Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 102, 2006
Effekter af pulseksponering med pyrethroider på vandløbsinvertebrater
- med særlig fokus på lambda-cyhalothrin
Indholdsfortegnelse
- 2.1 Metodebeskrivelse
- 2.2 Forsøgsorganismer
- 2.3 Bevægelsesadfærd i laboratoriet
- 2.3.1 Overordnet forsøgsstrategi
- 2.3.2 Forsøgsbetingelser
- 2.3.3 Registrering af bevægelsesadfærd i laboratoriet
- 2.3.4 Generelt forsøgsdesign
- 2.3.5 Relativ toksicitet af pyrethroider anvendt i Danmark
- 2.3.6 Pulsvarighedens betydning
- 2.3.7 Temperaturens betydning
- 2.3.8 Pesticidblanding
- 2.3.9 Koncentrations-respons-forsøg med udvalgte arter
- 2.3.10 Restitueringsforsøg
- 2.3.11 Rovdyr-byttedyrs-interaktion
- 2.3.12 Statistisk metode
- 2.4 Drift i strømrender
- 2.5 Oversigt over udførte forsøg
- 2.6 Pesticidvalg
- 2.7 Kvantificering af lambda-cyhalothrin
- 3.1 Kvantificering af lambda-cyhalothrin
- 3.2 Bevægelsesadfærd i laboratoriet
- 3.3 Drift i strømrender
- 4.1 Generalisérbarhed af projektresultater
- 4.2 Ekstrapolering fra laboratorie til strømrender
- 4.3 Forsinkede effekter efter endt pulseksponering
- 4.4 Interspecifikke interaktioner og flerarts-forsøg
- 4.5 Sammenstilling af laboratorie- og strømrendeforsøg
Bilag A: Statistik for strømrendeforsøg
Forord
Nærværende rapport indeholder resultater fra et pesticidforskningsprojekt med titlen “Effekter af pulseksponering med pyrethroider på bevægelsesadfærd og drift hos vandløbsinvertebrater”. Projektet, der blev igangsat af Miljøstyrelsen inden for indsatsområdet “Ferskvand og Pesticider”, er blevet udført i perioden august 2003 til december 2005, og er løbende blevet evalueret af en følgegruppe bestående af: Jørn Kirkegaard (formand) og Alf Aagaard, Miljøstyrelsen; Gitte Larsen og Lars Kjellerup Larsen, Skov- og Naturstyrelsen; Peter Wiberg-Larsen, Fyns Amt; Niels-Martin Frost, DuPont Danmark ApS; Poul Henning Petersen og Hans Roust Thysen, Dansk Landbrugsrådgivning; Jørgen Jakobsen og Niels Henrik Spliid, DJF, Forskningscenter Flakkebjerg; Merete Styczen og Dorthe Lærke Baun, DHI – Institut for Vand og Miljø; Jens Carl Streibig, Nina Cedergreen Forchhammer, Carsten Petersen, Christian Bender Koch, Birgitte Gjettermann og Lars Bo Pedersen, KVL; Dean Jacobsen, Ferskvandsbiologisk Laboratorium, KU; Anders Baun og Tina Slothuus, Miljø & Ressourcer, DTU; René Kastbjerg Juhler, GEUS.
Forfatterne ønsker at takke Jørgen Jakobsen, Alf Aagaard, Peter Wiberg-Larsen, Anders Baun, Hans Roust Thysen og Niels-Martin Frost, der i forbindelse med udarbejdelsen af nærværende rapport skriftligt kommenterede rapportudkastet.
Ulrik Nørum og Poul Bjerregaard
Biologisk Institut
Syddansk Universitet
Nikolai Friberg og Søren Erik Larsen
Afd. for Ferskvandsøkologi
Danmarks Miljøundersøgelser
Marts 2006
Sammenfatning
Formål
Et af de centrale fænomener, der observeres i forbindelse med kortvarige pesticidpulse i vandløb, er drift, hvor en lang række vandløbsinvertebrater føres med strømmen og helt eller delvist forsvinder fra den belastede vandløbsstrækning. Det primære formål med projektet var at koble laboratoriestudier af pyrethroid-insekticiders subletale effekter på bevægelsesadfærd med driftstudier under mere miljørealistiske forhold i strømrender. Hypotesen var, at såfremt denne kobling kunne dokumenteres, ville de mindre omkostningstunge laboratorieforsøg kunne bruges til direkte at ekstrapolere til effekter i naturlige vandløb.
Metoder
Bevægelsesadfærden hos vandløbsinvertebrater anbragt i glaspetriskåle i laboratoriet blev registreret ved hjælp af et computerstyret videosporingsystem før, under og efter forskellige former for pulseksponering for pyrethroider, primært lambda-cyhalothrin. Effekten på bevægelsesadfærden blev sammenholdt med lambda-cyhalothrins indvirkning på drift og effekter i flerarts-forsøg i udendørs strømrender.
Forsøgsoversigt
I laboratoriet blev eksponeringsforsøg med pyrethroiderne esfenvalerat, alpha-cypermethrin, cypermethrin, lambda-cyhalothrin og tau-fluvalinat gennemført, og lambda-cyhalothrin blev valgt som modelpyrethroid. Betydningen af pulseksponeringens varighed og vandtemperaturen for adfærdsresponset blev undersøgt. Endelig blev et forsøg gennemført med lambda-cyhalothrin i kombination med det i danske vandløb hyppigt forekommende glyphosat.
Pulseksponeringsforsøg med krebsdyrene Gammarus pulex og Asellus aquaticus, slørvingen Leuctra nigra, døgnfluen Heptagenia sulphurea, vårfluen Sericostoma personatum og sneglen Ancylus fluviatilis blev gennemført i laboratoriet, ligesom langtidsvirkninger af pulseksponering med lambda-cyhalothrin blev undersøgt i restitueringsforsøg af tre ugers varighed med G. pulex og A. aquaticus. Effekter på interspecifikke interaktioner blev undersøgt ved et studie af rovdyr-byttedyrs-interaktionen mellem G. pulex og L. nigra.
Der blev gennemført to typer af eksperimenter i strømrender: Drift og flerarts-forsøg. I drifteksperimenter blev det umiddelbare driftrespons hos G. pulex, L. nigra, H. sulphurea, A. fluviatilis og S. personatum undersøgt under og efter pulseksponering med lambda-cyhalothrin i semi-naturlige vandløbsrender.
I eksperimenterne med flere arter blev de længerevarende effekter af eksponering med lambda-cyhalothrin undersøgt med hensyn til direkte og indirekte effekter. Undersøgelsen inkluderede G. pulex, L. nigra, H. sulphurea, og A. fluviatilis. Forsøget forløb i 10 dage med daglig måling af drift. Derudover blev omsætning af organisk stof og algebiomassen målt for at få indblik i indirekte effekter af pyrethroideksponeringen.
Resultater
I nedenstående tabel er resultater fra laboratoriet og strømrenderne sammenlignet på baggrund af NOEC- (no observable effect concentration) og LOEC- (lowest observable effect concentration) værdier. De laveste observerede effektkoncentrationer (LOEC) for L. nigra (1 ng l-1), G. pulex (10 ng l-1), A. aquaticus (10 ng l-1) og H. sulphurea (100 ng l-1) er alle indenfor forventede miljørealistiske niveauer.
Tabel 1. Ekstrapolering fra laboratorie til strømrender. NOEC- og LOEC-lambda-cyhalothrin-koncentrationer (ng l-1) for hyperaktivitet (Hyper), hypoaktivitet (Hypo) og immobilisering (I) i laboratoriet og for drift (D) i strømrenderne efter 90 minutters eksponering.
| Laboratoriet | Strømrender | ||||||
| Art | NOEC | LOECHyper | LOECHypo | LOECI | NOECD | LOECD | |
| Leuctra Nigra |
0,1 | - | 1 | 10 | - | 1 | |
| Gammarus pulex | 1 | 10 | 1000 | 1000 | 1 | 10 | |
| Asellus aquaticus | 1 | 10 | 100 | 1000 | ikke undersøgt | ikke undersøgt | |
| Heptagenia sulphurea | 10 | 100 | 10000 | 10000 | 10 | 100 | |
| Sericostoma personatum | 10000a | - | - | - | 1000 a | - | |
| Ancylus fluviatilis | 10000 a | - | - | - | 1000 a | - | |
a: Højeste testede koncentrationer i henholdsvis laboratoriet og strømrenderne.
I såvel laboratoriet som i strømrenderne var rangordningen af de anvendte arters følsomhed over for lambda-cyhalothrin:
L. nigra > G. pulex > H. sulphurea > S. personatum > A. fluviatilis
Laboratorieresultaterne viser, at de fem undersøgte pyrethroider har både samme kvalitative virkning på bevægelsesadfærden (hyperaktivitet og immobilisering) og meget sammenlignelige effektkoncentrationer hos G. pulex. Generelt observeres hyperaktivitet ved 10 ng l-1. Ved 100 ng l-1 er G. pulex ikke i stand til at opretholde hyperaktiviteten, og ved 1000 ng l-1 registreres immobilisering.
Konklusioner
På baggrund af resultaterne i nærværende projektrapport konkluderes:
1) at såvel de observerede effektkoncentrationer som rangordningen af de anvendte ferskvandsinvertebraters følsomhed over for lambda-cyhalothrin kan ekstrapoleres direkte fra laboratoriestudier af bevægelsesadfærd til studier af drift i de mere miljørealistiske strømrender. Når sammenhængen mellem ændringerne i bevægelsesadfærd i laboratoriet og driftresponset i strømrenderne hos den enkelte art er dokumenteret, vurderes laboratoriestudierne at kunne erstatte strømrendestudierne. Dette gælder dermed for de i projektet anvendte arter.
2) at alpha-cypermethrin, cypermethrin, lambda-cyhalothrin og tau-fluvalinat alle påvirker adfærden hos G. pulex markant ved en koncentration på 10 ng l-1, mens esfenvalerat fører til adfærdsændringer allerede ved 1 ng l-1. Alle de undersøgte pyrethroider har dermed effekter ved koncentrationer, der knapt kan måles; kravet til detektiongrænsen i de analysekemiske metoder, man normalt anvender til at monitere pesticider i grundvand og overfladevand, er til sammenligning 10 ng l-1.
3) at kortvarige pulseksponeringer med lambda-cyhalothrin fører til forsinkede effekter på adfærd og overlevelse hos ferskvandsinvertebrater; effekter der kan registreres i 2-3 uger efter pulseksponeringens ophør.
4) at rovdyr-byttedyrs-interaktioner påvirkes ved miljørealistiske pulseksponeringer for lambda-cyhalothrin.
5) at pulseksponering med lambda-cyhalothrin udløser klare indirekte effekter på tilvæksten i algebiomassen og på omsætningen af organisk stof, hvilket indikerer, at miljørealistiske eksponeringer af vandløbsinvertebratsamfundet med lambda-cyhalothrin kan give effekter på økosystemniveau, der går ud over påvirkningen på specifikke arter og populationer.
Projektets resultater dokumenterer, at de anvendte metoder komplementerer hinanden, således at de omkostningseffektive laboratoriestudier kan anvendes til hurtige screeninger af mulige effekter af pesticider på ferskvandsinvertebraters bevægelsesadfærd med henblik på fastlæggelse af effektkoncentrationer og rangordning af arternes følsomhed. Disse data kan bl.a. anvendes i planlægningen af strømrendeforsøg. Samtidigt kan observationer fra mere komplekse strømrendestudier, som f.eks. effekter i flerarts-forsøg, opklares ved efterfølgende laboratoriestudier af bevægelsesadfærd, restituering og interspecifikke interaktioner.
Summary
Aim
Drift is one of the key phenomena observed in relation to short-term pesticide pulses in freshwater streams. During these events a diverse range of stream invertebrates are carried along with the current and disappear completely or partially from the contaminated stretch of the stream. The main aim of the project was to link laboratory studies of the sublethal effects of pyrethroide insecticides on locomotory behaviour with drift studies under more environmentally realistic conditions in stream microcosms. The hypothesis was that if this link could be established, the cost-effective laboratory studies could be utilized to extrapolate directly to effects in natural streams.
Methods
The locomotory behaviour of stream invertebrates placed in glass petri dishes in the laboratory was quantified by using a computerized video tracking system before, during and following various kinds of pulse exposures to pyrethroids, especially lambda-cyhalothrin. The effects on locomotory behaviour were compared with the effects of lambda-cyhalothrin on drift and the effects in a multi-species system in outdoor stream microcosms.
Experiments
In the laboratory, behavioural toxicity tests were performed using the pyrethroids esfenvalerate, alpha-cypermethrin, cypermethrin, lambda-cyhalothrin and tau-fluvalinate, and lambda-cyhalothrin was selected as model pyrethroid. The influence of pulse duration and water temperature on the behavioural response was investigated. Finally, an experiment with lambda-cyhalothrin in combination with glyphosate, which is commonly occurring in Danish streams, was performed.
Pulse exposures using the crustaceans Gammarus pulex and Asellus aquaticus, the stonefly Leuctra nigra, the mayfly Heptagenia sulphurea, the caddisfly Sericostoma personatum and the snail Ancylus fluviatilis were performed in the laboratory. The long-term effects of pulse exposure to lambda-cyhalothrin were investigated in recovery studies lasting three weeks using G. pulex and A. aquaticus. The effects on interspecific interactions were investigated in a study of the predator-prey-relationship between G. pulex and L. nigra.
Two types of experiments were carried out in the stream microcosms: Drift and effects on a multi-species system. In the drift experiments, the immediate drift response in G. pulex, L. nigra, H. sulphurea, A. fluviatilis and S. personatum during and following pulse exposure to lambda-cyhalothin was studied in semi-natural stream microcosms.
In the multi-species studies, the long-term effects of exposure to lambda-cyhalothrin were investigated to elucidate both direct and indirect effects. This study included G. pulex, L. nigra, H. sulphurea and A. fluviatilis. The study lasted 10 days, with daily measurements of drift. Furthermore, the breakdown of organic matter and change in algal biomass was determined in order to clarify indirect effects of pyrethroid exposure.
Results
In the table below, a comparison of the laboratory and stream microcosm results based on NOEC (no observable effect concentration) and LOEC (lowest observable effect concentration) values is given. The LOECs for L. nigra (1 ng l-1), G. pulex (10 ng l-1), A. aquaticus (10 ng l-1) and H. sulphurea (100 ng l-1) are all within expected environmentally realistic levels.
Table 1. Extrapolation from laboratory to stream microcosms. NOEC and LOEC lambda-cyhalothrin-concentrations (ng l-1) for hyperactivity (Hyper), hypoactivity (Hypo) and immobilisation (I) in the laboratory and for drift (D) in the stream microcosms after 90 minutes of exposure.
| Laboratory | Stream microcosm | ||||||
| Species | NOEC | LOECHyper | LOECHypo | LOECI | NOECD | LOECD | |
| Leuctra nigra |
0.1 | - | 1 | 10 | - | 1 | |
| Gammarus pulex | 1 | 10 | 1000 | 1000 | 1 | 10 | |
| Asellus aquaticus | 1 | 10 | 100 | 1000 | no data | no data | |
| Heptagenia sulphurea | 10 | 100 | 10000 | 10000 | 10 | 100 | |
| Sericostoma personatum | 10000a | - | - | - | 1000 a | - | |
| Ancylus fluviatilis | 10000 a | - | - | - | 1000 a | - | |
a: Highest concentrations tested in the laboratory and in the stream microcosms.
In the laboratory as well as in the stream microcosms, the rank ordering of the sensitivity of the species towards lambda-cyhalothrin was:
L. nigra > G. pulex > H. sulphurea > S. personatum > A. fluviatilis
The laboratory results indicate that the five pyrethroids investigated have the same qualitative effects on locomotory behaviour (hyperactivity and immobilisation) as well as very comparable effect concentrations in G. pulex. In general, hyperactivity is observed at 10 ng l-1. At 100 ng l-1 G. pulex is not capable of maintaining the hyperactivity, and at 1000 ng l-1 immobilisation is observed.
Conclusions
Based on the results of the present report it is concluded:
1) that both the observed effect concentrations as well as the rank ordering of the species sensitivities towards lambda-cyhalothrin can be extrapolated directly from laboratory studies of locomotory behaviour to studies of drift in the more environmentally realistic stream microcosms. Once the relation between the changes in locomotory behaviour in the laboratory and drift in the microcosms has been documented for the individual species, the laboratory studies may replace the stream microcosm studies. Consequently, this applies to the species studied in the present project.
2) that alpha-cypermethrin, cypermethrin, lambda-cyhalothrin and tau-fluvalinate all markedly affect the behaviour of G. pulex at a concentration of 10 ng l-1, while esfenvalerate causes behavioural changes at 1 ng l-1. All of the pyrethroids studied consequently have effects at concentrations which are barely quantifiable; for comparison, the requirements for the detection limit in the analytical techniques used for monitoring pesticides in freshwater is 10 ng l-1.
3) that brief pulse exposure to lambda-cyhalothrin results in delayed effects on behaviour and survival in freshwater invertebrates; effects which may be observed for 2-3 weeks after the pulse exposure has ceased.
4) that predator-prey-interactions are affected at environmentally realistic pulse exposures to lambda-cyhalothrin.
5) that pulse exposure to lambda-cyhalothrin results in marked indirect effects on the growth of the algal biomass and the breakdown of organic matter, indicating that environmentally realistic exposure of stream invertebrate communities to lambda-cyhalothrin may lead to effects at the ecosystem level, beyond the direct effects on specific species and populations.
The results of the project demonstrate that the methods utilized complement each other, i.e. the cost-effective laboratory studies can be employed for rapid screening of potential effects of pesticides on the locomotory behaviour of freshwater invertebrates in order to establish effect concentrations and rank ordering of species sensitivities. These data may be utilized in e.g. the planning of stream microcosm experiments. Concurrently, observations from more complex stream microcosm studies, such as effect studies at the community level, may be elucidated by subsequent laboratory studies of locomotory behaviour, recovery and interspecific interactions.
1 Indledning
1.1 Baggrund og nuværende videngrundlag
1.1.1 Pesticider i vandløbssystemer
Anvendelse af pesticider på dyrkede arealer kan ved afstrømning fra behandlede marker føre til pulskontaminering af vandløb (Mathiessen et al. 1995; Felding et al. 1997; Liess et al. 1999; Schulz & Liess 1999a, b). Pesticidtilledning til vandløbene sker typisk, når en marksprøjtning efterfølges af større mængder nedbør, hvorved pesticiderne transporteres til vandløbet ved overfladisk afstrømning eller eventuelt via drænrør og giver anledning til kortvarige pesticidpulse. Pesticider tilføres endvidere ved ulovlige forureningshændelser, f.eks. direkte oversprøjtning eller udslip fra punktkilder som vaske- og fyldepladser.
I Danmark er der i perioden 1989-1996 påvist 32 forskellige pesticider i vandløb (Bichel-udvalget 1999), og undersøgelser i bl.a. fynske vandløb har ydermere dokumenteret forekomst af op til 18 pesticider i samme vandløb inden for ét døgn (Fyns Amt 1997). Forskellige undersøgelser antyder, at mellem 0,01 og 0,3% af den anvendte pesticidmængde - typisk i forbindelse med nedbørshændelser - ender i vandløb (Bichel-udvalget 1999). Som det fremgår af Bichel-rapporten (Bichel-udvalget 1999), er effekterne af disse pesticider i overfladevandet endnu kun ufuldstændigt klarlagte.
For de fleste pesticider fundet i danske vandløb gælder, at de målte koncentrationer ikke kan forventes at være akut letale. For visse insekticider, især pyrethroiderne, har en række nyere udenlandske studier imidlertid vist, at pulskontaminering af vandløb i forbindelse med almindelig landbrugsdrift kan føre til effekter på populations- og økosystemniveau (oversigt i Schulz & Liess 1999b).
1.1.2 Behov for nye metoder
Traditionelle metoder til vurdering af pesticiders toksicitet i akvatiske systemer bygger primært på mortalitetsstudier med kontinuert/kronisk eksponering af et begrænset antal arter. Der tages i disse studier ikke højde for pesticid-kontamineringens kortvarige natur. Hidtil har kun begrænset information om pesticidpulsvarighed og –niveau været tilgængelig, men et nyudviklet modelbaseret værktøj til at vurdere risikoen for pesticideksponering af vandløb udfra oplandsoplysninger kan delvist anvendes til at forudsige disse forhold (Projektet ”Model based tool for evaluation of exposure and effects of pesticides in surface waters” støttet af Miljøstyrelsen siden 1998). Derfor vil et værktøj, der kan forudsige økosystemeffekter af de modellerede koncentrationer af pesticider, være meget værdifuldt.
1.1.3 Drift hos vandløbsinvertebrater
Et af de centrale fænomener, der observeres i forbindelse med kortvarige insekticidpulse i vandløb, er drift, hvor en lang række vandløbsinvertebrater føres med strømmen og helt eller delvist forsvinder fra den belastede vandløbsstrækning. Drift kan opdeles i to hovedgrupper: Adfærdsdrift og katastrofedrift (Waters 1965). Adfærdsdrift er et resultat af invertebraternes aktivitet på overfladen af substratet som regel i forbindelse med fødesøgning eller f.eks. flugt fra en prædator. Katastrofedrift kan skyldes forstyrrelser af substratet eller forurening med miljøfremmede stoffer og er karakteriseret ved, at et meget stort antal individer går i drift samtidig og ikke nødvendigvis på et tidspunkt, hvor invertebraterne normalt er aktive p.gr.a. fødesøgning. Driftpotentialet varierer fra art til art, hvilket bl.a. hænger sammen med arternes varierede evne til at genfinde substratet (Ciborowski & Corkum 1980; Elliott 2002a). Hos mange arter af invertebrater observeres den største drift ved lave lysintensiteter (f.eks. Allan et al. 1986). Denne natdrift afspejler, at risikoen for prædation generelt er mindst ved lave lysintensiteter, hvorfor hovedparten af vandløbsinvertebrater henlægger deres fødesøgning til natten (f.eks. Allan 1995).
1.1.4 Bevægelsesadfærd og pyrethroideksponering
Kontaminering af vandløb med methoxychlor, cypermethrin, permethrin, fenvalerat og ethylparathion har i feltundersøgelser forårsaget drift hos ferskvandsinvertebrater (oversigt i Schulz & Liess 1999b), og i mesocosmforsøg er bl.a. lindan og kobber vist at inducere drift (Girling et al. 2000). Pesticider kan påvirke vandløbsdyrenes adfærd på følgende to måder: Enten detekterer dyret pesticidet via dets sansesystem og ændrer adfærd; eller også absorberes pesticidet og har en intern virkning, der fører til ændret adfærd. Påvisning af sådanne subletale effekter på vandløbsinvertebrater har nødvendiggjort udviklingen af nye metoder til kvantificering af pesticidinducerede adfærdsændringer.
I forbindelse med to afsluttede pesticidforskningsprojekter (Nørum & Bjerregaard 2003; Møhlenberg et al. 2004) er der udviklet nye laboratorie- og strømrendebaserede metoder, der bør kobles for at realisere/validere omkostningseffektive metoder til påvisning af subletale effekter på økologisk relevante adfærdsparametre hos vandløbsinvertebrater. De udviklede metoder har i nævnte projekter vist sig at være særdeles følsomme, og markante effekter af pyrethroiderne cypermethrin, esfenvalerat og lambda-cyhalothrin i koncentrationer ned til få ng/l er blevet observeret. Til sammenligning er der i Århus Amt fundet koncentrationer på op til 660 ng/l, og i Fyns Amt op til 200 ng/l, af pyrethroidet esfenvalerat i vandløb (Fyns Amt 1997; Århus Amt 1999). I Århus Amt blev esfenvalerat fundet i koncentrationer højere end 100 ng/l i 20% af alle udtagne vandprøver. Pyrethroiderne er arealmæssigt set den mest anvendte insekticidstofgruppe, og i 2002 udgjorde pyrethroidernes andel 80% af det samlede teoretiske antal insekticidbehandlede hektarer (Miljøstyrelsen 2003).
1.2 Formål
Det primære formål med projektet har været at koble subletale effektstudier i laboratoriet med effektstudier under mere miljørealistiske forhold i strømrender. Hypotesen har været, at såfremt denne kobling kunne dokumenteres, ville de mindre omkostningstunge laboratorieforsøg kunne bruges til direkte at ekstrapolere til effekter i naturlige vandløb. Hypotesen er blevet testet ved at gentage eksponeringsscenarier undersøgt i laboratoriet i kontrollerede eksperimenter i strømrender, der har en højere grad af overensstemmelse med naturlige forhold end laboratoriet.
Derudover havde projektet til formål at belyse mere komplekse effekter af pyrethroider på de økologiske forhold i vandløb. Hypotesen har været, at samspillet mellem organismer, og den funktion organismerne har i økosystemet, påvirkes af subletale eksponeringer. Hypotesen er blevet testet ved både i laboratoriet og strømrender at undersøge samspillet mellem rovdyr og byttedyr, samt i strømrender at undersøges hvorledes en gruppe af organismer responderer på en pulseksponering med pyrethroid, målt som adfærdsændringer, overlevelse og funktion.
2 Metoder
- 2.1 Metodebeskrivelse
- 2.2 Forsøgsorganismer
- 2.3 Bevægelsesadfærd i laboratoriet
- 2.3.1 Overordnet forsøgsstrategi
- 2.3.2 Forsøgsbetingelser
- 2.3.3 Registrering af bevægelsesadfærd i laboratoriet
- 2.3.4 Generelt forsøgsdesign
- 2.3.5 Relativ toksicitet af pyrethroider anvendt i Danmark
- 2.3.6 Pulsvarighedens betydning
- 2.3.7 Temperaturens betydning
- 2.3.8 Pesticidblanding
- 2.3.9 Koncentrations-respons-forsøg med udvalgte arter
- 2.3.10 Restitueringsforsøg
- 2.3.11 Rovdyr-byttedyrs-interaktion
- 2.3.12 Statistisk metode
- 2.4 Drift i strømrender
- 2.5 Oversigt over udførte forsøg
- 2.6 Pesticidvalg
- 2.7 Kvantificering af lambda-cyhalothrin
2.1 Metodebeskrivelse
Bevægelsesadfærden hos vandløbsinvertebrater anbragt i glaspetriskåle i laboratoriet blev registreret ved hjælp af et computerstyret videosporingsystem før, under og efter forskellige former for pulseksponering for pyrethroider, primært lambda-cyhalothrin. Effekten på bevægelsesadfærden blev sammenholdt med lambda-cyhalothrins indvirkning på drift og effekter i flerarts-forsøg i udendørs strømrender. Laboratorieforsøgene blev udført ved Biologisk Institut, SDU, mens strømrendeforsøg blev udført hos Afd. for Ferskvandsøkologi, DMU, Silkeborg. For at opnå realistiske koncentrations-respons-sammenhænge blev de reelle koncentrationer af pyrethroidet lambda-cyhalothrin bestemt i såvel laboratorie- som strømrendestudierne.
