|
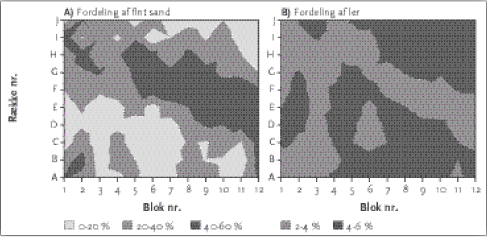
Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetation 3 Eksperimentel adskillelse af herbicid- og N-påvirkning3.1 Materiale og metoder Den eksperimentelle adskillelse af herbicidbehandling og N-tilførsel afspejler til en vis grad et successionsstudie under forskellige påvirkninger af N og herbicid. I relation til hegnsundersøgelserne kunne forsøgets tre første år afspejle billedet af naturlig indvandring af plantearter i nyplantede hegn på en dybdepløjet jord med forskellige grad af landbrugspåvirkning. I eksperimentet øgede vi dog 'successionshastigheden' ved at udså en lang række arter for hurtigt at få en ensartet vegetation til statistiske sammenligninger. Formålet med undersøgelsen var imidlertid ikke at belyse ændringerne over tid, så der vil for vegetationens vedkommende blive fokuseret på resultaterne efter tredje år. For leddyrenes vedkommende præsenteres resultater for alle tre år (2001-2003), men dog med størst fokus på 2003. 3.1 Materiale og metoder3.1.1 ForsøgsetableringEn eksperimental vegetation blev etableret på nærings- og kalkfattig tør sandjord ved Ebdrup på Djursland. Marken havde tidligere været anvendt til korndyrkning, men havde ligget brak et år. Jorden blev dybdepløjet (60 cm dybt), harvet og tromlet for at begrave eksisterende frøbank fra pløjelaget. 120 permanente prøvefelter á 7x7m med 1,4m mellem prøvefelterne blev udlagt med 9m bufferzone rundt om eksperimentet, der er omgivet af trævækst på alle fire sider. 10 replikater af 12 unikke behandlinger blev etableret i et randomiseret blok design (Mead 1990). Behandlingene var 0, 1, 5 og 25 % af markdosis af glyphosat og 0, 25 og 100 kg N/ha/år. For at undgå begrænsning af andre næringsstoffer fik alle felter hvert år i maj tilført 53 kg Fosfor, 141 kg Kalium, 50 kg Svovl og 0,7 kg Kobber /ha/år. Glyphosat (RoundupBio, glyphosat-isopropylaminsalt, 360 g a.i./l) blev første gang udbragt efter etablering af tæt vegetation 24. august 2001 med vindhastighed på ca. 2 m/s, 50 cm over jordoverflade. Sprøjtebommen var 3 m bred med 50 cm mellem dyserne (Lurmark Lo-drift LD 015 Green) med et tryk på 2.0 bar. Anbefalet markdosis er ifølge producentens vejledning 4-6 l/ha. Vi valgte at bruge 4 l/ha (1440 g a.i./ha) som udgangspunkt for 0, 1, 5 og 25 % markdosis som resulterede i hhv. 0, 4, 20 og 100 ml glyphosat pr. 15 l vand. Anden og tredje glyphosatdosering fandt sted i april 2002 og 2003. 3.1.2 Artssammensætning af eksperimentel vegetationI alt blev 30 arter med forskellige funktionelle egenskaber udsået. Udvælgelsen af arter er foretaget så objektivt som muligt med fokus på, at det skulle blive en græsdomineret vegetation, hvor de vigtigste livsstrategier (funktionelle grupper af planter) var repræsenteret. Denne sammensætning blev suppleret med enkelte kendte herbicid- og/eller nitrogenfølsomme arter, arter i tilbagegang i agerlandet og en sjælden art. Endvidere har aktuel forekomst i læhegn/græsland og tilgængelighed af kommercielle frø spillet en rolle Udfra en bruttoliste på 255 arter blev der indhentet information om arternes livsstrategier. Karplanternes livsstrategier kan forklares med tre funktionelle syndromer, som betegnes CSR-strategier (Konkurrence-, Nøjsomheds- og Pionersyndromet): C for konkurrence (competition) tolerance og S for stresstolerante og R for ruderat/pionertilpassede eller forstyrrelsestolerante arter (Grime 1988, 1997; Ejrnæs 2000). Der findes ”rene” C-strateger som fx alm. kvik, men de fleste arter er en kombination af to eller tre syndromer. CSR-strategierne gives en værdi fra 0-12 for alle tre syndromer, hvor den maksimale sumværdi er12. For at få repræsenteret forskellige strategier i den udsåede vegetation blev arterne udtrukket stratificeret tilfældigt i de forskellige grupper, hvor der først udvalgtes en græsart for hver strategi for at sikre græsdominans. Derefter udvalgtes øvrige urter i de forskellige strategier. 3.1.3 Udsåning og etablering af vegetationSpiringsevnen hos de udvalgte arter blev testet eksperimentelt i petriskåle før udsåning. På trods af at marken stort set var plan og tilsyneladende meget homogen, opstod der en vis vand- og vinderosion efter dybdepløjning og såning. 3.1.4 Dataindsamling3.1.4.1 Jordbundsdata Prøverne for 01 og 03 blev analyseret for pHH2O, ledningsevne, glødetab samt total N og P. Frisk jord extraheredes for uorganisk N og P i 1 time i 50 ml 0,5 M K2SO4. Ekstraktet blev filtreret gennem Whatman GF-D-filter og nedfrosset til analyse for NH4+-N med indophenol metoden, NO3--N med cadmium-reduktionsmetoden og PO4-P med molybdæn-blå-metoden (Allen 1989). PO4-P ekstrationen var meget lav og er derfor udeladt. En delmænge af jorden tørredes ved 70 grader C og blev knust. 4 gram heraf blev brændt ved 550 grader C. En anden fraktion (200 mg tørvægt) blev opløst i svovlsyre i en time for analyse af total N og P (Kedrowski 1983) med Hitachi U-2000 spektophotometer. C-total måltes med LECO CNS-1000 med metoden efter Sørensen & Bülow-Olsen (1984). Kornstørrelsesfordeling blev foretaget af Forskningscenter Foulum og de kemiske analyser af Økologisk Kemisk Laboratorium, Botanisk Institut, Københavns Universitet. 