, hvor Hc er variationen inden for enkelte plots og Dcs er variationen mellem enkelte plots. Den del af variationen, der stammer fra variation mellem plots, kan bestemmes som Gcs = Dcs / Ht (Nei, 1973).
3.4 Resultater og diskussion
Genotypefordelingen viste at der fandtes op til syv genotyper af hver af de tre svampe (Fig 4.1). De hyppigste genotyper var lige hyppige i de to marker, og der var ingen forskel på antallet af genotyper i de to marker hos G.caledonium og G.geosporum. Hos G.mosseae var der færre genotyper i den konventionalt dyrkede mark sammenlignet med den økologisk dyrkede.
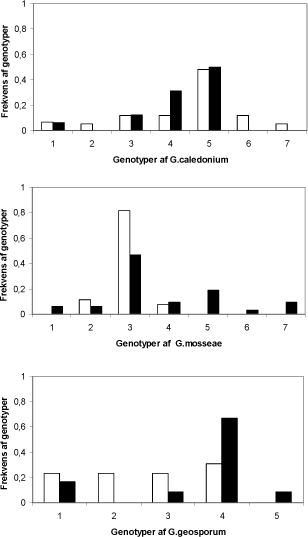
Figur. 3.1. Genotypefordeling af de tre arter af mykorrhizasvampe i konventionelt (hvide søjler) og økologisk dyrkede marker (sorte søjler).
Den totale heterogenitet Ht var af samme størrelseorden for alle tre svampe (Tabel 3.1). Gst som beskriver variationen mellem de to marker var lille og kunne kun forklare 2 - 7 % af variationen hos G.mosseae og G.caledonium. Den største del af variationen kom fra mellem plot (Gcs). Det vil sige at dyrkningspraksis, her eksemplificeret ved de to marker med konvetionel og økologisk dyrkning, havde ringe betydning for den genetiske diversitet af mykorrhizasvampe. Sammenligningen viste at variationen mellem de enkelte plots indenfor markerne bidrog meget mere til heterogeniteten end variation mellem de to marker.
Dyrkningspraksis betyder altså ikke så meget for den genetiske diversitet af mykorrhizasvampe. En vigtig undtagelse var dog G.mosseae, hvor det tyder på, at der var færre genotyper i den konventionelt dyrkede mark, sammenlignet med den økologisk dyrkede mark. Imidlertid er det vigtigt at undersøgelsen ikke havde til formål at undersøge forskelle på konventionel og økologisk dyrkning. En sådan undersøgelse ville kræve at flere par af konventionel og økologiske marker blev undersøgt. Variation mellem de enkelte plots bidrog meget mere til heterogeniteten end variation mellem de to marker. Den relativt høje ’mellem marker’ variation på 16 % hos G.geosporum kan skyldes at værdien er baseret på et sub-optimalt antal sporer indsamlet fra den økologiske mark.
Den observerede fordeling af heterogeniteten skal ses som et resultat af svampenes klonale vækst. Undersøgelse af de tre DNA markører viste, at de tre markører var koblede, hvilket tyder på fravær af rekombination hos svampene (Rosendahl & Taylor, 1997). Svampene vokser således i afgrænsede kloner, uden at der sker en opblanding af deres gener. Dyrkningspraksis påvirker altså ikke fordelingen af svampegenotyper nævneværdigt, men det kan ikke udelukkes, at konventionel dyrkning kan reducere den genetiske diversitet i marken ved at reducere antallet af genotyper (Stukenbrock & Rosendahl, 2005a). Det er uvist, om dette har betydning for jordens frugtbarhed (van der Heijden et al., 1998). Hvorvidt konventionel dyrkning også kan reducere artsantal og diversitet undersøges i kapitel 6.
Tabel 3.1. Den totale heterogenitet udtrykt som Ht, samt fordeling af variation mellem marker Gst og mellem plots Gcs for de tre undersøgte arter af mykorrhizasvampe. Det ses af det meste af variationen kan forklares som variation mellem plots i de enkelte marker.
| Ht | Gst | Gcs | |
| G. mosseae | 0.62 | 7% | 52% |
| G.caledonium | 0.75 | 2% | 49% |
| G.geosporum | 0.70 | 16% | 56% |
Det kan konkluderes, at genotyper af mykorrhizasvampe indsamlet fra blot én af markerne vil være repræsentative for begge marker. Dermed er det muligt at begrænse antallet af forsøgsfaktorer, og dermed forsøgets samlede volumen, i nogle af de efterfølgende dyrkningseksperimenter med indsamlet jord og indsamlede svampe.
4 Fungiciders indflydelse på mykorrhiza i parvise kombinationer mellem planter og mykorrhizasvampe
- 4.1 Dosis-responsmålinger med forskellige arter af mykorrhizasvampe
- 4.2 Effekt af fungicider på forskellige genotyper (isolater) af samme mykorrhizasvampeart
Tidligere undersøgelser har vist, at effekten af et givet fungicid kan variere mellem forskellige mykorrhizasvampe (Dodd & Jeffries, 1989); (Schreiner & Bethlenfalvay, 1997). Det er ikke undersøgt, hvorledes sådanne forskelle afhænger af fungiciddosis, og om forskelle i fungicidfølsomhed også kan erkendes inden for forskellige genotyper af den samme mykorrhizasvampeart. Disse spørgsmål er belyst i dette kapitel med anvendelse af svampe isoleret fra forsøgsmarken i Tåstrup.
4.1 Dosis-responsmålinger med forskellige arter af mykorrhizasvampe
4.1.1 Formål
Det var formålet at eksponere forskellige mykorrhizasvampe fra forsøgsmarken for forskellige doser mancozeb og carbendazim med henblik på at teste følgende hypoteser: (i) forskellige mykorrhizasvampe reagerer forskelligt på en given fungicidbehandling, (ii) der er forskel på skadevirkningen af carbendazim og mancozeb og (iii) fungicideffekter på mykorrhizasvampe er ikke målbare ved doseringer lavere end No Observed Adverse Effect Level (NOAEL). De opnåede resultater skulle i øvrigt anvendes til at fastlægge fungicid-dosering i de øvrige projektaktiviteter.
4.1.2 Eksperimentelt
Eksperimentet blev gennemført som et potteforsøg med Solare-ært og fire mykorrhizasvampe fra forsøgsmarkens økologiske sædskifte: Glomus claroideum (Schenck & Smith) (BEG88), G. mosseae (Nicol. & Gerd.) Gerdemann & Trappe (BEG84), G. caledonium (Nicol. & Gerd.) Trappe & Gerd (H07-1) og G. geosporum (Nicol. & Gerd.) Walker (BEG90). BEG-numre refererer til svampens nummer i den europæiske kultursamling af mykorrhizasvampe, International Bank for the Glomeromycota. Svampene blev opformeret i potter med jordkløver (Trifolium subterraneum L.) og jord fra disse kulturer blev anvendt som inokulum.
Ærteplanter blev dyrket i potter med 1000g bestrålet jord (10 MeV elektronstråling, 15 kGy) som blev tilført alle nødvendige plantenæringsstoffer, bortset fra fosfat (Pearson & Jakobsen, 1993). Bestrålingen var nødvendig for eliminering af de naturligt forekommende mykorrhizasvampe og for dermed at kunne gennemføre kontrollerede studier af fungiciders indflydelse på specifikke svampe. Nogle mikroorganismer overlever bestrålingen, og andre vil kolonisere potterne i løbet af vækstperioden. Jorden er derfor ikke steril. Jorden til de enkelte potter blev blandet med 50 g inokulum af en af de fire mykorrhizasvampe og andre potter modtog ikke inokulum. Hver af de fem mykorrhizabehandlinger blev kombineret med syv fungicidbehandlinger: ubehandlet, mancozeb (1, 5 eller 25 x markdosis) og carbendazim (0,01; 0,1 eller 1 x markdosis). Den valgte dosering af mancozeb var baseret på indledende undersøgelser, der tydede på, at mykorrhiza ikke blev påvirket af mancozebdoser lavere end markdosis. Tidligere studier har derimod vist, at carbendazim-doser lavere end markdosis har en skadelig effekt på mykorrhizasvampe (Schweiger & Jakobsen, 1998). Doseringerne blev opnået ved iblanding af 3.3, 16.7 og 83,3 µg mancozeb g-1 jord samt 0,006; 0,060 og 0,602 µg carbendazim g-1 jord. Der var fire gentagelser for hver behandling. Det beskedne antal doserings-niveauer tillod ikke en optimal analyse af dosis-respons forløb fx ved hjælp af Gompertz regressionsmodellen. Resultaterne er derfor analyseret ved hjælp af variansanalyse.
Jorden i det rodfri hyfekammer blev blandet med 5 kBq carrier-fri 33PO4 g-1 og kammeret blev placeret midt i potten. Der var 3 gentagelser for hver behandling, og de 95 potter blev placeret i vækstkammer med 16 t lys ved 21°C og 8 t mørke ved 16°C. Forsøget blev høstet 39 dage efter udplantning af spirede ærtefrø, og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, længde af hyfer i 33P-mærket jord samt planternes indhold af 33P og totalt P. Kolonisering af rødder med G. geosporum var 7 % eller mindre og dermed for lav til at en troværdig måling af fungicideffekter kunne opnås. G. geosporum data er derfor udeladt i det følgende.
4.1.3 Resultater og diskussion
Uden fungicidtilførsel havde 28 - 42 % af rodlængden udviklet mykorrhiza, og de to laveste doser af begge fungicider forøgede markant koloniseringen hos alle tre svampe (P < 0,001), medens den højeste dosis af begge fungicider førte til en reduceret mykorrhizadannelse i forhold til ubehandlet (Figur 4.1).
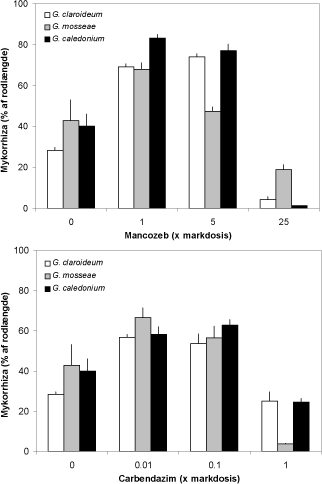
Figur 4.1. Effekt af forskellige doser af mancozeb og carbendazim på rodkolonisering hos ært dyrket med tre forskellige mykorrhizasvampe. Middelværdier og standardfejl er angivet.
Dosis-respons kurven havde således et omvendt U- eller J-forløb hos alle tre svampe, og kurvens form afhang af både svamp og fungicid. Mancozeb førte generelt til den kraftigste stimulering, og effekten af mancozeb på G. mosseae var anderledes end effekten på de to øvrige svampe. Dosering med 5x mancozeb førte til en reduceret G. mosseae-kolonisering, men ved den højeste dosering blev de to andre svampe hæmmet mest. I modsætning hertil, havde den højeste carbendazim-dosis en kraftigere negativ effekt på G. mosseae end på de to øvrige svampe.