2.2 Forsøgsorganismer
2.2.1 Valg of forsøgsorganismer
De anvendte forsøgsorganismer er blevet udvalgt på baggrund af følgende kriterier:
1. De skulle være repræsentative for de mest udbredte taxonomiske grupper af vandløbsinvertebrater i Danmark.
2. De skulle have forskellig tolerance overfor pesticider og andre påvirkninger.
3. De skulle være forskellige med hensyn til livshistorie, fødebiologi og deres forventede adfærdsrepons overfor pesticider (nogle forventedes at øge aktiviteten, andre ikke).
4. De skulle være robuste under forhold med ikke-strømmende vand, så de kunne anvendes i videosporingssystemet i laboratoriet.
Da der ikke var vandbevægelse i de til videosporingsforsøgene anvendte petriskåle, ville visse vandløbsinvertebrater ikke kunne overleve i laboratorieopstillingen. De fem testorganismer, der blev anvendt i såvel laboratorie- som strømrendeforsøg, inkluderede ferskvandstangloppen Gammarus pulex, slørvingen Leuctra nigra, døgnfluen Heptagenia sulphurea, vårfluen Sericostoma personatum, og huesneglen Ancylus fluviatilis. Endvidere blev vandbænkebideren Asellus aquaticus anvendt til laboratorieforsøg. Indsamlingslokaliteterne er angivet i Tabel 2.1.
Tabel 2.1. Indsamlingslokalitet for forsøgsorganismer anvendt i laboratoriet (L) og strømrenderne (S)
| Art | Indsamlingslokalitet |
| Gammarus pulex | Lindved Å (L), Hagenstrup Møllebæk (S) |
| Asellus aquaticus | Lindved Å (L) |
| Leuctra nigra | Velling Skov (Unavngivne skovvandløb) (L, S) |
| Heptagenia sulphurea | Brende Å (L), Mattrup Å (S) |
| Sericostoma personatum | Rold Kilde (L, S) |
| Ancylus fluviatilis | Havbæk (L, S) |
2.2.2 Opmåling af forsøgsdyr
Hovedparten af alle invertebrater anvendt i laboratorieforsøgene samt fra strømrendeforsøgene blev ethanolkonserveret og efterfølgende målt, således at deres størrelsesfordeling mellem de enkelte forsøg eventuelt kunne inddrages i vurderingen af resultaterne. For at kunne relatere størrelsen af de anvendte invertebrater til andre undersøgelser blev tørvægten beregnet på baggrund af tidligere beskrevne relationer mellem længden af en given morfologisk enhed (f.eks. bredden af hovedkapslen) og dyrenes tørvægt (Zelinka & Marvan 1976; Iversen & Jessen 1977; Friberg et al. 2002). For visse arter var det nødvendigt at etablere nye relationer (A. aquaticus og A. fluviatilis). Dette blev gjort ved at indsamle nye individer på indsamlingslokaliteterne og dernæst måle den relevante morfologiske enhed. Derefter blev dyrene tørret, uden først at være konserveret, til konstant vægt ved 60 °C. Ved hjælp af simpel lineær regression blev der dernæst opstillet en model, hvorved den morfologiske enhed kan konverteres til en tørvægt. Tabel 2.2 viser gennemsnitsvægt og standard error på de individer af invertebrater, der er indgået i de enkelte eksperimenter.
Tabel 2.2. Tørvægt af de anvendte individer af invertebrater (gns ± SEM), samt længde-tørvægts-relationer
| Art | Eksperiment | Tørvægt (mg) |
| Laboratorieforsøg | ||
| Gammarus pulex | Koncentrations- responsforsøg med udvalgte arter |
6,55±0,38 |
| Asellus aquaticus | 2,31±0,09 | |
| Leuctra nigra | 0,25±0,01 | |
| Heptagenia sulphurea | 0,34±0,08 | |
| Sericostoma personatum | 5,09±0,11 | |
| Ancylus fluviatilis | 4,97±0,21 | |
| Strømrendeforsøg | ||
| Gammarus pulex | 1-10 ng l-1 100-1000 ng l-1 Nat 10-100 ng l-1 Fisk 1-10 ng l-1 |
4,29±0,10 4,45±0,10 4,65±0,11 4,84±0,10 |
| Leuctra nigra | 1-10 ng l-1 100-1000 ng l-1 |
0,29±0,007 0,30±0,007 |
| Heptagenia sulphurea | 1-10 ng l-1 100-1000 ng l-1 |
1,64±0,07 0,51±0,02 |
| Sericostoma personatum | 100-1000 ng l-1 | 2,32±0,08 |
| Ancylus fluviatilis | 100-1000 ng l-1 | 14,01±0,23 |
2.3 Bevægelsesadfærd i laboratoriet
2.3.1 Overordnet forsøgsstrategi
Der blev gennemført en række forsøg med G. pulex, der havde det dobbelte formål at optimere forsøgsdesignet til eksponering af de øvrige arter af invertebrater samt at dokumentere generalisérbarheden af de opnåede resultater med henblik på ekstrapolering af observerede effekter på bevægelsesadfærden i laboratoriet til effekter på drift i strømrenderne.
Indledningsvis blev eksponeringsforsøg med fem pyrethroider gennemført for at undersøge, hvorvidt det var muligt at evaluere eventuelle skadevirkninger af pyrethroider som stofgruppe på baggrund af studier af ét modelpyrethroid. Betydningen af pulseksponeringens varighed for adfærdsresponset blev undersøgt med henblik på fastlæggelse af modelpulsvarigheden. Da der i de planlagte laboratorieforsøg ville blive anvendt forskellige vandtemperaturer, og da der forventeligt ville være visse temperatursvingninger i strømrendeforsøgene, blev temperaturens indvirkning på adfærdsresponset hos G. pulex undersøgt i det temperaturinterval, som forventedes anvendt, d.v.s. ca. 5-15 °C. Endelig blev et mindre forsøg gennemført med G. pulex eksponeret for modelpyrethroidet i kombination med et af de hyppigst forekommende pesticider i vandløb, glyphosat, for at vurdere, hvorvidt denne samtidige tilstedeværelse ændrede adfærdsresponset.
Efter valget af modelpyrethroid og modelpulsvarighed blev eksponeringsforsøg med krebsdyrene G. pulex og A. aquaticus, slørvingen L. nigra, døgnfluen H. sulphurea, vårfluen S. personatum og sneglen A. fluviatilis gennemført for at øge vidensgrundlaget for vurderingen af effekter på vandløbsinvertebrater. Med henblik på at opnå større indsigt i mulige langtidsvirkninger af kortvarige pulseksponeringer blev restitueringsforsøg af tre ugers varighed gennemført med G. pulex og A. aquaticus. Mulige effekter af pyrethroider på interspecifikke interaktioner blev undersøgt ved et studie af rovdyr-byttedyrs-interaktionen mellem G. pulex (rovdyr) og L. nigra (byttedyr).
2.3.2 Forsøgsbetingelser
Opbevaring af dyr og udførelsen af forsøg foregik ved SDU i Biologisk Instituts akvarierum ved konstant temperatur med en fotoperiode på 12 timers lys og 12 timers mørke. Ved hjemkomst til akvarierummet blev dyrene, efter et døgns temperaturakklimering i gennemluftet åvand, overført til kunstigt ferskvand (ISO 6431 test water, OECD 2000) (Tabel 2.3). G. pulex og A. aquaticus blev herefter akklimeret i mindst en uge ved 15±1 °C inden forsøgsstart. For at minimere dødeligheden af de indsamlede forsøgsorganismer blev L. nigra, S. personatum og A. fluviatilis i stedet akklimeret i 1-3 dage ved 6±1 °C inden forsøgsstart, mens H. sulphurea blev akklimeret 1 dag ved 10±1 °C.
Tabel 2.3. Kunstigt ferskvand (ISO 6431 test water, OECD 2000)
| Salt | Koncentration (mg/l) |
| CaCl2×2H2O | 294 |
| MgSO4×7H2O | 123,25 |
| NaHCO3 | 64,75 |
| KCl | 5,75 |
2.3.3 Registrering af bevægelsesadfærd i laboratoriet
Et state-of-the-art videosporingssystem, EthoVision Pro (Noldus Information Technology, Holland) blev anvendt til kvantificering af bevægelsesadfærd. Hardware- og softwareopsætning samt adfærdsarena- og eksponeringsdesign er tidligere blevet optimeret i forbindelse med et pesticidforskningsprojekt (Nørum & Bjerregaard 2003) (Figur 2.1).
Figur 2.1. Videosporingssystemet (EthoVision Pro).
Med videosporingssystemet kvantificeres dyrenes bevægelsesmønstre ved vektorregning (Figur 2.2) og nedbrydes i en række adfærdsparametre. F.eks. er følgende adfærdsparametre blevet anvendt for Gammarus pulex i et tidligere projekt (Nørum & Bjerregaard 2003):
- tilbagelagt afstand
- gennemsnitshastighed i bevægelse
- andel af henholdsvis svømmeadfærd, kravleadfærd og inaktivitet
- gennemsnitlig varighed af en svømmebevægelse
- antal svømmebevægelser
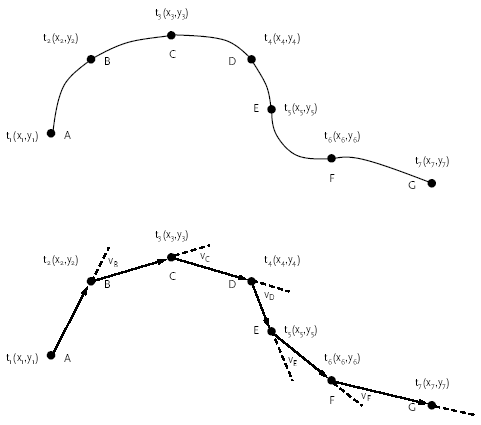
Figur 2.2. Kvantificering af bevægelsesadfærd ved vektorregning.
Den faktiske bevægelsesadfærd (øverst) estimeres ved vektoregning (nederst) baseret på rådatafiler indeholdende information om ændringer i objektets x,y-koordinater over tid.
En lang række yderligere adfærdsparametre kan beregnes v.hj.a. videosporingssystemets software. I nærværende projekt præsenteres kun resultater for parameteren ”tilbagelagt afstand”, da det er det mest integrerede og lettest fortolkelige respons til sammenligning af arternes relative følsomhed over for pyrethroideksponering.
2.3.4 Generelt forsøgsdesign
Designet af et typisk forsøg er følgende: Dyrene akklimeres natten over i glaspetriskåle (diameter: 9 cm) indeholdende 80 ml kunstigt ferskvand. Alle 16 dyrs baggrundsadfærd registreres derefter i en kontrolperiode (enten 30 eller 90 minutter) i vand, der ikke er tilsat pyrethroid. Dernæst afbrydes videosporingen midlertidigt. Det udvalgte pyrethroid, opløst i ethanol + vand (20 ml), tilføres 8 af petriskålene (den eksponerede gruppe). De resterende 8 petriskåle (kontrolgruppen) tilføres tilsvarende mængde ethanol + vand. Det anvendte volumen på 20 ml er tilstrækkeligt til at sikre fuldstændig opblanding i petriskålene. Tilsætningen af væsken foretages længst muligt fra dyrets position, og videosporingen genoptages 2 minutter efter afbrydelsen. Kontrolgruppens og den eksponerede gruppes adfærd følges derefter i endnu en periode (typisk 90 minutter), hvorefter forsøget afsluttes, og en poolet vandprøve (ca. 700 ml) med vand fra den eksponerede gruppes 8 petriskåle indsamles til bestemmelse af aktuelle koncentrationer. Den endelige koncentration af ethanol var 100 µl/l, hvilket ikke overstiger grænserne for opløsningsmidler i OECD Test Guidelines for dafnietest (OECD 2000).
2.3.5 Relativ toksicitet af pyrethroider anvendt i Danmark
Den relative toksicitet af fem pyrethroider (alpha-cypermethrin, cypermethrin, lambda-cyhalothrin, tau-fluvalinat og esfenvalerat) blev undersøgt ved at registrere adfærdsresponset hos eksponerede G. pulex ved 4 forskellige koncentrationer (1, 10, 100 og 1000 ng/l). Eksponeringen varede 90 minutter. Baseret på de opnåede resultater blev lambda-cyhalothrin udvalgt som modelpyrethroid til de efterfølgende studier. Forsøgene blev udført ved 15±1 °C.
2.3.6 Pulsvarighedens betydning
Betydningen af pulseksponeringens varighed for adfærdsresponset blev fastlagt ved eksponering af G. pulex for 4 lambda-cyhalothrin-koncentrationer (1, 10, 100 og 1000 ng/l) med en varighed på op til 5 timer. Baseret på de opnåede resultater er en modelpulsvarighed på 90 minutter blevet valgt. Forsøgene blev udført ved 15±1 °C.
2.3.7 Temperaturens betydning
En undersøgelse af temperaturens indflydelse på toksiciteten blev gennemført ved eksponering af G. pulex for 2 lambda-cyhalothrin-koncentrationer (10 og 100 ng/l) ved 3 temperaturer (5, 10 og 15 °C). Eksponeringen varede 90 minutter.
2.3.8 Pesticidblanding
Effekten af samtidig eksponering for lambda-cyhalothrin og glyphosat hos G. pulex blev undersøgt. Glyphosat blev valgt, da det hyppigt forefindes i danske vandløb. Indledningsvis blev G. pulex eksponeret for glyphosat ved 3 koncentrationer (100, 1000 og 10000 ng/l). Dernæst blev G. pulex eksponeret for lambda-cyhalothrin ved 4 koncentrationer (1, 10, 100 og 1000 ng/l) i kombination med glyphosat (1000 ng/l). Eksponeringen varede 90 minutter. Forsøgene blev udført ved 15±1 °C.
2.3.9 Koncentrations-respons-forsøg med udvalgte arter
Lambda-cyhalothrins akutte effekt på de valgte arter af forsøgsorganismer blev undersøgt ved 5 koncentrationer (0,1-10000 ng/l afhængig af arten, se Tabel 2.6) med henblik på beskrivelse af koncentrations-respons-sammenhænge ved miljørealistiske eksponeringsniveauer. Eksponeringen varede 90 minutter. Forsøgene blev udført ved 10±1 °C, da visse af de anvendte arter forventeligt ville have en dårlig overlevelse ved 15 °C. Da vandtemperaturen kunne tænkes at påvirke lambda-cyhalothrins toksicitet, blev alle invertebraterne testet ved samme temperatur for at sikre sammenligneligheden m.h.t. toksicitetsvurderingen.
2.3.10 Restitueringsforsøg
Restitueringsforsøg er blevet gennemført med G. pulex og A. aquaticus ved 3 lambda-cyhalothrin-koncentrationer (10, 100 og 1000 ng/l) for at vurdere dyrenes evne til at komme sig efter en pulseksponering. Grupper af forsøgsdyr (30 stk fordelt i tre glaspetriskåle med 250 ml vand) blev eksponeret for lambda-cyhalothrin i enten 1½ eller 4½ time i kunstigt ferskvand. Dernæst blev de skyllet forsigtigt, men grundigt, i rindende hanevand, inden de blev overført til ukontamineret kunstigt ferskvand. Overlevelsen og graden af immobilisering blev herefter fulgt i en 21-dages restitueringsperiode, idet hvert individ ved visuel observation blev registreret som værende død, immobil eller aktiv. Døde dyr blev fjernet løbende. Ved restitueringsperiodens udløb blev overlevende, aktive individer videosporet. Forsøgene blev udført ved 15±1 °C.
2.3.11 Rovdyr-byttedyrs-interaktion
Effekten af lambda-cyhalothrin på rovdyr-byttedyrs-interaktionen mellem G. pulex og L. nigra blev undersøgt ved 3 koncentrationer (1, 10 og 100 ng/l). I dette forsøg blev G. pulex akklimeret natten over i glaspetriskålene, mens L. nigra blev akklimeret som samlet gruppe i ét akvarie natten over. Eksponeringen foregik på følgende måde: umiddelbart efter tilsætningen af lambda-cyhalothrin til en petriskål med en G. pulex blev et mindre plastikrør (diameter: 2,7 cm) placeret midt i petriskålen (diameter: 9 cm), og en L. nigra blev overført til røret. Videosporingen blev påbegyndt, og dyrene blev indledningsvis eksponeret i 30 minutter, mens de blev holdt adskilt. Herefter blev plastikrøret fjernet, og interaktionen mellem G. pulex og L. nigra blev fulgt i yderligere 60 minutter under fortsat eksponering. I alt blev følgende antal par observeret: Kontrolgruppe: n=24, 1 ng/l: n=25, 10 ng/l: n=18, 100 ng/l: n=16. Da eventuelle ændringer i rovdyrsadfærden hos G. pulex kunne skyldes, at eksponerede L. nigra ikke var spiselige for G. pulex, blev endnu en forsøgsrække gennemført: L. nigra blev her eksponeret separat for 100 ng/l i 30 minutter, inden de blev overført til ukontamineret vand i plastikrørene, mens G. pulex ikke blev eksponeret. Herefter blev plastikrøret fjernet, og interaktionen mellem G. pulex og L. nigra blev fulgt i 60 minutter. I alt blev 9 par observeret. Forsøgene blev udført ved 15±1 °C. Denne temperatur er i overensstemmelse med den, som generelt blev anvendt til forsøg med G. pulex. For L. nigra’s vedkommende blev en temperatur på 10±1 °C anvendt i ovennævnte koncentrations-respons-forsøg med udvalgte arter. Der var ingen observerbare ændringer i L. nigra’s adfærd i rent vand ved 15±1 °C.
2.3.12 Statistisk metode
Resultater fra videosporingsforsøgene blev primært evalueret ved repeated measures ANOVA. Regressionsanalyse blev anvendt i restitueringsforsøget og i studiet af rovdyr-byttedyrs-interaktionen. Den statistiske analyse blev foretaget ved hjælp af SAS®. I alle tilfælde blev et signifikansniveau på a=0,05 anvendt.
2.4 Drift i strømrender
2.4.1 Overordnet forsøgsstrategi
Der blev gennemført to typer af eksperimenter i strømrender: Drift og flerarts-studier. I drifteksperimenter blev det umiddelbare driftrespons hos vandløbsinvertebrater undersøgt under og efter pulseksponering med lambda-cyhalothrin i semi-naturlige vandløbsrender. Undersøgelserne inkluderede krebsdyret G. pulex, slørvingen L. nigra, døgnfluen H. sulphurea, sneglen A. fluviatilis og vårfluen S. personatum. Vandløbsinvertebraterne blev eksponeret for lambda-cyhalothrin i 90 minutter, hvorefter driften fortsat blev registreret indtil 150 minutter fra eksponeringens start. Der blev målt drift 60 min før eksponering, ved start af eksponeringen og herefter hver 30. minut, i alt 7 gange per forsøgsserie (-60, 0, 30, 60, 90, 120 og 150 minutter). Vandløbsinvertebraterne blev eksponeret for 2 eller 4 koncentrationer af lambda-cyhalohtrin afhængig af arten. Alle eksperimenter med undtagelse af et enkelt med G. pulex blev gennemført om dagen. Eftersom langt de fleste vandløbsinvertebrater udviser størst drift om natten, blev der ligeledes gennemført et forsøg om natten for at undersøge om organismernes, her G. pulex, højere aktivitetsniveau og tilbøjelighed til at gå i drift om natten påvirkede deres driftrespons ved lambda-cyhalothrin-eksponering.
Eksperimenterne med samspil, eller interaktioner, mellem organismerne blev udført i såvel laboratoriet som i strømrenderne. Det samspil, der blev undersøgt, var rovdyr-byttedyrsadfærd. I laboratoriet blev prædations-effektiviteten af krebsdyret G. pulex på slørvingen L. nigra undersøgt ved forskellige koncentrationer af lambda-cyhalothrin. G. pulex ernærer sig primært som detritivor men kan også fungere som et effektivt rovdyr. I strømrender blev det undersøgt, hvorledes duftstoffer fra ørred (Salmo trutta) påvirker G. pulex’s driftadfærd ved forskellige koncentrationer af lambda-cyhalothrin. Det vides fra tidligere undersøgelser, at individer af G. pulex nedsætter deres driftaktivitet, når der er duftstoffer fra ørreder tilstede i vandet (Friberg et al., 1994). Dette er en antiprædator adfærd, der har til formål at reducere sandsynligheden for, at det enkelte individ bliver spist, da ørreder primært indtager deres føde fra driften. Samtidig er det kendt fra tidligere undersøgelser, at G. pulex øger deres driftaktivitet, når de bliver eksponeret for lambda-cyhalothrin (Lauridsen & Friberg, 2005). Ved at tilsætte ørred-duftstoffer til vand, der blev tilledt renderne, kunne det undersøges, hvilke af disse to modsatrettede adfærdsstimuli, der dominerer, og dermed sandsynliggøres om eksponering med lambda-cyhalothrin øger prædationsrisikoen i vandløb.
I flerarts-eksperimenterne blev de længerevarende effekter af eksponering med lambda-cyhalothrin undersøgt med hensyn til direkte og indirekte effekter. Undersøgelsen inkluderede krebsdyret G. pulex, slørvingen L. nigra, døgnfluen H. sulphurea, og sneglen A. fluviatilis udsat i de samme strømrender efter eksponering for to koncentrationer af lambda-cyhalothrin. Eksponering (90 minutter) foregik i laboratoriet dagen før eksperimentets start for at undgå det umiddelbare tab i drift. Forsøget forløb i 10 dage med daglig måling af drift. Derudover blev omsætning af organisk stof og algebiomassen målt for at få indblik i indirekte effekter af pyrethroideksponeringen.
2.4.2 Strømrender
Undersøgelserne blev udført på DMU’s udendørs vandløbsrende-faciliteter i Lemming, 8 km nord for Silkeborg. Systemet består af 12 vandløbsrender som konstant fødes med frisk grundvand, der hele året har en temperatur på 7-8 °C (Figur 2.3). Inden vandet ledes i renderne, føres det ned af en iltningstrappe og gennem et trykfilter, hvor bl.a. partikler og jern bindes (se Tabel 2.4). Fra trykfiltret føres vandet til en fødekasse (50x70x60cm), hvorfra det, via et rørsystem, fordeles ligeligt til alle 12 render, så de har en konstant vandføring på 3,2 l min-1. De 12 vandløbsrender er fire meter lange og 10 cm brede. Faldet på vandløbsrenderne er indstillet til 1%, hvilket svarer til et naturligt fald på strygsekvenser i mindre danske vandløb. Substratet i vandløbsrenderne er udlagt således, at det imiterer en strygsekvens i et mindre dansk vandløb med grus og sten samt bladpakker i partierne med strømlæ (Figur 2.4). Ved den anvendte substratfordeling er vandets gennemsnitlige opholdstid i renderne 58,2 ± 1,3 sekunder og den gennemsnitlige strømhastighed 0,064 m sek-1 (Tabel 2.4). Terminalt i hver vandløbsrende falder/løber hele vandføringen gennem et driftnet (maskevidde 1 mm) som opfanger alle invertebrater større end 1 mm, der føres med strømmen.

Figur 2.3. Foto af forsøgsopstillingen med de 12 vandløbsrender. Driftnettene er placeret i de lodrette nedløbsrør forrest i billedet.
Tabel 2.4: Fysiske og kemiske parametre for vandet i strømrenderne
| Kemiske parametre | Værdi/koncentration |
| NH4 | 0,009 mg l-1 |
| NO2 + NO3 | 2,720 mg l-1 |
| Total N | 2,830 mg l-1 |
| PO4 | 0,003 mg l-1 |
| Total P | 0,003 mg l-1 |
| Total Fe | 0,005 mg l-1 |
| pH | 7,0 |
| Fysiske parametre | |
| Vandføring per rende | 3,2 ± 0,03 l min.-1 |
| Vandets gennemsnitlige opholdstid i renderne | 58,2 ± 1,3 sek. |
| Gennemsnitlig strømhastighed | 0,064 ± 0,001 m sek.-1 |
| Vandtemperatur (range) | 8,1-10,4 °C |
Såvel vandløbsrenderne som sedimentet i vandløbsrenderne blev udskiftet mellem alle forsøgsserier, således at hver forsøgsserie indledtes med at etablere sedimentet. Først blev hver vandløbsrende tilført 4,5 kg småsten (1-3 cm) fordelt jævnt på en 3 meter lang strækning. Småstenene var forinden konditioneret 7 dage i vandløbsvand for at skabe en naturlig biofilm på substratet. Efter fordelingen af gruset blev der udlagt knytnævestore sten for hver 30. centimeter. Disse sten blev indsamlet samme dag som etableringen af vandløbsrenderne i Hulbæk, et lille vandløb 5 km øst for Silkeborg. Inden stenene blev udlagt i vandløbsrenderne, blev de renset for dyr og skidt. Efter udlægningen af sten blev der i strømlæet bag hver sten placeret 2 elleblade (Alnus glutinósa (L)), som forinden var konditioneret 5 dage i kildevand ved 10°C. Efter udlægningen af elleblade blev en Stowaway Tidbit temperatur logger placeret i nedstrømsenden af substratsstrækningerne.

Figur 2.4. Foto af de 3 meter lange substrat-strækninger bestående af grus, sten og elleblade.
2.4.3 Dosering af lambda-cyhalothrin i strømrenderne
Lambda-cyhalothrinopløsningerne blev fremstillet ved at opløse en given mængde stof i 100 ml ethanol (96%). Ud fra denne stamopløsning blev der fremstillet en fortyndingsrække med ethanol til de ønskede koncentrationer. Opløsningerne blev fremstillet således, at der skulle tilsættes 10 ml opløsning til 1 L demineraliseret vand for at opnå de ønskede koncentrationer.
Lambda-cyhalothrin blev doseret med 10 ml min-1 til enkelte render ved hjælp af en peristaltisk pumpe (Ole Dich Type 110 med 12 kanaler). For hvert eksperiment blev den aktuelle vandføring målt i renderne for at beregne, hvor stort et volumen af pesticidopløsning, der skulle doseres til den enkelte rende. Fra hvert eksperiment blev der udtaget vandprøver til pesticidanalyse.
2.4.4 Drifteksperimenter
Drifteksperimenterne blev gennemført i strømrender om dagen med undtagelse af et enkelt forsøg med G. pulex, der blev udført om natten. Alle forsøgsdyr blev introduceret til renderne dagen før eksperimenterne. Om morgenen før eksperimentet blev dyrene, der var driftet ud om natten, erstattet. Inden eksponeringsstart blev der yderligere målt drift i 60 minutter for at sikre, at driften var stabiliseret. Var driften i denne periode meget høj, blev forsøget ikke gennemført.