3.1.4.2 Vegetation 3.1.4.3 Leddyr 3.1.5 Databehandling3.1.5.1 Vegetation DCA analysen af den eksperimentelle vegetation blev foretaget med følgende nødvendige forholdsregler, som anbefalet af Økland (1990). I alt 20 sjældne arter blev udeladt (forekom i mindre end 5 prøvefelter), ét prøvefelt blev udeladt (< fem arter til stede). Efter første DCA kørsel skilte fire prøveflader sig klart ud ift andre felter med > 1 s.d. enhed. De fire outliers blev fjernet fra analysen som anbefalet af Økland (1990). Der indgår derfor i alt indgår 115 prøvefelter (af 120 i alt) og i alt 29 arter. Ordinationen blev foretaget i PC-ORD (McCune & Mefford 1999) med standardopsætning. Kendall Tau-korrelationer blev beregnet for diskrete variable og produkt-moment korrelationer for kontinuerte variable. DCA-plottene er afbildet med middelværdien for en behandlingsgruppe som centrum (radius = 1 S.E.) i en cirkel. CCA-analyse blev foretaget på samme datamatrice med CANOCO version 4.02 (ter Braak & Smilauer 1997-99) efterfulgt af fortløbende udvælgelse og Monte Carlo permutationstest for at vise den relative betydning af de enkelte variable og hvorvidt de forklarende variable afviger fra permuterede tilfældige variable (Sokal & Rohlf 1995). Der blev anvendt 999 permutationer uden restriktioner. De forskellige variable blev standardiseret for at sikre ens vægtning af alle variable. For at analysere responsen af enkeltarter blev der lavet en indikator-artsanalyse (INSPAN) (Dufrêne and Legendre 1997) i PC-ORD. INSPAN blev udført på tre undergrupper af data, hvor enten herbicidbehandling eller N-tilførsel blev holdt konstant pga signifikante interaktionseffekter af de to faktorer i variansanalysen. 3.1.5.2 Leddyr Det fulde datasæt, hvor tæger, jordlopper og snudebiller er bestemt til art, blev undersøgt vha. den multivariate programpakke PRIMER version 5.2.9 (se 1.3.1). Data for jordbunds- og plantevariable blev inddraget som forklaringsvariable. Data blev om nødvendigt transformeret inden analyserne (primært for at bringe alle variable til nogenlunde samme størrelsesorden). Den indbyrdes korrelation mellem de tilgængelige data for plantebiomasse og –diversitet samt jordbundsforhold blev undersøgt, og for stærkt korrelerede variables vedkommende blev en enkelt af disse valgt ud. PI en 120120 matrice blev korrelationen mellem artssammensætningen og de udvalgte miljøvariable (relateret til jordbund og planter) undersøgt vha. BIOENV. Ved hjælp af MDS blev fremstillet plots af artssammensætningen i relation til behandlingsvariablene (glyphosat og kvælstof). To-vejs ANOSIM blev anvendt til at teste, om artssammensætningen af leddyrene var signifikant forskellig mellem prøvefelter behandlet med forskellige doser glyphosat og kvælstof. SIMPER blev brugt til at undersøge de enkelte arters bidrag til forskelligheden i artssammensætningen mellem prøvetagningsfelter med forskellig behandling undersøgt. Alle tests er vurderet på et 5 % signifikansniveau. 3.2 Resultater3.2.1 Jordbund og blok-effekterEn variansanalyse på jordbundskarakteristika viste signifikante effekter af kvælstoftilførslen på en række målte jordbundskarakteristika (ledningsevne, pH, NO3 og NH4 og uorganisk N). Det kunne dermed konstateres, at N-tilførslen har givet forskelle i tilgængeligt næringstofmængder. Endvidere viste der sig visse forskelle i kornstørrelsesfordelingen i forsøgsområdet (Figur 3.1).
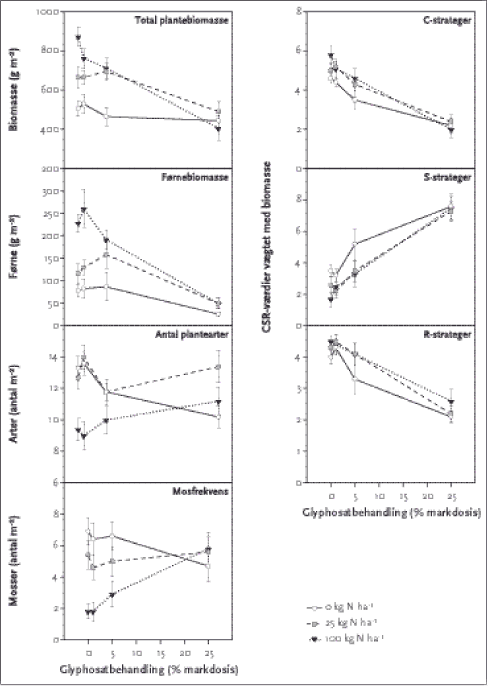
Figur 3.1. Fordeling af fint sand (venstre) og ler (højre) i forsøgsarealet. Langs 'x- aksen' er de 12 behandlinger fordelt på de 10 tilfældigt placerede replica på 'y-aksen' 3.2.2 Etablering af plantearterI Bilag B findes fotos af den eksperimentelle vegetation. Tre udsåede arter (langbladet vortemælk Euphorbia esula, almindelig agermåne Agrimonia eupatoria og almindelig torskemund, Linaria vulgaris) spirede aldrig i petri-skålene, men blev etableret i felten. Fire måneder efter såning i 2001 var 23 ud af de 30 (77 %) udsåede arter etableret. Efter 3 vækstsæsoner var over 90 % af de udsåede arter etableret og alle fire grupper (C, S, R og blandet) af plantestrategier var vel repræsenteret (Tabel 3.1). Tabel 3.1. De udsåede arters højde, etablering og total biomasse efter 3 vækstsæsoner. (C = Competitiv strategi, S = Stress tolerant strategi, R = Ruderat strategi) ifølge Grime (1988) og Ejrnæs and Bruun (2000). Højde iflg. Lid (1987). Frøspiring er procent spiring af 20 frø i petriskål efter 59 dage (n=2). Etablering er udtrykt som procent af prøvefelter med succes fire mdr. og 2,5 år efter udsåning. De udsåede arters relative dominans er udtrykt som % af total høstet biomasse. Mindst 3 arter fra hver gruppe blev etableret i min. 40 % af felterne efter 4 mdr. En græsart fra alle fire funktionelle grupper etablerede sig, men kvik (Elytriga repens) blev dog dårligt etableret. Dette blev delvist kompenseret ved en kraftig etablering af stortoppet hvene (Agrostis gigantea med en CR strategi), der var iblandet frø af alm. hvene (A. capillaris). De tre dominerende græsser udgjorde 86 % af den totale høstede biomasse. Af de enårige blev almindelig dværgløvefod (Aphanes arvensis) aldrig etableret, mens enårig rapgræs (Poa annua) og kornblomst (Centaurea cyanus) blev etableret, men forsvandt stort set igen efter 2,5 år. Derimod har to andre enårige, salomons lysestage (Lepidium campestre) og mark-forglemmigej (Myosotis arvensis), formået at forblive omend kun som en sporadisk del af biomassen. 