Miljøprojekt, 1120
Sandsynlige konsekvenser af klimaændringer på artsudbredelser og biodiversitet i Danmark
Indholdsfortegnelse
- 2.1 Bioklimatisk habitat-modellering
- 2.2 Beregning af effekten af klimaændringer i Danmark
- 2.3 Fordele og ulemper ved den anvendte metode
- 2.4 Valg af arter
- 3.1 Udvalgte artseksempler
- 3.2 Diversitet
- 3.3 Geografisk variation i Danmark
- 3.4 Forskelle inden for naturtyperne
- 4.1 Hvor troværdig er den bioklimatiske habitat-modellering?
- 4.2 Sandsynlighed for tab af arter
- 4.3 Indvandring af nye arter
- 4.4 Er arterne repræsentative for dansk biodiversitet?
- 4.5 Anbefalinger
Forord
Denne rapport er udarbejdet i forbindelse med Regeringens beslutning om at igangsætte et udredningsarbejde om Danmarks tilpasning til fremtidens klima.
Projektet er foregået som et samarbejde mellem Afdeling for Vildtbiologi og Biodiversitet, DMU og Biologisk Institut, Århus Universitet. Fra DMU har Flemming Skov deltaget. Fra Århus Universitet har Jens-Christian Svenning og Signe Normand deltaget.
Der har været nedsat en følgegruppe bestående af: Ditte Holse, Miljøstyrelsen og Hans Erik Svart, Skov- og Naturstyrelsen.
Sammenfatning og konklusioner
I denne rapport analyseres de potentielle konsekvenser af de klimaændringer, der forudsiges frem til år 2100 ifølge klimascenarierne A2 og B2 fra FN’s klimapanel (IPCC), for udbredelsen af 104 udvalgte arter af karplanter i Danmark. De udvalgte arter omfatter 88 såkaldt karakteristiske arter under EU’s Habitatdirektiv. Disse arter repræsenterer et bredt spektrum af danske vegetationstyper fra klit og overdrev over eng og mose til skov. Derudover analyseres de potentielle konsekvenser for de to danske plantearter, der står på Habitatdirektivets Bilag 2, samt for en række arter fra Mellem- og Sydeuropa med henblik på at vurdere i hvilket omfang, klimaændringerne vil give mulighed for sydlige arters indvandring.
Ved hjælp af en bioklimatisk habitatmodel beregnes hver arts potentielle udbredelsesområde i Europa nu og under klimascenarierne B2 og A2. Resultaterne sammenholdes og for Danmark beregnes det, hvorvidt ændringerne er negative, neutrale eller positive.
Resultaterne i denne rapport tyder på, at klimaændringerne over det 21. århundrede under både A2- og B2-scenarierne må forventes at have en overvejende negativ effekt på de undersøgte arter og sandsynligvis på den botaniske biodiversitet i Danmark som helhed. Resultaterne viser, at under klimascenarium B2 forventes det, at 67% af arterne vil påvirkes negativt og 12% positivt. For 21% af arterne er påvirkningen neutral. For klimascenarium A2 er de tilsvarende tal 78%, 10% og 12%.
Inden for Danmarks grænser varierer den forventede påvirkning imidlertid betydeligt. Generelt påvirkes den sydøstlige del af landet langt mere negativt end den vestlige del, hvor negative og positive påvirkninger stort set ophæver hinanden.
Der er forskel, dog ikke statistisk signifikant, på, hvor meget arter tilhørende forskellige naturtyper påvirkes. Tallene tyder dog på, at arter, der tilhører ferske enge og især høj- og lavmoser vil være mest sårbare overfor den globale opvarmning.
Resultaterne viser, at hhv. 4% og 7% af de 84 hjemmehørende karakteristiske arter helt vil miste passende klimatiske forhold under B2- og A2-scenariet. Det vurderes sandsynligt, at disse arter over tid vil uddø i Danmark under disse scenarier. I alt forventes hhv. 6% og 15% af de hjemmehørende karakteristiske arter at blive udsat for klimatiske forhold under B2- og A2-scenariet, der er mere end 80% dårligere end dem, de lever under i Danmark i dag. Disse arters udbredelse i Danmark må forventes at komme under pres.
Spredning forventes at blive en særdeles vigtig begrænsende faktor for planters positive udbredelsesmæssige respons på fremtidige klimaændringer. De fleste arter har ikke en spredningskapacitet, der sætter dem i stand til at følge klimaændringer af den hastighed og styrke, som beskrives i de brugte klimascenarier for det 21. århundrede. Den udstrakte menneskeskabte fragmentering af de naturlige habitater i Danmark og det meste af det øvrige Europa forstærker dette problem.
I rapporten konkluderes det, at man med nogen forsigtighed godt kan bruge karplanter som indikatorer for følsomheden af den generelle biodiversitet overfor klimaændringer. Det anbefales at udarbejde en rullende klimatilpasningsstrategi baseret på en overvågning af naturtyper og arter. Denne overvågning kunne bl.a. basere sig på det nye NOVANA-program, hvor et stort antal habitattyper og arter overvåges. Ved nøje at følge frem- og tilbagegang for hjemmehørende og nyindvandrede arter, kan denne overvågning anvendes som et afsæt for egentlige handlingsplaner.
Summary and conclusions
The present report analyses the potential consequences of the climatic changes predicted to have occurred by year 2100 according to the A2 and B2 climate scenarios produced by the UN Intergovernmental Panel on Climate Change (IPCC) for 104 selected vascular plant species in Denmark. The chosen species include 88 so-called characteristic species under the EU Habitats Directive. These species represent a broad span of Danish vegetation types, from dunes and dry grasslands to meadows, bogs, and forests. In addition, the potential consequences are analysed for the two Danish plant species on the Habitats Directive’s annex 2, and for a suite of species from Central and Southern Europe, in order to evaluate to which degree the climatic changes might allow southern species to invade.
Using a bioclimatic habitat model we compute the potential distribution of each species under current conditions and under the B2 and A2 scenarios. Combining the results, we compute whether the changes in Denmark will be negative, neutral or positive.
The results in the present report suggest that 21st century climate change, as represented by the B2 and A2 scenarios, must be expected to have a predominantly negative effect on the study species and probably also on the floristic diversity in Denmark in general. The results show that 67% of the species are expected to be negatively affected and 12% positively affected under the B2 scenario. For 21% of the species the impact will be neutral. Under the A2 scenario the corresponding figures are 78%, 10%, and 12%.
The expected impact varies considerably within Denmark. Generally, the south-eastern part of the country is more negatively affected than the western part, where negative and positive impacts largely balance out.
The impact also differ, albeit not significantly, between species characterising different habitat types. The figures suggest that species belonging to humid meadows and mesophile grasslands and, in particular, raised bogs, mires and fens will be most vulnerable to global warming.
The results indicate that 4% and 7% of the 84 native characteristic species will completely loose suitable climatic conditions under the B2 and A2 scenario, respectively. It is suggested that these species eventually would go extinct in Denmark. A total of 6% and 15% of the native characteristic species can be expected to experience climatic conditions under the B2 and A2 scenario, respectively, which are 80% less suitable than those they occur under in Denmark today. The occurrence of these species in Denmark would then be expected to come under pressure.
Dispersal is expected to become a particularly important constraint on the positive range response of plant species to future climatic changes. Most species will not have sufficient dispersal capacity to track climatic changes of the speed and strength described in the 21st century climate scenarios. The extensive anthropogenic fragmentation of the natural habitats in Denmark and most of Europe exacerbates this problem.
It is concluded that it, with some care, is possible to use vascular plants as indicators of the climate change sensitivity of biodiversity in general. It is recommended to develop a progressive climate adaptation strategy based on the monitoring of habitat types and species. Such a monitoring could at least partly be based on the new NOVANA program, which monitors a large number of habitat types and species. By careful monitoring the expansions and declines of the long-term native as well as newly arrived species it becomes possible to use this monitoring as a basis for developing plans of actions.
1 Introduktion
Det forventes, at globale klimaændringer over de næste 100 år vil ændre levevilkårene for planter og dyr i Danmark og dermed påvirke den biologiske sammensætning af økosystemer og i sidste ende biodiversiteten. Nogle arter vil få sværere ved at klare sig i Danmark og vil måske helt forsvinde, mens andre kan få nye muligheder for at trives og eventuelt sprede sig.
Denne rapport har til formål at vurdere den mulige effekt af de globale klimaændringer på udvalgte danske plantearter. Denne viden er nødvendig for at kunne lave en langsigtet planlægning af naturbeskyttelsen (Hannah et al. 2002). Rapporten vil også give et skøn over i hvilket omfang, varmekrævende arter vil få mulighed for at etablere sig i den danske natur. Udredningen vil ydermere indeholde en generel vurdering af behovet for en øget indsats i naturforvaltningen, herunder naturgenopretning, for at sikre den biologiske mangfoldighed i Danmark samt de danske skoves robusthed.
Analyserne i rapporten bygger på en fremskrivning af klimaet over det næste århundrede baseret på klimascenarier udarbejdet af det internationale klimapanel Intergovernmental Panel on Climate Change (IPCC 2001). IPCC blev dannet i 1988 på initiativ af FN’s verdensorganisation for meteorologi (WMO) og FN’s miljøprogram (UNEP). IPCC har foreslået fire samfundsøkonomiske scenarier, der beskriver mulige globale udviklingslinier mht. befolkningstal, økonomisk vækst og teknologisk udvikling. Med udgangspunkt i disse scenarier kan udledningen af drivhusgasser og andre klimapåvirkende faktorer beregnes. Disse data indgår igen i komplicerede klimamodeller, der beregner, hvilket klima vi kan forvente op gennem det næste århundrede. Det er ikke muligt at vurdere sandsynligheden for de fire klimascenarier, og IPCC anbefaler, at der anvendes mindst to scenarier for at belyse det mulige udfaldsrum for det fremtidige klima.
1.1 Klimascenarier
De to scenarier, der anvendes i denne rapport, betegnes hhv. A2 og B2 (IPCC, 2001){IPCC, 2001 1233 /id}{IPCC, 2001 1233 /id}{IPCC, 2001 1233 /id}. A2 fører i 2100 til en relativt kraftig global opvarmning med en gennemsnitlig temperaturstigning på 2,8–4,8°C i forhold til 1990. B2 repræsenterer et mindre dramatisk scenarium, hvor det forudses, at den globale temperatur stiger 1,9–3,4°C i forhold til 1990. Tidligere undersøgelser har vist, at klimaet under scenarium A2 vil ændre sig så meget, at det klimatiske optimum for en række skovbundsurter vil flytte sig 700-1400 km mod nord i Europa (Skov & Svenning 2004). Hvilken indflydelse sådanne forskydninger reelt vil få for den enkelte arts udbredelse i Danmark vil afhænge af mange faktorer, bl.a. artens klimatiske tolerance, dens spredningsevne, dens krav til habitat, respons på eutrofiering mm.
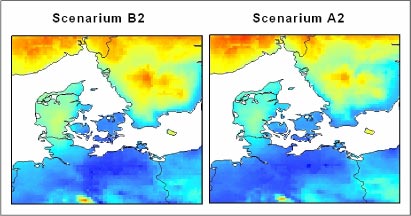
For bedre at illustrere hvilket klima vi kan forvente i Danmark om 100 år, viser Figur 1.1, hvor i Europa vi i dag finder et klima, der svarer til fremskrivningerne for Danmark i år 2100 under de to scenarier
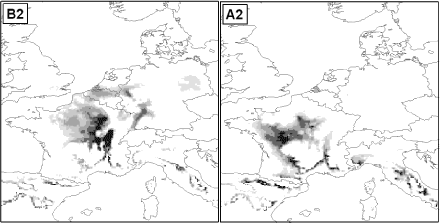
Figur 1.1 Hvor ”kommer” fremtidens klima fra? De to kort viser, hvor der i dag findes et klima, der svarer til hhv. B2 og A2 scenarierne i Danmark i år 2100. Jo mørkere gråtone, jo mere ligner klimaet, fremtidens danske klima under de to scenarier.
Kortene viser, at under det relativt milde B2 scenarium kan vi forvente et klima i år 2100, som vi i dag hovedsageligt finder i det nordlige og centrale Frankrig. Under A2 scenariet kan vi forvente et klima, som man i nutiden har i det centrale og sydlige Frankrig samt i bjergegne i det nordlige Spanien og i Italien.
1.2 Artsdiversitetsmønstre i Europa
Hvilken artsdiversitet finder man så i disse områder; og kan vi forvente, at Danmark vil opleve en tilsvarende stigning eller et tilsvarende fald i diversitet? Kortet i Figur 1.2 viser planternes artsrigdom i Europa. Kortet er baseret på Atlas Florae Europaeae (www.fmnh.helsinki.fi/english/botany/afe/index.htm), der på nuværende tidspunkt omfatter udbredelseskort for mere end 4.000 europæiske plantearter, -underarter og -varieteter. Kortet viser, at vi i dag finder den største diversitet af planter i et område langs de sydlige skråninger af Alperne, ved Pyrenæerne og i Balkanområdet. Sammenholdes dette med kortene på Figur 1.1 ses det, at de områder der i dag har et klima svarende til det, der er forudsagt for Danmark i år 2100, har en artsrigdom af samme størrelse eller lidt større end den, vi har i Danmark i dag.
Dette betyder imidlertid ikke, at der ikke vil ske noget med den danske flora. En del arter er fælles for de to områder, men mange er forskellige og det endelige resultat af klimaændringer vil afhænge af mange faktorer, bl.a. hvorvidt arterne vil være i stand til at overleve eller tilpasse sig klimaændringerne på stedet, eller sprede sig i et tempo, der svarer til den rate, hvormed klimaet ændrer sig.
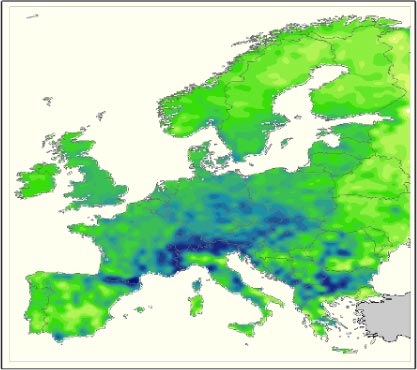
Figur 1.2 Fordelingen af artsrigdom for planter i Europa. Farverne indikerer antallet af arter for hvert område på 50x50 km, der er undersøgt. Skala: fra gul (0-100 arter) over grøn (100-400 arter) til mørkeblå (400-700 arter).
I denne rapport vil denne problemstilling blive belyst gennem bioklimatisk habitat-modellering af en række arters potentielle udbredelse nu og under de to valgte klimascenarier.
2 Metoder
- 2.1 Bioklimatisk habitat-modellering
- 2.2 Beregning af effekten af klimaændringer i Danmark
- 2.3 Fordele og ulemper ved den anvendte metode
- 2.4 Valg af arter
Udbredelsen af plantearter styres af en række forskellige faktorer, der vekselvirker på forskellig skala i tid og rum. Disse faktorer deles typisk i tre grupper: (1) Abiotiske faktorer, dvs. omgivelsernes fysisk-kemiske beskaffenhed. (2) Biotiske faktorer, f.eks. konkurrence med andre planter eller samspil med bestøvere eller skadedyr. (3) Historiske faktorer, dvs. fortidige hændelser og processer, hvor virkningerne akkumulerer over tid. Blandt de historiske faktorer, der regnes for vigtige for den geografiske fordeling af plantearter, er bl.a. effekter af istider og andre fortidige klimaændringer, evolutionære processer, samt, især på mindre geografisk skala, menneskets fortidige påvirkning af naturen.
På større skala anses klima for langt den vigtigste plantefordelende faktor (Hawkins et al. 2003). Klimaets betydning kommer da også tydeligt til udtryk, når man betragter klodens vegetationszoner (tropisk regnskov, savanne, ørken, tempereret skov osv.), hvis udbredelser alle er tæt koblede til de overordnede klimamønstre. Der er selvfølgelig mange andre abiotiske faktorer med stor indvirkning på fordelingen af planter. Topografi, jordbundens struktur og dens indhold af næringsstoffer og vand kan have stor betydning. Det samme gælder en lang række biotiske faktorer. Disse faktorer opererer dog primært på mindre geografisk skala (= 200 km eller endog =10 km) end der normalt anvendes i forbindelse med modellering af klimaets betydning for artsudbredelser (Pearson & Dawson 2003).
2.1 Bioklimatisk habitat-modellering
Bioklimatisk habitat-modellering er en fællesbetegnelse for en række forskellige metoder, der dog alle tager udgangspunkt i den observerede udbredelse af en art. De fleste metoder arbejder ud fra den antagelse, at artens udbredelse har opnået en vis grad af ligevægt med klimaet, dvs. at klimaet således i betydeligt omfang afgrænser udbredelsen. Den observerede udbredelse kan derfor bruges til at beregne artens klimatiske ’rum’ eller niche (’bioclimatic envelope’). En oversigt over de forskellige metoder kan findes i Guisan & Zimmermann (2000).