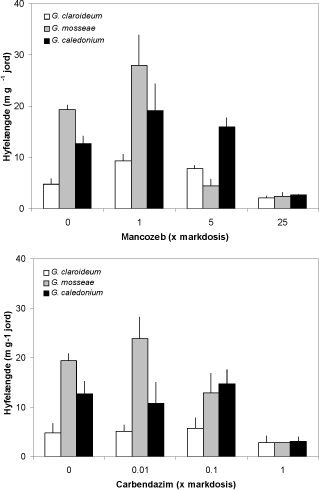
Figur 4.2. Effekt af stigende dosis af mancozeb og carbendazim på længden af rod-eksterne hyfer produceret af tre forskellige mykorrhizasvampe dyrket i symbiose med ært. Hyfelængden blev målt i 0-1 cm jordlaget tættest på nylonnettet, der adskilte den 33P-mærkede jord fra resten af pottevolumen. Middelværdier og standardfejl er angivet.
Det høje niveau af begge fungicider havde en stærkt hæmmede effekt på hyfevæksten af alle tre svampe (P < 0,001), og de lave værdier i højre side af Figur 4.2svarede til den målte hyfelængde i jord med mykorrhiza-fri kontrolplanter. Svampemyceliet kolonisering af den 33P-mærkede jord varierede i øvrigt, således at G. mosseae opnåede de største hyfelængder, men G. mosseae var samtidig den eneste af de tre svampe der reagerede negativt på de næsthøjeste doser af begge fungicider. De to lave fungiciddoseringer havde ikke nogen effekt på hyfelængden, bortset fra en stimulering af G. claroideum ved 1 x mancozeb. Data i Figur 4.2 stammer fra 0-1 cm jordlaget tættest på rødderne, men samme tendens blev observeret i 1-4 cm jordlaget (data ikke vist), hvor G. caledonium producerede den største hyfelængde uden fungicidbehandling (~50 m g-1 jord). Til gengæld blev G. caledonium signifikant hæmmet af ikke blot det højeste fungicidniveau men også af de to laveste niveauer af begge fungicider. Spredning af G. mosseae i dette lag blev også hæmmet af den midterste dosering af begge fungicider.
De lave niveauer af fungicidtilførsel stimulerede ikke blot svampenes vækst i rødder og i jord, men også svampenes optagelse af fosfat. Uden fungicid tilførsel var G. caledonium den eneste svamp, der optog signifikant mere 33P end niveauet i den mykorrhizafri kontrol, hvor 33P optagelsen var 0,26 kBq (Fig. 4.3). Markdosis
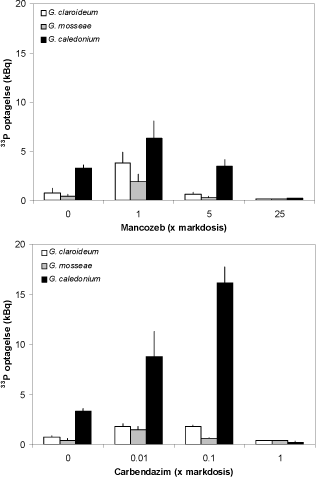
Figur 4.3. Effekt af stigende dosis af mancozeb og carbendazim på 33P-indhold hos ærteplanter, som blev dyrket i symbiose med tre forskellige mykorrhizasvampe. 33P er transporteret til planten via mycelium der vokser gennem nylonnettet og ind i den radioaktive jord. Middelværdier og SE er angivet.
af mancozeb resulterede i en general forøgelse af 33P optagelsen (P < 0,001 for alle svampe), og for G. claroideum var der tale om en 4x forøgelse (P < 0,01). En 5 x markdosis af mancozeb havde ikke nogen effekt på 33P optagelsen, medens en 25x dosis fuldstændig hæmmede svampenes 33P optagelse (P < 0,001). De laveste carbendazim doser forøgede ligeledes optagelsen af 33P (P < 0,001) og denne effekt var særlig udtalt hos G. caledonium, hvor optagelsen ved 0,1 x markdosis var 4,5 x så høj som i fraværet af fungicider. Markdosis af carbendazim hæmmede fuldstændigt optagelsen af 33P uptake (P = 0,02) i overensstemmelse med hæmningen af hyfevækst (P < 0,001).
Planternes vækst og totale P optagelse blev ikke signifikant påvirket af mykorrhiza, hvorimod 25 x mancozeb markdosis havde en fytotoksisk effekt (data ikke vist). Det kan ikke udelukkes, at den reducerede plantevækst ved 25 x mancozeb har påvirket etablering og funktion af mykorrhiza. Mancozeb i 5 x dosis havde en negativ effekt på P indholdet i symbiosen med G. mosseae, medens markdosis af carbendazim havde en generelt negativ indflydelse på planternes totale P indhold (data ikke vist).
En stimulerende effekt af en lav fungiciddosis blev bekræftet af resultater opnået i efterfølgende eksperimenter, hvor 1,0 x markdosis carbendazim blev anvendt som referencebehandling (se afsnit 4.2 og kapitel 5).
De opnåede resultater bekræfter, at mykorrhizasvampe er følsomme i forskellig grad for behandling af jorden med fungicider (Dodd & Jeffries, 1989); (Schreiner & Bethlenfalvay, 1997). Nærværende studie udvider denne erkendelse til at gælde hyfernes vækst i jorden og især hyfernes transport af P til planten. Resultaterne viser tydeligt, at skadetærskelen (NOAEL) er langt højere for behandling med mancozeb end med carbendazim. Det skal bemærkes, at denne rapport estimerer NOAEL vha. ekstrapolering af responskurver til nul-behandlings niveauet. Resultaterne kunne derimod ikke bekræfte hypotesen om, at fungiciddoser lavere en NOAEL ikke påvirker svampene. Små mængder fungicid gav således en markant stimulering af mykorrhizasvampes vækst og P optagelse. Dette er tidligere antydet for lave doser af carbendazim (Schweiger & Jakobsen, 1998) og for lave doser af en fenpropimorph-propiconazol blanding under markforhold (Schweiger et al., 2001). Der er flere mulige forklaringer på den uventede stimulering af mykorrhizasvampene ved de lave fungiciddoser. En mulighed er at mykorrhizasvampenes funktion påvirkes negativt af andre mikroorganismer (McAllister et al., 1997), og en relativt lavere skadetærskel (NOAEL) for eventuelle antagonister ville dermed kunne resultere i en positiv effekt af små fungiciddoser på mykorrhizasvampene. Denne forklaringsmodel forekommer dog mindre sandsynlig, idet lavdosis-stimuleringerne er blevet observeret med to fungicider i en række uafhængige eksperimenter og med anvendelse af forskellige mykorrhizasvampe. Forekomsten af eventuelle antagonistiske grupper må forventes at have varieret betydeligt imellem de nævnte eksperimenter.
En anden, og mere sandsynlig forklaring er, at de observerede stimuleringer skyldes hormesis, som er et velkendt økotoksikologisk fænomen (Calabrese, 2005). Hormesis (’excite’ eller stimulere) betegner den situation, hvor toksiner har en gunstig indflydelse på en organismes vækst, reproduktion eller levetid (Calabrese & Baldwin, 2003). Den biologiske baggrund for hormesis er ikke klarlagt. Der kunne være tale om en direkte effekt, men det er mere sandsynligt, at hormesis skyldes en over-kompensation som reaktion på en forstyrrelse af homeostasis (Trewavas & Stewart, 2003; Calabrese, 2005).
En sikker påvisning af hormesis kræver, at dosis-respons studier inddrager et tilstrækkeligt antal sub-NOAEL doser, samt at effekten af de lave doser studeres over tid. Et typisk hormesis-forløb viser en tidlig negativ effekt af lave toksin-doser, hvorefter organismens kompensering/overkompensering fører til at toksineffekten fremtræder som gunstig for organismen (Calabrese, 2005). Den klokkeformet ’sub-NOAEL’-responskurve, der i nærværende studie blev observeret for flere af de målte variable, svarer til resultater af dosis-respons studier af en lang række toksiner på stort set alle grupper organismer, herunder svampe (Calabrese, 2005). Hormesis-effekter på mykorrhizasvampe kan entydigt undersøges i dosis-respons studier på petriskål-baserede kulturer af rødder og mykorrhizasvampe (se (Nielsen et al., 2002). Disse kulturer indeholder ikke andre mikroorganismer end mykorrhizasvampe og er velegnede til direkte studier af tidsforløb.
Dosis-respons mønsteret var i nogle tilfælde væsentligt påvirket af arten af mykorrhizasvamp. Dette eksemplificeres tydeligst af den meget kraftige positive effekt af 0,01 og 0,1 x markdosis carbendazim på 33P optagelsen hos G. caledonium, hvor 1 x markdosis hæmmede P optagelsen fuldstændigt. Det er sandsynligt, at de doser, som mykorrhizasvampene eksponeres for under markforhold, er lavere end markdosis som følge af afgrødens opfangning af størstedelen af et sprøjtemiddel samt midlets binding til jorden i større eller mindre grad. Heraf følger, at, i det omfang mykorrhizasvampe i markjord eksponeres for fungicider, så er der sandsynligvis tale om sub-NOAEL doser, der har positive, snarere end negative effekter.
Hormesis-effekterne på mykorrhizasvampenes vækst og P optagelse er i denne undersøgelse observeret hos planter høstet 39 dage efter såning. Det er derfor ikke muligt at udtale sig om den langsigtede konsekvens af hormesis for svampenes vækst, næringsstoftransport og produktion af sporer. Det er muligt, at dosis-responskurven ændrer form over tid og at den fungicid-inducerede stimulering er et tidsbegrænset fænomen.
4.2 Effekt af fungicider på forskellige genotyper (isolater) af samme mykorrhizasvampeart
4.2.1 Formål
Vi har tidligere påvist en betydelig variation i vækstmønster af 15 isolater af G. mosseae (Munkvold et al., 2004), men det er uvist, om der også eksisterer intraspecifik variation i mykorrhizasvampenes tolerance for fungicider. Dette blev undersøgt ved anvendelse af G. caledonium isolater fra Snubbekorsgård forsøgsmarken samt nogle enkelte reference-svampe. Undersøgelsen testede hypotesen, at effekten af mancozeb og carbendazim ikke blot varierer mellem arter af mykorrhizasvampe men også mellem genotyper af den samme art.
4.2.2 Eksperimentelt
Eksperimentet afveg fra de øvrige i projektet ved at anvende Medicago truncatula som værtplante for mykorrhizasvampene i stedet for ært. Medicago truncatula er en bælgplante ligesom ært, og de to arter er temmelig homologe mht. til de gener der er centrale for mykorrhizasvampes kolonisering af planteroden (Oldroyd et al., 2005). Væksten af M. truncatula er relativt stærkt afhængig af mykorrhiza, og dermed kan eventuelle negative effekter af fungicider monitoreres i løbet af vækstperioden. Dette er ikke muligt hos ært, hvor vækstrespons på mykorrhiza er meget mindre end hos M. truncatula.