31 arter indvandrede til prøvefelterne, heraf 11 mosser, men ingen af de indvandrede arter formåede at dominere vegetationen (99,5 % af biomassen stammer fra de udsåede arter). De fem mest almindelige mosser var Ceratodon purpureus (104 prøvefelter), Bryum sp. (24 prøvefelter), Brachythecium rutabulum (17 prøvefelter), Ditrichum heteromallum (9 prøvefelter) og Polytrichyum piliferum (8 prøvefelter). 3.2.3 Klassisk analyse af vegetationsdataResultater af variansanalysen på vegetationsdata viste en signifikant interaktion af herbicidbehandling og N-tilførsel (Tabel 3.2). Interaktionen bestod i, at forskellene mellem N-niverauerne mindskedes med øget herbiciddosering, eller sagt på en anden måde, at herbicideffekten (skønt overordnet set ikke signifikant) steg med N-tilførslen (Figur 3.2). For førnemængden var der ligeledes signifikant interaktion mellem herbicid- og kvælstofeffekterne (Tabel 3.2). Der var en markant stigning i førneproduktionen med øget N- tilførsel ved lave herbicidtilførsler, og ligesom for den totale biomasse øgedes herbicideffekten ved stigende N-tilførsel (Figur 3.2). Tabel 3.2. Resultater af variansanalyse (Generaliseret lineær model LME -Linear Mixed Effect model) for vegetation på de eksperimentelle forsøgsfelter som funktion af herbicid behandling x produktivitet (N-tilførsel). Fed angiver statistisk signifikans. Ved signifikant interaktion mellem herbcid- og kvælstofeffekt giver det ikke mening at se på de separate effekter.
For karplantediversiteten bestod interaktionen mellem de to variable i, at artsantallet faldt med øget N-tilførsel, når der ingen herbicidtilførsel var, mens N-effekten faldt ved øget herbiciddosering (Figur 3.2). Sagt på en anden måde var der en negativ effekt af øget herbiciddosering i de ugødede felter, mens der ved 25 kg N/ha stort set ingen herbicideffekt var, og ved 100 kg N/ha var der en positiv effekt af øget herbiciddosering på antallet af karplantearter. Mosfrekvensen var signifikant påvirket af inteaktionen mellem N-tilførsel og herbiciddosering (Tabel 3.2, Figur 3.2), idet der i de ugødede felter var en negativ effekt af øget herbicidpåvirkning (især 25 % markdosis), mens der ved 25 kg N/ha ingen herbicideffekt var, og ved 100 kg N/ha var der en positiv effekt af herbiciddoseringen på mosfrekvensen.
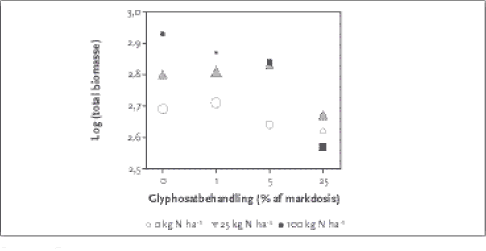
Figur 3.2. Effekter af glyphosatbehandling (markdosis 1440 g a.i./ha) på totalbiomasse, førneproduktion, antal karplanter og mosfrekvens samt effekter på konkurrenceplanter (C-strrateger), nøjsomhedsplanter (S-strateger) og ruderatplanter (R-strateger) beregnet som vægtet biomassse ved tre niveauer af N-tilførsel (0, 25 og 100 kg N/ha/år). For de tre hovedgrupper af plantestrategier blev der fundet en signifikant effekt af både herbicidbehandling og N-tilførsel på den fremherskende plantestrategi (Tabel 3.2). Herbicideffekten var for alle grupper uafhængig af N-tilførslen. Den biomassevægtede C- og R-værdi faldt med øget herbiciddosering, mens S-værdien steg (Figur 3.2). Det betyder, at øget pesticidanvendelse giver lavere C- og R-værdier; men højere S-værdi i plantesamfundet. Glyphosat kunne alene reducerede artsantallet med maks. 23 %, og kvælstofgødskning reducerede artsantallet med maks. 29 % (Figur 3.2). De to faktorer indvirker således klart og nogenlunde lige meget rent kvantitativt på diversiteten. N alene virkede stærkt på plantebiomassen (fra ca. 500 til 830 g/m²), mens glyphosat ved højt N kunne reducere biomassen tilsvarende. Af Figur 3.2 kan det endvidere ses, at ved 0, 1 og 25 % herbicid var der ingen klar effekt af kvælstofgødskning på de tre strategier, mens der ved 5 % glyphosat var en tendens til at ved øget N-tilførsel steg andelene af C og R, mens S faldt.Ved 0 Herbicid: R var næsten konstant ved øget N-tilførsel, mens C udkonkurrerede S ved højere N.Ved 25 % herbicid: R næsten konstant ved øget N-tilførsel, S udkonkurrerede C ved højere N. S-strateger udkonkurrerede altså C- og til dels R-strateger ved høj herbicidbelastning ved såvel oligo- som eutrofe forhold. Her skal man dog være opmærksom på, at de enkelte strategiers biomasseproduktion primært udgjordes af fåresvingel, stortoppet hvene og rejnfan. En tilsvarede situation med få arter, der dominerer, er dog ikke ualmindeligt i agerlandet. En analyse af gennemsnitsværdier af total biomasse og diversitet for de 12 forskellige kombinationsbehandlinger viste, at uden N-tilførsel var der maksimal biomasse og diversitet ved ingen eller 1 % herbicid (Figur 3.3). Endvidere var der ved 25 kg N-tillførsel maximal biomasse ved 5 % herbicid, mens artsantallet var relativt ens for de fire herbicid-niveauer. Ved 100 kg N-tilførsel var der højst artsantal ved 25 % herbicid, men biomasse maximum ved 0 herbicid.