Der er mange måder at beskrive den klimatiske niche på, men i langt de fleste studier indgår mål for temperatur og fugtighed. I denne rapport anvendes tre klimatiske variable af velkendt betydning for mange plantearters udbredelser (Figur 2.1):
Graddage over 5°C (Growing Degree Days el. GDD) – Dette er et mål for den mængde energi, som planterne har til rådighed for vækst.
Absolut minimums-temperatur – Mange planters udbredelse begrænses af kuldestress, og den absolutte minimums-temperatur er et godt mål herfor.
Vandbalance – Dette indeks angiver forskellen mellem nedbør og potentiel fordampning (evapotranspiration) opsummeret måned for måned over året. Negative værdier angiver et underskud af vand, mens positive værdier viser, at nedbøren overstiger den potentielle evapotranspiration.
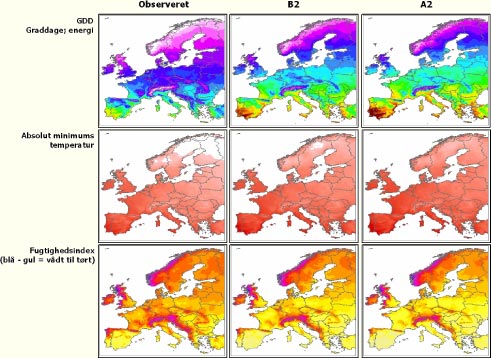
Figur 2.1 Kortene viser de tre anvendte klimavariable i Europa. Den første kolonne viser de observerede klimadata (perioden 1961-1991) og de to næste kolonner værdierne for klimascenarierne B2 og A2 i år 2100.
Den anvendte metode beregner hver arts minimum, optimum og maksimum i forhold til hver af de tre klimavariable. Modellens output er en kontinuert værdi mellem 0 og 1, der angiver hvor egnet klimaet er for arten (herefter kaldet klima-egnethed). Værdien 1 opnås hvis alle tre klimavariable ligger inden for deres optimum, mens værdien 0 fås, hvis blot én af de tre variable ligger uden for de beregnede minimums- og maksimumsværdier. Jo tættere værdien ligger på 1, jo mere egnet anses klimaet at være for den pågældende art. Yderligere detaljer om metoden kan findes i Skov & Svenning (2004) og Svenning & Skov (2004).
Ved hjælp af et Geografisk Informations System (GIS) kan man beregne og visualisere en arts potentielle nutidige udbredelse, dvs. hvor arten klimatisk set vil kunne trives i dag ifølge den bioklimatiske habitat-model, og dens potentielle udbredelse i år 2100 under klimascenarierne B2 og A2 (Figur 2.2).
Det er vigtigt at slå fast, at de beregnede værdier kun viser i hvilke områder, der findes et egnet klima for arten, ikke hvor man aktuelt finder den. Der kan som nævnt være mange årsager til, at en art ikke findes på en given lokalitet, selvom klimaet er passende (Pearson & Dawson 2003; Svenning & Skov 2004).
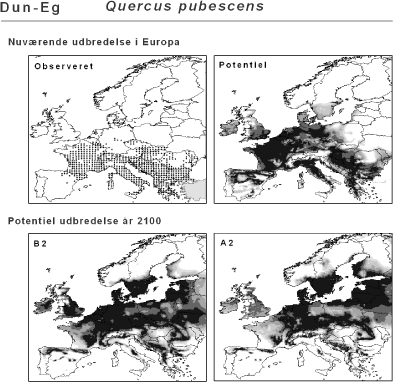
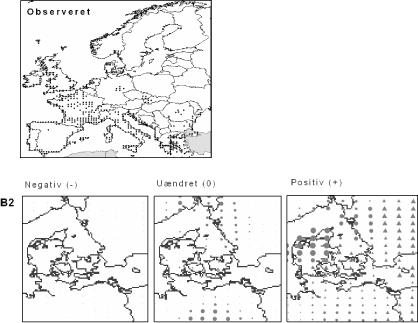
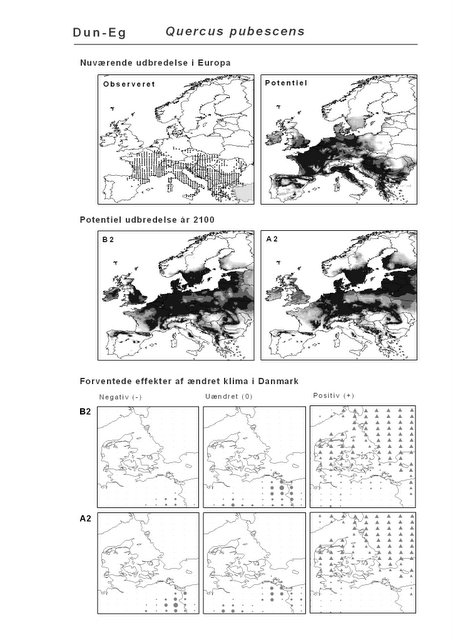
Figur 2.2 Eksempel på en bioklimatisk modellering. Kortudsnittet øverst til venstre viser den observerede udbredelse (sorte prikker). Disse data er anvendt til at beregne den klimatiske niche, som derefter er brugt til at beregne artens potentielle udbredelse under hhv. det nuværende klima og de to klimascenarier. Dun-Eg er en tørketålende, varmekrævende, men løvfældende træart, der er vigtig i de sydeuropæiske skove.
2.2 Beregning af effekten af klimaændringer i Danmark
For hver art vurderes den sandsynlige respons på de forudsagte klimaændringer i forskellige områder af Danmark (Figur 2.3):
- Positiv effekt – Arten forekommer i eller i umiddelbar nærhed af området og vil få væsentlig bedre klimatiske betingelser.
- Neutral/uændret effekt – Arten vil ikke få væsentligt ændrede klimatiske betingelser i Danmark. Det er vigtigt at bemærke, at arter der ikke forekommer i området i dag, og ifølge modelleringerne heller ikke vil komme til det i år 2100, også opfattes som neutralt påvirkede.
- Negativ effekt – Arten forekommer i området, men vil få væsentligt dårligere klimatiske betingelser end i dag.
- Potentiel positiv effekt – Arten forekommer ikke i nærheden af området nu, men vil potentielt kunne trives der efter en klimaændring, såfremt den har mulighed for at spredes dertil samt at egnede habitater forefindes.
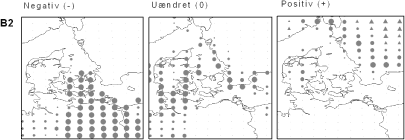
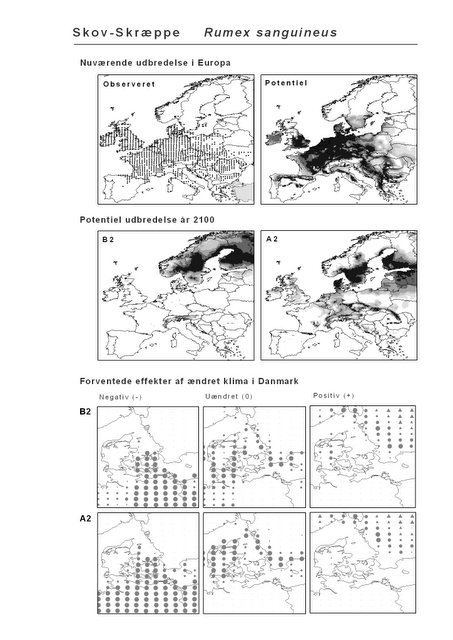
Figur 2.3 Forventede effekter af ændret klima i Danmark på Skov-Skræppe (Rumex sanguineus) under klimascenarium B2. Skov-Skræppe er almindelig i østdanske løvskove på næringsrig muldbund. Kortet tv. viser, hvor i Danmark arten vil få væsentligt dårlige betingelser, end den har i dag. Prikkernes størrelse angiver relativt, hvor meget dårligere betingelserne bliver. Kortet i midten viser, hvor klimaændringerne ikke vil få den store indflydelse på artens forekomst. Endelig viser kortet th., hvor klimaændringerne vil have en forventet positiv indflydelse på Skov-Skræppe. En trekantet signatur angiver potentiel positiv effekt, dvs. en positiv effekt langt fra artens nuværende udbredelsesområde.
2.3 Fordele og ulemper ved den anvendte metode
Den valgte metode har en række fortrin og begrænsninger, som det er vigtigt at kende, når resultaterne skal vurderes. For det første forudsætter modellen en betydelig ligevægt mellem klima og nutidig udbredelse. Denne betingelse er ikke altid opfyldt. F.eks. har Svenning og Skov (2004) vist, at en lang række træarter stadig ikke opfylder hele det område i Europa, deres klimatiske niche tillader; sandsynligvis på grund af langsom spredning ud fra deres istidsrefugier (små områder i Sydeuropa, hvor de overlevede sidste istids kulde og tørke). For det andet er modellen statisk og indeholder derfor ingen eksplicit håndtering af tidsmæssige forløb, f.eks. spredningsforløb. For det tredje tager modellen ikke højde for, om planterne ville kunne tilpasse sig det ændrede klima. Hertil skal det dog bemærkes, at modellen baserer sig på en given arts fulde europæiske udbredelse og derfor må forventes nærmere at overestimere end at underestimere den lokale tilpasningsevne.
Disse mangler til trods, vurderer vi, at den valgte model på indeværende tidspunkt er det bedste eksisterende redskab til vurdering af, hvordan det 21. århundredes klimaændringer vil kunne påvirke den danske floras mangfoldighed og de enkelte arters udbredelser. Selvom det er ønskeligt at udvikle dynamiske modeller, der også inddrager spredning og konkurrence, er de eksisterende modeller, der gør det, endnu under udvikling og langt fra operationelle, hvis de skal være velfunderede empirisk set. Den brugte model har også det store fortrin, at den har en relativt simpel opbygning, der stiller små krav til datainput, hvilket gør det muligt at analysere et stort antal arter. Metoden anvendes da også generelt, som et bedste ’første bud’ når der laves konsekvensvurderinger i forbindelse med klimaændringer og biodiversitet (f.eks. Thuiller et al. 2005; Thomas et al. 2004). Se Pearson & Dawson (2003) og Skov & Svenning (2004) for yderligere diskussion af metoden.
2.4 Valg af arter
Udvalget af arter er primært baseret på Habitatdirektivets liste over karakteristiske arter (Søgaard et al. 2003). Disse arter repræsenterer for det første et bredt udsnit af danske naturtyper og omfatter arter med meget forskellig hyppighed, udbredelse og økologi. I alt modelleres udbredelsen af 88 karakteristiske arter (se Appendiks I), heraf fire, der forekommer i Danmark, men ikke er hjemmehørende (Alnus incana, Picea abies, Salix alba og Salix fragilis).
| Hav og strand | Klitter | Søer og vandløb | Overdrev | Ferske enge | Lav- og højmoser | Klipper og huler | Skove |
| 20 | 6 | 9 | 8 | 10 | 5 | 7 | 29 |
Tabel 2.1 Fordelingen af karakteristiske arter på naturtyper. Bemærk, at summen overstiger 88, da en del arter kan være karakteristiske for mere end én naturtype.
I forbindelse med modelleringen er det kun muligt at anvende arter, der er kortlagt i Atlas Florae Europaeae (AFE). Dette atlasprojekt er imidlertid ikke færdiggjort, og det har derfor ikke været muligt at modellere vigtige grupper som f.eks. græsser, halvgræsser, kurve- og skærmblomstrede arter.
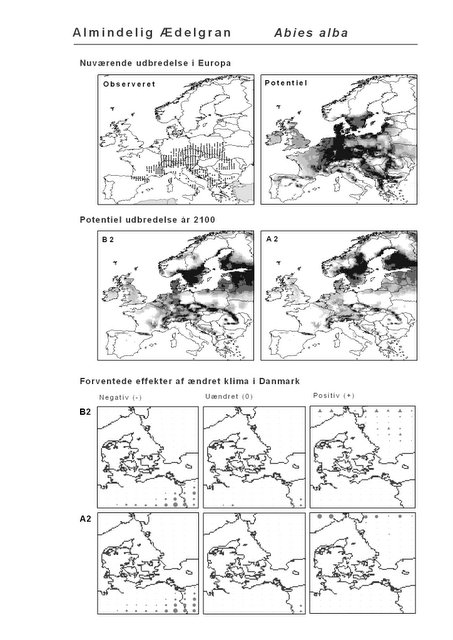
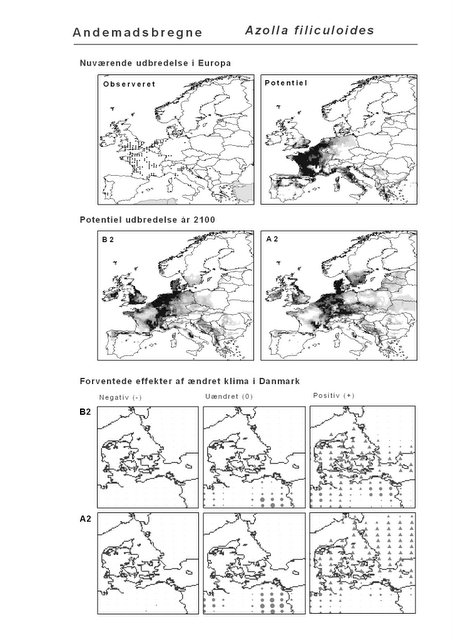
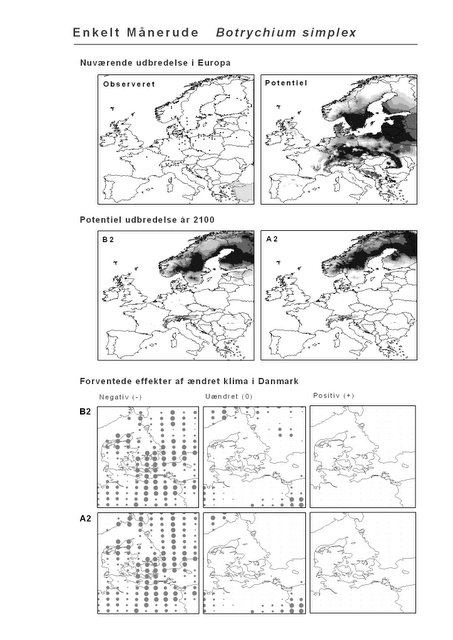
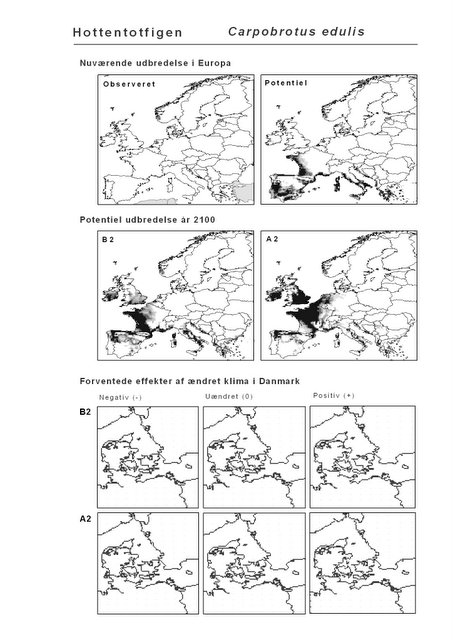
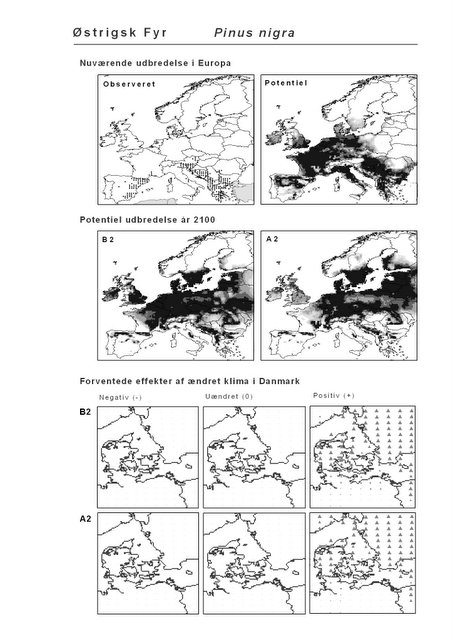
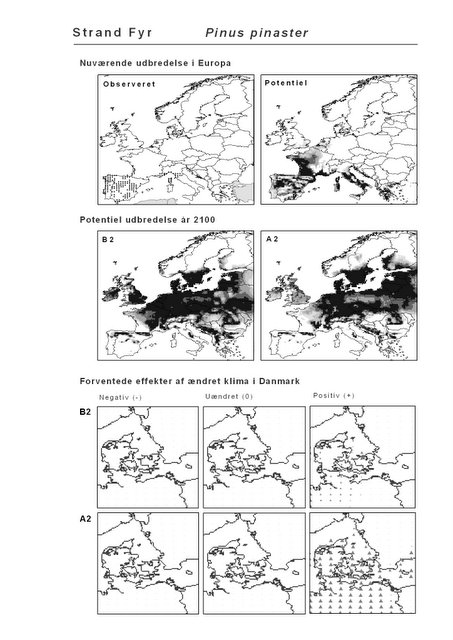
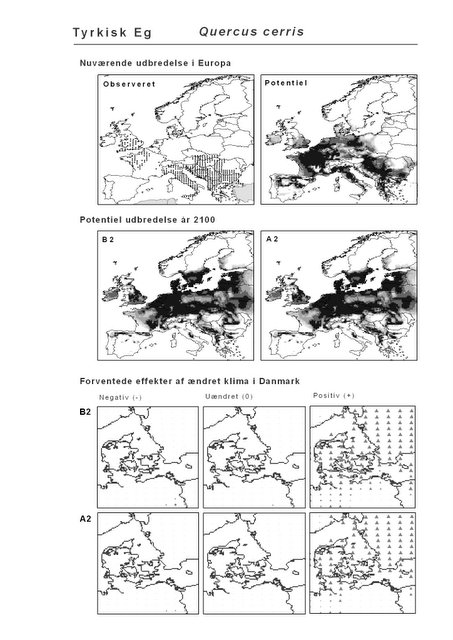
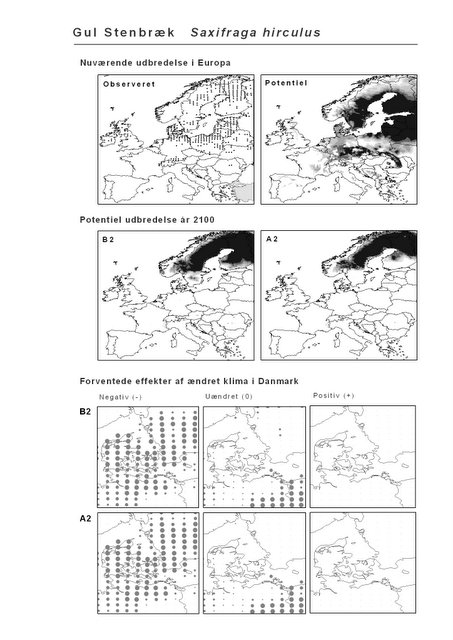
Udover de 88 karakteristiske arter indgår 16 andre arter i undersøgelsen. Det drejer sig dels om to arter fra Habitatdirektivets Bilag 2, nemlig Enkelt Månerude (Botrychium simplex) og Gul Stenbræk (Saxifraga hirculus) samt hjemmehørende danske træer, der er kortlagt i AFE, men som ikke optræder på listen over karakteristiske arter. Endelig modelleres der også en række arter, der ikke vokser i Danmark nu, men hvor en klimaændring kan forventes at gøre det muligt i fremtiden. Denne liste kunne selvsagt blive meget lang, men der er her valgt at fokusere på et begrænset sæt eksempler på arter, der enten har vist potentiale som invasive arter sydpå i Europa eller træer, der måske vil kunne indgå i danske skove i en varmere fremtid. Det drejer sig om vandplanten Andemadsbregne (Azolla filiculoides) og den lave sukkulent Hottentotfigen (Carpobrotus edulis), der oprindelig kommer fra hhv. fra Nordamerika og Sydafrika, samt de hjemmehørende mellem- og sydeuropæiske træarter Østrigsk Fyr (Pinus nigra), Strand-Fyr (Pinus pinaster), Laurbær (Laurus nobilis), Almindelig Valnød (Juglans regia), Frynse-Eg (Quercus cerris), Dun-Eg (Quercus pubescens), Sten-Eg (Quercus ilex) og Ægte Kastanje (Castanea sativa). Herudover er Alm. Ædelgran (Abies alba) medtaget. Dels fordi den er almindelig plantet i de danske skove, dels fordi den er et eksempel på en sydlig art, der især hører til i bjergrige områder.