2.4.5 Flerarts-forsøg
Eksperimentet med effekter på flere arter sammen blev gennemført i april 2005 i perioden fra 11.-21. april. Renderne blev 3 dage før eksperimentets start forsynet med substrat bestående af småsten (1-3 cm i diameter) samt knytnævestore sten. Småstenene blev fordelt jævnt på en strækning på 2,5 meter i hver rende. Småstenene var forinden blevet konditioneret i en måned i næringsrigt vand fra Lemming Å. Småstenene blev her fordelt i et tyndt lag i plastikkar og opbevaret i et klimarum (14 ºC) på DMU’s faciliteter i Silkeborg. Over karrene blev der ophængt UV-lamper, der gav konstant belysning i konditioneringsperioden. Konditioneringen af småstenene skulle sikre etableringen af en naturlig biofilm på stenene. 21 af stenene blev taget fra til måling af baggrundsniveauet i algebiomasse. De knytnævestore sten blev hentet i et mindre unavngivet vandløb ved Sdr. Vissing samme dag som etableringen af renderne. Inden udlægninen blev de vasket rene for snavs og dyr. Der blev placeret 7 sten pr. rende med 25 cm mellemrum. Stenene nummereredes 1-7, startende længst opstrøms. Ved den anvendte substratfordeling var vandets gennemsnitlige opholdstid i renderne 51,6 ± 2,7 sekunder og den gennemsnitlige strømhastighed 0,068 ± 0,003 m sek-1.
Der blev placeret fem bladpakker i hver rende foran de knytnæve store sten (nr. 2-6). Hver bladpakke bestod af 10 bøgeblade (Fagus sylvatica). Bladene blev indsamlet i et lille unavngivet skovvandløb i Velling skov og var således allerede konditioneret inden forsøget. Bladene blev forsynet med en snor, der holdt dem sammen i pakker, og snoren blev fæstnet under de knytnævestore sten, således at bladpakkens placering var sikret under hele forsøget. Bladpakkerne blev placeret samtidig med etableringen af vandløbsrenderne dvs. 3 dage inden forsøget start. Forinden var bladpakkerne blevet vådvejet og 15 blade desuden taget fra til referencemålinger. De fem bladpakker i hver rende blev benævnt A-E i nedstrøms retning. Efter etableringen af vandløbsrenderne blev en Stowaway Tidbit temperatur logger placeret i nedstrømsenden af substratstrækningerne, og temperaturfluktuationerne blev målt i 24 timer (Tabel 2.4).
Lambda-cyhalothrin stamopløsningen blev blandet i 1 L kildevand i plastikkar, og dyrene blev nedsænket heri umiddelbart efter. Dyrene til kontrolforsøget blev nedsænket i kar med den tilsvarende mængde ethanol som blev benyttet i opløsningen med den højeste koncentration. Eksponeringen foregik i klimarum (4 ºC), og karrene med dyr blev beluftet under hele forløbet. De nominelle koncentrationer af lambda-cyhalothrin var 10 og 100 ng l-1. Eksponeringen fandt sted dagen før forsøgets start (d. 9/4-05) og varede 90 minutter. Efter eksponering blev dyrene overflyttet til beluftede kar med rent kildevand i 24 timer. Karrene med G. pulex og L. nigra blev forsynet med bøgeblade, og karrene med H. sulphurea og A. fluviatilis blev forsynet med biofilmbelagte sten.
På forsøgets første dag blev dyrene introduceret i vandløbsrenderne umiddelbart opstrøms for sektionen med substrat. I hver rende blev der udsat 65 G. pulex, 45 L. nigra, 16 H. sulphurea samt 25 A. fluviatilis, hvilket gav de i Tabel 2.5 viste individtætheder. Disse individtætheder ligger indenfor, hvad der naturligt kan forekomme i danske vandløb (Iversen, 1988; Friberg et al. 1994; Friberg, 1997). Der var 4 replikater af hhv. kontrol og behandlingerne med 10 og 100 ng l-1 lambda-cyhalothrin.
Tabel 2.5 Individtætheder i vandløbsrenderne
| G. pulex | L. nigra | H. sulphurea | A. fluviatilis | |
| # individer per rende | 65 | 45 | 16 | 25 |
| Individtæthed (ind. m-2) | 433 | 300 | 107 | 167 |
Igennem hele forsøgsperioden blev driftnettene tømt hver 24 timer, og alle levende individer blev reintroduceret til strømrenderne for at opretholde passende tætheder. Dette var nødvendigt, da renderne ikke er lange nok til, at invertebraterne kan have en naturlig bevægelsesadfærd i et længere tidsrum, herunder at opstrømsmigration ikke er mulig, når først dyrene er fanget i driftnettene. Såfremt invertebraterne ikke var blevet reintroduceret, ville de indirekte effekter på græsning og omsætning af organisk stof alene afspejle forskelle i tætheder i renderne og ikke påvirkningen fra eksponeringen med lambda-cyhalothrin. Ved tømning af hvert driftnet blev dyrene overført til en hvid fotobakke, hvor de blev talt op og deres tilstand vurderet på tre forskellige niveauer; upåvirket (velbevaret flugtrespons ved fysisk berøring), påvirket (hæmmet flugtrespons ved fysisk berøring) og døde. Desuden blev tre småsten sorteret fra i hver rende på 5., 8., og 10. forsøgsdag til klorofylanalyser. Efter den sidste tømning af driftnettene på 10. forsøgsdag blev alle bladpakkerne taget fra til vægtanalyser, og vandløbsrenderne blev tømt. Substratet blev konserveret i ethanol og blev senere grovsorteret under luplampe for at genfinde de tilbageværende dyr. Bladpakkerne blev efterfølgende tørret i 24 timer ved 60 ºC, hvorefter tørvægten blev bestemt. Herefter blev en kendt fraktion af hver bladpakke forasket ved 600 ºC i 24 timer, og den askefri tørvægt blev bestemt. Væksten af mikroalger på småstenene blev undersøgt ved klorofylekstrahering i et kendt volumen ethanol (96%) i 16 timer i mørke, hvorefter en kendt delprøve fra hver ekstrahering blev centrifugeret i 10 minutter ved 2000 omdr. min-1 og målt i et spektrofotometer ved to forskellige bølgelængder (665 nm samt 750 nm). Klorofylindholdet af hver ekstrahering blev beregnet i forhold til hver småstens projicerede areal. Det projicerede areal blev bestemt ved at tegne et omrids af den enkelte sten på papir, hvorefter arealet blev bestemt ved vejning.
2.4.6 Statistisk metode
Alle statistiske analyser er foretaget indenfor en enkelt forsøgsserie bestående af kontrol og to behandlinger. Der er ikke foretaget analyser mellem forsøgsserierne, da de blev gennemført på forskellige tidspunkter i projektperioden og derfor ikke er direkte sammenlignelige. De statistiske metoder, som er blevet anvendt til analyse af data fra strømrenderne (både enkeltarts- og flerarts-forsøgene) i nærværende projekt, er primært traditionel ANOVA samt repeated measures ANOVA, idet de forskellige responsvariable observeres henover tiden for de samme render. Variansanalysen har ligeledes indeholdt parvise sammenligninger med fokus på sammenligninger til kontrolgruppen. Parvise sammenligninger blev gennemført ved brug af en Bonferroni-korrigeret t-test. Yderligere uddybning af den statistiske metode er angivet i Bilag A. De statistiske analyser blev foretaget ved brug af SAS®. For at lette læsbarheden af resultatafsnittet er F- samt t-værdier angivet i tabeller, mens p-værdierne er angivet i selve teksten.
2.5 Oversigt over udførte forsøg
Tabel 2.6 Oversigt over udførte forsøg
| Forsøg | Art | Lambda-cyhalothrin-koncentration (ng/l) |
| Bevægelsesadfærd i laboratoriet | ||
| Relativ toksicitet af fem pyrethroider | Gammarus pulex | 1, 10, 100, 1000 (for hvert af de fem pyrethroider) |
| Pulsvarighedens betydning | Gammarus pulex | 1, 10, 100, 1000 |
| Temperaturens betydning | Gammarus pulex | 10, 100 (ved tre temperaturer) |
| Koncentrations-respons-forsøg | Gammarus pulex Asellus aquaticus Leuctra nigra Heptagenia sulphurea Sericostoma personatum Ancylus fluviatilis |
1, 10, 100, 1000, 10000 1, 10, 100, 1000, 10000 0,1, 1, 10, 100, 1000 1, 10, 100, 1000, 10000 1, 10, 100, 1000, 10000 1, 10, 100, 1000, 10000 |
| Restituering (21 dage) | Gammarus pulex Asellus aquaticus |
10, 100, 1000 |
| Pesticidkombination: lambda-cyhalothrin & glyphosat | Gammarus pulex | 1, 10, 100, 1000 |
| Rovdyr-byttedyrs-interaktion | Gammarus pulex Leuctra nigra |
1, 10, 100 |
| Drift i strømrender | ||
| Koncentrations-respons-forsøg | Gammarus pulex Leuctra nigra Heptagenia sulphurea Sericostoma personatum Ancylus fluviatilis |
1, 10, 100, 1000 1, 10, 100, 1000 1, 10, 100, 1000 100, 1000 100, 1000 |
| Eksponering om natten | Gammarus pulex | 1, 10, 100, 1000 |
| Rovdyr-byttedyrs-interaktion | Salmo trutta Gammarus pulex |
10, 100 |
| Flerarts-forsøg | Gammarus pulex Leuctra nigra Heptagenia sulphurea Ancylus fluviatilis |
10, 100 |
2.6 Pesticidvalg
Valget af pyrethroider var primært baseret på Bekæmpelsesmiddelstatistikken for 2002 (Miljøstyrelsen 2003), hvoraf det fremgik, at pyrethroider blev anvendt til sprøjtning af 80% af alle insekticidbehandlede arealer. Pyrethroiderne udgjorde 4 ud af 9 anvendte insekticider; nemlig alpha-cypermethrin, cypermethrin, lambda-cyhalothrin og tau-fluvalinat, der hver blev sprøjtet på ca. 100.000 ha. Laboratorieforsøg med disse pyrethroider blev gennemført. Endvidere blev det i 2002 forbudte pyrethroid, esfenvalerat, anvendt. Esfenvalerat ønskedes undersøgt, da der under forsøg udført i forbindelse med et tidligere pesticidforskningsprojekt var opnået resultater, der detaljeret beskriver det adfærdstoksikologiske respons bl.a. hos esfenvalerat-eksponerede G. pulex og A. aquaticus (Nørum & Bjerregaard 2003). Endelig foreligger der en række målinger af esfenvalerat-koncentrationer i danske vandløb. Esfenvalerat tænktes derfor anvendt som reference-pyrethroid. De fem valgte pyrethroider er syntetiske insekticider, der virker agonistisk på spændingsafhængige natriumkanaler i nervesystemet hos både target og non-target organismer. Til pesticidkombinationsforsøget blev glyphosat anvendt, da det er hyppigt forekommende i danske vandløb.
Alpha-cypermethrin
CAS Nummer: 67375-30-8
Navn: (R,S)-alpha-cyano-3-phenoxybenzyl(1-R,S)-cis-3-(2,2-dichlorovinyl)-2,2-dimethylcyclopropancarboxylat
Molekylærformel: C22H19Cl2NO3
Molvægt: 416,31
Cypermethrin
CAS Nummer: 52315-07-8
Navn: (R,S)-alpha-cyano-3-phenoxybenzyl(1-R,S)-cis,trans-3-(2,2-dichlorovinyl)-2,2-dimethylcyclopropancarboxylat
Molekylærformel: C22H19Cl2NO3
Molvægt: 416,31
Lambda-cyhalothrin
CAS Nummer: 91465-08-6
Navn: (RS)-alpha-cyano-3-phenoxybenzyl 3-(2-chloro-3,3,3-trifluoropropenyl)-2,2-dimethylcyclopropancarboxylat
Molekylærformel: C23H19Cl F3NO3
Molvægt: 449,86
Tau-fluvalinat
CAS Nummer: 102851-06-9
Navn: (RS)-alpha-cyano-3-phenoxybenzyl N-(2-chloro-a,a,a-trifluoro-p-tolyl)-D-valinat
Molekylærformel: C26H22Cl F3 N2O3
Molvægt: 502,92
Esfenvalerat
CAS Nummer: 66230-04-4
Navn: (S)-alpha-cyano-3-phenoxybenzyl(S)-2-(4-chlorophenyl)-3-methylbutyrat
Molekylærformel: C25H22ClNO3
Molvægt: 419,91
Glyphosat
CAS Nummer: 1071-83-6
Navn: N-phosphomethylglycin
Molekylærformel: C3H8NO5P
Molvægt: 169,07
2.7 Kvantificering af lambda-cyhalothrin
2.7.1 Metodebeskrivelse
For at opnå realistiske koncentrations-respons-sammenhænge mellem pyrethroidkoncentration og indvirkning på bevægelses- og driftadfærd er de reelle koncentrationer af lambda-cyhalothrin blevet bestemt i såvel laboratorie- som strømrendeeksponeringssystemerne. Metoden bygger på erfaringer fra et tidligere pesticidforskningsprojekt (Nørum & Bjerregaard 2003), hvor metoder til bestemmelse af bl.a. pyrethroiderne cypermethrin og esfenvalerat blev udviklet. Metodens princip er baseret på fastfaseekstraktion af pesticidkontamineret ferskvand efterfulgt af opkoncentrering ved inddampning, samt kvantificering ved omvendt fase HPLC-MS. Metoden er baseret på vandprøver med et volumen på 100-1000 ml.
2.7.1.1 Forbehandling af vandprøver
Umiddelbart efter opsamling af afmålt vandprøve indeholdende lamdba-cyhalothrin tilsættes en kendt mængde intern standard (esfenvalerat).
2.7.1.2 Fastfaseekstraktion
Ekstraktion af lambda-cyhalothrin fra vandprøven foretages på en C18-søjle (Sep Pak Vac, 6 cc, 1 g, C18 cartridges, fra Waters). Søjlen konditioneres med 5 ml methanol og vaskes med 5 ml Milli-Q vand. Prøven påsættes søjlen ved 20 kPa vacuum, svarende til ca. 3 ml/min. Dernæst vaskes søjlen med 5 ml Milli-Q vand og tørres i 1-2 minutter ved 30-40 kPa vacuum. Pesticidet elueres med 4 ml methanol. Eluatet inddampes efterfølgende til tørhed under luftstrøm og genopløses i 0,300 ml 75% methanol.
2.7.1.3 LC-MS analyse
Det anvendte Hewlett Packard LC-MSD-system består af en HP Series 1100 HPLC (solvent degasser, binær pumpe, autosampler og termostatreguleret kolonneafdeling) og et G1946A MSD quadropole massespektrometer udstyret med electrospray ionisation (ESI) i positiv mode. Til kvantificeringen anvendes en HPLC-kolonne (C18, 150×2,1 mm, Phenomenex fra Subware) med forkolonne af samme materiale. Der anvendes et væskeflow på 0,4 ml/min, en injection af 50 µl prøve, og en kolonnetemperatur på 25 °C. Følgende LC solventer blev brugt: Eluent A: 10 mM ammoniumacetat:methanol, 990:10 (v:v). Eluent B: 10 mM ammoniumacetat:methanol, 10:90 (v:v). Der elueres med følgende gradient (Tid, % B): (0 min, 75%); (3 min, 100%); (14 min, 100%); (14,1 min, 75%); postrun-tid: 6 min, 25% B. Massespektrometer-opsætning: Mode: ESI positiv (SIM: m/z 467 for lambda-cyhalothrin). Der benyttes intern standardisering med esfenvalerat (SIM: m/z 437). Drying gas temperature: 350 °C. Drying gas flow: 10 l/min. Nebulizer pressure: 30 psig. Capillary voltage: 3500 V. Fragmentor: 50 V. Standardkurven beregnes udfra standarderne: 0,7; 3,5; 35,0; 70,0 og 350 ng/ml injiceret standard. Lambda-cyhalothrin og esfenvalerat elueres efter henholdsvis 7,4 min og 8,0 min.
3 Resultater
- 3.1 Kvantificering af lambda-cyhalothrin
- 3.2 Bevægelsesadfærd i laboratoriet
- 3.3 Drift i strømrender
3.1 Kvantificering af lambda-cyhalothrin
Nominelle og aktuelle koncentrationer af lambda-cyhalothrin i de udførte laboratorie- og strømrendeforsøg er angivet i Tabel 3.1 og 3.2. De aktuelle koncentrationer ligger typisk på 70-80% af de nominelle, hvilket er acceptabel i betragtning af stoffets fysisk-kemiske egenskaber. Tab af stof fra vandfasen er forventelig bl.a. på grund af adsorption til forsøgsopstilling. I resultat-afsnittet angives de nominelle koncentrationer i tekst, tabeller og figurer.
Tabel 3.1. Nominelle og aktuelle koncentrationer i laboratorieforsøgene
| Lambda-cyhalothrin-konc. (ng/l) | |||
| Forsøg | Art | Nominel | Aktuel |
| Relativ toksicitet af fem pyrethroider | Gammarus pulex | 1, 10, 100, 1.000 | iv |
| Pulsvarighedens betydning | Gammarus pulex | 1, 10, 100, 1.000 | iv |
| Temperaturens betydning | Gammarus pulex | 10 (5 °C) | 8,8 |
| 10 (10 °C) | 10,7 | ||
| 10 (15 °C) | 10,1 | ||
| 100 (5 °C) | 60,7 | ||
| 100 (10 °C) | 53,9 | ||
| 100 (15 °C) | 53,9 | ||
| Koncentrations-respons-forsøg | Gammarus pulex | 1 | iv |
| 10 | 10,8 | ||
| 100 | 78,3 | ||
| 1.000 | 582 | ||
| 10.000 | 6.793 | ||
| Asellus aquaticus | 1 | iv | |
| 10 | 14,1 | ||
| 100 | 75,9 | ||
| 1.000 | 587 | ||
| 10.000 | 7.868 | ||
| Leuctra nigra | 0,1 | iv | |
| 1 | iv | ||
| 10 | ik | ||
| 100 | 99,5 | ||
| 1.000 | 862 | ||
| Heptagenia sulphurea | 1 | iv | |
| 10 | 12,5 | ||
| 100 | iv | ||
| 1.000 | 1.100 | ||
| 10.000 | 11.576 | ||
| Sericostoma personatum | 1 | iv | |
| 10 | 11,0 | ||
| 100 | 96,2 | ||
| 1.000 | 721 | ||
| 10.000 | 7.283 | ||
| Ancylus fluviatilis | 1 | iv | |
| 10 | ik | ||
| 100 | 132,3 | ||
| 1.000 | 839 | ||
| 10.000 | 5.887 | ||
| Restituering (21 dage) | Gammarus pulex Asellus aquaticus |
10, 100, 1000 | iv |
| Pesticidkombination: lambda-cyhalothrin & glyphosat | Gammarus pulex | 1, 10, 100, 1000 | iv |
| Rovdyr-byttedyrs-interaktion | Gammarus pulex Leuctra nigra |
1 | iv |
| 10 | 6,6 | ||
| 100 | 62,1 | ||
iv: ingen vandprøve, ik: ikke kvantificeret
Tabel 3.2. Nominelle og aktuelle koncentrationer i strømrendeforsøgene
| Lambda-cyhalothrin-konc. (ng/l) | |||
| Forsøg | Art | Nominel | Aktuel (gns±SEM, n=4) |
| Koncentrations-respons-forsøg | Gammarus pulex | 0 | id |
| 1 | id | ||
| 10 | ik | ||
| 100 | 58,4 ± 3,5 | ||
| 1.000 | 488 ± 45 | ||
| Leuctra nigra | 0 | id | |
| 1 | id | ||
| 10 | ik | ||
| 100 | 79,1 ± 14,3 | ||
| 1.000 | 760 ± 87 | ||
| Heptagenia sulphurea | 0 | id | |
| 1 | iv | ||
| 10 | iv | ||
| 100 | 81,0 ± 15,9 | ||
| 1.000 | 713 ± 78 | ||
| Sericostoma personatum | 0 | id | |
| 100 | 45,2 ± 6,7 | ||
| 1.000 | 426 ± 177 | ||
| Ancylus fluviatilis | 0 | id | |
| 100 | 80,7 ± 26,2 | ||
| 1.000 | 777 ± 102 | ||
| Eksponering om natten | Gammarus pulex | 0 | id |
| 10 | 7,4 ± 4,7 | ||
| 100 | 79,5 ± 3,3 | ||
| Rovdyr-byttedyrs-interaktion | Salmo trutta Gammarus pulex |
0 | id |
| 10 | ik | ||
| 100 | 96,5 ± 11,4 | ||
| Flerarts-forsøg | Gammarus pulex Leuctra nigra Heptagenia sulphurea Ancylus fluviatilis |
0 | id |
| 10 | ik | ||
| 100 | 92,0 ± 3,0 | ||
id: ikke detekteret, iv: ingen vandprøve, ik: ikke kvantificeret
3.2 Bevægelsesadfærd i laboratoriet
3.2.1 Relativ toksicitet af pyrethroider anvendt i Danmark
Figur 3.1 viser effekten af fem pyrethroider på den tilbagelagte afstand hos G. pulex ved 4 forskellige koncentrationer (1, 10, 100 og 1000 ng l-1). For alle fem pyrethroider observeres et bifasisk adfærdsrespons. Ved lave koncentrationer (1 ng l-1 for esfenvalerat og 10 ng l-1 for de øvrige) observeres en markant forøgelse af den tilbagelagte afstand til et konstant maksimum på ca. 500-600 cm/5 min; dog noget lavere for alpha-cypermethrin. Denne hyperaktivitet opretholdes i resten af eksponeringsperioden. Med stigende koncentration nås dette maksimale, hyperaktive niveau tidligere og tidligere, mens varigheden af maksimumsniveauet aftager. Ved 100 ng l-1 efterfølges hyperaktiviteten af et gradvist fald i tilbagelagt afstand, og ved 1000 ng l-1 observeres immobilisering efter ca. 45-60 minutters eksponering for alle pyrethroiderne pånær tau-fluvalinat. For tau-fluvalinats vedkommende ses ved 1000 ng l-1 et tydeligt reduceret aktivitetsniveau, men ikke fuldstændig immobilisering inden eksponeringsperiodens udløb. Overordnet set vurderes forskellen mellem de fem pyrethroider at være relativt lille, og lambda-cyhalothrin er i de efterfølgende studier anvendt som modelpyrethroid, da det på daværende tidspunkt var det mest anvendte af pyrethroiderne målt på sprøjtet areal (Miljøstyrelsen 2003).
3.2.2 Pulsvarighedens betydning
Figur 3.2 viser betydningen af pulseksponeringens varighed for den tilbagelagte afstand hos G. pulex ved 4 lambda-cyhalothrin-koncentrationer (1, 10, 100 og 1000 ng l-1) med en varighed på op til 5 timer. Det bifasiske adfærdsrespons efter 90 minutters eksponering er sammenligneligt med det tidligere observerede for lambda-cyhalothrin (Figur 3.1). Der er ingen markante effekter af 1 ng l-1. Hyperaktivitet observeres ved 10 ng l-1, hvilket efterfølges af et gradvist fald i bevægelsesaktivitet ved 100 ng l-1 og en hurtig immobilisering ved 1000 ng l-1. Ved længere eksponeringsvarigheder bliver de observerede adfærdsændringer mere markante. Ved 10 ng l-1 kan hyperaktiviteten ved længere tids eksponering ikke opretholdes, og aktiviteten falder til et niveau, der svarer til niveauet i kontrolperioden. Reduktionen i tilbagelagt afstand ved 100 ng l-1 bliver mere markant ved længere eksponeringsvarigheder, og efter ca. 2½ time nås et aktivitetsniveau, hvor dyrene tilsyneladende ikke længere udviser svømmeadfærd, men stadig ikke er fuldt immobiliserede. Ved 1000 ng l-1 resulterer en forlængelse af pulsvarigheden ud over ca. 60 minutter ikke i yderligere ændringer, da alle dyrene allerede er immobiliserede. På baggrund af ovenstående blev det besluttet at anvende en modelpulsvarighed på 90 minutter til de efterfølgende studier, da der ved længere eksponerings-varigheder ikke observeres et kvalitativt anderledes adfærdsrespons hos G. pulex.
Figur 3.1. Effekt af pyrethroider på tilbagelagt afstand
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 90 min, og adfærden i pyrethroid-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=6-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
Figur 3.2. betydningen af pulseksponeringens varighed for den tilbagelagte afstand
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 5 timer (300 min). Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=6-8). Asterisk angiver signifikant forskel mellem de to grupper ved en eksponeringsvarighed indtil den stiplede linie umiddelbart til højre for asterisken, d.v.s. ½, 1, 1½, 2, 3, 4 eller 5 timers eksponering.
3.2.3 Temperaturens betydning
Figur 3.3 viser betydningen af temperaturen for den tilbagelagte afstand hos G. pulex ved 2 lambda-cyhalothrin-koncentrationer (10 og 100 ng l-1) ved 3 temperaturer (5, 10 og 15 °C). De foregående studier til udvælgelse af modelpyrethroid og –pulsvarighed blev gennemført ved 15 °C. Effekten af en lavere temperatur er hos G. pulex et mindre markant og lidt forsinket adfærdsrespons. Ved såvel 10 som 100 ng l-1 aftager maksimumsniveauet for den tilbagelagte afstand med faldende temperatur, samtidig med at tidspunktet for maksimalværdien forsinkes lidt.
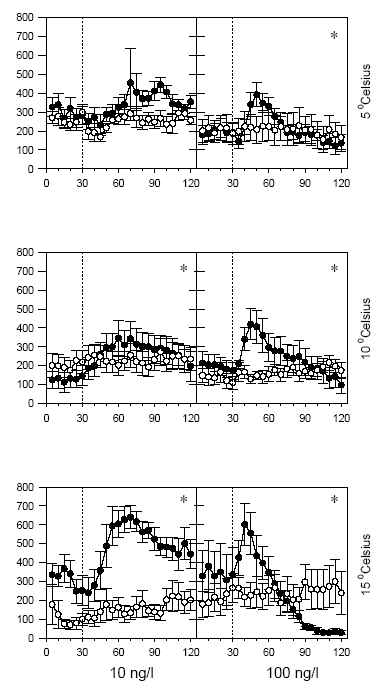
Figur 3.3. Temperaturens betydning for den tilbagelagte afstand hos G. pulex
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=5-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
3.2.4 Pesticidblanding
Figur 3.4 viser effekten af glyphosat på den tilbagelagte afstand hos G. pulex ved 3 koncentrationer (100, 1000 og 10000 ng l-1). Der blev ikke observeret forskelle mellem kontrolgruppen og de tre eksponeringsgrupper. Alle fire grupper udviste en tendens til forøget aktivitet i sidste halvdel af eksponeringsperioden. Denne tendens kan ikke tilskrives glyphosat-eksponering og er således et eksempel på, at G. pulex ikke nødvendigvis udviser konstant adfærd i en normal 2-timers observationsperiode.
Figur 3.5 viser den tilbagelagte afstand hos G. pulex eksponeret for lambda-cyhalothrin ved 4 koncentrationer (1, 10, 100 og 1000 ng l-1) alene eller i kombination med glyphosat (1000 ng l-1). Der var ingen forskel på de pyrethroidinducerede adfærdsændringer ved samtidig eksponering for glyphosat, og det observerede adfærdsrespons stemmer overens med det tidligere fundne (Figur 3.1 og 3.2).