Figur 3.3: Gennemsnitsværdier af total biomasse for de 12 forskellige kombinationsbehandlinger (markdosis 1440 g a.i./ha) med indikation af artsrigdom. Symbol størrelsen er proportional med artsantallet, cirkler = 0 N, trekanter = 25 Kg N ha-1,firkant 100 kg N ha-1 Analysen af plantebiomasse, artsrigdom og mosfrekvens mod de forklarende variable viste, at biomassen korrelerede kraftigt og positivt med førne, C- og R-strategier samt ler, men korrelerer negativt med S- strategien (Tabel 3.3). Artsrigdommen og mosfrekvensen korrelerede ikke stærkt med nogle af de viste variable, men mosser havde dog en vis positiv korrelation med fint sand og negativ korrelation med førne. Tabel 3.3. Korrelationskoefficienter mellem biomasse, artsdiversitet, mosfrekvens og forklarende variable. Grime indikator værdier er baseret på vægtet gennemsnit af prøvefelter udregnet på biomassen.
3.2.4 Multivariat behandling af vegetationsdata3.2.4.1 DCA-analyse Tabel 3.4. Korrellationskoefficienter mellem de to første ordinationsakser og forklarende variable. Alle de viste variable er signifikant korrelleret til en af de to akser med p-værdier < 0.01. De forklarende variable er sorterede ift værdien i DCA 1 søjlen. Fed indikerer de variable med størst korrellation med DCA 1 og 2.
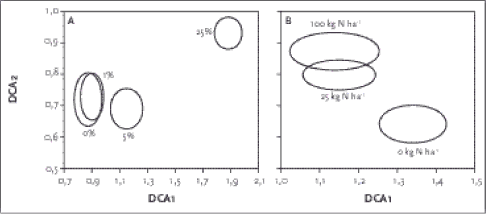
Dette afspejledes tydeligt i DCA plottene, når de fire glyphosatbehandlinger afbildedes (Figur 3.4a). Her fremgår det, at DCA 1 primært er en glyphosatrelateret gradient, men også at glyphosat i et vist omfang påvirker anden-aksen, hvor 25 % herbicid placerer sig relativt isoleret fra de øvrige herbicidbehandlinger. Andenaksen korrelerede svagt positivt med nitrogentilførsel og nitrat i jorden og svagt negativt med pH i 2003 og andelen af fint sand. Dette afspejledes mere tydeligt i DCA-plottene (Figur 3.4b) hvor de tre N-tilførselsniveauer blev afbildet. Der var en tydelig N-gradient i materialet på begge akser, hvor især 0 kg N isolerede sig fra 25 og 100 kg N (interaktion mellem herbiciddosering og kvælstoftilførsel).
Figur 3.4. DCA analyser af vegetationsdata for eksperimentet. Hver ring har middelværdien af de 10 replika som centrum og 1 standardfejl som radius. A viser herbicidbehandlingen (markdosis 1440 g a.i./ha), B de forskellige N-tilførselsniveauer. 3.2.4.2 CCA-analyse Tabel 3.5. Forklarende variable og ranket varians forklaret af de enkelte variable beregnet med fortløbende selektion af variable enkeltvis efterfulgt at Monte Carlo Permutations test. Variable der forklarer < 1 % er opført alfabetisk. N.S.= Ikke signifikant, * betyder p<0,05, *** betyder p<0,001. 3.2.4.3 Indikatorartsanalyse phosatfølsom, mens fåresvingel var meget robust over for glyphosat. Tabel 3.6. Indikatorartsanalyse i de forskellige herbicid-doseringsgrupper (markdosis 1440 g a.i./ha) under oligotrofe (0 kg N) forhold. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans (p < 0,05).
Tabel 3.7. Indikatorartsanalyse i de forskellige herbicid-doseringsgrupper (markdosis 1440 g a.i./ha) under eutrofe (100 kg N) forhold. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans (p < 0,05).
Andre arter reagerede forskelligt under de to N-niveauer – omend billedet ikke var så statistisk klart. Alm. kongepen (Hypochoeris radiata), hvidmelet gåsefod (Chenopodium album) og mosset Ceratodon purpureus klarede sig signifikant bedre i konkurrencen med højt glyphosatniveau og høj N-tilførsel, mens dette ikke var tilfældet ved lavt N-niveau, hvor der var en ikke-signifikant tendens til det modsatte. Uden herbicidbehandling kunne vi finde indikatorarter med klar respons på forskellig N- tilførsel (Tabel 3.8). Alm. kongepen (Hypochoeris radiata), alm. kællingetand (Lotus coniculatus) og mosset Ceratodon purpureus klarede sig signifikant bedre i konkurrencen uden N. Kvik (Elytrigia repens) og rejnfan (Tanacetum vulgare) reagerede derimod positivt på N- tilførsel. Prikbladet perikon (Hypericon perforatum) reagerede intermediært ved at have optimum ved 25 kg N. Tabel 3.8. Indikatorartsanalyse i de forskellige N-tilførselsniveauer uden herbicidbehandling. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans.
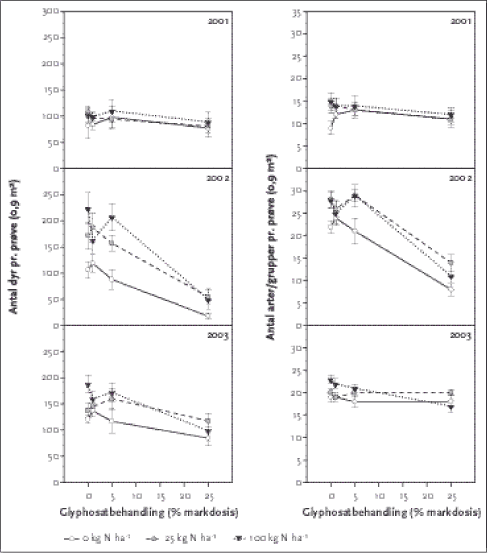
3.2.5 LeddyrBlandt de grupper af leddyr, der er sorteret (dvs. herbivore gruppe, løbebiller og rovbiller), var tæger de talrigeste med i gennemsnit 67 dyr per prøve i 2003, hvoraf de fleste var blomstertæger (48 per prøve) efterfulgt af frøtæger (12 per prøve). Cikaderne var også talrige (gennemsnitligt 56 per prøve), rovbiller forekom med et gennemsnit på 7 per prøve, Clavicornia-gruppen (skimmelbiller, mariehøns og glimmerbøsser) havde et gennemsnit på 4 per prøve, hvoraf 3 var skimmelbiller, og de øvrige grupper (bier/hvepse, svirrefluer, sommerfugle, bladlopper, løbebiller, smeldere, bladbiller, jordlopper og snudebiller) forekom alle i et gennemsnitligt antal under 1 per prøve. 3.2.5.1 Udvikling over år Set over alle tre år var der signifikante pesticideffekter på bier og hvepse, svirrefluer, cikader, de fleste tægegrupper, jordlopper, små rovbiller, rovbillelarver, alle Clavicorniagrupper (skimmelbiller, glimmerbøsser og mariehøns), det samlede antal leddyr og antallet af arter (Tabel 3.9). Der var i alle tilfælde tale om et fald i antallet ved højeste pesticiddosering sammenlignet med de usprøjtede felter (se Figur 3.5). For nogle grupper var der tale om et begyndende fald i antalllerede ved 1 % af markdosis, mens der for andre først var en negativ effekt ved 25 % af markdosis. Effekten af kvælstof var signifikant for næsten de samme grupper som pesticideffekten, med undtagelse af bier/hvepse og svirrefluer, som ikke var påvirket af kvælstofniveauet, og sommerfugle og løbebillelarver, som var signifikant påvirket af kvælstof, men ikke af herbicidet (Tabel 3.9). For de fleste grupper steg det gennemsnitlige antal med stigende kvælstoftilførsel, men for Scutelleridae-/Pentatomidae-tægenymfer var effekten omvendt, og løbebillelarvernes antal steg ved 25 kg N/ha sammenlignet med 0, men faldt så ved 100 kg N/ha. Tabel 3.9. P-værdier for effekt af pesticid- og N-behandling på leddyrfaunaen. Testen er udført på data fra årene 2001-2003. I de tilfælde, hvor interaktionleddet har en signifikant effekt, kan effekten af de enkelte variable ikke vurderes ud fra tabellen.
