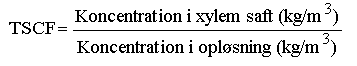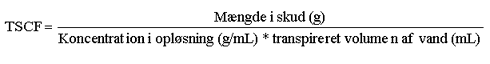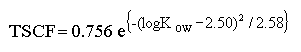|
Fytoremediering af forurening med olie- og tjćreprodukter 2. Generelle aspekter: Planter, mikroorganismer og fytoremediering2.1 Vegetationens effekt pĺ forurenet jord2.1.1 TranspirationUdplantning af trćer og andre planter har flere forskellige effekter pĺ jorden og de kemiske forbindelser, der er her i. Lukket vegetation med trćer har en střrre transpiration end blandinger af urter, grćsser og buske, som vil udvikle sig i de fřrste ĺr af den naturlige succession (Larcher 1995). Střrre transpiration betyder reduceret infiltration (vandring af stof til grundvandet). Det medfřrer ogsĺ et sćnket grundvandsspejl og en mere gennemluftet jord. Evapotranspirations-delen af vandbalancen er omkring 2/3 for en lřvskov i den tempererede zone og omtrent det samme for en nĺleskov (Larcher 1995). Pil groet i det sydlige Sverige evapotranspirerede mellem 365 og 495 mm vand per ĺr, hvilket svarede til mellem 60% og 85% af den gennemsnitlige ĺrlige nedbřr (Perrson 1995). I vinterhalvĺret sker der dog ingen evapotranspiration, da der er tale om lřvfćldende trćer. 2.1.2 AfdampningAfdampning fra jord til luft er en vćsentlig proces for flygtige forbindelser (Jury et al. 1983, 1987, 1990, Sanders & Stern 1994). Afdampningen kan reduceres med vegetation pga. den yderligere modstand, som planter og deres nedfaldne blade og lřv giver mellem jordoverfladen og atmosfćren (Trapp & Matthies 1997). Planteoptag og translokering efterfulgt af afdampning er dog en potentiel rute til at fjerne visse forbindelser fra jord (se afsnit 2.3.6 om fytoafdampning). 2.1.3 Iltforsyning til jordenIltforsyning til jorden anses for at vćre den vigtigste begrćnsende faktor for biologisk nedbrydning af střrstedelen af organiske kontaminanter. Nogle planter ćndrer jordmiljřet ved at skabe mulighed for ilttransport nedad (Grosse et al. 1996). Biologisk forstćrket transport af ilt ned i iltfrie zoner kan vćre ganske effektiv: Tagrřr, Phragmites australis, transporterer op til 11 L O2 m-2 h-1 til sediment (Brix et al. 1996). Bĺndpil, Salix viminalis, er kendt for at kunne gro under forhold med periodisk eller permanent oversvřmmelse (Ellenberg 1979, Jackson & Attwood 1996). Grunden til, at bĺndpil kan gro under disse forhold, er, at piletrćer generelt er tilpasset til iltmangel i jorden ved hjćlp af et ventilerende vćv (aerenchyma), som har store intercellulćre hulrum (op til 60%). Disse porer gřr det muligt for ilten at diffundere nedad. Iltforsyning til rřdderne via diffusion gennem jorden virker kun over korte afstande. Nogle arter, der kan gro i sumpomrĺder (herunder pil, Salix), er i stand til effektivt at pumpe luft til aerenchyma porene ved at udnytte trykforskelle, der er forĺrsaget af solopvarmning (Grosse et al. 1996). Bćvreasp, Populus tremula, har ligeledes evnen til at tilfřre ilt til sine rřdder (Grosse et al. 1992). Denne trćart kan řge ilttilfřrslen med 44% (sammenlignet med ilttilfřrsel via diffusion om natten). Nĺletrćet Taxodium distichum, som vokser i sumpomrĺder, kan řge ilttilfřrslen med 77%, hvorimod lind (Tilia), birk (Betula), ask (Fraxinus) og lřn (Acer) kun kan řge tilfřrslen med mindre end 20%. 2.1.4 RodudsondringerUdsondring fra planterřdder er defineret som frigivelse af substanser fra roden til den omgivende jord, rhizosfćren. Denne frigivelse sker via diffusion langs en elektrokemisk gradient. Det er en almindeligt forekommende proces i alle levende planter (Curl & Truelove 1986). Der er ikke helt overensstemmelse mellem jord-mikrobiologer og botanikere angĺende den prćcise definition af rhizosfćre. Rhizo- kommer fra det grćske ord for rod, men "sfćre" kan fortolkes pĺ flere forskellige mĺder. En mĺde at definere rhizosfćren pĺ er: Det volumen af jord, der sidder fast pĺ rřdderne. Mere specifikt kan den defineres som det lille omrĺde af jorden, der pĺvirkes af tilstedevćrelsen af levende rřdder; en pĺvirkning, der sker via lćkage eller udsondring af substanser, som pĺvirker den mikrobielle aktivitet (Curl & Truelove 1986). Udsondringen kan vćre gavnlig for planten af mange forskellige ĺrsager, heriblandt tiltrćkning af insekter, frastřdning af herbivorer og forsvar mod infektion (Larcher 1995). Udsondringer kan bĺde virke hćmmende og fremmende pĺ mikrobiel vćkst (Curl & Truelove 1986). Den gavnlige effekt opnĺs, f.eks. hvis udsondringen kan stimulere vćksten af de bakterier, som kan nedbryde forurening i rhizosfćrejorden. Dette menes at vćre tilfćldet ved frigivelse af fenol-forbindelser, som fremmer vćksten af PCB-nedbrydende bakterier (Fletcher & Hegde 1995). Rodudsondringer