Der blev anvendt 7 pottekulturer af G. caledonium: fire enkeltsporekulturer fra mark 29 (K209, K225, K219 og K230), en enkeltsporekultur fra mark 21 (M21), en multisporekultur fra mark 29 (H-07) og en multisporekultur fra UK (BEG20). Enkeltspore- og multisporekultur betyder, at pottekulturen er initieret fra en enkelt eller fra flere sporer. Kun når kulturen er startet fra en enkelt spore, kan man være sikker på, at den kun indeholder en genotype. DNA sekvensering af svampene tyder på at K209 og K225 er en og samme genotype (type 1) medens K219 og K230 repræsenterer en anden genotype (type 2). M21 kulturen er også en type 2 genotype. De to sidste G. caledonium svampe (H-07 og BEG 20) er hverken type 1 eller 2. For at muliggøre sammenligning af fungicideffekter på variation inden for en art med variation imellem arter, blev der inddraget yderligere to svampe: En enkeltsporekultur af G. mosseae fra mark 29 (K218) og en multisporekultur af G. intraradices isolat fra sjællandsk overdrev (BEG87). Svampene blev opformeret i potter med jordkløver (Trifolium subterraneum L.) og jord fra disse kulturer blev anvendt som inokulum.
Medicago truncatula (Jemalong) blev dyrket i potter med 1000 g bestrålet jord tilført næringsstoffer, bortset fra fosfat (Pearson & Jakobsen, 1993). Jorden til hver potte blev iblandet 50 g inokulum af en af de ni svampe eller forblev fri for mykorrhizainokulum. Hver af de 10 mykorrhizabehandlinger blev kombineret med tre fungicidbehandlinger: ubehandlet, mancozeb (0,5 x markdosis) og carbendazim (0,5 x markdosis). Doseringerne blev opnået ved iblanding af 1,67 µg mancozeb g-1 jord samt 0,301 µg carbendazim g-1 jord. Fuld markdosis af carbendazim til ekstra potter med G. intraradices blev anvendt som referencebehandling til kontrol af modelsystemets funktion. Tidligere studier har vist at denne dosis hæmmer mykorrhizasvampes 33P optagelse med mere end 95 % (Schweiger & Jakobsen, 1998). Der blev yderligere inddraget en 0,1 x carbendazim markdosis for at undersøge eventuelle hormesis effekter på G. intraradices.
Jorden i de rodfri hyfeindvækstkamre blev blandet med 5 kBq carrier-fri 33PO4 g-1 samt med den relevante fungicidmængde, og kammeret blev placeret midt i potten. Der var 4 gentagelser for hver behandling, og de 128 potter blev placeret i vækstkammer med 16 t lys ved 21 °C og 8 t mørke ved 16 °C. Der blev tilført 100 mg NH4NO3-N i løbet af vækstperioden. Forsøget blev høstet 48 dage efter udplantning af spirede M. truncatula-frø, og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, længde af hyfer i 33P-mærket jord samt planternes indhold af 33P og totalt P.
4.2.3 Resultater og diskussion
Som forventet havde alle mykorrhizasvampe en betydelig effekt på plantevæksten, som i alle tilfælde blev mere end fordoblet (Figur 4.4). Mancozeb havde ikke nogen klar effekt på plantevæksten, hvorimod carbendazim påvirkede væksten negativt i alle tilfælde bortset fra G. intraradices symbiosen (BEG87, ingen effekt) og planter uden mykorrhiza (NM, en svag stimulering).
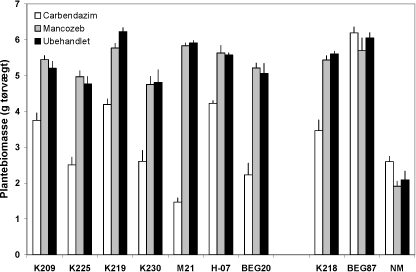
Figur 4.4. Effekt af mancozeb og carbendazim på biomasseproduktion hos M. truncatula dyrket i symbiose med 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218), G. intraradices (BEG87) eller uden mykorrhizapodning (NM). Middelværdier og standardfejl er vist. Effekten af svamp, af fungicidbehandling og interaktionen mellem de to faktorer var signifikant (P < 0,0001).
Resultaterne bekræfter således, at skadeeffekten af et fungicid afhænger af svampearten, og som noget nyt viser de, at effekten også varierer betydeligt inden for den samme art. Isolat M21 var fx betydelig mere følsomt for carbendazim end de to øvrige genotype 2 isolater (K219 og K230). Der var et nøje sammenfald mellem variationer i plantebiomasse (y) og planternes P indhold (x): y = 4,27Ln(x) - 2,36 (R² = 0,93), hvilket er i god overensstemmelse med, at P var eksperimentelt fastlagt som det vækstbegrænsende næringsstof. Det er et vigtigt resultat, at plantearter med en tydelig vækstrespons på mykorrhiza kan anvendes til direkte visualisering af skadeeffekter af fungicider og andre miljøfaktorer på mykorrhizasvampes funktion i næringsstofoptagelsen (Fig. 4.4).
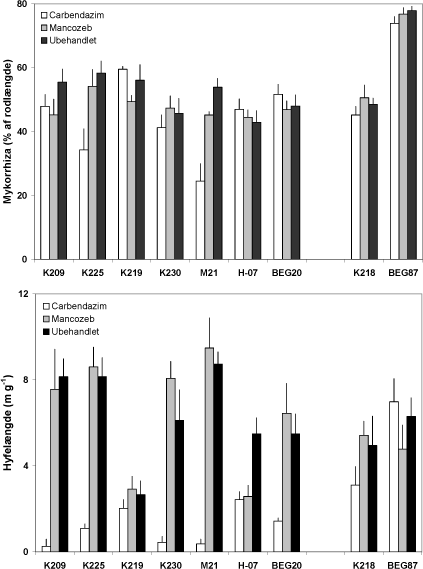
Figur 4.5. Effekt af mancozeb og carbendazim på mykorrhizaudvikling i rødder og i jord i symbioser mellem M. truncatula og 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218), G. intraradices (BEG87). Middelværdier og standardfejl er vist. Signifikansniveauer: mykorrhiza, P < 0,001 for fungicidbehandling og P< 0,0001for både svamp og interaktion; hyfelængde, P< 0,0001 for begge faktorer og for interaktionen.
Effekten af fungicidbehandlingerne blev undersøgt i større detalje ved analyse af svampenes vækst i rødder og jord samt af svampenes optagelse af 33P fra det rodfri mærkningskammer. Mykorrhizaen var veludviklet i rødderne af alle G. caledonium podede planter (44 - 58 % af rodlængden) og G. intraradices (BEG87) gav den kraftigste mykorrhizaudvikling (Figur 4.5). Mancozeb havde en mindre negativ effekt på rodkoloniseringen hos enkelte af G. caledonium isolaterne, medens carbendazim havde en tydelig negativ effekt hos K225 og M21 (forskellige genotyper). Hos de øvrige svampe kunne der ikke observeres nogen skadevirkning af carbendazim på rodkoloniseringen. Planter uden tilførsel af inokulum udviklede ikke mykorrhiza, og NM-behandlingen er derfor udeladt i både Fig. 4.5 og Fig. 4.6.
Hyfelængden i 33P mærket jord fra de mykorrhiza-fri planter var 1.01-1.21 m g-1, og de viste resultater i Figur 4.5 er alle justeret med baggrundsværdien for den respektive fungicidbehandling. Længden af mycelium i den 33P-mærkede jord varierede stærkt inden for G. caledonium isolaterne. Type 1 isolaterne (K209 og K225) var meget ens, hvorimod type 2 isolaterne varierede mellem 2.5 og 9 m hyfer g-1 jord (K219 og M21). Selv genotype 2 enkeltspore-isolaterne K219 og K230 fra samme mark var meget forskellige.
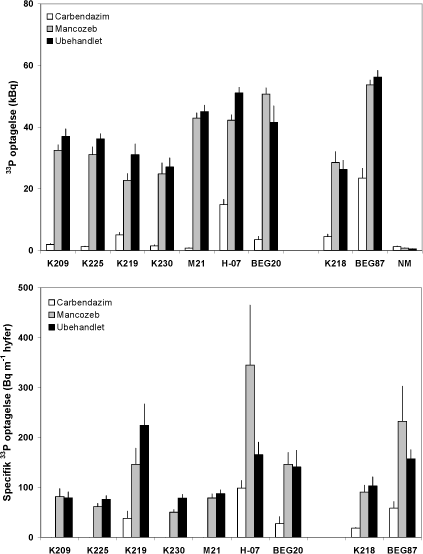
Figur 4.6. Effekt af mancozeb og carbendazim på mykorrhizasvampes totale og hyfelængde-specifikke optagelse og transport af 33P fra jord til M. truncatula, som blev dyrket i symbiose med 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218) eller G. intraradices (BEG87). Middelværdier og standardfejl er vist. Signifikansniveauer: 33P optagelse, P<0,0001 for begge faktorer og for interaktionen; specifik 33P optagelse, P<0,0001 for både svamp og fungicidbehandling og P<0,01 for interaktion.
Mancozeb gav en svag forøgelse i hyfelængden hos seks af symbioserne, men den eneste signifikante effekt var en reduktion i hyfelængde hos H-07. Carbendazim hæmmede hyfevæksten næsten 100 % hos K209, K230 og M21 symbioserne, men havde en ringe eller ingen effekt hos G. caledonium K219 og G. intraradices BEG87. Hos H-07 hæmmede begge fungicider hyfelængden med ~ 50 %. Der var en stor inter- og intraspecifik variation i fungiciders effekt på mykorrhizasvampenes vækst i jorden. Den mere markante effekt af fungicider på svampens vækst i jord end i rodkoloniseringen bekræfter tidligere resultater (Kjøller & Rosendahl, 2000b), og disse forskelle er formodentlig knyttet til en forskel i eksponeringsgrad over for fungicidet, således at svampen inde i roden i højere grad er beskyttet mod fungicidet end svampemyceliet i jorden.
Mykorrhizasvampene optog forskellig mængde 33P fra den radioaktive jord, også på det intraspecifikke niveau. En af genotype 2 svampene (M21) optog således tydeligt mere end de to øvrige type 2 svampe (K219 og K230), som havde optaget næsten lige meget 33P på trods af en markant mindre hyfelængde hos K219 (Figur 4.5 og 4.6). De to type 1 svampe havde optaget samme mængde 33P. Manzoceb havde en svagt negativ effekt på 33P optagelsen hos de fleste svampe, hvorimod carbendazimn havde en stærkt hæmmende effekt, også hos de svampe, hvor hyfevæksten ikke var påvirket eller blev hæmmet i mindre omfang. Dette illustreres godt i Figur 4.6, der viser 33P optagelsen pr meter hyfe i den 33P mærkede jord. Dette udtryk tager ikke hensyn til eventuelle tidsbetingede forskelle i hyfevæksten. Det fremgår at type 2 svampene K219 og K230 har meget forskellig kapacitet for P optagelse pr. hyfelængdeenhed. Det fremgår også, at carbendazim ikke alene påvirker mykorrhizasvampes funktion via en effekt på svampenes vækst, men også via en effekt på myceliets kapacitet til at optage og transportere P til planten. Carbendazim havde således en ringe effekt på hyfelængden af K219 og BEG87, men hyfernes P optagelse var stærkt hæmmet. Manzoceb forøgede kapaciteten for P optagelse hos H-07 og BEG87, og disse svampe var samtidig mest tolerante over for carbendazim behandlingen.