* Testet med interaktionsleddet Af Tabel 3.10 fremgår, at blokeffekten maksimalt udgjorde 10 % af den samlede tilfældige varians, mens forskelle i forekomsten af leddyr mellem årene gennemgående udgjorde en større del (op til 56 %). Blokeffekten varierede mellem årene (interaktion), idet der i 2001 og 2003 var en kraftigere blokeffekt end i 2002. Variationen mellem felter inden for blokken (felt(blok) i Tabel 3.10) var af cirka samme størrelse som variationen mellem blokke. Tabel 3.10. Procentvis bidrag til den samlede ”tilfældige” variation for det samlede leddyrdatasæt (2001-2003). Data er testet uden interaktionsleddet glyphosat*NH4 som systematisk variabel.
* Data for alle år 3.2.5.2 Leddyr år 2003 Tabel 3.11. P-værdier (Proc MIXED) for effekt af glyphosatbehandling og kvælstoftilførsel på leddyr indsamlet i 2003. Effekten af hver af de to variable kan kun vurderes i de tilfælde, hvor interaktionen ikke var signifikant. Tal i parentes indikerer problemer med testen (for få dyr, manglende konvergens el. lign.).
* Testet med interaktionsleddet glyphosat*kvælstof som systematisk variabel. Af Figur 3.5 ses, at antal arter og antal dyr per prøve er reduceret ved den højeste glyphosatdosering. Felterne, der modtog 100 kg N/ha, havde flere dyr og arter i den usprøjtede situation end de andre kvælstofniveauer, men faldet i antal dyr og arter som funktion af stigende pesticiddosering var også størst i 100 kg N-felterne (Figur 3.5). Den maksimale herbicideffekt var 47 % reduktion i antallet af dyr og 26 % reduktion af artsantallet. Kvælstoftilførsel resulterede i en øgning af antal leddyr på maks. 55 % og maks. 21 % flere arter (cf. Figur 3.5). Den pesticideffekt, der ses for antal dyr ialt, går igen for de fleste øvrige grupper med signifikant pesticideffekt. For bier/hvepse var der dog ingen klar tendens, mens store løbebiller (for få dyr), og skimmelbiller forekom i mindre antal ved 1 og 5 % markdosis end i de usprøjtede plots, men i højest antal i plots behandlet med 25 % af markdosis (middelværdier hhv. 3,1; 2,3; 1,8 og 4,1). Antallet af de forskellige grupper steg generelt med stigende kvælstoftilførsel, med undtagelse af Scutelleridae-/Pentatomidaetægenymfer, som forekom i faldende antal med stigende kvælstoftilførsel. Den tilfældige variation mellem prøverne udgjordes for op til 30 %s vedkommende af variation mellem blokke (Tabel 3.12), mens variationen mellem felter inden for blokke udgjorde op til 64 %. Begge variationsparametre varierede meget mellem de forskellige grupper af leddyr.
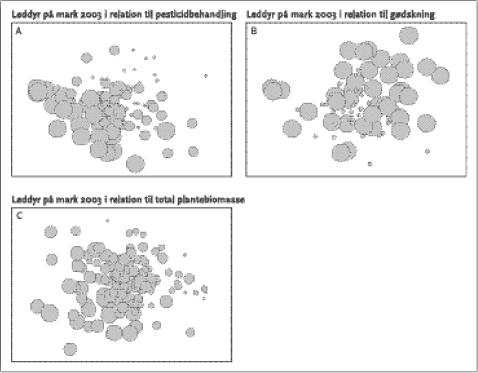
Figur 3.5. Antal leddyr (minus bladlus og trips) og antal leddyrarter per prøve i årene 2001-2003 som funktion af glyphosatdosis (markdosis 1440 g a.i./ha) ved de tre forskellige gødskningsniveauer. Punkterne repræsenterer rmiddelværdier af 10 prøver, de vertikale linjer er standardfejlen på middeltallet. MDS-plottene (Figur 3.6) viser, at der er en sammenfaldende gradient i artssammensætning på den ene side og glyphosatbehandling eller plantebiomasse på den anden, mens det ikke er tilfældet for kvælstoftilførslen. Med andre ord er forskelle i artssammensætning bedre korreleret med pesticidbehandlingen og plantebiomasse end med kvælstoftilførslen. Den mest forklarende enkeltvariabel blandt de undersøgte jordbunds- og plantevariable (miljøparametre) var total plantebiomasse (korrelation 0,247 i BIOENV), efterfulgt af pesticidbehandlingen (korrelation 0,202). Ingen andre miljøvariable havde en korrelation på over 0,2 med artssammensætningen af leddyr. Når det maksimale antal forklarende variable blev sat til tre, var artssammensætningen af leddyr bedst korreleret med pesticidbehandlingen, den totale plantebiomasse og førnebiomassen (korrelation 0,351). Da der var en vis sammenhæng mellem de to biomassemål, blev førnebiomassen efterfølgende udelukket fra analysen, hvorefter pesticidbehandlingen, den totale plantebiomasse og det i 2003 målte NO3-N forklarede artsfordelingen bedst (korrelation 0,336). Når maksimalt fem forklarende variable indgik i analysen, var korrelationen bedst med en kombination af pesticidbehandling, kvælstoftilførsel, antal plantearter, førnebiomasse og bloknummer (korrelation 0,396). En opdeling af plantebiomassen i græsser, øvrige urter (ikke-græsser), mosser og træer og efterfølgende analyse viste, at artssammensætningen af leddyr var bedst forklaret af græsbiomassem (korrelation 0,14 i BIOENV), efterfulgt af den samlede plantebiomasse (0,127) og biomassen af øvrige urter (0,077).