fra planter kan ogsĺ ćndre oplřseligheden af forbindelser, der ellers ville vćre uoplřselige i jorden og fast bundne til partikler i jorden eller til planterřdder. Kompleksbindende molekyler i udsondringen kan danne oplřselige komplekser f.eks. med metalioner (Larcher 1995). Courgetter (Cucurbita pepo) og grćskar (Cucurbita maxima), begge grćskararter, kan oplřse og optage lipofile stoffer som f.eks. polyklorerede dibenzodioxiner og -furaner (Huelster et al. 1994). Ĺrsagen til dette er sandsynligvis et kompleksdannende protein, som findes i rod- og xylem-udsondring og ogsĺ i plantesaft fra courgetter (Huelster & Marschner 1995, Neumann et al. 1996). I vedplanter er der fundet glykosid og glukuron konjugater af 1-hydroxypyren (Nakajima et al. 1996). Konjugater er det normale produkt af fase II reaktioner i planter (Komossa et al. 1995), og mange af dem er oplřselige i jorden. Den ćndrede oplřselighed kan pĺvirke optag og transport ind i planter, men ogsĺ nedbrydning i jord og transport til grundvand (Se ogsĺ afsnit 2.2.1 om biotilgćngelighed). 2.2 Begreberne omkring fytoremedieringFytoremediering er blevet defineret som: „Anvendelsen af grřnne planter og disses mikrobiota, jordtilsćtninger, og agronomiske teknikker til at fjerne, indeholde eller uskadeliggřre uřnskede kemiske stoffer i miljřet " (Cunningham et al. 1996). Det vides, at planter kan optage mange forurenende forbindelser, isćr dem med middel lipofilicitet og/eller svage elektrolytter (Briggs et al. 1982, Briggs et al. 1987, Trapp & Pussemier 1991, Burken & Schnoor 1998, Hsu et al. 1991, Trapp 2000). Fytoremediering kan virke i kraft af mange faktorer, heriblandt vegetationens pĺvirkning af fysiske (vandbalance, transportprocesser), kemiske (redox potentiale, pH, kompleksdannende stoffer) og biologiske (enzymer, rřdder, bakterier og svampe) parametre i jorden. 2.2.1 BiotilgćngelighedEn stor udfordring, der mĺ overkommes i fytoremedieringsprojekter, er at sikre forureningens tilgćngelighed for planter (isćr rřdder) og bakterier (Alexander 1994). 2.2.1.1 Biotilgćngelighed for mikroorganismer Betegnelsen bioremediering er i litteraturen blevet anvendt om processen, hvor mikroorganismer i jorden omsćtter forurening. Bioremediering kan derfor godt vćre en del af fytoremedierings-processerne. For forbindelser med ringe oplřselighed i vand er det diffusive flow af kontaminanter til bakterieceller begrćnset (Koch 1990, Bosma et al. 1997, Harms & Zehnder 1994, Harms 1996, Harms & Bosma 1996, Rijnaarts et al. 1990, Volkering et al. 1993), og optagelsesgraden kan blive ubetydelig, hvis bakterierne ikke er i direkte fysisk kontakt med forureningen. I bioremedieringsprojekter er det blevet forsřgt at forřge bakteriers optag ved jćvnligt at blande jorden rundt og derudover inokulere jorden med et stort antal bakterier. Mikrobiel nedbrydning i rhizosfćrejord har fordele overfor ren bakteriel nedbrydning i utilplantet jord. Af disse fordele kan nćvnes:
2.2.1.2 Biotilgćngelighed for rřdder En forudsćtning for succesfuld nedbrydning i rhizosfćren er et godt rodnet, da roddybden bestemmer den jorddybde, hvori fytoremedieringen kan ske. Sćdvanligvis befinder rřdder sig mest i de řverste jordlag. Det er sjćldent at finde rřdder under 3 meters dybde. Ifřlge Dobson & Moffat (1995) er den gennemsnitlige roddybde for trćer 1–2 m (poppeltrćer 0,8–2,43 m), hvilket er meget mindre end det almindeligvis forventes, og nćsten 90% af antallet af alle trćrřdder befinder sig i de řverste 60 cm af jorden. Ringe gennemluftning af jorden vil medfřre en mindre roddensitet. Selvom piletrćer kan overleve permanent oversvřmmelse og vandmćttet jord, vil rřdderne ikke gro dybere ned i grundvandet (Larcher 1995), og over 60% af de tyndeste rřdder vil ikke ligge dybere end 20 cm (Rytter & Hansson 1996). Der findes dog teknikker, hvormed man kan tvinge rřdderne til at vokse dybere ned i jorden, f.eks. gennem en tildćkning af jorden, hvilket medfřrer udtřrring i de řvre jordlag, eller ved at vikle plastictape omkring stiklinger, fřr de plantes ud (Lars C. Larsen, Hedeselskabet, personlig meddelelse). 