3 Resultater
- 3.1 Udvalgte artseksempler
- 3.2 Diversitet
- 3.3 Geografisk variation i Danmark
- 3.4 Forskelle inden for naturtyperne
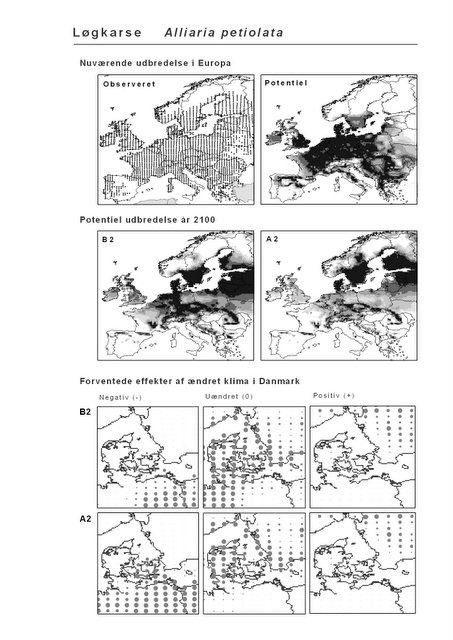
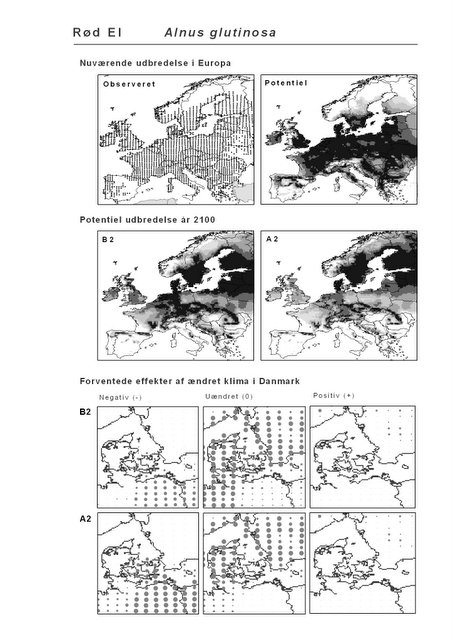
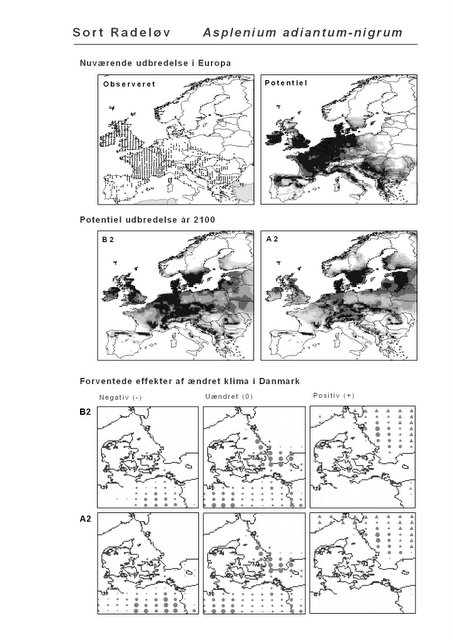
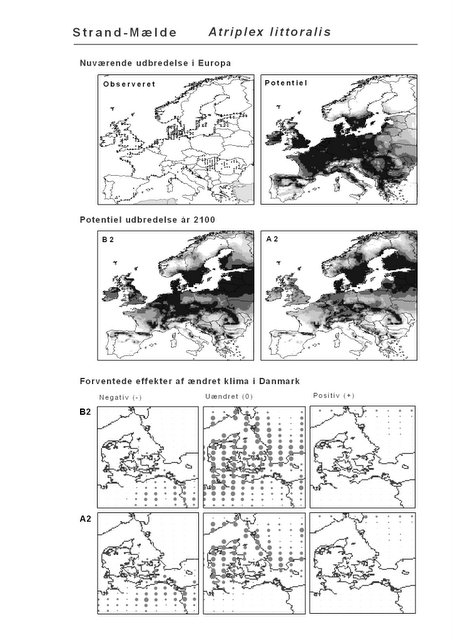
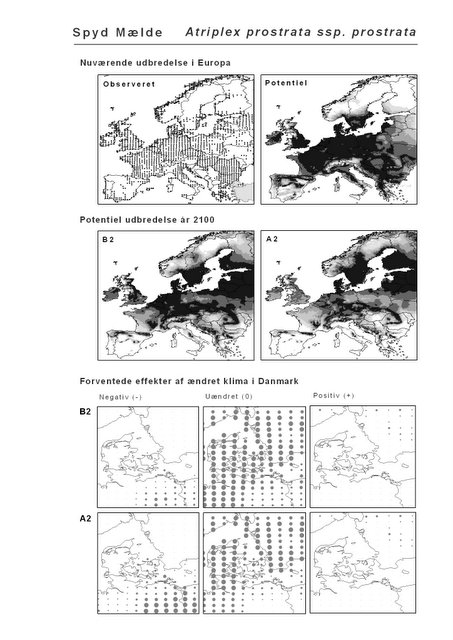
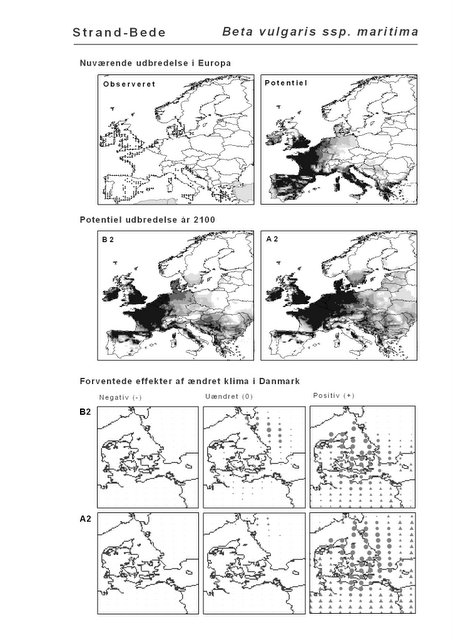
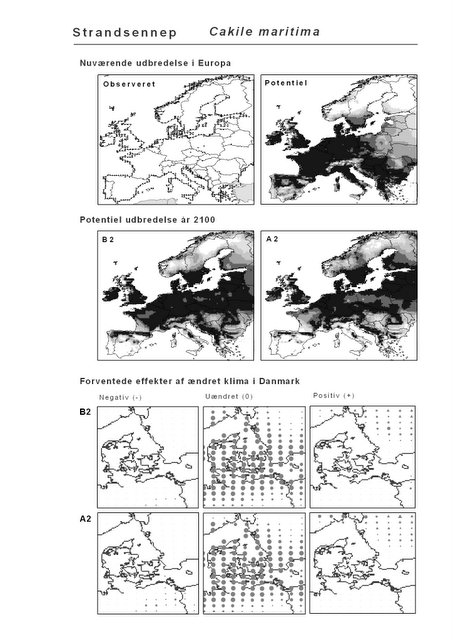
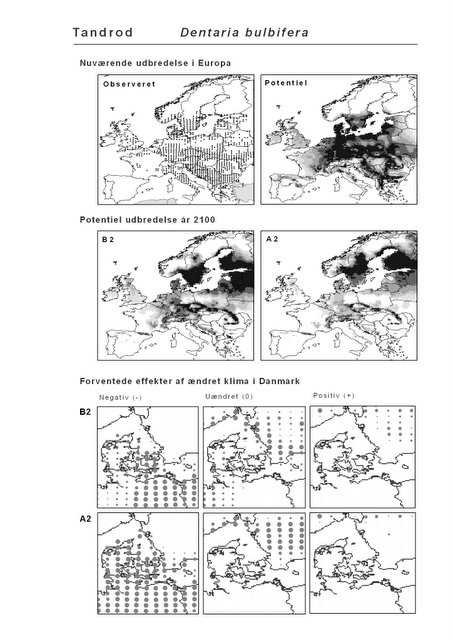
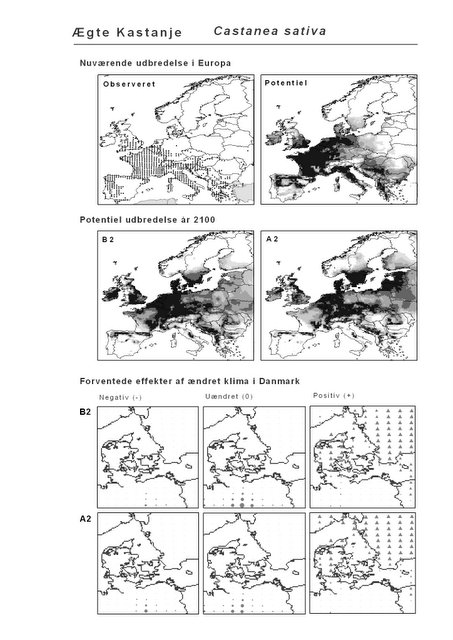
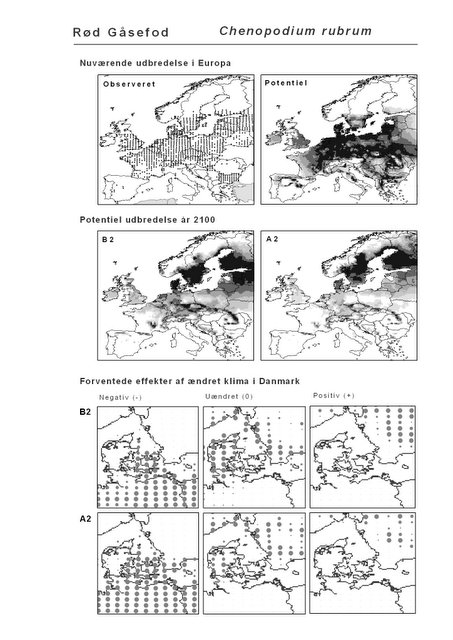
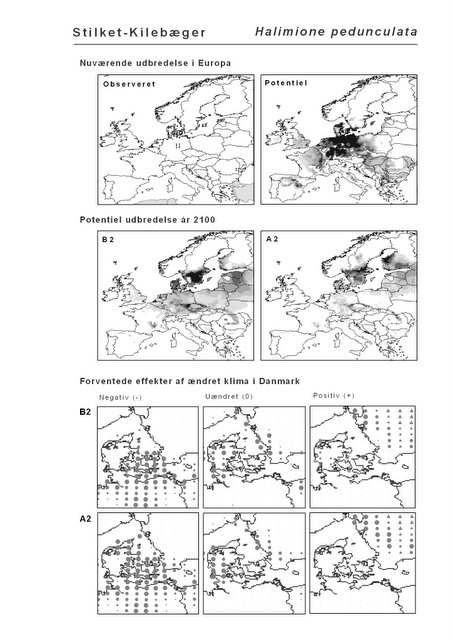
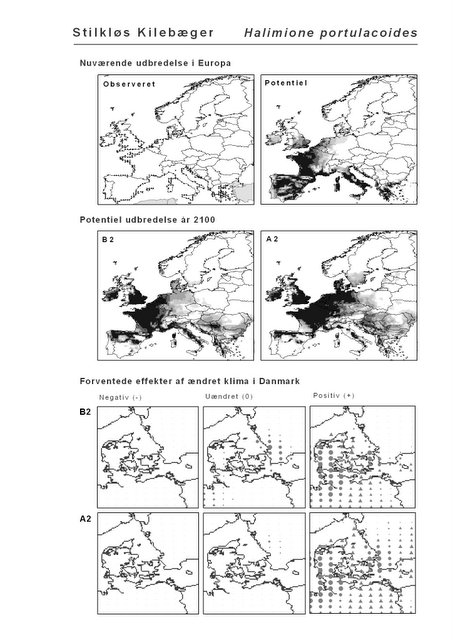
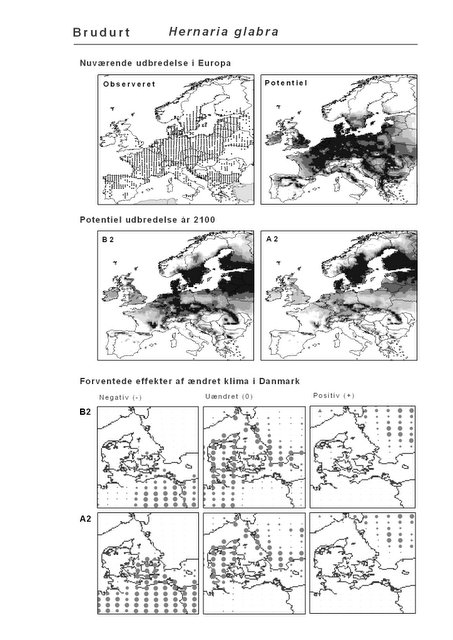
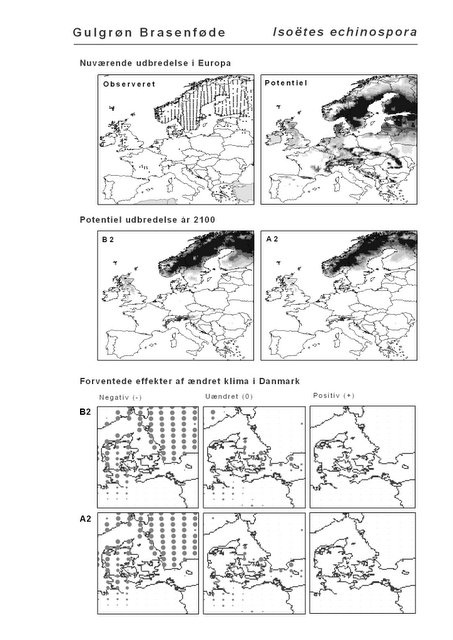
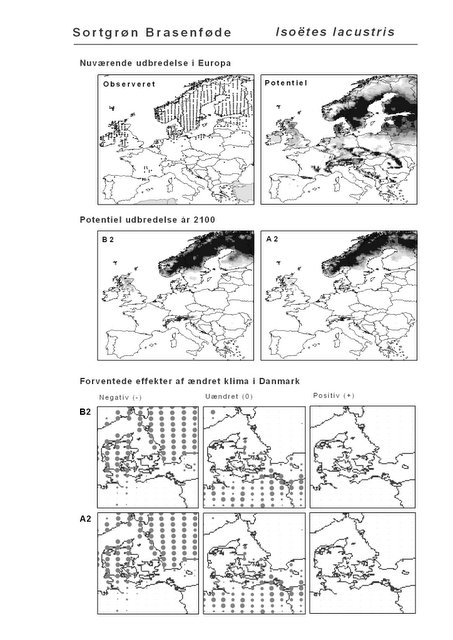
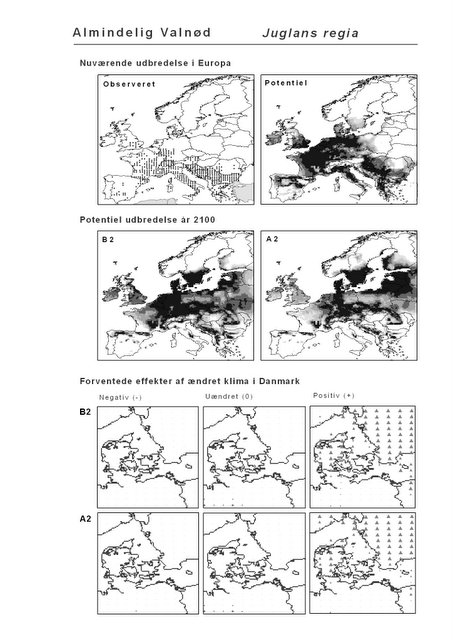
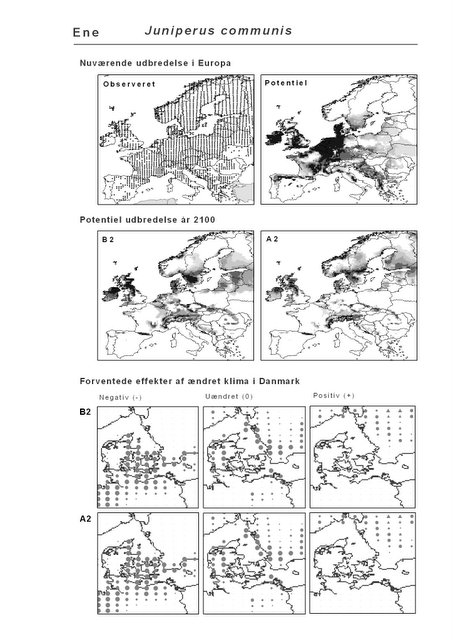
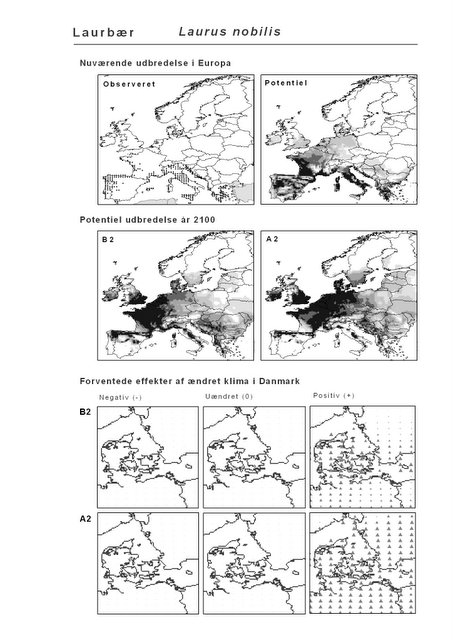
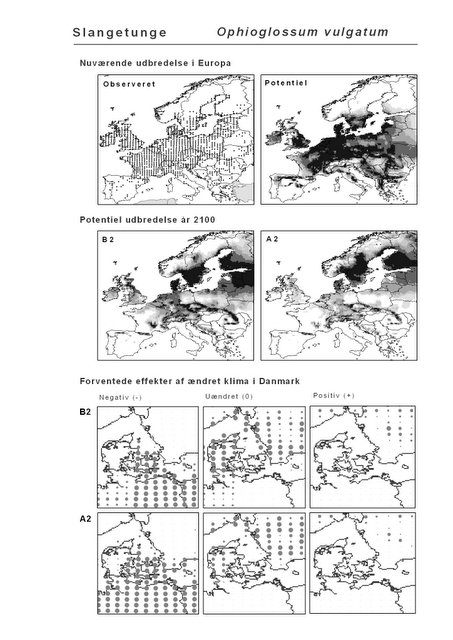
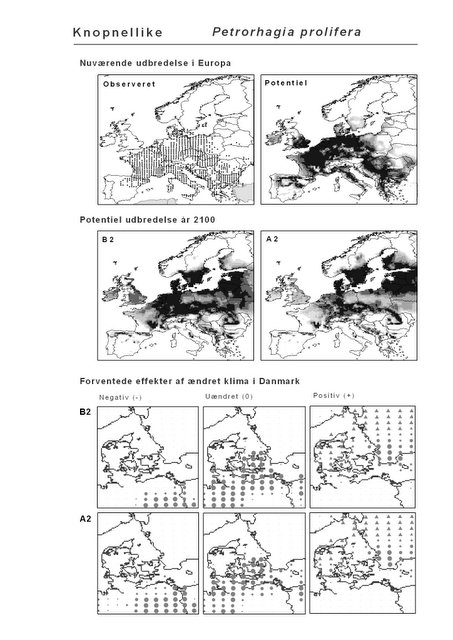
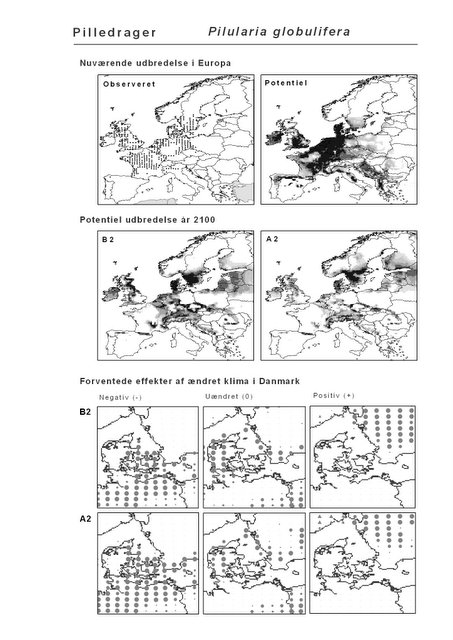
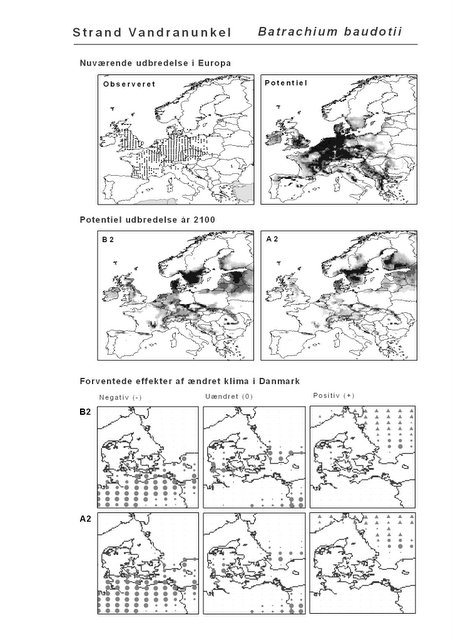
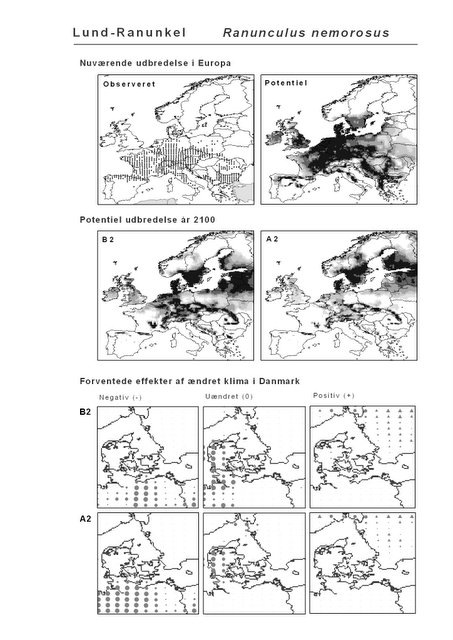
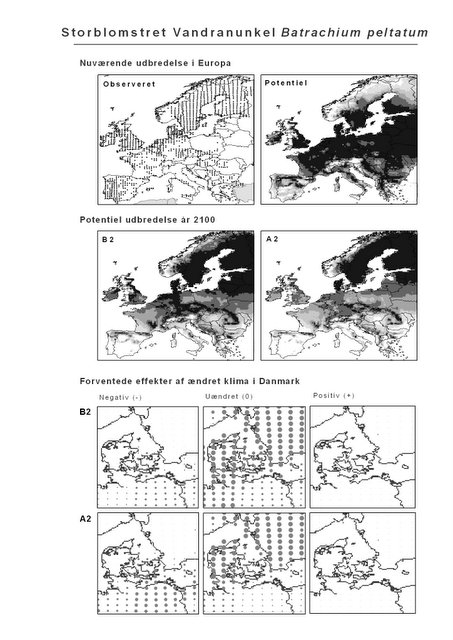
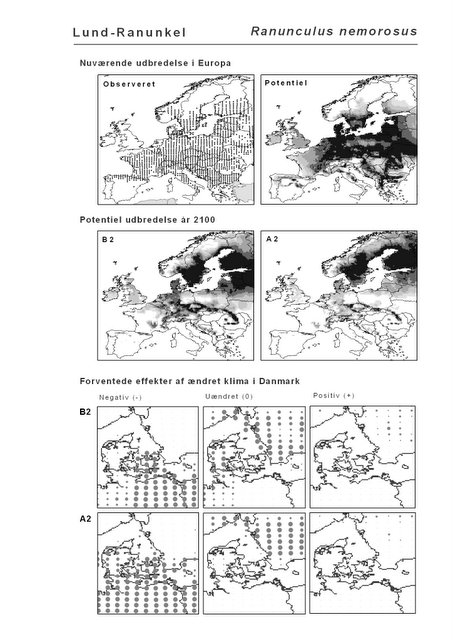
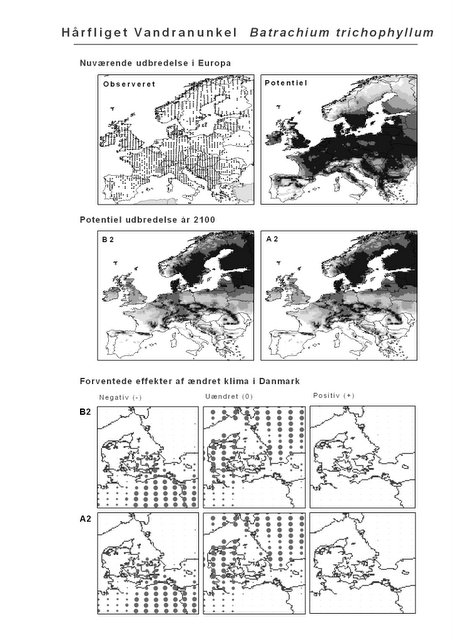
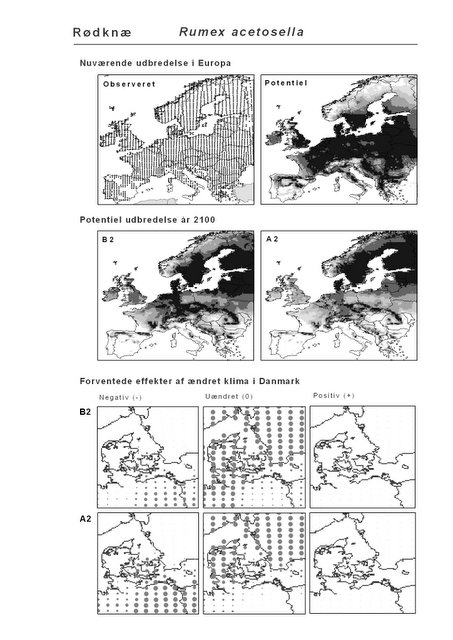
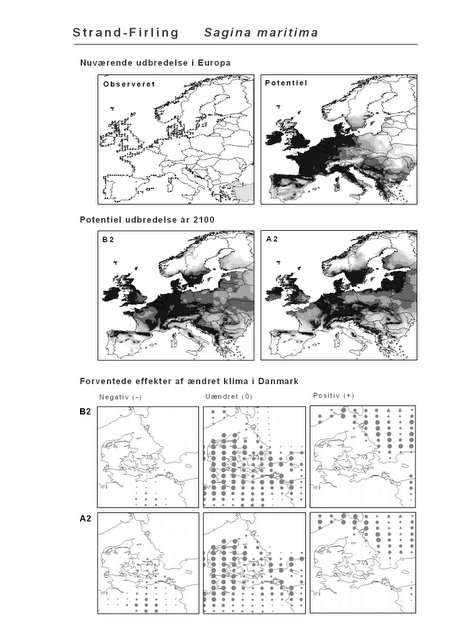
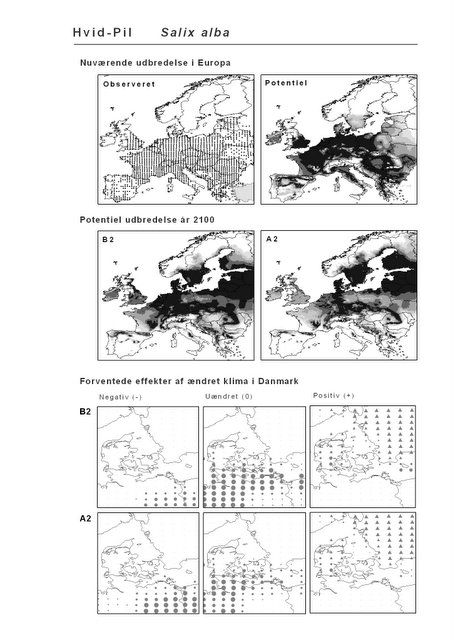
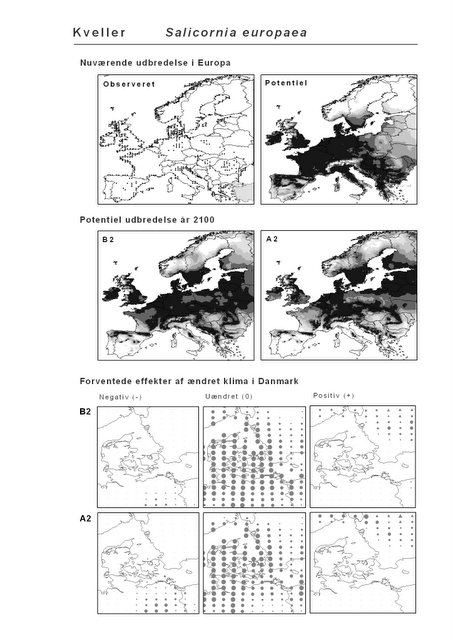
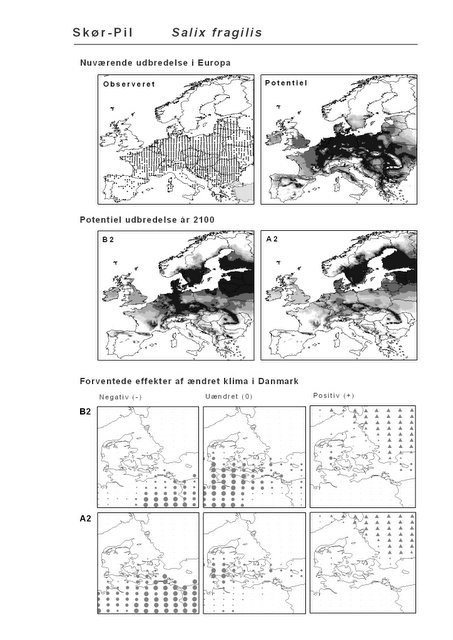
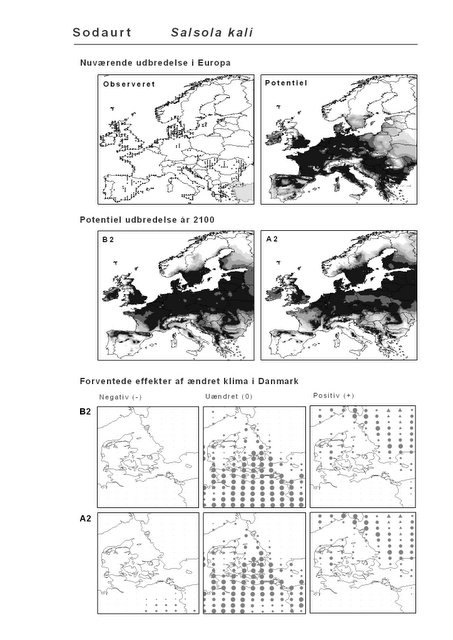
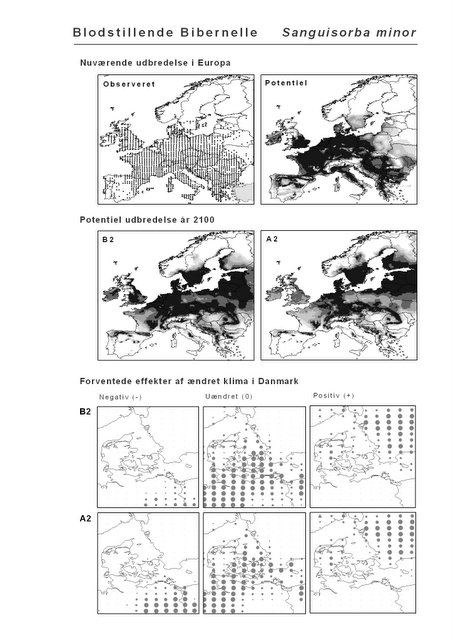
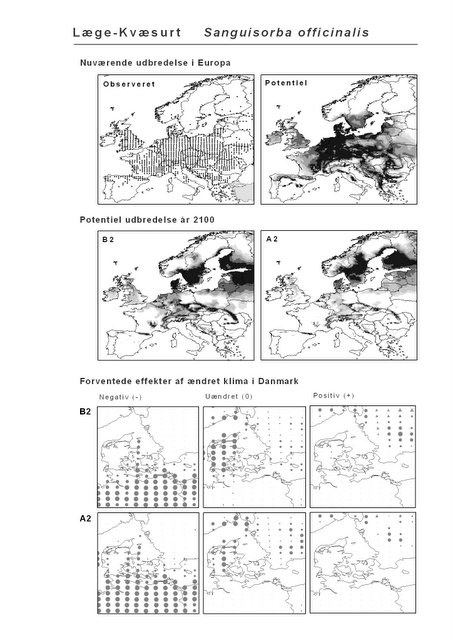
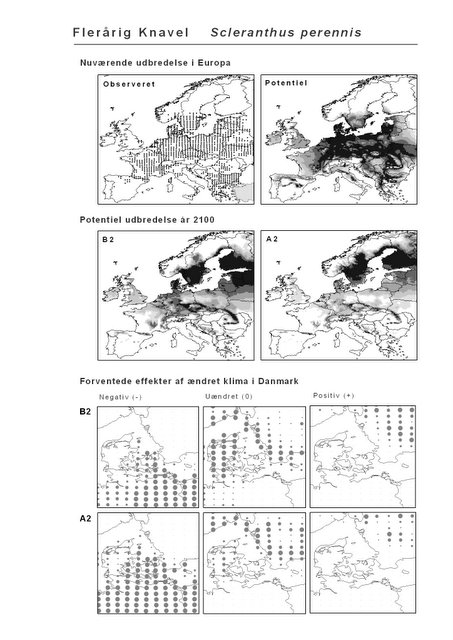
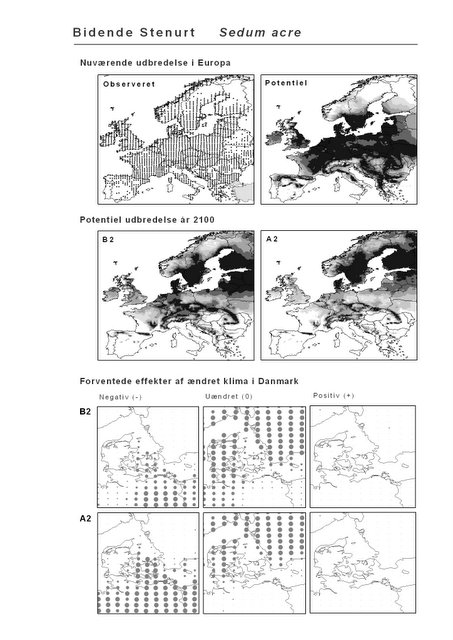
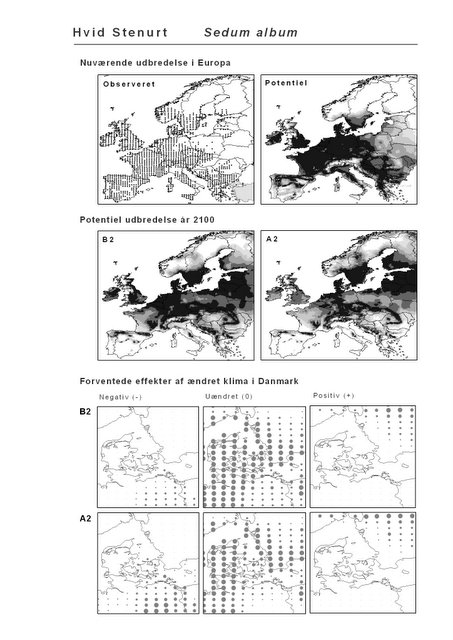
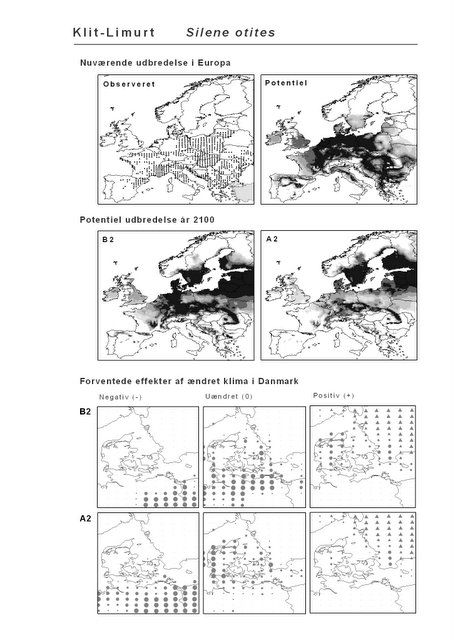
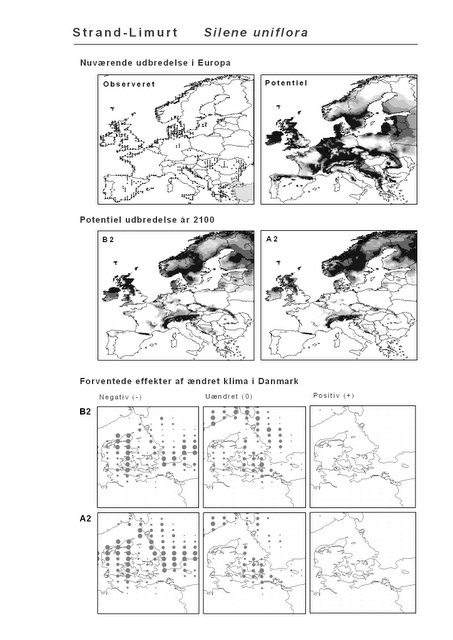
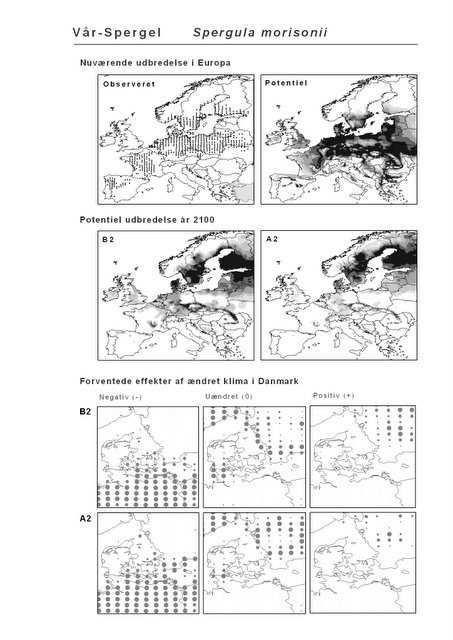
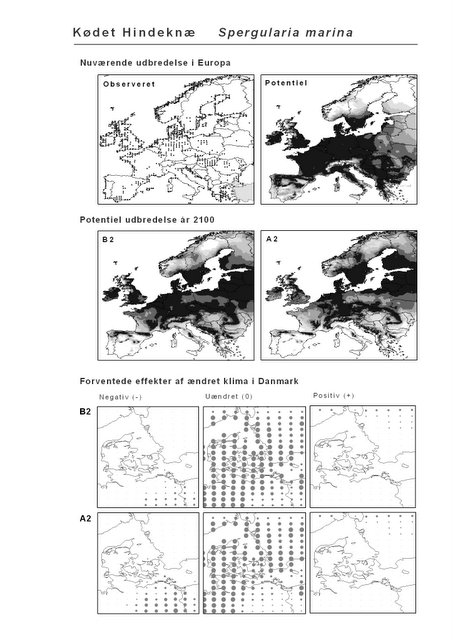
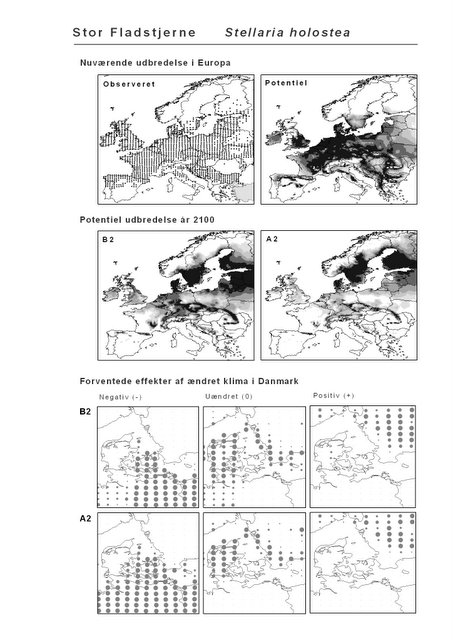
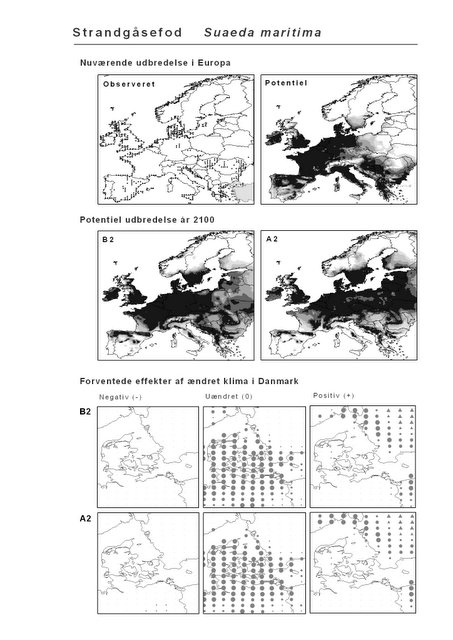