Figur 3.6. Multidimensionel scalings-plots med hhv. pesticidbehandling (A), gødskning (B) og total plantebiomasse (C) lagt over, dvs. afstanden mellem boblerne er proportional med forskellen i artssammensætning, og størrelsen af boblerne svarer til behandlingsintensiteten/plantebiomassen. To-vejs ANOSIM (Tabel 3.13) gav en overordnet R på 0,188 for effekten af glyphosatbehandlingen (maks. 1,0) på artssammensætningen og et signifikansniveau på 0,001 (testet på tværs af kvælstoftilførselsniveauerne). Artssammensætningen i de usprøjtede felter samt felter behandlet med 1 og 5 % af markdosis var signifikant forskellig fra felterne behandlet med 25 % af markdosis. For kvælstoftilførslen var der en R på 0,177 og et overordnet signifikansniveau på 0,001 (testet på tværs af pesticidniveauerne). Der var signifikante forskelle i artssammensætningen mellem alle kvælstoftilførselsniveauer. Ingen enkeltarter bidrog iøjnefaldende til forskellene i artsammensætning mellem felter behandlet med forskellige glyphosat- og kvælstofdoser (SIMPER). Det maksimale bidrag var 11 % af forskellen mellem felter sprøjtet med 25 og 5 % af markdosis forklaret af Stenodema-nymfer (blomstertæge, lever på græsser), baseret på en gennemsnitlig frekvens på hhv. 4,21 og 6,12 i de to pesticidniveauer. Denne type tæge var i alle, undtagen et, tilfælde den mest forklarende for artsammensætningsforskelle mellem såvel pesticid- som kvælstoftilførselsniveauer, efterfulgt af andre tæger, cikader og små rovbiller i varierende rækkefølge. Tabel 3.12. Procentvis bidrag til den samlede ”tilfældige” variation for leddyr indsamlet i 2003.
Tabel 3.13. ANOSIM-test af effekten af glyphosat (markdosis 1440 g a.i./ha) og kvælstof på artssammensætningen af leddyr.
3.3 Diskussion af felteksperimentets resultater3.3.1 Jordbund og forsøgsetableringEksperimentet har generelt været meget vellykket i forhold til etablering og udvikling af vegetationen. Enkelte arter havde svært ved at etablere sig fra frø (fx kvik, stor nælde, gråbynke og agertidsel som er udbredte og konkurrencestærke hegnsplanter). Såbedet var stærkt soleksponeret, og få dages udtørring af frø kan have haft afgørende betydning for nogle arters etablering. Flere af C-strategerne formerer sig vegetativt, så de er vanskelige at etablere fra frø. De udsåede arter dominerer vegetationen og græsser opnåede den ønskede dominans – mens indvandringen af arter, specielt mosser, gav yderligere dimensioner til eksperimentet mht. koloniseringsmuligheder for planter. Selvom arealet var udvalgt som værende homogen mht. jordbund, viste der sig dog visse forskelle i jordbunden. Den tilførte kvælstof har derudover haft tilsigtede signifikante effekter efter 3 år på kvælstofniveauet og dertil knyttede sideeffekter på pH og ledningsevne. 3.3.2 Effekter af herbicidbehandling og N-tilførsel på vegetation og leddyrHovedformålet med eksperimentet var at belyse effekten af herbicidafdrift og kvælstoftilførsel. Eksperimentet har påvist klare interaktioner mellem N- og glyphosatpåvirkningen for plantebiomasse, førne, karplantediversitet, mosfrekvenser og dominansen af konkurrencestærke plantearter. For plantebiomasse og førne var der en meget markant reduktion ved 25 og 100 kg N i kombination med 25 % glyphosat, men denne effekt udeblev ved 0 kg N. For mosserne var frekvensen ved høje kvælstofniveauer klart højere ved 25 % glyphosat end ved lavere doser, hvorimod det omvendte var tilfældet ved lavere kvælstofdoser (Figur 3.2). Glyphosat ændrer totalt konkurrenceforholdene og dermed artssammensætningen af planter. Det skyldes sandsynligvis, at fåresvingel er både nøjsom og tilsyneladende relativt glyphosat-tolerant, således at den på trods af 25 % markdosis opnår en rimelig biomasse og dominerer felterne (Tabel 3.6), hvorimod de herbicidfølsomme arter forsvinder. Glyphosat forhindrer en række arter i at etablere sig og mindsker artsdiversiteten ved 0 kg N-tilførsel. Ved 25 kg N er der stort set ingen effekt af glyphosatbehandlingen, mens der ved 100 kg N er en positiv effekt af stigende glyphosatdosering, således at artsantallet ved 25 % af markdosis er sammenligneligt med antallet ved 0 og 25 kg N (Figur 3.2). Dette skyldes formentlig, at glyphosaten virker så negativt på nogle arter (konkurrencestrategerne), at deres tilbagegang giver nicher til relativt mere glyphosattolerante arter (bl.a. R- og S-strateger). Vi skal dog bemærke, at det især er meget få arter fra hver strategi, der dominerer vegetationen, så billedet kunne være anderledes med andre arter, selvom få arter, der dominerer i hver strategi, ikke vil være usædvanligt i hegnsvegetation. Glyphosat kan alene reducere artsantallet med ca ¼, og en tilsvarende reduktion i artsantal ses af kvælstofgødskning (Figur 3.2). Begge faktorer indvirker således klart og nogenlunde lige meget rent kvantitativt på diversiteten. Glyphosatens isolerede betydning på plantebiomassen er meget klar ved højt N-nivau, men næsten fraværende ved 0 kg N. En mulig forklaring på, at C-strategerne tilsyneladende påvirkes mest af glyphosatbehandlingen kunne være, at C-strategerne oftest er højere og derfor måske kan 'skygge' for glyphosatafdrift på R- og S-strategerne. Vi har ingen målinger af afsat mængde glyphosat på de forskellige planter, men det er ikke vort indtryk, at der har været væsentlig forskel i afsætningen af glyphosat på de enkelte arter. Udbringningen er sket om foråret (maj), hvor de fleste planter endnu er små. Den potentielle forskel i eksponering bliver først mere udpræget senere i vækstsæsonen. Måske er konkurrenceplanterne generelt mere følsomme end de øvrige, fx pga. deres højere vækstrate. En sådan forklaring stemmer overens med, at ved stigende glyphosatdosering