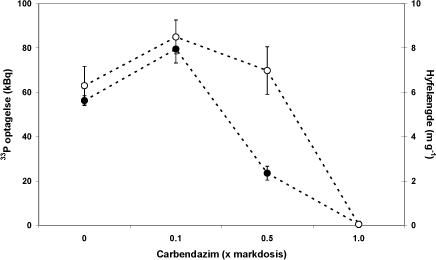
Figur 4.7. Dosis-respons for carbendazim effekter på hyfelængde (åbne cirkler) og 33P optagelse (fyldte cirkler) hos G. intraradices i symbiose med M truncatula.
Den negative kontrolbehandling (markdosis af carbendazim) medførte som forventet en >95 % hæmning i 33P optagelsen hos G. intraradices, hvilket bekræfter at modelsystemet fungerede optimalt (Fig. 4.7). Dosis-responskurver for carbendazimdoserne 0; 0,1; 0,5 og 1 x markdosis viser at den laveste dosis havde en stimulerende og hormesis-lignende effekt på både hyfelængden og 33P optagelsen hos G. intraradices, på samme måde som påvist for tre andre Glomus svampe i afsnit 5.1
Disse resultater fører til konklusionen, at tilførsel af 0,5 x markdosis mancozeb har en begrænset indflydelse på mykorrhizasvampes vækst og funktion i planters P optagelse. Derimod har 0,5 x markdosis carbendazim markante skadevirkninger, som dog afviger væsentligt imellem svampe, også imellem svampe af samme genotype. Det er bemærkelsesværdigt at en ensartet toksisk effekt på P optagelsen hos to svampe af samme genotype kunne være forårsaget af forskellige effekter af fungicidet: i det ene tilfælde blev hyfelængden reduceret kraftigt (K230) medens især P optagelseskapaciteten pr. hyfelængdeenhed blev påvirket hos den anden svamp (K219). Resultaterne har bekræftet den testede hypotese, at mykorrhizasvampe responderer forskelligt på fungicider, ikke blot imellem arter, men også blandt isolater eller genotyper inden for den samme art. Der konkluderes, at resultater opnået med en enkelt svamp ikke nødvendigvis gælder for andre svampe, heller ikke når der er tale om svampe inden for én enkelt art.
5 Fungiciders indflydelse på funktion af mykorrhiza baseret på rekonstruerede svampesamfund
5.1 Formål
Det var formålet at undersøge fungiciders effekt på kolonisering og fosfatoptagelse for de fire mulige kombinationer af tre mykorrhizasvampe isoleret fra det økologiske sædskifte; hver af de fire kombinationer blev sammenlignet med behandlinger, hvor svampene forekommer enkeltvis. Eksperimentet blev designet til test af følgende hypotese: Fungicider påvirker ikke blot funktionen af mykorrhizasvampe i monokultur, men kan også forskyde balancen i kombinationer af to eller tre svampe og dermed også sammensætning og funktion af marksamfund.
5.2 Eksperimentelt
Forsøget blev gennemført som vist i Tabel 5.1. Der blev anvendt 0,5 x markdosis af mancozeb og carbendazim ligesom i afsnit 5.2 To ekstra carbendazim-behandlinger (0,1 x og 1 x markdosis) blev inddraget med henblik på at konstruere responskurve for carbendazim og dermed opnå en kontrol af testsystemets funktion.
Tabel 5.1. Design af faktorielt eksperiment til undersøgelse af fungiciders effekt på enkeltkulturer og blandinger af tre mykorrhizasvampe. Hver af de otte svampebehandlinger var kombineret med de tre fungicidbehandlinger
| Svampeart (mark 29) | Mykorrhiza-inokulum (g potte-1) | Fungicid |
| A | 150 | 0,5x Carbendazim |
| B | 150 | 0,5x Mancozeb |
| C | 150 | Ubehandlet |
| AB | 75+75 | |
| AC | 75+75 | |
| BC | 75+75 | |
| ABC | 50+50+50 | |
| NM | 0 | |
| B | 150 | 0,1x Carbendazim |
| B | 150 | 1x Carbendazim |
De anvendte svampe er isoleret fra det økologiske sædskifte (mark 29, Snubbekorsgaard). A og B er Glomus mosseae (hhv. kultur nr. L 541, BEG84 og kultur nr. L 545, BEG85) og C er G. caledonium (kultur nr. L 554, BEG86). Den forudgående kvalitetstest af inokulum viste at A, B, og C havde koloniseret hhv. 64, 61 og 54 % af den totale rodlængde hos jordkløver-testplanterne. Der blev anvendt samme mængde inokulum af hver svamp og den totale mængde inokulum var ens i alle mykorrhizabehandlinger.
Forsøgsjorden var en 1:1 blanding (vægt/vægt) af Rørrendegård-jord og kvartssand, som var elektronstråle-bestrålet (10 MeV, 15 kGy) og derefter iblandet næringsstoffer (Pearson & Jakobsen, 1993). Rørrendegård er naboejendom til Snubbekorsgård, og jordtypen er den samme, men indholdet af plantetilgængeligt P er lavere i den anvendte Rørrendegård-jord.
Jorden blev påfyldt 1.5 L potter indeholdende et rodfrit mycelie-indvækstkammer i form af et 50 mL plastrør, hvor begge åbninger var lukket med et nylon net med 25 µm maskestørrelse. Kammeret var fyldt med 59 g jord mærket med 3.5 kBq carrier-fri 33PO4 g-1. Med fem gentagelser af hver behandling, blev der opstillet 130 potter i vækstkammer med 16 t lys ved 21 °C og 8 t mørke ved 16 °C. Potter med to ærteplanter (Solare) i hver blev vandet til 65 % af vandkapaciteten og blev gødet med 4 x 50 mg N i form af en NH4NO3-opløsning i løbet af vækstperioden. Forsøget blev høstet 56 dage efter såning og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, længde af hyfer i 33P-mærket jord samt planternes indhold af 33P og totalt P.
I behandlinger med mere end én svamp blev de enkelte svampe kvantificeret ved at amplificere LSU-rDNA fra rodprøver med specifikke primere. PCR prøverne blev derefter analyseret med SSCP (Single Strand Conformational Polymorphism) for at kunne adskille de tre svampe (Kjøller & Rosendahl 2000a). Svampene blev rangeret på skalaen 0, 1 eller 2 i forhold til SSCP-båndenes intensitet. De enkelte svampes bidrag til den samlede kolonisering blev derefter beregnet (se tekst til Figur 5.2).
5.3 Resultater
5.3.1 Vekselvirkning mellem svampe i fravær af fungicider
De tre svampe koloniserede ærteplanternes rødder i forskelligt omfang, når de var hver for sig. Mere end 50 % af rodlængden havde udviklet mykorrhiza i planter podet alene med svamp C (G. caledonium), medens podning med G. mosseae svampene (A eller B) førte til koloniseringsniveauer på kun omkring 13 % (Figur 5.1). Denne overraskende afvigelse fra de forventede koloniseringsniveauer, baseret på data for kvalitetskontrol af inokulum, skal formodentlig forklares ved en vis grad af vært-svamp præference mykorrhiza-symbiosen (Helgason et al., 2002).
Tilsyneladende eksisterede der en gunstig vekselvirkning mellem A og B, således at de to svampe i forening koloniserede en større andel af rodlængden end når svampene var alene. Idet inokulummængder var halveret, hvor svampene blev blandet to og to, må dette resultat skyldes en vekselvirkning i en eller flere af de signal-pathways, der er involveret i mykorrhizadannelsen. Den positive vekselvirkning mellem A og B førte til en fordobling af hyfelængden og en tredobling af myceliets 33P optagelse (Figur 5.1). Kvantificeringen af de enkelte svampe i kombinations-behandlingerne tyder på, at svamp A bliver stimuleret når den kombineres med svamp B, og i særdeleshed i kombination med svamp C (Figur 5.2, ubehandlet). Kombinationer af svamp A og B med svamp C førte til en svag reduktion i kolonisering af rødderne i forhold til behandlingen med svamp C alene (Figur 5.1). Dette kunne skyldes en fortynding af C-inokulum med de svagere A- og/eller B-inokula. Den PCR-baserede kvantificering af de enkelte svampe tyder dog på en egentlig hæmning af svamp C, når A og/eller B var til stede samtidig (Figur 5.2, ubehandlet). En hæmning af svamp C induceret af svamp A og/eller svamp B er i overensstemmelse med, at både hyfelængde og 33P optagelse var lavere i kombinationer med C, end hvor C var alene (Figur 5.1). Planter uden tilførsel af inokulum af mykorrhizasvampe udviklede ikke mykorrhiza.
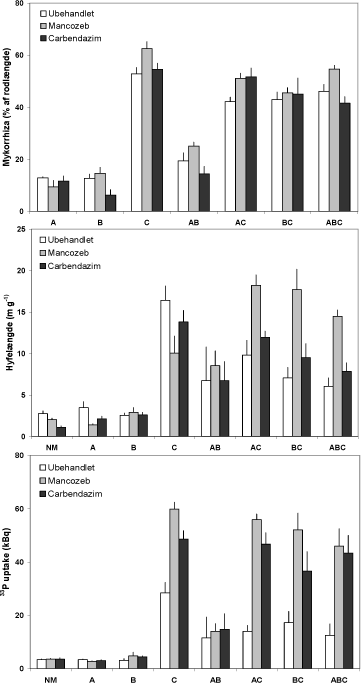
Figur 5.1. Effekt af mancozeb og carbendazim på tre mykorrhizasvampes kolonisering af ærterødder og af jord samt på svampenes 33P-optagelse. Rodkolonisering er målt ved mikroskopi af farvede rodprøver. Svampene var til stede enkeltvis (A, B og C) eller i kombination (AB, AC, BC, ABC) og den anvendte mængde inokulum var ens i de syv behandlinger. Én behandling modtog ikke svampeinokulum (NM). Middelværdier og standardfejl er vist.
5.3.2 Fungicideffekter på vekselvirkning mellem mykorrhizasvampe
Fungicidernes indflydelse på mykorrhizasvampenes vækst og 33P optagelse varierede betydeligt mellem de forskellige svampebehandlinger. Mancozeb stimulerede rodkoloniseringen i C, AC og ABC behandlingerne, medens en modsat rettet tendens kunne ses i A behandlingen (Figur 5.1). Hyfelængder og 33P optagelser hos A og B behandlingerne er vanskelige at tolke, idet værdier har samme niveau som målt hos planter uden mykorrhiza. Imidlertid havde manzoceb en negativ effekt på hyfelængden i alle 4 behandlinger med C svampen. Hvor C var kombineret med de to andre svampe var hyfelængden fordoblet i mancozeb-behandlingen, som dog havde en negativ effekt, hvor svamp C var alene.