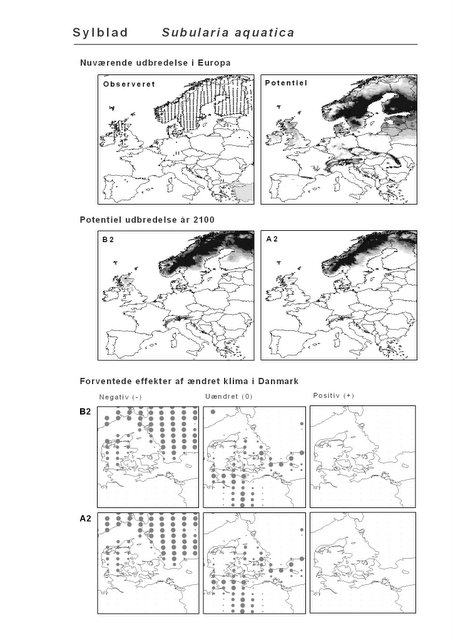
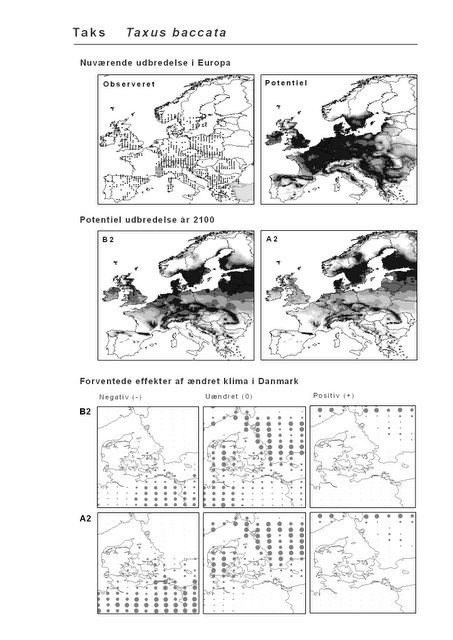
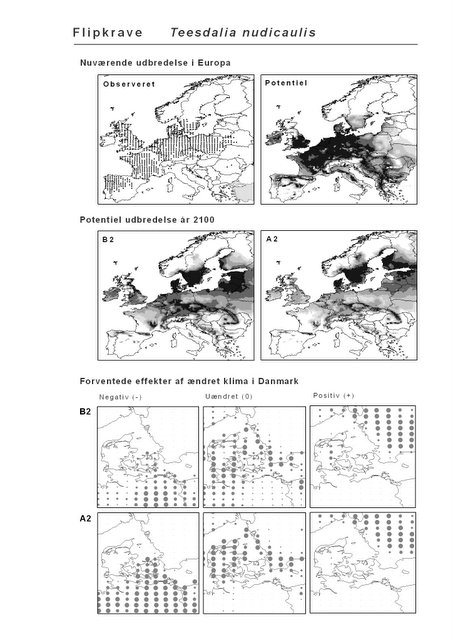
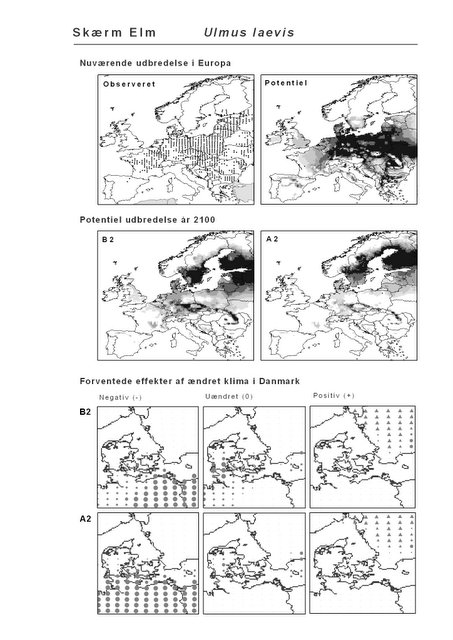
Resultaterne af den bioklimatiske habitat-modellering kan findes i Appendiks II. Hver art repræsenteres på et artsblad med følgende information:
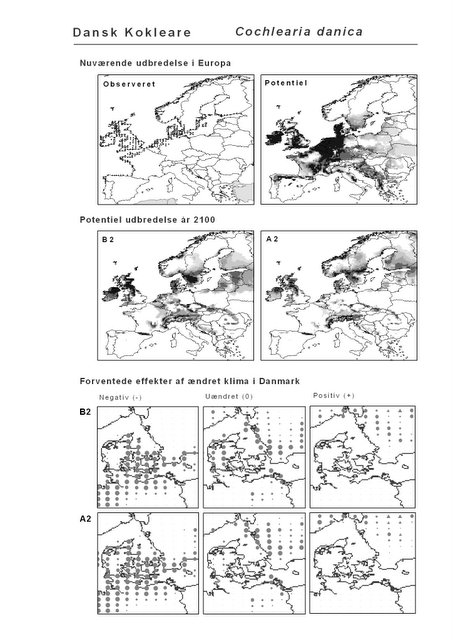
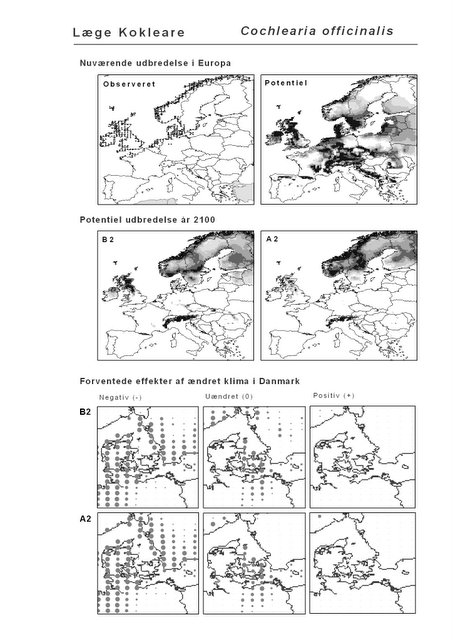
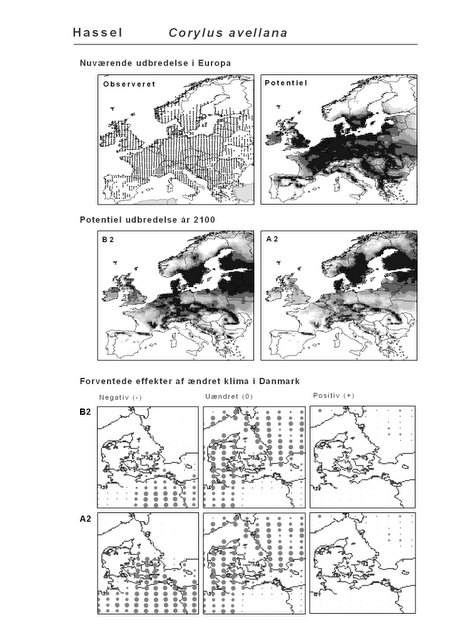
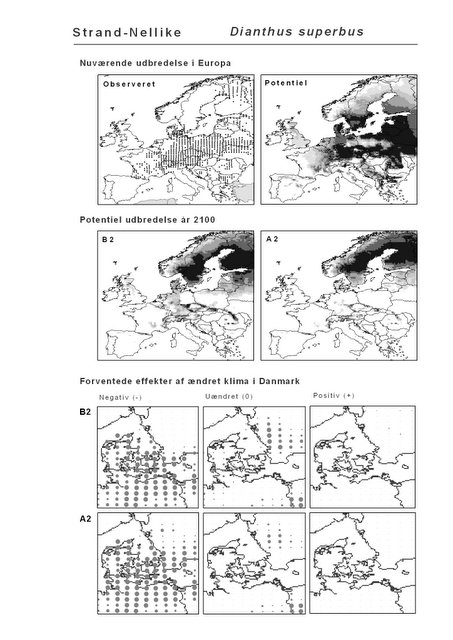
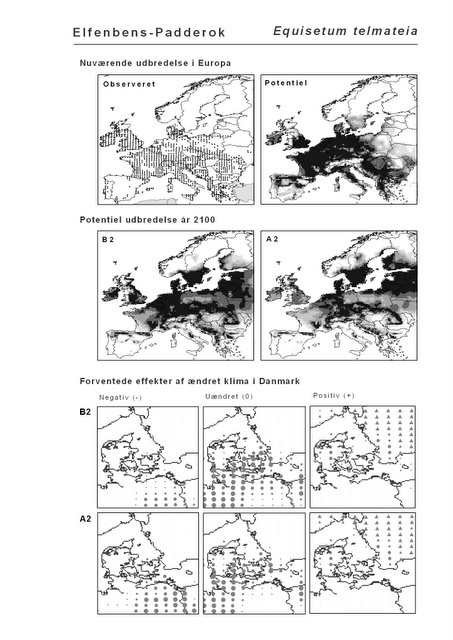
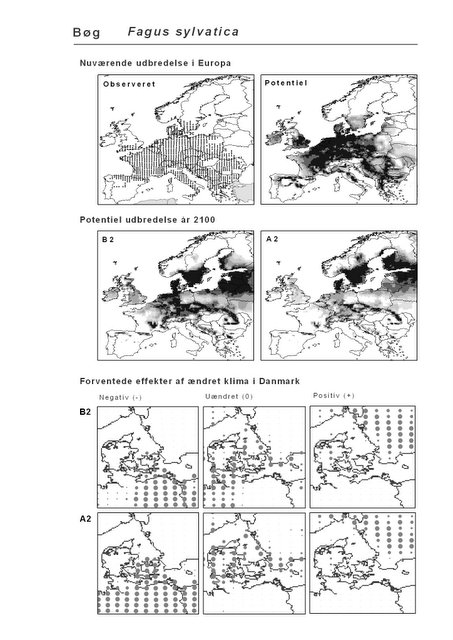
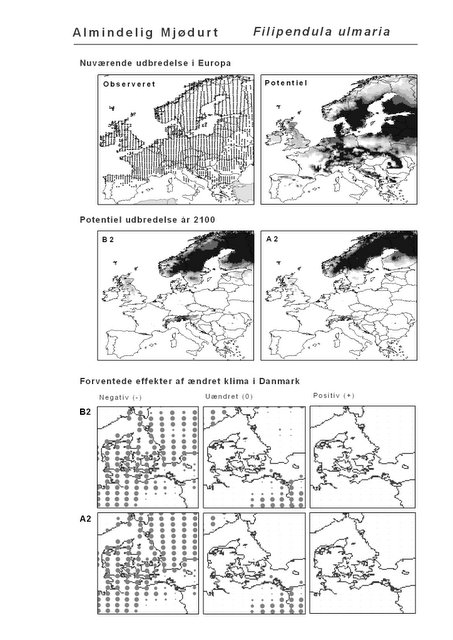
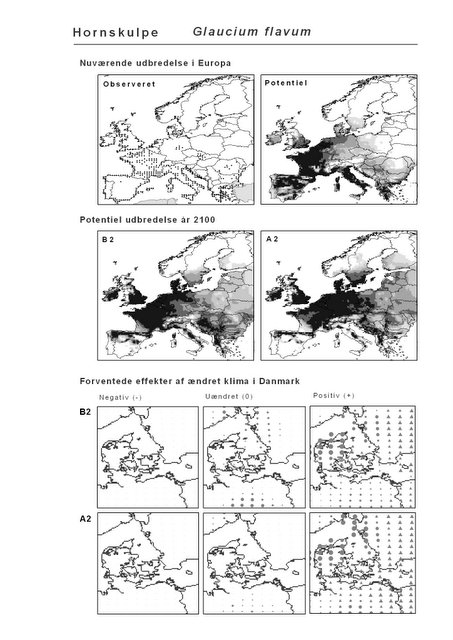
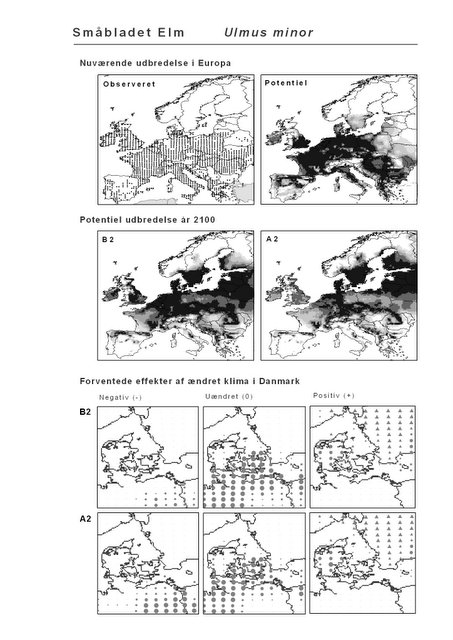
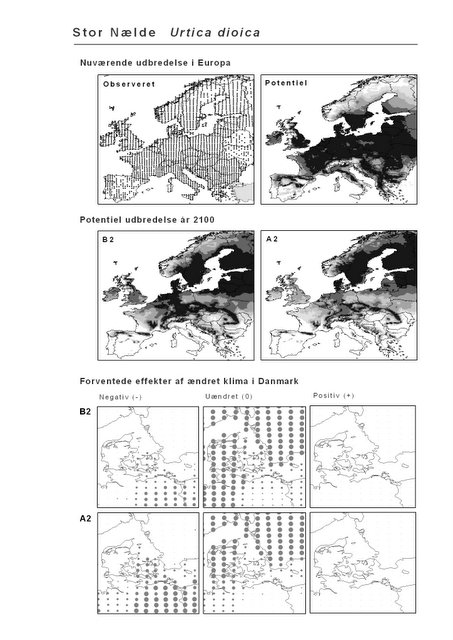
De øverste fire kort (se Figur 3.1 og Figur 2.2) viser dels nuværende observeret og potentiel udbredelse i Europa, dels den potentielle udbredelse i år 2100 under hhv. klimascenarium B2 og A2
De nederste seks kort viser de forventede effekter af et ændret klima i Danmark for de to scenarier (se Figur 3.1 og Figur 2.3).
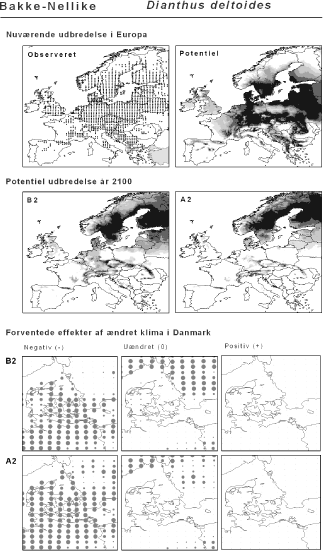
Figur 3.1 Et eksempel på et artsblad. Bakke-Nellike er karakteristisk for tør, kalkfattig sandet bund på bl.a. overdrev, skrænter og i grønklitter.
3.1 Udvalgte artseksempler
3.1.1 Negativ påvirkning
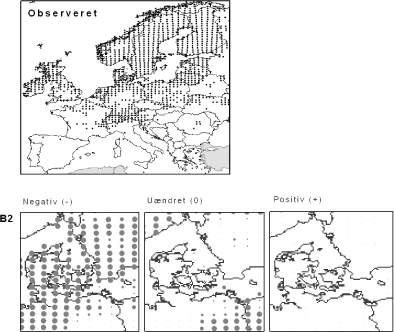
Figur 3.2 Langbladet Soldug (Drosera anglica). Øverst: Den observerede udbredelse iflg. Atlas Florae Europaeae. Nederst: Den forventede påvirkning i Danmark under B2-scenariet.

Langbladet Soldug er et eksempel på en art med en overvejende nordlig eller alpin udbredelse. Langbladet Soldug findes på våd tørvebund i bl.a. højmoser og fattigkær. Den er sjælden i Danmark, men temmelig almindelig i det øvrige Norden. Arten forventes at få ringere klimatiske kår i Danmark efter både scenarium B2 og A2. For landet som helhed forventes det, at begge klimascenarier