forsvinder den positive effekt af N-tilførsel på den totale plantebiomasse (Figur 3.2). En anden mulig forklaring på den interaktive effekt af N-tilførsel og glyphosatdosering er, at selv beskedne glyphosatdoser hindrer en konkurrencerelateret uddøen (competitive exclusion), som ellers typisk er resultatet af tre år med øget N-tilførsel, fordi de dominerende nitrofile arter bortskygger små og lave arter. Glyphosatbehandlingen vil på den måde kunne åbne vegetationen for mindre konkurrencestærke arter selv ved 100 kg N/ha/år. Med de multivariate analyseteknikker viste det sig, at glyphosat er den væsentligste variabel til ændringer i vegetationssammensætningen (Tabel 3.6). Jordbundskarakteristika og N-tilførsel viste sig således at være af sekundær betydning i forhold til de anvendte glyphosatkoncentrationer. Det skyldes, at glyphosat ved 0 kg kvælstof øger uddøen af herbicidfølsomme arter og ved 100 kg N virker glyphosat især ved at reducere biomasse og førne. Dette åbner mulighed for øget indvandring/etablering af nye (nitrofile) arter, som ikke findes ved 0 kg N. Glyphosat kan således virke både reducerende og fremmende afhængigt af N-niveauet. Indikatorartsanalysen viste, at nogle arter reagerer meget forskelligt på herbicidbehandling afhængig af N-tilførslen. Dette kan have været medvirkende årsag til, at de forskellige funktionelle plantegrupper har reageret så markant, idet deres biomasse ofte er domineret af en eller meget få arter. Hvis vi eksempelvis havde udsået en anden, mere glyphosatfølsom nøjsomhedgræs end fåresvingel, havde billedet måske set anderledes ud. Hvis stor nælde, agertidsel, gråbynke eller kvik var slået an, kunne billedet også have været anderledes. Derfor skal dette resultat tolkes med varsomhed. Det ændrer dog ikke ved, at arter har forskellige reaktionsmønstre på glyphosat og næringsberigelse, og at en vegetation, der udsættes for herbicidafdrift, meget vel kan være domineret af en eller få arter. For leddyrene var der signifikante effekter af glyphosatbehandlingen på en lang række grupper, på antal dyr og på diversiteten, uafhængigt af N-tilførslen (Tabel 3.9, Figur 3.5). Specielt 25 % glyphosat slog kraftigt igennem på leddyrene. Eftersom glyphosat formentlig ikke er giftigt for leddyrene, må der være tale om en indirekte effekt via effekten på planterne, førnen og (indirekte) mikroklimaet. Dette underbygges af, at artsammensætningen af leddyrene i høj grad er korreleret med plantebiomasse og førnebiomasse. Den overvejende negative effekt af herbicidbehandlingen på leddyrene stemmer overens med fx resultaterne i Reddersen m.fl. (1998). Der findes kun ganske få studier, der tidligere har beskæftiget sig med effekter af lav- herbiciddosis på vegetation sammensat af arter, der findes i (halv-) naturen og småbiotoper i agerlandet (Bicheludvalget 1999). Langt den overvejende del af pesticidforskningen har været rettet mod arter på markfladen, såvel ukrudtsarter som skadedyr og svampe. Der er lavet talrige dosis-respons studier af de forskellige midlers effekter på veldefinerede målorganismer som en del af godkendelsesprocedurer for pesticider. Derimod er denne markante effekt på dyre- og plantesamfund ikke påvist før. Endvidere har der været lavet en række studier af indirekte effekter af herbicider på leddyr og fugle i agerlandet (se review i Bicheludvalget 1999), men kun et enkelt studie (Kleijn & Snoeijing 1997) har tidligere forsøgt at kvantificere effekten af et herbicid (Starane 200; fluoroxypyr) på agerlandets kantvegetation. Kleijn & Snoeijing (1997) fandt et fald i artsrigdom med øget fluoroxypyrdosering, mens biomasse produktionen steg for græssers vedkommende og faldt for andre urter under højproduktive forhold. De fandt endvidere, at 5 og 10 % fluoroxypyr (af markdosis) reducerede biomasseproduktionen af indvandrende urter og øgede ekstinktionsraten af de tilstedeværende arter. Forsøget anvendte årlig slåning og fjernelse af biomassen, hvilket også påvirkede konkurrencen signifikant, så derfor kan resultaterne ikke direkte sammenlignes med nærværende forsøg. Eksperimentet i dette projekt har ved at tildele varierende doser af N og ensartede doser af P og K kunnet påvise en række forventede effekter af kvælstof på levende biomasse og på førne. Nitrogentilførsel alene (dvs. uden glyphosatbehandling) øgede produktiviteten af en sådan vegetation med ca. 65 %, når vegetationen som i dette tilfælde var N-begrænset. Konkurrencestrategerne (især repræsenteret ved stortoppet hvene) viste en stigende dominans med øget N-tilførsel, mens Nøjsomhedsplanterne (S-strateger især repræsenteret ved fåresvingel) viste en vigende tilstedeværelse. Ruderaterne viste ingen signifikante ændringer. Artsrigdommen for karplanter og frekvensen for mosser er højst ved 0 kg N/ha og stærkt faldende ved stigende N-tilførsel. Dette bekræfter således teorierne om tålegrænser for luftbårent kvælstof til N-begrænsede økosystemer (Bobbink m.fl. 1998). Der er næppe tvivl om at effekterne af øget N-tilførsel på de undersøgte leddyrgrupper, som jo primært består af herbivore arter, er afledt af den øgede plante- og førnebiomasse og faldende plantediversitet ved øget N (Figur 3.2 og 3.5). De fleste arter har gavn af den øgede mængde tilgængelig føde, bedre mikroklima og mere stratificerede vegetationsstruktur, der opstår ved øget vækst, mens nogle få arter kan tænkes at være negativt påvirkede i kraft af, at plantediversiteten falder. I nærværende studie sås kun et konsekvent fald for en type tæger (Scutelleridae/Pentatomidae), som lever på græs, hvilket ikke umiddelbart kan forklares af ovenstående argumentation. Den overvejende positive effekt i dette eksperiment kunne meget vel hænge sammen med, at vegetationen