Bekæmpelsesmiddelforskning fra Miljøstyrelsen Nr. 121, 2008
Sprøjtepraksis i sædskifter med og uden glyphosattolerante afgrøder
Indholdsfortegnelse
- 1.1 Herbicidtolerante afgrøder
- 1.2 Landmænds sprøjteadfærd
- 1.3 Undersøgelsens formål
- 1.4 Arbejdshypoteser
- 1.5 Valg af afgrøder
- 1.6 Valg af plantearter
- 1.7 Valg af dyrkningsscenarier
Forord
Projektet Herbicidtolerante GM-afgrøder: Langtidseffekter af sprøjtestrategier på flora og fauna i mark og tilstødende biotoper er forløbet over 3 år 2004-2007. Den oprindelige projektleder, Kathrine H. Madsen, blev på grund af jobskifte afløst af Niels Holst midtvejs i projektet.
Projektet har bygget på et tværfagligt samarbejde, idet der har indgået forskning med baggrund i såvel naturvidenskab (biologi og agronomi) som humaniora (sociologi og filosofi).
Projektet har resulteret i to rapporter: Den første fokuserede på den sociologiske ramme for GM-afgrøder, Miljøvenlige genmodificerede afgrøder? Vil landmændene have dem, og vil de blive brugt til gavn for naturen? (Bekæmpelsesmiddelforskning fra Miljøstyrelsen, 112, 2007) skrevet af Jesper Lassen m.fl. Den anden, nærværende rapport, fokuserer på projektets biologiske og agronomiske aspekter men inddrager resultaterne fra den første rapport til definition af relevante problemstillinger samt perspektivering af resultaterne.
Vi takker de skiftende medlemmer af Miljøstyrelsens Følgegruppe for råd og vejledning undervejs. Der rettes en særlig tak til Claus J. Christensen, Claus Hansen, Jens E. Jensen, Helle Ø. Nielsen, Nis Schmidt, Jens C. Sørensen og Sonja Graugaard for gennemlæsning og kommentering af første rapportudkast.
Sammenfatning
En indførelse af genetisk modificerede afgrøder med tolerance over for glyphosat vil sandsynligvis ændre herbicidanvendelsen i dansk planteavl i form af ændrede valg af midler og sprøjtetidspunkter. I så fald vil forholdene for flora og fauna i mark og hegn blive ændret som følge af den ændrede herbicidpåvirkning. I dette projekt undersøgte vi, hvorledes indførelsen af glyphosattolerante (GT) afgrøder, vinterraps, majs og roer, vil påvirke floraen i mark og hegn, idet det forventes, at den senere sprøjtning, som GT-afgrøder giver mulighed for, kan have afledte positive effekter på floraen.
Forsøgsarbejdet udført i mark og udyrkede habitater blev suppleret med matematisk modellering til forudsigelse af langtidseffekten af forskellige dyrkningsscenarier. En sociologisk undersøgelse tidligt i projektet1 dannede grundlag for at opstille en række dyrkningsscenarier.
Vi konkluderer, at hvis blot GT-afgrøder dyrkes i sædskifte med ikke-GT-afgrøder, så vil der gennem årene ikke opstå problemer med udbrud af særligt glyphosattolerante ukrudtsarter i marken eller dominans af glyphosattolerante arter i hegnet. Danske landmænd ville da også med deres nuværende viden og holdninger sandsynligvis vælge at kombinere glyphosat med andre sprøjtemidler under dyrkningen af GT-afgrøder, netop for at forhindre udviklingen af glyphosattolerant ukrudt.
Hvis man ønsker at forøge ukrudtsdækket i forsommeren gennem en forsinkelse af herbicidbehandlingen, så vil det i GT-afgrøder være muligt at gøre dette med en mindre forøgelse af behandlingsindeks end muligt i konventionelle afgrøder. Men i konkurrencesvage afgrøder som majs og roer kræves et større forsøgsarbejde for at kortlægge de præcise muligheder for at udsætte sprøjtningen i herbicidtolerante sorter uden, at det går ud over dyrkningssikkerheden.
Mere ukrudt i marken kan give flere insekter, hvilket potentielt også kan have en positiv effekt længere op i fødekæden. Vi kan imidlertid ikke forudsige mængden af insekter blot ud fra ukrudtsmængden.
Summary
The introduction of genetically-modified, glyphosat-tolerant crops would likely alter the pattern of pesticide use in Danish agriculture, both in terms of herbicide choice and timing of treatments. This would change the living conditions for the flora and fauna in fields and field edges because of the change in herbicide exposure. In this project we investigated how the introduction of glyphosat-tolerant (GT) crops, winter oilseed rape, maize and beets, could affect the flora in fields and field edges. It was hypothesised that the delayed spraying, that GT crops make possible, can have positive effects on the flora.
Experimental work in fields and uncultivated habitats was supported by mathematical modelling to predict the long-term effect of different cultivation scenarios. A sociological investigation early on in the project1 provided a basis to define these scenarios.
We conclude that, if just GT crops are grown in rotation with non-GT crops then this will prevent the evolution of glyphosat-tolerant weeds in the fields and of dominance by glyphosat-tolerant species in the field edges. But in fact Danish farmers are, with their current knowledge and attitude, most likely to combine glyphosate with other herbicides when growing GT crops, exactly to prevent the evolution of glyphosate-tolerant weeds.
If the goal is to increase weed coverage in the early summer then GT crops will allow this at a lower increase in the treatment frequency index than conventional herbicides. However, in competitively weak crops, like maize and beet, much experimental work will be needed to define more precisely the possibilities for postponing the spraying in GT cultivars without compromising yields.
An increased density of weeds in the field may increase the density of insects too – with derived, potentially positive effects higher up in the food chain. But it is not possible to predict the density of insects just from the density of weeds.
1 Indledning
- 1.1 Herbicidtolerante afgrøder
- 1.2 Landmænds sprøjteadfærd
- 1.3 Undersøgelsens formål
- 1.4 Arbejdshypoteser
- 1.5 Valg af afgrøder
- 1.6 Valg af plantearter
- 1.7 Valg af dyrkningsscenarier
1.1 Herbicidtolerante afgrøder
På verdensplan er herbicidtolerance den mest udbredte egenskab i genmodificerede afgrøder. I 2007 dyrkedes således herbicidtolerante afgrøder på 75 mio. ha, insekticidresistente afgrøder udgjorde 15 mio. ha, mens afgrøder med begge disse egenskaber blev dyrket på yderligere 15 mio. ha (International Service for the Acquisition of Agri-biotech Applications, www.isaaa.org). I EU og dermed i Danmark er det aktuelt kun tilladt landmænd at dyrke én genmodificeret afgrøde, en majssort med indbygget insektresistens. Blandt herbicidtolerante afgrøder er glyphosattolerance (GT) den mest udbredte egenskab². GT-afgrøder har den umiddelbare dyrkningsmæssige fordel, at de kan sprøjtes med det meget effektive og billige ukrudtsmiddel, glyphosat, uden at tage skade. Dette giver flere muligheder for at tilrettelægge ukrudtsbekæmpelsen.
Over for landbrugets praktiske og økonomiske afvejninger af genteknologiens nytte står befolkningens skeptiske holdning. Teknologien ses i en større sammenhæng og kun samfundsmæssigt nyttige anvendelser (f.eks. til ny medicin) anses for at være acceptable3,4. Selvom vi i dette projekt fokuserer på de landbrugsfaglige og miljømæssige aspekter af GT-afgrøder, vil den endelige beslutning om dyrkningen af disse afgrøder bero på en bredere debat, som går videre end konkrete overvejelser af GT-afgrøders nytteværdi og effekter på biodiversitet.
Glyphosat er i modsætning til hovedparten af de konventionelle herbicider ikke selektivt. Alle plantearter er mere eller mindre følsomme over for glyphosat5,6. Blandt de herbicidtolerante afgrøder kan de glyphosattolerante afgrøder derfor forventes at have den største effekt på agerlandets plantesamfund både i marken og i tilstødende biotoper.
Den største herbicideffekt opnås, når planterne er små, derfor sprøjtes der som regel tidligt i vækstsæsonen eventuelt fulgt op af senere sprøjtninger efter behov. Behovet afgøres primært af, om afgrøden har kunnet udkonkurrere det resterende ukrudt. Glyphosat er imidlertid i modsætning til andre herbicider effektiv over for de fleste ukrudtsarter, også når de er blevet store6,7. Det giver mulighed for i GT-afgrøder at lægge den første sprøjtning senere end i konventionelle afgrøder. Dermed giver GT-afgrøder både landmanden større frihed med hensyn til sprøjtetidspunkt og større dyrkningssikkerhed, da en mislykket sprøjtning kan følges op af en senere glyphosatsprøjtning.
Vil de dyrkningsmæssige fordele ved GT-afgrøder være fristende for landmanden? I givet fald vil indførelse af disse nye afgrøder føre til en større anvendelse af glyphosat, idet anvendelse af glyphosat i de nuværende konventionelle afgrøder er stærkt begrænset. En sådan ændret herbicidanvendelse vil kunne påvirke agerlandets flora både negativt (hvis det medfører en mere effektiv bekæmpelse) og positivt (hvis sprøjtningen falder senere end i konventionelle afgrøder). Da vegetationen tjener som næringskilde for jordbundsorganismer, insekter, fugle m.m., kan man forestille sig ganske omfattende, afledte effekter på agerlandets fauna afhængigt af, hvorledes GT-afgrøderne indpasses i sædskiftet.
Man kan umiddelbart forestille sig to modsatrettede konsekvenser af den forøgede fleksibilitet, som GT-afgrøder giver. På den ene side vil en udsætning af den første sprøjtning føre til, at en større mængde ukrudt står i længere tid i marken. På den anden side kan en fuld udnyttelse af glyphosats effektivitet føre til, at ukrudtet sjældent når at udvikle sig frem til frøsætning, hvorved ukrudtsbestanden vil blive reduceret på længere sigt. Dette er blevet påvist både i GT-roer8 og GT-majs9. GT-afgrøder giver således både mulighed for at forøge og at reducere biodiversiteten
De første danske undersøgelser af ændringer i flora og fauna ved dyrkning af herbicidtolerante afgrøder blev gennemført af Danmarks Miljøundersøgelser (DMU) i GT-roer10. I sammenligning med konventionelle roer var der mere ukrudt i juni måned, når den første sprøjtning blev foretaget senere end i konventionelle roer, men senere på sommeren kunne der være mindre (på grund af glyphosats større effektivitet)10-12. Tætheden af hvirvelløse dyr i marken blev påvirket af, om der dyrkedes GT eller konventionelle roer. I det første studium10 varierede effekten mellem forskellige dyregrupper og fra sted til sted (seks lokaliteter), så der ikke var nogen generel positiv eller negativ effekt på faunaen af at dyrke GT-roer. I det andet studium, som blev udført på én lokalitet, var der konsekvent en positiv effekt af den sene sprøjtning i GT-roer på tætheden af hvirvelløse dyr11,12. Effekten af sprøjtetidspunktet varierede mellem de forskellige taksonomiske grupper, men der sås positive effekter på både herbivore og ikke-herbivore grupper, hvilket formentlig skyldes, at ukrudtet ikke alene er en fødekilde men også har en for dyrene gavnlig effekt på f.eks. mikroklima og habitatstruktur11. Samlet set viser disse undersøgelser, at GT-roer kan have en umiddelbar positiv effekt på biodiversiteten, men at langtidseffekten bør undersøges nærmere.
De britiske Farm Scale Evaluations var særdeles omfattende og sammenlignede økosystemeffekter af konventionelle og herbicidtolerante vårafgrøder (raps, majs og roer)13. Hver afgrøde blev testet på 60-70 lokaliteter over tre år. I raps og roer fandtes som i de danske studier, at den sene sprøjtning i GT-afgrøden gav en forøget ukrudtsbiomasse indtil sprøjtning, men en mindre biomasse senere hen i forhold til den konventionelle afgrøde. I majs var biomassen høj både før og efter sprøjtning14. Hvis man betragtede effekten på de enkelte ukrudtsarter, fremgik det dog, at nogle var positivt påvirkede af sprøjtepraksis i GT-afgrøden og andre negativt15. Effekten på de forskellige taksonomiske grupper af overfladeaktive hvirvelløse dyr fordelte sig ligeligt på positive og negative effekter16. Ved at dele de hvirvelløse dyr op i funktionelle grupper fremkom et langt klarere billede. I de afgrøder og på de tidspunkter, hvor ukrudtsbiomassen var høj, havde det positive effekter på dyr højere oppe i fødekæden17. Hovedeffekten på flora og hvirvelløse dyr fandtes i selve marken, medens lignende mindre effekter fandtes i tilstødende biotoper18. Hvorvidt dyr findes i marken skal fortolkes i sammenhæng med deres levevis og mobilitet: Blomstersøgende insekter som sommerfugle og bier påvirkedes negativt af GT-afgrøder, men om dette har nogen væsentlig effekt på bestandene i agerlandet kommer an på, hvor vigtigt ukrudt i marken er som fødekilde19.
Sammenfattende om de danske og engelske undersøgelser kan man sige, at det ikke er overraskende, at en reduktion af det nederste led i fødekæden (ukrudt) har afledte effekter højere oppe. Herbiciders afledte negative effekter på insekter og fugle er således demonstreret flere gange20. På grund af økosystemers kompleksitet overrasker det heller ikke, at sådanne effekter ikke altid kan detekteres, eller at sammenhængen sommetider findes at være modsat. De tydeligste effekter fandtes, hvor sprøjtetidspunktet for GT-afgrøden afveg mest markant fra den konventionelle afgrøde. Størrelsen af dette herbicidfrie tidsvindue er vigtig for, hvor stor den umiddelbart positive effekt på flora og fauna bliver12. En positiv effekt afhænger således af, at landmanden i begyndelsen af vækstsæsonen er villig til at acceptere en større mængde ukrudt i en GT-afgrøde end i en tilsvarende konventionel afgrøde.
Den forsinkede herbicidsprøjtning, som er mulig i GT-afgrøder, har en umiddelbart positiv virkning på floraen, men på grund af glyphosats høje effektivitet over for mange arter, så kan effekten på lang sigt godt vise sig at være negativ på grund af mindsket frøsætning. Den effektive bekæmpelse i GT-afgrøder forårsager en mindre overlevende ukrudtsbiomasse, og det er velkendt, at der er en meget stabil sammenhæng mellem ukrudtets frøproduktion (y) og biomasse (x), y=axb, som ikke eller kun i ringe grad påvirkes af årsagen til, at plantemassen er lille eller stor, det være sig konkurrence med forskellige afgrøder21 eller behandling med forskellige herbicider22. Ydermere kan man forvente forskydninger i floraen på grund af plantearternes forskellige følsomhed over for herbiciderne anvendt i konventionelle afgrøder i forhold til følsomheden for glyphosat.
Floraen i hegnets fodpose (det vil sige den urte- og græsdominerede vegetation, som findes i det upløjede område mellem træer og buske og marken) påvirkes meget af markdriften, især brugen af gødning og herbicider23. Artsdiversiteten er lavere, og andelen af mosser og naturlige floraelementer er lavere end i tilsvarende usprøjtede hegn ved samme næringsstofforhold24. Vegetationen, der overvejende er flerårig, afspejler således den måde marken drives på. Afdrift af glyphosat forekommer i konventionelle afgrøder som oftest, hvor midlet bruges til nedvisning eller til kvikbekæmpelse, det vil sige fra sommer til efterår. I en GT-afgrøde vil glyphosat blive anvendt tidligere på året, og afdriften herfra må forventes at have større konsekvenser på floraen i fodposen, da planterne endnu ikke har sat frø. I de britiske Farm Scale Evaluations påvirkede herbiciderne floraen i markkanten. I kanten af vårraps- og roemarker var der en negativ påvirkning af floraen ved dyrkning af GT-sorter sammenlignet med de tilsvarende konventionelle afgrøder, mens der i kanten af majsmarker var en positiv virkning18.
Hvorledes de naturmæssige sideeffekter af at dyrke GT-afgrøder udmøntes vil afhænge af, hvorledes landmanden vælger at udnytte de dyrkningsmæssige muligheder ved GT-afgrøder. En undersøgelse af GT-afgrødedyrkningens konsekvenser på Danmarks flora og fauna må derfor inddrage både sociologiske og økologiske aspekter.
1.2 Landmænds sprøjteadfærd
Det sociologiske studium i landmænds sprøjteadfærd, som var en del af dette projekt, er beskrevet i en særskilt rapport1. Her gennemgår vi de resultater, som har særlig betydning for det agronomisk-biologiske delstudium, beskrevet i nærværende rapport.
En forudsætning for at opnå de eventuelle miljømæssige fordele ved GT-afgrøder er, at landmændene ændrer deres adfærd henimod færre og senere sprøjtninger. Et vigtigt formål med interviewene i det sociologiske studium var derfor at undersøge, om landmænd faktisk vil ændre deres adfærd i den retning.
Et argument, der taler for den ønskede adfærdsændring, er driftsøkonomien. Det synes at give god mening ud fra en tankegang om at optimere driften at følge sprøjteplaner med brug af kun et enkelt herbicid samt færre og senere sprøjtninger. At der så oven i købet er fordele for miljøet burde tale yderligere for, at landmændene skulle ændre deres adfærd.
I lyset af interviewene må vi dog konkludere, at landmænd med den viden og de holdninger de har i dag formodentlig kun i meget begrænset omfang vil ændre deres adfærd i den ønskede retning. Dette beror på flere forhold:
§ Landmændene opfatter det slet ikke som en fordel for miljøet, at man lader ukrudtet vokse sig stort, inden man sprøjter. For landmændene drejer miljøbeskyttelse sig først og fremmest om at beskytte grundvandet mod forurening.
§ Landmændene optimerer økonomisk ved at følge ”tommelfingerregler”, som via tradition og erfaring har vist sig nyttige; og en vigtig tommelfingerregel for de interviewede landmænd er, at rene (dvs. ukrudtsfrie) marker er en god ting. Reglen er indarbejdet i en grad, så rene marker også for landmændene er en æstetisk kvalitet og ses som tegn på godt landmandskab.
§ I forhold til driftsøkonomien synes landmændene også at prioritere produktionssikkerhed – her at de undgår at ende i store ukrudtsproblemer – højere end en langsigtet økonomisk optimering.
§ Landmændene er skeptiske over for helt at opgive de traditionelle pesticider. Med deres nuværende viden og holdninger vil de i forbindelse med dyrkning af GT-afgrøder vælge at kombinere glyphosat med et eller flere andre sprøjtemidler.
Landmændene vil dog efter alt at dømme sprøjte færre gange samt lægge den første sprøjtning en smule senere i forhold til, hvad der med de nuværende systemer er normen.
1.3 Undersøgelsens formål
Undersøgelsen havde til formål at undersøge konsekvenserne for flora og fauna i mark og hegn ved at inddrage GT-afgrøder i sædskiftet, idet indførelsen af sådanne afgrøder ville forårsage en ændring i landmandens sprøjtepraksis.
Korttidseffekterne, det vil sige inden for en dyrkningssæson, blev belyst direkte ved forsøg i væksthus, mark og naturlige biotoper, mens langtidseffekterne blev beregnet ved hjælp af modeller, som inddrog konkurrenceforhold og populationsdynamik for ukrudt og anden flora både i mark og fodpose. Konklusionerne samles via en række scenarier, som sammenligner typiske sædskifter bestående af konventionelle afgrøder alene eller i kombination med GT-afgrøder. I scenarier med GT-afgrøder sammenlignes endvidere de forskellige sprøjtestrategier, som landmanden kunne tænkes at anvende.
1.4 Arbejdshypoteser
Overordnet arbejdede vi ud fra hypotesen, at
§ den sprøjtepraksis, som dyrkningen af GT-afgrøder vil afstedkomme, kan på afgørende vis ændre levevilkårene for flora og fauna i mark og hegn.
På kort sigt
§ vil anvendelsen af glyphosat i GT-afgrøder påvirke floraens frøproduktion i mark og hegn markant i forhold til traditionelle herbicider;
§ vil en sen glyphosatsprøjtning forbedre levevilkårene i forsommeren for flora og fauna i marken i forhold til traditionel sprøjtepraksis.
På lang sigt
- vil ændringen i floraens konkurrenceforhold, forårsaget af den ændrede herbicidanvendelse. medføre, at plantesamfundene i mark og hegn ændres markant, muligvis med afledte effekter på faunaen.
1.5 Valg af afgrøder
Vi har valgt at sammenligne dyrkningen af GT og konventionelle typer af tre afgrøder: Vinterraps, majs og sukkerroer. Af disse GT-afgrøder betragter vi kun GT-majs som en sandsynlig fremtidig afgrøde i Danmark. De øvrige afgrøder inddrages for at have et bedre sammenligningsgrundlag for de opnåede resultater for GT vs. konventionel majs.
Ifølge Danmarks Statistik for perioden 2002-2007 er dyrkningen af sukkerroer faldet jævnt fra 60 til 40 tusinde ha, mens fodermajs steg fra 100 til 140 og vinterraps fra 80 til 180 tusinde hektar. Vårraps dyrkes knapt mere (1 ha i 2007). Tallene skal sammenholdes med det samlede dyrkede areal på 2.7 mio. ha i 2007, hvoraf kornafgrøder udgjorde ca. halvdelen.
1.5.1 Vinterraps
Raps er meget villig til at krydse med andre rapsplanter, vilde slægtninge og beslægtede kulturafgrøder, hvilket afspejles i loven om sameksistens, som gør dyrkning af GM-raps i Danmark noget nær umulig. Men globalt set er GM-raps en vigtig afgrøde:
- I 2006 udgjorde GM-raps 5% af det samlede areal i hele verden dyrket med GM-afgrøder25.
1.5.2 Fodermajs
GM-majs er allerede godkendt til dyrkning i Danmark. Der er stor interesse for majs til bioenergi, og majs som GM-afgrøde er dyrkningsmæssigt umiddelbart let at acceptere, da der i Danmark hverken er problemer med overvintrende frø eller krydsning.
§ GT-majs er et eksempel på, at sprøjtningen kan udsættes væsentligt i forhold til den nuværende almindelige praksis. Den kan derfor hjælpe til at undersøge de dyrknings- og naturmæssige konsekvenser af sprøjtestrategier basereret på GT.
1.5.3 Sukkerroer
Ligesom majs er sukkerroer godkendt til dyrkning i Danmark. Problemer med krydsninger begrænser sig til én vild slægtning (strandbeden), og der er ikke problemer med overvintrende frø. Selvom markedet for foder- og sukkerroer forventes at være begrænset26, er der alligevel gode grunde til at inddrage dem i undersøgelsen:
§ Ligesom for majs giver GT-sukkerroer større frihed i valg af sprøjtetidspunkter end konventionelle sukkerroer, og de kan ligeledes belyse effekten af sprøjtestrategier baseret på GT.
1.6 Valg af plantearter
Ukrudt er i denne sammenhæng floraen i marken, af hvilke vi har valgt fem enårige arter: Agersennep, burresnerre, fuglegræs, hvidmelet gåsefod og lugtløs kamille (videnskabelige navne på disse og andre plantearter kan ses i Bilag A). De er valgt bredt og repræsenterer således fem forskellige plantefamilier samt forskellige livsstrategier. F.eks. spirer agersennep og hvidmelet gåsefod primært om foråret, de andre både forår og efterår27. Vi valgte en starttæthed i scenarierne på 100 frø pr. m² af hver art, fordelt jævnt i pløjelaget (til 20 cm dybde) ved scenariets begyndelse det vil sige den 1. januar år 1. Denne tæthed er tilstrækkelig til at udgøre et truende ukrudtsproblem ved mangelfuld bekæmpelse.
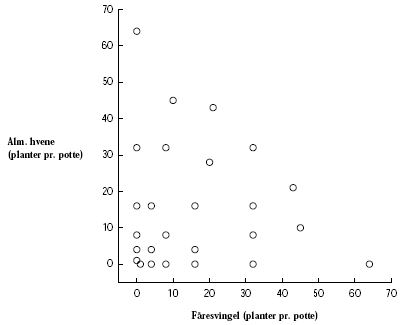
Som repræsentanter for den vilde flora i markens tilstødende arealer er valgt en række af både græsser og tokimbladede, i alt otte arter: Fåresvingel, rejnfan, alm. hvene, stor nælde, alm. kørvel, agertidsel, mælkebøtte og kvik. Disse er valgt for at repræsentere forskellige livsstrategier bl.a. er agertidsel og kvik også alvorlige ukrudtsarter i marken.
Det er disse 13 arter, der fokuseres på i dette projekt, både eksperimentelt og i scenariestudierne. I forsøg, som skal afdække generelle biologiske sammenhænge, er der dog i visse tilfælde af praktiske årsager valgt at arbejde med andre plantearter.
1.7 Valg af dyrkningsscenarier
Scenarierne er bygget op om fire sædskifter, hvor GT- og ikke-GT-typer af vinterraps, majs og roer kan indgå i sædskifte med konventionelle afgrøder. I praksis vil man ikke fortløbende dyrke GT-afgrøder, da ensidig brug af glyphosat vil fremprovokere glyphosatresistent ukrudt. Vi har valgt fire forskellige sædskifter:
- Vinterraps-vårbyg-vinterbyg.
- Majs-vårbyg.
- Vedvarende majs.
- Roer-vårbyg-vinterhvede-vinterhvede.
I det første sædskifte vil man i praksis ofte indskyde vinterhvede mellem vinterraps og vårbyg, men det valgte sædskifte giver mulighed for en mere ekstrem effekt af GT-afgrøden (vinterraps), da den her dyrkes hvert tredje og ikke hvert fjerde år.
Dyrkningsscenarierne defineres i form af dyrkningskalendere for pløjning, såning, herbicidbehandlinger og høst. I realiteten vil dyrkningskalenderen variere afhængigt af vejrliget, men i simuleringerne arbejdes med dansk standardvejr (30 års gennemsnit), og derfor afspejler dyrkningskalenderne nedenfor de typiske datoer for de forskellige markoperationer.
I ikke-GT-afgrøder er herbicider og dosis valgt ud fra Planteværn Onlines anbefalinger, som er givet ud fra afgrøde, ukrudtsarterne og deres vækststadium (www.planteinfo.dk). I GT-afgrøder er glyhosatdosis skønnet ud fra et effektkrav svarende til Planteværn Onlines anbefalinger i kombination med arternes dosisresponskurver7.
GT-afgrøderne indgår i sædskifte med korn som vår- eller vintersæd. I alle tilfælde sprøjtes kornet i foråret med ioxynil+bromoxynil. I vintersæd vil man ofte indlede med en sprøjtning efterår, opfulgt af en forårssprøjtning efter behov. Vi nøjedes med forårssprøjtningen, da indledende simuleringsstudier viste, at dette gav en tilstrækkelig bekæmpelse med den valgte ukrudtsflora.
Sædskiftet i scenarium 1 (tabel 1.1) findes i to variationer med hensyn til rapsen:
a. Med konventionel raps og herbicidbehandlinger med jordmiddel og opfølgende sprøjtning efterår samt en sprøjtning mod kamille forår.
b. Med GT-raps og glyphosatbehandling efterår.
Sædskifterne i scenarium 2 (tabel 1.2) og 3 (tabel 1.3) findes begge i fire variationer med hensyn til majsen:
a. Splitsprøjtning med MaisTer.
b. Splitsprøjtning med glyphosat.
c. Normaltidig glyphosatbehandling.
d. Sen glyphosatbehandling.
Sædskiftet i scenarium 4 (tabel 1.4) findes i to variationer med hensyn til roerne:
a. Med konventionelle roer og fire traditionelle herbicidbehandlinger april-maj.
b. Med GT-roer og to glyphosatbehandlinger maj-juni.
I majs anbefales det, at anvende MaisTer i kombination med et andet herbicid for at forebygge udviklingen af resistent ukrudt. Risikoen er særlig stor, fordi majs ofte dyrkes på samme areal år efter år. For at begrænse antallet af scenarier nøjedes vi med MaisTer.
I roer anvendes traditionelt herbicidblandinger. Vi udelod ethofumesat i blandingerne, da dette primært iblandes til bekæmpelse af pileurter, som ikke indgik i scenarierne.
I alt arbejdes der således med 2+4+4+2=12 scenarier. Af disse scenarier er det kun de forskellige sprøjtestrategier i majs, som vi afprøvede i praksis (2.1.2). Sædskifterne og deres variationer blev belyst gennem modelstudier i mark (2.1.7) og hegn (2.2.5).