Figur 3.4. Effekt af glyphosat på tilbagelagt afstand hos Gammarus pulex
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 30 min, og adfærden i glyphosat-kontamineret vand dernæst fulgt i yderligere 90 min. (Gennemsnit ± SEM, n=7-8)
Figur 3.5. Effekt af lambda-cyhalothrin i kombination med glyphosat på tilbagelagt afstand hos Gammarus pulex
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 30 min, og adfærden i kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: eksponeret for lambda-cyhalothrin i angivne koncentrationer; lukkede symboler: eksponeret for lambda-cyhalothrin i angivne koncentrationer i kombination med glyphosat i en koncentration på 1000 ng/l. (Gennemsnit ± SEM, n=6-8)
3.2.5 Koncentrations-respons-forsøg med udvalgte arter
Figur 3.6-3.11 viser effekten af lambda-cyhalothrin på den tilbagelagte afstand hos 6 arter af ferskvandsinvertebrater eksponeret for 5 lambda-cyhalothrin-koncentrationer ved 10±1 °C.
Adfærdsændringerne svarer hos G. pulex (Figur 3.6) til de tidligere observerede. Ved 1 ng l-1 ses ingen ændringer, mens der ved 10 ng l-1 observeres en markant forøget aktivitet, der opretholdes i resten af eksponeringsperioden. Ved 100 ng l-1 efterfølges hyperaktiviteten af et gradvist fald i den tilbagelagte afstand, og ved 1000 ng l-1 observeres immobilisering efter ca. 55-60 minutters eksponering. Ved 10000 ng l-1 ses hyperaktivitet ganske kortvarigt i det første 5-minuttersinterval efter påbegyndt eksponering, mens immobiliseringen indtræffer efter ca. 40-45 minutter.
A. aquaticus reagerer umiddelbart som G. pulex ved lambda-cyhalothrin-eksponering, og effektkoncentrationerne er sammenlignelige (Figur 3.7). Ved 1 ng l-1 ses ingen ændringer, mens 10 ng l-1 forårsager hyperaktivitet, der opretholdes i resten af eksponeringsperioden. Ved 100 ng l-1 udvises hyperaktivitet kortvarigt, hvilket efterfølges af et markant fald i den tilbagelagte afstand. Ved 1000 ng l-1 observeres immobilisering efter ca. 35 minutters eksponering. Ved 10000 ng l-1 registreres ikke hyperaktivitet, da A. aquaticus ikke kan opretholde et forhøjet aktivitetsniveau i hele det første 5-minuttersinterval efter påbegyndt eksponering, og immobilisering observeres efter kun 15 minutters eksponering.
Figur 3.8 viser, at L. nigra er den mest følsomme af de testede arter. Mens der ingen effekt er ved 0,1 ng l-1, observeres allerede efter ca. 40 minutters eksponering ved 1 ng l-1 en markant reduktion i tilbagelagt afstand til et lavt aktivitetsniveau, der opretholdes eksponeringsperioden ud. Ved højere koncentrationer registreres immobilisering ca. 75, 60 og 60 minutter efter påbegyndt eksponering ved henholdsvis 10, 100 og 1000 ng l-1. L. nigra udviser tilsyneladende ikke hyperaktivitet som et tidligt tegn på pyrethroideksponering og adskiller sig derfor fra de andre arter, hvor lambda-cyhalothrin inducerer adfærdsændringer, d.v.s. G. pulex, A. aquaticus og H. sulphurea, hvor hyperaktivitet ved lave koncentrationer efterfølges af immobilisering ved højere koncentrationer.
Adfærdsresponset hos H. sulphurea er vist i Figur 3.9. Der observeres ingen adfærdsændringer ved 1 og 10 ng l-1. Ved 100 ng l-1 udvises hyperaktivitet efterfulgt af et gradvist fald i den tilbagelagte afstand. Ved både 1000 og 10000 ng l-1 er hyperaktiviteten af meget kort varighed, men ved 10000 ng l-1 efterfølges dette af immobilisering efter ca. 50-55 minutters eksponering. Videosporingsresultaterne for H. sulphurea adskiller sig fra de øvrige arters, idet kontrolgruppernes aktivitetsniveau fluktuerer meget i løbet af observationsperioden.
Det viste sig vanskeligt at opnå et konstant aktivitetsniveau i kontrolgrupperne hos S. personatum, og et gradvist fald blev typisk registreret (Figur 3.10). For A. fluviatilis vedkommende udviste dyrene et meget lavt aktivitetsniveau med tilbagelagte afstande på mindre end 2 cm/5 min (Figur 3.11). En relativ stor andel (op til 50%) af A. fluviatilis bevægede sig slet ikke og blev udeladt ved videre databehandling. Hos hverken S. personatum eller A. fluviatilis blev der registreret ændringer i adfærd som følge af lambda-cyhalothrin-eksponering på op til 10000 ng l-1.
Figur 3.6. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Gammarus pulex
Baggrundsadfærden for G. pulex i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=6-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
Figur 3.7. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Asellus aquaticus
Baggrundsadfærden for A. aquaticus i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=7-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
Figur 3.8. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Leuctra nigra
Baggrundsadfærden for L. nigra i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=7-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
Figur 3.9. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Heptagenia sulphurea
Baggrundsadfærden for H. sulphurea i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr (gennemsnit ± SEM, n=6-8). Asterisk angiver signifikant forskel mellem de to grupper under eksponering.
Figur 3.10. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Sericostoma personatum
Baggrundsadfærden for S. personatum i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr. (Gennemsnit ± SEM, n=5-8)
Figur 3.11. Effekt af lambda-cyhalothrin på tilbagelagt afstand hos Ancylus fluviatilis
Baggrundsadfærden for A. fluviatilis i ukontamineret vand blev registreret i 30 min, og adfærden i lambda-cyhalothrin-kontamineret vand dernæst fulgt i yderligere 90 min. Åbne symboler: kontroldyr; lukkede symboler: eksponerede dyr. (Gennemsnit ± SEM, n=4-8)
3.2.6 Restitueringsforsøg
Figur 3.12 viser overlevelsen, mens Figur 3.13 viser andelen af immobiliserede blandt de overlevende, hos G. pulex og A. aquaticus i en 21-dages restitueringsperiode efter en pulseksponering (1½ eller 4½ time) for lambda-cyhalothrin (10, 100 eller 1000 ng l-1). Figur 3.14 viser den tilbagelagte afstand hos de aktive individer ved afslutningen af restitueringsperioden.
For G. pulex resulterer eksponering for 1000 ng l-1 i såvel 1½ som 4½ time i immobilisering og mortalitet hos langt de fleste individer inden for den første uge efter eksponeringens ophør, og ingen individer overlever til restitueringsperiodens afslutning. Ved 100 ng l-1 ses forøget dødelighed under restitueringsperioden efter en 4½-times puls, mens immobiliseringen er forbigående og varer 1-3 dage. Der er ingen effekter på dødelighed og immobilisering ved 10 ng l-1 hos G. pulex.
A. aquaticus immobiliseres ved lavere eksponeringskoncentrationer end G. pulex; 13% og 63% af A. aquaticus er immobiliserede umiddelbart efter eksponering for 10 ng l-1 i henholdsvis 1½ og 4½ time. Efter endt eksponering aftager andelen af immobiliserede blandt de overlevende individer, og der observeres markant lavere dødelighed efter en pulseksponering for 1000 ng l-1 i såvel 1½ som 4½ time end hos G. pulex. Ved restitueringsperiodens udløb, d.v.s. efter 3 uger i ukontamineret vand er der for begge arter ingen immobiliserede individer blandt de overlevende, pånær for A. aquaticus eksponeret for 1000 ng l-1. Her er ca. 50% af de overlevende efter en 4½-times puls fortsat ude af stand til at bevæge sig, mens dette kun gælder for ca. 10% pulseksponeret i 1½ time. Videosporingen af aktive individer ved restitueringsperiodens udløb viser, at der for begge arter gælder, at der hverken var forskel i aktivitetsniveauet mellem eksponeringsgrupperne eller i forhold til aktitetsniveauet af nyligt indsamlede, ueksponerede dyr.
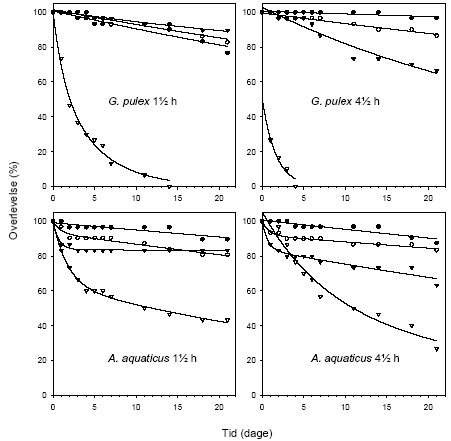
Figur 3.12. Overlevelsen (%) hos A. aquaticus og G. pulex i en 21-dages restitueringsperiode efter en pulseksponering for lambda-cyhalothrin
lukkede cirkler = kontrolgruppe, åbne cirkler = 10 ng l-1, lukkede trekanter = 100 ng l-1, åbne trekanter = 1000 ng l-1. (n=30)
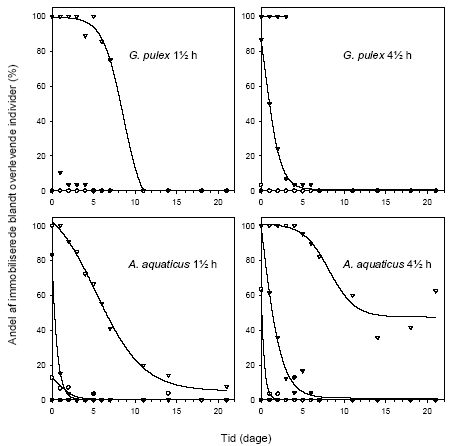
Figur 3.13. Andelen (%) af immobiliserede individer blandt overlevende A. aquaticus og G. pulex i en 21-dages restitueringsperiode efter en pulseksponering for lambda-cyhalothrin
lukkede cirkler = kontrolgruppe, åbne cirkler = 10 ng l-1, lukkede trekanter = 100 ng l-1, åbne trekanter = 1000 ng l-1. (n=30)
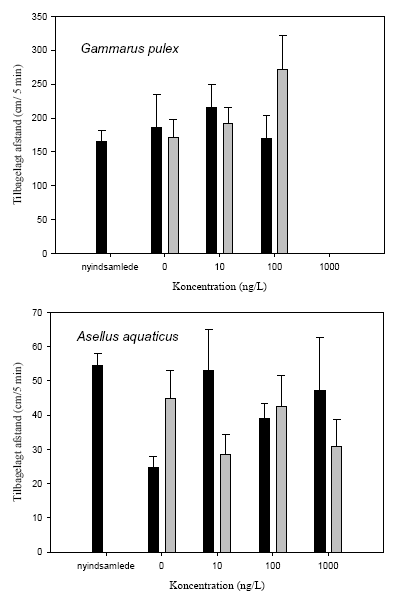
Figur 3.14. Tilbagelagt afstand hos Gammarus pulex og Asellus aquaticus efter 21 dages restituering efter pulseksponering for lambda-cyhalothrin
Adfærden hos de aktive individer blev registreret i ukontamineret vand 60 min, og den gennemsnitlige tilbagelagte afstand er præsenteret. Adfærden hos nyligt indsamlede, ueksponerede dyr er præsenteret til sammenligning. Sorte søjler: 4½-times puls, grå søjler: 1½-times puls. (Gennemsnit ± SEM, n=13-16, dog A. aquaticus 1000 ng l-1: 4½-times puls: n=2, 1½-times puls: n=8)
3.2.7 Rovdyr-byttedyrs-interaktion
Figur 3.15 viser repræsentative eksempler på rovdyr-byttedyrs-interaktioner mellem G. pulex (rovdyr) og L. nigra (byttedyr) under samtidig eksponering for lambda-cyhalothrin. Tilbagelagte afstande for G. pulex og L. nigra samt afstanden mellem dyrene er vist, idet kun de 60 minutters interaktion er medtaget. Figuren viser enkelteksempler fra kontrolgruppen, og grupper eksponeret for 1, 10 eller 100 ng/l. Endvidere vises et eksempel fra gruppen, hvor kun L. nigra blev pre-eksponeret for 100 ng/l i 30 min, inden de 60 minutters interaktion i ukontamineret vand.
I eksemplerne fra kontrolgruppen, gruppen eksponeret for 1 ng/l samt gruppen, hvor kun L. nigra blev pre-eksponeret, observeres, at L. nigra fanges inden for få minutter. G. pulex tilbringer derefter ca. 35-40 min med at spise byttet (der indtages fuldstændigt), hvorefter aktivitetsniveauet igen stiger. Ved 10 ng/l var der generelt stor variation i rovdyradfærden hos G. pulex, der udviser et markant forhøjet aktivitetsniveau. Eksemplet for 10 ng/l viser en interaktion, hvor L. nigra først nedlægges efter 30 min. I dette tilfælde spiser G. pulex kun halvdelen af byttet i løbet af ca. 10 min, og genoptager herefter det forhøjede aktivitetsniveau. Ved 100 ng/l er G. pulex hyperaktiv, og byttedyret L. nigra ignoreres fuldstændigt, på trods af gentagne møder.
Figur 3.16 opsummerer overlevelsestiden for L. nigra for et større antal interaktioner. I kontrolgruppen nedlægger G. pulex størstedelen af L. nigra inden for få minutter efter interaktionens påbegyndelse, og næsten alle byttedyr er døde inden interaktionsperiodens udløb. Det samme gør sig gældende ved 1 ng l-1, og for gruppen hvor L. nigra eksponeres separat for 100 ng l-1 i en halv time inden interaktionen. Ved 10 ng l-1 sker der en markant forøgelse af overlevelsen hos L. nigra, og ca. 50% overlever interaktionen med G. pulex. Ved 100 ng l-1 overlever alle L. nigra, og G. pulex udviser slet ingen rovdyrsadfærd. Regressionsanalysen viser, at selv om overlevelsen af L. nigra ved interaktionsperiodens udløb således først påvirkes ved 10 ng l-1, øges tiden til 50% nedlagte fra 2 min i kontrolgruppen til 4 min ved såvel 1 ng l-1 som i gruppen, hvor kun L. nigra blev eksponeret for 100 ng l-1. Ved 10 ng l-1 nås 50% nedlagte lige netop ikke ifølge regressionsanalysen.
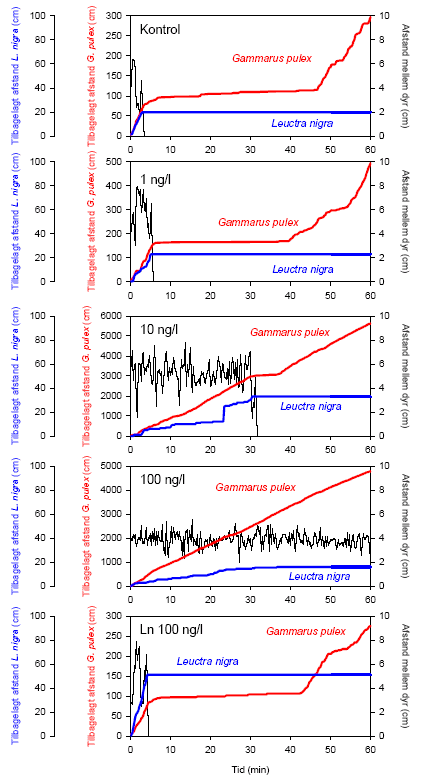
Figur 3.15. Eksempler på rovdyr-byttedyrs-interaktioner mellem L. nigra og G. pulex under samtidig eksponering for lambda-cyhalothrin
Tilbagelagte afstande for G. pulex (rovdyr) og L. nigra (byttedyr) samt afstand mellem dyrene er vist. Dyrene blev indledningsvist eksponeret i 30 minutter, mens de blev holdt adskilt. Herefter blev adskillelsen fjernet, og interaktionen mellem G. pulex og L. nigra blev fulgt i yderligere 60 minutter under fortsat eksponering. Kun de 60 minutters interaktion er vist. Figuren viser enkelt-eksempler fra kontrolgruppen (n=24), 1 ng/l (n=25), 10 ng/l (n=18) og 100 ng/l (n=16). Endeligt vises et eksempel fra gruppen, hvor kun L. nigra blev pre-eksponeret for 100 ng/l i 30 min, inden 60 minutters interaktion i ukontamineret vand (n=9)
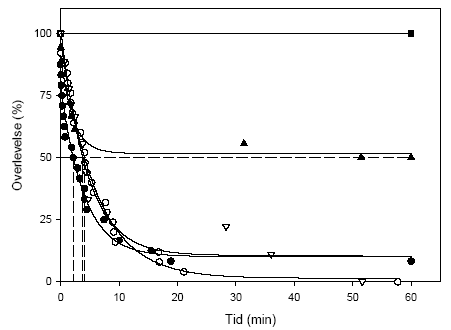
Figur 3.16. Overlevelsestid for L. nigra under prædation fra G. pulex ved samtidig eksponering for lambda-cyhalothrin
Dyrene blev indledningsvist eksponeret i 30 minutter, mens de blev holdt adskilt. Herefter blev adskillelsen fjernet, og interaktionen mellem G. pulex og L. nigra blev fulgt i yderligere 60 minutter under fortsat eksponering. Overlevelsen af L. nigra under de 60 minutters interaktion er vist. Lukkede cirkler = kontrolgruppe (n=24), åbne cirkler = 1 ng/l (n=25), lukkede trekanter = 10 ng/l (n=18), lukkede firkanter = 100 ng/l (n=16), åbne trekanter = kun 30 minutters eksponering af L. nigra for 100 ng/l efterfulgt af overførsel til G. pulex i ukontamineret vand (n=9)
3.3 Drift i strømrender
3.3.1 Koncentrations-respons-forsøg med udvalgte arter
3.3.1.1 Driftrespons
Allerede ved en lambda-cyhalothrin koncentration på 1 ng l-1 ses en tendens til forøget drifttab (p>0.05) for G. pulex efter 90 minutter (Figur 3.17). Ved 10 ng l-1 sker der en mindre forøgelse i driften indenfor de første 30 minutter af eksponeringen, hvorefter drifttabet forøges eksponentielt. Efter 90 minutter er alle G. pulex driftet ud af renderne ved 10 ng l-1, mens der ved forsøgets afslutning (150 min) var ca. 60% af individerne tilbage i renderne behandlet med 1 ng l-1 lambda-cyhalothrin. Driften af G. pulex er signifikant påvirket af behandlingen (p<0,0001), af prøvetagningstidspunktet (p<0,0001) og interaktionen mellem behandling og tidspunkt (p<0,0001). Drifttabet var signifikant højere for både 1 og 10 ng l-1 sammenlignet med kontrol (p<0,0001) og for 10 sammenlignet med 1 ng l-1 (p<0,0001). Ved eksponering med 100 og 1000 ng l-1 lambda-cyhalothrin sker der et meget kraftigt drifttab inden for for de første 30 min, mest markant for 1000 ng l-1 hvor alle G. pulex drifter ud af renderne (Figur 3.18). Ved 100 ng l-1 er de sidste G. pulex først driftet ud efter 120 minutter, hvilket er langsommere end for 10 ng l-1 (Figur 3.17). Denne forskel kan formentlig tilskrives variation i forsøgsdyrenes tilstand i de enkelte eksperimenter og ikke reelle forskelle i respons mellem de to koncentrationer. Som for de to lavere koncentrationer ses en signifikant effekt af behandlingen (p<0,0001), af prøvetagnings-tidspunktet (p<0,0001) og en signifikant interaktion mellem behandling og tidspunkt (p<0,0001). Drifttabet var signifikant højere for både 100 og 1000 ng l-1 sammenlignet med kontrol (p<0,0001) mens der ikke var signifikant forskel for 1000 sammenlignet med 100 ng l-1 (p>0,05).
L. nigra udviser et signifikant drifttab ved eksponering med 1 og 10 ng l-1 lambda-cyhalothrin sammenlignet med kontrol (Figur 3.19). Faldet er størst for 10 ng l-1 men i modsætning til G. pulex udviser L. nigra et lineært forløb i drifttabet og først efter 150 minutter er alle L. nigra driftet ud af renderne behandlet med 10 ng l-1 lambda-cyhalothrin. For 1 ng l-1 er der stadig ca. 20% af individerne tilbage efter 150 minutter. Driften af L. nigra er signifikant påvirket af behandlingen (p<0,0001) og prøvetagningstidspunktet (p=0,035), mens der ikke er signifikant interaktion mellem behandling og tidspunkt (p>0,05). Drifttabet var signifikant højere for både 1 og 10 ng l-1 sammenlignet med kontrol (p<0,0001) mens der ikke var signifikant forskel for 10 sammenlignet med 1 ng l-1 (p>0,05).Ved 100 og 1000 ng l-1 lambda-cyhalothrin øges drifttabet eksponentielt og effekten af de to koncentrationer er ikke signifikant forskellige (Figur 3.20). Faldet er dog for begge koncentrationer mindre markant end for G. pulex, hvilket bevirker, at der er individer, om end få, tilbage i render i hele forsøgsperioden. Ved 100 og 1000 ng l-1 er der signifikant effekt af behandlingen (p<0,0001), af prøvetagningstidspunktet (p<0,0001) og en signifikant interaktion mellem behandling og tidspunkt (p<0,0001).
Ved koncentrationer på 1 og 10 ng l-1 lambda-cyhalothrin ses ingen effekt på driften af H. sulphurea (Figur 3.21). Derimod ses der er et signifikant drifttab ved 100 og 1000 ng l-1 lambda-cyhalothrin (Figur 3.22). Dette drifttab er dog ikke så markant som for G. pulex og L. nigra. Ved 100 ng l-1 starter driften svagt efter 60 minutter og er ved 90 minutter signifikant forskelligt fra kontrolrenderne (p=0.0022). Ved forsøgets afslutning er ca. 50% af nymferne driftet ud af renderne. For 1000 ng l-1 er ca. 80% driftet ud efter 150 minutter. For de to høje koncentrationer er der en signifikant effekt af behandling (p<0,0001), af prøvetagningstidspunktet (p<0,0001) og interaktionen mellem behandling og tidspunkt (p<0,0001). Drifttabet var signifikant højere for både 100 og 1000 ng l-1 sammenlignet med kontrol (p<0,0001) og for 1000 sammenlignet med 100 ng l-1 (p<0,0001).
A. fluviatilis udviser ingen drift ved behandling med lambda-cyhalothrin koncentrationer på 100 og 1000 ng l-1 (Figur 3.23). Stort set samme billede ses for S. personatum, hvor kun enkelte individer går i drift i renderne eksponeret med lambda-cyhalothrin koncentrationer på 100 og 1000 ng l-1 (Figur 3.24). Der er derfor ingen signifikante effekter med hensyn til disse to arter, selvom der en tendens til effekt af lambda-cyhalothrin på driften hos S. personatum (p=0,067).
Resultaterne fra den statistiske analyse er vist i Tabel 3.3.