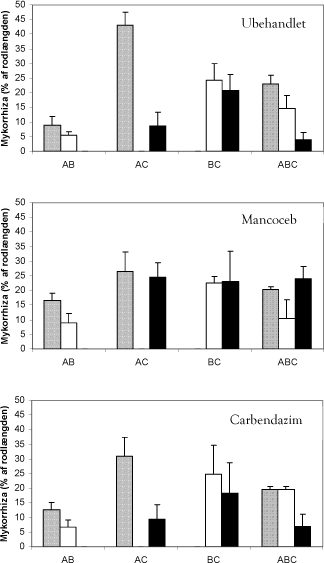
Figur 5.2. Forekomst af de tre svampe A (skraverede søjler), B (åbne søjler) og C (sorte søjler) i forsøg med mere end én svamp. Bidraget til mykorrhizadannelse (% af rodlængde) fra den enkelte svamp er beregnet på baggrund af total rodkolonisering (Figur 5.1) og den relative intensitet af de genetiske markører.
En samlet vurdering af effekter på hyfelængder og rodkolonisering tyder på, at mancozeb havde en modsatrettet effekt på det rodeksterne og det rodinterne mycelium (Figur 5.1). Mancozeb forøgede 33P optagelsen meget tydeligt i alle fire C-behandlinger: Optagelsen blev fordoblet, hvor C var den eneste svamp, tredoblet i BC kombinationen og næsten firedoblet i AC og ABC kombinationerne (Figur 5.1).
Behandling med mancozeb medførte, at balancen mellem A og C blev ændret, således at svamp C klarede sig væsentlig bedre end i tilsvarende kombinationer uden fungicidtilførsel (Figur 5.2, mancozeb). Behandling med carbendazim ændrede derimod ikke på svampenes indbyrdes hyppighed i forhold til i ubehandlet jord (Figur 5.2, carbendazim og ubehandlet). Data for P optagelsen viser, at kombinationer med svamp C havde en lavere P optagelse, end når svamp C var alene (Figur 5.1). Denne negative effekt af kombinationerne på svampenes P optagelse blev tydeligvis modvirket af mancozeb. Dette skyldes sandsynligvis at mancozeb øger svamp C’s andel af koloniseringen (Figur 5.2). Dette underbygges af, at hyfelængden i fravær af fungicider var mindre i kombinationer med svamp C, end når svamp C var alene; det omvendte forhold gjorde sig gældende i behandlinger med mancozeb, hvor den største hyfelængde blev målt i kombinationerne (Figur 5.1).
Effekten af carbendazim på rodkolonisering var begrænset til en svag stimulering i AC behandlingen, medens hyfelængden blev stimuleret i behandlinger med svamp C. Ligesom mancozeb, havde carbendazim en kraftig positiv virkning på 33P optagelsen i behandlinger omfattende C svampen. De kraftigste effekter, op til tre ganges forøgelse, blev opnået i AC og ABC kombinationerne. Sammenligner man de enkelte svampes kolonisering, ses det at svamp C ikke klarer sig godt i konkurrencen med svamp A, men at balancen mellem svamp A og svamp C svarer til hvad man ser i ubehandlede planter (Figur 5.2). Dvs at stimuleringen ikke kan forklares som en stimulering af svamp C, som har vist sig at øge planternes fosfor- optagelse, men må tilskrives hormesis-effekter som beskrevet i kapitel 4.
5.3.3 Effekter på plantevækst samt dosis-respons forløb for Glomus mosseae
Plantebiomassen varierede i området 10 – 12 g tør vægt pr potte. Den mindste vækst blev opnået i behandlinger med alene A eller B svampen, medens de største planter blev høstet i potter uden mykorrhizapodning samt i C og AB behandlingerne (data ikke vist). Der blev observeret en tendens til vækstreduktion i fungicidbehandlede A, B og AB potter. Glomus mosseae svampen ”B” blev anvendt til test af modelsystemets funktion. Valget var ikke ideelt pga. det lave koloniseringsniveau i fravær af fungicid. Imidlertid viser dosis-respons kurverne, at testsystemet fungerede som forventet, idet både 0,1 og 0,5 x markdosis resulterede i en tydeligt forøget rodkolonisering, hvor 1 x markdosis førte til en svag reduktion (Figur 5.3). Uden fungicid afviger hyfelængde og 33P optagelse ikke fra værdier opnået uden mykorrhiza (Se Figurer 5.1 og 5.3), men 0,1 x markdosis carbendazim resulterer i en stimulering af begge variable. Dette hormesis-lignende responsmønster er i overensstemmelse med resultaterne beskrevet i afsnit 4.1, 4.2 samt i dette afsnit.
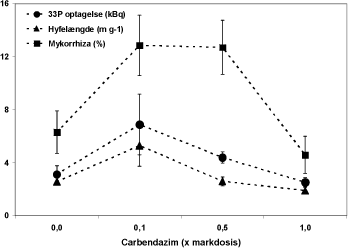
Figur 5.3. Dosis-respons for carbendazim effekter på rodkolonisering, hyfelængde og 33P optagelse hos G. mosseae ”B” i symbiose med Solare-ært.
5.4 Diskussion
Forsøget viste, at effekten af flere mykorrhizasvampe på fosforoptagelsen ikke kunne forudsiges ud fra de enkelte svampes effekt på fosforoptagelsen. Dette skyldes sandsynligvis, at der er en konkurrence mellem svampene, samtidig med at svampene også kan påvirke hinanden positivt. Konkurrence mellem AM svampe er tidligere vist ved at anvende protein (enzym) markører, hvor det blev vist at visse svampe kan udelukke andre svampe fra rodsystemet (Hepper et al., 1988).
Fungicideffekter på mykorrhizasvampe enkeltvis og i blanding er tidligere blevet undersøgt, og resultaterne viste en større fungicidtolerance når svampene var i blanding end når de forekom enkeltvis (Schreiner & Bethlenfalvay, 1997). De anvendte doser var dog ~50 x højere end markdosis og derfor ikke realistiske. Stimulerende effekter er tidligere blevet påvist af anbefalede doser carbendazim og en fenrpopimorph – propiconazol blanding (Schweiger et al., 2001) samt anbefalede doser af metalaxyl (Groth & Martinson, 1983).
Begge fungicider havde en begrænset indflydelse på mykorrhizasvampenes totale kolonisering af ærterødderne. Mancozeb havde dog en effekt på konkurrencen og samspillet mellem de enkelte svampe. I de fleste tilfælde var der tale om en fungicid-induceret stimulering af svampenes aktivitet. En del af denne effekt kunne forklares som en forskydning mellem de enkelte svampes rodkolonisering. Dette resultat bekræfter hypotesen om, at fungicider kan påvirke sammensætningen af mykorrhizasvampe samfund. Både carbendazim- og især mancozeb- behandlingerne resulterede her i en højere fosforoptagelse, men dette bør ikke tolkes som et generelt billede på effekter af fungicidbehandling på mykorrhizasvampe, idet svampenes indbyrdes samspil vil have stor betydning for udfaldet.
6 Fungiciders indflydelse på forsøgsmarkernes naturlige mykorrhizasvampe-samfund
6.1 Formål
Det var formålet at undersøge effekten af fungicider på sammensætning og funktion af naturlige samfund af mykorrhizasvampe. Der blev anvendt jord fra to forskellige sædskifter og inden for hvert sædskifte blev jorden indsamlet som fire delprøver langs en 50 m transekt. Eksperimentet skulle teste hypotesen, at effekten af fungicider ville være større end effekten af dyrkningspraksis eller stedvariationen.
6.2 Eksperimentelt
Forsøget blev gennemført som et potteforsøg med de 24 behandlinger, der fremkommer ved at kombinere dyrkningspraksis, markprøver og fungicidbehandling som vist i Tabel 6.1. Der var 5 gentagelser for hver behandling og der blev i alt opstillet 120 potter.
Tabel 6.1. Design af eksperiment til undersøgelse af fungicideffekter på samfund af mykorrhizasvampe i markjord.
| Dyrkningspraksis | Markprøver | Fungicidbehandling (x markdosis) |
| 1. Økologisk Mark 29 2. Konventionel Mark 21 |
A B C D |
Mancozeb (1) Carbendazim (0,1) Ubehandlet |
Forsøgsjorden blev indsamlet langs 50 m transekter i mark 21 og mark 29 (M21 og M29), som er vist i Figur 2.1. Hvert transekt blev opdelt i fire afsnit á 12,5 m, og der blev udtaget delprøver i 12 tilfældigt placerede punkter i hvert afsnit; den mindste afstand mellem punkterne var 0,25 m. De 12 delprøver fra hvert afsnit blev slået sammen til én og dermed blev der indsamlet 4 prøver (A, B, C og D) fra hver mark. Hver af de 8 markprøver blev sigtet gennem et 5 mm trådnet og derefter blandet med en tilsvarende vægtmængde kvartssand (0,3 - 0,6 mm) for at sænke jordens P indhold og dermed opnå en øget mykorrhizaudvikling. Alle 8 blandinger blev tilført en fosfatfri gødningsblanding (Pearson & Jakobsen, 1993).
Hver prøve blev delt i tre portioner svarende til de tre fungicidbehandlinger, mancozeb (1 x markdosis eller 3,3 µg g-1 jord), carbendazim (0,1 x markdosis eller 0,06 µg g-1 jord) og ubehandlet. Beregning af markdosisækvivalenter er beskrevet i afsnit 5.1. De i alt 24 jordblandinger blev påfyldt potter uden afdræn med 1500 g jord i hver. Hver potte indeholdt et hyfe-indvækstkammer med jord, der var iblandet 5 kBq carrier-fri 33P g-1.
To ærteplanter (Solare) blev dyrket i hver potte og de i alt 120 potter blev placeret i et vækstkammer med 16 t lys ved 21°C og 8 t mørke ved 16°C. Potterne blev vandet efter behov til en vægt svarende til 65 % af vandkapaciteten.
Planter blev høstet efter 6 uger, og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, samt planternes indhold af 33P. Diversiteten af mykorrhizasvampe i rødderne blev bestemt på delprøver af rodsystemet fra hver potte. Prøverne blev opbevaret ved –20 °C og T-RFLP blev anvendt til at bestemme antal forskellige genotyper af mykorrhizasvampe. DNA blev ekstraheret fra samtlige rodprøver, og generne som koder for den lille ribosomale subunit (SSU-rDNA) blev mangfoldiggjort vha. PCR og mykorrhizaspecifikke primere. Prøverne blev oprenset og skåret med to restriktionsenzymer, TAQ og SAU. Fragmenterne blev mærket med en fluoriserende primer og blev kørt på en ABI 377 sekvenator. Fragmenternes størrelse (længde) og intensitet blev målt. De blev derefter sammenlignet med fragmentlængder hos alle registrerede mykorrhizasvampe for at eliminere fragmenter, der ikke kunne identificeres.
Forekomsten af arter blev registreret for hver gentagelse og derefter analyseret med en principal komponent analyse (PCA). Diversiteten blev beregnet som Shannon diversitets indeks for de to marker og for de tre fungicidbehandlinger.