Tabel 1.1. Dyrkningskalender for scenarium 1.
| Afgrøder og behandlinger | Dato | |
| Vinterraps (år 1) Pløjning Såning (a) Sprøjtning i konventionel raps 90 g clomazon pr. ha 500 g propyzamid pr. ha 80 g clopyralid pr. ha (b) Sprøjtning i GT-raps 540 g glyphosat pr. ha Høst |
14.8.1 15.8.1 15.8.1 5.11.1 1.5.2 25.9.1 20.7.2 |
|
| Vårbyg (år 2) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
9.4.2 10.4.2 24.4.2 15.8.2 |
|
| Vinterbyg (år 3) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
17.9.3 18.9.3 24.4.3 1.8.4 |
|
Tabel 1.2. Dyrkningskalender for scenarium 2.
| Afgrøder og behandlinger | Dato | |
| Majs (år 1) Pløjning Såning (a) Splitsprøjtning i konventionel majs 100 g MaisTer pr. ha 50 g MaisTer pr. ha (b) Splitsprøjtning i GT-majs 360 g glyphosat pr. ha 360 g glyphosat pr. ha (c) Normaltidig sprøjtning i GT-majs 720 g glyphosat pr. ha (d) Sen sprøjtning i GT-majs 720 g glyphosat pr. ha Høst |
1.11.0 1.5.1 25.5.1 8.6.1 25.5.1 8.6.1 8.6.1 20.6.1 19.9.1 |
|
| Vårbyg (år 2) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
9.4.2 10.4.2 24.4.2 15.8.2 |
|
Tabel 1.3. Dyrkningskalender for scenarium 3.
| Afgrøder og behandlinger | Dato | |
| Majs (vedvarende) Pløjning Såning (a) Splitsprøjtning i konventionel majs 100 g MaisTer pr. ha 50 g MaisTer pr. ha (b) Splitsprøjtning i GT-majs 360 g glyphosat pr. ha 360 g glyphosat pr. ha (c) Normaltidig sprøjtning i GT-majs 720 g glyphosat pr. ha (d) Sen sprøjtning i GT-majs 720 g glyphosat pr. ha Høst |
1.11.0 1.5.1 25.5.1 8.6.1 25.5.1 8.6.1 8.6.1 20.6.1 19.9.1 |
|
Tabel 1.4. Dyrkningskalender for scenarium 4.
| Afgrøder og behandlinger | Dato | |
| Sukkerroer (år 1) Pløjning Såning (a) Sprøjtning i konventionelle roer 240 g phenmedipham pr. ha 160 g phenmedipham + 5 g triflusulfuron- methyl pr. ha 160 g phenmedipham + 5 g triflusulfuron- methyl pr. ha 240 g phenmedipham + 1 L metamitron pr. ha. (2) Sprøjtning i GT-roer 270 g glyphosat pr. ha 270 g glyphosat pr. ha Høst |
1.11.0 21.4.1 23.4.1 1.5.1 11.5.1 25.5.1 17.5.1 5.6.1 28.10.1 |
|
| Vårbyg (år 2) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
9.4.2 10.4.2 24.4.2 15.8.2 |
|
| Vinterhvede (år 3) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
17.9.2 18.9.2 24.4.2 1.8.3 |
|
| Vinterhvede (år 4) Pløjning Såning Herbicidsprøjtning 76 g ioxynil + 76 g bromoxynil pr. ha Høst |
17.9.3 18.9.3 24.4.3 1.8.4 |
|
Valg af modelleringsmetode
Indflydelsen af dyrkningsscenarierne på floraen i mark og hegn blev beregnet ved hjælp af simuleringsmodeller baseret på planternes basale biologiske egenskaber. For marken anvendtes modellen FieldWeeds og for hegnet modellen FieldEco. Begge modeller forefandtes inden dette projekt gik i gang, men blev videreudviklet for at kunne beskrive dynamikken af netop de arter, som indgik i dette projekt. I et tidligere miljøstyrelsesprojekt28,29 om frøpuljens dynamik anvendtes en statistisk beskrivelse baseret på et omfattende datamateriale. En sådan metode og dog mere bagudskuende end fremadskuende, og i forbindelse med scenarierne har vi netop brug for den forudsigelseskraft, som simuleringsmodeller har.
FieldWeeds er en frøpuljemodel, som inkluderer effekterne af markoperation-erne i løbet af sædskiftet. Da alle de fem udvalgte ukrudtsarter er enårige, ansås denne model for passende. Som anbefalet af andre ukrudtsmodellører, holder modellen styr på ukrudtets demografi (antal planter)30 og biomasse31. Manglende viden om ukrudtets fremspiring (som afhænger af vejret på kompliceret vis) er tidligere udpeget som væsentligt usikkerhedsmoment i frøpuljemodeller32,33, og det indebærer også for FieldWeeds en iboende usikkerhed. For at imødegå dette opererer FieldWeeds med en fast fremspiringskalender, som repræsenterer den typiske fremspiring gennem et år.
I modsætning til FieldWeeds er FieldEco en mere detaljeret, fysiologisk model af planters vækst og konkurrence. Den blev valgt til simulering af de otte hegnsarter, der alle er flerårige. Da flerårige planter, i modsætning til enårige, viderefører reserver fra et år til det næste, kræver de et nøjere regnskab med plantens kulstof og kvælstof. I hegnet er konkurrenceforholdene mere indviklede end i marken, da der dér er mange arter i indbyrdes konkurrence og ikke som i marken én dominerende art (afgrøden), som ukrudtet konkurrerer med. Økofysiologiske modeller som FieldEco anbefales netop til at modellere plantekonkurrence33.
Ukrudtsmodeller vil altid være usikre på grund af grænsebetingelser, som er mere eller mindre uden for menneskelig kontrol (f.eks. vejr, dyrkningspraksis, startfrøbanken), og fordi modeller nødvendigvis altid er simplere end virkeligheden. Denne usikkerhed fører til kravet om validering. Valideringsbegrebet kan forvirre meget, og man bør tænke grundigt over dets rødder og rationale34, inden en validering sættes i værk. Vi bruger udtrykket ”validering” for det at sammenligne modeloutput med uafhængige feltdata.
Inden valideringen blev FieldEco, men ikke FieldWeeds, kalibreret. Det betyder, at man efter at have bygget modellen sammenligner den med uafhængige feltdata. Man får da modellen til at passe med disse feltdata ved at indstille på modellens frie parametre. Frie parametre er enten parametre, som man har dårligt belæg for værdien af, eller hvis værdi forventes at variere fra sted til sted. Efter kalibreringen følger da valideringen med et nyt, uafhængigt datasæt.
Validering af populationsdynamiske ukrudtsmodeller, som de to anvendt i dette projekt, er meget datakrævende og dyrt. I et nyligt review35, som omfattede 134 artikler om sådanne modeller, fandtes kun 3 modeller at være grundigt validerede. Samme review konkluderede da også, at ukrudtsmodellerne indtil videre er bedst egnede som forskningsværktøjer. De er gode til at formulere og teste hypoteser om ukrudtets dynamik, mens de er mindre velegnede til at give konkret vejledning. Ikke desto mindre er det med dette formål, at modellerne anvendes i nærværende projekt. Med bevidstheden om den store usikkerhed i modellernes beregninger anvender vi derfor modellerne til at sammenligne ukrudtsarternes dynamik i de forskellige scenarier. Man kan med større sikkerhed forudsige, hvorvidt én art vil blive begunstiget og en anden hæmmet i et scenarium, end man kan sige præcist, hvorledes dynamikken vil forløbe. Modellerne blev udviklet og anvendt med dette for øje. Endvidere blev alle projektets konklusioner (kapitel 5) draget med belæg i empiriske data, i nogle tilfælde med støtte i modellernes forudsigelser, men aldrig med belæg kun i modellerne.
2 Metoder
Konsekvenserne af at indføre herbicidresistente afgrøder i dansk landbrug afhænger af den praksis de vil indgå i. Hvor fleksible landmænd vil være i deres dyrkningspraksis, hvis de inddrager herbicidresistente afgrøder, blev undersøgt i projektets sociologiske del, som beskrevet i den særskilte rapport1.
For at kunne beregne langtidseffekterne i marken udførte vi dosisresponsforsøg med herbicider anvendt på udvalgte ukrudtsarter (2.1.1) samt markforsøg med glyphosatsprøjtning i majs (2.1.2). Desuden målte vi ukrudtets vækst og frøproduktion i majs (2.1.3), vårbyg (2.1.4) og vinterraps (2.1.5). Mens resultaterne fra disse forsøg kunne anvendes til direkte modellering af ukrudtets populationsdynamik i marken, måtte vi beskrive dynamikken i den tilknyttede fauna mere indirekte. På basis af markforsøg med forskellige afgrøder beregnede vi den statistiske sammenhæng mellem biomassen af planter (afgrøde og ukrudt) og leddyr (insekter, edderkopper m.v.) (2.1.6).
Forsøgsarbejdet i marken gav kun et mål for effekterne over en enkelt dyrkningssæson. For at udvide fortolkningsmulighederne sammenfattede vi i en matematisk model (FieldWeeds) resultaterne fra vore forsøg med resultater fra tidligere undersøgelser i Danmark og udlandet. Dette muliggjorde simulering af langtidseffekterne i marken i de forskellige dyrkningsscenarier (2.1.7).
I biotoper, som støder op til marken, påvirkes floraen af herbicidernes afdrift fra marksprøjten. Dette kan medføre at nogle planter dør, mens andre påvirkes negativt i deres vækst og formering (subletale effekter). De subletale effekter udmøntes både direkte og ved ændring i planternes indbyrdes konkurrenceforhold.
Ligesom for markukrudtet udførte vi dosisresponsforsøg med herbicider anvendt på udvalgte arter fra hegnets fodpose (2.2.1), hvor vi især så på de vækststandsninger af midlertidig karakter, som optræder ved subletale doser (2.2.2). I et nyskabende forsøg beregnede vi dosis-respons på en kombination af to arter voksende i konkurrence, hvilket krævede en ny matematisk tilgang (2.2.3).
Et langvarigt studium af et plantesamfunds succession under varierende grad af herbicid- og gødningspåvirkning var blevet etableret under et tidligere projekt. Vi fulgte op på dette i nærværende projekt med målinger af både frøpulje og artsammensætning, således at de langsigtede ændringer som følge af konkurrenceforholdene under disse påvirkninger kunne belyses eksperimentelt (2.2.4). For udvalgte arter fra hegnsfloraen udviklede vi en matematisk model (FieldEco), som muliggjorde simulering af langtidseffekterne på floraen i biotoper stødende op til marker i de forskellige dyrkningsscenarier (2.2.5).
2.1 Herbicideffekter på markens biodiversitet
2.1.1 Ukrudtsbiomasse og frøproduktion
I standardtests gøres planters respons på en herbicidbehandling normalt op som reduktionen i overjordisk biomasse tre uger efter behandling. Mere relevant for den langsigtede effekt af behandlingen er imidlertid effekten på frøsætningen. Specielt for sent høstede afgrøder, hvor der går lang tid fra sidste sprøjtning til høst, er det interessant at undersøge planternes mulighed for genvækst og frøsætning.
For at belyse disse forhold udførte vi forsøg med sort natskygge og hanespore, som begge forekommer som ukrudt i majs. Hanespore kræver i modsætning til de fleste andre græsser ikke vernalisering (kuldepåvirkning) for at producere frø, hvilket er en fordel i forsøgsmæssig sammenhæng. Vi supplerede standardtesten (biomasserespons) med en opgørelse af frøproduktionen på de fuldt udviklede planter.
Frø af sort natskygge og hanespore blev sået i 2 L potter i en blanding af jord, sand og sphagnum (2:1:1 vægtforhold) indeholdende alle nødvendige mikro- og makronæringsstoffer. Der blev anvendt frø fra Forskningscenter Flakkebjergs frøbank. Potterne blev placeret på udendørs borde og blev automatisk vandet 3 gange dagligt. Efter fremspiring blev antallet af planter pr. potte reduceret til 4 i potter til biomassemåling og 2 i potter til frøhøst.
Glyphosat og MaisTer blev udsprøjtet på to forskellige udviklingstrin af hver planteart. Begge plantearter blev behandlet på 4-bladstadiet. Sort natskygge blev desuden behandlet på 8-10-bladstadiet, mens hanespore havde 6-8 blade ved den sene behandling.
Herbiciderne blev udsprøjtet i en væskemængde på ca. 150 L/ha ved hjælp af en pottesprøjte. MaisTer blev udsprøjtet i blanding med 2,68 L/ha MaisOil. Der blev anvendt 6 doseringer af hvert herbicid, og hver behandling blev udført med 3 gentagelser. Doseringerne blev varieret i forhold til plantearternes forventede følsomhed på de forskellige udviklingstrin. Den højeste dosering af glyphosat varierede fra 288 g/ha til 432 g/ha ved de forskellige behandlinger, mens den højeste dosering af MaisTer varierede fra 7,75 til 15,5 g as./ha. Behandlingsindeks kan ses i Bilag B.
Biomassemålingerne blev udført ca. 3 uger efter sprøjtning, hvor frisk- og tørvægt af planterne blev målt. Planter til frøhøst blev placeret i væksthus, og hanespore, som har en meget kraftig vækst, blev omplantet i 10 L spande. Frøene af hanespore modnede meget uens, og frøhøsten strakte sig over 4 måneder, hvor modne frøstande ugentligt blev klippet af og lagt i papirsposer. Bær af sort natskygge blev høstet i november. Bærrene blev fermenteret ved opbevaring i vand i 7 dage for at frigøre frøene fra frøgemmet. Den fermenterede bærmasse blev herefter sigtet under rindende vand, hvorved frøene blev sorteret fra. Frøene af begge plantearter blev tørret, og den samlede vægt af frø pr. potte blev målt. Tusindkornsvægten blev bestemt ud fra optælling og vejning af 3´200 frø fra hver potte.
Visuelle bedømmelser viste, at på sort natskygge med 8-10 blade var doseringerne af begge herbicider for lave, mens doseringerne af MaisTer på hanespore på 4-bladstadiet var for høje. Disse behandlinger blev derfor gentaget i et efterfølgende forsøg. I dette forsøg blev sort natskygge behandlet på 6-8 bladstadiet, og hanespore blev behandlet med MaisTer på 4-bladstadiet. Biomassemålingerne blev udført ca. 3 uger efter sprøjtning (frisk- og tørvægt), og frøhøst blev foretaget i en periode på 3 måneder.
2.1.2 Glyphosatsprøjtning i majs
Forsøget blev gentaget over to vækstsæsoner, 2005 og 2006. Begge år udlagdes 20 parceller á 10´3 m² på Forskningscenter Flakkebjergs jorder. Der blev afprøvet fire sprøjtestrategier med 4 gentagelser i alt 16 parceller (tabel 2.1). Desuden holdtes 4 parceller sprøjtefrie til bedømmelse af ukrudtsfloraen.
2.1.2.1 Etablering
Forsøgsarealet blev pløjet om efteråret og dernæst fældet, gødet med 680 kg/ha 22N-3P-8K, såbedsharvet og tilsået den 12. maj 2005 og den 15. maj 2006. Forsøget blev sået – i 2005 med NAXOS hybridmajs og i 2006 med Banguy hybridmajs – med majssåmaskine i 4-5 cm dybde og med 75 cm rækkeafstand i 4 rækker. Der tilstræbtes et plantetal på 8,5 planter pr. m². Under såningen blev der givet yderligere 160 kg/ha 12N-23P-0K i rækkerne.
Tabel 2.1. Sprøjtestrategier afprøvet i 2005 og 2006. For splitsprøjtning er dosis angivet for hver behandling.
| Strategi | Behandling (pr. ha) | Stadium | 2005 | 2006 | ||||
| MaisTer splitsprøjtning | 2 ´ 15,5 g as. |
Ukrudt med 2-4 blade og 10-14 dage senere | 8. juni og 20. juni |
14. juni og 29. juni |
||||
| Glyphosat- splitsprøjtning | 2 ´ 360 g | Ukrudt med 2-4 blade og 10-14 dage senere | 8. juni og 20. juni |
14. juni og 29. juni |
||||
| Normaltidig glyphosat- sprøjtning |
720 g | Medio juni | 20. juni | 29. juni | ||||
| Sen glyphosat- sprøjtning |
720 g | Medio juli | 11. juli | 12. juli | ||||
| Usprøjtet | – | – | – | – |
Ukrudt blev ikke sået. Den naturlige bestand var domineret af hvidmelet gåsefod, agersennep og burresnerre.
2.1.2.2 Målinger
Ukrudtet blev gjort op i 4 kategorier, de tre dominerende plantearter samt resten, inden hver sprøjtning samt 1 måned efter sidste sprøjtning. På hver prøvetagningsdag afhøstedes ukrudtet i 3 plots á 0,5 m² i hver af de 20 parceller. I hvert plot optaltes antal planter. Desuden måltes overjordisk frisk- og tørvægt samt bladareal i ét af plottene. Målingerne blev foretaget den 7.6, 20.6, 11.7, 9.8 og 3.10 i 2005 og den 13.6, 27.6, 10.7, 17.8 og 5.10 i 2006.
I 2005 blev der løbende målt gennemsnitlig højde af majsplanterne i de enkelte parceller.
Den 3. oktober 2005 og 9. oktober 2006 blev parcellerne høstet, og antal kolber samt frisk- og tørvægt af kolber og stængel+blade blev målt.
2.1.3 Ukrudtets vækstforhold i majs
Forsøget blev gentaget over to vækstsæsoner, 2005 og 2006. Begge år udlagdes 20 parceller á 10´3 m² på Forskningscenter Flakkebjergs jorder. Der etableredes 5 typer parceller, karakteriseret af varierende tætheder af afgrøde og ukrudt, med 4 gentagelser.
2.1.3.1 Etablering
Forsøgsarealet blev pløjet om efteråret og dernæst fældet, gødet med 680 kg/ha 22N-3P-8K, såbedsharvet og tilsået den 12. maj 2005 og den 15. maj 2006. Forsøget blev sået – i 2005 med NAXOS hybridmajs og i 2006 med Banguy hybridmajs – med majssåmaskine i 4-5 cm dybde og med 75 cm rækkeafstand i 4 rækker. Under såningen blev der givet yderligere 160 kg/ha 12N-23P-0K i rækkerne.
I 2005 blev frø af burresnerre og fuglegræs anskaffet hos Herbiseed, UK, mens lugtløs kamille var fra egen høst. I 2006 var alle ukrudtsfrø af egen høst. Efter spiretest af ukrudtet blev det håndsået dagen efter afgrødens såning i 0-1 cm dybde. Det blev tilsigtet at opnå specifikke plantetætheder (tabel 2.2 og 2.3). Dette opnåedes dels ved passende udsædsmængde og dels ved efterfølgende lugning af kimplanterne. Herunder lugedes også andre ukrudtsarter væk. I hver parcel blev hver ukrudtsart sået i 5 runde plots á 0,5 m², 15 plots i alt pr. parcel. I yderligere 5 sådanne plots pr. parcel fulgtes naturligt fremspiret hvidmelet gåsefod. I 2005 var såbedet meget tørt i fremspiringsperioden, hvilket forsinkede ukrudtets fremspiring.
Tabel 2.2. Tilsigtede plantetætheder 2005 (kimplanter pr. m²). Hvor der står flere arter, gælder tætheden for hver art.
| Parceltype | Majs | Burresnerre, fuglegræs | Lugtløs kamille | |||
| 1 | 8,5 | 0 | 0 | |||
| 2 | 0 | 500 | 250 | |||
| 3 | 8,5 | 20 | 20 | |||
| 4 | 8,5 | 100 | 40 | |||
| 5 | 8,5 | 500 | 250 |
Tabel 2.3. Tilsigtede plantetætheder 2006 (kimplanter pr. m²). Hvor der står flere arter, gælder tætheden for hver art.
| Parceltype | Majs | Burresnerre, lugtløs kamille, fuglegræs, hvidmelet gåsefod | ||
| 1 | 8,5 | 0 | ||
| 2 | 0 | 60 | ||
| 3 | 8,5 | 15 | ||
| 4 | 8,5 | 30 | ||
| 5 | 8,5 | 60 |
2.1.3.2 Målinger
På hver prøvetagningsdag afhøstedes 1 plot på 0,5 m² af hver art i hver af de 20 parceller. I hver plot optaltes antal planter. Desuden måltes overjordisk frisk- og tørvægt samt bladareal af ukrudtet (parceltype 2-5). Afhængigt af planternes udvikling kunne alle målinger ikke altid foretages på alle arter på alle datoer. Målingerne blev foretaget den 27.5, 4.7, 12.7, 26.7 og 4.10 i 2005 og den 5.7, 17.7, 1.8 og 10.10 i 2006.
Arternes udviklingstrin blev løbende bestemt i 3 kategorier: Vegetativ, blomstrende og afmodnet. Parcellerne blev høstet den 4. oktober 2005 og den 10.oktober 2006, og der opgjordes antal kolber samt frisk- og tørvægt af kolber og stængel+blade. Desuden opgjordes ukrudtets frøsætning den 4. oktober 2005, men ikke i 2006.
2.1.4 Ukrudtets vækstforhold i vårbyg
Forsøget blev gentaget over to vækstsæsoner, 2005 og 2006. Begge år udlagdes 20 parceller á 10´2 m² på Forskningscenter Flakkebjergs jorder. Der etableredes 5 typer parceller, karakteriseret af varierende tætheder af afgrøde og ukrudt, med 4 gentagelser.
2.1.4.1 Etablering
Forsøgsarealet blev pløjet om efteråret og dernæst fældet, såbedsharvet og tilsået den 11. april 2005 og den 24. april 2006. Vårbyg (Barke maltbyg) blev sået med kornsåmaskine i 2-4 cm dybde. Samtidigt med såning blev der givet 420 kg/ha 22N-3P-8K.
Ukrudtsfrø blev fremskaffet og udsået som i majs (2.1.4.1), blot var småparcellerne i vårbyg mindre, 0,1 m² og ikke 0,5 m² som i majs. De tilsigtede tætheder var en anelse højere (tabel 2.4 og 2.5) end i majs, da vårbyg er en stærkere konkurrent. I 2005 var såbedet meget tørt i fremspiringsperioden, hvilket forsinkede ukrudtets fremspiring.
De 4 ukrudtsfrie parceller blev sprøjtet, som anbefalet af Planteværn Online, den 23. maj 2005 med 1,4 g Harmony + 0,2 tablet Express pr. ha og den 27. maj 2006 med 0,2 L Basagran 480 + 1,5 tablet Harmony Plus St. + 0,15 L Agropol pr. ha.
Tabel 2.4. Tilsigtede plantetætheder 2005 (kimplanter pr. m²).
| Parceltype | Vårbyg | Burresnerre, fuglegræs |
Lugtløs kamille | |||
| 1 | 400 | 0 | 0 | |||
| 2 | 0 | 500 | 250 | |||
| 3 | 400 | 40 | 40 | |||
| 4 | 400 | 100 | 80 | |||
| 5 | 400 | 500 | 250 |
Tabel 2.5. Tilsigtede plantetætheder 2006 (kimplanter pr. m²).
| Parceltype | Vårbyg | Burresnerre, lugtløs kamille, fuglegræs, hvidmelet gåsefod | ||
| 1 | 400 | 0 | ||
| 2 | 0 | 60 | ||
| 3 | 400 | 15 | ||
| 4 | 400 | 30 | ||
| 5 | 400 | 60 |
2.1.4.2 Målinger
På hver prøvetagningsdag afhøstedes 1 plot på 0,1 m² af hver art i hver af de 20 parceller. I hvert plot optaltes antal planter. Desuden måltes overjordisk frisk- og tørvægt samt bladareal af både afgrøde (parceltype 1 og 3-5) og ukrudt (parceltype 2-5). Afhængigt af planternes udvikling kunne alle målinger ikke altid foretages på alle arter på alle datoer. Målingerne blev foretaget den 1.6, 15.6, 22.6, 29.6 og 4.7 i 2005 og den 12.6, 3.7, 21.8 i 2006.
Arternes udviklingstrin blev løbende bestemt i 3 kategorier: Vegetativ, blomstrende og afmodnet. Slutbiomasse af afgrøde og ukrudt samt ukrudtsarternes frøsætning blev målt den 18. august 2005 og den 26. oktober 2006.
2.1.5 Ukrudtets vækstforhold i vinterraps
Forsøget blev gentaget over to vækstsæsoner, 2005-2006 og 2006-2007. Begge sæsoner udlagdes 20 parceller á 10´2½ m² på Forskningscenter Flakkebjergs jorder. Der etableredes 5 typer parceller, karakteriseret af varierende tætheder af afgrøde og ukrudt, med 4 gentagelser.
2.1.5.1 Etablering
I første vækstsæson blev forsøgsarealet pløjet og jordpakket den 28. august. Dagen efter blev der rotorharvet, sået 3,5 kg/ha af sorten Elna i 3-4 cm dybde og efterfølgende tromlet. Der blev gødet samtidigt med såning, 30 kg/ha 22N-3P-8K, samt i foråret den 12. april, 135 kg/ha 22N-3P-8K.
I anden vækstsæson blev forsøgsarealet pløjet og jordpakket den 23. august. To dage efter blev der rotorharvet, sået 3,8 kg/ha af sorten Elna i 3-4 cm dybde og efterfølgende tromlet. Der blev gødet samtidigt med såning, 30 kg/ha 22N-3P-8K, samt i foråret den 28. april, 668 kg/ha 22N-3P-8K.
I 2005 blev frø af burresnerre anskaffet hos Herbiseed, UK, mens fuglegræs, lugtløs kamille og kornvalmue var fra egen høst. I 2006 var alle ukrudtsfrø af egen høst. Efter spiretest af ukrudtet blev det håndsået den 30. august 2005 og den 29. august 2006 i 0-1 cm dybde. Der sigtedes efter fastlagte plantetætheder (tabel 2.6). Dette opnåedes dels ved passende udsædsmængde, dels ved efterfølgende lugning af kimplanterne. Herunder lugedes også andre ukrudtsarter væk. I hver parcel blev hver ukrudtsart sået i 5 runde plots á 0,25 m², 20 plots i alt pr. parcel.
De 4 ukrudtsfrie parceller blev ikke herbicidsprøjtet i første vækstsæson, kun i den anden som anbefalet af Planteværn Online, den 14. marts 2007 med 1 L Kerb 500 SC pr. ha.
Tabel 2.6. Tilsigtede plantetætheder i begge vækstsæsoner (kimplanter pr. m²).
| Parceltype | Raps | Burresnerre, lugtløs kamille |
Fuglegræs | Kornvalmue | ||||
| 1 | 80 | 0 | 0 | 0 | ||||
| 2 | 0 | 250 | 500 | 250 | ||||
| 3 | 80 | 50 | 15 | 20 | ||||
| 4 | 80 | 100 | 30 | 100 | ||||
| 5 | 80 | 250 | 60 | 250 |
2.1.5.2 Målinger
På hver prøvetagningsdag afhøstedes 1 plot på 0,25 m² af hver art i hver af de 20 parceller. I hvert plot optaltes antal planter. Desuden måltes overjordisk frisk- og tørvægt samt bladareal af både afgrøde (parceltype 1 og 3-5) og ukrudt (parceltype 2-5). Afhængigt af planternes udvikling kunne alle målinger ikke altid foretages på alle arter på alle datoer. Målingerne blev foretaget den 17.11, 3.5, 10.5, 22.5, 19.7 og 6.11 i 2005-2006 og den 6.11,2.4, 18.4 og 2.7 i 2006-2007.
Afgrødens udviklingstrin blev løbende bestemt. Den endelige biomasse af afgrøde og ukrudt opgjordes den 20. juli 2006 og den 2. juli 2007.
2.1.6 Faunarespons på plantebiomasse
Til sammenligning med tidligere publicerede12 og upublicerede data (M. Bruus) blev der indsamlet data for insekt-, afgrøde- og ukrudtsbiomasse i to majsmarker. Disse marker var begge beliggende lige øst for Silkeborg. Den ene mark var på 10,4 ha og den anden på 16,5 ha. Begge marker blev dyrket konventionelt. Ved begge marker var der enkelte buske og træer i skellet, og markerne lå i et område med marker blandet med beboelse. Majsen blev sået ca. 1. maj. Den 14. maj blev der foretaget ukrudtsbekæmpelse med 0,5 L/ha Calaris og 10.2 g as./ha MaisTer, og den 26. maj med 0,5 L/ha Calaris og 31 g as./ha MaisTer.
2.1.6.1 Indsamlingsdesign
Prøverne blev indsamlet på fem datoer fra begyndelsen af juni til begyndelsen af august. I hver mark blev der indsamlet 12 prøver på hver prøvetagningsdato, 6 felter 1 m inde og 6 felter 30 m inde (figur 2.1).
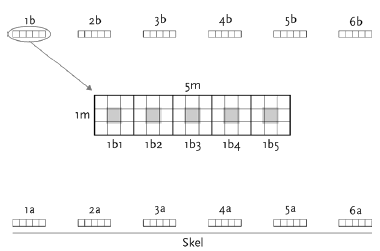
Figur 2.1. Design for indsamling af leddyr og vegetation i majs.
De to rækker af prøvetagningsfelter lå hhv. 1 m og 30 m fra markskellet. I et prøvetagningsfelt (1´1 m²) indsamledes leddyr med 9 sug i et 3´3 mønster, hvorefter vegetationen afhøstedes fra den midterste ½´½ m² (gråt areal). De 5 prøvetagningsfelter på stribe blev anvendt på 5 forskellige prøvetagningsdage.
2.1.6.2 Prøvetagning
En leddyrprøve dækkede 0,81 m² og bestod af 9 sug med D-vac taget inden for 1 m² (figur 2.1). Hvert sug varede 10 sekunder. Prøven blev først grovsigtet for at frasortere plantedele m.v. og derefter sigtet i en 1,8 mm sigte, idet den fraktion, der bliver tilbage i sigten, repræsenterer mulig fugleføde. Efter tørring ved 80oC i ca. 24 timer blev prøverne vejet.
Vegetationsprøverne blev taget efter leddyrprøverne fra de midterste ¼ m² (figur 2.1), som indeholdt én række majs:
- Højden af afgrøden og ukrudtet blev målt (uden at strække planterne).
- Dækningsgraden af henholdsvis afgrøde og den samlede ukrudtsflora blev bedømt.
- De tilstedeværende ukrudtsarter blev noteret.
- Der høstedes biomasse af afgrøde og total ukrudt (alle ukrudtsarter i én pose, afgrøde i en anden).
Planterne blev tørret ca. 48 timer ved 80oC og derefter vejet.
2.1.6.3 Dataanalyse
Sammenhængen mellem insektbiomasse og de forskellige mål for ukrudt og afgrøde blev analyseret ved Spearmans ikke-parametriske korrelationsanalyse36, da denne ikke forudsætter en speciel type fordeling af data, og heller ikke forudsætter en lineær sammenhæng, men blot en monoton sammenhæng. Data for de to majsmarker blev analyseret hver for sig, idet data for de forskellige datoer ikke er uafhængige inden for den enkelte mark, og resultaterne siger derfor ikke med sikkerhed noget generelt om majsmarker men beskriver udviklingen i de to undersøgte marker.