vil forværre situationen for arten med 73% (Appendiks I). På kortet ovenfor (og i Appendiks II) ses, at tilstanden forventes at forværres ensartet i Danmark. Bemærk, at grunden til, at grænseområdet mellem Polen og Tyskland er angivet som uændret og ikke negativt, er, at området allerede i dag har en lav klimatisk egnethed.
3.1.2 Neutral påvirkning
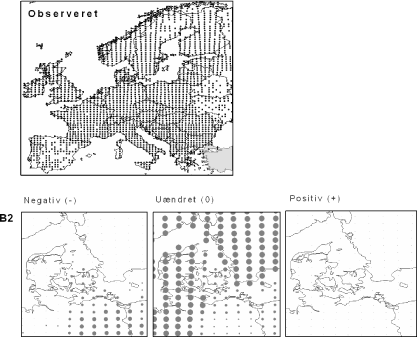
Figur 3.3 Stor Nælde (Urtica dioica). Øverst: Den observerede udbredelse iflg. Atlas Florae Europaeae. Nederst: Den forventede påvirkning i Danmark under B2-scenariet.

Stor Nælde har en meget vid udbredelse, der dækker det meste af Europa med undtagelse af særligt tørre områder i Middelhavsområdet og det centrale Europa. Arten har en stor klimatisk tolerance, og det forventes da også, at klimaet i Danmark fortsat vil være egnet for Stor Nælde i år 2100 under både scenarium B2 og A2. For landet som helhed forventes det, at klimascenarierne vil forværre situationen for Stor Nælde med hhv. 7% og 16% (Appendiks I). På kortet ovenfor (og Appendiks II) kan det ses, at det især er i det sydøstlige Danmark, at tilstanden vil forværres.
3.1.3 Positiv påvirkning
Figur 3.4 Hornskulpe (Glaucium flavum). Øverst: observerede udbredelse iflg. Flora Atlas Europaeae. Nederst: Den forventede påvirkning i Danmark.

Hornskulpe er et eksempel på en sydlig art, der er sjælden i Danmark i dag. Arten har en udpræget sydlig udbredelse og er sjælden i hele Norden. På vore breddegrader er Hornskulpe en udpræget kystnær art, der trives på strande eller strandvolde. For landet som helhed forventes det, at klimascenarierne vil forbedre situationen for Hornskulpe med hhv. 12% og 15% (Appendiks I). På kortet ovenfor (og i Appendiks II) kan det ses, at det især er i det vestlige Danmark, at tilstanden vil forbedres.
3.1.4 Potentiel positiv påvirkning
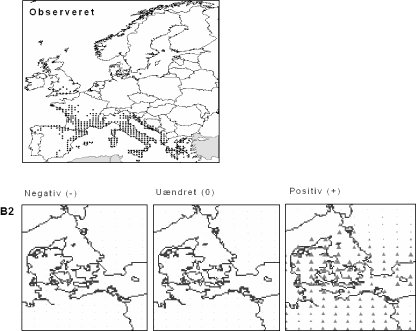