6.3 Resultater
Planter fra konventionelt dyrket jord (M21) havde 5-10 % større biomasse end de tilsvarende planter fra økologisk dyrket jord (P = 0,08; data ikke vist). Denne forskel var lavere end forventet på baggrund af forskelle i jorden fosforindhold som er 25 og 45 mg P kg-1 i ublandet jord fra henholdsvis den økologisk og den konventionelt dyrkede mark. Den begrænsede forskel i væksten mellem de to marker skyldes formodentlig at mykorrhiza udbalancerede en negativ effekt af et lavt P niveau. Mellem 25 og 40 % af rodlængden havde udviklet mykorrhiza, og kolonisering var signifikant højest (P<0,0001) i jord fra den økologiske mark med det laveste P indhold (Figur 6.1, M29). Koloniseringen varierede også mellem delprøver inden for både mark 21 og mark 29 (P<0,05). Der var en signifikant interaktion mellem delprøve og fungicidbehandling i mark 21 (P<0,05); det tilsvarende signifikansniveau for mark 29 var P=0,052.
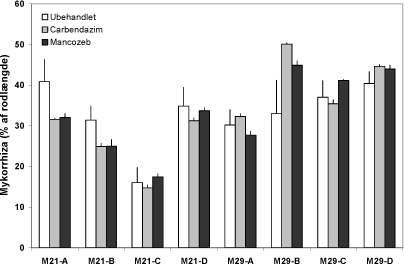
Figur 6.1. Mykorrhizaudvikling i rødder hos Solare-ært dyrket 6 uger i jord fra det konventionelle (M21) og det økologiske sædskifte (M29) ved Snubbekorsgaard. A, B, C og D angiver fire forskellige prøver langs transekter. Søjler angiver den procentvise andel af rodlængden som var koloniseret af mykorrhizasvampe.
Mykorrhizadannelsen er noget lavere end de maksimumværdier på 60-70 %, der tidligere er rapporteret i markforsøg med ært i det økologiske sædskifte (Schweiger et al., 2001; Bodker et al., 2002). I nærværende projekt er koloniseringen tilsvarende blevet målt på ærterødder indsamlet langs de to transekter i juli 2003. Disse rødder udviste en kolonisering på hhv. 62 % og 46 % i det konventionelle og det økologiske sædskifte. Disse data følger ikke det forventede mønster med den kraftigste kolonisering i jorden med det laveste P niveau, men forskelle i sædskifter og jordbehandling kan have spillet en rolle. Der var en tendens til, at fungicidbehandling øgede koloniseringen i det økologiske sædskifte og reducerede koloniseringen i det konventionelle. Disse forskelle var dog ikke signifikante. Derimod varierede koloniseringen signifikant mellem de fire deltransekter (A – D) i hver mark (P<0,001). Denne variation skyldtes ikke forskelle i jordens fosfatindhold og årsagen er ikke identificeret.
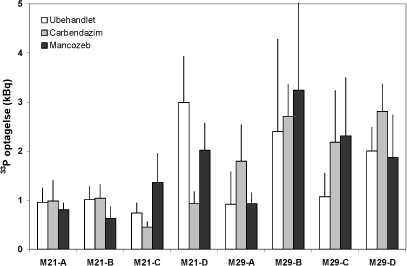
Figur 6.2. 33P indhold i Solare-ært dyrket 6 uger i jord fra det konventionelle og det økologiske sædskifte ved Snubbekorsgaard.
Det var hensigten at forsøg 2 skulle give information om fungicidernes betydning for mykorrhizasvampenes optagelse af P fra jord til plante. Hertil blev der anvendt et 33P-mærket jordvolumen som var adskilt fra resten af jorden vha. et 25µm nylon net. Svampehyfer, men ikke rødder, kan vokse igennem nettet og derved få adgang til den radioaktive fosfor. Planterne indeholdt overraskende små mængder 33P, formodentlig pga. en begrænset hyfeindvækst inden for de 6 ugers vækstperiode (Figur 6.2); de små 33P værdier skal vurderes på baggrund af en samlet tilførsel på 110 kBq pr. potte. De lave værdier for optagelse af 33P er behæftet med store standardfejl på middelværdier, og der kan ikke udledes nogen signifikant effekt af fungicidbehandling. Derimod var der en signifikant forskel i 33P optagelse mellem de to marker (P<0,05).
Sammensætning af svampesamfundet i rødderne er analyseret vha. T-RFLP (Terminal Restriction Fragment Length Polymorphism), som er baseret på ekstraktion af rDNA og derefter PCR amplificering ved anvendelse af en terminalt placeret fluorescerende primer. PCR produktet skæres derefter med restriktionsenzymer. Som følge af sekvensvariation, vil det terminale restriktions-site være forskelligt for hver art i samfundet. Det digitale output giver oplysning om produktets længde i form af basepar (f. eks. for en art) og intensiteten af fluorescenssignalerne er et mål for hyppigheden af arterne i samfundet.
For at fjerne mulige falske mykorrhizasekvenser fra analyserne, blev samtlige kendte sekvenser af SSU rDNA samlet og på computer skåret med de to restriktionsenzymer. De resulterende fragmenter blev samlet i en database. Kun de fundne fragmenter som havde samme længde som sekvenser i databasen blev anvendt til PCA og til udregning af diversitetsindeks.
For yderligere at bekræfte forskellen mellem betydningen af dyrkningspraksis: økologisk og konventionel, markens heterogenitet og fungicidbehandling, blev det yderligere undersøgt, om forskellene kunne bekræftes på baggrund af tilstedeværelsen af kendte svampe. Dertil blev LSU rDNA fra prøverne amplificeret, og PCR produkterne derefter klonet og sekvenseret. Dette er meget arbejdskrævende og kostbart, blev kun få af prøverne blev derfor analyseret.
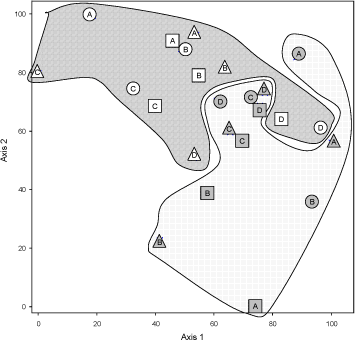
Figur 6.3. PCA på T-RFLP data efter skæring med restriktions-enzymet TAQ. Punkter fra de to marker er samlet. Mark 29 er linie-skraveret og mark 21 er prikket. Cirkler er med carbendazin, trekanter med mancoceb og firkanter er ubehandlede. Bogstaverne henviser til delprøve langs transektet.
PCA analyserne på T-RFLP data viste at det der primært grupperede data var hvilken mark prøverne var taget fra. Dette var tilfældet med begge restriktionsenzymer (Figur 6.3, TAQ og 6.4 (SAU). Den næste faktor som skiller data er svær at vurdere, men der er ikke noget der tyder på at fungicidbehandlingen skulle have større betydning end den heterogenitet, der findes inden for marken. Især når PCA blev udført med fragmenter skåret med TAQ, var punkter fra samme plot grupperer tættere sammen end punkter fra samme fungicidbehandling (Figur 6.3).
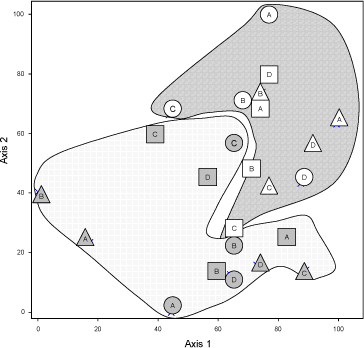
Figur 6.4. PCA på T-RFLP data efter skæring med restriktions-enzymet SAU. Punkter fra de to marker er samlet. Mark 29 er linie-skraveret og mark 21 er prikket. Cirkler er med carbendazin, trekanter med mancozeb og firkanter er ubehandlede. Bogstaverne henviser til delprøve langs transektet.
Resultaterne kunne bekræftes ved at amplificere og klone DNA direkte fra prøverne. Også her viste det sig også at markerne betød mest for fordelingen af arterne (Figur 6.5).
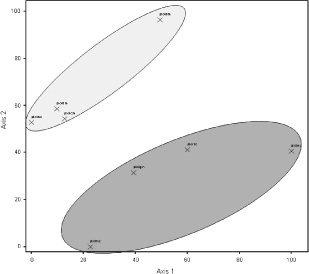
Figur 6.5. PCA af diversitets data opnået ved PCR, kloning og sekvensering af LSU rDNA. Mørk gråtone indeholder data fra mark 29, lys gråtone fra mark 21.
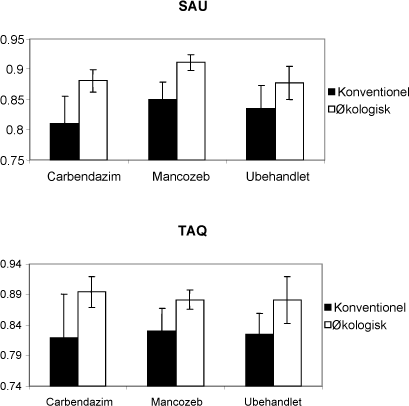
Figur 6.6. Diversitetsindeks udregnet på baggrund af T-RFLP data. Øverst udregnet for fragmenter skåret med TAQ og nederst skåret med SAU.
Diversiteten af mykorrhizasvampe i de to marken viste sig at være forskellig, idet der var en højere diversitet i den økologiske mark (Figur 6.6). Diversiteten blev ikke påvirket af fungicidbehandlingen. Der blev opnået de samme resultater uanset hvilket restriktionsenzym der blev anvendt. Resultaterne bekræfter PCA analyserne, idet forskellen mellem markerne giver de største forskelle, mens fungicidbehandlingens betydning ikke var målbar.
Samlende viste undersøgelserne, at der var stor forskel på de to marker, og at disse forskelle også gjaldt svampesamfundets sammensætning. Denne forskel i samfundenes sammensætning forklarer også den målte forskel i mykorrhizaudvikling og fosforoptagelse. Prøverne blev indsamlet langs et 50 m transekt, og det var interessant, at der var en betydelig variation indenfor markerne, både hvad angik mykorrhizakolonisering og fosforoptagelse. Dette blev også afsløret af PCA-analyserne, hvor punkter fra samme plot i transektet lå tæt sammen. Fungicid behandlingen havde kun lille effekt på svampesamfundene, og det var også svært at se en generel effekt af fungicider på kolonisering og fosforoptagelsen. Konkluderende kan vi sige at dyrkningspraksis er afgørende for svampesamfundets sammensætning og funktion, mens effekten af fungicid behandling vil være mindre end sted-variationen i marken.
7 Samlet diskussion
Denne undersøgelse anlægger en bred synsvinkel på fungiciders indflydelse på mykorrhizasvampe. Tidligere studier har især sammenlignet forskellige fungiciders effekt på enkelte svampeisolater og har kun i begrænset omfang inddraget dosis-respons undersøgelser. Eksperimenterne beskrevet i denne rapport har undersøgt fungicideffekter på mykorrhizasvampe på individ-, populations- og samfundsniveau. Denne integrerede tilgang giver et mere realistisk billede af de mulige konsekvenser af landbrugets anvendelse af fungicider for betydningen af mykorrhiza som plantevækstfaktor.
Projektet har så vidt muligt kun anvendt mykorrhizasvampe, som er isoleret fra forsøgsmarken i Tåstrup i dette projekt eller i et tidligere projekt under Den Tværministerielle Pesticidforskningsindsats. De anvendte svampe hører til de mest almindeligt forekommende i marken, og deres respons på fungicider, enten som monokulturer eller i blandinger, må forventes at være repræsentative for fungicideffekter på mykorrhizasvampe i en dansk landbrugsjord.