2.1.7 Modellering af ukrudtets populationsdynamik
2.1.7.1 Parameterestimater og simulering
Modellen FieldWeeds (figur 2.2 og 2.3) anvendtes til at modellere populationsdynamikken af de fem udvalgte ukrudtsarter: Agersennep, burresnerre, fuglegræs, hvidmelet gåsefod og lugtløs kamille (afsnit 1.6).
Modellen indeholder en række parametre, som beskriver ukrudtets biologi, afgrødevækst og andre dyrkningsmæssige faktorer. I de følgende afsnit redegøres for, hvorledes værdierne for disse parametre er blevet estimeret.
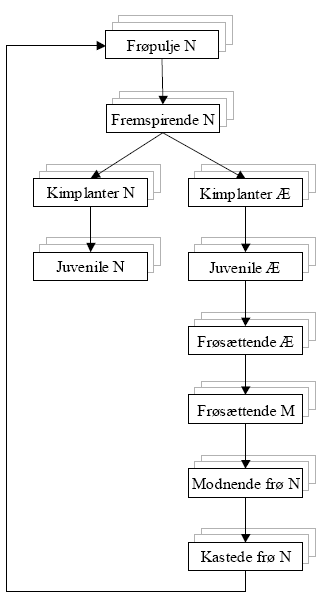
Figur 2.2. Livscyklusmodellen i FieldWeeds.
Kasser indeholder det angivne livsstadium for én ukrudtsart. Bagvedliggende kasser symboliserer samme model for andre ukrudtsarter. Pile angiver fysiologisk udvikling (ældning), som forløber efter graddagsskala.N = Antal pr. m². Æ = Tæthedsækvivalenter pr. m², som korrigerer antallet (N) for ukrudtets konkurrenceevne i forhold til afgrødens bladarealindeks på ukrudtets fremspiringstidspunkt (Æ≤N), se figur 2.3. M = Ukrudtsbiomasse g/m². Beregnes ud fra Æ i forhold til den aktuelle afgrødes konkurrenceevne.
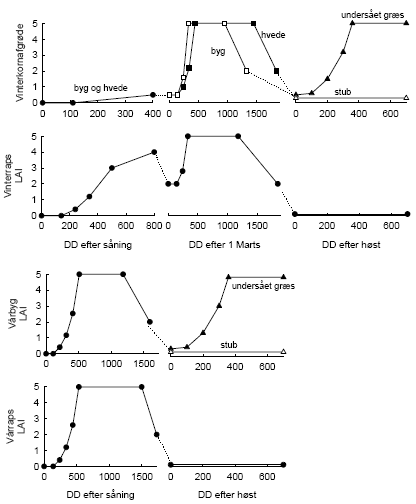
Figur 2.3. Simpel afgrødemodel anvendt i FieldWeeds.
Denne simple vækstmodel for afgrøden (J.E. Olesen, upubliceret) indvirker på konkurrenceevnen af det ukrudt, som spirer på et bestemt tidspunkt. Jo større LAI (bladarealindeks), desto mindre tæller ukrudtet udtrykt i tæthedsækvivalenter (Æ). oD= Graddage over 0oC
2.1.7.2 Frøpulje
Frøene opbevares i modellen i 20 jordlag á 1 cm. Ved jordbearbejdning skifter de plads mellem lagene, som beskrevet af Cousens & Moss 37.
I modellen forsvinder frøene enten som følge af mortalitet eller fremspiring (tabel 2.7). Frøene antages at dø med en fast rate, det vil sige, at henfaldet er eksponentielt.
For hvert tidsskridt (1 døgn) spirer fra hvert jordlag en del af frøene i forhold til frøets spirevillighed (i) i denne dybde (tabel 2.8), (ii) på denne dato (tabel 2.9) og (iii) under det aktuelle bladdække (LAI) af afgrøden (figur 2.3). Denne relative spirevillighed skaleres, så den samlede årlige fremspiring kommer til at matche den empirisk kendte årlige fremspiring (tabel 2.7).
Tabel 2.7. Frøpuljemodel: Estimering af årlig mortalitet og fremspiring.
| Ukrudtsart | Mortalitet og fremspiring | |
| Agersennep | Mortalitet og fremspiring: 0,77±0,033 år-1 38. Mortalitet og fremspiring: 1–0,050.2=0,45 år–1; heraf 35% fremspiring 39. Spiring af nyhøstede frø: 62%40 til 80% 41. Første år spirer 94-99%42. Estimater: Optimal fremspiring sættes til 0,95 år–1; mortalitet findes ved simulering så at mortalitet og fremspring bliver 0,45 år–1. |
|
Burresnerre |
Mortalitet målt til 0,2 år–1 43. Mortalitet sat til 0,6 år–1 i model44. Årlig fremspiring sat til 3,7% i vintersæd, 3,6% i vårsæd og 1,0% i majs i model45,46. I en anden model blev den sat til 40% for nye frø og 50% for ældre frø44. Kuldebehandling (14 dage ved 3oC) fremmer spiringen fra 10 til 90%47. Total årlig fremspiring i marken 60-80%48. Estimater: Optimal fremspiring sættes til 0,8 år–1; mortalitet sættes til 0,2 år–1. |
|
Fuglegræs |
Eksponentielt henfald over 20 år i uforstyrret jord var 0,122 år–1 49. Eksponentielt henfald over 10 år i uforstyrret jord var 0,287 år–1 50. Frøhvile varierer meget mellem forskellige populationer51. Spiringen falder med varigheden af stratificeringen, fra 100 til 10% (op til 6 uger ved 3oC)52. I væksthus spirede 75% af frøene53. Fremspiring i marken 0,06 til 0,36 år–1 varierende med sted og år54. Estimater: Optimal fremspiring sættes til 0,4 år–1; mortalitet sættes til 0,2 år–1. |
|
| Hvidmelet gåsefod | Eksponentielt henfald over 10 år i uforstyrret jord var 0,317 år–1 50. Fremspiring om foråret er større jo koldere vinter (2 til 25%)54. Fremspiring over 2 år var 6.6%55. Fremspiring 0,06-0,18 år–1 56. Kuldebehandling (–5 til 5oC) fremmede spiringen fra 30 til 64%57. Estimater: Optimal fremspiring sættes til 0,25 år–1; mortalitet sættes til 0,3 år–1. |
|
| Lugtløs kamille | Eksponentielt henfald over 20 år i uforstyrret jord (vellugtende kamille) var 0,105 år–1 49. Over 5 år i uforstyrret jord forsvandt 47%, det vil sige 1–0,530.2 = 0,12 år–1. Fremspiring over 2 år var 7,1%55. Spireevnen i laboratoriet var 28-47%40. Estimater: Optimal fremspiring sættes til 0,3 år–1; mortalitet sættes til 0,1 år–1. |
Tabel 2.8. Frøpuljemodel: Iboende evne til fremspiring i forhold til jorddybde.
| Ukrudtsart | Fremspiringsdybde | |
| Agersennep58 | 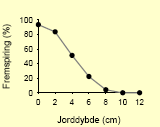 |
|
| Burresnerre58 | 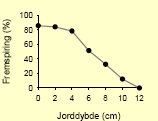 |
|
| Fuglegræs58 | 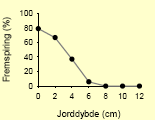 |
|
| Hvidmelet gåsefod58 | 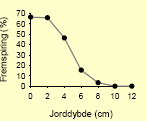 |
|
| Lugtløs kamille55 | 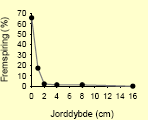 |
Tabel 2.9. Frøpuljemodel: Iboende evne til fremspiring efter
kalendermåned.
| Ukrudtsart | Fremspiringskalender | |
| Agersennep59 | 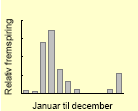 |
|
| Burresnerre48 | 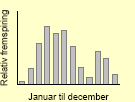 |
|
| Fuglegræs49 | 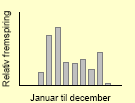 |
|
| Hvidmelet gåsefod59 | 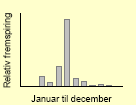 |
|
| Lugtløs kamille60 | 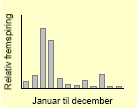 |
2.1.7.3 Udvikling
Efter fremspiring beskrives plantens udvikling (tabel 2.10.) på en graddagsskala (”oD” = graddage).
Tabel 2.10. Varigheden af ukrudtsarternes livsstadier.
| Ukrudtsart og stadium | Varighed | |
| Agersennep | ||
| Juvenil | 800oD 41 over 4oC | |
| Frøsætning | 400oD 41 over 4oC | |
| Frømodning | 200oD 41 over 4oC | |
| Frøkastning | 20 dage (gæt) | |
| Burresnerre | ||
| Juvenil | 721oD over 5.3oC, hvor kun de lyse timer tælles med i dagene61. | |
| Frøsætning | 375oD 62 over 4oC | |
| Frømodning | 150oD 62 over 4oC | |
| Frøkastning | 20 dage (gæt) | |
| Fuglegræs | ||
| Juvenil | 700oD over 0.5oC63 | |
| Frøsætning | 2-3 måneder51. Planten har ikke-determineret vækst; varigheden sættes arbitrært til 1 år. | |
| Frømodning | 10 dage64. Sættes til 150oD over 0.5oC. | |
| Frøkastning | 15 dage (gæt) | |
| Hvidmelet gåsefod |
||
| Juvenil | 436oD over 5.5oC65 500oD over 2.0oC66 |
|
| Frøsætning | 220oD over 5.5oC65 | |
| Frømodning | 225oD over 5.5oC65 | |
| Frøkastning | 50 dage (gæt) | |
| Lugtløs kamille |
||
| Juvenil | 600oD over 4oC 67 | |
| Frøsætning | 375oD over 4oC 67 | |
| Frømodning | 4 uger67. Sættes til 400oD over 4oC. | |
| Frøkastning | 50 dage (gæt) | |
2.1.7.4 Vækst og udbyttetab
Udbyttetab beregnes ud fra sammenhængen mellem fremspiringstæthed og procent udbyttetab68(tabel2.11). Plantetætheden omregnes til tæthedsækvivalenter, som afhænger af afgrødens bladdække (LAI) på fremspiringstidspunktet. Øget skygning resulterer i færre tæthedsækvivalenter. Udbyttetabet omregnes til ukrudtsbiomasse ud fra en fast relation på 80% udbyttetab af den endelige ukrudtsbiomasse.
Tabel 2.11. Udbyttetab forårsaget af ukrudt. Ukrudtsplantetætheden regnes i tæthedsækvivalenter, som korrigerer for fremspiringstidspunktet. (a) Værdier for vårraps. *Værdier skønnet ud fra de øvrige værdier.
| Ukrudt og afgrøde | Udbyttetab pr. ukrudtsplante; i (%) |
Max. udbyttetab; a (%) |
||
| Agersennep | ||||
| Vinterkorn | 0,469 | 4o* | ||
| Vinterraps (a) | 2,070 | 10070 | ||
| Vårkorn | 0,4* | 40* | ||
| Majs | 3,0* | 40* | ||
| Roer | 3,641 | 60* | ||
| Burresnerre | ||||
| Vinterkorn | 1,569,71-74 | 10071,73,74 | ||
| Vinterraps | 2,0* | 100* | ||
| Vårkorn | 0,575 | 100* | ||
| Majs | 3,0* | 100* | ||
| Roer | 3,0* | 100* | ||
| Fuglegræs | ||||
| Vinterkorn | 0,269,71,72 | 571 | ||
| Vinterraps | 0,776 | 5576 | ||
| Vårkorn | 0,2* | 5* | ||
| Majs | 3,0* | 40* | ||
| Roer | 3,0* | 60* | ||
| Hvidmelet gåsefod | ||||
| Vinterkorn | 0,269 | 36* | ||
| Vinterraps (a) | 0,870 | 2570 | ||
| Vårkorn | 0,577 | 3677 | ||
| Majs | 3,578-80 | 4079,80 | ||
| Roer | 2,541,81 | 6081 | ||
| Lugtløs kamille | ||||
| Vinterkorn | 169,71 | 1671 | ||
| Vinterraps | 1* | 16* | ||
| Vårkorn | 1* | 16* | ||
| Majs | 3,0* | 40* | ||
| Roer | 3.0* | 60* | ||
Parametrene for udbyttetab (tabel 2.11) kan sammenlignes med den rangordning af ukrudtet, som blev foretaget i et tidligere Miljøstyrelsesprojekt82. F.eks. fandtes burresnerre også da at være mere tabsgivende i vinter- end i vårsæd. Den åbne afgrøde, majs og roer, var ikke omfattet af dette tidligere studium.
2.1.7.5 Frøproduktion
Der forventes en allometrisk relation83 mellem endelig ukrudtsbiomasse og frøproduktion af formen, y=axb, hvor b eventuelt kan være 1 (tabel 2.12). For agersennep valgtes parameterværdierne, a=41,7 og b=1,14. For de øvrige fire arter anvendtes de værdier, som blev målt i dette projekt (figur 3.11).
Tabel 2.12. Model for frøproduktion.
| Ukrudtsart | Frøproduktion (y; antal frø) afhængig af ukrudtsbiomasse (x; g) | |
| Agersennep | y = 139x 21 y = 41,7x1,14 84 |
|
| Burresnerre | y = 21,4x 85 y = 22,5x 86 |
|
| Fuglegræs | y = 177x+23 87 y = 64,6x1,15 84 |
|
| Hvidmelet gåsefod | y = 1574x 21 y = 420x1.19 88 y = 735x 89 y = 430x–62 90 |
|
| Lugtløs kamille | y = 789x 86 y = 562x1,05 84 |
2.2 Herbicideffekter på hegnets flora
2.2.1 Ukrudtsbiomasse og frøproduktion
Mælkebøtte, alm. hvene og fåresvingel blev valgt som repræsentative testplanter for flerårige arter, som forekommer i hegnets fodpose.
Frø af mælkebøtte, alm. hvene og fåresvingel blev i slutningen af februar sået i 2 L potter i en blanding af jord, sand og sphagnum (2:1:1 vægtforhold) indeholdende alle nødvendige mikro- og makronæringsstoffer. Alle frø var indkøbt hos HerbiSeed, U.K. Potterne blev placeret på borde i et opvarmet væksthus. Efter fremspiring blev antallet af planter pr. potte reduceret til 4 i potter til biomassemåling og 2 i potter til frøhøst. Potterne blev herefter flyttet til et uopvarmet væksthus med henblik på vernalisering.
Glyphosat og MaisTer blev udsprøjtet på tre forskellige udviklingstrin af hver planteart (tabel 2.13).
| Tabel 2.13. Sprøjtedatoer og udviklingstrin ved sprøjtning af tre arter fra hegnets fodpose. | ||||||||
| Dato | Mælkebøtte | Alm. hvene | Dato | Fåresvingel | ||||
| 14/4 | 6 blade | 3 blade, 1 sideskud |
29/3 | 6 sideskud | ||||
| 25/4 | 9 blade | 4 sideskud | 12/4 | 20 cm høj, begyndende strækning | ||||
| 4/5 | 12 blade | Begyndende strækning | 27/4 | 30 cm høj | ||||
Herbiciderne blev udsprøjtet i en væskemængde på ca. 150 L/ha ved hjælp af en pottesprøjte. Der blev anvendt 6 doseringer af hvert herbicid, og hver behandling blev udført med 3 gentagelser. Doseringerne blev varieret i forhold til plantearternes forventede følsomhed på de forskellige udviklingstrin. Den højeste dosering af glyphosat varierede fra 360 g/ha til 1080 g/ha ved de forskellige behandlinger, mens den højeste dosering af MaisTer varierede fra 31 til 124 g as./ha.
Biomassemålingerne blev udført ca. 3 uger efter sprøjtning, hvor frisk- og tørvægt af planterne blev målt. For mælkebøtte blev der foretaget biomassemåling af både overjordiske og underjordiske dele. Rødderne blev således vasket fri for jord, og frisk- og tørvægt blev bestemt. De mælkebøtteplanter, som skulle anvendes til frøhøst, blev omplantet i 10 L’s spande. Frøhøst af alle arter blev foretaget løbende over en periode på 3 måneder, hvor modne frøstande blev afklippet i poser. Efter afsluttet høst blev antallet af frøstande talt, og frøene blev tørret og renset ved sigtning og gennemblæsning med luft. Den samlede vægt af frø per potte blev målt. Tusindkornsvægten blev bestemt udfra optælling og vejning af 3 x 200 frø fra hver potte.
2.2.2 Vækststandsning
2.2.2.1 Væksthusforsøg
Forsøgene blev udført på stor nælde og mælkebøtte, som repræsenterer to flerårige arter, der forekommer i hegn. Frøene var indkøbt hos HerbiSeed, UK. Effekten af subletale doseringer på planternes vækst blev undersøgt på henholdsvis frøfremspirede og rodfremspirede skud. Begge plantearter blev sået i 2 L potter i en blanding bestående af markjord, sand og sphagnum (2:1:1 vægt %). Potterne blev placeret i væksthus. Efter fremspiring blev antallet af planter pr. potte reduceret til 1. Da planterne af stor nælde var ca. 15 cm høje, og mælkebøtte havde 8-10 blade, blev den overjordiske plantemasse klippet af i de potter, hvor der skulle frembringes rodfremspirede skud.
Senest dagen før sprøjtning blev potterne placeret på et automatisk vandebord i væksthus. Her blev vægten af hver enkelt potte registreret. Herbicideksponering blev foretaget, da stor nælde havde 6 til 8 blade og mælkebøtte 8 til 12 blade, uanset om der var tale om frø- eller rodfremspirede planter.
Herbiciderne blev udsprøjtet i en væskemængde på ca. 150 L/ha ved hjælp af en pottesprøjte. Der blev ved alle behandlinger anvendt 6 doseringer af glyphosat og 4 doseringer af MaisTer. Alle behandlinger blev gentaget 3 gange. Doseringerne blev varieret ud fra den forventede følsomhed af de enkelte plantearter og udviklingstrin. På stor nælde blev der som maksimaldosering anvendt fra 720 til 1800 g glyphosat pr. ha og fra 12,4 til 31 g as./ha MaisTer, mens de tilsvarende doseringer på mælkebøtte var 360 g glyphosat pr. ha og 7,8 til 15,5 g as./ha MaisTer.
De 15 gentagelser af hver behandling blev opdelt i 5 hold potter med 3 gentagelser pr. behandling. De første 3 uger efter sprøjtning blev der ugentligt høstet et hold planter, hvor frisk- og tørvægt blev målt. Desuden blev et hold planter anvendt til måling af frøproduktion, hvor modne frøstande blev klippet af ugentligt i op til 4 måneder efter behandling. Frøene blev tørret og renset og den samlede vægt af frø pr. potte blev målt. Tusindkornsvægten blev bestemt udfra optælling og vejning af 100 frø fra hver potte. Det femte hold planter blev anvendt til biomarkørtest 7 dage efter sprøjtning. Ved denne test blev der afprøvet et test-kit, som er under udvikling i et innovationsprojekt finansieret af Danmarks FødevareErhverv, Danmarks Miljøundersøgelser og Aarhus Universitet. Det aktuelle test-kit kan forudsige om en herbicidbehandling medfører plantens død eller en vækststandsning.
I op til 3 uger efter behandling blev der ugentligt foretaget ikke-destruktive målinger i form af visuelle bedømmelser, registrering af vandforbrug og måling af fluorescens. Det daglige vandforbrug for hver enkelt potte blev registreret, idet potterne automatisk blev vejet flere gange dagligt og fik tilført næringsvæske til et fast sætpunkt. Fluorescensmålingerne (lys-responskurver) blev foretaget ugentligt med et HansaTech PAM på de rodfremspirede planter, dog blev der kun målt på det hold planter, som blev anvendt til frøhøst. Inden målingerne blev planterne stillet i et mørkt rum i mindst 1 time for at standardisere fotosynteseniveauet i planten i forhold til solskins- og gråvejrsdage. Alle målinger blev udført på samme blad af hver plante, idet bladet var afmærket med en bomuldssnor.
Der blev ugentligt foretaget visuelle bedømmelser af det hold planter, som fluorescensmålingerne blev udført på, samt det hold planter, som skulle høstes. Ved de visuelle bedømmelser blev der anvendt en skala fra 0 til 10, hvor 0 betyder ingen symptomer, 5 betyder 50% reduktion af biomasse, og 10 betyder, at planten er død.
2.2.2.2 Markforsøg
Forsøget blev udlagt på Forskningscenter Flakkebjergs jorder i 2005 og 2006.
I 2005 lå forsøget i en hegnsfodpose neden for et hvidtjørnehegn (mod nord). Selve fodposen var ca. 4 m bred og lå op til en majsmark (mod syd). I maj-juni måned var den østlige del af fodposen domineret af mælkebøtte, mens den vestlige del var domineret af stor nælde. Efter observationsperioden blev gråbynke dominerende i pletter. Forsøget blev delt i to, den ene del placeret i stor nælde og den anden i mælkebøtte. Der var 4 blokke med 4 plots svarende til antal behandlinger. Hver parcel var 0,2-0,3 m². Behandlingerne var randomiseret i hver blok.
I 2006 lå forsøget i en brakbræmme på 48´5 m² langs et skel. I marken i øvrigt dyrkedes vinterraps. Der blev udlagt 32 plots á 3´2½ m². I hvert plot udplantedes først i maj 6 planter, enten mælkebøtte eller stor nælde, 16 plots af hver art. Planterne var forinden drevet frem i potter i drivhus af lokalt indhøstede frø. Der var 4 blokke med 4 plots svarende til antal behandlinger. Behandlingerne lå systematisk i hver blok. Planterne voksede op i det naturligt forekommende ukrudt, domineret af agersennep og hvidmelet gåsefod.
Parcellerne blev sprøjtet den 12. maj 2005 med en rygsprøjte og den 19. juni 2006 (mælkebøtte) og den 20. juni 2006 (brændenælde) med forsøgssprøjte. De fire behandlinger var
- Usprøjtet.
- MaisTer: Stor nælde 100 g a.i. pr. ha, mælkebøtte 25 g a.i. pr. ha.
- Glyphosat: Stor nælde 540 g/ha, mælkebøtte 120 g/ha.
- Glyphosat: Stor nælde 1080 g/ha, mælkebøtte 360 g/ha.
Målinger 2005
Fra hvert plot blev der udtaget 3 planter til måling af frisk- og tørvægt, bladareal og antal blade: den 12. maj (før sprøjtning) og den 26. maj, for stor nælde tillige den 29. juni.
Fluorescens blev målt som fluorescens-induktionskurver og lys-responskurver med et HansaTech PAM på det øverste udvoksede blad på en plante i hver parcel, som blev udtaget tilfældigt. Inden målingerne blev planterne skygget i minimum 2 minutter for at standardisere fotosynteseniveauet i planten. Parcellerne blev målt dagligt lige før og lige efter sprøjtning og med længere tidsinterval fra 4 dage efter sprøjtning. Den sidste måling blev foretaget den 26. maj i mælkebøtte og den 7. juni i stor nælde.
Plantehøjde samt blomstring og aktiv vækst (kun for stor nælde) blev bedømt jævnligt indtil 5. august.
Målinger 2006
På hver prøvetagningsdag blev der høstet 3 planter af begge arter fra hver behandling, det vil sige 24 planter i alt fra 24 forskellige plots. Rødderne blev vasket rene for jord. Planterne blev lagt langs en lineal og fotograferet. Over- og underjordisk tørvægt samt bladareal blev målt pr. plante. De 8 prøvetagningsdage var den 14.6, 27.6, 10.7, 24.7, 7.8, 4.9, 2.10 og 30.10.
2.2.3 Planters konkurrenceforhold
2.2.3.1 Konkurrenceforsøg i væksthus
Frø af fåresvingel og alm. hvene blev sået i priklekasser i væksthus. Frøene var indkøbt fra HerbiSeed, UK. Efter fremspiring blev planterne udplantet i monokultur og binære blandinger i polystyrenkasser (40´40 cm²) i et faktorielt design (figur 2.4). Som vækstmedium blev anvendt en blanding af markjord, sand og sphagnum (2:1:1 vægtforhold).
Figur 2.4. Design af konkurrenceforsøg med to arter.
De viste 26 kombinationer af de to arters tætheder blev kombineret med 2 niveauer af hvert af de to herbicider samt 1 usprøjtet. Tre gentagelser gav i alt 3×26×3×2=468 plantekasser.
Glyphosat (22 og 61 g/ha) og MaisTer (1,2 og 2,4 g as./ha) blev udsprøjtet på planternes buskningsstadium (4-6 sideskud). Doseringerne var udvalgt, så den forventede effekt var henholdsvis 20 og 50% effekt på alm. hvene, som i dosis-respons forsøgene havde vist sig at være den mest følsomme af de pågældende arter over for begge herbicider.
Der blev anvendt en laboratoriesprøjte og udsprøjtning blev foretaget i en væskemængde på ca. 150 L/ha. MaisTer blev udsprøjtet i blanding med Maisoil.
Sideløbende med konkurrenceforsøgene blev der udført dosis-responsforsøg med begge arter i potter, således at det var muligt at bestemme den nøjagtige effekt af doseringerne under de givne betingelser. I dosis-responsforsøgene blev planterne behandlet med 6 doseringer af hvert herbicid. Forsøgsdetaljer i øvrigt som i afsnit 2.2.1.
Begge forsøg blev udført med 3 gentagelser pr. behandling, og planterne blev høstet 4-6 uger efter sprøjtning. Ved høst blev et på forhånd fastlagt antal planter af hver art fra hver kasse afklippet ved jordoverfladen, således at minimum de planter, som voksede i de yderste 5 cm af kassen, ikke blev høstet. Frisk- og tørvægt af de enkelte arter blev målt.
2.2.3.2 Konkurrencemodellen PCENV
PCENV er en statistisk regressionsmodel, som blev fittet til det kombinerede dosisrespons- og konkurrenceforsøg (2.2.3.1). Modellen beskriver effekten af herbicid og plantetæthed på biomassen af to arter ved hjælp af en generaliseret hyperbolsk regressionsmodel, som i adskillige studier har vist sig at være en god empirisk beskrivelse af biomasse hos konkurrerende planter91-97:
Den gennemsnitlige individbiomasse af de to arter kan beskrives som:
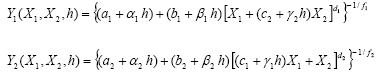 [1]
[1]
hvor indeks i=1...2 angiver de to arter.
Modellen beskriver sammenhængen mellem biomasse (Yi, g pr. plante) og tæthed (Xi, planter pr. m–2) for de to arter ved forskellige herbicidkoncentrationer (h, g/ha). Den empiriske model ser umiddelbart kompliceret ud men kan tænkes opbygget af tre simple komponenter:
1. Hyperbolsk aftagende biomasse som funktion af tæthed,
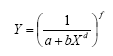
med formparametrene d og f, som begge er positive og antages at være uafhængige af herbicidkoncentrationen; (1/ai)fi er et mål for biomassen af art i ved grænsetilfældet af lav tæthed. Hvis di = fi = 1, da er 1/bi et mål for grænsetilfældet af biomassen af art i pr. areal ved høj tæthed i monokultur.
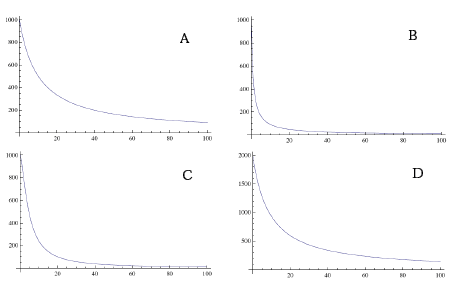
Eksemplerne i figur 2.5 viser (A) en art med maksimum biomasse på (1/0,001)1= 1000 g. I (B) ses, at kurven bliver stejlere jo mindre b er. Hvis d>1 (C) er konkurrencen ekstra stærk ved høj tæthed, mens f>1 (D) giver mindre konkurrence ved lav tæthed og maksimum biomasse på (1/0,001)1,1= 1995 g.
2. En kvantificering af interspecifik konkurrence i ækvivalenter af den målte art, f.eks. måler X1+c2X2 tætheden af art 1 og art 2 i ”art 1-ækvivalenter”.
3. En lineær effekt af herbicidet på de enkelte parametre, f.eks. måler a+ah effekten af herbicidet på parameteren a. Effekten af herbicidet (glyphosat eller MaisTer) på de to arters vækst og konkurrenceevne er således parameteriseret ved αi , βi og γi.
Figur 2.5. Eksempler på biomassemodel.
Parametrenes betydning for plantens biomasse (Y) afhængigt af dens tæthed (X). Følgende standardværdier er anvendt for modellens fire parametre: a=0,001; b=0,0001; d=1 og f=1. Eksemplerne (B-D) viser, hvorledes modellen reagerer på at ændre én parameter: (B) b=0,001; (C) d=1,5 og (D) f=1,1.
Fitning af model og beregning af ED10
Både data og modelrespons blev Box-Cox transformeret98 for at opnå varianshomogenitet og normalfordelt residualvarians. Modelparametre og de sammensatte funktioner af modelparametre og h blev eksponentielt transformerede for at sikre positive værdier, fx ![]() og
og ![]() , og fittet til data ved hjælp af både likelihood og Bayesianske metoder. Den Bayesianske fælles posteriorfordeling af parametrene i modellen [1] blev simuleret ved hjælp af MCMC metoder ved brug af Metropolis-Hastings algoritmen med 500.000 trækninger, hvor priorfordelingen af parametrene var antaget at være uniform99.
, og fittet til data ved hjælp af både likelihood og Bayesianske metoder. Den Bayesianske fælles posteriorfordeling af parametrene i modellen [1] blev simuleret ved hjælp af MCMC metoder ved brug af Metropolis-Hastings algoritmen med 500.000 trækninger, hvor priorfordelingen af parametrene var antaget at være uniform99.