'Effektiv roddybde' defineres som dybden med en rodtćthed střrre end 0,1 cm rod/cm3. For pil blev den mĺlt pĺ grovsandet jord og lerblandet sandjord til henholdsvis 65–95 cm og 115–125 cm (Mortensen et al 1998, Jřrgensen & Schelde 2001). Der er dog enkelte rřdder, som gĺr dybere. Iritz (1996) angiver roddybden af pil (ikke nćrmere defineret) til 50 cm i begyndelsen af vćkstsćsonen stigende til 1,3 m per 1. juli. Det er oplagt, at planter med dybtvoksende rřdder ville vćre at foretrćkke til in situ fytoremediering af organisk kontaminering, fordi denne oftest findes flere meter under overfladen. Derudover ville det vćre at foretrćkke at benytte flerĺrige planter. Almindelige trćer, buske eller afgrřde planter ville vćre fordelagtige m.h.t. tilgćngelighed af frř eller stiklinger. Udover roddybden kan rodens struktur vćre en bestemmende faktor for resultatet af en remedieringsproces. I et studie af PAH nedbrydning i rhizosfćrejord blev det konkluderet, at et meget forgrenet rodnet med tynde rřdder tilsyneladende var mere effektivt end pćlerřdder til at řge nedbrydningen (Schwab et al. 1995). Det er muligt at fĺ planter til at řge deres antal af tynde rřdder. Dette er vist med polejmynte (Mentha pulegium). Disse planter blev inokulerede med Agrobacterium rhizogenes (Kalidas Shetty, personlig meddelelse). Det samme er blevet observeret for johannesbrřd trć Robinia pseudoacacia L. (Han et al. 1993). Hĺrfine rřdder blev dannet udfra segmenter inokuleret med Agrobacterium rhizogenes R1601 pĺ under en uge. Opsamlende kan det pĺpeges, at der er en mangel pĺ viden om planters roddybde. Det er mest agronomer, der beskćftiger sig med afgrřde planter som korn og kartofler, der har undersřgt emnet. For trćer peger de fĺ undersřgelser, der findes, pĺ, at de fleste trćers roddybde er yderst ringe for střrsteparten af roden. Dette udelukker dog ikke, at nogle trćarter har rřdder i dybereliggende jordlag. Indtil dette er blevet nřjere undersřgt, mĺ det dog antages, at omkring 2 m dybde er grćnsen for rhizo- og fytonedbrydning. Hydraulisk kontrol er dog mulig til střrre dybder, eftersom flere forskere har pĺpeget, at de har fundet pćlerřdder fra forskellige planter i forbavsende dybder, f.eks. lucernerřdder ned til 18 m (G. Gissel-Nielsen, personlig meddelelse) og tilsvarende dybder for individuelle tilfćlde for andre planter (tabel 1). Tabel 1:
* freatofytisk betyder, at rřdderne kan nĺ grundvandet (Jordahl et al., 1997) Det ser ud til, at planterodens morfologi er kraftigt pĺvirket af miljřmćssige forhold. Nogle af disse faktorer kan favorisere at bringe rřdderne i nćrheden af kontaminering, mens andre faktorer kan have den modsatte effekt. Střrstedelen af de hĺrfine rřdder forventes at vćre i den jordzone, som er fugtig og nćringsrig. Dette er sćdvanligvis de řverste jordlag af landbrugsjord, men ikke nřdvendigvis pĺ forurenet jord. Enkelte pćlerřdder til at sikre vandforsyning mĺ nřdvendigvis nĺ ned til overfladen af grundvandsspejlet, hvilket vil sige ca. 2–20 m, men disse rřdder vil ikke sprede sig sĺ meget ud i jorden. Eftersom det meste forurening forekommer ujćvnt fordelt i jorden, er det muligt, at planterřdderne vil prřve at undgĺ den kontaminerede jord, hvis denne er toksisk for planten. Pćlerřdder vokser dog overvejende i revner og sprćkker pga. den mindre mekaniske modstand. Hovedparten af de i morćneler udsivende stoffer pĺ olie- og tjćreforurenede lokaliteter befinder sig ligeledes i sprćkkerne (Klint & Fredericia 1995, Jakobsen & Klint 1999). Dette břr medfřre, at de dybe rřdder vokser i umiddelbar nćrhed af de dybtliggende jordforureninger. 2.2.2 Rhizosfćre mikrobiotaRhizosfćren er en zone med hřj mikrobiel aktivitet (Cunningham et al. 1996). Bakterier vokser i kolonier, der dćkker 4 til 10% af rodens overflade (Shimp et al. 1993). I rhizosfćren er antallet, diversiteten og střrrelsen af bakterierne střrre end i den řvrige jord, hvilket formodentlig afspejler deres bedre ernćring (Campbell & Greaves 1990). Per vćgtenhed af jord indeholder rhizosfćren af de fleste planter typisk 10 til 20 gange sĺ mange bakterier som den omgivende jord, men der kan vćre helt op til 100 gange sĺ mange (Bazin et al. 1990). Som eksempel pĺ dette er det vist, at mikrobielle koncentrationer af denitrificerende bakterier, pseudomonader og monoaromatiske kulbrinte-nedbrydere var signifikant hřjere i jordprřver fra rhizosfćren af poppeltrćer end i den nćrliggende landbrugsjord (Jordahl et al. 1997). Dette formodes at vćre forĺrsaget af bakteriel vćkst pĺ rodudsondringer (Boyle & Shann 1995), som dannes af unge rřdder. Estimater for den ĺrlige produktion af rhizoaflejring (substanser, der kommer fra henfald af dřdt plantevćv) og rodudsondringer varierer fra 7 til 27% af den totale producerede plantemasse (Moser & Haselwandter 1983). Da rodudsondringer primćrt ikke dannes som respons pĺ stimuli men nćrmere er et resultat af rodcellelćkage, er udsondringernes kemiske sammensćtning udefinerbar og mangfoldig, hvilket afspejler planternes primćre og sekundćre metabolisme. Primćre metabolitter (kulhydrater, aminosyrer) kan forstćrke den uspecifikke heterotrofe mikrobiota (Newman 1985), hvorimod de sekundćre metabolitter (fenoler, terpenoider, steroider) tilsyneladende