På individniveau bekræftede projektet, at fungicider kan hæmme mykorrhizasvampe (Trappe et al., 1984) og resultaterne viste, at der kan være stor forskel på, hvordan forskellige arter og selv forskellige genotyper indenfor samme art reagerer på fungicider. Resultaterne beskrevet i kapitel 4 og 5 viser, at carbendazim er yderst toksisk for mykorrhizasvampe og carbendazim er af samme årsag blevet anvendt som kontrolbehandling i mange undersøgelser af mykorrhizas effekt på plantevæksten (Kahiluoto & Vestberg, 2000; Thingstrup et al., 2000). NOAEL-værdier (No Observed Adverse Effect Level) var således også langt lavere for carbendazim (0,1 – 0,5 x markdosis) end for mancozeb (5 – 25 x markdosis) (Kapitel 4.1). Det var derimod uventet, at tilførsel af små doser (< NOAEL) af begge fungicider havde en generelt stimulerende effekt på en række forskellige mykorrhizasvampe undersøgt i kapitel 4 og 5. Stimulerende effekter af metalaxyl på mykorrhiza er tidligere blevet rapporteret hos majs og sojabønne (Groth & Martinson, 1983), og et markforsøg med ært i Tåstrup-marken antydede at en propiconazol-fenpropimorph blanding og carbendazim i små doser stimulerede mykorrhizasvampes P optagelse (Schweiger et al., 2001). Resultaterne i kapitel 4 og 5 viser en klassisk hormesis effekt, dvs. at dosis-respons kurven er omvendt J-formet, således at små doser har en positiv effekt, som forsvinder gradvist og bliver negativ, når dosis forøges yderligere. Den fysiologiske baggrund for stimuleringen blev ikke klarlagt, men involverer formodentlig overkompensation i svampens reaktion på en svag stress påført af fungiciderne. Studier med andre organismer har vist, at hormesis-effekten er tidsafhængig, således at der initialt er en negativ effekt af de små toksin-doser, men at dette billede over tid ændres til en stimulering (Calabrese, 2005). Alternativt, kunne den observerede stimulering måske skyldes en større skadevirkning af små fungiciddoser på andre mikroorganismer med en mulig antagonistisk effekt over for mykorrhizasvampe end på mykorrhiza-svampene selv. Denne forklaring forekommer mindre sandsynlig, dels fordi mykorrhiza-svampe er biotrofe og derfor ikke konkurrerer om kulstofsubstrat med andre mikroorganismer, dels fordi andre former for interaktion mellem mykorrhiza-svampe og saprotrofe mikroorganismer er komplekse og hyppigst til gavn for mykorrhizasvampene (McAllister et al. 1997; Garcia-Romera et al. 1998; Fracchia et al. 2000). Antagonisme fra saprotrofe mikroorganismer over for mykorrhizasvampe ville derfor næppe optræde i tre uafhængige eksperimenter.
På populationsniveau viste undersøgelsen, at der er et betydeligt antal genotyper af forskellige arter af mykorrhizasvampe til stede i marken. Den genotypiske variation og fordeling indenfor de enkelte marker kunne forklare det meste af variationen, når nabomarker med økologisk eller konventionelt dyrkningssystem blev sammenlignet. Forskellene mellem konventionel og økologisk dyrkning gav altså ikke anledning til en ændret genetisk diversitet indenfor arterne. Derfor skal det ikke forventes, at eventuelle effekter af forskelle i jordens indhold af pesticidrester slår igennem på populationsniveau.
Dyrkningshistorien (konventionel eller økologisk dyrkning) har derimod en væsentlig effekt på diversiteten på samfundsniveau, selvom diversiteten også varierer inden for marken. Kapitel 6 viser, at de mark- og stedbetingede forskelle var tydeligt større end små og ikke-signifikante effekter af fungicider og dette er en ny opdagelse. Resultaterne fra kapitel 5, hvor fungicideffekterne blev undersøgt på rekonstruerede samfund af få (2 – 3) mykorrhizasvampe viser, at fungicid-behandling markant kunne ændre udfaldet af interaktionerne i de mulige kombinationer af de tre svampe. Udfaldet var samtidig meget svært at forudsige ud fra viden om de enkelte svampe.
Populationsstudierne beskrevet i kapitel 3 har direkte relevans for markforhold, medens ekstrapolering af potteforsøgenes resultater til markforhold kræver en nærmere vurdering. Ét spørgsmål vedrører sandsynligheden for en faktisk eksponering af marksamfund af mykorrhizasvampe for fungicider, et andet vedrører de mulige begrænsninger, som anvendelsen af semisterile vækstmedier medfører.
En del af et udsprøjtet pesticid vil bliver opfanget af afgrøden, og den mængde der rammer jordoverfladen aftager med stigende plantedække (Probst et al., 2005).
I kartoffelmarker, der typisk sprøjtes 5-10 gange med mancozeb vil kun 5-15 % ramme jorden i de vækststadier hvor sprøjtningen er mest intensiv (Jensen og Spliid, 2003). Den intensive sprøjtning kan alligevel medføre, at mancozeb i løbet af vækstsæsonen akkumuleres til residual-koncentrationer på op til 10 mg kg-1 jord, selv når der er taget højde for nedbrydning af stoffet (Christensen og Helweg, 2003). Denne beregning antager en opblanding i et 6,6 cm jordlag. Selv om mancozeb adsorberes forholdsvis stærkt i jorden, må det forventes, at nedbør og mekanisk jordbearbejdning vil resultere i eksponering af mykorrhizasvampe i et 5-10 cm jordlag. Den estimerede residualkoncentration falder i området mellem de to laverste mancozeb-doseringer anvendt i nærværende projekt og dette forhold styrker muligheden for at ekstrapolere til markforhold. Det forekommer derfor sandsynligt at den målte mancozeb-inducerede stimulering af mykorrhiza også kan optræde under markforhold. Ligeledes må det forventes at den høje persistens af carbendazim vil kunne føre til effekter på mykorrhiza under markforhold, enten negative eller positive (hormesis). Carbendazim anvendes ikke i dansk planteavl. Sandsynligheden for at hormesis optræder under markforhold styrkes af to forhold: 1) de målte effekter i potteforsøg var meget signifikante og 2) stimulerende effekter af lave doser af en fenpropimorph-propiconazol blanding og af carbendazim er tidligere blevet observeret i markforsøg (Schweiger et al. 2001).
Semisterile vækstmedier blev anvendt i eksperimenter beskrevet i kapitel 4 og kapitel 5 for at kunne kontrollere den initiale forekomst af udvalgte mykorrhiza-svampe isoleret fra forsøgsmarken. Strålesteriliseringens påvirkning af jordens indhold af øvrige mikroorganismer ligger uden for rammerne af dette projekt. Det er dog klart, at mange bakterier overlever bestrålingen, at det anvendte mykorrhizasvampeinokulum indeholder mange mikroorgansimer, samt at bestrålet jord indeholder en stor mængde substrat for bakterievækst og derfor hurtigt bliver rekoloniseret af overlevende og invaderende bakterier (Cawse, 1975).
Disse forhold bidrager til at øge resultaternes relevans i forhold til almindelig dyrkningspraksis. Projektet var i øvrigt ikke rettet mod en afklaring af fungiciders generelle effekt på mykorrhizasvampe under markforhold, men havde fokus på fungicideffekter set i forhold til biologisk kompleksitet. Projektets påvisning af en tydelig gensidig påvirkning mellem de enkelte svampe i simple rekonstruerede samfund leder til konklusionen, at fungicideffekter målt på enkelte mykorrhiza-svampe ikke er brugbare til at forudsige effekter på komplekse marksamfund af mykorrhizasvampe. Resultater af direkte effektmålinger på marksamfund kan på den anden side ikke fortolkes på nuværende tidspunkt, hvor metoder til kvantificering af de enkelte svampe ikke er tilstrækkeligt udviklede.
7.1 Konklusioner
De vigtigste konklusioner fra dette projekt kan opsummeres således:
- Markjordens samfund af mykorrhizasvampe udvikles under påvirkning af svampenes gensidige hæmning eller stimulering i forbindelse med deres kolonisering af planterødder samt af en række eksterne faktorer (jordbundsforhold, genotype og næringsstofstatus for værtplanten, jordens indhold af miljøfremmede stoffer).
- Rodkolonisering med den enkelte mykorrhizasvamp er i høj grad påvirket af egenskaber hos andre svampe i samme samfund. Mekanismerne bag de observerede stimuleringer eller hæmninger er ikke afdækket.
- Små fungiciddoser kan stimulere funktionen af mykorrhizasvampes funktion.
- Disse hormesis-effekter optræder både, når mykorrhizaen er etableret fra en enkelt genotype, og når flere genotyper bidrager. Den hyppige behandling af kartoffelmarker med mancozeb sandsynliggør, at mancozeb-induceret hormesis også kan optræde under markforhold. Den langsigtede betydning af hormesis for mykorrhizasvampes aktivitet, herunder sporeproduktion og overlevelse, er ikke undersøgt.
- Dyrkningshistorie og variation inden for en mark har større indflydelse på diversiteten af mykorrhizasvampesamfundet end behandling med mancozeb eller carbendazim.
- De komplicerede samspil mellem mykorrhizasvampe gør det meget vanskeligt at ekstrapolere fra fungicideffekter på enkelte svampegenotyper til fungicideffekter på samfund af mykorrhizasvampe.
- Det må forventes, at anbefalede markdoser af mancozeb og carbendazim ikke har betydende negative effekter på funktion af mykorrhiza; carbendazim anvendes dog ikke i dansk landbrug. En generel konklusion vedrørende fungiciders indflydelse på mykorrhizasvampe skal baseres på videre studier der omfatter flere fungicider.
7.2 Perspektiver
Projektets resultater tyder ikke på, at anvendelsen af mancozeb i planteavlen har negativ indflydelse på forekomst og funktion af mykorrhizasvampe. Det er dog nødvendigt at søge de opnåede resultater verificeret ved anvendelse af ikke-steriliseret jord med en naturligt forekommende mikroflora, idet de observerede hormesis-effekter er observeret i potteforsøg med strålingssteriliseret jord. En videreførelse af projektet bør inddrage både potteforsøg og markforsøg, og der bør lægges vægt på studier af naturlige samfund af mykorrhiza i kartoffelmarker, der som oftest behandles med store mængder mancozeb. Fremtidige dosis-respons studier bør inddrage et større antal behandlings-doser for at muliggøre en optimal statistisk analyse af responsforløbet, fx ved anvendelse af Gompertz-regression.
Projektets metodemæssige tilgang fremstår som et eksempel for diversitets-funktions studier på andre organismegrupper. Undersøgelsen af effekter af fungicider på flere organisationsniveauer: individ, population og samfund, gjorde det muligt at forklare en del af de observerede effekter. Den anvendte brede tilgang viste tydeligt, at målte effekter af fungicider på komplekse biologiske systemer ikke kan forudsiges på baggrund af undersøgelser i forenklede modelsystemer.