For at kunne visualisere interaktionen mellem effekterne af konkurrence og herbicidsprøjtning blev den herbicidkoncentration, hvor den forventede biomasse er reduceret med 10% (ED10), beregnet ved at indsætte sammenhørende parameterværdier i modellen [1], for art i og løse ligningen,
![]() [2]
[2]
for ![]() . Ved at indsætte værdier fra den fælles posteriorfordeling af parametrene i [1] fandtes posteriorfordelingen af
. Ved at indsætte værdier fra den fælles posteriorfordeling af parametrene i [1] fandtes posteriorfordelingen af ![]() . Ved at beregne ED10 i stedet for f.eks. ED50 sikrer man sig, at effekten af herbicidet tilnærmelsesvis kan beskrives ved hjælp af lineære sammenhænge, som anvendt i [1].
. Ved at beregne ED10 i stedet for f.eks. ED50 sikrer man sig, at effekten af herbicidet tilnærmelsesvis kan beskrives ved hjælp af lineære sammenhænge, som anvendt i [1].
Effekten af at ændre tætheden af de to arter på ![]() ved f.eks. at ændre
ved f.eks. at ændre ![]() fandtes ved at beregne posteriorfordelingen af
fandtes ved at beregne posteriorfordelingen af ![]() og teste for, om fordelingen var forskellig fra 0.
og teste for, om fordelingen var forskellig fra 0.
2.2.4 Plantesamfundets succession
2.2.4.1 Forsøgets etablering og behandling
Forsøget gennemførtes på eksperimentelt etablerede felter, der tidligere havde indgået i en undersøgelse af effekter af glyphosat og kvælstof på vegetation og leddyr i hegn og engvegetation24. Inden udsåning af 33 plantearter, som kan findes i hegnets fodpose, blev forsøgsområdet dybdepløjet, hvilket efterlader de øvre jordlag så godt som fri for frø. I øvrigt fortsattes gødskning og sprøjtning som tidligere, det vil sige med en behandling hvert år sidst i maj (tabel 2.14).
Tabel 2.14. Behandlinger og dataindsamling i successionsforsøg. Forsøgsår 2001-2003 fra tidligere forsøg24.
| År | Kvælstoftilførsel | Herbicidbehandling | Dataindsamling | |||
| 2001 | Maj | 24. august | Juli og oktober | |||
| 2002 | Maj | April | August | |||
| 2003 | Maj | April | August | |||
| 2004 | Ingen | Ingen | Frøbankprøver den 24.9. | |||
| 2005 | 12. maj | 30. maj | Dækning og biomasse den 23.-25.5, 14.-20.6, 26.-31.8, 15.-16.9. | |||
| 2006 | 15. maj | April | Dækning og biomasse den 14.-20.6, 26.-31.8 | |||
| 2007 | 16. maj | April | Dækning og biomasse den 31.5 |
Kvælstof blev tilført i tre mængder: 0, 25 og 100 kg N/ha/år. Glyphosat blev tilført som Roundup Bio i fire mængder: 0, 14,4, 72 og 360 g as./ha. Sprøjtningerne blev foretaget med Lurmark Lo-drift LD 015 Green udstyr med 3 m sprøjtebom, 50 cm mellem dyserne og et tryk på 2.0 bar (figur 2.6). De 12 forskellige kombinationer af glyphosat og kvælstof blev udført i 10 gentagelser i et komplet randomiseret blokdesign med en blokstørrelse på 7´7 m², i alt 120 blokke.
MaisTer blev blot tilført i én dosering, 112 g as./ha og kun sammen med den højeste kvælstofmængde på 100 kg/ha/år. Denne behandling blev gentaget i 10 blokke á 4´7 m². Disse 10 blokke og 3 kontrolblokke uden tilførsel af MaisTer blev randomiseret og placeret langs den nordlige kant af forsøgsområdet. Den mindre blokstørrelse er acceptabel, da der ikke skulle foretages indsamlinger af f.eks. insekter og udelukkende anvendtes ikke-destruktive indsamlingsmetoder til vegetationsundersøgelserne.
2.2.4.2 Målinger
Frøbanken blev undersøgt én gang ved projektets begyndelse i de 120 blokke med glyphosat- og kvælstofbehandling, mens dækning (% dækning af jordoverfladen) og biomasse blev registreret flere gange årligt (tabel 2.14) i alle blokke.

Figur 2.6. Sprøjtning af successionsforsøg.
Undersøgelser af frøbank og dimensionering af sådanne undersøgelser vanskeliggøres ofte af den meget store rumlige variation, der er i frøbanker såvel i marken som i naturen. I nærværende projekt er denne variation imidlertid forsøgt elimineret ved at ’nulstille’ frøbanken ved en forudgående dybdepløjning, som omtalt ovenfor. Frøbanken, der findes i et plot, kan således primært tilskrives den eksperimentelle behandling plottet har været udsat for, det vil sige forskelle i glyphosatpåvirkning og kvælstoftilførsel. Frøbanken blev undersøgt ved at bestemme mængden af spiringsdygtige frø i jordprøver indsamlet med et jordbor med en diameter på 6 cm fra de øverste 5 cm af jorden. I hvert forsøgsplot indsamledes 4 prøver, hvorved opnåedes et samlet prøvetagningsareal på 0,0112 m² pr. plot. Prøverne blev indsamlet den 8. og 13. september 2004.
Efter indsamling blev prøverne opbevaret ved 0°C i 10 dage, herefter blev de sigtet med en 2 mm sigte for at sortere de største sten, rødder og grene fra. Jordprøven blev derpå bredt ud i et ca. 1 cm tykt lag i spiringsbakker med jord. Spiringsbakkerne blev stillet randomiseret på vandingsbord i væksthus ved minimum 8°C og lys 16 timer i døgnet. Bakkerne blev kontrolleret for nye planter to gange ugentligt igennem to måneder, hvorefter de fik lov at tørre ud. Ved hver optælling blev bakkens placering på ny randomiseret. Spiringsbakken blev inddelt i 6 mindre felter for at lette optællingen af planter (figur 2.7). Så snart en plante var identificeret til art, blev den registreret og derefter forsigtigt fjernet. Efter udtørring blev bakkerne stillet i fryser 6 døgn ved
-18°C, hvorefter prøverne blev optøet og jorden genopfugtet og atter sat til spiring i en måned. Alle fremspirede planter, det var muligt at identificere til art, var på dette tidspunkt undersøgt, og opgørelsen blev afsluttet. Til artsbestemmelsen anvendtes Dansk Felflora100.
Planternes dækning blev bestemt ved brug af point-intercept metoden101,102: En 75´75 cm ramme med et 15´15 cm net blev placeret over plantedækket, og 25 pinde blev ført vertikalt ned gennem plantedækket. For hver pind blev alle plantearter, som pinden berørte, registreret. Derved får man et mål for plantens dækningsgrad i procent.
For tre plantearter (alm. hvene, fåresvingel og kvik) blev den overjordiske biomasse desuden estimeret. Dette blev udført ikke-destruktivt ved at anvende en modifikation af point-intercept metoden103-105: I stedet for kun at registrere berøring eller ej optælles antallet af berøringer mellem pinden og planterne. Ud fra antallet af berøringer kan biomassen beregnes104, når man forud har bestemt sammenhæng mellem antal berøringer og biomasse. For at etablere denne sammenhæng blev der for hver af de 3 plantearter udvalgt 15 40´40 cm² kvadrater uden for forsøgsområdet, inden for hvilke tætheden af planten og dermed antallet af berøringer varierede så meget som muligt. Antallet af berøringer mellem pinden og den udvalgte planteart blev talt i 9 punkter i et 15´15 cm² net, og derefter blev planterne høstet ved jordoverfladen. Optælling og høst blev foretaget den 24. august 2005. Efter høst blev plantematerialet tørret ved 80°C i 24 timer og tørvægten bestemt.

Figur 2.7. Optælling af planter i spirebakke.
2.2.4.3 Databehandling
Effekten af herbicid- og kvælstoftilførsel på det samlede antal spirede planter i frøbankundersøgelsen blev analyseret ved hjælp af variansanalyse (proc mixed i SAS, med herbiciddosis og N-tilførsel som systematiske variable og transekt som tilfældig variabel). For at opnå varianshomogenitet blev data først log-transformeret.
Dækningsdata for de forskellige arter blev opdelt i alm. hvene, fåresvingel, kvik og en samleklasse med andre arter. Dette reducerede datasæt blev analyseret med en log-lineær model (SAS: PROC GENMOD med dist=poisson).
Ændringer i dækningen af de forskellige arter over tid blev undersøgt inden for hver kombination af kvælstof- og glyphosatbehandlingerne. Modellen kunne ikke generelt reduceres ved at udelade tredje ordens interaktionen (år´art´plot), og de plot, hvor der var signifikante (år´art´plot), blev effekten af behandlingen på dækningen undersøgt på det enkelte plot.
Effekten af kvælstof og glyphosat samt en eventuel interaktion mellem de to undersøgt i en apriori reduceret model (dækning = nit´art gly´art nit´gly´art år´nit´art år´gly´art år´nit´gly´art år´plot´art), hvor leddet år´plot´art indeholder den variation, som ikke kan forklares med effekten af kvælstof- og glyphosatbehandlingerne.
2.2.5 Modellering af fodposeplanternes populationsdynamik
Til modellering af fodposeplanterne anvendte vi modelleringsværktøjet, FieldEco, som er blevet udviklet over en årrække på Danmarks Miljøundersøgelser (DMU). I FieldEco simuleres planternes fotosyntese og vækst i detaljer under indflydelse af vejr samt indbyrdes konkurrence. Der indgik otte plantearter i modelleringen, som baseredes på grundige feltstudier (tabel 2.15).
2.2.5.1 Modelstruktur
Modellen styres af planternes nedarvede vækstmønster, som for hvert art beskrives ved hjælp af rater for respiration, vækst og reproduktion. Disse rater udmøntes på ethvert tidspunkt i plantens vækst i et behov for fotosynteseprodukter til brug for respiration, vækst (af rod, blade osv.) og reproduktion (knopper, blomster, frø). Alle planteorganer opdeles i modellen i to stadier: juvenile (f.eks. unge blade, der stadig vokser) og adulte (f.eks. gamle blade, der ikke vokser længere).
For hvert simuleringstrin, som er én dag, opererer modellen i fire trin:
(1) Beregning af behovet for fotosynteseprodukter (g tørstof). Dette behov udtrykker hvor meget tørstof, der skal fotosyntetiseres, for at planten kan opnå sin maksimale vækst inden for dette døgn. Det samlede behov gøres op som summen af, hvor meget der skal bruges til respiration, vækst og reproduktion af alle plantens organer: Rødder, blade, stængler, knopper, blomster, frø og eventuelle oplagringsorganer. For planteorganer som rødder og blade forstås ”reproduktion” som tilvæksten af nye blade og rødder.
(2) Beregning af hvor stor en del af behovet, der kan opfyldes. Produktionen af nyt tørstof via fotosyntesen foregår på grundlag af CO2, vand og lys. I modellen antages, at CO2 og vand ikke er begrænsende faktorer, hvorimod lysoptagelsen beregnes detaljeret (se nedenfor).
(3) Fordeling af ressourcen (det just producerede tørstof) til plantens forskellige organer i prioriteret rækkefølge. Prioriteringen varierer gennem året. I den periode, hvor plantearten foretager den reproduktive vækst (frøsætning), benyttes følgende prioritering: (i) frugter/frø, (ii) blomster, (iii) knopper, (iv) stængler, (v) blade og rødder (ressourcen deles ligeligt) og (vi) oplagringsorganer. Blade og rødder har dog reserveret 5% af ressourcen på forhånd. Når den reproduktive vækst er afsluttet, og frøene/frugterne er færdigudviklede, skifter prioriteringen til at give oplagringsorganer førsteprioritet samtidig med, at det ikke er muligt at prioritere reproduktiv vækst. Fordelingen indenfor de enkelte planteorganer giver de ældste stadier højeste prioritet, dvs. at de simulerede planter altid prioriterer det, der er skabt, frem for at skabe nyt.
(4) Beregning af vækst. Det beregnes hvor meget vækst, der kommer ud af de tildelte ressourcer, og hvor stor reproduktionen er for de enkelte organer (det vil sige hvor mange nye rødder, blade, stængler eller knopper, der skabes). Samtidigt beregnes ældning af alle organer.
2.2.5.2 Plantens omgivelser i modellen
Modellen drives af input angående temperatur og lysmængde. Hertil anvendes klimafiler fra DMU i Silkeborg med timemålinger af temperatur og solindstråling.
Temperaturen benyttes til at beregne tiden i fysiologisk tid (grad-dage, °D), som er en mere relevant tidsskala end dage og uger for vekselvarme organismer som planter. Den daglige graddagstilvækst beregnes ved at trække en tærskeltemperatur, hvorunder tilvæksten er så ringe, at den kan ignoreres, fra dagens gennemsnitstemperatur. Alle processer i planten relateres til graddage, f.eks. udtrykkes vækstraten i g/°D, reproduktionsrater i g/°D eller antal/°D; udviklingstider (f.eks. udviklingstiden for en knop) udtrykkes blot i °D. Ved at benytte graddage i stedet for dage og uger som tidsmåler opfanges temperaturens indflydelse på planternes vækst altså automatisk. Temperaturen kommer ind i modellen via en klimafil med timemålinger.
Lysoptagelsen er i modellen afhængig af plantens behov for fotosynteseproduktion, hvilket betyder, at planten under gode lysforhold maksimalt kan optage lys svarende til den optimale vækstrate. Hvis der derimod ikke er nok lys til rådighed til at tilfredsstille plantens behov, bliver plantens efterspørgsel kun delvist imødekommet. Derved bliver vækst- og reproduktionsrater tilsvarende lavere og kommer til at ligge under det optimale. Solindstrålingen sætter dagligt grænsen for fotosyntesen, og den kommer ligesom temperaturen fra en klimafil.
Solindstrålingen vil dog kunne ramme toppen af de højeste planter uhindret. Jo længere nede i plantemassen plantedelene befinder sig, jo mindre lys vil de have til rådighed på grund af skygning. I modellen simuleres lyskonkurrence imellem plantearter ved at inddele vegetationen i horisontale zoner, hvori det samlede bladarealindeks (planteareal pr. jordareal) er bestemmende for, hvor meget af lyset, der optages. I den øverste zone foretages beregningen ud fra den målte lysmængde (fra klimafilen), i næste zone ud fra den målte lysmængde minus det, der blev optaget i øverste zone, i næste zone igen fratrækkes den lysmængde, der blev optaget i de to overliggende zoner osv. Denne beregningsmetode betyder, at der kan opfanges så meget lys af en vegetation af tætte, høje planter, at der ikke er lys nok til små planter, der derfor vil dø. Når modellen har beregnet, hvor meget lys der er blevet optaget i de forskellige horisontale zoner, fordeles lyset til de forskellige plantearter proportionalt med deres bladarealindeks i zonen. Således er en plantepopulations samlede lysoptagelse summen af lysoptagelsen i de forskellige zoner. Den optagne lysmængde omregnes til fotosynteseprodukter (g tørstof) ved hjælp af en konstant omregningsfaktor.
Effekten af herbicider beregnes, afhængig af herbicidtype, ved at lade herbicidet nedsætte eller nulstille planteorganernes vækstrate og reproduktionsrate, eller ved at lade det nedsætte fotosynteseaktiviteten. Størrelsen af effekterne er afhængige af mængden af herbicid i forhold til anbefalet markdosis og blev beregnet ud fra forsøgene 2.4.1-2.4.4.
2.2.5.3 Vækstdata
Der blev i 2004-2005 indsamlet vækstdata for 8 plantearter karakteristiske for hegnets fodpose (tabel 2.15). Indsamlingerne blev foretaget med 3-5 ugers mellemrum for at samle data fra hele plantens overjordiske vækstcyklus.
Indsamlingerne blev foretaget på plots á ¼´¼ m² dog ikke for fåresvingel, som blev indsamlet pr. tue; tuens grundareal blev da også målt. Ved indsamling af store tuer, blev tuen delt op i halve eller kvarte tuer, og data korrigeret med denne faktor. For hver planteart blev der indsamlet data fra 10 plots, der blev valgt tilfældigt i områder, hvor den ønskede planteart var klart dominerende. En undersøgelse af variationen på de første indsamlinger viste en standard error på 10-20%. På den baggrund besluttedes det at fortsætte indsamlingen i 10 plots.
Tabel 2.15. Indsamlingslokaliteter og –datoer for de enkelte arter.
| Art | Datoer | Lokalitet | Lokalitetsbeskrivelse | |||
| ·fåresvingel ·rejnfan ·alm. hvene |
21.12 (kun fåresvingel), 11.4, 2.5, 23.5, 16.6, 7.7, 3.8, 24.8, 15.9, 18.10 | Brakmark på Djursland | Udyrket mark domineret af de tre indsamlede arter. | |||
| ·stor nælde ·alm. kørvel | 12.4, 3.5, 25.5, 14.6, 12.7, 1.8, 23.8, 13.9 og 17.10 | Levende hegn ved Harbovad vest for Silkeborg | Hegn af seljerøn, hvidtjørn og hyld; fodpose domineret af stor nælde og alm. kørvel med bunddække af kvik. | |||
| ·agertidsel ·mælke-bøtte ·kvik |
14.4, 3.5, 24.5, 17.6, 13.7, 2.8, 23.8, 13.9, 14.11 | Udyrket overdrev på kanten af Funder Ådal vest for Silkeborg | Prøveområder valgt, hvor de tre arter hver især var dominerende. |
Hvert plot blev inddelt vertikalt i højdezoner. I hver højdezone blev der for planterne af den pågældende art optalt antallet af blade, aks/kurve/skærme med knopper, aks/kurve/skærme med blomster og aks/kurve/skærme med frugter. Derudover blev højden af plantearten målt, og antallet af rodfæstede planter i plottet optalt. Fra hver højdezone blev de forskellige organer efter optællingen puttet i papirposer, som blev bragt tilbage til laboratoriet, hvor de blev tørret og vejet. Tørring foregik i tørreskab ved 60°C, indtil en konstant vægt var opnået. Ud over de optalte organer indgik også stænglerne i tørvægtsbestemmelserne. I nogle tilfælde var det uoverkommeligt at optælle alle bladene (f.eks. for alm. hvene og kvik), og antallet af blade blev da anslået ud fra tørvægten af 100 blade og den samlede tørvægt af alle bladene.
Ved to af indsamlingerne medio juli og ultimo august blev der foretaget målinger af den fotosyntetisk aktive stråling i toppen, midten og bunden af vegetationen for at kunne beregne lysoptagelsen.
2.2.5.4 Modelparametre
For hver af de 8 plantearter krævede modellen 60 parametre (tabel 2.16). Hvert planteorgan beskrives med 4-7 parametre, afhængig af hvilket organ, der er tale om, hvortil kommer 6 parametre, der gælder for planten som helhed. Det store datakrav til bestemmelse af disse mange parametre afspejles i det store arbejde i felten med at indsamle vækstdata (tabel 2.15). Med 8-11 indsamlingstidspunkter hver med 10 målepunkter pr. art opnåedes flere målepunkter end parametre, hvilket er nødvendigt for at kunne bestemme parametrene.
Modellen blev tilpasset de observerede data ved hjælp af masser af simuleringer med tilfældigt valg af parametre (indenfor et givet interval). Summen af de kvadrerede afvigelser imellem simulerede og observerede værdier for tørvægt (g/m²) og antal (pr. m²) blev udskrevet, og de parameterværdier, der gav den bedste sammenhæng imellem observerede og simulerede data, blev valgt. Ved gentagne tilpasninger opnåedes en god modelbeskrivelse. Tilpasningen blev udført for ét planteorgan ad gangen. I figur 2.8 ses den resulterende model for vild kørvel.
Planteartens gennemsnitshøjde gennem tiden simuleres af modellen, mens fordelingen omkring gennemsnittet beskrives ved hjælp af en fast parameter, idet højdefordelingen antages at følge en normalfordelingskurve med en fast spredning. Tilpasningen af højdefordelingskurven udførtes for hver art og hvert organ efter samme metode, som beskrevet ovenfor, ud fra data indsamlet i op til seks højdezoner.
Tabel 2.16. FieldEco-parametre for vild kørvel. Tilsvarende parametre blev bestemt for de øvrige 7 arter.
| Parametre for planteorganer | Stadium | Værdi | ||
| Rødder | ||||
| 1. Udviklingstid (°D) | Juvenile | 150 | ||
| 2. Udviklingstid (°D) | Adulte | 250 | ||
| 3. Startvægt (g) | Juvenile | 0,005 | ||
| 4. Reproduktionsrate (antal pr. °D pr. g total plantevægt) | Nyskabte juvenile | 0,003 | ||
| 5. Starttidspunkt (°D efter nytår) | Begge | 0 | ||
| 6. Vækstrate (pr. °D) | Juvenile | 0,02 | ||
| Blade | ||||
| 7. Udviklingstid (°D) | Juvenile | 165 | ||
| 8. Udviklingstid (°D) | Adulte | 250 | ||
| 9. Startvægt (g) | Juvenile | 0,0045 | ||
| 10. Reproduktionsrate (antal pr. °D pr. g total plantevægt) | Nyskabte juvenile | 0,0035 | ||
| 11. Starttidspunkt (°D efter nytår) | Begge | 0 | ||
| 12. Vækstrate (pr. °D) | Juvenile | 0,02 | ||
| 13. Areal pr. tørvægt (m²/g) | Begge | 350 | ||
| 14. Højdefordeling, faktor a | Begge | 0,03 | ||
| 15. Højdefordeling, faktor b (cm) | Begge | 30 | ||
| Stængler | ||||
| 16. Udviklingstid (°D) | Juvenile | 260 | ||
| 17. Udviklingstid (°D) | Adulte | 550 | ||
| 18. Startvægt (g) | Juvenile | 0,006 | ||
| 19. Reproduktionsrate (antal pr. °D pr. g total plantevægt) | Nyskabte juvenile | 0,0005 | ||
| 20. Starttidspunkt (°D efter nytår) | Begge | 0 | ||
| 21. Vækstrate (pr. °D) | Juvenile | 0,035 | ||
| 22. Omregningsfaktor, areal pr. tørvægt (m²/g) | Begge | 50 | ||
| 23. Højdefordeling, faktor a | Begge | 0,0085 | ||
| 24. Højdefordeling, faktor b (cm) | Begge | 50 | ||
| Knopper el. knopstande | ||||
| 25. Udviklingstid (°D) | Juvenile | 100 | ||
| –. Udviklingstid (°D) | Adulte | Overføres til blomster | ||
| 26. Startvægt (g) | Juvenile | 0,004 | ||
| 27. Reproduktionsrate (antal pr. °D pr. g total plantevægt) | Nyskabte juvenile | 0,015 | ||
| 28. Starttidspunkt (°D efter nytår) | Begge | 50 | ||
| 29. Vækstrate (pr. °D) | Juvenile | 0,03 | ||
| 30. Bremse blomster (°D) | Juvenile | 20 | ||
| 31. Bremse frugter (°D) | Juvenile | 50 | ||
| 32. Omregningsfaktor, areal pr. tørvægt (m²/g) | Begge | 80 | ||
| 33. Højdefordeling, faktor a | Begge | 1,4 | ||
| 34. Højdefordeling, faktor b (cm) | Begge | 40 | ||
| Blomster el. blomsterstande | ||||
| 35. Udviklingstid (°D) | Juvenile | 90 | ||
| –. Udviklingstid (°D) | Adulte | Overføres til frugter | ||
| 36. Starttidspunkt (°D efter nytår) | Begge | 50 | ||
| 37. Vækstrate (pr. °D) | Juvenile | 0,0 | ||
| 38. Højdefordeling, faktor a | Begge | 0,95 | ||
| 39. Højdefordeling, faktor b (cm) | Begge | 40 | ||
| Frugter el. frugtstande | ||||
| 40. Udviklingstid (°D) | Juvenile | 200 | ||
| 41. Udviklingstid (°D) | Adulte | 200 | ||
| 42. Starttidspunkt (°D efter nytår) | Begge | 100 | ||
| 43. Vækstrate (pr. °D) | Juvenile | 0,0 | ||
| 44. Omregningsfaktor, areal pr. tørvægt (m²/g) | Begge | 80 | ||
| 45. Højdefordeling, faktor a | Begge | 0,95 | ||
| 46. Højdefordeling, faktor b (cm) | Begge | 30 | ||
| Oplagringsorganer | ||||
| 47. Udviklingstid (°D) | Juvenile | 400 | ||
| 48. Udviklingstid (°D) | Adulte | 600 | ||
| 49. Startvægt (g) | Juvenile | 0,0003 | ||
| 50. Reproduktionsrate (antal pr. °D pr. g total plantevægt) | Nyskabte juvenile | 0,0006 | ||
| 51. Starttidspunkt (°D efter nytår) | Begge | 0 | ||
| 52. Vækstrate (pr. °D) | Juvenile | 0,018 | ||
| 53. Højdefordeling, faktor a | Begge | 0,95 | ||
| 54. Højdefordeling, faktor b (cm) | Begge | 50 | ||
| Alle organer | ||||
| 55. Temperaturtærskel, T0 (oC) | Begge | 5 | ||
| 56. Respirationsrate (pr. °D) | Begge | 0,01 | ||
| Hele planten | ||||
| 57. Reproduktiv periode start (°D efter nytår) | 0 | |||
| 58. Reproduktiv periode slut (°D efter nytår) | 700 | |||
| 59. Maksimal højde (cm) | 115 | |||
| 60. Højdevækstfaktor (proportionalitetsfaktor) | 0,75 |
Figur 2.8. FieldEco-model af vild kørvels organer.
Simulering (kurver) af feltdata (r) fra 2005, som beskriver de forskellige planteorganers tørvægt(A - E, G) og antal (H – M) samt plantens højde (F). Feltdata vist som gns.±s.e. Dato angivet månedsvist fra april til november.
3 Resultater
3.1 Herbicideffekter på markens biodiversitet
3.1.1 Ukrudtsbiomasse og frøproduktion
Både sort natskygge og hanespore udviste det typiske, S-formede dosis-respons på biomasse efter behandling med både glyphosat og MaisTer. Generelt var de anvendte doseringers effekt på frøproduktion lavere end på biomasse, men også denne parameter kunne beskrives ved hjælp af S-formede doseringskurver. Udfra doseringskurverne var det muligt at beregne hvilken dosering, der var nødvendig for at opnå en given effekt.
I relation til ukrudtsbekæmpelse er 90% effekt et ønsket mål og tabel 3.1 viser de beregnede ED90-doseringer af glyphosat og MaisTer på henholdsvis sort natskygge og hanespore med biomasse og frøproduktion som måleparameter. For begge ukrudtsarter krævede en sen sprøjtning (større udviklingstrin) for begge herbicider en højere dosering for at opnå 90% reduktion af biomassen. Doseringsbehovet øgedes mere for MaisTer end for glyphosat, hvilket sandsynligvis kan tilskrives, at glyphosat er mere systemisk end MaisTer.
Tabel 3.1. Reduktion af overjordisk plantebiomasse og frøsætning som respons på herbiciddosering. ED90 er den beregnede dosis for 90% reduktion. 95%-konfidensintervallet er vist i parentes. aEkstrapolerede doseringer. bMaksimalt anvendte doseringer (ekstrapolerede værdier var langt større).
| Ukrudtsart og herbicid | Udviklingstrin | Biomasse ED90 |
Frøsætning ED90 |
|||||
| Sort natskygge Glyphosat |
4 blade | 79,2 | (72,0...101) | 180a | ||||
| (g as./ha) | 8-10 blade | 263 | (169...360) | 443a | ||||
| MaisTer | 4 blade | 0,601 | (0,499...0,701) | >1,00b | ||||
| (g as./ha) | 8-10 blade | 4,10 | (2,40...5,80) | >2,00b | ||||
| Hanespore Glyphosat |
4 blade | 130 | (119...137) | 245 | (180...305) | |||
| (g as./ha) | 6-8 blade | 248 | (173...324) | 115 | (7,20...180) | |||
| MaisTer | 4 blade | 0,800 | (0,499...1,20) | 2,10 | (1,60...2,70) | |||
| (g as./ha) | 6-8 blade | 1,60 | (0,400...3.69) | 2,79 | (2,29...3,29) | |||
I forsøgene blev biomassen målt 3-4 uger efter sprøjtning og afspejler således korttidseffekten af behandlingerne, mens frøproduktionen blev målt flere måneder efter behandling og således afspejler langtidseffekten. Generelt skulle der højere doseringer til at reducere frøproduktionen end til at reducere biomassen (tabel 3.1). Det betyder, at plantens reproduktive evne er mindre følsom over for herbiciderne end dens vegetative vækst. Dvs. at selvom man opnår en vækstreduktion på kort sigt, så er planterne ofte i stand til på langt sigt at komme i vækst igen og producere frø.
Forskellen på ED90-doseringen for biomasse og frøproduktion var større, jo tidligere sprøjtningen blev udført, og var desuden generelt større med MaisTer end med glyphosat.
Det skal bemærkes, at forsøgene er udført som potteforsøg med planter i renbestand. Betydningen af konkurrence med afgrøden om lys, vand og næring er således ikke inddraget. Vi formoder at denne konkurrence vil forøge effekten af sprøjtningen, men at de relative forskelle fremhævet ovenfor vil gælde også i marken.
3.1.2 Glyphosatsprøjtning i majs
Forsøgene med forskellige sprøjtestrategier i majs faldt mere tydeligt ud i 2006 end i 2005 (figur 3.1-3.2). Når der ses på udbyttet, kan det konkluderes, at splitsprøjtning med glyphosat fungerede lige så godt som splitsprøjtning med MaisTer. Med blot én glyphosatsprøjtning var høstresultatet lige så godt, men kun med en tidlig sprøjtning (20. eller 29. juni), ikke med en sen (11. eller 12. juli).
Som ventet var ukrudtets tørvægt faldet ved prøvetagningen, som fulgte efter sprøjtningen. Den eneste undtagelse (agersennep ved tidlig glyphosatsprøjtning i 2006) skyldtes et enkelt afvigende plot – måske med blot en enkelt overlevende stor plante; dette fremgår af den store usikkerhed på gennemsnittet vist i figur 3.2.