er i stand til specifikt at stimulere aktiviteten af bakterielle nedbrydere. F.eks. er det vist, at fenoler er i stand til at stimulere aktiviteten af PCB-nedbrydende bakterier (Fletcher & Hegde 1995), og planteafledte terpenoider som l-carvon (der dannes af grřn mynte, Mentha spicata) kan vćre gavnlige for at řge nedbrydning af PCB ved hjćlp af Arthrobacter. Stimuleringen formodes at vćre baseret pĺ strukturel analogi mellem plantens metabolitter og kontaminanter (Gilbert & Crowley 1997, Boyle & Shann 1995, Siciliano & Germida, 1998). Man kunne forestille sig, at bakterielle nedbrydere kunne blive hćmmet af katabolitter i et miljř med god forsyning af kulstof (Yuste et al. 1998). Selvom hřjt bakterietal i rhizosfćren er blevet beskrevet mange gange i litteraturen, stĺr det mindre klart, om den metaboliske aktivitet er hřjere i rhizosfćrejord end i den omgivende jord. Nogle undersřgelser af den cellulćre aktivitet af P. fluorescens har indikeret, at den metaboliske aktivitet hos rodkoloniserende bakterier er lav (sammenlignelig med aktiviteten hos sultede celler) (Normander et al. 1999). I jord vil bakterielle nedbrydere, specielt i sultet tilstand, ikke omsćtte en enkelt kulstofkilde ad gangen, men er derimod i stand til samtidigt at nedbryde sukkerstoffer og miljřfremmede stoffer (Bally et al. 1994, Egli 1995, Kovarova et al. 1997). Matematisk modellering af kulstofstrřmning i en typisk forurenet jord viser, at nedbrydning af forurening alene (pga. den ringe masseoverfřrselshastighed) ikke er nok til at understřtte vćkst og vedligeholdelse af en stor biomasse af mikrobielle nedbrydere (Harms 1999). 2.2.3 Optag af organiske stoffer i planter2.2.3.1 Rodkoncentrationsfaktor, RCF Optag af organiske forbindelser i rřdder afhćnger af egenskaberne for stoffet og planten. Generelt sker optaget hurtigt, indtil ligevćgt er opnĺet. Ligevćgten er blevet beskrevet med rodkoncentrationsfaktoren (RCF) (Shone & Wood 1974):
Enheden for RCF er udfra denne definition: mL oplřsning optaget per g rod. Nĺr optaget sker fra en vandig oplřsning (f.eks. i forsřg med hydroponiske oplřsninger som vćkstmedie), vil RCF for ikke-elektrolytter stige med vćrdien af log KOW. Ved kemisk ligevćgt vil koncentrationerne af lipofile forbindelser i rřdderne vćre tusind gange hřjere end i det omgivende vand. KOW er fordelingskoefficienten mellem n-oktanol og vand (koncentration i oktanol divideret med koncentration i vand). Afhćngigheden af log KOW kan forklares med lipofil adsorption af stoffet til plantens lipider i rřddernes endodermis-celler. Begrebsmćssigt er rodoptag forskellig fra translokering til ledningsvćvet (xylemet) (Shone & Wood 1974). Nĺr optag sker fra jord, vil koncentrationerne i rřdderne sjćldent overstige koncentrationerne i jorden, fordi adsorption til rodlipider konkurrerer med adsorption til humuspartikler. Disse har en adsorptionskapacitet, der ligner rřddernes, hvilket ikke er overraskende, eftersom de er udviklet primćrt af plantepolymere. Ved kemisk ligevćgt vil koncentrationerne i rod og jord derfor vćre ens. Kemisk ligevćgt opnĺs dog kun for de tynde rřdder, ikke for de tykke rřdder eller for rodfrugter som f.eks. gulerod (se figur 5 i afsnit 3.5), idet stofferne ikke trćnger ind igennem de yderste cellelag (Trapp 1995). 2.2.3.2 Transpirations-strřmnings-koncentrations-faktor, TSCF Translokering ind i plantens skud kan beskrives med transpirations-strřmnings-koncentrations-faktoren, TSCF:
Denne faktor er uden enhed, da den beskriver et volumen oplřsning per volumen af xylem-saft. TSCF reprćsenterer ideelt set koncentrationsforholdet mellem xylem-saften og den eksterne oplřsning. Eftersom dette er vanskeligt at mĺle direkte, bestemmes TSCF ofte eksperimentelt ud fra den stofmasse, der translokeres til plantens skud (Shone & Wood 1974):
I denne model er det antaget, at der ikke sker hverken nedbrydning eller andre tab fra skuddene i lřbet af mĺleperioden, og at transporten tilbage til rřdderne er ubetydelig. Hvis dette alligevel foregĺr, er det nřdvendigt at korrigere de eksperimentelle data. Translokeringen af ikke-dissocierbare kemikalier er mest effektiv for forbindelser med middel lipofilicitet (Briggs et al. 1982). En regressionsformel for TSCF for poppeltrćer er udarbejdet af Burken og Schnoor (1998) (vćr opmćrksom pĺ en fejl i originalartiklen):
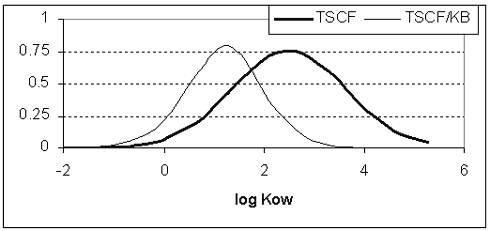
For at fĺ udtrykt koncentrationsforholdet mellem xylem og jord, mĺ man dividere TSCF med fordelingskoefficienten mellem jord og vand, her kaldet KB. Lipofile forbindelser med en hřj log Kow adsorberes til jord og vil ikke oplřses i vand. Derved vil optimum pĺ figur 1 flyttes mod venstre (figur 1).