Det har været foreslået at anvende in vitro dyrkning af rødder og mykorrhizasvampe som modelsystemer til en overordnet screening af fungiciders økotoksikologiske effekter (Wan et al., 1998). Metoden er dog behæftet med begrænsninger: dels er det kun få mykorrhizasvampe, der kan etableres i disse systemer, dels vil plantemedierede effekter ikke kunne måles i et biologisk system, der optager det nødvendige kulstof fra glukose opløst i dyrkningsmediet. Det vil dog være oplagt at anvende modelsystemet til at en nærmere eftervisning af hormesis ved tilførsel af små fungiciddoser. Modelsystemer med de mykorrhiza-afhængige Medicago truncatula planter fremstår som et muligt alternativt screeningssystem. Projektet viste således, at væksten af M. truncatula planterne i sig selv gav et temmelig realistisk billede af fungicideffekter på svampene. Det må dog igen understreges, at projektet viste, at effekten af pesticider ikke kan forudsiges, så snart kompleksiteten øges fra en til to svampe.
Der er klart et behov for bedre at forstå de signal- og metabolisme-pathways, der er involveret i mykorrhizadannelse. Denne indsigt er nødvendig for vores bestræbelser på at forstå, hvorfor en svamp kan kolonisere helt forskelligt, når den er sammen med andre end når den er alene. Der er ingen tvivl om, at dette rummer en meget stor grad af kompleksitet, og at det eksperimentelt vil være en stor udfordring.
8 Referencer
Anon. 2001. Bekæmpelsesmiddelstatistik 2000. Orientering fra Miljøstyrelsen.
Anon. 2005. Bekæmpelsesmiddelstatistik 2004. Orientering fra Miljøstyrelsen.
Bødker L, Kjøller R, Kristensen K, Rosendahl S. 2002. Interactions between indigenous arbuscular mycorrhizal fungi and Aphanomyces euteiches in field-grown pea. Mycorrhiza 12: 7-12.
Bødker L, Kjøller R, Rosendahl S. 1998. Effect of phosphate and the arbuscular mycorrhizal fungus Glomus intraradices on disease severity of root rot of peas (Pisum sativum) caused by Aphanomyces euteiches. Mycorrhiza 8: 169-174.
Calabrese EJ. 2005. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environmental Pollution 138: 378-411.
Calabrese EJ, Baldwin LA. 2003. Toxicology rethinks its central belief - Hormesis demands a reappraisal of the way risks are assessed. Nature 421: 691-692.
Cawse PA. 1975. Microbiology and biochemistry of irradiated soils. In Soil Biochemistry, Vol 3, (EA Paul, AD McLaren, Eds), pp. 213-267. Marcel Dekker, New York.
Christensen P, Helweg A. 2003. Forventelige pesticidkoncentrationer i jord efter erhvervsmæssig pesticidanvendelse. Miljøstyrelsen, Miljøprojekt No. 844, 61pp.
Dodd JC, Jeffries P. 1989. Effect of fungicides on three vesicular-arbuscular mycorrhizal fungi associated with winter wheat (Triticum aestivum L.). Biology and Fertility of Soils 7: 120-128.
Gordon TR, Okamoto D, Milgroom MG. 1992. The structure and interrelationship of fungal populations in native and cultivated soils. Molecular Ecology 1: 241-249.
Fracchia S, Garcia-Romera I, Godeas A, Ocampo JA. 2000. Effect of the saprophytic fungus Fusarium oxysporum on arbuscular mycorrhizal colonization and growth of plants in greenhouse and field trials. Plant and Soil 223: 175-184
Garcia-Romera I, Garcia-Garrido JM, Martin J, Fracchia S, Mujica MT, Godeas A, Ocampo JA. 1998. Interactions between Saprotrophic Fusarium strains and arbuscular mycorrhizas of soybean plants. Symbiosis 24: 235-245
Groth DE, Martinson CA. 1983. Increased Endomycorrhizal Infection of Maize and Soybeans After Soil Treatment and Metalaxyl. Plant Disease 67: 1377-1378.
Helgason T, Merryweather JW, Denison J, Wilson P, Young JPW, Fitter AH. 2002. Selectivity and functional diversity in arbuscular mycorrhizas of co-occurring fungi and plants from a temperate deciduous woodland. Journal of Ecology 90: 371-384.
Hepper CM, Azcon-Aguilar C, Rosendahl S, Sen R. 1988. Competition between 3 species of Glomus used as spatially separated introduced and indigenous mycorrhizal inocula for leek (Allium-porrum L). New Phytologist 110: 207-215.
Jacobsen OS, Bossi R. 1997. Degradation of ethylenethiourea (ETU) in oxic and anoxic sandy aquifers. FEMS Microbiology Reviews 20: 539-544.
Jakobsen I, Rosendahl S. 1993. Planter har ikke rødder - de har mykorrhiza! Naturens Verden 9: 329-336.
Jakobsen I, Abbott LK, Robson AD. 1992. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum. 1: Spread of hyphae and phosphorus inflow into roots. New Phytologist 120: 371-380.
Jensen PK, Spliid, NH. 2003.Deposition of pesticides on the soil surface. Miljøstyrelsen, Pesticides Research No.65, 60pp.
Kahiluoto H, Vestberg M. 2000. Creation of a non-mycorrhizal control for a bioassay of AM effectiveness 2. Benomyl application and soil sampling time. Mycorrhiza 9: 259-270.
Kjøller R, Rosendahl S. 2000a. Detection of arbuscular mycorrhizal fungi (Glomales) in roots by nested PCR and SSCP (Single Stranded Conformation Polymorphism). Plant and Soil 226: 189-196.
Kjøller R, Rosendahl S. 2000b. Effects of fungicides on arbuscular mycorrhizal fungi: differential responses in alkaline phosphatase activity of external and internal hyphae. Biology and Fertility of Soils 31: 361-365.
Kjoller R, Rosendahl S. 2001. Molecular diversity of glomalean (arbuscular mycorrhizal) fungi determined as distinct Glomus specific DNA sequences from roots of field grown peas. Mycological Research 105: 1027-1032.
McAllister CB, Garcia-Garrido JM, Garcia-Romera I, Godeas A, Ocampo JA. 1997. Interaction between Alternaria alternata or Fusarium equiseti and Glomus mosseae and its effects on plant growth. Biology and Fertility of Soils 24: 301-305.
Michelsen A, Rosendahl S. 1990. The effect of VA mycorrhizal fungi, phosphorus and drought stress on the growth of Acacia nilotica and Leucaena leucocephala seedlings. Plant and Soil 124: 7-13.
Munkvold L, Kjøller R, Vestberg M, Rosendahl S, Jakobsen I. 2004. High functional diversity within species of arbuscular mycorrhizal fungi. New Phytologist 164: 357-364.
Nei M. 1973. Analysis of Gene Diversity in Subdivided Populations. Proceedings of the National Academy of Sciences of the United States of America 70: 3321-3323.
Newman EI. 1966. A method for estimating the total length of root in a sample. Journal of Applied Ecology 3: 139-145.
Nielsen JS, Joner EJ, Declerck S, Olsson S, Jakobsen I. 2002. Phospho-imaging as a tool for visualisation and nonnvasive measurement of P transport dynamics in arbuscular mycorrhizas. New Phytologist 154: 809-820.
Oldroyd GED, Harrison MJ, Udvardi M. 2005. Peace talks and trade deals. Keys to long-term harmony in legume-microbe symbioses. Plant Physiology 137: 1205-1210.
Pearson JN, Jakobsen I. 1993. The relative contribution of hyphae and roots to phosphorus uptake by arbuscular mycorrhizal plants, measured by dual labelling with P-32 and P-33. New Phytologist 124: 489-494.
Probst M, Berenzen N, Lentzen-Godding A, Schulz R. 2005. Scenario-based simulation of runoff-related pesticide entries into small streams on a landscape level. Ecotoxicology and Environmental Safety 62: 145-159.
Rosendahl S. 1985. Interactions between the vesicular-arbuscular mycorrhizal fungus Glomus fasciculatum and Aphanomyces euteiches root-rot of peas. Phytopathologische Zeitschrift-Journal of Phytopathology 114: 31-40.
Rosendahl S, Taylor JW. 1997. Development of multiple genetic markers for studies of genetic variation in arbuscular mycorrhizal fungi using AFLP(TM). Molecular Ecology 6: 821-829.
Schreiner RP, Bethlenfalvay GJ. 1997. Plant and soil response to single and mixed species of arbuscular mycorrhizal fungi under fungicide stress. Applied Soil Ecology 7: 93-102.
Schweiger PF, Jakobsen I. 1998. Dose-response relationships between four pesticides and phosphorus uptake by hyphae of arbuscular mycorrhizas. Soil Biology & Biochemistry 30: 1415-1422.
Schweiger PF, Jakobsen I. 1999. Direct measurement of arbuscular mycorrhizal phosphorus uptake into field-grown winter wheat. Agronomy Journal 91: 998-1002.
Schweiger PF, Spliid NH, Jakobsen I. 2001. Fungicide application and phosphorus uptake by hyphae of arbuscular mycorrhizal fungi into field-grown peas. Soil Biology & Biochemistry 33: 1231-1237.
Schweiger PF, Thingstrup I, Jakobsen I. 1999. Comparison of two test systems for measuring plant phosphorus uptake via arbuscular mycorrhizal fungi. Mycorrhiza 8: 207-213.
Smith SE, Smith FA, Jakobsen I. 2003. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiology 133: 16-20.
Stukenbrock EH, Rosendahl S. 2005a. Clonal diversity and population genetic structure of arbuscular mycorrhizal fungi (Glomus spp.) studied by multilocus genotyping of single spores. Molecular Ecology 14: 743-752.
Stukenbrock EH, Rosendahl S. 2005b. Development and amplification of multiple co-dominant genetic markers from single spores of arbuscular mycorrhizal fungi by nested multiplex PCR. Fungal Genetics and Biology 42: 73-80.
Thingstrup I, Rubaek G, Sibbesen E, Jakobsen I. 1998. Flax (Linum usitatissimum L.) depends on arbuscular mycorrhizal fungi for growth and P uptake at intermediate but not high soil P levels in the field. Plant and Soil 203: 37-46.
Thingstrup I, Kahiluoto H, Jakobsen I. 2000. Phosphate transport by hyphae of field communities of arbuscular mycorrhizal fungi at two levels of P fertilization. Plant and Soil 221: 181-187.
Trappe JM, Molina R, Castellano M. 1984. Reactions of mycorrhizal fungi and mycorrhiza formation to pesticides. Annual Review of Phytopathology 22: 331-359.
Trewavas A, Stewart D. 2003. Paradoxial effect of chemicals in the diet on health. Current Opinion in Plant Biology 6: 185-190.
van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR. 1998. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69-72.
Wan MT, Rahe JE, Watts RG. 1998. A new technique for determining the sublethal toxicity of pesticides to the vesicular-arbuscular mycorrhizal fungus Glomus intraradices. Environmental Toxicology and Chemistry 17: 1421-1428.
Version 1.0 September 2006 • © Miljøstyrelsen.