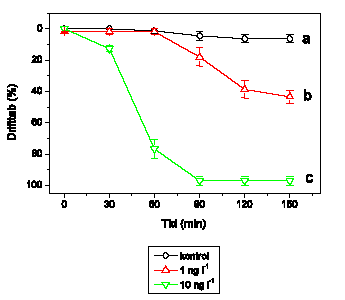
Figur 3.17. Det procentuelle tab ved drift af Gammarus pulex fra render behandlet med 1 og 10 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
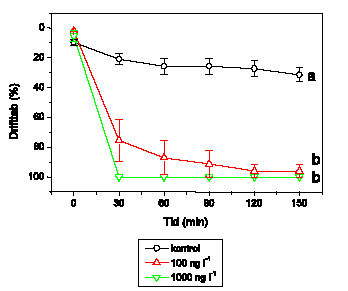
Figur 3.18. Det procentuelle tab ved drift af Gammarus pulex fra render behandlet med 100 og 1000 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
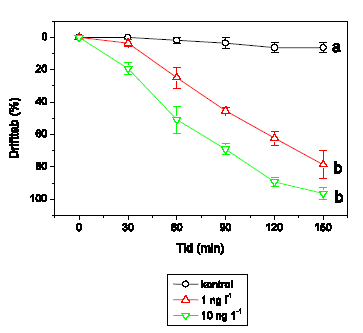
Figur 3.19. Det procentuelle tab ved drift af Leuctra nigra fra render behandlet med 1 og 10 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
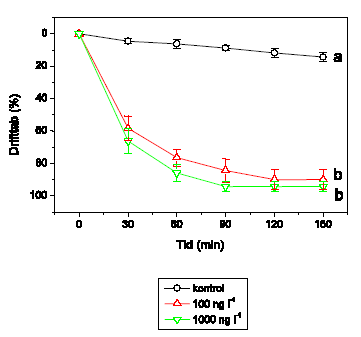
Figur 3.20. Det procentuelle tab ved drift af Leuctra nigra fra render behandlet med 100 og 1000 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
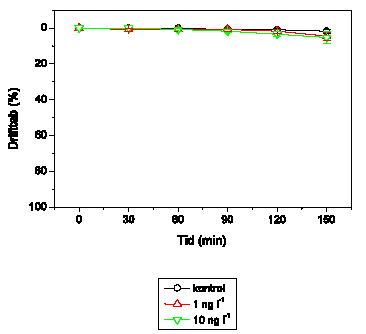
Figur 3.21. Det procentuelle tab ved drift af Heptagenia sulphurea fra render behandlet med 1 og 10 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
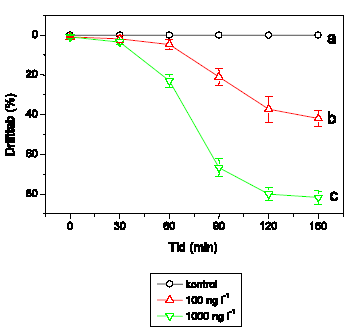
Figur 3.22. Det procentuelle tab ved drift af Heptagenia sulphurea fra render behandlet med 100 og 1000 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
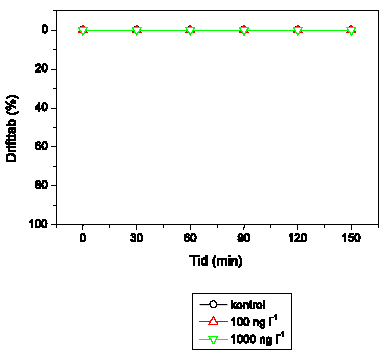
Figur 3.23. Det procentuelle tab ved drift af Ancylus fluviatilis fra render behandlet med 100 og 1000 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
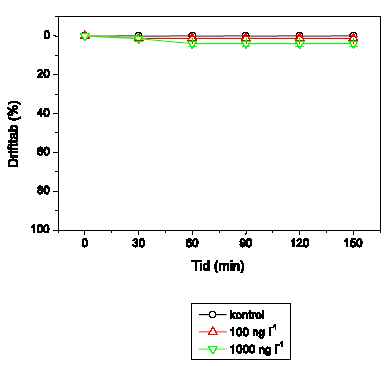
Figur 3.24. Det procentuelle tab ved drift af Sericostoma personatum fra render behandlet med 100 og 1000 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
Tabel 3.3. Test af antallet af individer i drift ved 90 min, 150 min og mellem 90 og 150 min
| Art | Tid | F-test | Parvise tests |
| Gammarus pulex (1-10 ng l-1) |
Ved 90 min | F2,9=52,93, P<0,0001 | 0 - 10: t=-0,82 P=0,4335 0 - 100: t=-9,29 P<0,0001 10 – 100: t=-8,47 P<0,0001 |
| Ved 150 min | F2,9=44,41, P<0,0001 | 0 - 10: t=-2,89 P=0,018 0 - 100: t=-9,21 P<0,0001 10 – 100: t=-6,33 P=0,0001 |
|
| Mellem 90 og 150 min | F2,9=22,29, P=0,0003 | 10: t=-6,67 P<0,0001 0 - 100: t=-3,64 P=0,0054 10 – 100: t=3,03 P=0,0142 |
|
| Gammarus pulex (100-1000 ng l-1) |
Ved 90 min | F2,9=25,12, P<0,0001 | 0 - 10: t=-6,69 P<0,0001 0 - 100: t=-5,37 P=0,004 10 – 100: t=1,32 P=0,2202 |
| Ved 150 min | F2,9=26,59, P=0,0002 | 0 - 10: t=-7,05 P<0,0001 0 - 100: t=-5,15 P=0,0006 10 – 100: t=1,89 P=0,0909 |
|
| Mellem 90 og 150 min | F2,9=0,98, P=0,4125 | 0 - 10: t=-0.78 P=0,4579 0 - 100: t=0,62 P=0,5503 10 – 100: t=1,40 P=0,1962 |
|
| Leuctra nigra (1-10 ng l-1) |
Ved 90 min | F2,9=45,92, P<0,0001 | 0 - 10: t=-5,34 P=0,0005 0 - 100: t=-9,56 P<0,0001 10 – 100: t=-4,23 P=0,0022 |
| Ved 150 min | F2,9=34,58, P<0,0001 | 0 - 10: t=-5,64 P=0,0003 0 - 100: t=-8,11 P<0,0001 10 – 100: t=-2,48 P=0,0353 |
|
| Mellem 90 og 150 min | F2,9=5,53, P=0,0271 | 0 - 10: t=-2,88 P=0,0181 0 - 100: t=-2,88 P=0,0181 10 – 100: t=0 P=1 |
|
| Leuctra nigra (100-1000 ng l-1) |
Ved 90 min | F2,9=54,81, P<0,0001 | 0 - 10: t=-8,57 P<0,0001 0 - 100: t=-9,49 P<0,0001 10 – 100: t=-0,92 P=0,3823 |
| Ved 150 min | F2,9=55,43, P<0,0001 | 0 - 10: t=-8,95 P<0,0001 0 - 100: t=-9,27 P<0,0001 10 – 100: t=-0,32 P=0,7564 |
|
| Mellem 90 og 150 min | F2,9=1,33, P=0,3111 | 0 - 10: t=-1,41 P=0,0,1909 0 - 100: t=0 P=1 10 – 100: t=1,41 P=0,1909 |
|
| Heptagenia sulphurea (1-10 ng l-1) |
Ved 90 min | F2,9=0,30, P=0,7479 | 0 - 10: t=0 P=1 0 - 100: t=-0,67 P=0,5192 10 – 100: t=-0,67 P=0,5192 |
| Ved 150 min | F2,9=0,66, P=0,5397 | 0 - 10: t=0,78 P=0,4548 0 - 100: t=1,95 P=0,0826 10 – 100: t=2,34 P=0,0438 |
|
| Mellem 90 og 150 min | F2,9=0,30, P=0,7509 | 0 - 10: t=-0,50 P=0,6267 0 - 100: t=-0,76 P=0,4694 10 – 100: t=-0,25 P=0,8069 |
|
| Heptagenia sulphurea (100-1000 ng l-1) |
Ved 90 min | F2,9=122,48, P<0,0001 | 0 - 10: t=-4,25 P=0,0022 0 - 100: t=-15,17 P<0,0001 10 – 100: t=-10,92 P<0,0001 |
| Ved 150 min | F2,9=544,89, P<0,0001 | 0 - 10: t=-15,42 P<0,0001 0 - 100: t=-32,99 P<0,0001 10 – 100: t=-17,57 P<0,0001 |
|
| Mellem 90 og 150 min | F2,9=164,37, P<0,0001 | 0 - 10: t=-10,51 P<0,0001 0 - 100: t=-18,05 P<0,0001 10 – 100: t=-7,54 P<0,0001 |
|
| Sericostoma personatum (100-1000 ng l-1) |
Ved 90 min | F2,9=3,50, P=0,0751 | 0 - 10: t=-0,87 P=0,4090 0 - 100: t=-2,60 P=0,0288 10 – 100: t=-1,73 P=0,1173 |
| Ved 150 min | F2,9=3,50, P=0,0751 | 0 - 10: t=-0,87 P=0,4090 0 - 100: t=-2,60 P=0,0288 10 – 100: t=-1,73 P=0,1173 |
|
| Mellem 90 og 150 min | Kan ikke testes | Kan ikke testes |
t-tests har 9 frihedsgrader.
3.3.1.2 Tilstand af organismer i drift
I de eksponerede render var antallet af aktive G. pulex fanget i drift markant lavere end i kontrolrenderne (Figs. 3.25 og 3.26). Denne effekt var signifikant for 1 og 10 ng l-1 (p<0,0001) men ikke for de to høje koncentrationer (p>0,05). Andelen af aktive G. pulex var signifikant lavere for 10 ng l-1 sammenlignet med kontrol (p<0,0001) og for 1 sammenlignet med 10 ng l-1 (p<0,0001). Samtidig var der en stigning i antallet af individer, der udviste en påvirket adfærd med nedsat mobilitet med stigende koncentration. Denne effekt var signifikant for både de lave (p<0,0001) og høje koncentrationer (p<0,0001). Ved både 100 og 1000 ng l-1 var der flere påvirkede end upåvirkede individer af G. pulex. Parvise sammenligninger viste, at andelen af hæmmede individer var signifikant højere for både 1, 10, 100 og 1000 ng l-1 sammenlignet med kontrol (hhv. p=0,0013, p=0,0005, p=0,0011og p=0,0017). Antallet af døde G. pulex var derimod ikke signifikant forskellige. Der var en signifikant effekt af tidspunkt og G. pulex tilstand (data ikke vist): I løbet af forsøgsperioden udviste et stigende antal af individerne fanget i drift i de behandlede render nedsat aktivitet og hæmmet adfærd. Dette var signifikant for både de lave (p=0,0004) og høje koncentrationer (p<0,0001) og begge tilfælde var der en signifikant interaktion mellem tid og koncentration (p<0,0001).
Andelen af L. nigra der var aktive i render behandlet med lambda-cyhalothrin var signifikant lavere sammenlignet med kontrol for både 1 og 10 ng l-1 (p<0,0001) og 100 og 1000 ng l-1 koncentrationer (p<0,0001). Parvis sammenligninger viste at andelen af hæmmede individer var signifikant højere for både 1, 10, 100 og 1000 ng l-1 sammenlignet med kontrol (p<0,0001). Udover denne effekt var der ingen L. nigra, som blev vurderet til have påvirket adfærd (Figs. 3.27 og 3.28), og der var ingen signifikant effekt af behandlingen på dødeligheden (p>0,05). Kun ved de høje koncentrationer (100 og 1000 ng l-1) faldt andelen af aktive L. nigra signifikant over tid (p<0,0001).
Ingen H. sulphurea blev vurderet til være hæmmede ved behandlingen med 1 og 10 ng l-1 (Figur 3.29). Der var en tendens til flere døde i de behandlede render, men dette var ikke signifikant (p>0,05). Ved eksponering med 100 og 1000 ng l-1 lambda-cyhalothrin ses derimod et stort antal påvirkede H. sulphurea og en øget dødelighed (Figur 3.30). Både andelen af H. sulphurea som var hæmmet og andelen af døde individer var signifikant højere i de behandlede render sammenlignet med kontrol (hhv. p<0,0001 og p=0,0019). Der var signifikant forskel 1000 ng l-1 sammenlignet med kontrol (p=0,0018) og for 1000 sammenlignet med 100 ng l-1 (p<0,0001). Både antallet af påvirkede nymfer og døde individer i drift steg signifikant i løbet af forsøgsperioden (hhv. p<0,0001 og p=0,0025), og der var en signifikant interaktion mellem tid og koncentration (hhv. p<0,0001 og p=0.003).
Ingen A. fluviatilis blev fundet i drift og alle individer, der blev indsamlet i renderne efter forsøget, blev vurderet til at have normal adfærd, og der var ingen døde (Figur 3.31).
Der var et stigende antal S. personatum, der udviste påvirket adfærd med stigende koncentration, og ved 1000 ng l-1 var langt hovedparten af individerne påvirket (Figur 3.32).
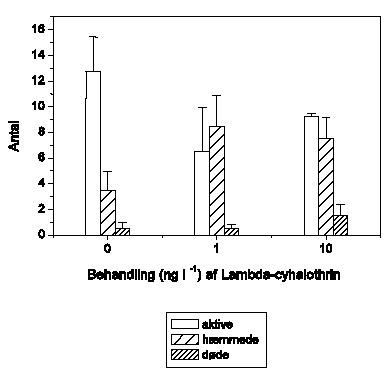
Figur 3.25. Tilstanden af Gammarus pulex fanget i drift/indsamlet efter forsøget ved behandling med 1 og 10 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
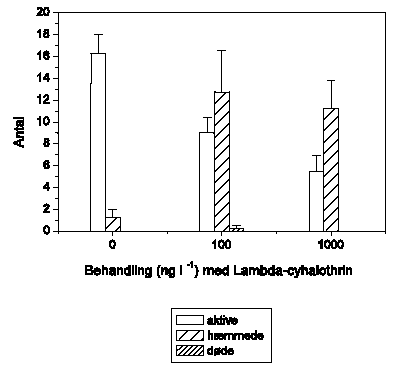
Figur 3.26. Tilstanden af Gammarus pulex fanget i drift/indsamlet efter forsøget ved behandling med 100 og 1000 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
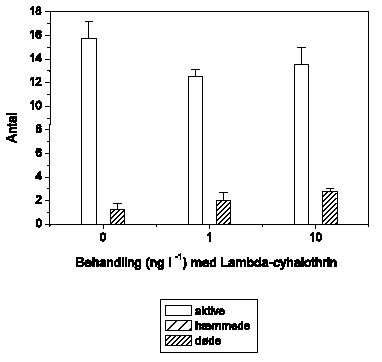
Figur 3.27. Tilstanden af Leuctra nigra fanget i drift/indsamlet efter forsøget ved behandling med 1 og 10 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
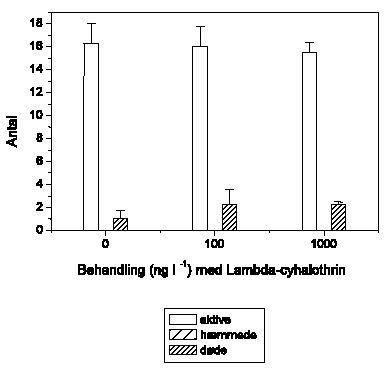
Figur 3.28. Tilstanden af Leuctra nigra fanget i drift/indsamlet efter forsøget ved behandling med 100 og 1000 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
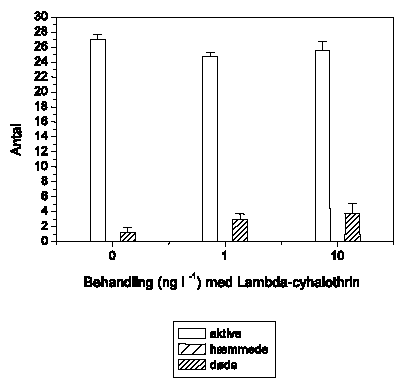
Figur 3.29. Tilstanden af Heptagenia sulphurea fanget i drift/indsamlet efter forsøget ved behandling med 1 og 10 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
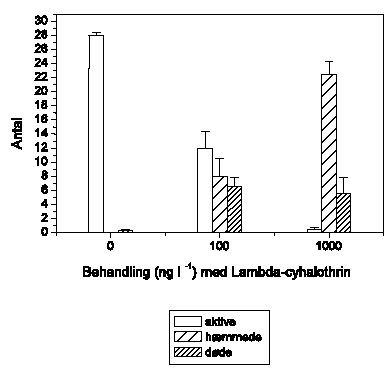
Figur 3.30. Tilstanden af Heptagenia sulphurea fanget i drift/indsamlet efter forsøget ved behandling med 100 og 1000 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
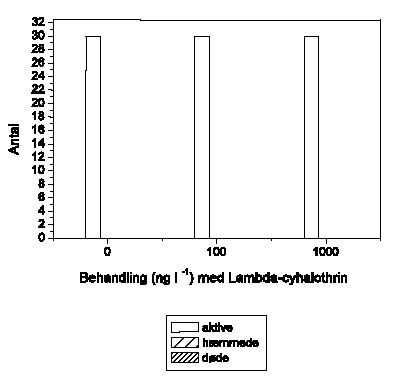
Figur 3.31. Tilstanden af Ancylus fluviatilis fanget i drift/indsamlet efter forsøget ved behandling med 100 og 1000 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
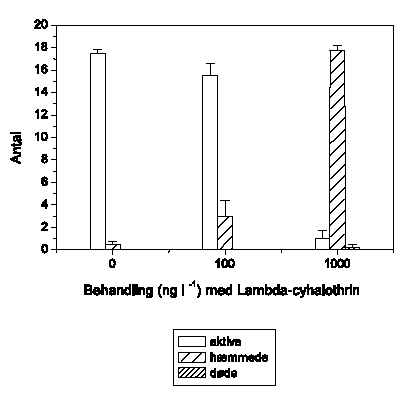
Figur 3.32. Tilstanden af Sericostoma personatum fanget i drift/indsamlet efter forsøget ved behandling med 100 og 1000 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
Tilstanden af de undersøgte invertebrater er angivet i detaljer i Tabel 3.4-3.6.
Tabel 3.4. Tilstanden af de undersøgte invertebrater efter 90 min (3 drift-prøvetagninger af 30 min). Værdierne er angivet som et gennemsnit af 4 replikater for hver behandling/kontrol (antal per 30 min±SEM). A=aktive; H=hæmmede; D=døde.
| Lambda-cyhalothrin-koncentration (ng l-1) | ||||||
| 0 | 1 | 10 | 100 | 1000 | ||
| Gammarus pulex | A | 0,25 ±0,18 0,92±0,36 |
0,083±0,083 | 3,08±0,84 | 2,83±1,25 | 1,58±0,81 |
| H | 0 0,25±0,18 |
0,58±0,31 | 2,50±0,73 | 3,50±1,41 | 3,75±1,72 | |
| D | 0 0 |
0,083±0,083 | 0,33±0,19 | 0,08±0,08 | 0 | |
| Leuctra nigra | A | 0,17±0,11 0,5±0,15 |
2,17±0,52 | 3,75±0,59 | 5,17±1,34 | 5,67±1,47 |
| H | 0 0 |
0 | 0 | 0 | 0 | |
| D | 0,17±0,11 0,08±0,08 |
0,42±0,15 | 0,58±0,19 | 0,08±0,08 | 0,50±0,23 | |
| Heptagenia sulphurea | A | 0,08±0,08 0 |
0,08±0,08 | 0,17±0,11 | 1,75±0,58 | 6,25±1,55 |
| H | 0 0 |
0 | 0 | 0,67±0,36 | 4,75±1,27 | |
| D | 0,17±0,11 0 |
0,08±0,08 | 0,17±0,11 | 0,92±0,38 | 1,42±0,50 | |
| Sericostoma personatum | A | 0 | - | - | 0,08±0,08 | 0,25±0,13 |
| H | 0 | - | - | 0 | 0 | |
| D | 0 | - | - | 0 | 0,08±0,08 | |
| Ancylus fluviatilis | A | 0 | - | - | 0 | 0 |
| H | 0 | - | - | 0 | 0 | |
| D | 0 | - | - | 0 | 0 | |
Tabel 3.5. Tilstanden af de undersøgte invertebrater efter 150 min (5 drift-prøvetagninger af 30 min). Værdierne er angivet som et gennemsnit af 4 replikater for hver behandling/kontrol (antal per 30 min±SEM). A=aktive; H=hæmmede; D=døde.
| Lambda-cyhalothrin-koncentration (ng l-1) | ||||||
| 0 | 1 | 10 | 100 | 1000 | ||
| Gammarus pulex | A | 0,15±0,11 0,60±0,18 |
0,20±0,12 | 1,85±0,60 | 1,70±0,80 | 0,95±0,51 |
| H | 0,05±0,05 0,15±0,11 |
1,00±0,28 | 1,50±0,52 | 2,35±0,92 | 2,25±1,10 | |
| D | 0 0 |
0,05±0,05 | 0,20±0,12 | 0,05±0,05 | 0 | |
| Leuctra nigra | A | 0,20±0,09 0,5±0,11 |
2,25±0,38 | 3,15±0,43 | 3,30±0,95 | 3,40±1,08 |
| H | 0 0 |
0 | 0 | 0 | 0 | |
| D | 0,15±0,08 0,10±0,07 |
0,30±0,11 | 0,45±0,14 | 0,15±0,95 | 0,30±0,15 | |
| Heptagenia sulphurea | A | 0,10±0,07 | 0,25±0,16 | 0,30±0,13 | 2,15±0,47 | 4,60±1,08 |
| H | 0 | 0 | 0 | 0,90±0,33 | 3,65±0,88 | |
| D | 0,10±0,07 | 0,10±0,07 | 0,25±0,10 | 1,15±0,30 | 0,90±0,33 | |
| Sericostoma personatum | A | 0 | - | - | 0,05±0,05 | 0,15±0,08 |
| H | 0 | - | - | 0 | 0 | |
| D | 0 | - | - | 0 | 0,05±0,05 | |
| Ancylus fluviatilis | A | 0 | - | - | 0 | 0 |
| H | 0 | - | - | 0 | 0 | |
| D | 0 | - | - | 0 | 0 | |
Tabel 3.6. Tilstanden af de undersøgte invertebrater ved forsøgets afslutning (invertebrater der var tilbage i renderne). Værdierne er angivet som et gennemsnit af 4 replikater for hver behandling/kontrol (antal±SEM). A=aktive; H=hæmmede; D=døde.
| Lambda-cyhalothrin-koncentration (ng l-1) | ||||||
| 0 | 1 | 10 | 100 | 1000 | ||
| Gammarus pulex | A | 12,00±2,58 11,50±1,50 |
5,25±3,20 | 0 | 0 | 0 |
| H | 3,25±1,25 0.5±0,50 |
3,50±2,18 | 0 | 1,00±1,00 | 0 | |
| D | 0,50±0,50 0 |
0,25±0,25 | 0,50±0,50 | 0 | 0 | |
| Leuctra nigra | A | 15,5±1,32 14,75±1,31 |
2,5±1,15 | 0 | 0,25±0,25 | 0 |
| H | 0 0 |
0 | 0 | 0 | 0 | |
| D | 0,5±0,29 0,5±0,5 |
0,5±0,29 | 0,5±0,5 | 1,50±0,96 | 0,75±0,48 | |
| Heptagenia sulphurea | A | 27,00±0,71 28±0,41 |
24,00±0,82 | 24,75±1,25 | 11,50±2,50 | 0 |
| H | 0 0 |
0 | 0 | 3,50±1,19 | 4,25±1,11 | |
| D | 0,75±0,75 0,25±0,25 |
2,50±0,50 | 2,50±1,04 | 0,50±0,29 | 1,00±0,71 | |
| Sericostoma personatum | A | 17,50±0,29 | - | - | 15,25±1,25 | |
| H | 0,50±0,29 | - | - | 3,00±1,41 | ||
| D | 0 | - | - | 0 | ||
| Ancylus fluviatilis | A | 0 | - | - | 30 | 30 |
| H | 0 | - | - | 0 | 0 | |
| D | 0 | - | - | 0 | 0 | |
3.3.2 Eksponering om natten
Ved driftundersøgelse om natten ses der en umiddelbar stigning i drifttabet for lambda-cyhalothrin-koncentrationer på 10 og 100 ng l-1 (Figur 3.33). Denne stigning er mest markant for 100 ng l-1, hvor 90% af individerne er driftet ud af renderne 30 minutter efter eksponeringens start. I modsætning til driftforsøgene udført om dagen ses der et tydeligt drifttab i kontrolrenderne, hvilket formentlig afspejler G. pulex generelt højere aktivitetsniveau om natten. Responset på behandlingen med lambda-cyhalothrin svarer godt overens med, hvad der blev fundet om dagen, dog med en tendens til at være lidt mere markant om natten (se også Figur 3.17 og 3.18). Imidlertid er effekten af behandling med lambda-cyhalothrin på driften om natten lige præcis ikke signifikant (p=0,0051) på grund af det store drifttab i kontrolrenderne.
Antallet af aktive G. pulex fanget i drift om natten var stort set konstant med stigende koncentration af lambda-cyhalothrin (Figur 3.34). Der var ligeledes stort set ingen forskelle mellem hæmmede og døde G. pulex i de tre grupper og som konsekvens heraf ingen signifikante (p>0,05) effekter på G. pulex tilstand i drifteksperimentet foretaget om natten. Sammenlignet med eksperimenterne foretaget om dagen var der markant færre påvirkede G. pulex om natten.

Figur 3.33. Det procentuelle tab ved drift om natten af Gammarus pulex fra render behandlet med 10 og 100 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrender. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
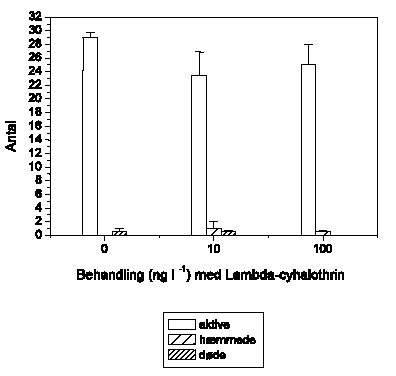
Figur 3.34. Tilstanden af Gammarus pulex fanget i drift/indsamlet efter forsøget om natten ved behandling med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
Resultaterne fra den statistiske analyse er vist i Tabel 3.7, mens tilstanden af G. pulex er angivet i detaljer i Tabel 3.8.
Tabel 3.7. Test af antallet af individer i drift ved 90 min, 150 min og mellem 90 og 150 min
| Art | Tid | F-test | Parvise tests |
| Gammarus pulex - Nat (10-100 ng l-1) |
Ved 90 min | F2,9=2,67, P=0,1230 | 0 - 10: t=-1,78 P=0,1080 0 - 100: t=-2,16 P=0,0588 10 - 100: t=-0,38 P=0,7138 |
| Ved 150 min | F2,9=1,30, P=0,3193 | 0 - 10: t=-1,21 P=0,2567 0 - 100: t=-1,53 P=0,1611 10 - 100: t=-0,32 P=0,7593 |
|
| Mellem 90 og 150 min | F2,9=3,44, P=0,0778 | 0 - 10: t=2,21 P=0,0546 0 - 100: t=2,33 P=0,0449 10 - 100: t=0,12 P=0,9076 |
t-tests har 9 frihedsgrader.
Tabel 3.8. Tilstanden af de nateksponerede G. pulex efter 90 min (3 drift-prøvetagninger af 30 min) ), 150 min (5 drift prøvetagninger af 30 min) og ved forsøgets afslutning (invertebrater der var tilbage i renderne). Værdierne er angivet som et gennemsnit af 4 replikater for hver behandling/kontrol (antal per 30 min±SEM). A=aktive; H=hæmmede; D=døde.
| Lambda-cyhalothrin-koncentration (ng l-1) | ||||
| 0 | 10 | 100 | ||
| Gammarus pulex (90 min) |
A | 2,25±0,64 | 6,92±1,79 | 7,50±2,78 |
| H | 0 | 0,33±0,26 | 0,17±0,11 | |
| D | 0 | 0 | 0 | |
| Gammarus pulex (150 min) |
A | 1,90±0,45 | 4,20±1,31 | 4,5±1,84 |
| H | 0 | 0,20±0,16 | 0,1±0,07 | |
| D | 0 | 0,05±0,05 | 0 | |
| Gammarus pulex (ved forsøgets afslutning) |
A | 13,50±2,18 | 0 | 0 |
| H | 0 | 0 | 0 | |
| D | 0,50±0,50 | 0,25±0,25 | 0 | |
3.3.3 Rovdyr-byttedyrs-interaktion
G. pulex udsat for ørredlugt i renderne udviste et respons på behandlingen med lambda-cyhalohtrin-koncentrationer på 10 og 100 ng l-1, der overordnet svarede til det respons, som var fundet ved de samme koncentrationer i eksperimenter uden fiskelugt (Figur 3.35 samt 3.17 og 3.18). Der var derfor ingen umiddelbar indikation af, at fiskelugt påvirkede G. pulex’ driftrespons ved en puls med lambda-cyhalothrin. For 10 ng l-1 skete der kun et lille drifttab indenfor de første 30 minutter, hvorefter det øgedes eksponentielt, og alle G. pulex var stort set driftet ud af renderne efter 90 minutter. For 100 ng l-1 var driften markant allerede efter 30 minutter, og også her var stort set alle individer driftet ud efter 90 minutter. Effekten af behandling med lambda-cyhalothrin på driften hos G. pulex udsat for fiskelugt var signifikant (p<0,0001), og det samme var prøvetagningstidspunktet (p<0,0001) og interaktionen mellem behandling og tidspunkt (p<0,0001). Drifttabet var signifikant højere for både 10 og 100 ng l-1 sammenlignet med kontrol (p<0,0001) men ikke mellem 10 og 100 ng l-1 (p>0,05).
Selvom om det absolutte antal af aktive G. pulex var sammenligneligt mellem grupperne (Figur 3.36), var andelen af aktive individer signifikant lavere for de to behandlede grupper (p<0,0001). Det lavere antal aktive individer skyldtes især en signifikant forøgelse i G. pulex, der udviste en hæmmet adfærd (p=0,0045). Der var ingen signifikante forskelle mellem antallet af døde G. pulex mellem grupperne (p>0,05). Der blev færre aktive og flere hæmmede individer i løbet af forsøget, hvilket var signifikant (hhv. p<0,0001 og p=0,0004), og med hensyn til aktive dyr var der en signifikant interaktion mellem tid og koncentration (p<0,0001).