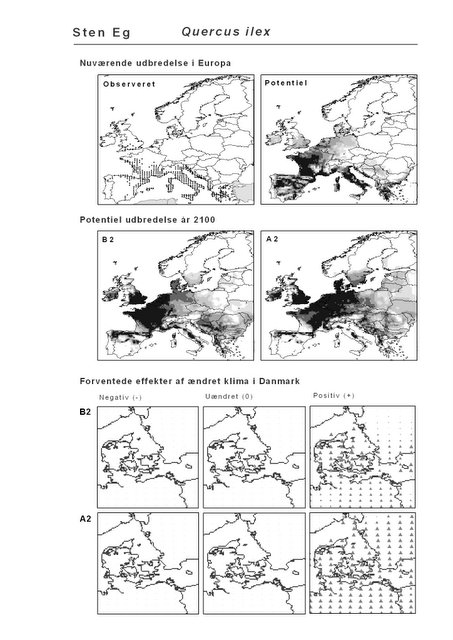
Figur 3.5 Sten-Eg (Quercus ilex). Øverst: Den observerede udbredelse iflg. Atlas Florae Europaeae. Nederst: Den forventede påvirkning i Danmark.

Sten-Eg er et eksempel på en sydlig art, der ikke vokser i Danmark nu, men må forventes at kunne finde egnet klima her om 100 år. Denne stedsegrønne træart forekommer naturligt vidt udbredt i Sydeuropa, men findes også naturaliseret så langt mod nord som England (Atlas Florae Europaeae). De trekantede symboler på kortet viser, at arten potentielt vil kunne trives i det meste af Danmark, hvis de beregnede klimaændringer indtræffer. Det er dog ikke sandsynligt, at Sten-Eg vil kunne indfinde sig af sig selv i løbet af den relativt korte periode pga. den store afstand til nutidens bestande.
3.2 Diversitet
Alle artsbladene i Appendiks II er gennemgået og for hver art er det vurderet, hvorvidt påvirkningen af klimaændringerne skønnes at være negativ, delvist negativ, neutral, delvist positiv, positiv eller potentielt positiv under de to klimascenarier. Resultaterne er opsummeret i tabel 3.1 nedenfor.
| Klimascenarium B2 | Klimascenarium A2 | ||||||||
| Ne- ga- tiv |
Del- vis ne- ga- tiv |
Neu- tral |
Del- vis po- si- tiv |
Po- si- tiv |
Ne- ga- tiv |
Del- vis ne- ga- tiv |
Neu- tral |
Del- vis po- si- tiv |
Po- si- tiv |
| 16 | 45 | 19 | 6 | 5 | 31 | 40 | 11 | 5 | 4 |
| 67,0% | 20,9% | 12,1% | 78,0% | 12,1% | 9,9% | ||||
Tabel 3.1 Antal arter, der forventes at blive påvirket negativt, delvist negativt, neutralt, delvist positivt og positivt under klimaændringerne under hhv. klimascenarium B2 og A2. Tallet i den midterste række angiver hvor mange arter, der er i hver kategori. Kun de 84 hjemmehørende arter i undersøgelsen er inkluderet. Den nederste række angiver i procent andelen af arter, der forventes overvejende at blive påvirket negativt, neutralt eller positivt af de forudsagte klimaændringer.