3.1.3 Ukrudtets vækstforhold i majs
Den naturlige forekomst af hvidmelet gåsefod og lugtløs kamille i marken gjorde det svært at opnå de lave tæthedsniveauer i 2005 (tabel 3.2), men i 2006 lykkedes det (tabel 3.3). Som ventet for en afgrøde som majs, der i lang tid står åben, begrænsede den kun ukrudtets vækst lidt. Samtidigt led den selv betydeligt i konkurrencen (figur 3.3-3.4).
Tabel 3.2. Fremspiring af ukrudt i majsforsøg 2005 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
71,0 | ±31,22 | 49,5 | ±27,29 | 65,0 | ±82,31 | 99,5 | ±16,76 | ||||
| Med afgrøde Lav tæthed |
14,5 | ±14,18 | 6,00 | ±5,889 | 64,0 | ±88,18 | 34,0 | ±11,78 | ||||
| Mellem tæthed | 32,5 | ±19,49 | 24,5 | ±3,416 | 70,0 | ±106,9 | 34,0 | ±4,320 | ||||
| Høj tæthed | 65,5 | ±22,65 | 60,0 | ±16,73 | 36,0 | ±50,20 | 74,0 | ±36,11 | ||||
Tabel 3.3. Fremspiring af ukrudt i majsforsøg 2006 (gns.±s.d. kimplanter pr.
m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
31,5 | ±11,00 | 34,5 | ±19,69 | 28,5 | ±6,61 | 30,0 | ±12,96 | ||||
| Med afgrøde Lav tæthed |
10,5 | ±7,90 | 10,5 | ±3,79 | 13,5 | ±9,15 | 5,5 | ±1,00 | ||||
| Mellem tæthed | 8,5 | ±6,81 | 17,0 | ±6,63 | 22,5 | ±9,57 | 18,0 | ±7,12 | ||||
| Høj tæthed | 30,0 | ±7,48 | 32,5 | ±15,18 | 37,5 | ±11,00 | 24,5 | ±5,74 | ||||
3.1.4 Ukrudtets vækstforhold i vårbyg
Det lykkedes ikke helt at styre fremspiringstætheden af hvidmelet gåsefod, som i 2005 forsvandt helt fra to af tæthedsniveauerne (tabel 3.4), og af lugtløs kamille, som forsvandt i ét af niveauerne i 2006 (tabel 3.5).
Vårbyg var i alle tilfælde i stand til at trykke ukrudtet til betydeligt lavere tæthed, end når ukrudtet voksede i renbestand (figur 3.5-3.6). Det kan undre, at vårbyggen selv led så betydeligt i konkurrencen (figur 3.5-3.6 øverst), når man betænker, at ukrudtet aldrig (bortset fra burresnerre i 2005) blev særlig stort. Udbyttetabet er sandsynligvis et artefakt, idet lugearbejdet for at opretholde de ønskede ukrudtsbestande var ganske omfattende, og dette kunne ikke undgå at skade afgrøden.
Tabel 3.4. Fremspiring af ukrudt i vårbygforsøg 2005 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
658 | ±234,6 | 65,0 | ±94,34 | 67,5 | ±18,93 | 128 | ±40,31 | ||||
| Med afgrøde Lav tæthed |
87,5 | ±25,00 | 12,5 | ±9,574 | 0,0 | ±0,00 | 57,5 | ±37,75 | ||||
| Mellem tæthed | 123 | ±43,49 | 57,5 | ±22,17 | 0,0 | ±0,00 | 90,0 | ±24,49 | ||||
| Høj tæthed | 573 | ±131,0 | 205 | ±93,99 | 77,5 | ±17,08 | 220 | ±57,74 | ||||
Tabel 3.5. Fremspiring af ukrudt i vårbygforsøg 2006 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
65,0 | ±19,15 | 30,0 | ±42,43 | 60,0 | ±28,28 | 65,0 | ±25,17 | ||||
| Med afgrøde Lav tæthed |
12,5 | ±5,00 | 22,5 | ±20,62 | 42,5 | ±37,75 | 0,0 | ±0,00 | ||||
| Mellem tæthed | 45,0 | ±30,00 | 57,5 | ±9,57 | 12,5 | ±12,58 | 20,0 | ±33,67 | ||||
| Høj tæthed | 72,5 | ±53,15 | 62,5 | ±12,58 | 37,5 | ±25,00 | 35,0 | ±44,35 | ||||
3.1.5 Ukrudtets vækstforhold i vinterraps
Det var svært at styre fremspiringstætheden af ukrudtet især i efteråret 2005, hvor 5 ud af de 12 kombinationer mellem ukrudt og vinterraps endte uden ukrudt (tabel 3.6). For kornvalmue kom i alle tilfælde kun ganske få planter op (tabel 3.6-3.7).
Fremspiringen var stort set tilendebragt ved den første optælling af kimplanter i november i begge forsøg. Planterne fik imidlertid i de fleste tilfælde først rigtig gang i deres vækst i juni (figur 3.7-3.8). I 2006-2007 havde burresnerre og især fuglegræs dog opnået en betydelig størrelse allerede i november. De aftog i størrelse henover vinteren, hvorefter burresnerre atter tog fat fra juni af, mens fuglegræs helt uddøde. Denne uddøen må have skyldtes afgrødens konkurrence, da fuglegræs voksende alene genoptog sin vækst i juni akkurat som burresnerre (figur 3.8).
Tabel 3.6. Fremspiring af ukrudt i vinterrapsforsøg 2005-2006 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Kornvalmue | Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
194 | ±78,55 | 133 | ±115,7 | 35,0 | ±32,23 | 17,0 | ±21,26 | ||||
| Med afgrøde Lav tæthed |
35,0 | ±20,75 | 0 | ±0 | 0 | ±0 | 5,00 | ±5,030 | ||||
| Mellem tæthed | 83,0 | ±26,81 | 0 | ±0 | 0 | ±0 | 0 | ±0 | ||||
| Høj tæthed | 167 | ±54,59 | 77,0 | ±135,5 | 1,00 | ±2,000 | 16,0 | ±24,66 | ||||
Tabel 3.7. Fremspiring af ukrudt i vinterrapsforsøg 2006-2007 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Kornvalmue | Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
229 | ±132,1 | 350 | ±100,0 | 19,0 | ±22,72 | 91,0 | ±6,83 | ||||
| Med afgrøde Lav tæthed |
21,0 | ±16,45 | 33,0 | ±31,56 | 1,00 | ±2,000 | 6,00 | ±5,16 | ||||
| Mellem tæthed | 63,0 | ±17,09 | 64,0 | ±41,70 | 0 | ±0 | 15,0 | ±12,38 | ||||
| Høj tæthed | 26,0 | ±14,79 | 30,0 | ±25,82 | 4,00 | ±8,000 | 43,0 | ±17,70 | ||||
Figur 3.1. Sprøjtescenarier i majs 2005.
I øverste række vises tørvægten (kg) ved høst af 4 m af parcellen: Kolber (venstre søjler) og stængler (højre søjler). Søjler med samme bogstav er ikke signifikant forskellige (P=0,05). Nedenunder vises overjordisk tørvægt (g pr. ½ m²) for tre ukrudtsarter, samt andet ukrudt totalt. Sprøjtetidspunkter er angivet ved stiplede linier. Data vist som gns.±s.e.
Figur 3.2. Sprøjtescenarier i majs 2006.
I øverste række vises tørvægten (kg) ved høst af 4 m af parcellen: Kolber (venstre søjler) og stængler (højre søjler). Søjler med samme bogstav er ikke signifikant forskellige (P=0,05). Nedenunder vises overjordisk tørvægt (g pr. ½ m²) for tre ukrudtsarter, samt andet ukrudt totalt. Sprøjtetidspunkter er angivet ved stiplede linier. Data vist som gns.±s.e.
Figur 3.3. Vækst af afgrøde og ukrudt i majs 2005.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,5 m²) vist som gns.±s.e.
Figur 3.4. Vækst af afgrøde og ukrudt i majs 2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,5 m²) vist som gns.±s.e.
Figur 3.5. Vækst af afgrøde og ukrudt i vårbyg 2005.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,1 m²) vist som gns.±s.e.
Figur 3.6. Vækst af afgrøde og ukrudt i vårbyg 2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,1 m²) vist som gns.±s.e.
Figur 3.7. Vækst af afgrøde og ukrudt i vinterraps 2005-2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,25 m²) vist som gns.±s.
Figur 3.8. Vækst af afgrøde og ukrudt i vinterraps 2006-2007.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,25 m²) vist som gns.±s.e.
3.1.6 Faunarespons på plantebiomasse
Væksten af afgrøde og ukrudt samt tætheden af leddyr fulgtes ad gennem sæsonen på begge de undersøgte marker i 2005 (figur 3.9; Spearmans r gav P<0,0001).
På en given dag var der imidlertid generelt ringe sammenhæng mellem biomassen af ukrudt (eller afgrøde) og insekter i de to majsmarker (figur 3.10). Det var således ikke muligt ud fra ukrudtsbiomassen at forudsige insektbiomassen en given dag.
Figur 3.9. Afgrøde- og ukrudtbiomasse (g pr. 0,25 m²), afgrøde- og ukrudtshøjde (cm), samt leddyrbiomasse (mg pr. prøve à 0,81 m²). Punkter viser gns.±s.e. for 12 prøver.

Figur 3.10. Ukrudt og insekter i to majsmarker 2005.
Sammenhængen mellem biomassen af ukrudt og insekter er vist på fem datoer for begge marker (Kalbybaard og Linaa). Y-skala er den samme i alle figurer. Målingerne med nulværdier er udeladt.
3.1.7 Modellering af ukrudtets populationsdynamik
3.1.7.1 Ukrudtets frøproduktion
De supplerede data om ukrudtets frøproduktion, som blev indsamlet for at understøtte estimeringen af modellens parametre, gav robuste sammenhænge mellem biomasse og frøproduktion (figur 3.11).
![Figur 3.11. Ukrudtets frøproduktion. Tørvægt og frøproduktion af ukrudt høstet i plots á 0,1-0,5 m². Ukrudt enten uden konkurrence (o) eller i konkurrence med majs (?·) eller vårbyg (r). Lineære regressionslinier beregnet på log-transformerede data, punktet [·] ikke medtaget.](images/fig3_11.gif)
Figur 3.11. Ukrudtets frøproduktion.
Tørvægt og frøproduktion af ukrudt høstet i plots á 0,1-0,5 m². Ukrudt enten uden konkurrence (o) eller i konkurrence med majs (?·) eller vårbyg (r). Lineære regressionslinier beregnet på log-transformerede data, punktet [·] ikke medtaget.
3.1.7.2 Modelvalidering
Modellens parametre blev estimeret fra litteraturen samt figur 3.11, som beskrevet i afsnit 2.1.7. Det blev valgt ikke at tilpasse parameterværdierne, således at modellen bedst muligt beskrev resultatet af de seks valideringsforsøg (tre afgrøder gentaget over to år, se figur 3.3-3.8) – en såkaldt ”kalibrering” af modellen. Datagrundlaget i litteraturen blev anset for større og bredere, end det som kunne fremskaffes i dette projekt, og modellen baseret på litteraturen skulle således dække bredere, hvad angår forskelle i dyrkningsforhold og vejrlig.
Som testvariabel for valideringen anvendtes ukrudtsbiomassen inden høst, det vil sige på det tidspunkt, hvor den normalt topper, og hvor frøkastet sker i forhold til den opnåede biomasse. Derved testes modellens evne til at simulere resultatet af konkurrencen mellem afgrøde og ukrudt. Inden hver simulering indstilledes frøbanken til en tæthed, som gav den fremspiringstæthed for hver art, som var observeret i forsøget i afgrøden i det pågældende år (tabel 3.2-3.7). Valideringen checkede altså ikke, om modellen gav den rette fremspiring i forhold til vejr og jordbearbejdning, men kun slutresultatet af konkurrenceforløbet fra fremspiring og frem til høst.
Ser vi først på det generelle mønster, så ramte modellen godt i majs, mens der var en klar tendens til overestimering af ukrudtets vækst i vårbyg og vinterraps, især ved lave tætheder – under ca. 10 g/m², hvor modellen forudsagde en ukrudtsvægt på 10-100 g/m² (figur 3.12). Planteværn Online angiver for de fire anvendte ukrudtsarter i disse to afgrøder et effektkrav for behandling på 80-95%. Det vil sige, at man uden bekæmpelse ville forvente en betydelig vækst af ukrudtet gennem sæsonen og dermed et uacceptabelt udbyttetab, men ser man på vækstkurverne for ukrudtet i vårbyg (figur 3.5 og 3.6) og vinterraps (figur 3.7 og 3.8), fremgår det, at ukrudtet stort set blev udkonkurreret af afgrøden (forsøgene blev jo ikke sprøjtet). Det modellen forudsiger er altså, at ukrudtet i disse situationer ville opnå en vægt på 10-100 g/m², hvilket ville have været tabsgivende, men i de konkrete forsøg i Flakkebjerg de to år stod ukrudtet i stampe. Således opstår den konkrete overestimering.
Sammenligner man ukrudtets vækst i majs (figur 3.3 og 3.4) og vårbyg (figur 3.5 og 3.6), ser man, at ukrudtet både i 2005 og 2006 groede langt bedre i majs (konkurrencesvag) end i vårbyg (konkurrencestærk). Når ukrudtet trivedes dårligt i vårbyg, skyldtes det altså afgrødens vækst, som igen var styret af vejret. Man må forestille sig, at under andre vejr- og dyrkningsforhold ville ukrudtet have vokset sig stort og tabsgivende i vårbyggen. Modellen er imidlertid så simpel, at vejret ikke medregnes, og den kan derfor umuligt forudsige forskelle mellem år betinget af forskelle i vejret.
Modellen havde et særligt problem med vinterraps, hvor ukrudtets vækst afbrydes af vinteren. Overvintringen gør det mere kompliceret at simulere planters vækst korrekt. Det ses af valideringen, at særligt fuglegræs i 2006-2007 drillede (de tre punkter længst mod venstre i figur 3.12). Fuglegræs opnåede da sin maksimale vægt inden vinteren og ikke om sommeren før høst, som modellen antager. Dette er et væsentligt problem for modellen, som udbygges på dette punkt, hvis den skal anvendes til at modellere ukrudt i vintersæd i fremtiden. En yderligere komplikation opstår, når frøproduktionen skal modelleres. Det er ukendt, hvorvidt fuglegræs i 2006-2007 nåede at producere frø.
En sammenligning af de to år viste ingen forskel i modellens evne til at ramme observationerne (figur 3.12). Sammenfattende kan man sige, at modellen i disse tre afgrøder over to år havde en generel misvisning henimod en overestimering af ukrudtets vækst og dermed af dets frøsætning. Hvis denne misvisning gælder generelt, vil modellen have tendens til at overvurdere ukrudtsproblemet både inden for sæsonen og på længere sigt ved en overestimering af henholdsvis ukrudtets konkurrenceevne og populationsvækstrate. Imidlertid skal modellen her anvendes til at sammenligne forskellige dyrkningsstrategier og ikke til at forudsige absolutte ændringer i frøbanken. Til dette brug mener vi, at modellen er et pålideligt værktøj, selvom den med henblik på modellering af ukrudt i vintersæd bør udbygges.
3.1.7.3 Modellens antagelser
Konsekvensen af de fire dyrkningsscenarier (tabel 1.1 til 1.4) på frøbankdynamikken af de fem udvalgte ukrudstarter (afsnit 1.6) blev simuleret over 20 år, som vist i figur 3.13 til 3.16, begyndende med en moderat infestation på 100 frø pr. m² af hver art. Modellen har én fri parameter, frøpredation på jordoverfladen, som blev sat til 5% pr. dag. Frøpredationen er i virkeligheden meget variabel fra år til år og fra sted til sted, og den anvendte 5% pr. dag er den øvre grænse for det observerede106-108. Mens frøpredationen således blev sat temmelig højt, så blev herbicideffekten sat på den nedre grænse for det forventede. Modellen anvendte de målte effekter tre uger efter sprøjtning i væksthus, men effekten må for disse enårige arter forventes som regel at være større, da den overlevende biomasse vil være svækket og nemt vil kunne bukke under for konkurrence med afgrøden. For glyphosat anvendtes dosisresponskurver fra litteraturen7, mens Planteværn Onlines anvendtes for de øvrige herbicider.
Imidlertid er formålet med simuleringerne at forudsige forskelle i udviklingen af ukrudtsfloraen som følge af scenariernes forskellige sprøjtestrategier, snarere end at forudsige den absolutte størrelse af ukrudtspopulationerne. Ændringer af disse antagelser, frøpredationens størrelse og herbicidernes endelige effekt, vil ikke ændre på ukrudtsarternes relative position i konkurrencen, men blot flytte populationerne i samlet flok (og uændret rækkefølge) op eller ned ad y-aksen (figur 3.13-3.16).
3.1.7.4 Modellens forudsigelser
Dyrkning af GT-raps udløste ikke nogen ændring i ukrudtsfloraens sammensætning (figur 3.13). Hyppigheden af de fem ukrudtsarter efter 20 år var den samme som i sædskiftet med konventionel raps. Hvis sædskiftet havde været mindre anstrengt, og der kun havde været dyrket raps hvert fjerde år og ikke hvert tredje, så ville det blot have gjort ændringen om muligt endnu mindre.
Ved dyrkning af majs i sædskifte med vårbyg havde sprøjtestrategien indflydelse på ukrudtsfloraens udvikling (figur 3.14). Splitsprøjtning var mere effektiv med glyphosat end med MaisTer. Én sprøjtning med glyphosat den 8. juni havde mindre effekt, mens udsættelse til den 20. juni tillige forårsagede en voldsom udvikling af hvidmelet gåsefod. I praksis ville landmanden ikke acceptere en sådan udvikling men ændre sin sprøjtestrategi i majs. Det ses tydeligt, at frøbanken stiger efter år med majsdyrkning efterfulgt af et mindre fald i året med vårbyg.
Ensidig majsdyrkning viste sig at være en problematisk strategi i konventionel majs, da den fremmede lugtløs kamille som et problemukrudt (figur 3.15). Dette kan skyldes kamilles biologi. Dens frøpulje er mere persistent end de fem arters (mortalitetsrate på blot 0,1 år–1, tabel 2.7). Samtidigt har den den anden laveste fremspiringsrate (0,3 år–1), kun hvidmelet gåsefods er lavere (0,25 år–1) (tabel 2.7). Dyrkning af GT-majs med anvendelse af glyphosat som splitsprøjtning gav tilfredsstillende kontrol med ukrudtet, men med blot én sprøjtning fik hvidmelet gåsefod dog et voldsomt udbrud akkurat som i sædskiftet, hvor majs vekslede med vårbyg (figur 3.14).
I sædskifterne med roer gjorde anvendelse af glyphosat i GT-roerne ingen forskel på udviklingen af ukrudtsfloraen i forhold til den traditionelle sprøjtestrategi (figur 3.16).
Figur 3.12. Validering af modellen for markukrudt. Ukrudtsbiomassen ved høst.
Sammenligning mellem modellens forudsigelser og biomassen observeret i markforsøgene i Flakkebjerg (figur 3.3-3.8). Punkter over/under diagonalen angiver over-/underestimering af modellen. De samme data er vist seks gange: På lineær og logaritmisk skala (venstre og højre), for ukrudtsarterne (øverst), afgrøderne (midt) og forsøgsårene (nederst).
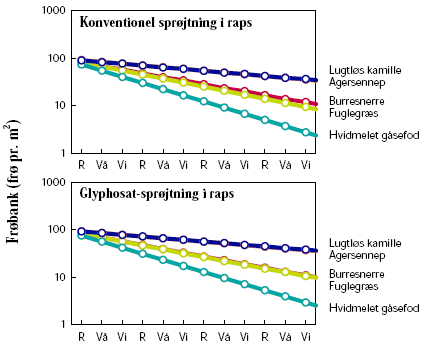
Figur 3.13. Scenarie 1: Sædskifte med konventionel vs. GT-vinterraps.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”R”=Vinteraps, ”Vå”=Vårbyg, ”Vi”=Vinterhvede.
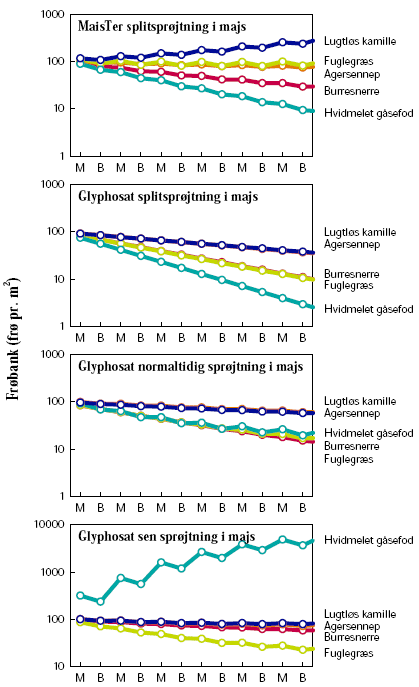
Figur 3.14. Scenarie 2: Sædskifte med konventionel vs. GT-majs.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”M”=Majs, ”B”=Vårbyg.
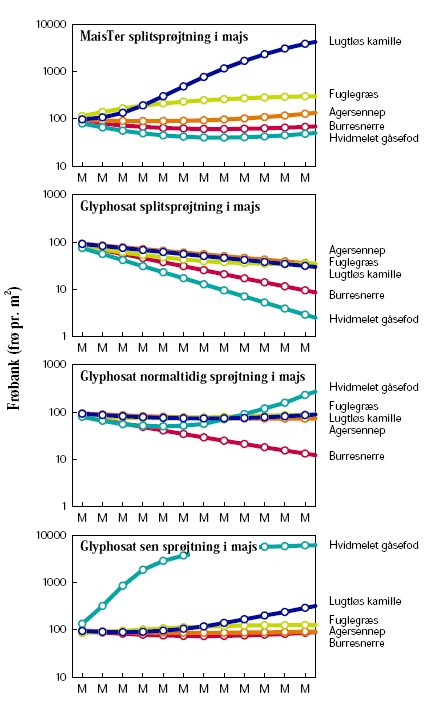
Figur 3.15. Scenarie 3: Ensidigt sædskifte, konventionel vs. GT-majs.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”M”=Majs.
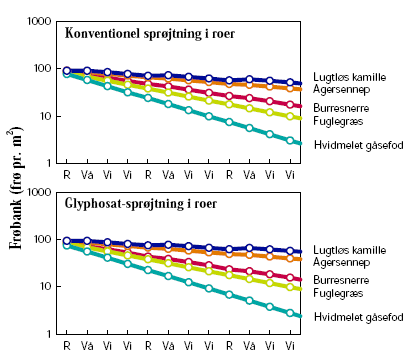
Figur 3.16. Scenarie 4: Sædskifte, med konventionelle vs. GT-roer.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”R”=Roer, ”Vå”=Vårbyg, ”Vi”=Vinterhvede.
3.2 Herbicideffekter på hegnets flora
3.2.1 Ukrudtsbiomasse og frøproduktion
Effekten af glyphosat og MaisTer på biomasseproduktionen af alm. hvene, fåresvingel og mælkebøtte kunne beskrives ved hjælp af S-formede doseringskurver (tabel 3.8). Derimod var det kun muligt at beskrive frøproduktionen af mælkebøtte og alm. hvene (kun MaisTer) med S-formede kurver. På alm. hvene var effekten af glyphosat på frøproduktionen for høj, mens effekten af MaisTer var meget variabel. Fåresvingel satte slet ikke frø sandsynligvis på grund af utilstrækkelig vernalisering.
På mælkebøtte øgedes ED50-doseringen af begge herbicider med stigende udviklingstrin både for biomasse og frøproduktion. Effekten var mere udtalt på den overjordiske end den underjordiske (resultater ikke vist) biomasse.
For mælkebøtte var der ikke nogen forskel på ED90-doseringen af glyphosat for biomasse og frøsætning, hvilket indikerer, at planterne ikke kunne kompensere for skaden på lang sigt. Det modsatte var tilfældet for MaisTer, hvilket tyder på, at for dette herbicid var vækststandsningen kun midlertidig. Senere kunne planterne genoptage væksten og sætte frø. Det betyder, at der skal opnås en større effekt af MaisTer sammenlignet med glyphosat på biomassen for at hindre frøproduktion.
Resultaterne viser, at for græsserne havde udviklingstrinnet langt mindre betydning for effekten end for de tokimbladede arter (tabel 3.1 og 3.8). Undtagelsen var fåresvingel, hvor ED90-doseringerne af begge herbicider øgedes stærkt i strækningsfasen. En af årsagerne til, at der ikke var så stor forskel på doseringsbehovet på de to første udviklingstrin af græsserne, kan være, at græsserne på de tidlige udviklingstrin har meget oprette blade, hvor der kun sker en begrænset afsætning.
Tabel 3.8. Reduktion af overjordisk plantebiomasse og frøsætning som respons på herbiciddosering. ED90 er den beregnede dosis for 90% reduktion. 95%-konfidensintervallet er vist i parentes.
| Ukrudtsart og herbicid | Udviklingstrin | Biomasse ED90 |
Frøsætning ED90 |
|||||
| Alm. hvene Glyphosat |
1 sideskud | 50,4 | (39,6...61,2) | |||||
| (g as./ha) | 6 sideskud | 43,2 | (28,8...64,8) | |||||
| strækning | 25,2 | (18,0...36,0) | ||||||
| MaisTer | 1 sideskud | 1,30 | (0,992...1,61) | 2,08 | (0,310...4,50) | |||
| (g as./ha) | 6 sideskud | 2,08 | (1,71...2,39) | 0,900 | (0...5,80) | |||
| strækning | 2,08 | (1,71...2,48) | ||||||
| Fåresvingel Glyphosat |
6 sideskud | 115 | (93,6...137) | |||||
| (g as./ha) | 20 cm høj | 122 | (101...162) | |||||
| strækning | 227 | (180...317) | ||||||
| MaisTer | 6 sideskud | 6,60 | (5,49...7,69) | |||||
| (g as./ha) | 20 cm høj | 7,91 | (5,49...10,4) | |||||
| strækning | >124 | |||||||
| Mælkebøtte Glyphosat |
6 blade | 32,4 | (25,2...39,6) | 64,8 | (54,0...79,2) | |||
| (g as./ha) | 9 blade | 61,2 | (43,2...101) | 101 | (82,8...130) | |||
| 12 blade | 82,8 | (54,0..220) | 122 | (101...155) | ||||
| MaisTer | 6 blade | 0,279 | (0,217...0,775) | 0,868 | (0,713...0,992) | |||
| (g as./ha) | 9 blade | 0,589 | 1,18 | (1,18...1,61) | ||||
| 12 blade | 2,11 | (2,11...2,79) | ||||||
Alm. hvene og mælkebøtte var væsentlig mere følsomme over for begge herbicider end fåresvingel. Følsomheden af alm. hvene og mælkebøtte (tabel 3.8) var ikke væsentlig forskellig fra følsomheden af ukrudtsarter som sort natskygge og hanespore (tabel 3.1). Af de undersøgte arter vil fåresvingel således være den art, der vil blive påvirket mindst af sprøjtedrift fra disse to herbicider.
3.2.2 Vækststandsning
På flerårige arter fremkalder sublethale doseringer ofte en vækststandsning i en periode efter sprøjtning, hvorefter planten genoptager væksten ved at translokere reserver fra de underjordiske forrådsorganer. En sådan midlertidig væksthæmning kan have betydning for plantens konkurrenceevne.
Generelt var der stor variation i resultaterne af dosisresponsforsøgene på mælkebøtte og stor nælde. Det er et velkendt fænomen, at variationen i herbicidforsøg med flerårige arter er større end variationen i forsøg med enårige arter. Normalt vil man derfor øge antallet af gentagelser i forsøget for at opnå en større statistisk sikkerhed, men i dette forsøg, hvor der var inkluderet 5 forskellige høsttider (4 forskellige tidspunkter efter sprøjtning samt et hold planter til biomarkørtest), ville et øget antal gentagelser betyde en voldsom stor forøgelse af antallet af potter. Resultaterne må således tages med forbehold og kan udelukkende indikere, om det er muligt at afgøre, om der er tale om midlertidig væksthæmning og i givet fald anslå varigheden af en sådan væksthæmning.
3.2.2.1 Væksthusforsøg
En midlertidig vækststandsning er kendetegnet ved, at sluteffekten er mindre end effekten målt kort tid efter sprøjtning. Da forsøgene er opgjort som tørvægt relativt til ubehandlede planter (figur 3.17 og 3.18), er en midlertidig vækststandsning tilstede, når kurven for sluteffekt ligger over kurverne for relativ tørvægt 1, 2 og 3 uger efter sprøjtning.
3.2.2.2 Effekt af glyphosat i væksthus
For glyphosat på frøfremspiret mælkebøtte ses, at den relative biomasse 1, 2 og 3 uger efter sprøjtning var højere end eller lig med slutbiomassen (figur 3.17). Der er her tale om en stigende effekt med tid efter sprøjtning, hvilket også for de højeste doseringers vedkommende ses af, at biomassen 2 og 3 uger efter sprøjtning er lavere end 1 uge efter sprøjtning. På rodfremspiret mælkebøtte har de samme doseringer ikke givet nogen effekt på slutbiomassen. Her ses op til 60% effekt af flere doseringer ved høst kort tid efter sprøjtning. Væksthæmningens varighed afhænger af doseringen, og i de 4 laveste doseringer genoptog planterne væksten efter 3 uger.