Figur 1: RCF og TSCF for elektrolytter afhćnger af pH i den eksterne oplřsning (Shone & Wood 1974, Briggs et al. 1987, Rigitano et al. 1987, Inoue et al. 1998). Dette skyldes pH-forskelle mellem den eksterne oplřsning og planteceller. Hvis et molekyle dissocierer i det fysiologisk relevante interval, vil det pĺvirke bĺde optagelseshastigheden og -niveauet. Generelt vil optag af svage baser řges med den eksterne pH, mens optag af svage syrer vil sćnkes. Modellen břr derfor ikke benyttes til dissocierede forbindelser. En model, der kan benyttes for elektrolytter, er blevet publiceret for nyligt (Trapp 2000). 2.2.3.3 Overfřrselsfaktor, (transfer factor) TF Overfřrselsfaktoren, TF, er defineret som koncentrationsforholdet mellem plantevćv pĺ hřsttidspunktet divideret med koncentrationen i vćkstmediet. Ofte er koncentrationen i planter pĺ hřsttidspunktet relateret til den initielle eller den nominelle koncentration i jord eller oplřsning. I sĺ fald kan den reelle overfřrselsfaktor blive underestimeret. Desuden er TF ofte givet for třrvćgt, som er střrre end for vĺd vćgt af planterne. I almindelighed har overfřrselsfaktoren ikke noget at gřre med kemisk ligevćgt, men refererer sćdvanligvis til steady-state situationer (balance mellem optag og frigivelsesprocesser). 2.2.3.4 Hvem nedbryder – plante eller bakterie? Udfra fysisk-kemiske data kan der drages nogle konklusioner angĺende hvilke kemiske forbindelser, der vil blive pĺvirket mest af planters tilstedevćrelse i en forurenet jord. De relativt vandoplřselige forbindelser, som har en relativt hřj TSCF/KB (figur 1), vil forholdsmćssigt hurtigt blive optaget med transspirations-strřmningen, og vil blive fřrt opad mod stćnglen/stammen og bladene. Der kan de enten blive akkumuleret, omdannet, nedbrudt, eller afdampet fra bladene. Sidstnćvnte proces afhćnger af damptrykket. Planter kan ogsĺ pĺvirke de mere lipofile forbindelser, som ikke fřres op i planten. Disse forbindelser kan i stedet blive adsorberet til eller optaget i rřdderne, forudsat at forbindelsen er biotilgćngelig. I sĺ fald vil plantens metabolisme konkurrere med mikroorganismernes. Hvis man antager, at biomassen af rřdder og mikroorganismer er omtrent ens, bliver det et spřrgsmĺl om nedbrydningshastigheder, hvilken organisme, der vil dominere nedbrydningsprocesserne. Hvis nedbrydningen inden i planten er hurtigere end den mikrobielle nedbrydning i jorden, vil der vćre en konstant koncentrationsgradient mellem inder- og ydersiden af rodcellerne. De forurenende molekyler vil diffundere langs denne gradient, enten i vandfasen eller i gasfasen. Diffusion i gasfasen er ca. 10000 gange hurtigere end i vandfasen. Derfor, hvis fordelingskoefficienten mellem luft og vand, KAW, er střrre end 10-4, vil diffusion i gasfasen normalt dominere transporten. Der kan kun siges lidt om nedbrydningshastigheden i planterřdder sammenlignet med hastigheden for mikrobiel omsćtning. Ĺrsagen til dette er, at de rent eksperimentelt er sĺ godt som umulige at skelne. Under reduceret ilttryk (anoxiske forhold) vil plantens metabolisme ikke spille nogen rolle. Planter kan overleve under sĺdanne forhold i en hviletilstand, men de er ikke metabolsk aktive (Jackson & Attwood 1996). Tabel 2 viser nogle tilfćlde, hvor det kunne dokumenteres, at enten planter eller bakterier forĺrsagede nedbrydningen. Tabel 2:
2.2.4 Erfaringer med fytoremedieringDen lange tabel, der udgřr bilag C viser udvalgte resultater fra fytoremedieringsprojekter med organisk forurening, isćr med vćgt pĺ PAH’er, olie-kulbrinte og forbindelser i bilbrćndstof. Den angivne mekanisme, hvis der er angivet en, kan godt vćre én ud af flere. 2.3 Forskellige fytoremedieringsteknikkerAdskillige fytoremedieringsteknikker er blevet beskrevet (EPA 2000, Lombi & Wenzel 1999, Flathman & Lanza 1998): Fytoekstraktion, rhizofiltrering, fytostabilisering, rhizonedbrydning, fytonedbrydning, fytoafdampning, hydraulisk kontrol, vegetativt dćkke og bufferstriber. Alle disse teknikker er baseret pĺ ekstraktion, tilbageholdelse i planten, nedbrydning eller en kombination af disse. Beskrivelserne af de forskellige teknikker břr ikke opfattes som fuldstćndigt forskellige, da der kan forekomme overlap af betydninger. 2.3.1 FytoekstraktionBetegnelsen for optag (translokering) og akkumulering af forurenende forbindelser er fytoekstraktion. Denne teknik anvendes mest til oprensning af tungmetal. Kontaminanterne fjernes fra en forurenet lokalitet, nĺr planterne, som har groet i den forurenede jord, hřstes (EPA, 2000). 2.3.2 RhizofiltreringRhizofiltrering involverer adsorption af kontaminanter til rřdder, eller deres udfćldning i rodzonen. F.eks. kan tungmetaller eller lipofile forbindelser ekstraheres fra vand med denne teknik (EPA, 2000). 