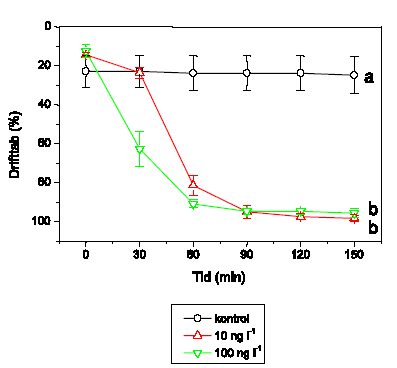
Figur 3.35. Det procentuelle tab ved drift af Gammarus pulex fra render eksponeret for ørredlugt og behandlet med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol. Tid 0 svarer til starttidspunktet for eksponeringen med lambda-cyhalohtrin (varighed 90 min). Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe. Forskellige bogstaver angiver signifikante forskelle mellem behandlingerne.
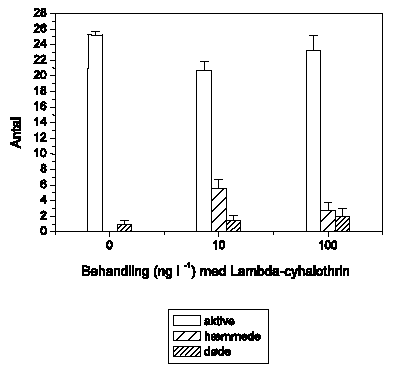
Figur 3.36. Tilstanden af Gammarus pulex fanget i drift/indsamlet efter forsøget i render eksponeret for ørredlugt og behandlet med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol (0). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
Resultaterne fra den statistiske analyse er vist i Tabel 3.9, mens tilstanden af G. pulex er angivet i detaljer i Tabel 3.10.
Tabel 3.9. Test af antallet af individer i drift ved 90 min, 150 min og mellem 90 og 150 min
| Art | Tid | F-test | Parvise tests |
| Gammarus pulex - Fisk (10-100 ng l-1) |
Ved 90 min | F2,9=186,93, P<0,0001 | 0 - 10: t=-16,25 P<0,0001 0 - 100: t=-17,20 P<0,0001 10 - 100: t=-0,94 P=0,3694 |
| Ved 150 min | F2,9=186,99, P<0,0001 | 0 - 10: t=-16,56 P<0,0001 0 - 100: t=-16,93 P<0,0001 10 - 100: t=-0,37 P=0,7184 |
|
| Mellem 90 og 150 min | F2,9=7,66, P=0,0114 | 0 - 10: t=-3,73 P=0,0047 0 - 100: t=-0,83 P=0,4290 10 - 100: t=2,90 P=0,0176 |
t-tests har 9 frihedsgrader.
Tabel 3.10. Tilstanden af G. pulex udsat for ørredlugt efter 90 min (3 drift-prøvetagninger af 30 min) ), 150 min (5 drift prøvetagninger af 30 min) og ved forsøgets afslutning (invertebrater der var tilbage i renderne). Værdierne er angivet som et gennemsnit af 4 replikater for hver behandling/kontrol (antal per 30 min±SEM). A=aktive; H=hæmmede; D=døde.
| Lambda-cyhalothrin-koncentration (ng l-1) | ||||
| 0 | 10 | 100 | ||
| Gammarus pulex (90 min) |
A | 0,08±0,08 | 7,25±1,86 | 7,67±1,87 |
| H | 0 | 1,58±0,32 | 0,92±0,45 | |
| D | 0 | 0,33±0,14 | 0,25±0,13 | |
| Gammarus pulex (150 min) |
A | 0,10±0,07 | 4,55±1,34 | 4,65±1,39 |
| H | 0 | 1,05±0,26 | 0,55±0,29 | |
| D | 0 | 0,25±0,10 | 0,15±0,08 | |
| Gammarus pulex (ved forsøgets afslutning) |
A | 18,75±2,32 | 0 | 0 |
| H | 0 | 0,25±0,25 | 0 | |
| D | 1,00±0,41 | 0,25±0,25 | 1,25±0,63 | |
3.3.4 Flerarts-forsøg
3.3.4.1 Direkte effekter på drift
I flerarts-eksperimentet blev levende dyr fanget i drift genudsat til renderne for at opretholde passende tætheder igennem hele forsøgsperioden. For kontrolrender var tæthederne meget konstante med et lille fald for G. pulex til sidst i perioden (Figur 3.37). Der var således gennemsnitligt 61,3 G. pulex , 44,5 L. nigra og 14,8 H. sulphurea tilbage ved forsøgets afslutning mod hhv. 65, 45 og 15 ved forsøgets start. For A. fluviatilis var antallet konstant på 25 individer i alle 4 replikatrender i hele forsøgsperioden. For renderne behandlet med 10 ng l-1 lambda-cyhalothrin var forløbet i tætheden af de 4 makroinvertebrater under forsøget stort set identisk med kontrolrenderne (Figur 3.38). I disse render var der gennemsnitligt 60,3 G. pulex, 43,0 L. nigra, 25,0 A. fluviatilis og 14,5 H. sulphurea tilbage ved forsøgets afslutning. For renderne behandlet med 100 ng l-1 lambda-cyhalothrin var der et mere markant fald i antallet af individer af G. pulex og L. nigra til sidst i perioden end for de to andre grupper (Figur 3.39). Dette afspejler den større dødelighed i denne behandling i forhold til de øvrige og dermed at færre individer blev genudsat som forsøget skred frem (se senere). Ved afslutning af forsøget var der i renderne behandlet med 100 ng l-1 l følgende gennemsnitlige antal makroinvertebrater tilbage: 57,0 G. pulex, 38,3 L. nigra, 25,0 A. fluviatilis og 13,5 H. sulphurea.
Andelen af G. pulex i drift steg for kontrol og 10 ng l-1 i midten af forsøgsperioden for dernæst at stabilisere sig på et forhøjet niveau i forhold til starten af forsøget (Figur 3.40). Driften var højest i kontrolrenderne, intermediær i 10 ng l-1 behandlingen og lavest i 100 ng l-1 behandlingen. Driften i kontrol renderne var signifikant højere end både driften ved 10 og 100 ng l-1 (p<0,0001), ligesom driften var signifikant højere ved 10 end ved 100 ng l-1 (p<0,0001). Som for G. pulex steg driften overordnet i midten af perioden for L. nigra (Figur 3.41), men i modsætning til G. pulex faldt driften lidt igen for så at stige til sidst i forsøgsperioden. Der var ingen forskel på driftmønstret mellem de tre grupper (kontrol, 10 og 100 ng l-1) og derfor heller ikke nogle signifikante forskelle (p>0,05). Meget få H. sulphurea var generelt i drift og der var intet mønster i driften, hverken overordnet eller mellem grupperne (Figur 3.42) og ingen signifikante forskelle (p>0,05). Ingen A. fluviatilis gik i drift i de 12 render uanset tidspunkt og behandling (data ikke vist).
Resultaterne fra den statistiske analyse er vist i Tabel 3.11.
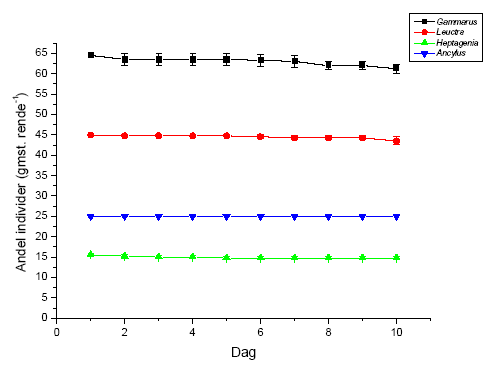
Figur 3.37. Gennemsnitligt (± standard error, n=4) antal individer af de fire undersøgte arter i kontrolrenderne i forsøgsperioden (10 dage). Start-individantallet i renderne var 65 for Gammarus pulex, 45 for Leuctra nigra, 25 for Ancylus fluviatilis og 15 for Heptagenia sulphurea.
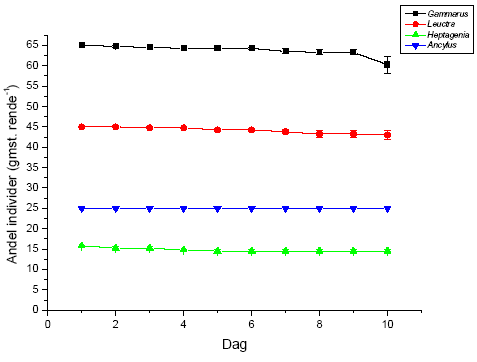
Figur 3.38. Gennemsnitligt (± standard error, n=4) antal individer af de fire undersøgte arter i renderne behandlet med 10 ng l-1 lambda-cyhalothrin i forsøgsperioden (10 dage). Start-individantallet i renderne var 65 for Gammarus pulex, 45 for Leuctra nigra, 25 for Ancylus fluviatilis og 15 for Heptagenia sulphurea.

Figur 3.39. Gennemsnitligt (± standard error, n=4) antal individer af de fire undersøgte arter i renderne behandlet med 100 ng l-1 lambda-cyhalothrin i forsøgsperioden (10 dage). Start-individantallet i renderne var 65 for Gammarus pulex, 45 for Leuctra nigra, 25 for Ancylus fluviatilis og 15 for Heptagenia sulphurea.
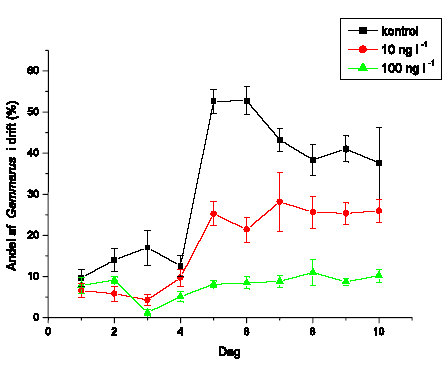
Figur 3.40. Andelen af Gammarus pulex i drift i forsøgsperioden (10 dage) for render behandlet med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.

Figur 3.41. Andelen af Leuctra nigra i drift i forsøgsperioden (10 dage) for render behandlet med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
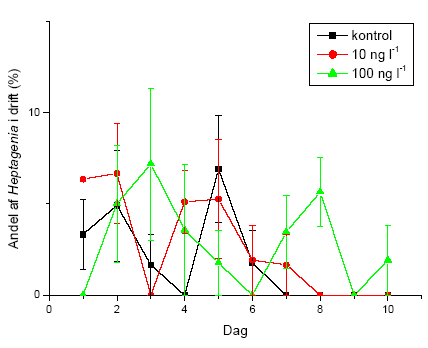
Figur 3.42. Andelen af Heptagenia sulphurea i drift i forsøgsperioden (10 dage) for render behandlet med 10 og 100 ng l-1 lambda-cyhalothrin samt kontrol. Hver behandling og kontrol havde 4 replikater. Figuren viser gennemsnit og standard error for de 4 render i hver gruppe.
3.3.4.2 Direkte effekter på organismernes tilstand
I flerarts-eksperimentet blev makroinvertebraternes tilstand undersøgt dagligt i driftprøverne og ved forsøgsafslutning, da renderne blev tømt for dyr. I det følgende er tilstanden af individer fanget i drift og tilstanden af de individer, der var tilbage i renderne ved forsøgets afslutning, behandlet separat.
For G. pulex faldt andelen af aktive individer i drift markant med behandlingen (Figur 3.43). Der var samtidig en mindre stigning i antallet af hæmmede individer med lambda-cyhalothrin koncentrationen, mens antallet af døde var sammenligneligt mellem de tre grupper (kontrol, 10 og 100 ng l-1). Både andelen af aktive og hæmmede individer var signifikant forskellige mellem grupperne (hhv. p<0,0001 og p=0,0034). Der var en signifikant lavere andel af aktive G. pulex for både ved 10 og 100 ng l-1 (p<0,0001) sammenlignet med kontrol, ligesom andelen var signifikant lavere ved 100 end ved 10 ng l-1 (p<0,0001). Aktiviteten var signifikant faldende over forsøgsperioden i de behandlede render (p<0,0001) og med signifikant interaktion mellem tid og koncentration (p<0,0001) (data ikke vist). Andelen af hæmmede individer var ligeledes signifikant stigende gennem forsøgsperioden (p=0,002).
Andelen af aktive L. nigra i drift faldt ligeledes med behandlingen, mens andelen af hæmmede individer steg (Figur 3.44). Samtidig steg antallet af døde individer især for 100 ng l-1 . Den stigende andel af hæmmede individer var signifikant forskellig mellem grupperne (p=0,0079), og der var signifikant forskel mellem kontrol og 100 ng l-1 (p=0,0025) samt mellem 10 ng l-1 og 100 ng l-1 (p=0,032). Der var over tid signifikant færre aktive L. nigra og samtidig flere hæmmede individer (hhv. p=0,0001 og p=0,0052) og med signifikant interaktion mellem tid og koncentration for aktivitet (p=0,019)(data ikke vist).
Ud fra de relativ få individer af H. sulphurea fundet i drift ses, at færre individer er aktive ved den højeste koncentration, samtidig med at flere er hæmmede eller døde (Figur 3.45). Andelen af aktive nymfer er signifikant forskellig mellem grupperne (p=0,024), ligesom der er en signifikant effekt af prøvetagningstidspunktet (p=0,029).
Ingen A. fluviatilis blev fundet i drift (data ikke vist).
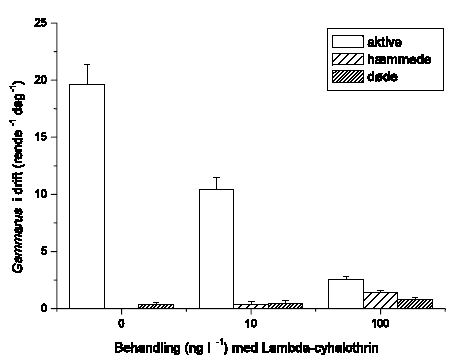
Figur 3.43. Tilstanden af Gammarus pulex fanget i drift i løbet af forsøgsperioden (10 dage). Gennemsnit og standard error er baseret på 10 observationer per rende med 4 replikater for hver gruppe (kontrol, 10 og 100 ng l-1 lambda-cyhalothrin). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde.
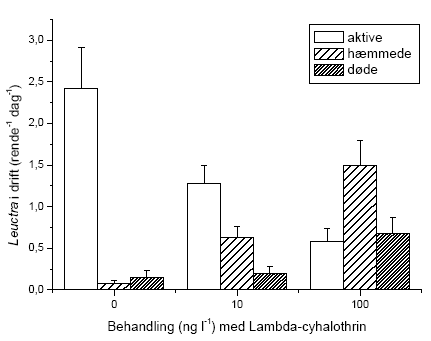
Figur 3.44. Tilstanden af Leuctra nigra fanget i drift i løbet af forsøgsperioden (10 dage). Gennemsnit og standard error er baseret på 10 observationer per rende med 4 replikater for hver gruppe (kontrol, 10 og 100 ng l-1 lambda-cyhalothrin). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde.
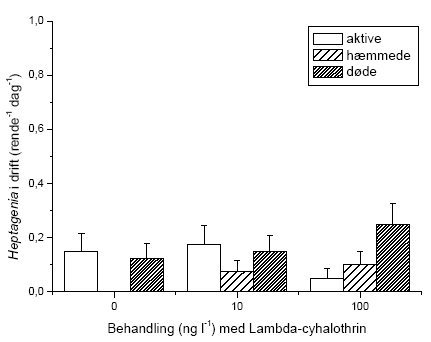
Figur 3.45. Tilstanden af Heptagenia sulphurea fanget i drift i løbet af forsøgsperioden (10 dage). Gennemsnit og standard error er baseret på 10 observationer per rende med 4 replikater for hver gruppe (kontrol, 10 og 100 ng l-1 lambda-cyhalothrin). Alle individer blev visuelt klassificeret til at være normalt aktive (aktive), udvise reduceret mobilitet (hæmmede) eller være døde.
Resultaterne fra den statistiske analyse er vist i Tabel 3.11.
Tabel 3.11. Test antal individer i drift og deres tilstand i flerarts-forsøgene
| Art | Status | F-tests | Parvise tests |
| Total | Drift | F2,9=47,85, P<0,0001 F9,81=33,46, P<0,0001 F18,81=7,88, P<0,0001 |
0 - 10: t=5,90 P=0,0002 0 - 100: t=9,71 P<0,0001 10 - 100: t=7,18 P<0,0001 |
| Påvirkede | F2,9=15,38, P=0,0013 F9,81=5,10, P<0,0001 F18,81=1,24, P=0,2511 |
0 - 10: t=-1,90 P=0,0899 0 - 100: t=-5,46 P=0,0004 10 - 100: t=-3,56 P=0,0061 |
|
| Døde | F2,9=10,28, P=0,0047 F9,81=3,59, P=0,0008 F18,81=0,86, P=0,6305 |
0 - 10: t=-0,69 P=0,5089 0 - 100: t=-4,23 P=0,0022 10 - 100: t=-3,54 P=0,0063 |
|
| Gammarus pulex | Drift | F2,9=140,43, P<0,0001 F9,81=26,44, P<0,0001 F18,81=7,31, P<0,0001 |
0 – 10: t=9,54 P<0,0001 0 – 100: t=16,70 P<0,0001 10 – 100: t=7,16 P<0,0001 |
| Aktive | F2,9=123,26, P<0,0001 F9,81=20,21, P<0,0001 F18,81=7,11, P<0,0001 |
0 – 10: t=25,50 P<0,0001 0 – 100: t=15,68 P<0,0001 10 – 100: t=7,22 P<0,0001 |
|
| Påvirkede | F2,9=11,39, P=0,0034 F9,81=3,26, P=0,0020 F18,81=0,80, P=0,6960 |
0 – 10: t=-1,28 P=0,2312 0 – 100: t=-4,62 P=0,0012 10 – 100: t=-3,34 P=0,0087 |
|
| Døde | F2,9=1,59, P=0,2555 F9,81=2,59, P=0,0113 F18,81=1,14, P=0,3319 |
0 – 10: t=-0,40 P=0,6972 0 – 100: t=-1,71 P=0,1219 10 – 100: t=-1,31 P=0,2240 |
|
| Tilbage | F2,9=1,58, P=0,2573 F9,81=18,45, P<0,0001 F18,81=2,34, P=0,0051 |
0 – 10: t=-0,48 P=0,6420 0 – 100: t=1,24 P=0,2449 10 – 100: t=1,72 P=0,1186 |
|
| Leuctra nigra | Drift | F2,9=0,30, P=0,7513 F9,81=5,73, P<0,0001 F18,81=0,78, P=0,7161 |
0 – 10: t=0,60 P=0,5609 0 – 100: t=-0,11 P=0,9150 10 – 100: t=-0,71 P=0,4936 |
| Aktive | F2,9=2,61, P=0,1278 F9,81=4,42, P=0,0001 F18,81=0,99, P=0,3461 |
0 – 10: t=1,41 P=0,1933 0 – 100: t=2,26 P=0,0500 10 – 100: t=0,86 P=0,4143 |
|
| Påvirkede | F2,9=8,71, P=0,0079 F9,81=2,88, P=0,0052 F18,81=1,04, P=0,4237 |
0 – 10: t=-1,60 P=0,1447 0 – 100: t=-4,14 P=0,0025 10 – 100: t=-2,54 P=0,0317 |
|
| Døde | F2,9=7,24, P=0,0134 F9,81=2,70, P=0,0085 F18,81=1,09, P=0,3797 |
0 – 10: t=-0,33 P=0,7502 0 – 100: t=-3,45 P=0,0073 10 – 100: t=-3,12 P=0,0123 |
|
| Tilbage | F2,9=7,10, P=0,0141 F9,81=14,03, P<0,0001 F18,81=3,27, P=0,0001 |
0 – 10: t=0,59 P=0,5723 0 – 100: t=3,52 P=0,0066 10 – 100: t=2,93 P=0,0168 |
|
| Heptagenia sulphurea | Drift | F2,9=0,82, P=0,4692 F9,81=2,65, P=0,0097 F18,81=1,55, P=0,0955 |
0 - 10: t=-1,11 P=0,2950 0 - 100: t=-1,11 P=0,2950 10 - 100: t=0 P=1 |
| Aktive | F2,9=5,73, P=0,0249 F9,81=2,22, P=0,0287 F18,81=1,15, P=0,3182 |
0 - 10: t=-0,64 P=0,5384 0 - 100: t=2,56 P=0,0308 10 - 100: t=3,20 P=0,0109 |
|
| Påvirkede | Ikke muligt at teste | Ikke muligt at teste | |
| Døde | F2,9=1,47, P=0,2813 F9,81=1,47, P=0,1719 F18,81=1,12, P=0,3506 |
0 - 10: t=-0,32 P=0,7537 0 - 100: t=-1,62 P=0,1402 10 - 100: t=-1,29 P=0,2279 |
|
| Tilbage | F2,9=0,29, P=0,7573 F9,81=15,84, P<0,0001 F18,81=2,04, P=0,0162 |
0 - 10: t=0,20 P=0,8431 0 - 100: t=0,73 P=0,4819 10 - 100: t=0,53 P=0,6091 |
t-tests har 9 frihedsgrader.
3.3.4.3 Indirekte effekter på algebiomasse og omsætningen af organisk stof
Nedbrydningen af det organiske stof i bladpakkerne var mere end dobbelt så høj i kontrolrenderne sammenlignet med de to grupper af behandlede render (10 ng l-1 og 100 ng l-1) (Figur 3.46). Forskellen i nedbrydning var signifikant forskellig mellem grupperne (p=0,017) og den var signifikant højere i kontrolrenderne sammenlignet med både 10 ng l-1 og 100 ng l-1 (hhv. p=0,016 og p=0,035). Der var ingen signifikant forskel mellem de to grupper af behandlede render (p>0,05).
Algebiomassen på sten i renderne steg i løbet af forsøgsperioden for alle grupper, dog klart mest tydeligt for renderne behandlet med 100 ng l-1 lambda-cyhalothrin (Figur 3.47). Der var en signifikant effekt af behandlingen og prøvetagningstidspunktet på algebiomassen (hhv. p=0,033 og p=0,0086), og algebiomassen var signifikant højere i renderne behandlet med 100 ng l-1 end 10 ng l-1 og kontrol (hhv. p=0,035 og p=0,016).
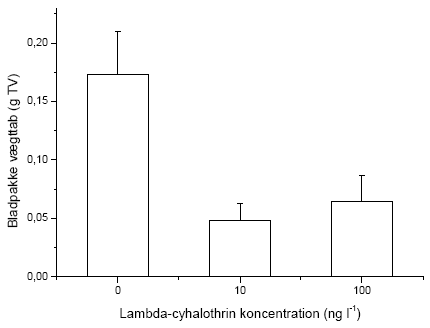
Figur 3.46. Vægttab af bladpakker i løbet af forsøgsperioden (10 dage). Gennemsnit og standard error er baseret på 5 bladpakker i hver rende (n=4, kontrol, 10 og 100 ng l-1 lambda-cyhalothrin).
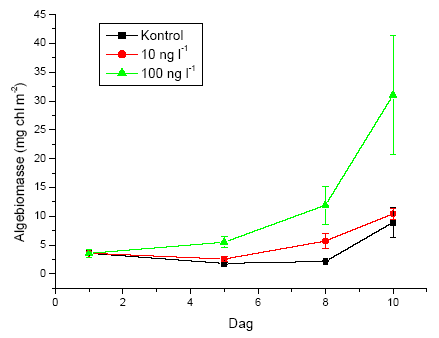
Figur 3.47. Ændringer i algebiomassen i forsøgsperioden (10 dage). Sten til algeanalyser blev indsamlet dag 1, 5, 8 og 10. Hvert punkt repræsentere gennemsnit og standard error for hver gruppe (n=4, kontrol, 10 og 100 ng l-1 lambda-cyhalothrin).
Resultaterne fra den statistiske analyse er vist i Tabel 3.12 og 3.13.
Tabel 3.12. Test af bladpakkernes vægttab efter 10 dage.
| Test | Test-værdi | p-værdi |
| Test for behandlingseffekt | F2,9=6,67 | P=0,0167 |
| 0 mod 10 ng l-1 | t9=3,36 | P=0,0084 |
| 0 mod 100 ng l-1 | t9=2,92 | P=0,0169 |
| 10 mod 100 ng l-1 | t9=-0,43 | P=0,6757 |
Tabel 3.13. Test af udviklingen i algebiomassen
| Test | Test-værdi | p-værdi |
| Interaktionsled | F4,9=2,40 | P=0,1271 |
| Behandlingseffekt | F2,9=7,48 | P=0,0122 |
| Dagseffekt | F2,9=5,85 | P=0,0236 |
| 0 mod 10 ng l-1 | t9=-1,20 | P=0,2613 |
| 0 mod 100 ng l-1 | t9=-3,78 | P=0,0043 |
| 10 mod 100 ng l-1 | t9=-2,59 | P=0,0294 |
4 Diskussion
- 4.1 Generalisérbarhed af projektresultater
- 4.2 Ekstrapolering fra laboratorie til strømrender
- 4.3 Forsinkede effekter efter endt pulseksponering
- 4.4 Interspecifikke interaktioner og flerarts-forsøg
- 4.5 Sammenstilling af laboratorie- og strømrendeforsøg
4.1 Generalisérbarhed af projektresultater
Baseret på projektets indledende forsøg med G. pulex kan der drages en række konklusioner vedrørende generalisérbarheden af projektresultaterne.
4.1.1 Relativ toksicitet af pyrethroider anvendt i Danmark
Indledningsvis blev eksponeringsforsøg med fem pyrethroider gennemført for at undersøge, hvorvidt det var muligt at evaluere eventuelle skadevirkninger af pyrethroider som stofgruppe på baggrund af studier af ét modelpyrethroid. Resultaterne viser, at de undersøgte pyrethroider har både samme kvalitative virkning på bevægelsesadfærden (hyperaktivitet og immobilisering) og meget sammenlignelige effektkoncentrationer hos G. pulex.
Generelt observeres hyperaktivitet ved 10 ng l-1, og dette forhøjede aktivitetsniveau opretholdes i resten af eksponeringsperioden. Ved 100 ng l-1 er G. pulex ikke i stand til at opretholde hyperaktiviteten, da der sker et gradvist tab af evnen til at udvise koordineret svømning, og den tilbagelagte afstand aftager. Immobilisering registreres generelt ved 1000 ng l-1 efter ca. 45-60 minutters eksponering. Der er enkelte afvigelser fra dette generelle billede for esfenvalerat og tau-fluvalinat. For esfenvalerats vedkommende observeres markant hyperaktivitet allerede ved 1 ng l-1. Ved eksponering for tau-fluvalinat ved 1000 ng l-1 registreres ikke fuldstændig immobilisering inden eksponeringsperiodens udløb.
For cypermethrin og esfenvalerat er de observerede adfærdsændringer samt de fundne effektkoncentrationer hos G. pulex i fuld overensstemmelse med tidligere videosporingsstudier (Nørum & Bjerregaard 2003).