På stor nælde blev der opnået høj sluteffekt på de frøfremspirede planter. Doseringerne blev reduceret på de rodfremspirede planter, og her blev der kun opnået ca. 20% effekt. Generelt er effekten af samme dosering væsentlig lavere på de rodfremspirede sammenlignet med frøfremspirede planter. I lighed med effekterne på mælkebøtte er der for de frøfremspirede planters vedkommende ikke tale om midlertidig vækststandsning, men om en fremadskridende og permanent vækststandsning. Det noget varierende forløb af kurverne for biomasse på rodfremspirede planter gør det vanskeligt at drage konklusioner. Sluteffekten er i flere tilfælde mindre end effekten kort tid efter behandling, hvilket indikerer, at der er tale om midlertidige væksthæmninger. Det parallelle forløb og placeringen af kurverne 1, 2 og 3 uger efter sprøjtning indikerer, at væksten er genoptaget 3 uger efter sprøjtning.
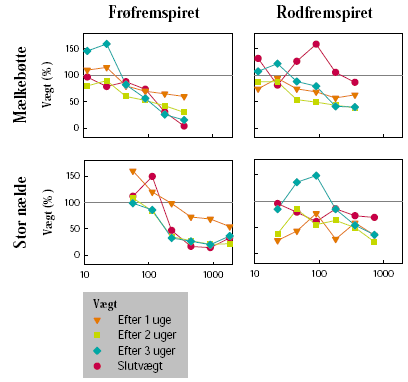
Figur 3.17. Glyph osateffekt på væksten af biomasse.
Biomassen blev målt 1, 2 og 3 uger efter sprøjtning på planter fremspiret fra frø eller rod. Slutvægten måltes efter 11 (fra frø) eller 24 (fra rod) uger for mælkebøtte og efter 13 (fra frø) eller 20 (fra rod) uger for stor nælde. Vægten er angivet som procent af vægten af ubehandlede planter.
3.2.2.3 Effekt af MaisTer i væksthus
På begge arter blev der anvendt højere doseringer af MaisTer på de rodfremspirede end på de frøfremspirede planter. Effekten over for mælkebøtte var væsentlig lavere på rodfremspirede end på frøfremspirede planter (figur 3.18). På de frøfremspirede planter var der kun i de to laveste doseringer tale om midlertidig vækststandsning, og væksten synes at være genoptaget 3 uger efter sprøjtning. På de rodfremspirede planter var der midlertidig vækststandsning i de 3 laveste doseringer, og her var væksten stadig hæmmet 3 uger efter sprøjtning. På stor nælde var sluteffekten på både frø- og rodfremspirede planter større end eller lig med effekten kort tid efter sprøjtning, og der var således ikke tale om midlertidig væksthæmning.
3.2.2.4 Effekter i væksthuset generelt
Generelt må det konkluderes, at frøfremspirede planter af flerårige arter er væsentligt mere følsomme end rodfremspirede planter. Midlertidige væksthæmninger blev i forsøgene observeret hos såvel frø- som rodfremspirede planter men er formentlig hyppigt forekommende hos planter, som kan translokere næring fra forrådsorganer til nye skud. I flere tilfælde synes væksten i forsøgene genoptaget 3 uger efter sprøjtning, men varigheden afhænger formodentlig af effekten af de anvendte doseringer, således at jo større effekt der opnås, jo længerevarende vækststandsning må forventes. Resultaterne viser, at der i modeller for flerårige arters vækst med herbicidpåvirkning bør indbygges en vækststandsning i minimum 3 uger, når planterne eksponeres for herbiciddoseringer, som forventes at give mellem 15 og 50% sluteffekt. Ved sluteffekter højere end 50% er der oftest tale om en permanent vækststandsning, og ved mindre end 15% effekt vil vækststandsningen være så kort, at den ikke skal indregnes.
En permanent langtidseffekt på flerårige arter forudsætter, at de underjordiske organer ødelægges. Translokationen af aktivstof ud i disse organer afhænger af, hvor systemisk herbicidet er, men flerårige arter med et udbredt rodsystem vil kunne overleve adskillige års eksponering af selv et meget systemisk herbicid som glyphosat.
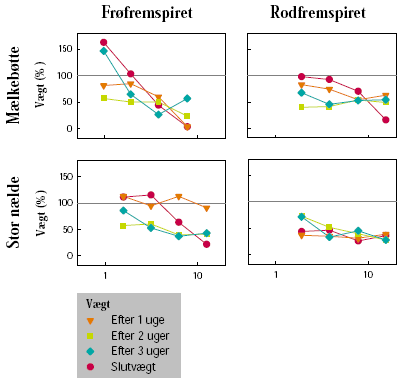
Figur 3.18. MaisTer-effekt på væksten af biomasse.
Biomassen blev målt 1, 2 og 3 uger efter sprøjtning på planter fremspiret fra frø eller rod. Slutvægten måltes efter 11 (fra frø) eller 24 (fra rod) uger for mælkebøtte og efter 13 (fra frø) eller 20 (fra rod) uger for stor nælde. Vægten er angivet som procent af vægten af ubehandlede planter.
3.2.2.5 Markforsøg
Den uforstyrrede vækst var for begge arter karakteriseret ved en tilvækst gennem hele sæsonen. I efteråret afsattes tørstof fra bladmassen ned i rødderne, som fungerer som reserveorgan vinteren over (figur 3.19).
De to arter reagerede modsat på herbiciderne: Mælkebøtte blev efter en kortvarig hæmning stimuleret til vækst og opnåede derved en større slutvægt end usprøjtede planter. For stor nælde derimod havde væksthæmningen blivende effekt, og for den art blev slutvægten mindre som følge af sprøjtningen (figur 3.19).
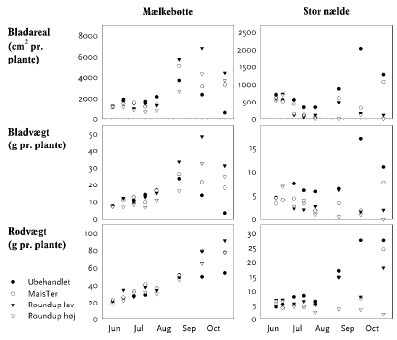
Figur 3.19. Herbicidforsøg i marken, Flakkebjerg 2006.
MaisTer: Mælkebøtte (25 g aktivstof/ha) og stor nælde (100 g aktivstof/ha).
Glyphosat: Mælkebøtte (lav 120 g aktivstof/ha, høj 360 g aktivstof/ha) og stor nælde (lav 540 g aktivstof/ha, høj 1080 g aktivstof/ha). Data vist som gns.±s.e.
3.2.3 Planters konkurrenceforhold
3.2.3.1 Simpelt respons uden konkurrence
Resultaterne af enkeltartsforsøg med alm. hvene og fåresvingel udført sideløbende med konkurrenceforsøget er vist i tabel 3.9. Fåresvingel var mere modstandsdygtig end alm. hvene og krævede en dosis, der var 2 til 9 gange større for at opnå samme effekt.
3.2.3.2 PCENV-estimering af respons på glyphosat
Efter en Box-Cox transformation (λ1 =-0,8λ2 =1) af både data og model viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 88% af den observerede variation.
Tabel 3.9. Reduktion af overjordisk plantebiomasse som respons på herbiciddosering. ED10, ED50 og ED90 er de beregnede doser for 10%, 50% og 90% reduktion. 95%-konfidensintervaller er vist i parentes.
| Herbicid og ukrudtsart |
ED10 | ED50 | ED90 | ||||||
| Glyphosat (g as./ha) Alm. hvene |
18,0 | (10,8...25,2) | 36,0 | (28,8...46,8) | 75,6 | (61,2...86,4) | |||
| Fåresvingel | 36,0 | (10,8...57,6) | 115 | (79,2...151) | 374 | (256...486) | |||
| MaisTer (g as./ha) Alm. hvene |
0,372 | (0,186...0,589) | 0,992 | (0,682...1,30) | 2,88 | (2,20...3.60) | |||
| Fåresvingel | 3,29 | (1,80...4,90) | 6,11 | (4,71...7,60) | 36,5 | (8,90...13,5) | |||
De estimerede parametre i konkurrencemodellen måler effekten af glyphosat på hele responskurven, og glyphosat havde en signifikant positiv effekt på biomassen af fåresvingel ved lav tæthed i de beskedne doser, som anvendtes i forsøget og en signifikant negativ effekt på biomassen af alm. hvene ved høje tætheder. Der var en signifikant negativ effekt af glyphosat på alm. hvenes konkurrenceevne overfor fåresvingel (tabel 3.10). Glyphosat skubbede altså til konkurrenceforholdet mellem fåresvingel og alm. hvene til fordel for fåresvingel.
Tabel 3.10. PCENV-parameterværdier for responset på glyphosat for fåresvingel (art 1) og alm. hvene (art 2). Værdier estimeret ved hjælp af maksimum likelihood og testet ved hjælp af likelihood ratio.
| Parameter | Værdi | Signifikans | ||
| a1 | –0,17 | - | ||
| a2 | –9,29 | - | ||
| b1 | –4,51 | - | ||
| b2 | –47,58 | - | ||
| c1 | –2,365 | - | ||
| c2 | 1,152 | - | ||
| d1 | –0,163 | Exp(1); P = 0,011 | ||
| d2 | 1,983 | |||
| f1 | 0,0459 | Exp(1); P = 0,016 | ||
| f2 | 2,663 | |||
| 1 | –0,00684 | ?0; P = 0,007 | ||
| 2 | –0,0616 | = 0; P = 0,17 | ||
| 0,00204 | = 0; P = 0,67 | |||
| 0,0674 | 0; P < 0,001 | |||
| 0,0211 | = 0; P = 0,072 | |||
| –0,00633 | 0; P < 0,001 |
ED10 af glyphosat på alm. hvene ved forskellige tætheder af fåresvingel og alm. hvene blev beregnet ud fra ligning 2 i afsnit 2.2.3. Medianværdien af ED10 for alm. hvene afhang af tætheden af fåresvingel og alm. hvene (figur 3.20), dog var der en betydelig usikkerhed på de beregnede ED10-værdier(figur 3.21 og 3.22). Den beregnede ED10 fra konkurrenceforsøget er sammenlignelig med den estimerede ED10 fra standardtesten (= 19.0 med et 95% konfidensinterval mellem 12.1 og 25.9; fra tabel 3.9).
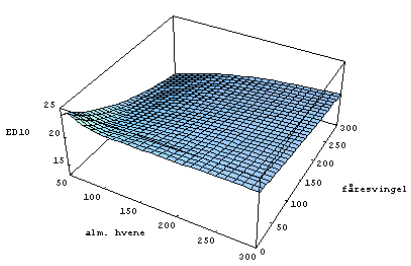
Figur 3.20. Effekten af glyphosat på alm. hvene i konkurrence med fåresvingel.
Den beregnede medianværdi af ED10 (10% biomassetab) for alm. hvene som funktion af tætheden af fåresvingel og alm. hvene (m–2).
Trods den betydelige usikkerhed på den beregnede ED10 var der en signifikant men lille effekt af den intra-specifikke konkurrence på den beregnede ED10 for alm. hvene, således at der ved højere tæthed af planten kræves en relativ lavere dosis glyphosat for at reducere biomassen med 10% (ED10(0,100)-ED10(0,200) > 0, P = 0.03). Denne lavere dosis er dog ubetydelig (median = 0.06) i forhold til usikkerheden på de beregnede ED10 værdier (figur 3.21).
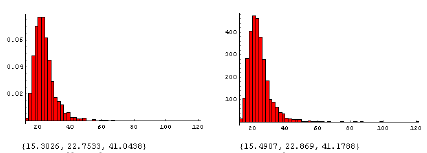
Figur 3.21. Effekt af glyphosat på alm. hvene i intraspecifik konkurrence.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 200 m-2 (til venstre) og 100 m-2 (til højre). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
Derimod var der en betydelig signifikant effekt af fåresvingel på ED10 for alm. hvene (ED10(0,100)-ED10(100,100) > 0, P = 0.03), således at glyphosat har en kraftigere effekt på alm. hvene, når alm. hvene konkurrerer med fåresvingel (figur 3.22). Den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10%, er ca. 16% lavere, hvis alm. hvene konkurrerer med fåresvingel i forholdet 1:1 i forhold til, hvis alm. hvene kun konkurrerer med sig selv. Hvis forholdet af fåresvingel er højere end 1:1, da reduceres den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10% yderligere (figur 3.20).
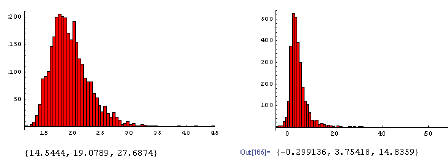
Figur 3.22. Effekt af glyphosat på alm. hvene i konkurrence med fåresvingel.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 100 m–2 for alm. hvene og 100 m–2 for fåresvingel (A) og fordelingen af ED10 (alm. hvene:100 m–2) – ED10 (alm. hvene:100 m–2, fåresvingel: 100 m–2). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
3.2.3.3 PCENV-estimering af respons på MaisTer
Efter en Box-Cox transformation ![]() af både data og model (2.4.2.2.1) viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 66% af den observerede variation.
af både data og model (2.4.2.2.1) viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 66% af den observerede variation.
De estimerede parametre i konkurrencemodellen måler effekten af MaisTer på hele responskurven, og MaisTer havde en signifikant positiv effekt på biomassen af fåresvingel ved både lav og høj tæthed i de beskedne doser, som anvendtes i forsøget, og en signifikant negativ effekt på biomassen af alm. hvene ved både lave og høje tætheder. Der var en signifikant positiv effekt af MaisTer på fåresvingels konkurrenceevne overfor alm. hvene og omvendt en signifikant negativ effekt af MaisTer på alm. hvenes konkurrenceevne overfor fåresvingel (tabel 3.11).
Tabel 3.11. PCENV-parameterværdier for responset på MaisTer for fåresvingel (art 1) og alm. hvene (art 2). Værdier estimeret ved hjælp af maksimum likelihood og testet ved hjælp af likelihood ratio.
| Parameter | Værdi | Signifikans | ||
| a1 | 9.68327 | - | ||
| a2 | –21.5456 | - | ||
| b1 | –67.8936 | - | ||
| b2 | –50.5823 | - | ||
| c1 | –0.988756 | - | ||
| c2 | 1.81707 | - | ||
| d1 | 2.76147 | ≠Exp(1); P < 0,001 | ||
| d2 | 2.13329 | |||
| f1 | 4.39852 | ≠Exp(1); P < 0,001 | ||
| f2 | 2.77588 | |||
| 1 | –12.6327 | 0; P = P < 0,001 | ||
| 2 | 4.95714 | 0; P = P < 0,001 | ||
| –2.03603 | = 0; P = 0,69 | |||
| 3.11466 | 0; P < 0,001 | |||
| 0.431216 | 0; P < 0,001 | |||
| –0.237765 | 0; P < 0,001 |
Effekten af MaisTer på alm. hvene ved forskellige tætheder af fåresvingel og alm. hvene blev beregnet ved ED10 [ligning 2]. Medianværdien af ED10 for alm. hvene afhang af tætheden af fåresvingel og alm. hvene (figur 3.23), dog var der en betydelig usikkerhed på de beregnede ED10 (figur 3.24 og 3.25). Den beregnede ED10 fra konkurrenceforsøget er sammenlignelig med den estimerede ED10 fra standardtesten (= 0,4 med et 95% konfidensinterval mellem 0,2 og 0,4; fra tabel 3.9).
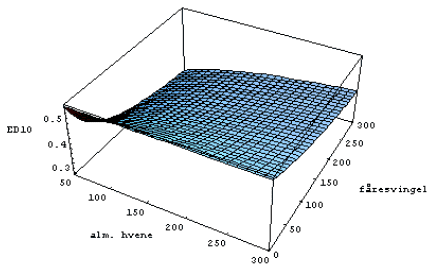
Figur 3.23. Effekten af MaisTer på alm. hvene i konkurrence med fåresvingel.
Den beregnede medianværdi af ED10 (10% biomassetab) for alm. hvene som funktion af tætheden af fåresvingel og alm. hvene (planter pr. m²).
I modsætning til glyphosat var der ingen signifikant effekt af den intraspecifikke konkurrence på den beregnede ED10 for MaisTer på alm. hvene (figur 3.24; P = 0,78). Men ligesom for glyphosat var der en betydelig signifikant effekt af fåresvingel på ED10 for MaisTer på alm. hvene (ED10(0,100)-ED10(100,100) > 0, P = 0,03), således at MaisTer havde en kraftigere effekt på alm. hvene, når den var i konkurrence med fåresvingel end uden (figur 3.25). Den dosis MaisTer, som kræves for at reducere biomassen af alm. hvene med 10%, er ca. 33% lavere, hvis alm. hvene konkurrerer med fåresvingel i forholdet 1:1 i forhold til, hvis alm. hvene kun konkurrerer med sig selv. Hvis forholdet af fåresvingel er højere end 1:1, da reduceres den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10% yderligere (figur 3.23).
ED10 for MaisTer på fåresvingel kunne ikke estimeres troværdigt ud fra ligning 2 på grund af for lave herbiciddoser i konkurrenceforsøget (resultater er ikke vist).
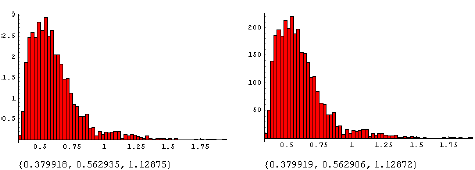
Figur 3.24. Effekt af MaisTer på alm. hvene i intraspecifik konkurrence.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 200 m-2 (til venstre) og 100 m-2 (til højre). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
Figur 3.25. Effekt af MaisTer på alm. hvene i konkurrence med fåresvingel.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 100 m–2 for alm. hvene og 100 m–2 for fåresvingel (A) og fordelingen af ED10 (alm. hvene:100 m–2) – ED10 (alm. hvene:100 m–2, fåresvingel: 100 m–2). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
3.2.4 Plantesamfundets succession
3.2.4.1 Frøbankens påvirkning af glyphosat og kvælstof
Det totale antal frø, der spirede og udviklede sig til identificerbare planter, var 7906. Fire spirer udviklede sig aldrig til planter, der kunne artsbestemmes. Tokimbladede arter var talrigest repræsenteret og udgjorde 70% af det samlede antal fremspirede planter. Knap 45% eller 3522 planter var en- eller toårige arter og tilhører således en gruppe, der for at være repræsenteret i floraen skal forekomme i frøbanken.

Figur 3.26. Fotoet viser 4 spiringsbakker udvalgt fra en tilfældig transekt. Prøverne er fra plot, der har modtaget 100 kg N/ha, og bakkerne er opstillet således, at de fra venstre mod højre indeholder prøver fra plot, der modtaget en stigende glyphosatdosering (0, 14, 4, 72 og 360 g as./ha).
Variansanalysen viste, at glyphosat reducerede frøbanken med stigende dosis (P=0,008, figur 3.26), mens gødskningen ingen effekt havde (P=0,12). Der var ingen interaktion mellem de to variable. Den samlede mængde af spiringsdygtige frø i frøbanken var afhængig af den behandling plottet havde modtaget (figur 3.27)
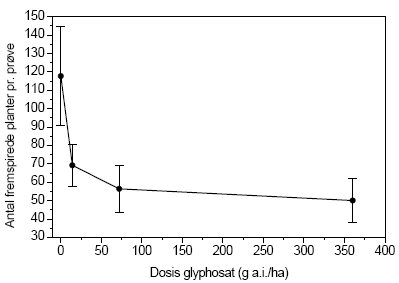
Figur 3.27. Total fremspiring af frø (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, som funktion af årlig glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
En opdeling af de fremspirede frø i en- og tokimbladede arter (figur 3.28) viste, at tokimbladede arter udgjorde langt den største del af planterne, og at antallet af disse faldt mest markant (godt 60 %) med stigende glyphosatdosis (P=0,03), mens antallet af enkimbladede planter var betydelig lavere og var relativt upåvirket af glyphosatdosis (P=0,07). Gødskning påvirkede frøbanken positivt for de tokimbladede (P=0,001), mens de enkimbladede var upåvirkede (P=0,18). Ved det højeste kvælstofniveau (100 kg N/ha) var der dog et tydeligt fald i antallet af spiringsdygtige frø med stigende glyphosatdosering.
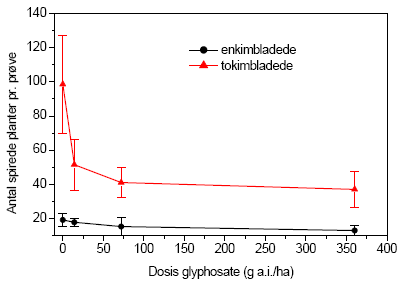
Figur 3.28. Fremspiring af planter (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, fordelt på en- og tokimbladede arter som funktion af glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
Den samlede fremspiring af en- og toårige arter (figur 3.29) viste en negativ påvirking af frøbanken med stigende glyphosatdosering (P=0,002) og en positiv påvirkning af gødskning (P=0,0002). Der var ingen interaktion mellem parametrene.
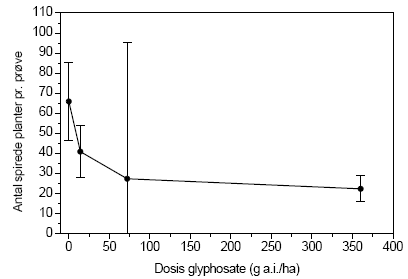
Figur 3.29. Fremspiring af en- og toårige arter (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, som funktion af glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
Der blev fundet 40 plantearter i frøbanken, hvoraf 33 var tokimbladede og 7 enkimbladede. Fordelingen af spirede frø på de enkelte arter og behandlinger fremgår af Bilag C. Blandt de enkimbladede var en art en hybrid mellem græsserne alm. hvene (Agrostis capillaris) og stortoppet hvene (A. gigantea). Blandt de oprindeligt såede 33 arter, hvoraf 27 var etablerede og fortsat tilstede i plottene ved sidste forudgående registrering i 200324, fandtes spiringsdygtige frø af 17 af disse arter. De øvrige 23 arter var dels almindelige markukrudtsarter, en almindelig skovukrudtsart (alm. gederams, Epilobium angustifolium) og træet dunbirk (Betula pubescens).
Fire arter, rejnfan (Tanacetum vulgare), filtbladet kongelys (Verbascum thapsus), alm. stedmoderblomst (Viola tricolor) og stortoppet hvene (Agrostis gigangtea), udgjorde samlet 85% af de fremspirede planter og henholdsvis 19, 24, 14 og 28%. De øvrige arter var ofte ganske fåtallige, og kun 4 arter (mælkebøtte, prikbladet perikon, fåresvingel og hvene-hybriden) blandt de resterende bidrog med mere end 1%.

Figur 3.30. Forsøglokaliteten nær Kalø i august 2006.
3.2.4.2 Vegetationens påvirkning af glyphosat og kvælstof
Selv med det blotte øje kunne man se, at vegetationen var påvirket af behandlingerne (figur 3.30). Sprøjtningen med glyphosat forårsagede større områder med bar jord eller død vegetation to uger efter behandlingen i sammenligning med usprøjtede plots (figur 3.31), og der skete ikke noget fald i andelen af bar jord over den treårige projektperiode (dvs. år 4-6 i successionen)(figur 3.32). For hovedparten af arterne faldt dækningen (figur 3.32) og biomassen (figur 3.34) med stigende glyphosatdosering. Det gjaldt f.eks. alm. hvene, kvik, stortoppet hvene og rejnfan. Fåresvingel var den eneste art, der havde en stigende dækning og biomasse med stigende glyphosatdosering.
Kvælstof havde en positiv virkning på dækning og biomasse af de to græsser, kvik og stortoppet hvene (P<0,0001), men for de øvrige arter var der ingen (f.eks. rejnfan) eller en negativ effekt af kvælstof.
Der var enkelte signifikante tredjeordens interaktioner (år´art´plot), hvor udviklingen af en enkelt art var anderledes end de viste gennemsnit (figur 3.32 og 3.34), men det drejede sig højst om ét plot pr. behandling.
Græsser udgjorde ved alle behandlinger og tidspunkter omkring 50% eller mere af den samlede dækning. Gennem årene var det dog forskellige græsarter, der bidrog til dækningen. I 2005 dominerede alm. hvene, fåresvingel og stortoppet hvene, mens kvik havde en ubetydelig dækning. Dækningen af kvik øgedes imidlertid betydeligt gennem projektperioden navnlig ved det højeste kvælstofniveau, hvor kvik i 2007 nåede en dækning på 1/3 af arealet undtagen ved den højeste dosis af glyphosat, hvor dækningen af de øvrige græsarter navnlig fåresvingel var højere (P<0,0001)(figur 3.32).
Fåresvingel var den dominerende græs ved højeste dosis af glyphosat uanset kvælstofniveauet. Ved lavere glyphosatdosis var alm. hvene dominerende, medens kvik var dominerende ved højt kvælstofniveau (figur 3.32).
Artsantallet var generelt faldende med stigende glyphosatdosis (figur 3.33). Blandt de 33 arter, der blev sået i 2001 (Bilag C), spirede og etablerede alle arter sig med undtagelse af knoldet mjødurt (Filipendula vulgaris). Agertidsel (Cirsium arvense) blev ikke registreret i det tidligere projekt24 men blev fundet i hele denne projektperiode 2005-2007. Desuden blev få levedygtige frø registreret i frøbanken i 2004. Hvorvidt agertidsel er etableret fra de oprindeligt såede frø eller fra frø, der efterfølgende er spredt til området, vides ikke, men agertidsel har frø, der overlever længe i frøbanken109. På trods af at der blev fundet en stor mængde spiringsdygtige frø i frøbanken, udgjorde andre arter end de tre græsser alm. hvene, fåresvingel og kvik en faldende andel af den samlede dækning ved alle behandlinger (figur 3.32).
Seksogtyve arter af de 33 såede arter findes fortsat i forsøgsområdet. Blandt de seks forsvundne arter er tre arter alm. dværgløvefod (Aphanes arvensis), enårig rapgræs (Poa annua) og kornblomst (Centaurea cyanus) ikke registreret inden for plottene siden 2001, haremad (Lapsana communis) og alm. kællingetand (Lotus corniculatus) ikke siden 2003 og alm. pimpinelle (Pimpinella saxifraga) og alm. agermåne (Agrimonia eupatoria) ikke siden 2005. Kun 9 arter forekommer hyppigt, se Bilag C.
Blandt de nye arter, der er registreret, forekommer kun alm. røllike (Achillea millefolim), gul snerre (Galium verum), smalbladet rapgræs (Poa annua) og rødknæ (Rumex acetosella) ved gentagne registreringer i den samme blok og kan dermed anses for etablerede. Blandt de sporadisk forekommende arter er en del enårige ukrudtsarter f.eks. hvidmelet gåsefod (Chenopodium album), burresnerre (Galium aparine), vej pileurt (Polygonum aviculare) og snerlepileurt (Fallopia convolvulus).

Figur 3.31. Glyphosateffekt på etableret vegetation.
Usprøjtet plot (venstre) og plot sprøjtet med 360 g glyphosat pr. ha (højre) 2 uger efter sprøjtning.
Den mest markante forandring i forsøgsperioden (2005-2007) var den voldsomme vækst af kvik (figur 3.32 og 3.34). I 2003 udgjorde kvik <0,1% af den høstede biomasse. I 2007 var biomassen steget til 35-45 g/m² ved høj kvælstoftilførsel uanset glyphosatdoseringen. Stigningen modsvaredes af et tilsvarende fald i biomassen af alm. hvene (figur 3.34) Biomassen af fåresvingel var højest ved de høje glyphosatdoseringer og nogenlunde uforandret gennem forsøgsperioden (figur 3.34), selvom dækningen steg i perioden (figur 3.32).
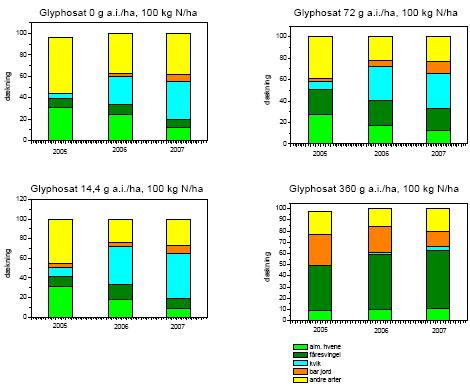
Figur 3.32. Udviklingen i dækningen fra 2005 til 2007 af udvalgte arter, andre arter samt af bar jord.
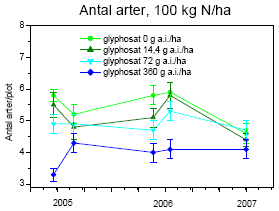
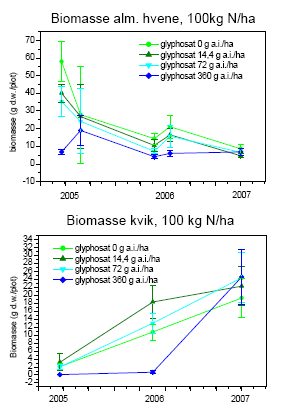
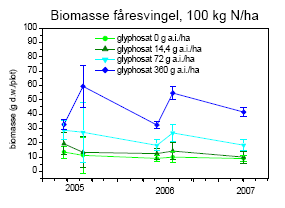
Figur 3.34. Udvikling i plantebiomasse fra 2005 til 2007. Data viser gns.±s.e. (n=10).
3.2.4.3 Vegetationens påvirkning af MaisTer
Undersøgelsen af effekter af MaisTer var ikke designet således, at vi kan adskille effekten af gødskning fra effekten af herbicidet. De effekter, vi ser, er kombinationseffekter. Ved sammenligning af udviklingen med og uden herbicidbehandling kan faldet i antal arter (figur 3.35) og i dækningen af fåresvingel og rejnfan (figur 3.36) umiddelbart efter første sprøjtning i 2005 bedst forklares som en herbicideffekt, mens alm. hvene i mindre grad påvirkedes af MaisTer. Gødskningen stimulerede tydeligvis både alm. hvene og rejnfan, hvorimod dækningen af fåresvingel reduceredes efter kvælstoftilførslen. Fåresvingel er en langsomt voksende græs, der er mest konkurrencedygtig ved lave kvælstofniveauer, hvilket kan forklare, at dækningen af fåresvingel reduceredes ved begge behandlinger.