2.3.3 FytostabiliseringFytostabilisering beskriver immobilisering af forbindelser i jorden, eller stabilisering af jorden i sig selv for at hindre jorderosion. Immobilisering kan gřres ved at fjerne transportvejene: vand eller vind. Immobilisering kan ogsĺ opnĺs ved, at kontaminanterne kemisk ćndres fra en oplřselig form til en uoplřselig (udfćldning i rodzonen). Kontaminanterne kan ogsĺ blive bundet til jordens humuspartikler eller til lignin i plantecellevćggen. Endelig kan immobiliseringen skyldes en adsorption til planterřdder (EPA 2000, Cunningham et al. 1996). 2.3.4 Hydraulisk kontrolEn sćrlig slags fytostabilisering, som ikke inkluderer ekstraktion af forurenende forbindelser fra jorden, er hydraulisk kontrol. Denne teknik bruges til at pĺvirke niveauet eller bevćgelsen af grundvand, overfladevand eller vand i jorden. Hydraulisk kontrol er fjernelse af vand via plantens normale optag, for at hindre udsivning, vandring og dermed spredning af kontaminering (EPA 2000). 2.3.5 "Pump and tree"En af de almindeligt anvendte remedieringsteknikker til grundvand er "pump og rens" (pump and treat): Oppumpning af vand med efterfřlgende teknisk rensning (stripning, adsorption, bioreaktor osv.). En ny idé er at bruge det oppumpede vand pĺ tidspunkter med negativ vandbalance (sommer) til vanding f.eks. af et skovomrĺde. Teknikken er ogsĺ blevet benćvnt "Trćmediering" (Gatliff 1994). En hektar piletrćer kan transpirere op til 3000 m3 vand gennem juli mĺned (Larcher 1995). Skovjord har en hřj metabolsk kapacitet og kan mĺske nedbryde mange forbindelser relativt hurtigt. Derfor kan i det mindste noget af vandet renses ved hjćlp af trćer. Kemikalier, som kan oprenses fra grundvand pĺ denne mĺde, er organiske oplřsningsmidler (f.eks. triklorethen), MTBE, olieprodukter, nćringsstoffer og muligvis flere endnu. Nogle forhold er endnu ikke afklarede. F.eks. kender man ikke den optimale dosering af vand (skovdřd forĺrsaget af toksiske effekter břr undgĺs). 2.3.6 FytoafdampningEfter optag i planter kan kontaminanterne i nogle tilfćlde (afhćngig af forbindelse og plante) blive transporteret til plantedele over jorden, blive metaboliseret, opbevaret eller fordampe fra bladene (Trapp 1995). Processen har vist sig relevant for m-xylene (Trapp & Christiansen 2001), klorbenzen (Baeder-Bederski et al. 1999), triklorethen (Orchard et al. 2000) og andre flygtige forbindelser (Burken & Schnoor 1998), og selv organisk bundet kviksřlv (EPA 2000). Teknikken er relevant for alle forbindelser, som hurtigt translokeres (log KOW < 3.5), og som har et hřjt damptryk eller en hřj Henrys lov-konstant (enhedslřs KAW >> 10-5). Afdampning flytter kun forureningsproblemet fra et sted til et andet, og den kan derfor opfattes som en uřnsket sideproces. De resulterende koncentrationer i atmosfćren er dog normalt ikke over baggrundsniveauet i byomrĺder (Trapp et al. 2000a). Translokering gennem planten, og dermed ogsĺ afdampning, sker hovedsageligt, nĺr solen skinner, hvilket giver mulighed for fotolyse. 2.3.7 FytonedbrydningFytonedbrydning er metabolisme i planter. Omsćtningen kan ske internt i planten, efter at forbindelsen er optaget fra jorden. Den kan ogsĺ ske eksternt, eftersom bl.a. enzymer, der produceres af planten, kan muliggřre nedbrydning ude i jorden (Cunningham et al. 1996). Velegnede kandidater til fytonedbrydning er moderat hydrofobe, organiske forbindelser med log Kow mellem 0,5 og 3, da disse forbindelser let optages og translokeres i planter (Schnoor et al. 1995, Cunningham et al. 1997). Identificering af metabolitter i planten bekrćfter, at omsćtning af forureningen er en realitet. 2.3.8 RhizonedbrydningRhizonedbrydning er defineret som nedbrydningen af organiske forbindelser i jorden gennem mikrobiel aktivitet, som er forřget pga. rodzonen (EPA 2000). Rodudsondringer fra planter pĺvirker (og řger i flere tilfćlde) den mikrobielle aktivitet i nćrheden af rřdderne. De fleste planter lever i symbiose med bakterier og/eller svampe. Mikroorganismetćtheden omkring rřdderne er meget střrre end i jord uden rřdder (Crowley et al. 1997). Mikroorganismer kan ogsĺ migrere til dybereliggende jordlag, mens de sidder fast pĺ planterřdder. Fyto- og rhizonedbrydning anvendes til oprensningen af organisk kontaminering, heriblandt forurening med olie, PAH, BTEX, TNT, klorerede oplřsningsmidler og pesticider (EPA 2000, Newman 2000). 