3.3 Geografisk variation i Danmark
Undersøgelsen viser, at der er relativt stor forskel på, hvor kraftig effekten af klimaændringerne forventes at være i forskellige dele af Danmark. Der er en klar tendens til, at det sydøstlige Danmark rammes langt hårdere end det vestlige Danmark. Tendensen er den samme for begge klimascenarier (se Figur 3.6). I det centrale Jylland er den totale ændring for samtlige arter kun på -4, hvilket betyder, at de negative konsekvenser for nogle arter stort set udlignes af positive konsekvenser for en andre arter. På det sydøstlige Sjælland og Lolland-Falster er den samlede effekt derimod stærkt negativ (-35 til -40).
Figur 3.6 Geografisk variation i effekterne af klimaændringer for de 88 karakteristiske arter. Ændringen i total klima-egnethed mellem nutiden og år 2100 under B2- og A2-scenariet indenfor det nuværende udbredelsesområde. Total klima-egnethed er beregnet ved for hver grid-celle at summere de 88 karakteristiske arters klima-egnethed i år 2100 og fratrække cellens summerede klima-egnethed i nutiden. Signaturen går fra blå (-40) til rød (+25).
3.4 Forskelle inden for naturtyperne
Det er interessant at vide, om planter i forskellige naturtyper reagerer på klimaændringer på samme måde, eller om nogle naturtyper er mere sårbare end andre? Tabel 3.2 opsummerer hvor egnet klimaet vil være for de forskellige naturtyper i Danmark om 100 år ifølge B2- og A2-scenarierne.
| Naturtype | I dag (%) | B2 (%) | A2 (%) | Δ B2 (%) | Δ A2 (%) |
| Alle | 65 ± 32 | 50 ± 32 | 40 ± 30 | -15 ± 21 | -26 ± 26 |
| Hav og strand | 65 ± 34 | 54 ± 35 | 48 ± 37 | -11 ± 23 | -18 ± 31 |
| Klitter | 59 ± 38 | 45 ± 30 | 35 ± 25 | -13 ± 23 | -24 ± 26 |
| Søer og vandløb | 65 ± 34 | 51 ± 40 | 42 ± 34 | -14 ± 11 | -23 ± 10 |
| Overdrev | 62 ± 30 | 52 ± 28 | 40 ± 28 | -10 ± 23 | -21 ± 29 |
| Ferske enge | 75 ± 20 | 50 ± 25 | 34 ± 25 | -25 ± 16 | -41 ± 19 |
| Lav og højmoser | 78 ± 14 | 40 ± 25 | 28 ± 24 | -38 ± 31 | -49 ± 33 |
| Klipper og huler | 69 ± 38 | 59 ± 36 | 47 ± 34 | -10 ± 8 | -22 ± 17 |
| Skove | 63 ± 36 | 48 ± 30 | 35 ± 27 | -15 ± 20 | -27 ± 27 |
Tabel 3.2 Middel klima-egnethed (± standard afvigelse) for en række danske habitattyper i dag og i 2100 efter klimascenarierne B2 og A2, samt den procentvise ændring (ΔB2, ΔA2) i klima-egnethed under B2- og A2-scenarierne i forhold til i dag. Den enkelte arts klima-egnethed er her beregnet som antallet af celler med at passende klima (klima-egnethed > 0) ud af det maksimalt mulige antal celler i Danmark.
Som det ses i Tabel 3.2 er spredningen på de enkelte naturtyper relativt stor, og da der kun er få arter i nogle af kategorierne (Tabel 2.1) kan der ikke påvises statistisk signifikante forskelle i de enkelte naturtypers klima-sensitivitet. Med en vis forsigtighed anes dog en tendens til, at ferske enge og især lav- og højmoser vil være mest sårbare overfor den globale opvarmning. En tilsvarende undersøgelse i England (Berry et al. 2002) har da også påvist et sammenligneligt resultat.
4 Diskussion
- 4.1 Hvor troværdig er den bioklimatiske habitat-modellering?
- 4.2 Sandsynlighed for tab af arter
- 4.3 Indvandring af nye arter
- 4.4 Er arterne repræsentative for dansk biodiversitet?
- 4.5 Anbefalinger
Resultaterne i denne rapport tyder på, at hvis klimaændringerne over det 21. århundrede bliver som beregnet i B2- og A2-scenarierne fra IPCC, må de forventes at have en helt eller delvist negativ effekt på forekomsten af mindst 2/3 af de udvalgte 84 hjemmehørende danske plantearter, der er karakteristiske for de danske naturtyper (jf. EU’s Habitatdirektiv, Buchwald & Søgaard 2000). Man må derfor forvente en relativt stærk negativ påvirkning af den hjemmehørende danske floras mangfoldighed såvel som den danske natur mere generelt.
4.1 Hvor troværdig er den bioklimatiske habitat-modellering?
Idet der ses bort fra den usikkerhed, der er knyttet til klimascenarierne i sig selv, i hvilket omfang kan man så fæste lid til de her rapporterede modelresultater? Der er betydelige huller i den videnskabelige forståelse af, hvordan arters udbredelse generelt begrænses. Selvom der er stor enighed om, at klima er en vigtig faktor på stor geografisk skala, så er det også klart, at mange andre faktorer kan spille ind, f.eks. spredning, jordbund og topografi (Svenning & Skov 2004; Pearson & Dawson 2003). Det er indlysende, at den usikkerhed, der ligger i den mangelfulde videnskabelige forståelse af arters udbredelse, vil blive overført til enhver prediktiv model for arters udbredelse i fremtiden. Det gælder også den bioklimatiske habitat-model, der er anvendt i denne rapport.
Den valgte model er relativt simpel og inddrager nogle få klimatiske parametre af veletableret betydning for planters udbredelse (Skov & Svenning 2004). Den er yderligere designet til at give nogle relativt brede estimater for en arts klimatiske niche og må ud fra det perspektiv anses for at give et velfunderet og ganske forsigtigt bud på de udbredelsesmæssige konsekvenser af ændringer i klimaet. Andre bioklimatiske modeller har da også givet mindst ligeså og ofte mere dramatiske forudsigelser af de biodiversitetsmæssige konsekvenser af den fremtidige opvarmning for en lang række geografiske regioner samt plante- og dyregrupper (se f.eks. Thuiller et al. 2005; Thomas et al. 2004).
Den bioklimatiske modellering er som tidligere nævnt ikke dynamisk (Pearson & Dawson 2003), dvs. den indeholder intet eksplicit tidsperspektiv, og en af dens vigtigste begrænsninger er derfor, at den ikke giver et direkte bud på, hvor hurtigt de forudsagte responser kan forventes realiseret. Overgangen fra sidste istid til den indeværende varme mellemistid skete i form af en kraftig temperaturstigning over nogle få tusinde år (med hurtige skift af større hastighed) frem til for ca. 10.000 år siden. Denne opvarmning var af samme størrelsesorden som den temperaturforøgelse, der forventes frem til år 2100 under f.eks. A2-scenariet. Skiftet fra istid til mellemistid gav anledning til meget dramatiske vegetationsændringer overalt i verden (se f.eks. Bush et al. 2004). Ud fra det perspektiv er det bestemt ikke urealistisk at forvente tilsvarende dramatiske vegetationsændringer i den nære fremtid.
Det skal også nævnes, at det stigende indhold af CO2 i atmosfæren må forventes at kunne få stor indvirkning på vegetation, men at de store usikkerheder, der er i den videnskabelige forståelse af dette emne (Ehleringer et al. 2005), ikke har tilladt at inkludere en sådan effekt i den bioklimatiske habitat-modellering.
4.2 Sandsynlighed for tab af arter
Dette studium viser at op mod 4% og 7% af de 84 hjemmehørende karakteristiske arter helt må forventes at miste passende klimatiske forhold i Danmark under hhv. B2- og A2-scenariet. Tilsvarende må hhv. 6% og 15% af disse arter forventes at blive udsat for klimatiske forhold, der er mere end 80% dårligere end dem, de lever under i Danmark i dag. Skal vi forvente, at disse arter uddør i Danmark over de næste hundrede år?
Som diskuteret ovenfor er tidsperspektivet noget usikkert, idet modellen ikke er dynamisk og derfor ikke har et eksplicit tidsperspektiv. Det er dog veldokumenteret, at mere mobile organismegrupper som fugle, havfisk og sommerfugle allerede har udvidet deres udbredelse i retning af polerne som reaktion på den temperaturstigning, der allerede er foregået over de sidste hundrede år (Thomas & Lennon 1999; Parmesan et al. 1999; Perry et al. 2005). Det er også vist, at mange plantearter mere lokalt har ekspanderet nær deres nordvendte grænse (Walther et al. 2002). Det kan derfor forventes, at vi vil se en forsat udvidelse af arternes udbredelse i retning af polerne, hvis den globale temperaturstigning fortsætter.
Det er derimod mere kontroversielt, om vi kan forvente en tilsvarende nordvendt forskydning af arternes grænse mod syd. Grunden hertil er, at mens det er alment accepteret, at arternes polvendte grænse kontrolleres af abiotiske faktorer, tillægges biotiske faktorer, f.eks. konkurrence, af nogle en større betydning for kontrollen af den ækvatorvendte grænse (Loehle 1998). I så fald vil en tilbagetrækning af nordlige arter nordenud af Danmark i et vist omfang afhænge af, om deres sydlige konkurrenter indfinder sig i Danmark og altså ikke blot af klimaændringerne i sig selv. I så tilfælde vil lokal naturpleje i et vist omfang kunne modvirke den generelle effekt af et varmere klima. Der er dog efterhånden samlet en del empiriske evidens for, at den ækvatorvendte grænse (eller den nedre grænse i bjergegne) kan være nogenlunde ligeså følsom overfor klimatisk opvarmning som den polvendte/øvre grænse (Perry et al. 2005; Bush et al. 2004). Samlet set er der derfor ikke umiddelbart belæg for, at den bioklimatiske habitat-modellering i særlig grad skulle overdrive effekten af den globale opvarmning på de danske arters sydgrænse og derved på de forudsagte artstab.
4.3 Indvandring af nye arter
Den bioklimatiske habitat-modellering viser, at sydlige arter, der ikke i dag forekommer naturligt i Danmark, må forventes at kunne trives i den danske natur i år 2100 under begge klimascenarier. Det drejer sig ikke blot om arter, der i dag er vidt udbredte i Mellemeuropa, f.eks. Ægte Valnød (Juglans regia) og Ægte Kastanie (Castanea sativa), men også udpræget sydeuropæiske arter som Strand-Fyr (Pinus pinaster) og de stedsegrønne løvtræer Laurbær (Laurus nobilis) og Sten-Eg (Quercus ilex). Laurbær og andre stedsegrønne løvtræer ses allerede nu invadere hidtidig løvfældende skov i det sydligste Mellemeuropa som følge af den allerede realiserede stigning i vintertemperaturen (Walther et al. 2000; 2002). I hvilket omfang en tilsvarende indvandring af sydlige arter til danske himmelstrøg vil blive realiseret under et fremtidigt varmere klima vil i høj afhænge af, om arterne kan sprede sig hertil fra deres nutidige udbredelsesområde på den tid, der er til rådighed. På samme vis vil spredning være af afgørende betydning for om arter, hvis klimaområde forskydes nord for Danmark, vil kunne følge med derop.
Generelt forventer vi, at spredningsrater i størrelsesordnen mindst 2 km per år er krævet for, at arterne kan følge med til de klimaændringer, som IPCC’s scenarier angiver for år 2100 på europæisk skala (Skov & Svenning 2004). Derved overstiger de påkrævede rater langt dem, der ses for mange planter i naturen i dag eller er estimeret for planternes ekspansion efter sidste istid (Skov & Svenning 2004; Clark et al. 1998; Honnay et al. 2002). Nye studier indikerer tilmed, at mange europæiske træarter kun i begrænset omfang har formået at vandre nordpå efter sidste istid trods det, at passende klimatiske forhold har eksisteret her i tusindvis af år (Svenning & Skov 2004; Svenning & Skov 2005). Derfor må spredning forventes at blive en særdeles vigtig begrænsende faktor for planters positive udbredelsesmæssige respons på fremtidige klimaændringer.
Dette gælder endda i endnu stærkere grad, når man også tager hensyn til den udstrakte menneskeskabte fragmentering af de naturlige habitater i Danmark og det meste af det øvrige Europa. Figur 4.1 viser et forsøg på at finde de mest sandsynlige spredningskorridorer gennem Europa for skovlevende planter. De angivne ruter går gennem områder med mindst ’friktion’ (dvs. mest skov). Kortet viser dels, at bevarelse af det nuværende skovdække er vigtigt, hvis det skal sikres, at forskellige organismer skal kunne spredes. Kortet viser også at lande med et lille skovbevokset areal – som f.eks. Danmark – formodentlig ikke vil komme til spille den store rolle som ’gennemgangsland’.
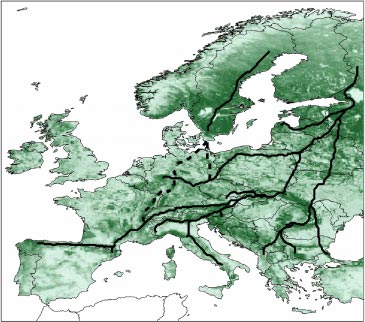
Figur 4.1 Mulige spredningskorridorer gennem Europa for skovlevende arter. Kortet viser tætheden af skov i dag. Linjerne går gennem de områder, hvor der i dag er mest skov og som derfor vil udgøre de vigtigste spredningskorridorer (fra Skov & Svenning 2004).
På grund af at mange planter har en begrænset kapacitet for hurtig langdistance spredning, må den aktive og passive langdistance transport af planter, der udføres af mennesker, bl.a. i forbindelse med skov- og havebrug, forventes at blive af særdeles stor vigtighed for den styrke og hastighed, hvormed vegetationen responderer på fremtidige klimaændringer. Aktiv flytning af plantebestande efter passende klimatiske forhold kan blive en nødvendighed for at sikre mange plantearters genetiske diversitet og måske deres overlevelse som sådan (Honnay et al. 2002; Skov & Svenning 2004).
4.4 Er arterne repræsentative for dansk biodiversitet?
I denne rapport er de såkaldt karakteristiske plantearter fra Habitatdirektivet brugt til at illustrere de potentielle effekter af fremtidige klimaændringer på biodiversiteten i Danmark. Dette rejser spørgsmålet, om resultaterne kan bruges til sige noget generelt om biodiversiteten i Danmark? Særligt kan man spørge i hvilket omfang karplanter kan bruges som indikatorer for andre organismegruppers respons på klimaforandringer?
Forskellige organismegrupper har forskellig økologi og er derfor særligt følsomme overfor forskellige typer af landskabsændringer. Dette fremgår tydelig af ”Rødliste 1997 for planter og dyr i Danmark” (Stoltze & Pihl 1998), der bl.a. viser, at laver er særligt følsomme overfor luftforurening, urter særligt følsomme overfor tilgroning af lysåbne naturtyper, mens mængden af gamle træer og dødt ved i skov er af særlig betydning for biller.
Trods de særlige økologiske krav hos forskellige organismegrupper er der dog megen evidens for, at klimaet har en meget stor og temmelig ensartet betydning for udbredelsen og diversiteten af de fleste organismegrupper på større geografisk skala (måske dog undtaget mikroorganismer). Således har man observeret tydelige fænologiske og i et vist omfang udbredelses-mæssige responser på den klimatiske opvarmning hos en bred vifte af dyre- og plantegrupper, se f.eks. Walther et al. (2002) eller Root et al. (2003). Studier af forskellige organismegruppers klimatiske niche vha. bioklimatisk habitat-modellering har også konkluderet, at både karplanter, insekter og fugle er begrænset på lignende vis af klimaet på stor skala (Huntley et al. 2004). Det må derfor også forventes, at de vil reagere nogenlunde sammenligneligt på fremtidige klimaændringer (Thomas et al. 2004).
Vi mener derfor, at man med nogen forsigtighed godt kan bruge karplanter som indikatorer for følsomheden af den danske ikke-mikrobielle biodiversitet overfor klimaændringer.
4.5 Anbefalinger
Rapporten viser, at de vegetationsændringer der forudsiges selv under det relativt milde B2-scenarium er ganske dramatiske. Fire procent af de karakteristiske arter forudsiges at forsvinde fra Danmark, i hvert fald på længere sigt, og 2/3 af arterne må forventes at gå tilbage i større eller mindre omfang. Det mere ekstreme A2-scenarium har dog en klart stærkere negativ effekt end B2-scenariet, så det fremgår tydeligt, at de biodiversitetsmæssige tab, vi må forvente, vil afhænge af styrken af de klimaændringer, der bliver realiseret.