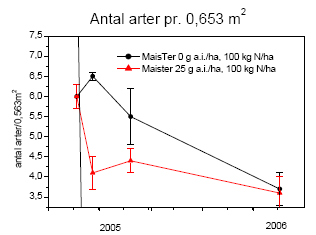
Figur 3.35. Artsantallet efter behandling med MaisTer (25 g as./ha) i forhold til ubehandlet; begge gødet med 100 kg N/ha. Den vertikale linie angiver behandlingstidspunktet. Punkter viser gns.±s.e.
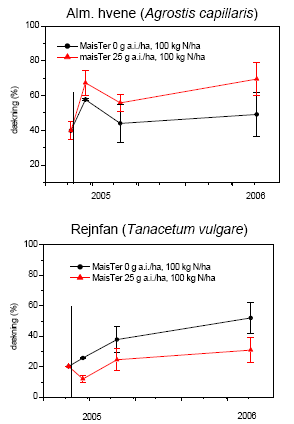
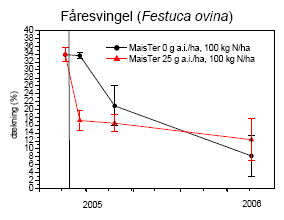
Figur 3.36. Dækningen af tre udvalgte arter efter behandling med MaisTer (25 g as./ha) i forhold til ubehandlet; begge gødet med 100 kg N/ha. Den vertikale linie angiver behandlingstidspunktet. Punkter viser gns.±s.e.
3.2.5 Modellering af fodposeplanternes populationsdynamik
3.2.5.1 Kalibrering af modellen
Modellen blev kalibreret til at simulere væksten af mælkebøtte, kvik, alm. hvene, fåresvingel, agertidsel, rejnfan, stor nælde og vild kørvel ved at tilpasse modellens parametre til beskrivelse af væksten til de indsamlede vækstdata på de otte arter (tabel 2.15). Tilpasningen blev foretaget ved at ændre på parametrene, indtil simuleringen af tørvægt og antal af plantens forskellige overjordiske organer (blade, stængler, knopper, blomster og frugter) stemte tilfredsstillende overens med observerede data. I denne proces blev benyttet både visuel sammenligning af feltdata og modeloutput samt optimering af modellens simuleringer ved hjælp af mindste kvadraters metode. Ved den sidste metode foretog modellen en lang række gennemregninger med tilfældigt valg af udvalgte parametre fra et interval, og efter hver beregning summerede den kvadraterne på forskellen mellem simulerede og observerede data. Den kombination af parametre, der gav det mindste kvadrat, blev så benyttet i den videre parameterisering. Da der er mange lokale minimumspunkter på sammenhængen imellem forskellige kombinationer af vækstparametre og kvadratet på afvigelsen, viste det sig hensigtsmæssigt at benytte en kombination af den visuelle bedømmelse af simuleringerne og mindste kvadraters metode.
Efter at modellen var blevet kalibreret til at simulere væksten af en planteart med hensyn til tørvægt og antal af de forskellige planteorganer, blev den kalibreret til at simulere højdefordelingen af disse organer ved hjælp af en tilsvarende kombination af den visuelle metode og mindste kvadraters metode.
Under kalibreringsprocessen blev der benyttet simuleringer over en seksårig periode, hvor der blev benyttet klimatisk input fra 2000 frem til 2006. Modellens output fra 2005 blev så benyttet i sammenligningerne med de observerede data, der stammede fra netop dette år. Modellen blev kalibreret ved hjælp af disse langtidssimuleringer, fordi de indsamlede data stammede fra flerårige planter, der havde vokset på de pågældende steder igennem flere år. FieldEco kan bringes til at simulere en plantes vækst ved hjælp af mange forskellige kombinationer af parametre, men ved at benytte tilpasning til både tørvægt og antal af alle overjordiske organer ved langtidssimuleringer er der sikret en realistisk parameterisering. Hvis parameteriseringen ikke lå indenfor realistiske rammer, opstod der ubalance imellem plantens forskellige organer i den simulerede plante, og simuleringerne kunne ikke forløbe over flere år.
Effekten af herbiciderne glyphosat og MaisTer blev kalibreret ved at simulere en sprøjtning på en given data og ved at justere effekten af herbicidet på plantens totale overjordiske biomasse til det observerede effektniveau (observeret i dette projekt eller fra andre kilder) fire uger senere.
Tilpasningsprocessen illustreres her med rejnfan som eksempel. Simuleringen af de observerede data (figur 3.37) viser, at modellen kunne simulere væksten igennem sæsonen ganske godt. Simuleringen af tørvægt af blade og stængler igennem sæsonen passer godt med data, mens det viste sig umuligt at få simuleringerne af antallet af blade sidst på sommeren til at passe med data. Dette skyldes, at rejnfan producerer en stor mængde relativt små blade i den sidste del af sommeren, og FieldEco er ikke programmeret til at simulere en variation i bladstørrelsen igennem sæsonen. Blev modellen kalibreret til at simulere antallet af blade korrekt, blev tørvægten af bladene for høj, og blev modellen kalibreret til at simulere tørvægten korrekt, blev antallet af blade midt på sommeren for lavt. Det var derfor nødvendigt at vælge enten det ene eller det andet, og der blev givet prioritet til tørvægten, da tørvægten er en drivende variabel i modellen. Dette problem gjorde sig også gældende for stor nælde og agertidsel, hvor der også blev givet prioritet til tørvægten. Simuleringen af tørvægten af blomsterstande passer heller ikke helt med data, da det ikke var muligt at få både antal og tørvægt af frugtstande på sidste dato til at passe godt. Det var ikke muligt at simulere et fald i tørvægt samtidig med en stigning i antal, hvilket må skyldes, at de sidst udviklede frugtstande bliver mindre end de først udviklede. Da FieldEco opererer med samme parameterisering af frugtudviklingen igennem hele sæsonen, var det ikke muligt at simulere dette. Der blev derfor også her givet prioritet til at simulere tørvægten godt.
FieldEco blev kalibreret til at simulere de øvrige syv plantearter på samme måde som rejnfan. Der var dog lidt forskel på, hvor mange planteorganer, der indgik i kalibreringen. Ved græsserne gav det ikke mening at skelne imellem stængler og blade, og ved kvik var blomstring og frøsætning så begrænset, at den blev ignoreret, det vil sige, at kun bladene indgik i parameteriseringen af kvik. Hos stor nælde var det meget svært at skelne imellem knopper, blomster (både han- og hunblomster) og frø under feltforhold, hvorfor de blev slået sammen som reproduktive organer.
3.2.5.2 Validering af modellen
Til validering af modellen anvendtes data, der var uafhængige af dem, der blev anvendt til kalibreringen. For mælkebøtte og stor nælde data fra Flakkebjerg 2006 (afsnit 2.2.2) og for alm. hvene, fåresvingel og kvik, anvendtes data fra successionsforsøget ved Kalø (afsnit 2.2.4). Valideringen blev foretaget uden ændring af modellens parameterværdier, hvor intet andet er anført.
For at simulere Flakkebjerg-data blev FieldEco blev sat op til at simulere et enkelt års vækst af enten mælkebøtte eller stor nælde med anvendelse af klimadata (temperatur og globalstråling) fra Forskningscenter Flakkebjergs klimastation
For at simulere Kalø-data blev FieldEco sat op til at simulere konkurrencen imellem fåresvingel, alm. hvene og kvik over en 6-årig periode, hvor der blev sprøjtet med RoundUp og gødsket, som angivet i tabel 2.14. Simuleringerne blev startet med 10 frø pr. ¼ m² af alm. hvene og fåresvingel den 11. april 2001, og med en enkelt kimplante af kvik den 25. juli 2004 ved høj kvælstoftilførsel (100 g N/ha/år). Kvik blev introduceret senere i simuleringen, da kvik indvandrede efter etableringen af forsøget. Der blev anvendt klimadata fra DMUs klimastation i Silkeborg. Simuleringerne blev foretaget med parameteriseringen af plantearterne, som beskrevet i afsnit 2.2.5.
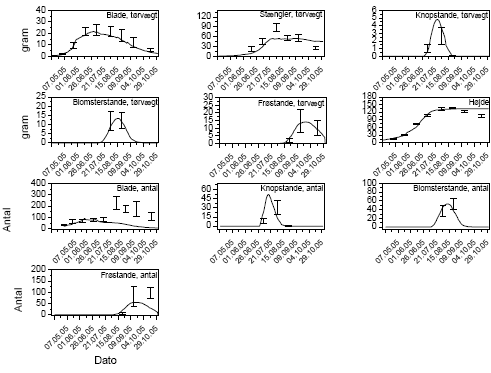
Figur 3.37. Tilpasning af modellen til at simulere parameteriseringsdata på rejnfan
(Tanacetum vulgare). Sammenligning imellem simulering (linien) og observerede værdier (gns.±s.e.).
Mælkebøtte
Feltdata viser en meget kraftig vækst af både rod og skud i august og september, hvilket modellen ikke er i stand til at simulere (figur 3.38). Der er dog foretaget en opjustering af kvælstofniveauet i simuleringen, da dette ikke kan antages at have været som i jorden, hvor planterne til parameterisering blev dyrket. Simuleringen uden anvendelse af herbicider viser stort set den observerede udviklingstendens, men når ikke det høje niveau fra observationerne, hvorimod simuleringen af rodbiomassen skyder klart under observationerne. Modellen er ikke i stand til at simulere faldet i skudbiomasse i juli-august, hvilket antagelig skyldes den tørkesituation, der forekom i denne periode, og som modellen ikke er programmeret til at tage i betragtning.
Modellen er blevet kalibreret til at simulere flerårige planter, mens der her er tale om planter, der er spiret fra frø det samme år. Der er søgt taget højde for dette ved at forhindre blomstring og udsætte allokering af ressourcer til oplagringsorganer til midt på sommeren, men modellen er faktisk ikke kalibreret til at håndtere førsteårsplanter. Tendenserne fra kontrollen gør sig i store træk også gældende for simuleringerne af situationerne med sprøjtninger med MaisTer og glyphosat, dog er modellen i højere grad i stand til at simulere skududviklingen i efterårsperioden. Simuleringen af sprøjteeffekterne fungerer tilfredsstillende ved at opfange tendenserne i udviklingen, det vil sige en reduktion af væksten efter sprøjtningen den 19. juni, hvorefter væksten genoptages. Tendenserne i simuleringerne af situationerne efter sprøjtning med glyphosat er i store træk sammenfaldende med observationerne, selv om niveauet i efteråret heller ikke her bliver fanget af simuleringen.
Det høje niveau i skudvægt i efteråret efter behandlingen med MaisTer oversteg niveauet fra kontrollen, hvilket måske skal tilskrives tilfældig variation af de indsamlede planter. Resultaterne skal i øvrigt ses i lyset af en meget stor variation imellem planterne i efterårsperioden og dermed følgende risiko for fluktuerende resultater.
Med hensyn til giftvirkningen i simuleringerne var det nødvendigt at reducere effekten af både glyphosat og MaisTer betydeligt i forhold til parameteriseringen ifølge resultaterne fra væksthus.
Stor nælde
Modellen simulerer kontrollen ganske godt men er ligesom ved mælkebøtte ikke i stand til at simulere vækststandsningen under tørken i juli-august, da den ikke er programmeret til det (figur 3.39). Simuleringen uden for denne periode er god. Med hensyn til simuleringen af effekten af glyphosat og MaisTer simuleres væksthæmningen ganske godt, men modellen overvurderer skudvæksten efter sprøjtningen. Dette skyldes antagelig igen tørken i juli-august, som sammen med giftvirkningen har nogle konsekvenser for planten, som FieldEco ikke tager med.
Simuleringerne underestimerer rodvægten i starten af sæsonen i alle simuleringer, hvilket hænger sammen med, at modellen ikke er blevet kalibreret til at simulere denne situation, hvor planterne er spiret fra frø det samme år. Ligesom for mælkebøtte er modellen kun blevet kalibreret til situationen med flerårige planter. Derfor er det faktisk ganske tilfredsstillende, at modellen alligevel fanger udviklingen i rodudvikling på disse førsteårsplanter, selv om den ikke rammer helt præcist fra starten, hvor spiringen er tættest på.
Almindelig hvene, fåresvingel og kvik
Der blev ikke foretaget ændringer i parameteriseringen af planternes vækst for at simulere de observerede data på fåresvingel, alm. hvene og kvik fra Kalø-eksperimenterne. Det var dog nødvendigt at foretage en justering af den tilgængelige mængde kvælstof i forhold til data. Dette blev udelukkende gjort i forhold til data uden sprøjtning med glyphosat (figur 3.40). De første simuleringer med glyphosatsprøjtninger viste, at planterne fra Kalø-felterne var klart mindre sensitive overfor dette stof, end data fra væksthuset viste. Det var derfor nødvendigt at reducere effekten af glyphosat for alle tre arter betydeligt.
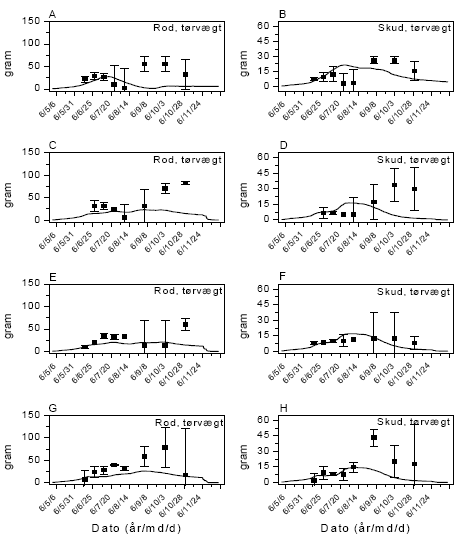
Figur 3.38. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af mælkebøtte. Kontrol (A,B), MaisTer (C, D), glyphosat lav (E,F) og glyphosat høj (G,H).
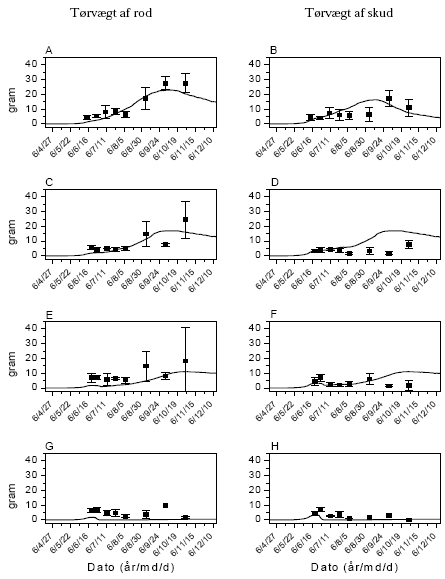
Figur 3.39. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af stor nælde. Kontrol (A,B), MaisTer (C, D), glyphosat lav (E,F) og glyphosat høj (G,H).
Modellen simulerer de observerede data ved lavt niveau af glyphosat for alle tre arter og viser, at FieldEco er i stand til at simulere plantekonkurrence over flere år (figur 3.41 og 3.42). Ved simulering af behandlingen med højeste dosis glyphosat opstår der imidlertid en underestimering af fåresvingels vækst, mens simuleringen af væksten af kvik og alm. hvene stadigvæk er god (figur 3.43). Problemerne med fåresvingel ved den høje belastning med glyphosat skyldes muligvis, at FieldEco ikke fanger fåresvingels tilvækst om vinteren. Dette bliver antagelig mere tydeligt ved høj belastning af glyphosat, hvor kvik og alm. hvene går tilbage (hvilket modellen fanger) og efterlader bedre plads til fåresvingel, som stort set ikke påvirkes af glyphosat. I simuleringen med høj glyphosatbelastning reagerer fåresvingel i overensstemmelse med observationerne med en højere biomasse i forhold til kontrollen men når ikke helt op på det observerede niveau. Denne mangel skyldes muligvis, at FieldEco ikke fanger den observerede vækst i efterårs- og vinterperioden, som derved mangles i simuleringen.
Sammenfaldet imellem simuleringer og observationer er dog generelt så godt, at det må konkluderes, at modellen er i stand til at simulere konkurrencen imellem de tre involverede arter over 6 år ved 4 forskellige påvirkninger af glyphosat (dog til dels med fåresvingel ved de højeste glyphosatniveauer som undtagelser).
Konklusion på validering
Simuleringerne af feltdata af fåresvingel, alm. hvene og kvik fra Kalø viser, at FieldEco er i stand til at simulere konkurrence imellem planter gennem flere år under påvirkning af glyphosat. Simuleringerne af feltdata fra Flakkebjerg faldt nogenlunde ud for stor nælde men mindre godt for mælkebøtte. Dette kan skyldes, at FieldEco ikke er blevet kalibreret til at simulere situationen med førsteårsplanter. Ydermere er mælkebøtte en meget variabel art, og mælkebøtterne fra Flakkebjerg kan meget vel være af en anden underart end den, som FieldEco er blevet kalibreret til.
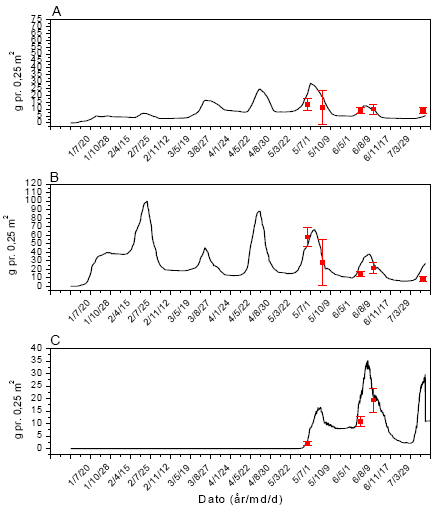
Figur 3.40. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) uden glyphosatpåvirkning.
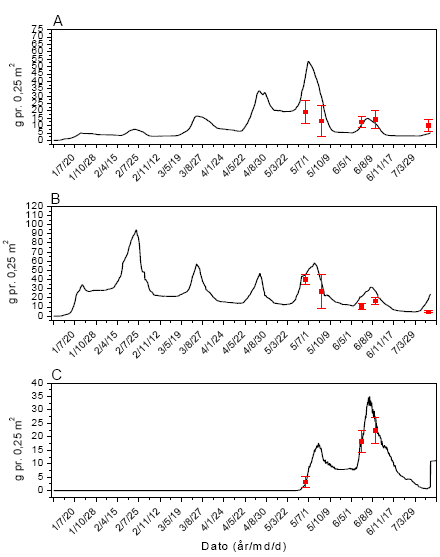
Figur 3.41. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 12 g glyphosat pr. ha.
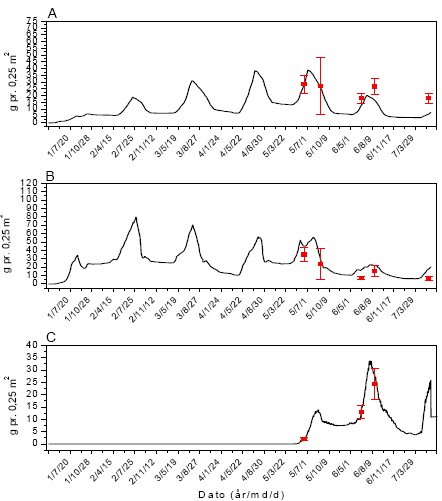
Figur 3.42. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 61 g glyphosat pr. ha.
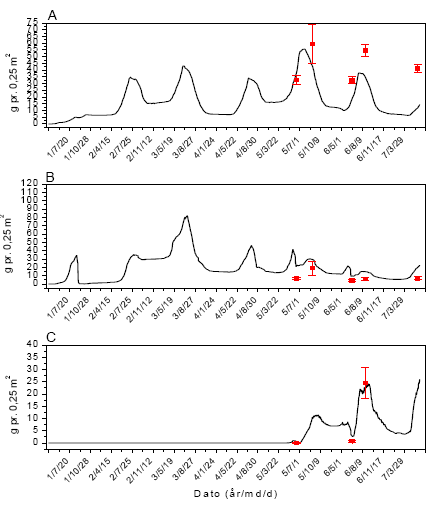
Figur 3.43. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 306 g glyphosat pr. ha.
3.2.5.3 Simulering af scenarier
Alle scenarier blev sat op som 7 års simuleringer af effekterne på hegnsvegetationen af sprøjtestrategierne relateret til 4 forskellige sædskifter (afsnit 1.7). En stor del af de danske levende hegn og markskel består af græsser, hvor kvik er meget stærkt repræsenteret, og spredte flerårige urter. Derfor er scenarierne sat op som scenarier, hvor de flerårige urter agertidsel, mælkebøtte, rejnfan, vild kørvel og stor nælde udsættes for en herbicidpåvirkning, mens de vokser i konkurrence med kvik. Herved opnås en simulering af den kombinerede effekt af herbicidpåvirkningen og konkurrencen med kvik. Dette gjorde sig dog i simuleringerne kun gældende for mælkebøtte og vild kørvel, mens det viste sig umuligt at simulere sameksistens imellem kvik og henholdsvis rejnfan, agertidsel og stor nælde. I alle disse tilfælde blev kvik skygget væk i løbet af 2-3 år. Dette er imidlertid ikke specielt overraskende, da især stor nælde og rejnfan men også i nogen grad agertidsel er bestanddannende monopoliserende arter i naturen. I naturen forekommer de som regel i tætte bestande og ikke blandet med græsset. Dermed er simuleringerne med FieldEco i overensstemmelse med, hvad man kan iagttage i naturen. Dog ses i naturen i nogle tilfælde sameksistens imellem agertidsel og kvik, men med parameteriseringen af agertidsel fra dette projekt var det ikke muligt at simulere.
I alle scenarier blev de involverede arter startet fra frø, og planterne fik to år til at etablere sig, før herbicidpåvirkningerne blev introduceret i simuleringerne. Dog blev scenarier med en herbicidbehandling om efteråret startet med en behandling i efteråret år 2. Til simuleringerne anvendtes hvert år klimadata fra 2001 fra DMUs klimastation i Silkeborg. Der blev anvendt gentagelser af samme klima for at undgå sløring af herbicideffekter på grund af variationer i temperatur og globalstråling (sollys).
Det blev valgt, at begrænse simuleringerne til at have cirka samme tidsmæssige udstrækning, som simuleringerne af feltdata fra Kalø, det vil sige 7 år, da der ikke er nogen sikkerhed for pålideligheden af simuleringer over længere tidsrum.
Mælkebøtte i konkurrence med kvik
Det var muligt at simulere sameksistens mellem kvik og mælkebøtte igennem 7 år ved lavt kvælstofniveau, mens kvik udkonkurrerede mælkebøtte ved højere niveauer. Alle scenarier med mælkebøtte blev kørt med den årlige maksimale tørvægt af reproduktive organer (knopstande, blomsterstande, frugtstande) som måleparameter for herbicideffekten. Denne parameter blev valgt, fordi den har med hegnsplantens (i dette tilfælde mælkebøtte) reproduktive output at gøre, og fordi blomstring af hegnsplanter kan anses for en naturkvalitetsmæssig værdi.
Korttidseffekter
Scenarierne viste en lille reduktion i mængden af reproduktive organer ved afdrift af glyphosat fra vinterraps i sædskifte 1, hvor vinterraps forekommer hvert 3. år (figur 3.44 øverst), det vil sige søjlen for sædskifte 1b er lavere end søjlen for sædskifte 1a, hver gang vinterraps forekommer. Denne effekt modvirkes dog i årene med vårbyg og vinterbyg, hvor der ikke er forskel i sprøjtningen imellem sædskifte 1a og 1b. Stigningen i mælkebøttens frugtsætning her skyldes modellens dynamiske allokering af ressourcer. Mælkebøttens frugtsætning er i modellen (og antagelig også i virkeligheden) følsom overfor størrelsen af overvintrende planter. Jo større plante, jo bedre frugtsætning. Hvis der et år er en reduceret frugtsætning, allokerer planten flere ressourcer til vækst af øvrige organer, hvilket betyder, at den overvintrende plante bliver større og derfor har relativt flere ressourcer at sætte ind i frugtsætning det følgende år. Derfor ses en kompensation i årene efter den reducerede frugtsætning på grund af glyphosat, en kompensation, der giver en svagt bedre frugtsætning ved anvendelse af glyphosatresistent raps end ved konventionel raps. Den negative effekt af glyphosat ved vinterrapsdyrkningen skyldes overvejende, at den reducerer mælkebøttens vækst i efteråret, hvorved den overvintrende plante bliver mindre. Denne negative effekt modsvarer fordelen af, at kvik rammes hårdere end mælkebøtte af glyphosat. Ved scenarier (ikke vist) med glyphosatsprøjtninger sidst på foråret er effekten omvendt, da mælkebøttens konkurrencefordel opvejer sprøjteeffekten.
Der er også i sædskifte 2 og 3 små korttidseffekter af sprøjtescenarierne med svag afdrift med en svagt forbedret blomstring ved brug af glyphosat i majs i stedet for MaisTer, hvilket skyldes den forbedrede konkurrencesituation for mælkebøtten efter sprøjtningen.
I sædskifte 4 er der en forbedret blomstring ved scenariet med glyphosat sammenlignet med konventionel sprøjtning, hver gang roer forekommer, men der kompenseres for denne effekt allerede det følgende år, hvor forholdet er omvendt, dog ikke ved den kraftigste afdrift på 25%, hvor mælkebøtte uddør i det konventionelle sprøjtemønster.
Langtidseffekter
Der ses ingen klare langtidseffekter af ændret sprøjtemønster pga. glyphosatresistente afgrøder i sædskifte 1, 2 og 3, men i sædskifte 4 er der en klar langtidsfordel af glyphosatanvendelsen ved høj afdriftsrate, idet mælkebøtten uddør ved det konventionelle scenarium. Dette skyldes den meget intensive sprøjtning, der foregår i forbindelse med roedyrkningen.
Vild kørvel i konkurrence med kvik
Det var muligt at simulere sameksistens af vild kørvel og kvik ved lave og medium koncentrationer af kvælstof. Ved høje koncentrationer af kvælstof blev kvik udkonkurreret på grund af skyggevirkningen fra den meget højere vild kørvel. Ved vild kørvel var der i simuleringerne tale om en plantepopulation under opbygning, da der fortsat var en stigning i mængden af reproduktive organer (figur 3.45) i de første år ved alle scenarier. Der var ligeledes en stigning i total biomasse med tiden i simuleringerne (ikke vist).
Korttidseffekter
Der er i vinterraps en svag tendens til en større produktion af reproduktiv biomasse hos vild kørvel ved sædskifte 1 med sprøjtescenariet ved anvendelse af glyphosatresistent raps end ved det konventionelle sprøjtescenarium. Effekten er dog ikke så stærk, at den holder sig ved de efterfølgende afgrøder ved alle afdriftskoncentrationer. Ved sædskifte 2 er der ligeledes en svagt positiv effekt på vild kørvel af scenarierne med glyphosat i sammenligning med scenarierne med MaisTer ved afdriftsniveauer på 5% og 25%. Ved 1% afdrift var effekten ikke stærk nok til at slå synligt igennem på figur 3.45.
Langtidseffekter
Der er en positiv langtidseffekt af den tidlige glyphosatsprøjtning (scenarium 3c), idet der er en klart højere maksimal reproduktiv biomasse i de sidste 4 år ved dette scenarium ved 25% afdrift end ved de øvrige scenarier i sædskifte 3. Denne tendens gør sig også gældende ved scenariet med 5% afdrift. En lignende tendens kunne forventes at slå igennem i sædskifte 2, men her bevirker den konventionelle sprøjtning i vårbyg hvert andet år en udjævning af effekten, som derfor ikke slår permanent igennem.
Stor nælde
Det var ikke muligt at simulere sameksistens imellem kvik og stor nælde, idet stor nælde i alle tilfælde konkurrerede kvik ud. Dette støttes af observationer fra naturen, hvor stor nælde er kendt for at være tæt bestanddannende. Ved præsentation af resultaterne fra scenarierne med stor nælde (figur 3.46) er den maksimale skudbiomasse præsenteret. Dette er blevet valgt, fordi skudbiomassen på grund af dens funktion som føde for adskillige almindelige sommerfuglearters larver synes at have større forvaltningsmæssig interesse end frøproduktionen.
Korttidseffekter
Der er en markant negativ korttidseffekt af glyphosat ved roedyrkningen hvert fjerde år i sædskifte 4. Effekten udjævnes allerede det følgende år. Der er også en mindre, negativ korttidseffekt af glyphosat i vinterrapsen i sædskifte 1, men kun ved 5 og 25% afdriftsniveauer.
Langtidseffekter
Ligesom for vild kørvel er der en klar tendens til, at den tidlige glyphosatsprøjtning er mindre skadelig end de øvrige sprøjtescenarier i sædskifte 3. Det er måske ikke så overraskende, at afdriften fra én sprøjtning er mindre skadelig end den halve afdrift fra 2 sprøjtninger, der derved påvirker planterne i længere tid. Men det er overraskende, at der er så stor forskel på den tidlige og den sene glyphosatsprøjtning. Denne forskel skyldes, at giftvirkningen i modellen holder i kortere tid i den tidlige sprøjtning end i den sene sprøjtning set på en graddagsskala. Planternes vækst simuleres på en graddagsskala med 5ºC som nulpunkt (ºD5), og giftens virkning antages i modellen af vare i 21 dage. På 21 dage efter den tidlige sprøjtning forløber der 166ºD5 og på 21 dage efter den sene sprøjtning forløber der 255ºD5. Dette skyldes, at der er en højere gennemsnitstemperatur i denne periode end efter den tidlige sprøjtning. Derved simuleres giftvirkningen til at være mere langvarig ved højere temperatur end ved lav målt på den graddagsskala, som planternes vækst simuleres efter. Hvorvidt det er korrekt at gøre sådan afhænger af, på hvilken tidsskala varigheden af glyphosats giftvirkning skal ses? Hvis nedbrydningen bedst beskrives ved hjælp af timer og dage, er simuleringsresultatet med en forskel imellem scenarium 3c og 3d i overensstemmelse med virkeligheden, men hvis nedbrydningen bedst beskrives på samme graddagsskala som planternes vækst, er forskellen et modelartefakt.