2.3.9 Andre teknikkerEn rensningsmetode, der er baseret pĺ nedbrydning i rodzonen, er landfarming med bestemte afgrřder. Denne metode kan f.eks. bruges til olieforurenet slam. Slammet hćldes pĺ jorden, marken gřdes og lucerne, Medicago sativa, eller grćs (som regel rajgrćs, Lolium) sĺs ud. Affaldsdepoter er ofte tilplantet med grćs, bĺde fordi det ser bedre ud, men ogsĺ for at hindre erosion (altsĺ en form for vegetativt dćkke). Andre gavnlige aspekter af tilplantningen: Forurening af grundvand reduceres, og mindre mćngder udslippende gas kan adsorberes af planterne (střrre mćngder gas er toksiske for planter) (EPA 2000). Kun i fĺ tilfćlde tillader myndighederne at tilplante med trćer, pĺ trods af at trćer kan transpirere mere. Det frygtes, at rřdderne kan gennemtrćnge dćkkende membraner under affaldsdepoter og derved skade dem. I fřlge Dobson og Moffat (1995) er der ingen basis for disse bekymringer. Tilplantning med trćer langs med floder som bufferstriber er ikke nogen ny idé. Det vides, at disse striber skaber barrierer for aflřb af nćringsstoffer fra landbrugsjord (EPA 2000). 2.4 Kriterier for anvendelse af fytoremedieringEn af ĺrsagerne til, at fytoremediering ikke er sĺ kendt, kan vćre, at teknologien er ny. Remedieringen krćver mange ĺr eller mĺske tiĺr. Der kan ikke siges at vćre mange gode resultater, fordi střrstedelen af projekterne er startet for relativt kort tid siden. Omkostningerne lader sig heller ikke endeligt evaluere, selvom der eksisterer oplysninger for nogle teknikker, der kan sammenlignes med konventionel behandling (EPA 1998). For tungmetalforurenet jord er omkostningerne ved fytoremediering omkring 1/3 af konventionelle teknikker, for bly under 1/20 og for olie under 1/10 (EPA 1998). Mere end halvdelen af omkostningerne skyldes monitering af jord, grundvand og vegetation. Til at beslutte, om fytoremediering kan vćre anvendelig i praksis pĺ en bestemt lokalitet med en bestemt type forurening, er der udviklet en metode til at lette overvejelserne. Resultatet er fytoremedierings-beslutningstrćet, se figur 2 (ITRC, 1999). I mange tilfćlde er det nřdvendigt at mĺle pĺ fytotoksicitet, planteoptag og metabolisme, fřr et feltforsřg igangsćttes. Dette krćver forudgĺende eksperimenter udfřrt i laboratoriet. Der er blevet udviklet en hurtig 4-trins test til dette formĺl. Testen medtager optag og toksicitetstest for trćer (pil og poppel) i forurenet jord eller i oplřsning (Trapp et al. 2000), en 14C-metabolisme-test, samt en model for langtidsprognose (efter Trapp et al. 1994). Figur 2: 2.5 RisikovurderingVed fytoekstraktion kan organisk forurening eller metabolitter heraf akkumulere i forskelligt plantevćv. Der mĺ trćffes foranstaltninger for at hindre spredning af forurening gennem fřdekćden, hvilket f.eks. kan ske, nĺr dyr spiser af vegetationen, der bruges til fytoremediering. Dette problem er dog kun relevant ved meget store fytoremedieringsprojekter. I et forsřg (Qiu et al. 1997) blev det dog konstateret, at PAH’erne opkoncentreres i planternes rřdder og ikke translokeres til plantedele over jorden. Derfor var der i dette tilfćlde ingen fare for opkoncentrering af PAH’er eller metabolitter gennem fřdekćden (i det mindste med grćsser) via grćssende dyr. Et andet problem kunne vise sig ved anvendelsen af trć fra fytoremedieringsprojekter til brćndsel. Visse forbindelser kunne tćnkes at blive emitteret til luften eller spredt med asken. Fytoremediering krćver tid. Derfor er det nřdvendigt at vurdere, hvad der kan ske pĺ en given lokalitet, indtil planterne er groet tilstrćkkeligt til at kunne stoppe spredning af kontamineringen (grundvandet). Under nedbrydning dannes metabolitter, og ikke alle af disse er uskadelige. Der kan ogsĺ dannes mere vandoplřselige og dermed mere biotilgćngelige forbindelser. Dette aspekt břr overvejes, inden feltanvendelse igangsćttes. Nĺr der kan ske afdampning fra plantens blade, kan det tćnkes, at giftige forbindelser frigives til atmosfćren i uřnskede koncentrationer. Herved ville en forurening blive spredt mere, end den vil blive begrćnset. Toksicitetstest kan hjćlpe med at fastslĺ, om toksiske metabolitter dannes. Test af denne type er udviklet pĺ Berlins Universitet (projektbeskrivelse kan ses pĺ http://dechema.de/biotech/brosch/einleit.htm), men ogsĺ pĺ Institut for Miljřteknologi pĺ Danmarks Tekniske Universitet (Ole Kusk, Řkotoksikologi). 