I strømrenderne er der tidligere udført forsøg med G. pulex eksponeret for esfenvalerat (Møhlenberg et al. 2004). Eksponeringstiden i dette studie var henholdsvis 30 og 150 minutter i modsætning til den i nærværende projekt anvendte på 90 minutter. En sammenligning af effektkoncentrationerne for esfenvalerat (Møhlenberg et al. 2004) og lambda-cyhalothrin (nærværende projekt) bekræfter dog, at esfenvalerat er noget mere toksisk end lambda-cyhalothrin.
De forholdsvis sammenlignelige effektkoncentrationer er bemærkelsesværdige, når man tager i betragtning, at de fem anvendte pyrethroider har varierende karakteristika m.h.t. molekylstruktur, molvægt, vandopløselighed, oktanol-vand-fordelingsforhold (log KOW) og indhold af aktive isomerer.
Effekten af en række pyrethroider på mortaliteten hos ferskvandinvertebrater er tidligere blevet sammenlignet. Siegfried (1993) eksponerede i et større studie en række akvatiske insekter (kvægmyggen Simulium vitattum, vårfluerne Hydropsyche og Cheumatopsyche spp., døgnfluer Heptageniidae, vandnymferne Enellagma og Ishnura spp. og vandkærer Hydrophilus spp.) for bl.a. tre pyrethroider, permethrin, cypermethrin og bifenthrin. Efter 24 timers kontinuert eksponering lå LC50-værdierne for de tre pyrethroider inden for en faktor 8 for alle de anvendte arter, hvilket understøtter de sammenlignelige effektkoncentrationer observeret i nærværende projekt.
4.1.2 Pulsvarighedens betydning
Betydningen af pulseksponeringens varighed for adfærdsresponset blev undersøgt i laboratoriet for at fastlægge modelpulsvarigheden. Baseret på de opnåede resultater for G. pulex blev en modelpulsvarighed på 90 minutter valgt. Selv om effekten af lambda-cyhalothrin ved længere eksponerings-varigheder bliver gradvis mere markant, forekommer der intet kvalitativt anderledes respons. Det mere markante adfærdsrespons svarer tilsyneladende til ændringen ved en forøgelse af eksponeringskoncentrationen.
Pulsvarighedens betydning for drift er tidligere blevet undersøgt i de i nærværende projekt anvendte strømrender, hvor G. pulex blev eksponeret for esfenvalerat i henholdsvis 30 og 150 minutter (Møhlenberg et al. 2004). Det blev konkluderet, at driftresponset ved en 150 minutters eksponering svarede til responset udløst af en 30-minutters eksponering for en ca. 10 gange højere koncentration. Driftresponset blev dermed fundet til at afhænge af såvel eksponeringskoncentrationen som –varigheden, hvilket er i overensstemmelse med laboratorieresultaterne fra nærværende projekt. Sammenhængen mellem pulsvarighed, -koncentration og mortalitet hos pyrethroideksponerede ferskvandsinvertebrater er tidligere dokumenteret (f.eks. Møhlenberg et al. 2004; Reynaldi & Liess 2005).
4.1.3 Temperaturens betydning
Temperaturens indvirkning på adfærdsresponset hos G. pulex blev undersøgt i det temperaturinterval, som blev anvendt i projektets øvrige forsøg, d.v.s. 5-15 °C. Der var en tendens til et lidt mere markant og tidligere påbegyndt adfærdsrespons med stigende temperatur. I denne situation, hvor toksiciteten stiger med temperaturen, siges pyrethroiderne at udvise en positiv temperaturkoefficient. I litteraturen er pyrethroider rapporteret at have enten negative, ingen eller positive temperaturkoefficienter afhængig af hvilket toksikologisk endpoint, man undersøger.
For pyrethroiders effekt på de spændingsafhængige natriumkanaler i dyrs nervesystemer er konklusionen entydig, idet der er tale om en negativ temperaturkoefficient, der primært skyldes en forlængelse af åbningen af de spændingsafhængige natriumkanaler, og deraf forøget natriuminflux, ved faldende temperatur (Ahn et al. 1987; Vijverberg & van den Bercken 1990; Narahashi 1996; Song & Narahashi 1996).
Temperaturens betydning for pyrethroidernes effekt på dødelighed og adfærd er derimod ikke entydig. En række grundige studier viser enten negative, ingen eller positive temperaturkoefficienter, og endda i visse tilfælde en kombination af negative og positive koefficienter i forskellige temperaturintervaller (Alzogaray & Zerba 1993; Punzo 1993; Alzogaray et al. 1997, 1998; Valles et al. 1998; Akkerhuis et al. 1999; Hodjati & Curtis 1999). Årsager til dette komplekse billede kan bl.a. være temperaturafhængig metabolisme, temperaturakklimering, en sammenhæng mellem temperatur og bevægelsesadfærd koblet til pesticidoptagelse, og endelig at der registreres forskellige typer adfærdsparametre (hyperaktivitet, ataksi eller immobilisering). Ofte kan man i samme adfærdsstudie samtidigt observere en negativ (f.eks. for EC50-værdier efter 24 timer) og en positiv temperaturkoefficient (f.eks. ET50-værdier) (Alzogaray & Zerba 1993).
Ovenstående er primært baseret på terrestriske arter, og der mangler således viden om temperaturkoefficienter hos akvatiske invertebrater. I nærværende projekt var temperaturens indvirkning på adfærdsresponset hos G. pulex dog relativt lille sammenlignet med effekten af at ændre eksponerings-koncentrationen. Dermed vurderes resultaterne fra de forskellige delforsøg i projektet at være direkte sammenlignelige, da der kun forekom relativt små udsving i de anvendte temperaturer.
4.1.4 Pesticidblanding
Pesticider forefindes ofte samtidigt i vandløbene, og undersøgelser i fynske vandløb har dokumenteret forekomst af op til 18 pesticider i samme vandløb inden for ét døgn, mens der i enkeltvandprøver blev påvist op til 13 forskellige stoffer (Fyns Amt 1997). I nærværende projekt blev G. pulex eksponeret for lambda-cyhalothrin i kombination med et hyppigt forekommende pesticid, glyphosat (Fyns Amt 1997; Århus Amt 1999). Der var ingen ændring i det pyrethroidinducerede adfærdsrespons ved denne samtidige eksponering for glyphosat (1000 ng l-1).
Ved anvendelse af samme metodik som i nærværende forsøg eksponerede Nørum & Bjerregaard (2003) G. pulex for kombinationer af cypermethrin og esfenvalerat. Adfærdsresponset blev registreret ved pyrethroidkoncentrationer på 10, 100 og 1000 ng l-1, enten med 100% cypermethrin, 100% esfenvalerat eller 50% cypermethrin sammen med 50% esfenvalerat (på vægtbasis). Effekten af kombinationseksponeringen var sammenlignelig med effekten af enkeltstofferne.
Disse studier af pesticidblandinger er ikke omfattende nok til, at man kan udtale sig generelt om effekten af miljørealistiske kombinationer. Forsøgene illustrerer imidlertid, at de laboratoriebaserede videosporingsmetoder kan danne udgangspunktet for evaluering af subletale effekter af komplekse pesticidblandinger.
4.1.5 Eksponering om natten
Langt de fleste invertebrater udviser tydelig natdrift, hvilket afspejler et generelt højere aktivitetsniveau om natten (f.eks. Brittain & Eikeland 1988). Dette højere aktivitetsniveau er primært en tilpasning for at nedsætte prædationsrisikoen fra især fisk, f.eks ørred, der jager ved synets hjælp (f.eks. Friberg et al. 1994). G. pulex udviser udpræget natdrift (Elliott 2002b; Heckmann & Friberg 2005), og den højere aktivitet kunne forventes at have indflydelse på dyrenes respons til en pesticideksponering.
Imidlertid viste forsøgene med pulseksponering om natten ingen overordnede forskelle i driftrespons hos G. pulex ved 10 og 100 ng l-1 lambda-cyhalothrin sammenlignet med forsøgene gennemført om dagen. Derimod var der, i modsætning til forsøgene udført om dagen, ingen signifikante forskelle mellem antal af dyr i drift i de behandlede render og kontrolrenderne. Det skyldtes, at driftniveauet var væsentlig højere i kontrolrenderne om natten end om dagen.
Resultaterne viser, at det respons G. pulex udviser om dagen afspejler katastrofedrift (Brittain & Eikeland 1988) og er uafhængigt af dyrenes aktivitetsniveau. Resultaterne indikerer samtidig, at driftforsøg både i laboratoriet og i felten skal gennemføres om dagen/i lys for at nedsætte driften i kontrolforsøgene og dermed øge kontrasten til behandlingen.
4.2 Ekstrapolering fra laboratorie til strømrender
Projektets resultater viser entydigt, at såvel de observerede effektkoncentrationer som rangordningen af de anvendte arters følsomhed over for pyrethroider kan ekstrapoleres direkte fra laboratoriet til strømrenderne.
4.2.1 Sammenligning af effektkoncentrationer
I nedenstående Tabel 4.1 er effektkoncentrationerne for lambda-cyhalothrin sammenlignet på baggrund af NOEC- (no observable effect concentration) og LOEC- (lowest observable effect concentration) værdier. NOEC- og LOEC-værdier har den iboende svaghed, at de afhænger af, hvilke koncentrationer man har valgt at teste; dette problem kunne afhjælpes ved regressionsanalyse til bestemmelse af ECx-værdier. NOEC- og LOEC-angrebsvinklen fastholdes imidlertid, da antallet af anvendte eksponerings-koncentrationer er for lavt (5 i laboratoriet, og 2 eller 4 i strømrenderne) til egentlig koncentrations-respons-regressionsanalyse, og da formålet med sammenligningen er at afgøre, hvorvidt effektkoncentrationerne i laboratoriet og strømrenderne er af samme størrelsesorden, og ikke direkte at teste de fundne effektkoncentrationer mod hinanden.
De laveste observerede effektkoncentrationer (LOEC) for L. nigra (1 ng l-1), G. pulex (10 ng l-1), A. aquaticus (10 ng l-1) og H. sulphurea (100 ng l-1) er alle indenfor forventede miljørealistiske niveauer.
Tabel 4.1. Ekstrapolering fra laboratorie til strømrender. NOEC- og LOEC-lambda-cyhalothrin-koncentrationer (ng l-1) for hyperaktivitet (Hyper), hypoaktivitet (Hypo) og immobilisering (I) i laboratoriet og for drift (D) i strømrenderne efter 90 minutters eksponering.
| Laboratoriet | Strømrender | ||||||
| Art | NOEC | LOECHyper | LOECHypo | LOECI | NOECD | LOECD | |
| Leuctra nigra |
0,1 | - | 1 | 10 | - | 1 | |
| Gammarus pulex | 1 | 10 | 1000 | 1000 | 1 | 10 | |
| Asellus aquaticus | 1 | 10 | 100 | 1000 | ikke undersøgt | ikke undersøgt | |
| Heptagenia sulphurea | 10 | 100 | 10000 | 10000 | 10 | 100 | |
| Sericostoma personatum | 10000a | - | - | - | 1000 a | - | |
| Ancylus fluviatilis | 10000 a | - | - | - | 1000 a | - | |
a: Højeste testede koncentrationer i henholdsvis laboratoriet og strømrenderne.
4.2.2 Rangordning af arternes følsomhed
I såvel laboratoriet som i strømrenderne var rangordningen af de anvendte arters følsomhed over for lambda-cyhalothrin efter 90 minutters eksponering:
L. nigra > G. pulex > H. sulphurea > S. personatum = A. fluviatilis
I laboratoriet blev A. aquaticus endvidere testet og fundet til at have en følsomhed sammenlignelig med G. pulex.
I strømrendeforsøgene blev dyrenes tilstand i drift og ved indsamlingen efter forsøget vurderet ved visuel observation. Denne tilstandsbedømmelse er mere subjektiv end registreringen af drift i renderne og bevægelsesadfærden i laboratoriet, men for dyr med meget lav mobilitet eller lav evne til at drifte kan disse tilstandsvurderinger imidlertid nuancere ovennævnte rangordning. Det drejer sig især om den relative følsomhed af S. personatum og A. fluviatilis, der ud fra de i drift- og videosporingsforsøgene observerede effektkoncentrationer (Tabel 4.1) vurderedes at være sammenlignelige. Tages dyrenes tilstand i drift/ved indsamling i betragtning observeres, at mens alle A. fluviatilis vurderedes normalt aktive ved 1000 ng l-1, var dette ikke tilfældet for S. personatum. For S. personatum var der derimod en stigende andel af individer med påvirket adfærd med stigende koncentration, især ved 1000 ng l-1 hvor størstedelen af individerne var påvirkede.
Dette understøttes af et studie af Lauridsen et al. (2006), der fandt, at S. personatum var mere følsomme overfor pyrethroider, når man bestemte effekten på deres omsætning af organisk stof, end når man registrerede drift i strømrenderne. For visse arter med lav mobilitet/driftpotentiale vurderes ændringer i bevægelsesadfærd derfor at være et uhensigtsmæssigt endpoint for toksicitetsbedømmelse.
Dermed fås følgende rangordning baseret på en vægtning af projektets resultater:
L. nigra > G. pulex > H. sulphurea > S. personatum > A. fluviatilis
Dette er i overensstemmelse med tidligere studier af pyrethroiders effekter på ferskvandinvertebrater (Anderson 1982; Stephenson 1982; Anderson & Shubat 1984; Mian & Mulla 1992; Breneman & Pontasch 1994; Giddings et al. 2001; Schulz & Liess 2001a; Beketov 2004; Møhlenberg et al. 2004; Lauridsen & Friberg 2005; van Wijngaarden et al. 2005). I disse studier er mortaliteten og adfærdsændringer blevet sammenlignet i forskelligartede eksponeringsforsøg, og overordnet set er krebsdyr blandt de mest følsomme overfor insekticider efterfulgt af slørvinger og døgnfluer, mens visse vårfluer og snegle er blandt de mindst følsomme.
Den i projektet fundne rangordning er i overensstemmelse med studier af samfundsstruktur og drift i pesticideksponerede vandløb (Kreutzweiser & Sibley 1991; Sibley et al. 1991; Davies & Cook 1993; Friberg et al. 2003; Berenzen et al. 2005; Heckmann & Friberg 2005), ligesom det er tilfældet for mere generelle artsfølsomhedsfordelinger baseret på følsomheden over for en række organiske toksiske forbindelser (Wogram & Liess 2001; von der Ohe & Liess 2004).
4.3 Forsinkede effekter efter endt pulseksponering
Strømrendeforsøgene viste for G. pulex, L. nigra og H. sulphurea, at der fortsat var forhøjet drift hos de eksponerede grupper i den time, som observationsperioden varede efter pulseksponeringens ophør. Restitueringsforsøg i laboratoriet med G. pulex og A. aquaticus viser, at der kan registreres forsinkede effekter i 2-3 uger efter pulseksponeringens ophør. Resultaterne viser, at det primært er koncentrationen og sekundært pulsvarigheden, der bestemmer effekterne på mortalitet og immobiliseringen i perioden efter overførslen til ukontamineret vand.
Forsinkede effekter af pyrethroider på overlevelse, reproduktion, vækst eller emergens er tidligere fundet hos G. pulex, vårfluerne Limnephilus lunatus og Anabolia nervosa, stankelbenslarven Tipula maxima og dansemyggen Chironomus riparius (Abel & Garner, 1986; Liess, 1994; Liess & Schulz, 1996; Schulz & Liess, 2000; Schulz & Liess, 2001a,b,c; Liess, 2002; Cold & Forbes, 2004; Beketov & Liess, 2005; Forbes & Cold, 2005; Heckmann et al., 2005).
4.4 Interspecifikke interaktioner og flerarts-forsøg
I naturlige vandløbsøkosystemer indgår de forskellige invertebratarter i et komplekst samspil. Mulige effekter af pyrethroider på disse interspecifikke interaktioner blev undersøgt ved studier af rovdyr-byttedyrs-interaktioner i såvel laboratoriet som i strømrenderne, mens effekter på flere arter blev belyst i flerarts-strømrendeforsøg.
4.4.1 Rovdyr-byttedyrs-interaktion
Resultaterne fra rovdyr-byttedyrs-interaktionen mellem G. pulex (rovdyr) og L. nigra (byttedyr) i laboratoriet viser en markant påvirkning ved 10 ng l-1, hvor ca. halvdelen af L. nigra overlever en 60-minutters interaktionsperiode, mens dette kun sker for 8% i kontrolgruppen. Ved 100 ng l-1 observeres ingen prædation. Denne ændring i rovdyrsadfærd hos G. pulex kunne skyldes, at eksponerede L. nigra ikke er attraktive som føde og således undgås. Dog viser interaktionen i gruppen, hvor kun L. nigra blev eksponeret, at dette ikke er tilfældet, da alle L. nigra blev nedlagt. I stedet ignorerer G. pulex tilsyneladende L. nigra ved 100 ng l-1, selv om de gentagne gange mødes i forsøgsarenaen. Regressionsanalysen viser, at selv om overlevelsen af L. nigra ved interaktionsperiodens udløb således først påvirkes ved 10 ng l-1, øges tiden til 50% nedlagte fra 2 minutter i kontrolgruppen til 4 minutter ved såvel 1 ng l-1 som i gruppen, hvor kun L. nigra blev eksponeret. Ved 10 ng l-1 nås 50% nedlagte lige netop ikke ifølge regressionsanalysen.
Invertebraters drift kan påvirkes direkte (prædation, mekanisk) eller indirekte (duftstoffer) ved tilstedeværelsen af en prædator, enten ved at driften nedsættes eller forøges (f.eks. Wooster & Sih 1995; McIntosh et al. 1996). I tidligere forsøg er det vist, at andelen af store G. pulex i natdrift steg, når de blev udsat fra duftstoffer fra ørreder (Friberg et al. 1994). Samtidig har Schulz & Dabrowski (2001) vist en synergistisk effekt mellem fenvalerat (200 ng l-1) og tilstedeværelsen af en fiskeprædator på driften af en døgnflue. Når døgnfluen blev eksponeret for fenvalerat alene, var driften større, end når der også var fisk til stede, hvorimod driften i kontrolgruppen (uden fenvalerat) blev forøget ved tilstedeværelsen af fisk.
Forsøgene med G. pulex viste imidlertid, som observeret i forsøgene udført om natten, ingen overordnede forskelle i driftresponset hos G. pulex ved 10 og 100 ng l-1 lambda-cyhalothrin sammenlignet med forsøgene gennemført uden ørredlugt. Eftersom G. pulex nedsætter aktiviteten, og dermed driften, ved tilstedeværelsen af en fiskeprædator, må den adaptive værdi være, at prædationsrisikoen nedsættes ved denne adfærd. Eksponering med lambda-cyhalothrin (og andre pyrethroider) overskygger denne adfærd, idet driften øges, hvilket kan forventes at give ophav til et forøget prædationstryk under naturlige forhold. En tidligere undersøgelse har vist, at ørredprædation kan have en markant indflydelse på G. pulex populationer i mindre, danske vandløb (Andersen et al. 1993).
4.4.2 Flerarts-forsøg
I det 10 dage lange forsøg vedrørende effekter af lambda-cyhalothrin på flere arter udviste tre af de fire arter et andet driftrespons end i kortidsforsøgene. G. pulex, L. nigra og H. sulphurea udviste enten lavere eller tilsvarende driftrater i de behandlede render sammenlignet med kontrolrenderne. A. fluviatilis udviste ingen driftadfærd i de to typer af forsøg. Den observerede forskel i drift kan tilskrives forskellige aktivitetsniveauer og afspejler derfor, at organismernes adfærd er påvirket i en længere periode efter eksponeringen, hvilket er i overensstemmelse med resultaterne af restitueringsforsøgene i laboratoriet. Tætheden af alle 4 arter var tilnærmelsesvis uændret i forsøgsperioden på grund af de generelt lave driftrater i de behandlede render og at dyrene blev genudsat efter, at de var fanget i drift. Derfor kan faldet i græsningsraten og i omsætningsraten af organisk stof ikke tilskrives nedsatte tætheder på grund af, at individer blev tabt fra renderne.
Fødefunktionelt er A. fluviatilis den væsentligste og mest effektive græsser i systemet efterfulgt af H. sulphurea (Elliott et al. 1988; Dall & Lindegaard 1995). Imidlertid kan både G. pulex og L. nigra udnytte biofilm, herunder bentiske alger, såfremt denne ressource er lettilgængelig (Marchant 1981; Henderson et al. 1990). At algebiomassen stiger i render, hvor invertebraterne var blevet behandlet med 100 ng l-1 lambda-cyhalothrin sammenlignet med kontrolrenderne og renderne eksponeret for 10 ng l-1 viser, at invertebraterne ikke er i stand til at regulere mængden af alger. Kjeldsen (1996) fandt i overensstemmelse hermed, at algebiomassen på sten jævnligt skyllet med et pyrethroid (permetrin) var højere end på sten skyllet med vand, fordi invertebratgræsserne blev holdt nede i antal.
I indeværende projekt er der ikke gennemført forsøg med arterne separat, og det er derfor ikke muligt at konkludere, hvilke arter der har nedsat fødeoptag og dermed bidraget til det observerede fald i græsningsrate. Imidlertid kan forsøget indikere, at A. fluviatilis muligvis er påvirket, selv om hverken driftadfærd eller dyrenes observerede tilstand er ændret, og at dette giver sig udtryk i en nedsat græsningsaktivitet. I tidligere undersøgelser er det vist, at S. personatum’s evne til omsætte organisk stof er et mere følsomt end-point end deres driftadfærd (Møhlenberg et al. 2004; Lauridsen et al. 2006). Alternativt kan resultatet tolkes, som at de øvrige arter under upåvirkede forhold spiller en større rolle som græssere, end hvad der kunne forventes udfra tidligere undersøgelser af deres fødefunktionalitet. Dette spørgsmål kan kun afgøres ved at udføre forsøg med hver enkelt art alene.
Omsætningen af organisk stof var lavere for begge behandlinger sammenlignet med kontrolrenderne. G. pulex og L. nigra, der dominerede tæthedsmæssigt i render, er begge primært iturivere og lever af at omsætte dødt partikulært organisk stof i vandløbsøkosystemer (f.eks. Iversen 1988). Da begge arter i alle typer af forsøg i nærværende projekt har udvist påvirket adfærd ved koncentrationer på 10 ng l-1 lambda-cyhalothrin understøttes flerarts-studiet af projektets øvrige resultater. De viser samtidig, at den påvirkning, der observeres i laboratoriet, direkte kan overføres til en påvirkning på flerarts-niveau og til at forudsige en ændring af den økologiske funktion.
For G. pulex er det tidligere vist, at omsætningsraten af organisk stof påvirkes negativt (falder) ved eksponering med esfenvalerat og lambda-cyhalothrin (Møhlenberg et al. 2004; Lauridsen et al. 2006). Andre undersøgelser har vist effekter af pesticider på samfunds- og økosystemniveau i søer og vandløb (f.eks. Lampert et al. 1989; Wallace et al. 1991). Fra fynske vandløb foreligger endvidere direkte observationer af masseopblomstringer af tråd- og kiselalger i forbindelse med udledninger af pyrethroider fra f.eks. gartnerier, når disse udledninger medfører at f.eks. samtlige G. pulex og andre invertebratgræssere slås ihjel (Wiberg-Larsen pers. kom.) Der er imidlertid ingen undersøgelser, der eksperimentelt har vist så klare indirekte effekter af en pesticidpåvirkning som nærværende forsøg. Ekstrapoleres projektets resultater til et egentlig vandløb, kan det forudsiges, at subletale eksponeringer med pyrethroider vil ændre energifluksene gennem økosystemets fødenet, hvilket kunne give ophav til f.eks. opblomstring af trådalger og nedsat sekundærproduktion.
4.5 Sammenstilling af laboratorie- og strømrendeforsøg
I projektet er der gennemført både korttids- og langtidsforsøg i såvel laboratoriet som i strømrenderne, og en tværgående analyse af resultaterne fører til en række konklusioner. Ved kobling af korttidsstudierne i laboratoriet og strømrenderne observeres, at der kan være kvalitative artsforskelle i adfærdsresponset. Ved de koncentrationer, der for hver art fører til forøget drift i strømrenderne, udviser G. pulex og H. sulphurea et forøget, og L. nigra et reduceret, aktivitetsniveau i laboratoriet. Overordnet viser korttidsstudierne derfor, at en ændring i bevægelsesadfærden i laboratoriet, uanset om det er en forøgelse eller en reduktion i aktiviteten, kan ekstrapoleres til forøget driftadfærd i strømrenderne.
I det længerevarende flerartsforsøg udviste eksponerede G. pulex og L. nigra reduceret driftadfærd sammenlignet med kontrolgrupperne, mens der ikke var nogen forskelle mellem eksponerede dyr og kontroldyr hos H. sulphurea. Samtidig viser forsøgene en forøget algebiomasse og en reduceret omsætning af organisk materiale ved sammenligning af eksponerede render og kontrolrender. Resultaterne indikerer dermed, at primært G. pulex, L. nigra og A. fluviatilis i denne posteksponeringsfase er påvirkede i en grad, der fører til ændringer i deres fødesøgningsadfærd men ikke i deres evne til at opretholde deres position. Den reducerede drift for G. pulex og L. nigra skyldes tilsyneladende et generelt reduceret aktivitetsniveau, hvilket understøttes af de direkte observationer af de driftende dyrs tilstand, hvor andelen af hæmmede G. pulex og L. nigra steg.
5 Konklusioner
Projektet dokumenterer, at det for de anvendte arter, ferskvandstangloppen Gammarus pulex, slørvingen Leuctra nigra, døgnfluen Heptagenia sulphurea, vårfluen Sericostoma personatum og huesneglen Ancylus fluviatilis, er muligt at ekstrapolere direkte fra effekter af pulseksponering med lambda-cyhalothrin i laboratoriestudier af bevægelsesadfærd til studier af drift i de mere miljørealistiske strømrender. Dette gælder for såvel de observerede effektkoncentrationer som rangordningen af de anvendte arters følsomhed over for lambda-cyhalothrin. Der kan imidlertid være kvalitative artsforskelle i adfærdsresponset. F.eks. observeres det i laboratoriet, at det for G. pulex er et forøget aktivitetsniveau, mens det for L. nigra er et reduceret aktivitetsniveau, der er baggrunden for den observerede forøgede drift i strømrenderne. Når sammenhængen mellem ændringerne i bevægelsesadfærd i laboratoriet og driftresponset i strømrenderne hos den enkelte art er dokumenteret, vurderes laboratoriestudierne at kunne erstatte strømrendestudierne. Dette gælder dermed for de i projektet anvendte arter.
Metoderne komplementerer hinanden, således at de omkostningseffektive laboratoriestudier kan anvendes til hurtige screeninger af mulige effekter af pesticider på ferskvandsinvertebraters bevægelsesadfærd med henblik på fastlæggelse af effektkoncentrationer og rangordning af arternes følsomhed. Disse data kan bl.a. anvendes i planlægningen af strømrendeforsøg. Samtidig kan observationer fra mere komplekse strømrendestudier, som f.eks. effekter i flerarts-forsøg, opklares ved efterfølgende laboratoriestudier af bevægelsesadfærd, restituering og interspecifikke interaktioner.