endnu ikke var fuldt udviklet, og biomassetilvæksten derfor har været den mest betydende faktor for leddyrene. En sådan effekt stemmer overens med resultaterne i Esbjerg & Petersen (2002), som fandt en positiv korrelation mellem plantebiomasse og leddyrbiomasse for visse herbivore grupper. Man kan sagtens forestille sig, at den forarmning af floraen, der sker ved længere tids N-påvirkning, på lang sigt vil resultere i en tilsvarende forarmning af leddyrfaunaen. Andre studier (Koricheva m.fl. 2000, Siemann 1998 og Haddad m.fl. 2000) tyder imidlertid på, at kun meget stationære leddyr vil påvirkes af en lokal forarmning af floraen som følge af fx N-tilførsel. I et studie af kvik-domineret flora sammenlignet med en artsrig ”naturlig” flora fandt Lagerlöf & Wallin (1993), at der godt nok var flere leddyrarter i plots med ”naturlig” flora, men kvik viste sig også at have en stor betydning som skjul for mange leddyrgrupper, primært rovlevende insekter. Størrelsen af afdriften af ammoniak fra markflader og fejlplaceret gødskning er meget dårligt dokumenteret, hvorimod effekten af gødskning er undersøgt meget grundigt i en lang række undersøgelser (se reviews af Waide m.fl. 1999, Bobbink m.fl. 1998). De fleste er enige, om at produktivitet påvirker diversitet, men de underliggende mekanismer og mønstre er man ikke enige om (Waide m.fl. 1999). En stor del af undersøgelserne opererer med NPK-gødskning, mens forskning i N-tilførsel alene hovedsageligt er lavet på N-følsom vegetation i kvælstofbegrænsede økosystemer (højmoser, heder, skove etc.) i relation til at fastsætte tålegrænser for N-tilførsel fra luften, (Bak m.fl. 1999). Kleijn & Snoeijing (1997) har i lighed med dette projekt undersøgt kombinationer af herbicid (fluoroxypyr, 0-50 % markdosis) med produktivitet (0, 25 og 50 % af markdosis NPK-tilførsel), hvor resultaterne som forventet gav meget signifikante udslag med øget biomasseproduktion og faldende artsdiversitet (specielt lave arter blev udkonkurreret) med øget produktivitet. Effekten her kan dog ikke umiddelbart sammenlignes med nærværende forsøg, da der er tale om et NPK-forsøg. 3.3.2.1 Funktionel fortolkning af resultaterne Resultaterne af glyphosatbehandlingen kan derudover tolkes som en forstyrrelse af økosystemet, der også har forskellig virkning på de forskellige plantestrategier. Forstyrrelser (fx. græsning, brand, slåning, herbicider) har oftest været undersøgt som enten-eller begivenheder, og en del undersøgelser påpeger, at mange halv-naturlige økosystemer har behov for forstyrrelser for at kunne opretholde artsdiversiteten (Mackey & Currie 2000). En eng har behov for græsning og/eller høslet for at opretholde sin struktur og funktion og de tilknyttede og tilpassede arter. Mangel på forstyrrelser går især ud over rosetplanter (typisk nøjsomhedsplanter) og ruderater samt mosser i græsland (Jacquemyn m.fl. 2003). Nogle typer af forstyrrelser forbedrer med andre ord vilkårene for visse plantearter og andre forringer mulighederne (Jutila & Grace 2002). Hvis man opfatter glyphosatbehandlingen som en forstyrrelse, kan resultatet af 25 % markdosis forklares med forbedrede konkurrencevilkår for en relativ glyphosattolerant nøjsomhedsplante (fåresvingel,) idet konkurrenceplanterne ikke kan klare forstyrrelsen. Resultaterne af den eksperimentelle undersøgelse viser, at indvandring (med mosser som eksempel) er størst ved lave N-niveauer pga. lav biomasse (og dækning) af vegetationen. Derimod har mange planter som udgangspunkt svært ved at etablere sig ved høj N-tilførsel pga. konkurrencen fra C-strategerne, som imidlertid viser sig mere følsomme over for glyphosat i eksperimentet. Derved får mosserne muligheden for at etablere sig ved moderate herbiciddoser. Ved 5 % glyphosat-dosis var effekten af kvælstofgødskning markant, således at andelen af C- og R-strateger stiger ved øget N-tilførsel , mens S-strategernes andel falder (Figur 3.2). Med andre ord: S-strateger udkonkurrerede altså C- og til dels R-strateger ved høj herbicidbelastning ved såvel lav som høj N-tilførsel. Vi har i denne rapport valgt at fokusere på år 3 (det sidste projektår), hvilket under alle omstændigheder kun er et billede i en succession, som kræver yderligere opfølgning, fx efter 5 og 10 år, specielt hvis resultaterne af eksperimentet skal sammenholdes med hegnsundersøgelsen (se kap. 4). Endvidere kunne det være særdeles interessant at indlægge en yderligere variabel, fx slåning i eksperimentet idet én af mulighederne for at forvalte eutrofieret vegetation er slåning og fjernelse af biomasse. Dette kan gælde for vegetationen i enge såvel som for vejkants- og hegnsfodposevegetation. 3.3.3 HypotesenePå baggrund af resultaterne af eksperimentet kan hypotesen om, at herbicidafdriften vil påvirke diversitet og dominans af en græslandsvegetation og de tilknyttede leddyr, bekræftes med den modifikation, at især 25 % af markdosis glyphosat gav de forventede effekter. I de fleste tilfælde var der ingen signifikant effekt af 1 % dosis, hvorimod 5 % fx gav signifikante ændringer i artsdominansen. Hypotese 2 vedrørende reduktion i artsantallet som følge af ammoniakafdrift simuleret ved N-tilførsel blev delvist bekræftet af experimentet. N-tilførslen reducerede artsantallet af en græslandsvegetation ved at øge dominansen af flerårige, konkurrencestærke plantearter på bekostning af mangfoldighed af planter og leddyr. Som forventet var denne reaktion stærkest hos planterne, da leddyrene formentlig reagerede på planternes vækst og tilstedeværelse. Eksperimentets hypotese H3, der forudsiger interaktioner mellem herbicid- og kvælstofpåvirkning, blev fuldt ud bekræftet af eksperimentet.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||