2.6 Ideer til at gřre fytoremediering mere effektiv2.6.1 Genetiske modifikationerAnvendelsen af genteknologi afhćnger af de naturligt forekommende mikroorganismer og deres evne til at kolonisere planterřdder. Rodkolonisering er et kritisk trin i spiringsfasen, hvilket skyldes, at nĺr rodoverfladen én gang er blevet koloniseret med en bestemt type bakterie, kan andre typer blive ekskluderet (David Dowling, personlig meddelelse). Tilfřrsel af specifikke bakterier, der kan nedbryde miljřfremmede stoffer, kan vćre en strategi for at gřre fytoremediering mere effektiv. En udvidet strategi kunne vćre at isolere, oprense og karakterisere specifikke gener, der er ansvarlige for nedbrydningen. Det pĺgćldende gen kunne herefter indfřres i en vćrtsorganisme, som efterfřlgende blev podet pĺ overfladen af steriliserede frř. Dette forudsćtter, at nedbrydende mikroorganismer kan identificeres, og at de er lige sĺ gode til at kolonisere rřdder som de typiske rodkoloniserende bakterier, der bruges som mikrobiel plantebeskyttelse i landbruget. Til dato er der kun begrćnset viden om fytoremediering med anvendelse af kendte rhizosfćre mikrobiota. PCB-nedbrydende gener er blevet indsat i en rodkoloniserende Pseudomonas fluorescens ved at opnĺ ekspression af genet i rhizosfćren af en sukkerroe, Beta vulgaris (Brazil et al. 1995). For nyligt er det blevet vist, at det bionedbrydende potentiale af organiske forbindelser er řget for P. putida (pWW0) i rhizosfćren (E. Duque og J.L. Ramos, personlig meddelelse). Et humant gen, der i leveren koder for et sćrligt enzym, er blevet indfřrt i tobak, Nicotiana tobacum.Enzymet oxiderer flere forskellige halogenerede, organiske forbindelser, heriblandt TCE, ethylen bromid, tetraklorkulstof, kloroform og vinylklorid. Transgene tobaksplanter var i stand til at nedbryde TCE 640 gange hurtigere end ikke-modificerede planter (Doty et al. 2000). Macek et al. (2000) peger ligeledes pĺ anvendelsen af genmodificerede planter for at řge effektiviteten ved fytoremediering. Stategier for modifikationer vćre transformerede planter, der danner specifikke proteiner, der kan binde og transportere xenobiotika. Man kunne ogsĺ forsřge at řge plantens udsondring af f.eks. enzymer, der i sig selv kan nedbryde xenobiotika; man kunne ogsĺ forsřge at řge udsondring af forbindelser, der stimulerer bakteriel vćkst i rhizosfćren og dermed rhizonedbrydning. I EU projektet ENDEGRADE (ENdophytic DEGRADEr bacteria for improving phytoremediation of organic xenobiotics: Endofytiske nedbrydende bakterier til at forbedre fytoremediering af organiske miljřfremmede forbindelser), bliver endofytiske bakterier modificeret til at udtrykke gener ansvarlige for nedbrydning af forbindelser, der kan translokeres via xylem. Endofytiske bakterier er dem, som lever inden i planter. For yderligere information om projektet, se websitet: http://www.ENDEGRADE.dmu.dk Muligvis er det slet ikke nřdvendigt med genetiske modifikationer. Kun en meget lille del af alle planter (omkring 400000 karplanter kendes i dag) er blevet undersřgt for deres evner til at omsćtte kemiske forbindelser. Planters sekundćre metabolisme varierer fra familie til familie, fra art til art og endda mellem varieteter (Frohne & Jensen 1985), og der findes et enormt antal forskellige enzymer. Det mĺ derfor forventes, at nĺr flere planter er blevet undersřgt, vil man finde flere plantearter, som kan vćre egnede til fytoremediering. 2.6.2 Nedbrydning ved hjćlp af svampeAnvendelse af svampe til at nedbryde forurening er blevet detaljeret studeret. Med svampe udnyttes kapaciteten af uspecifikke ekstracellulćre enzymer til at angribe xenobiotika. Indtil videre ser det ud til, at der er mulighed for dannelse af uřnskede, toksiske metabolitter, polymeriserede slutprodukter, og desuden ufuldstćndig mineralisering af kontaminanter (Aiken & Logan 1996, Dietrich et al. 1995, Öberg et al. 1990, Svenson et al. 1989, Lamar & Dietrich 1990). Dette tyder pĺ, at der er behov for yderligere grundlćggende forskning pĺ nuvćrende tidspunkt. Der er ved laboratorieforsřg opnĺet gode resultater med PAH nedbrydning vha. hvidmuldsdannende svampe fremdyrket pĺ halm. Disse svampe udskiller enzymer, der i svampenes naturlige miljř, trć, nedbryder lignin. Undersřgelserne har vist, at det er svćrt at overfřre resultaterne fra laboratoriet til stor skala, da det var vanskeligt at opnĺ tilstrćkkelig vćkst af de fleste hvidmuldsdannende svampe. Dette skyldes dels, at de er relativt langsomt voksende, og dels at der uundgĺeligt i denne skala kommer vćkst at skimmelsvampe i halmen. Sidstnćvnte kan yderligere medfřre forringet vćkst af de hvidmuldsdannende svampe (Miljřstyrelsen 1998c). I Tyskland evalueres muligheden for at nedbryde TNT in situ med svampe (Basidiomycetes). De fřrste forsřg viste, at svampene brugte TNT som kulstof- og nitrogenkilde og derefter sultede. Nye forsřg udfřres med symbiotiske svampe. De symbiotiske partnere, f.eks. hyld, Sambucus nigra, og skovfyr, Pinus sylvestris, er blevet udplantet (personlig meddelelse, Dr. W. Heyser, se ogsĺ http://dechema.de/biotech/brosch/einleit.htm). Resultaterne forventes at ligge klar til nćste ĺr, og de vil blive publiceret i Umweltwissenschaften und Schadstoff-forschung (UWSF) i fytoremedieringsserien, som redigeres af U. Karlson og S. Trapp.
|