Alpha-cypermethrin, cypermethrin, lambda-cyhalothrin og tau-fluvalinat påvirker alle markant adfærden hos G. pulex ved en koncentration på 10 ng l-1, mens esfenvalerat fører til adfærdsændringer allerede ved 1 ng l-1. Alle de undersøgte pyrethroider har dermed effekter ved koncentrationer, der knapt kan måles; kravet til detektiongrænsen i de analysekemiske metoder, man normalt anvender til at monitere pesticider i grundvand og overfladevand, er til sammenligning 10 ng l-1.
Kortvarige pulseksponeringer med lambda-cyhalothrin fører til forsinkede effekter på adfærd og overlevelse hos ferskvandsinvertebrater; effekter der kan registreres i 2-3 uger efter pulseksponeringens ophør.
Rovdyr-byttedyrs-interaktioner påvirkes ved pulseksponering for lambda-cyhalothrin. Under eksponering ændrer G. pulex sit naturlige driftrespons på tilstedeværelsen af en prædator (ørred), nemlig ved at driftraten ved lambda-cyhalothrin-eksponering øges, hvor den ellers normalt ville reduceres. Dette kan have negative konsekvenser for vandløbspopulationer. G. pulex’s prædation på L. nigra påvirkes endvidere markant ved 10 ng l-1.
Pulseksponering med lambda-cyhalothrin udløser klare indirekte effekter på tilvæksten i algebiomassen og på omsætningen af organisk stof. Dette indikerer, at miljørealistiske eksponeringer af vandløbsinvertebratsamfundet med lambda-cyhalothrin kan give effekter på økosystemniveau, der går udover påvirkningen af specifikke arter og populationer.
6 Perspektivering
6.1 Forskningsmæssige perspektiver
Projektets resultater fra flerarts-forsøget viser, at de undersøgte arter er i stand til at regulere algebiomassen og omsætte større mængder organisk stof, når de ikke er påvirket af lambda-cyhalothrin. Dette er et væsentligt resultat i vandløbsøkologisk henseende, da der især tidligere har været diskussion om, hvorvidt biologisk samspil har en betydning for struktur og funktion i vandløb (Sand-Jensen & Friberg, 2000). Undersøgelserne i dette projekt viser, at disse samspil er betydningsfulde, og det er væsentligt i vores vurdering af miljøpåvirkninger, som f.eks. pesticidbelastning, på vandløb. Med udgangspunkt i vores undersøgelser kan man forvente, at belastningen med pyrethroider vil have en række effekter, der ikke er direkte koblet til enkelte arters tolerance overfor stofferne. Sådanne effekter kan ikke ekstrapoleres udfra enkeltarts-test, men skal måles direkte som i indeværende projekt. Der er derfor et klart behov for, at undersøgelser med et højere kompleksitets-niveau, end det kendes fra standardtests, gennemføres med henblik på at etablere det nødvendige faglige grundlag for at vurdere risikoen for vandmiljøet ved brugen af pyrethroider. Da sådanne mere komplekse undersøgelser er ressourcekrævende, og generelt er kendetegnet ved en høj variabilitet og dermed et større behov for statistisk replikering, er det meget væsentlig at udvælge modelstoffer som i indeværende projekt, hvor såvel deres toksicitet overfor målorganismegrupper samt deres toksicitet i forhold til nært beslægtede stoffer/stofgrupper er kendt.
6.2 Administrative konsekvenser
De i projektet gennemførte forsøg illustrerer nødvendigheden af at inddrage pulseksponeringsproblematikken og subletale toksicitetsmål i risikovurderingssammenhæng, i stedet for fortsat at forlade sig på mortalitetsstudier under kontinuerte eksponeringsbetingelser, der for vandløbenes vedkommende åbenlyst ikke er miljørealistiske. Selv om dette behov i nærværende projekt udelukkende er dokumenteret for pyrethroid-insekticiderne, vurderes det ønskværdigt at gennemføre tilsvarende studier for andre insekticider med virkemekanismer, der retter sig mod invertebraters nervesystem. Mulige neurotoksiske insekticider inkluderer såvel allerede godkendte stofgrupper, f.eks. organophosphater og carbamater, som potentielle afløsere for eksisterende stoffer, f.eks. neonicotinoider.
For alle fem undersøgte pyrethroider viser resultaterne, at selv kortvarig pulseksponering for meget lave koncentrationer fører til markante adfærdsændringer hos ferskvandstangloppen Gammarus pulex. Disse lave effektkoncentrationer indikerer, at pyrethroiderne kan påvirke vandløbsinvertebratsamfundene ved niveauer, der knapt kan måles med de analysekemiske metoder, man normalt anvender til at monitere pesticider i grundvand og overfladevand. Koncentrationerne er samtidig væsentligt lavere end de foreliggende effektkoncentrationer fra standardtoksicitetstest, der anvendes i forbindelse med risikovurdering af pesticider. Projektet dokumenterer endvidere, at kortvarige pulseksponeringer med pyrethroidet lambda-cyhalothrin fører til forsinkede effekter på adfærd og overlevelse hos ferskvandsinvertebrater; effekter der kan registreres i 2-3 uger efter pulseksponeringens ophør. Sådanne forsinkede effekter er af central betydning i forbindelse med vurderingen af potentielle skadevirkninger i miljøet og bør følgelig tillægges større betydning i forbindelse med førnævnte risikovurdering.
Resultaterne afslører store artsforskelle i følsomhed over for pyrethroider, hvilket nødvendiggør studier af en bredere vifte af arter for i tilstrækkelig grad at kunne vurdere risikoen for vandløbsinvertebratfaunaen. Af projektets resultater fremgår, at det for de fem anvendte invertebratarter er muligt at ekstrapolere direkte fra pyrethroid-inducerede adfærdsændringer observeret i laboratoriet til drift under mere miljørealistiske betingelser. For disse arter kan de omkostningseffektive, laboratoriebaserede videosporingsstudier dermed erstatte strømrendestudierne, og videosporingssystemet vurderes at kunne anvendes til screeninger af mulige effekter af pesticider på ferskvandsinvertebraters bevægelsesadfærd. Projektets reultater indikerer, at sådanne adfærdsændringer kan være prædiktive for effekter på økosystemniveau. En mere formel anvendelse af adfærdsundersøgelserne som risikovurderingstestsystem i godkendelsesordningen for pesticider forudsætter imidlertid udarbejdelse af standardiserede testguidelines.
7 Referencer
Abel PD and Garner SM (1986) Comparisons of median survival times and median lethal exposure times for Gammarus pulex exposed to cadmium, permethrin and cyanide. Water Research 20 (5), 579-582.
Ahn YJ, Shono T and Fukami JI (1987) Effect of Temperature on Pyrethroid Action to Kdr-Type Housefly Adults. Pesticide Biochemistry and Physiology 28 (3), 301-307.
Akkerhuis GAJM, Kjaer C, Damgaard C and Elmegaard N (1999) Temperature-dependent, time-dose-effect model for pesticide effects on growing, herbivorous arthropods: Bioassays with dimethoate and cypermethrin. Environmental Toxicology and Chemistry 18 (10), 2370-2378.
Allan JD (1995) Stream Ecology. Chapman & Hall, London, UK.
Allan JD, Flecker AS and McClintock (1986) Diel epibenthic activity of mayfly nymphs and its nonconcordance with behavioral drift. Limnology and Oceanography 31, 1057-1065.
Alzogaray RA and Zerba EN (1993) Temperature Effect on the Insecticidal Activity of Pyrethroids on Triatoma infestans. Comparati ve Biochemistry and Physiology C-Pharmacology Toxicology and Endocrinology 104 (3), 485-488.
Alzogaray RA, Fontan A and Zerba EN (1997) Evaluation of hyperactivity produced by pyrethroid treatment on third instar nymphs of Triatoma infestans (Hemiptera: Reduviidae). Archives of Insect Biochemistry and Physiology 35 (3), 323-333.
Alzogaray RA, Picollo MI and Zerba EN (1998) Independent and joint action of cis- and trans-permethrin in Triatoma infestans (Hemiptera : Reduviidae). Archives of Insect Biochemistry and Physiology 37 (3), 225-230.
Andersen TH, Friberg N, Hansen HO, Iversen TM, Jacobsen D and Krøjgaard L (1993) The effects of introduction of brown trout (Salmo trutta L.) on Gammarus pulex L. drift and density in two fishless Danish streams. Archiv für Hydrobiologie 126, 361-371.
Anderson RL (1982) Toxicity of fenvalerate and permethrin to several nontarget aquatic invertebrates. Environmental Entomology 11 (6), 1251-1257.
Anderson RL and Shubat P (1984) Toxicity of Flucythrinate to Gammarus lacustris (Amphipoda), Pteronarcys dorsata (Plecoptera) and Brachycentrus americanus (Trichoptera) - Importance of Exposure Duration. Environmental Pollution Series A-Ecological and Biological 35 (4), 353-365.
Beketov MA (2004) Comparative sensitivity to the insecticides deltamethrin and esfenvalerate of some aquatic insect larvae (Ephemeroptera and Odonata) and Daphnia magna. Russian Journal of Ecology 35 (3), 200-204.
Beketov MA and Liess M (2005) Acute contamination with esfenvalerate and food limitation: Chronic effects on the mayfly, Cloeon dipterum. Environmental Toxicology and Chemistry 24 (5), 1281-1286.
Berenzen N, Kumke T, Schulz HK and Schulz R (2005) Macroinvertebrate community structure in agricultural streams: impact of runoff-related pesticide contamination. Ecotoxicology and Environmental Safety 60 (1), 37-46.
Bichel-udvalget (1999) Rapport fra Underudvalget for Miljø og Sundhed. 244 s.
Breneman DH and Pontasch KW (1994) Stream Microcosm Toxicity Tests: Predicting the Effects of Fenvalerate on Riffle Insect Communities. Environmental Toxicology and Chemistry 13 (3), 381-387.
Brittain JE and Eikeland TJ (1988) Invertebrate drift – A review. Hydrobiologia, 166, 77-93.
Ciborowski JJH and Corkum LD (1980) Importance of behaviour to the re-establishment of drifting Ephemeroptera. Advances in Ephemeroptera Biology, Plenum, New York, 321-330.
Cold A and Forbes VE (2004) Consequences of a short pulse of pesticide exposure for survival and reproduction of Gammarus pulex. Aquatic Toxicology 67 (3), 287-299.
Dall P and Lindegaard C (1995) En oversigt over danske ferskvandinvertebrater til brug ved bedømmelse af forurening i søer og vandløb. Ferskvandsbiologisk Laboratorium, Københavns Universitet.
Davies PE and Cook LSJ (1993) Catastrophic Macroinvertebrate Drift and Sublethal Effects on Brown Trout, Salmo-Trutta, Caused by Cypermethrin Spraying on A Tasmanian Stream. Aquatic Toxicology 27 (3-4), 201-224.
Der Ohe PC and Liess M (2004) Relative sensitivity distribution of aquatic invertebrates to organic and metal compounds. Environmental Toxicology and Chemistry 23 (1), 150-156.
Elliott JM (2002a) Time spent in the drift by downstream-dispersing invertebrates in a lake district stream. Freshwater Biology 47, 97-106.
Elliott JM (2002b) A continuous study of the total drift of freshwater shrimps, Gammarus pulex, in a small stony stream in the English Lake District. Freshwater Biology, 47, 75-86.
Elliott JM, Humpesch UH and Macan TT (1988) Larvae of the British Ephemeroptera: a key with ecological notes. FBA scientific publication no. 49, Ambleside, Cumbria, UK.
Felding G, Mogensen BB, Sørensen JB, Hansen AC (1997) Surface run-off of pesticides from farmland to streams and lakes. Bekæmpelsesmiddelforskning fra Miljøstyrelsen, nr. 29, 76 s.
Forbes VE and Cold A (2005) Effects of the pyrethroid esfenvalerate on life-cycle traits and population dynamics of Chironomus riparius - Importance of exposure scenario. Environmental Toxicology and Chemistry 24 (1), 78-86.
Friberg N, Andersen TH, Hansen HO, Iversen TM, Jacobsen D, Krøjgaard L and Larsen SE (1994) The effect brown trout (Salmo trutta L.) on stream invertebrate drift, with specieal reference to Gammarus pulex L. Hydrobiologia, 294, 105-110.
Friberg N, Kronvang B, Svendsen LM, Hansen HO and Nielsen MB (1994) Restoration of a channelized reach of the River Gelså, Denmark: effects on the macroinvertebrate community. Aquatic Conservation: Marine and Freshwater Ecosystems 4, 289-296.
Friberg N (1997) Benthic invertebrate communities in six Danish forest streams: impact of forest type on structure and function. Ecography 20, 19-28.
Friberg N, Larsen AD, Rodkjaer A, and Thomsen AG (2002) Shredder guilds in three Danish forest streams contrasting in forest type. Archiv für Hydrobiologie 153 (2), 197-215.
Friberg N, Lindstrom M, Kronvang B, and Larsen SE (2003) Macroinvertebrate/sediment relationships along a pesticide gradient in Danish streams. Hydrobiologia 494 (1-3), 103-110.
Fyns Amt (1997) De fynske vandløb. VANDMILJØovervågning. Tema: Ferskvand. Fyns Amt, Natur- og Vandmiljøafdelingen, 210 s.
Fyns Amt (2000) Fyns Vandmiljø. Status over 25 års indsats og resultater. Fyns Amt, Natur- og Vandmiljøafdelingen, 144 s.
Giddings JM, Solomon KR, and Maund SJ (2001) Probabilistic risk assessment of cotton pyrethroids: II. Aquatic mesocosm and field studies. Environmental Toxicology and Chemistry 20 (3), 660-668.
Girling AE, Pascoe D, Janssen CR, Peither A, Wenzel A, Schäfer H, Neumeier B, Mitchell GC, Taylor EJ, Maund SJ, Lay JP, Jüttner I, Crossland NO, Stephenson RR, Persoone G (2000) Development of methods for evaluating toxicity to freshwater ecosystems. Ecotoxicology and Environmental Safety 45, 148-176.
Heckmann LH and Friberg N (2005) Macroinvertebrate community response to pulse exposure with the insecticide lambda-cyhalothrin using in-stream mesocosms. Environmental Toxicology and Chemistry 24 (3), 582-590.
Heckmann LH, Friberg N, and Ravn HW (2005) Relationship between biochemical biomarkers and pre-copulatory behaviour and mortality in Gammarus pulex following pulse-exposure to lambda-cyhalothrin. Pest Management Science 61 (7), 627-+.
Henderson J, Hildrew AG and Townsend CR (1990) Detritivorous stoneflies in an iron-rich stream: food and feeding. I: I.C. Campbell (red.) Mayflies and stoneflies. Kluwer Academic Publishers, Dordrecht, The Netherlands, 249-254.
Hodjati MH and Curtis CF (1999) Effects of permethrin at different temperatures on pyrethroid-resistant and susceptible strains of Anopheles. Medical and Veterinary Entomology 13 (4), 415-422.
Iversen TM (1988) Secondary production and trophic relationships in a spring invertebrate community. Limnology and Oceanography, 33, 582-592.
Iversen TM and Jessen J (1977) Life-cycle, drift and production of Gammarus pulex (L.) (Amphipoda) in a Danish spring. Freshwater Biology 7(3), 287-296.
Kjeldsen K (1996) Regulation of algal biomass in a small lowland stream: field experiments on the role of invertebrate grazing, phosphorous and irradiance. Freshwater Biology, 36, 535-546.
Kreutzweiser DP and Sibley PK (1991) Invertebrate Drift in A Headwater Stream Treated with Permethrin. Archives of Environmental Contamination and Toxicology 20 (3), 330-336.
Lampert W, Fleckner W, Pott E, Schober U and Stùrkel KU (1989) Herbicide effects on planctonic systems of differenrent complexity. Hydrobiologia 188/189, 415-424.
Lauridsen RB and Friberg N (2005) Stream macroinvertebrate drift response to pulsed exposure of the synthetic pyrethroid lambda-cyhalothrin. Environmental Toxicology 20 (5), 513-521.
Lauridsen RB, Kronvang B and Friberg N (2006) Occurrence and impact of sediment-bound pyrethroids in Danish streams. Water, Air and Soil Pollution, in press.
Liess M (1994) Pesticide impact on macroinvertebrate communities of running waters in agricultural ecosystems. Verh.Internat.Verein.Limnol. 25 2060-2062.
Liess M (2002) Population response to toxicants is altered by intraspecific interaction. Environmental Toxicology and Chemistry 21 (1), 138-142.
Liess M and Schulz R (1996) Chronic effects of short-term contamination with the pyrethroid insecticide fenvalerate on the caddisfly Limnephilus lunatus. Hydrobiologia 324 (2), 99-106.
Liess M and Schulz R (1999) Linking insecticide contamination and population response in an agricultural stream. Environmental Toxicology and Chemistry 18 (9), 1948-1955.
Marchant R (1981) The ecology of Gammarus in running water. I: MA Lock and DD Williams (red.) Perspectives in running water ecology. Plenum Press, New York, US, 225-250.
Matthiessen P, D Sheahan, R Harrisson, M Kirby, R Rycroft, A Turnbull, C Volkner, R Williams (1995) Use of a Gammarus pulex bioassay to measure the effects of transient carbofuran runoff from farmland. Ecotoxicology and Environmental Safety 30, 111-119.
McIntosh AR and Townsend CR (1996) Interactions between fish, grazing invertebrates and algae in New Zealand stream: A trophic cascade mediated by fish-induced changes to grazer behaviour? Oecologia, 108, 174-181
Mian LS and Mulla MS (1992) Effects of Pyrethroid Insecticides on Nontarget Invertebrates in Aquatic Ecosystems. Journal of Agricultural Entomology 9 (2), 73-98.
Miljøstyrelsen (2002) Bekæmpelsesmiddelstatistik 2001. Orientering fra miljøstyrelsen Nr. 5, 43 s.
Miljøstyrelsen (2003) Bekæmpelsesmiddelstatistik 2002. Orientering fra miljøstyrelsen Nr. 5, 48 s.
Miljøstyrelsen (2004) Bekæmpelsesmiddelstatistik 2003. Orientering fra miljøstyrelsen Nr. 9, 47 s.
Miljøstyrelsen (2005) Bekæmpelsesmiddelstatistik 2004. Orientering fra miljøstyrelsen Nr. 6, 51 s.
Møhlenberg F, Kaas H, Schlüter L, Gustavson K, Andersen TT, Forbes V, Cold A, Friberg N, Larsen SE and Lauridsen RB (2004) Effekter af bekæmpelsesmidler på flora og fauna i vandløb. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 82.
Narahashi T (1996) Neuronal ion channels as the target sites of insecticides. Pharmacology & Toxicology 78 (1), 1-14.
Nørum U, P Bjerregaard (2003) Ferskvandsinvertebraters bevægelsesadfærd som biomarkør for pesticideksponering og -effekt. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 75. 66 s.
OECD (2000) OECD guidelines for testing of chemicals, revised proposal for updating guideline 202: Daphnia sp., acute immobilisation test.
Punzo F (1993) Detoxification Enzymes and the Effects of Temperature on the Toxicity of Pyrethroids to the Fall Armyworm, Spodoptera-Frugiperda (Lepidoptera, Noctuidae). Comparative Biochemistry and Physiology C-Pharmacology Toxicology and Endocrinology 105 (2), 155-158.
Reynaldi S and Liess M (2005) Influence of duration of exposure to the pyrethroid fenvalerate on sublethal responses and recovery of Daphnia magna Straus. Environmental Toxicology and Chemistry 24 (5), 1160-1164.
Schulz R and Liess M (1999a) Validity and ecological relevance of an active in situ bioassay using Gammarus pulex and Limnephilus lunatus. Environmental Toxicology and Chemistry 18, 2243-2250.
Schulz R and Liess M (1999b) A field study of the effects of agriculturally derived insecticide input on stream macroinvertebrate dynamics. Aquatic Toxicology 46, 155-176.
Schulz R and Liess M (2000) Toxicity of fenvalerate to caddisfly larvae: chronic effects of 1-vs 10-h pulse-exposure with constant doses. Chemosphere 41 (10), 1511-1517.
Schulz R and Dabrowski JM (2001) Combined effects of predatory fish and sublethal pesticide contamination on the behavior and mortality of mayfly nymphs. Environmental Toxicology and Chemistry 20 (11), 2537-2543.
Schulz R and Liess M (2001a) Acute and chronic effects of particle-associated fenvalerate on stream macroinvertebrates: A runoff simulation study using outdoor microcosms. Archives of Environmental Contamination and Toxicology 40 (4), 481-488.
Schulz R and Liess M (2001b) Toxicity of aqueous-phase and suspended particle-associated fenvalerate: Chronic effects after pulse-dosed exposure of Limnephilus lunatus (Trichoptera). Environmental Toxicology and Chemistry 20 (1), 185-190.
Schulz R and Liess M (2001c) Runoff simulation with particle-bound fenvalerate in multispecies stream microcosms: Importance of biological interactions. Environmental Toxicology and Chemistry 20 (4), 757-762.
Sibley PK, Kaushik NK, and Kreutzweiser DP (1991) Impact of A Pulse Application of Permethrin on the Macroinvertebrate Community of A Headwater Stream. Environmental Pollution 70 (1), 35-55.
Siegfried BD (1993) Comparative Toxicity of Pyrethroid Insecticides to Terrestrial and Aquatic Insects. Environmental Toxicology and Chemistry 12 (9), 1683-1689.
Song JH and Narahashi T (1996) Modulation of sodium channels of rat cerebellar Purkinje neurons by the pyrethroid tetramethrin. Journal of Pharmacology and Experimental Therapeutics 277 (1), 445-453.
Stephenson RR (1982) Aquatic Toxicology of Cypermethrin .I. Acute Toxicity to Some Fresh-Water Fish and Invertebrates in Laboratory Tests. Aquatic Toxicology 2 (3), 175-185.
Valles SM, Sanchez-Arroyo H, Brenner RJ, and Koehler PG (1998) Temperature effects on lambda-cyhalothrin toxicity in insecticide-susceptible and resistant German cockroaches (Dictyoptera : Blattellidae). Florida Entomologist 81 (2), 193-201.
Van Wijngaarden RPA, Brock TCM, and Van den Brink PJ (2005) Threshold levels for effects of insecticides in freshwater ecosystems: A review. Ecotoxicology 14 (3), 355-380.
Vijverberg HPM and Vandenbercken J (1990) Neurotoxicological Effects and the Mode of Action of Pyrethroid Insecticides. Critical Reviews in Toxicology 21 (2), 105-126.
Wallace JB, Cuffney TF, Webster JR, Lugthart GJ, Chung K and Goldowitz BS (1991) Export of fine organic particles from headwater streams: Effects of season, extreme discharges and invertebrate manipulations. Limnology and Oceanography, 36, 670-682.
Waters TF (1965) Interpretation of invertebrate drift in streams. Ecology 46, 327-334.
Wogram J and Liess M (2001) Rank ordering of macroinvertebrate species sensitivity to toxic compounds by comparison with that of Daphnia magna. Bulletin of Environmental Contamination and Toxicology 67 (3), 360-367.
Wooster D and Sih A (1995) A review of the drift and activity responses of stream prey to predator presence. Oikos, 73, 3-8.
Zelinka M and Marvan P (1976) Notes to estimating production of zoobenthos. Folia Fac. Sci. Nat. Univ. Purkynianae Brunensis 17, Biol. 58(10), 1-53,
Århus Amt (1999) Pesticider i vandløb, kilder og søer i Århus Amt.Teknisk Rapport. Århus Amt, Natur og Miljøkontoret, 51 s.
1 Statistik for strømrendeforsøg
1.1 Analyse af drift data fra koncentrations-respons-forsøg
Data indeholder en optælling af antallet af drift, aktive, påvirkede og døde individer fra 3 forskellige arter opgjort til forskellige tider før og efter eksponering – fra 12 render udsat for 2 forskellige koncentrationer af pesticid og kontrol (n=4).
Statistisk design:
Behandling: 2 doser af pesticid og kontrol
Replikater: Render
Respons: Antal driftede, aktive påvirkede eller døde individer for 3 arter. I Tabel X (se side X) er kun resultater for driftede dyr angivet. Antallet af driftede dyr er summeret op til tid 90 min efter eksponering og tilsvarende til tid 150 min. Ligeledes er antallet af driftede dyr mellem 90 og 150 min analyseret.
Statistisk metode: One-way ANOVA med behandling som fixed effekt. Parvise test udføres ved brug af lsmeans og t-test.
Model:
![]()
i: Angiver behandling (i=1,2,3)
j: Angiver rende (j=1,2,3,4)
1.2 Analyse af driftresultater i samfundsstruktur-eksperimentet
Data indeholder en optælling af antallet af drift, aktive, påvirkede og døde individer fra 3 forskellige arter samt de totale antal opgjort fra dag 1 til 10 – fra 12 render udsat for 2 forskellige koncentrationer af pesticid og kontrol (n=4).
Statistisk design:
Behandling: 2 doser af pesticid og kontrol
Replikater: Render
Respons: Antal driftede, aktive påvirkede eller døde individer for 3 arter – ligeledes er det totale antal individer i de 4 kategorier analyseret.
Statistisk metode: Repeated mixed effekt model med behandling som fixed effekt og render som random effekt og dag som repeated. Parvise test udføres ved brug af lsmeans og t-test. Der anvendes compound symmetri varians.
1.3 Analyse af vægttab hos bladpakker i samfundsstruktur-eksperimentet
Data indeholder målt tab fra bladpakker – fra 12 render udsat for 2 forskellige koncentrationer af pesticid og kontrol. Hver rende indeholder 5 bladpakker.
Statistisk design:
Behandling: 2 doser af pesticid og kontrol
Replikater: Render og bladpakker
Respons: Tab
Statistisk metode: Mixed effekt model med behandling som fixed effekt og render som random effekt. Parvise test udføres ved brug af lsmeans og t-test.
Model:
![]()
i: Angiver behandling (i=1,2,3)
j: Angiver rende (j=1,2,3,4)
k: Angiver bladpakke (k=1,2,3,4,5)
1.4 Analyse af udviklingen i algebiomasse i samfundsstruktur-eksperimentet
Data indeholder målt klorofyl – fra 12 render udsat for 2 forskellige koncentrationer af pesticid og kontrol. Klorofyl er målt til dag 5, 8 og 10.
Statistisk design:
Behandling: 2 doser af pesticid og kontrol.
Replikater: Render
Respons: klorofyl – der er gentagne målinger på samme rende
Statistisk metode: Repeated mixed effekt model med behandling som fixed effekt og render som random effekt og dag som repeated. Parvise test udføres ved brug af lsmeans og t-test. Der anvendes unstructered varians.
Version 1.0 Marts 2006 • © Miljøstyrelsen.