4.5.1 Overvågning
Som nævnt flere gange i rapporten er der stor usikkerhed i forbindelse med både klimascenarierne og den bioklimatiske modellering. Denne usikkerhed er problematisk i forbindelse med at beslutte og gennemføre bekostelige tiltag i forsøg på at modvirke og tilpasse sig til de negative effekter af de forudsagte klimaændringer. Skal man f.eks. i skovbruget anvende andre træarter end hidtil? Hvis det er tilfældet skal man allerede nu begynde at plante de nye arter, så de passer til klimaet om 50-100 år. På den anden side kunne det vise sig at være en fejlinvestering, hvis klimaet ikke ændrer sig som forudsagt.
Det anbefales derfor at udarbejde en rullende strategi baseret på en overvågning af naturtyper og arter. Denne overvågning kunne f.eks. basere sig på det nye NOVANA-program, hvori der hvert 6. år skal foreligge en statusrapport for en række naturtyper og arter baseret på et stort antal målestationer landet over. Denne statusopgørelse vil være velegnet til at følge udviklingen for et stort antal arter og konstatere, om de går frem eller tilbage, og hvorvidt nye arter kommer til. Baseret på denne viden vil en tilpasningsstrategi løbende kunne opdateres og tilrettes de reelle forhold.
4.5.2 Virkemidler
Sikring af sammenhængende naturområder. – Hvis de fremtidige klimaændringer bliver af den styrke som beskrives i B2- og A2-scenarierne, vil mange arter være nødt til at justere deres udbredelse betydeligt mod nord/nordøst. For at støtte arternes muligheder for at følge klimaet vha. spredning, vil det være vigtigt at sikre sammenhæng mellem naturområderne i det danske landskab. Man skal dog være opmærksom på det uundgåelige dilemma, at en øget sammenhæng mellem naturområder også vil lette spredning af sydlige arter, hvis forekomst i Danmark er muliggjort af en temperaturstigning, imellem de danske naturområder. Dette kan forårsage en hastigere og stærkere tilbagegang for klimatisk følsomme hjemmehørende arter. I det omfang at disse arter er hjemmehørende længere sydpå i Europa, og dér bliver truet af klimaændringerne vil en sådan spredning dog trods alt være hensigtsmæssig set fra et overordnet europæisk perspektiv. Ud fra det synspunkt vil det således være hensigtsmæssigt også at tilstræbe sammenhæng mellem naturområderne på tværs af landegrænserne i Europa (Skov & Svenning 2004 og Figur 4.1).
Sikring af store og vidt udbredte bestande. – For at øge en arts robusthed og dermed reducere dens risiko for at uddø er det vigtigt at sikre dens evolutionære tilpasningsevne ved at opretholde mange, genetisk forskellige populationer. Hvis arten også er vidt udbredt, vil man hermed også sikre miljømæssige tilpasningsmuligheder. Dette kan dog ikke gøres i Danmark alene, men kræver samarbejde på europæisk plan.
Aktiv flytning af bestande. – Et kontroversielt redskab, der sjældent anvendes i moderne naturforvaltning, er aktiv flytning af individer eller bestande. Det er der flere grunde til: Dels ønskes der ikke at gribe ind i naturlige processer og sløre naturlige udbredelsesmønstre, dels kan det føre til overdreven naturmanipulation, hvor man for eksempel giver tilladelse til, at ét naturligt økosystem ødelægges under henvisning til, at man kan oprette en erstatningsbiotop et andet sted. Aktiv flytning af bestande efter passende klimatiske forhold kan imidlertid blive nødvendig, hvis en art ikke vil være i stand til at følge de forudsagte klimaændringer på naturlig vis og derfor er i fare for at tabe uacceptabelt meget af sin genetiske diversitet eller endog helt at uddø.
Sikring af klimatisk stabile områder. – Under de kendte forhistoriske klimaændringer har en række områder i Europa fungeret som refugier. De er kendetegnet ved en vis klimatisk stabilitet som følge af stor terrænvariation, der sikrer mange klimatiske zoner indenfor et relativt lille område. I sådanne områder skal arterne ikke flytte sig ret meget for at finde gunstigt klima under en klimaændring, og de må tillægges derfor særlig vægt i naturbeskyttelsen. Udvælgelse og beskyttelse af sådanne områder bør ikke kun være et nationalt anliggende, men også udføres som et fælles europæisk initiativ.
4.5.3 Klimaændringer og skovbrug
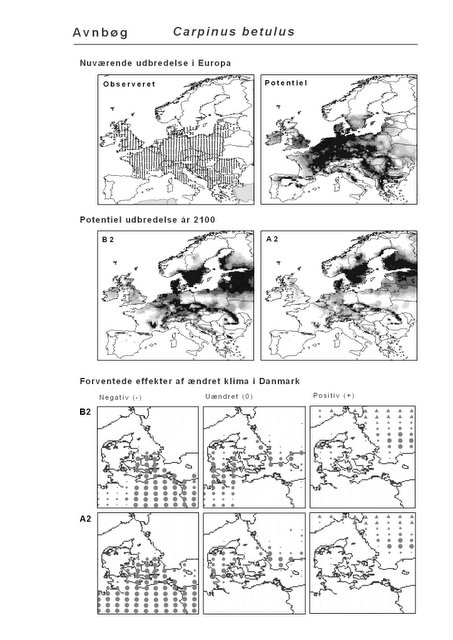
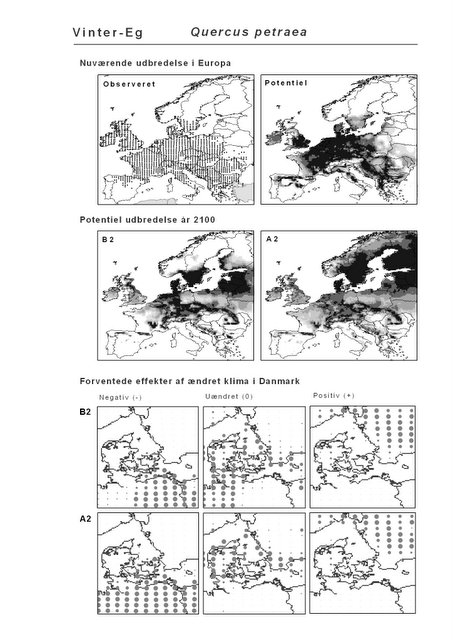
Det er bemærkelsesværdigt, at hjemmehørende danske træarter som Rød-El (Alnus glutinosa), Anvbøg (Carpinus betulus), Bøg (Fagus sylvatica), Stilk- og Vinter-Eg (Quercus robur og Q. petraea), modsat den indførte, men meget plantede Rød-Gran (Picea abies), forudsiges fortsat at finde passende forhold i Danmark om 100 år selv under A2-scenariet. Herved støtter den bioklimatiske habitat-modellering op om Skov- og Naturstyrelsens Handlingsplan for naturnær skovdrift i statsskovene, hvori et vigtigt argument for overgangen til naturnær drift med vægt på hjemmehørende træarter er forventningen om, at skove drevet efter naturnære principper vil være langt mere robuste overfor klimaændringer end konventionelt drevne skove (Skov- og Naturstyrelsen 2005).
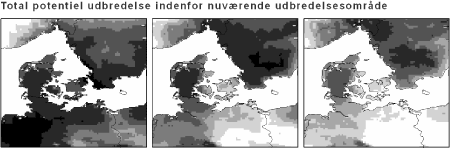
Figur 4.2 Total klima-egnethed for hjemmehørende danske skovtræer nu (tv) og under klimascenarierne B2 og A2 (hhv. i midten og th.). Beregnet som i Figur 3.6. Signaturen går fra 14 arter (sort) til 0 (hvid).
Resultaterne fra denne rapport viser også, at sydeuropæiske træarter som Strand-Fyr (Pinus pinaster) og Sten-Eg (Quercus ilex) muligvis vil kunne trives i Danmark om 100 år. Om det på et tidspunkt vil blive relevant, at benytte sig af disse i skovbruget må bl.a. afhænge af de biodiversitetsmæssige konsekvenser heraf. Det vil derfor til den tid være vigtigt at få belyst, om indførelsen af disse arter vil forstærke de negative effekter af klimaændringerne for den naturlige flora og fauna, f.eks. ved at udkonkurrere klimatisk pressede nordlige arter.
5 Litteratur
Berry,P.M., Dawson,T.P., Harrison,P.A. & Pearson,R.G. (2002). Modelling potential impacts of climate change on the bioclimatic envelope of species in Britain and Ireland. Global Ecology and Biogeography, 11, 453-462.
Buchwald, E. & Søgaard, B. (eds) (2000) Danske naturtyper i det europæiske NATURA 2000 netværk. Miljø- og Energiministeriet, Skov- og Naturstyrelsen, København, Danmark.
Bush, M. B., Silman, M. R., & Urrego, D. H. (2004) 48,000 years of climate and forest change in a biodiversity hot spot. Science, 303, 827-829.
Clark, J. S., Fastie, C., Hurtt, G., Jackson, S. T., Johnson, C., King, G. A., Lewis, M., Lynch, J., Pacala, S., Prentice, C., Schupp, E. W., Webb III, T., & Wyckoff, P. (1998) Reid's paradox of rapid plant migration: dispersal theory and interpretation of paleoecological records. BioScience, 48, 13-24.
Ehleringer, J.R., Cerling, T.E. & Dearing, M.D. (eds) (2005) A history of atmospheric CO2 and its effects on plants, animals, and ecosystems. Ecological studies 177. Springer Science+Business Media, Inc., New York, New York, USA.
Guisan, A. & Zimmermann, N. E. (2000) Predictive habitat distribution models in ecology. Ecological Modelling, 135, 147-186.
Hannah, L., Midgley, G. F., & Millar, D. (2002) Climate change-integrated conservation strategies. Global Ecology and Biogeography, 11, 485-495.
Hansen, K.(ed.) (1981) Dansk Feltflora. Gyldendalske Boghandel, Nordisk Forlag A/S, Copenhagen, Denmark.
Hawkins, B. A., Field, R., Cornell, H. V., Currie, D. J., Guegan, J. F., Kaufman, D. M., Kerr, J. T., Mittelbach, G. G., Oberdorff, T., O'Brien, E. M., Porter, E. E., & Turner, J. R. G. (2003) Energy, water, and broad-scale geographic patterns of species richness. Ecology, 84, 3105-3117.
Honnay, O., Verheyen, K., Butaye, J., Jacquemyn, H., Bossuyt, B., & Hermy, M. (2002) Possible effects of habitat fragmentation and climate change on the range of forest plant species. Ecology Letters, 5, 525-530.
Huntley, B., Green, R. E., Collingham, Y. C., Hill, J. K., Willis, S. G., Bartlein, P. J., Cramer, W., Hagemeijer, W. J. M., & Thomas, C. J. (2004) The performance of models relating species geographical distributions to climate is independent of trophic level. Ecology Letters, 7, 417-426.
IPCC (2001) Climate change 2001: Synthesis report - Summary for policy-makers. Intergovernmental Panel on Climatic Change, Cambridge University Press, Cambridge.
Loehle, C. (1998) Height growth rate tradeoffs determine northern and southern range limits for trees. Journal of Biogeography, 25, 735-742.
Parmesan, C., Ryrholm, N., Stefanescu, C., Hill, J. K., Thomas, C. D., Descimoin, H., Huntley, B., Kaila, L., Kullberg, J., Tammaru, T., Tennet, W. J., Thomas, J. A., & Warren, M. (1999) Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature, 399, 579-583.
Pearson, R. G. & Dawson, T. P. (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Global Ecology and Biogeography, 12, 361-371.
Perry, A. L., Low, P. J., Ellis, J. R., & Reynolds, J. D. (2005) Climate change and distribution shifts in marine fishes. Science, 308, 1912-1915.
Root, T. L., Price, J. T., Hall, K. R., Schneider, S. H., Rosenzweig, C., & Pounds, J. A. (2003) Fingerprints of global warming on wild animals and plants. Nature, 421, 57-60.
Skov- og Naturstyrelsen (2005) Handlingsplan for naturnær skovdrift i statsskovene. Miljøministeriet, Skov- og Naturstyrelsen, Copenhagen, Denmark.
Skov, F. & Svenning, J. C. (2004) Potential impact of climatic change on the distribution of forest herbs in Europe. Ecography, 27, 366-380.
Stoltze, M. & Pihl, S. (1998). Rødliste 1997 over planter og dyr i Danmark. Miljø- og Energiministeriet, Danmarks Miljøundersøgelser, Skov- og Naturstyrelsen, Copenhagen, Denmark.
Svenning, J. C. & Skov, F. (2004) Limited filling of the potential range in European tree species. Ecology Letters, 7, 565-573.
Svenning, J.C., Skov, F., & (2005) The relative roles of environment and history as controls of tree species composition and richness in Europe. Journal of Biogeography, 32, 1019-1033.
Søgaard, B., Skov, F., Ejrnæs, R., Nielsen, K.-E., Pihl, S., Clausen, P., Laursen, K., Bregnballe, T., Madsen, J., Baattrup-Pedersen, A., Søndergaard, M., Lauridsen, T. L., Møller, P. F., Riis-Nielsen, T., Buttenschøn, R. M., Fredshavn, J. R., Aude, E., & Nygaard, B. (2003) Kriterier for gunstig bevaringsstatus - Naturtyper og arter omfattet af EF-habitatdirektivet & fugle omfattet af EF-fuglebeskyttelsesdirektivet.- Faglig rapport fra DMU nr. 457. p. 463.
Thomas, C. D. & Lennon, J. J. (1999) Birds extend their ranges northward. Nature, 399, 213.
Thomas, C. D., Cameron, A., Green, R. E., Bakkenes, M., Beaumont, L. J., Collingham, Y. C., Erasmus, B. F. N., de Siqueira, M. F., Grainger, A., Hannah, L., Hughes, L., Huntley, B., van Jaarsveld, A. S., Midgley, G. F., Miles, L., Ortega-Huerta, M. A., Peterson, A. T., Phillips, O. L., & Williams, S. E. (2004) Extinction risk from climate change. Nature, 427, 145-148.
Thuiller, W., Lavorel, S., Araújo, M. B., Sykes, M. T., & Prentice, I. C. (2005) Climate change threats to plant diversity in Europe. Proceedings of the National Academy of Sciences USA, 0409902102.
Walther, G.-R. (2000) Climatic forcing on the dispersal of exotic species. Phytocoenologia, 30, 409-430.
Walther, G.-R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T. J. C., Fromentin, J.-M., Hoegh-Guldberg, O., & Bairlein, F. (2002) Ecological responses to recent climate change. Nature, 416, 389-395.
6 Appendiks I
6.1 Karakteristiske arter
Klik her for at se tabel til afsnit 6.1
6.2 Øvrige arter
| Ud- bre- del- se i Eu- ro- pa |
Hyp- pig- hed i Dan- mark |
B2 - æn- dring i for- hold til i dag |
A2 - æn- dring i for- hold til i dag |
|
| Almindelig Ædelgran (Abies alba) | 17,7% | 59,1% | ||
| Andemadsbregne (Azolla filiculoides) | 6,0% | |||
| Enkelt Månerude (Botrychium simplex)* | 4,3% | 1,5% | -18,3% | -18,3% |
| Hottentotfigen (Carpobrotus edulis) | 2,7% | |||
| Ægte Kastanje (Castanea sativa) | 27,6% | 52,1% | ||
| Hassel (Corylus avellana)* | 70,3% | 82,7% | -9,5% | -21,5% |
| Almindelig Valnød (Juglans regia) | 18,9% | 52,9% | ||
| Laurbær (Laurus nobilis) | 11,4% | |||
| Østrigsk Fyr (Pinus nigra) | 8,1% | 57,1% | ||
| Strand-Fyr (Pinus pinaster) | 18,9% | |||
| Tyrkisk Eg (Quercus cerris) | 19,5% | 42,9% | ||
| Sten-Eg (Quercus ilex) | 15,9% | |||
| Dun-Eg (Quercus pubescens) | 28,8% | |||
| Gul Stenbræk (Saxifraga hirculus)* | 15,3% | 2,3% | -40,6% | -40,6% |
| Skærm-Elm (Ulmus laevis)* | 25,4% | 28,6% | -0,6% | -2,8% |
| Småblomstret Elm (Ulmus carpinifolia)* | 53,3% | 3,8% | 9,2% | 6,2% |
* Arter naturligt forekommende i Danmark
7 Appendiks II
































Version 1.0 Oktober 2006 • © Miljøstyrelsen.