Agertidsel
Det var ikke muligt at simulere sameksistens imellem agertidsel og kvik i FieldEco med mindre kvælstofniveauet blev sat ekstremt lavt. Ved realistiske kvælstofniveauer udkonkurrerede agertidslen kvik i løbet af to eller tre år. Dette skyldes måske, at den tidselgruppe (måske en klon), der blev benyttet til parameterisering var meget kraftig og tæt bestandsdannende. Da der i naturen kan findes sameksisterende bestande af tidsler og kvikgræs, er der muligvis forskel på forskellige tidselkloners konkurrenceevne.
I scenarierne blev det besluttet at holde fast ved den parameterisering af tidsel, som var blevet gennemført, da der ikke fandtes oplysninger i litteraturen om, hvordan parameteriseringen skulle være for at kunne simulere sameksistens. Resultaterne i scenarierne er derfor baseret på effekterne af herbiciderne på agertidsel voksende uden interspecifik konkurrence (fig. 3.47).
Korttidseffekter
Der er korttidseffekter på blomstringen af agertidsel i sædskifte 2, hvor effekterne af sprøjtescenarierne med glyphosat er mindre skadelig end sprøjtescenariet med MaisTer, idet der er et forøget maksimum af reproduktive organer ved scenarierne med glyphosat i 5% og især i 25% afdriftsscenariet. En markant korttidseffekt findes i sædskifte 4, hvor sprøjtningen med glyphosat i forbindelse med roedyrkningen har en negativ effekt på tidslernes reproduktion; effekter er dog overvundet allerede det følgende år i forbindelse med vårbyggen.
Langtidseffekter
Det eneste sted, hvor der forekommer langtidseffekter på tidslernes produktion af reproduktive organer, er i sædskifte 3 med permanent majs. I dette sædskifte slår forskellen mellem scenarierne med MaisTer og glyphosat fra sædskifte 2 permanent igennem. De tilsvarende effekter i sædskifte 2, hvor majsdyrkningen kun forekommer hvert andet år, forsvinder i år uden majs.
Rejnfan
Det var ligesom for agertidsel og stor nælde ikke muligt at simulere sameksistens mellem rejnfan og kvik. Også i dette tilfælde blev kvik udkonkurreret af en tæt bestanddannende art, hvilket er i overensstemmelse med observationer fra naturen, hvor rejnfan som regel forekommer i tætte bestande.
Korttidseffekter
Der var ingen klare korttidseffekter af sprøjtescenarier med glyphosat på rejnfan.
Langtidseffekter
Der var en langtidseffekt i form af et generelt lavere frugtsætningsniveau ved 25% afdrift i alle sædskifter (fig. 3.48). Det vil sige, at pesticidet påvirker frugtsætningen, men at der generelt ikke er de store forskelle imellem sprøjtescenarierne inden for sædskifterne.
Figur 3.44.
Sammenligning imellem den årlige maksimale frugtsætning hos almindelig mælkebøtte i konkurrence med kvik ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.45.
Sammenligning imellem den årlige maksimale frugtsætning hos vild kørvel i konkurrence med kvikgræs ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.46.
Sammenligning imellem den årlige maksimale frugtsætning hos stor nælde ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.47.
Sammenligning imellem den årlige maksimale frugtsætning hos agertidsel ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.48. Sammenligning imellem den årlige maksimale frugtsætning hos rejnfan ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
3.2.5.4 Konklusion på simuleringer
Der blev i simuleringerne fundet små korttidsforskelle på de flerårige tokimbladede hegnsplanter i sædskifter med majs ved anvendelse af glyphosat i stedet for MaisTer, hvilket overvejende skyldes relativt svagere sensitivitet overfor glyphosat hos disse arter. I sædskifter med vinterraps og roer er billedet ikke så klart, da der blev fundet mindre skadelige effekter af glyphosat end af konventionel sprøjtning ved nogle kombinationer af arter og sædskifter og mere skadelige effekter ved andre kombinationer.
Hovedparten af de fundne forskelle imellem effekter af glyphosat i forhold til konventionel behandling var dog af kortvarig karakter og forsvandt igen i det følgende år, hvor der i tre af sædskifterne forekommer en kornafgrøde med konventionelt sprøjtemønster i alle scenarier. Simuleringerne peger kun på langtidsforskelle imellem effekterne af sprøjtningerne ved dyrkning af konventionelle afgrøder og glyphosatresistente afgrøder i forbindelse med sædskiftet med permanent majs, hvor der ikke er en kornafgrøde i sædskiftet til at rette forskellen op.
Der blev ikke fundet særlig markante forskelle imellem konventionel sprøjtning og glyphosatsprøjtninger ved 1% og 5% afdrift, men ved 25% begyndte forskellene at slå klart igennem. Det blev ikke undersøgt ved simulering, hvorledes planternes vækst ville have været helt uden herbicidpåvirkning.
4 Diskussion
På baggrund af de opnåede resultater og den eksisterende viden diskuteres i det følgende, hvilke ændringer dyrkning af glyphosattolerante afgrøder (GT-afgrøder) vil føre til – dels på kort sigt, det vil sige over én dyrkningssæson, og dels på lang sigt, det vil sige over flere år gennem hele sædskiftet. Projektets resultater forholdes til praksis gennem scenarierne opbygget over følgende fire sædskifter (se afsnit 1.7):
- Vinterraps-vårbyg-vinterbyg.
2. Majs-vårbyg.
3. Vedvarende majs.
4. Roer-vårbyg-vinterhvede-vinterhvede.
I hvert scenarium kan den fremhævede afgrøde enten være konventionel eller glyphosattolerant. Projektets sociologiske delstudium1 inddrages i diskussionen af, hvorledes de tekniske muligheder kan udmøntes i praksis, det vil sige hvilken sprøjteadfærd vi kan forvente landmændene vil have ved dyrkning af GT-afgrøder.
4.1 Ændringer på kort sigt
4.1.1 Hegnets flora
Ved afdrift kan floraen i hegn og andre marknære habitater påvirkes af herbicider. Den dosis, der afsættes i hegnet, varierer med en lang række forhold heriblandt vejrforhold og dysetype. Endvidere er afgrøden bestemmende for, hvilke midler og hvor mange behandlinger landmanden benytter. Ved den enkelte behandling kan afsættes op til 25% af markdosis110-112. For de fleste plantearter er 25% af markdosis ikke en dødelig dosis. Men små, nyligt spirede planter kan dø af påvirkningen, og større planter kan blive påvirkede selv af sublethale doseringer.
Der kan være stor forskel i plantearters følsomhed over for herbicider, og følsomheden kan afhænge af forsøgsbetingelserne. Vi fandt eksempelvis i potteforsøget, at alm. hvene er mere følsom over for glyphosat og MaisTer end fåresvingel. Hvis ED10 doseringen for begge herbicider på alm. hvene sættes til 1 N, når den vokser alene uden konkurrence fra andre arter, så er ED10 doseringen for fåresvingel af henholdsvis glyphosat og MaisTer 2 N og 10 N. Men hvis de to arter vokser i konkurrence med hinanden, forstærkes forskellen i følsomhed, idet ED10 for glyphosat på alm. hvene bliver 0,84 N MaisTer, og den tilsvarende værdi for MaisTer bliver 0,67.
Undersøgelse af herbicideffekter på enkeltarter kan altså ikke anvendes til præcist at forudse effekten i et plantesamfund, da konkurrencen mellem arterne ændrer deres respons på herbicidet. I et tidligere studium af mechlorprops indflydelse på konkurrenceforholdet mellem storkenæb og hyrdetaske fandt vi, at begge arters konkurrenceevne blev øget ved eksponering med lave doser af mechlorprop på trods af at der også her var stor forskel i arternes følsomhed over for herbicidet113. Det kan med andre ord være vanskeligt at forudsige, hvilken indflydelse en herbicidbehandling har på konkurrenceforholdet mellem plantearter.
Fåresvingel hører til de planter, som har en høj tolerance over for glyphosat. Derfor begunstiges denne art i konkurrencen med andre vilde planter ved afdrift af glyphosat fra GT-afgrøder. At fåresvingel således får forbedret sin konkurrenceevne under påvirkning af glyphosat fremgik meget tydeligt både af konkurrenceforsøget udført i væksthus og af successionforsøget i felten. Derimod var effekten af MaisTer på planternes konkurrenceforhold ikke entydig: I væksthusforsøget blev fåresvingel begunstiget i forhold til alm. hvene, mens dækningsgraden af fåresvingel under markforhold blev reduceret og dækningsgraden af alm. hvene øget. Det er dog uklart, i hvor høj grad denne effekt skyldes tilførsel af gødning (100 kg N) eller MaisTer, idet forsøget ikke var designet til at adskille effekterne.
Simuleringerne viste, at de flerårige tokimbladede planter i hegnet vil reagere svagt positivt i deres vækst ved anvendelse af glyphosat i stedet for MaisTer i majs. Dette skyldes overvejende en mindre sensitivitet overfor glyphosat, sammenlignet med MaisTer, hos disse plantearter. Der er ikke noget overraskende i, at forskellene imellem anvendelsen af de to sprøjtemidler er ret små, da sprøjtningerne med de to midler i scenarierne falder på stort set de samme tidspunkter og derfor også kan forventes at have nogenlunde samme effekt i hegnene. I majs betyder muligheden for at anvende glyphosat ikke ret meget for hegnsfloraen, da man i konventionel majs i forvejen anvender et bredspektret sprøjtemiddel (MaisTer), som ikke skader afgrøden.
I sædskifterne med vinterraps og roer gav simuleringerne et mindre entydigt svar for hegnsfloraen, idet effekten af glyphosat varierede med plantearten. I disse sædskifter spillede forsinkelsen af sprøjtetidspunktet den afgørende rolle i simuleringerne. Dette hænger sammen med timingen af sprøjtningen i forhold til plantens knopsætning. Hvis sprøjtningen falder således, at den reducerer plantens biomasse op mod knopsætning, giver dette planten færre kræfter til at danne og udvikle knopper .
Som målepunkt for simuleringerne af de flerårige urters vækst i hegn og skel er anvendt den maksimale biomasse af frugter/blomster, fordi denne parameter er vigtig for mange menneskers opfattelse af hegnets naturkvalitet. Derudover er denne parameter valgt, fordi blomstersætningen spiller en rolle for pollen- og nektarspisende insekter, samt fordi frugtsætningen spiller en rolle for frøædende arter (insekter, fugle, mus).
Modellen simulerer udelukkende knopsætning og blomstring ud fra beregning af, hvorhen de ved fotosyntesen producerede sukkerstoffer allokeres, og giftvirkningen simuleres alene ved effekter på fotosyntesen og vækstraten. Der er altså ikke inkluderet en direkte effekt af herbicidet på frøsætningen, hvilket muligvis er en forsimpling. Undersøgelser fra glyphosatresistent bomuld114 peger på, at glyphosat har en direkte virkning på frøsætning, hvorimod det samme ikke er tilfældet for glyphosatresistent majs. En sublethal effekt af glyphosat på blomstersætningen er også fundet hos den australske ukrudtsplante Senna obtusifolia115. Glyphosat kan altså meget vel have en betydelig større effekt på lige netop parameteren frøsætning end simuleringsmodellen beregner. Det bør derfor undersøges nærmere, hvor udbredt disse giftvirkninger er blandt plantearterne i og omkring markerne. Blandt afgrøder er der fundet herbicideffekt på blomstringen af rødkløver ved brug af Starane 180S137 og på raps ved brug af Express®138.
4.1.2 Markens biodiversitet og udbytte
Det er velkendt, at en udsættelse af herbicidbehandlingen medfører behov for en øget dosering, hvis bekæmpelseseffekten skal fastholdes. Det skyldes, at enårige ukrudtsplanter generelt er mest følsomme for herbicider i de tidlige udviklingsstadier, og at den nødvendige forøgelse af dosering er større for ikke-systemiske end for systemiske herbicider116. Både glyphosat og MaisTer er systemiske herbicider, men vi fandt, at stigningen i doseringsbehovet ved senere sprøjtning var større for MaisTer end for glyphosat. Tidligere forsøg har vist, at indflydelsen af ukrudtets størrelse på effekten af glyphosat er mest udtalt for de ukrudtsarter, som er mindst følsomme overfor glyphosat f.eks. liden nælde og snerle-pileurt7. Hvis man ønsker at forøge ukrudtsdækket i forsommeren gennem en forsinkelse af herbicidbehandlingen, vil det altså i GT-majs være muligt at gøre dette med en mindre forøgelse af behandlingsindekset end i konventionel majs. Det samme gør sig sandsynligvis gældende for andre GT-afgrøder (raps og roer), da majoriteten af de herbicider, der anvendes i raps og roer, er enten jordmidler eller ikke-systemiske herbicider.
Markforsøget med majs viste, at splitsprøjtning gav samme majsudbytte, hvad enten der anvendtes glyphosat eller MaisTer. Begge midler er da også effektive over for de dominerende ukrudtsarter i marken. Der erindres om, at der ikke anvendtes egentlig GT-majs, men at dette blev efterlignet ved afdækning af majsplanterne inden sprøjtning. Hvis man nøjedes med én glyphosatsprøjtning på det sene af de to tidspunkter i splitsprøjtningen, var der imidlertid risiko for udbyttetab. En udsættelse af glyphosatsprøjtningen på yderligere to uger gav klare udbyttetab. Erfaringerne med GT-majs i USA har vist, at netop fordi glyphosat kan bekæmpe stort ukrudt, er landmændene tilbøjelige til at udsætte sprøjtningen så længe, at de ikke opnår maksimalt udbytte som følge af ukrudtets konkurrence med afgrøden117.
Hvis GT-majs skal anvendes i praksis, og man ønsker, at landmanden skal udnytte mulighederne for senere sprøjtning end normalt, så forestår der altså et større forsøgsarbejde for at finde de rette sprøjtestrategier. Fra dette projekts sociologiske del ved vi ydermere, at landmændene ikke er risikovillige, når det gælder ukrudtsbekæmpelsen1. Man kan dog forestille sig, at hvis GT-majs indføres i dansk dyrkningspraklsis, så vil sprøjtestrategien umiddelbart ligne den gængse, når man ser på timingen af behandlingerne. Efterhånden som landmændene får erfaring med anvendelsen af glyphosat, og konsulenterne får datagrundlag til at lægge mere tidsmæssigt smidige sprøjteplaner, så vil der muligvis ske en forskydning hen mod lidt senere sprøjtetidspunkter.
Den samme forskydning mod senere sprøjtetidspunkt vil man kunne forvente, hvis andre GT-afgrøder tages op i dyrkningen. Jo mere konkurrencestærk GT-afgrøden er over for ukrudtet, jo mere rum vil det give for en forsinkelse af glyphosatsprøjtningen. Effekten på biodiversitet vil desuden være størst i forårssåede GT-afgrøder. I efterårssåede afgrøder vil landmanden foretrække at lægge første herbicidsprøjtning i efteråret for at få den største dyrkningssikkerhed, og sprøjtestrategien vil således ikke være forskellig fra den, der anvendes i konventionelle afgrøder.
Væksten af afgrøde og ukrudt samt tætheden af leddyr fulgtes ad gennem sæsonen på begge de undersøgte majsmarker. På en given dag var der imidlertid generelt ringe sammenhæng mellem biomassen af ukrudt (eller afgrøde) og insekter i de to majsmarker. Det er således ikke muligt ud fra ukrudtsbiomassen at forudsige insektbiomassen en given dag.
Majs er ligesom roer12 og til dels raps (Bruus et al., upubl.) en afgrøde, der giver god plads til ukrudt i begyndelsen af vækstsæsonen Til sammen tyder resultaterne af de tidligere undersøgelser og dette projekt på, at der i marker med lav afgrødedækning og forholdsvis meget ukrudt først på sæsonen (f.eks. majs og roer) er en bedre sammenhæng mellem udviklingen i ukrudt og insektbiomasse end i marker med lille ukrudtsbiomasse. I hvor høj grad sammenhængen skyldes en direkte fødekædeeffekt eller mere skyldes, at det øgede plantedække giver et mere favorabelt mikroklima, er dog ikke klarlagt.
Roeundersøgelsen12 samt de engelske undersøgelser af GM-afgrøder16,19 har desuden vist, at en forsinkelse af herbicidsprøjtningen kan resultere i flere insekter, formodentlig på grund af den øgede ukrudtsbiomasse. Det samme vil formentlig kunne ske ved senere sprøjtning i andre afgrøder med god plads til ukrudt, f.eks. hvis der dyrkes en herbicidtolerant sort, som kan sprøjtes senere end en konventionel sort uden konsekvenser for udbyttet. Hvis perioden med flere insekter falder sammen med yngleperioden for fugle, der søger insektføde til deres unger i marken, vil disse fugle også have fordel af en sådan dyrkningspraksis.
4.2 Ændringer på lang sigt
De to modelstudier af GT-afgrøders effekt på floraen viste samstemmende for både mark og hegn, at effekterne af glyphosat generelt blev udlignet allerede det følgende år i sædskifter, som skiftede mellem glyphosat og konventionel behandling. Der kunne derfor i de fleste tilfælde ikke iagttages langtidseffekter af GT-afgrøder i modellerne, hvis de indgik i sædskifte med konventionelle afgrøder.
4.2.1 Hegnets flora
Vi fandt, at hvis en veletableret flerårig plante (det vil sige skud fra rod eller stængel) rammes af herbicid, så vil langtidseffekten, 20 uger efter sprøjtning eller senere, være betydeligt lavere, end den effekt som måles 1-3 uger efter sprøjtning. Herbicidet forårsager altså en midlertidig væksthæmning. I modsætning hertil reagerede flerårige planter fremspiret fra frø mere som enårige planter, og der var god overensstemmelse mellem herbicideffekter målt fra 2 til 11 uger efter sprøjtning. Der kræves derfor betydeligt højere doseringer for at opnå en varig langtidseffekt på rodfremspirede planter af flerårige arter end på frøfremspirede planter. Det betyder, at veletablerede flerårige hegnsplanter har en naturlig modstandskraft mod herbicidafdrift, men til gengæld vil rekruttering af nye planter fra frø bliver hæmmet af afdriften.
I forbindelse med ukrudtsbekæmpelse er det velkendt, at flerårige planters reserver (i oplagringsorganer som rødder og stængler) gør bekæmpelse besværlig. Herbicidanvendelsen skal da times i forhold til plantens livscyklus for at udtømme dens reserver og endeligt slå den ihjel. Sådanne strategier er beskrevet for f.eks. agertidsel118. At målrettede sprøjtestrategier er nødvendige for at bekæmpe flerårigt ukrudt gør det yderligere usandsynligt, at tilfældig sprøjteafdrift vil have stor virkning på flerårige hegnsplanters vækst.
Herbicidafdrift udsætter marknære habitater for gentagne subletale doser, hvilket påvirker vegetationen. Antallet af arter reduceres, og artssammensætningen påvirkes til fordel for arter, der er mindre følsomme overfor herbicider (eksemplificeret ved fåresvingel i Kalø-forsøgene). For følsomme arter reduceres endvidere mængden af spiringsdygtige frø i jorden, således at rekrutteringsgrundlaget reduceres. Det betyder, at floraen i de marknære områder bliver artsfattig og domineres af få herbicidtolerante arter. Undersøgelserne underbygger således de iagttagelser, der er gjort i undersøgelser i danske hegn på konventionelle henholdsvis økologiske brug, hvor man har set, at hegnsfloraen på de konventionelle brug rummede færre arter, og at navnlig andelen af tokimbladede urter var lavere119,120.
Vi fandt, at ekstrem afdrift af glyphosat giver mere bar jord, hvilket også er fundet i andre danske afdriftsforsøg121. Den reducerede mængde af spiringsdygtige frø med stigende herbiciddosis forklarer, hvorfor der ikke sker en etablering af planter, selvom der skabes huller i vegetationen. I de tilfælde, hvor kimplanter er spiret, har det som oftest ikke ført til etablering af planterne. Dette kan meget vel skyldes, at kimplanterne er mere følsomme over for herbicider, end planterne er på senere vækststadier. Samlet set dokumenterer felteksperimentet, at den reduktion i biodiversiteten, som andre undersøgelser har antydet23,122-125, skyldes herbicidpåvirkningen.
Ud over afdriften af pesticider modtager markhegn også store mængder gødning, navnlig kvælstof, som sammen med herbicidanvendelsen påvirker floraen126. Når landbrugsdriftens virkninger på den omgivende natur skal bedømmes, er det derfor vigtigt at medregne effekten både af pesticider og af gødning.
4.2.2 Markens biodiversitet og udbytte
Herbiciders effekt bedømmes normalt som effekten på antal planter eller biomasse på forskellige tidspunkter efter sprøjtning, mens effekten på den reproduktive evne ikke indgår som en standardparameter127. I potteforsøgene fandt vi, at langtidseffekten målt som frøproduktion af en given dosis var mindre end effekten på biomasse målt 3 uger efter behandling. Samme resultat er tidligere fundet i andre potteforsøg128. Når planterne vokser uden konkurrence, kompenserer overlevende ukrudt ved genvækst efter behandlingen og sætter flere frø end forventet udfra den reduktion, som blev målt på biomassen.
Når effekten af MaisTer og glyphosat på vegetativ vækst og frøproduktion sammenlignes, så må man for en given effekt på biomassen forvente en mindre effekt på frøproduktion af MaisTer end af glyphosat. Et skifte fra konventionel til GT-majs vil derfor med uændret effektkrav føre til en nedsat frøproduktion hos ukrudtet.
Langtidseffekterne af at inkludere GT-afgrøder i sædskiftet eller ej blev belyst ved simuleringer af de fire dyrkningsscenarier, hvor ændringer i frøbanken blev anvendt som målevariabel. Med GT-raps eller GT-roer i sædskiftet adskilte udviklingen i ukrudtsfloraen sig ikke fra den, hvor henholdsvis konventionel raps eller konventionelle roer indgik. Men med GT-majs viste der sig en effekt af middelvalget, idet splitsprøjtning med glyphosat generelt var mere effektiv på lang sigt end med MaisTer (ved de valgte doseringer), især opnåedes med glyphosat en bedre langtidsbekæmpelse af lugtløs kamille. En enkelt glyphosatsprøjtning havde naturligvis mindre effekt på ukrudtsfloraens udvikling, men hvis den lå sent, kom hvidmelet gåsefod ud af kontrol. Dette er i overensstemmelse med amerikanske forsøg med ensidig dyrkning af GT-majs, hvor hvidmelet gåsefod ligeledes blev dominerende ved anvendelse af en lav dosering af glyphosat129. I et balanceret sædskifte vil hvidmelet gåsefod dog kunne kontrolleres med andre midler end glyphosat, hvoraf mange er effektive over for denne art.
Ensidig majsdyrkning inviterer i sig selv til ukrudtsproblemer, og simuleringerne pegede da også på lugtløs kamille som et problemukrudt i konventionel majs med MaisTer-splitsprøjtning. Men ved at skifte til ensidig GT-majs og glyphosat-splitsprøjtning var en tilstrækkelig ukrudtsbekæmpelse faktisk mulig. En enkelt glyphosatsprøjtning ville dog være utilstrækkelig og føre til opformering af hvidmelet gåsefod, jo hurtigere desto senere man lægger sprøjtningen.
Indenfor de seneste år er resultaterne fra de første sædskifteforsøg med GT-afgrøder blevet publiceret. Forsøgene viser, at hvis der dyrkes GT-afgrøder hvert eneste år, og der kun anvendes glyphosat, så vil der med tiden ske forskydninger i ukrudtsfloraens sammensætning, hvor de fra naturens side mindst følsomme ukrudtsarter bliver dominerende129,130. Disse resultater understøttes af erfaringerne fra praksis, hvor der i mange marker er observeret markante ændringer i ukrudtsfloraen131-135. Denne udvikling har betydet, at det i praksis nu anbefales at anvende glyphosat i kombination med andre herbicider f.eks. et jordmiddel ved såning134.
Foruden en ændring i ukrudtsfloraens sammensætning kunne man forestille sig, at diversiteten i ukrudstfloraen ville reduceres som følge af glyphosats bredspektrethed. Der foreligger meget få undersøgelser, som kan dokumentere, om dette er tilfældet. Der er fundet en reduceret diversitet i sædskifter med GT-majs og GT-sojabønne130 men ikke i sædskifter med GT-raps133, hvilket kan skyldes forskelle i hyppigheden af GT-afgrøder i sædskiftet. I det engelske BRIGHT projekt136 blev der ikke observeret nedgang i artsdiversiteten i nogen af sædskifterne efter 4 år, og frøbanken blev forøget i sædskifter med både GM-raps og GM-roer. GM-afgrøderne havde kun lille effekt på ukrudtsbekæmpelsen i de efterfølgende afgrøder og medførte reduceret forbrug af aktivstof i roer og lavere omkostning i både raps og roer. Et forhold man skal tage i betragtning, når man skal vurdere eventuelle effekter på diversiteten i ukrudstfloraen, er, at glyphosat ofte har afløst anvendelse af tankblandinger eller splitspøjtninger med 3 eller 4 forskellige herbicider.
En anden risiko ved intensiv dyrkning af GT-afgrøder er opformering af glyphosatresistente ukrudtsarter. Selv om risikoen for udvikling af resistens overfor glyphosat anses for at være lavere end for andre herbicider, så er der tidligere fundet glyphosatresistente ukrudtsarter i afgrøder og dyrkningssystemer, hvor der er en intensiv anvendelse af glyphosat, f.eks. plantager og hvede-rajgræssædskifter i Australien. Introduktionen af GT-afgrøder har imidlertid bidraget til fremkomsten af nye tilfælde af glyphosatresistente ukrudtsarter såsom canadisk bakkestjerne, hvidmelet gåsefod, italiensk rajgræs, bynke-ambrosius samt et par amarantarter117,131.
Landmanden er fokuseret på dyrkningssikkerhed, en praktisk hverdag i produktionen og økonomisk udbytte1. GT-afgrøder vil give ham flere frihedsgrader i ukrudtsbekæmpelsen, men disse var dog ikke indbygget i scenarierne, hvor sædskifte og sprøjtestrategi lå helt fast. I praksis må landmanden formode at korrigere sin sprøjtepraksis ud fra forholdene og ikke lægge sig fast på en strategi, som blot forøger ukrudtet fra år til år. Tænker man sig f.eks., at han på grund af belejlighed udskyder sprøjtningen ét år, og det medfører et stort frøkast, så vil han uden tvivl overveje at opjustere bekæmpelsen i de følgende år.
4.3 GT-afgrøder i praksis
På baggrund af resultaterne fra det sociologiske studium1 må det konkluderes, at indførelse af GT-afgrøder på kort sigt ikke af sig selv vil føre til, at de forventede miljømæssige fordele (i form af større ukrudtsflor i foråret) opnås i fuldt omfang. Landmændene vil dog efter alt at dømme sprøjte færre gange og vil lægge den første sprøjtning noget senere i forhold til, hvad der med de nuværende dyrkningssystemer er normen. Samtidig vil der være et ønske om at kombinere glyphosat med andre sprøjtemidler – både for at opnå en mere effektiv bekæmpelse og for at undgå udvikling af glyphosatresistent ukrudt. Dette vil forebygge de negative langtidseffekter som en ensidig anvendelse af glyphosat kan føre til. Ud over forebyggelsen af resistens handler det om at undgå en ensidig påvirkning af plantesamfundene i mark og hegn.
En introduktion af GT-afgrøder i Danmark vil kun udløse de mulige positive effekter på miljøet, hvis den følges af en indsats for at påvirke landmændenes holdninger og sprøjteadfærd. Når dette er sagt, skal det understreges, at nogle af de holdninger, der er afdækket i det foregående, formodentlig ligger meget dybt i danske landmænd. Der er derfor ingen grund til at tro, at det vil være en let sag at påvirke danske landmænd, så de følger foreskrevne sprøjteplaner om senere sprøjtning i GT-afgrøder. Dette understreges af udsagnene fra rådgiverne om, at det, der i almindelighed skal til for at skabe adfærdsændringer blandt landmændene, enten er lovmæssige forbud/påbud eller betydelige økonomiske gevinster.
5 Konklusioner
Undersøgelsen havde til formål at undersøge, hvilke konsekvenser det ville have for for flora og fauna i agerlandet at indføre dyrkningen af glyphosattolerante (GT) afgrøder. Ved dyrkning af GT-afgrøder vil glyphosat være det dominerende herbicid. En sådan ændring i forhold til den nuværende sprøjtepraksis, hvor glyphosat har stærkt begrænset anvendelse i landbruget, ville ændre levevilkårerne for agerlandets flora og fauna. Det var projektets formål at undersøge hvor voldsomme ændringer, der ville kunne forventes ved overgangen til dyrkning af GT-afgrøder. Sådanne ændringer kunne afstedkommes både af den direkte virkning af glyphosat i marken og af afdriften af glyphosat til det omgivende hegn. Selvom glyphosat kun har giftvirkning på planter, kunne derendvidere tænkes afledte effekter på faunaen.
På baggrund af undersøgelsens resultater (kapitel 3) og diskussion (kapitel 3.2.5.4) konkluderer vi følgende:
Hvis GT-afgrøder dyrkes ensidigt, vil dette indebære en risiko for, at ukrudtet gennem evolution udvikler resistens over for glyphosat. Endvidere vil der i det omgivende hegn, som modtager sprøjteafdrift fra marken, gennem plantesamfundets succession kunne ske en forarmning hen imod en flora sammensat af få, GT-tolerante plantearter. Begge disse risici kan imidlertid imødegås ved ikke at dyrke GT-afgrøde ensidigt, men i sædskifte med ikke-GT-afgrøder. Det skiftende valg af herbicider, som et sådant sædskifte indebærer, vil modvirke den uønskede ensretning af begge økologiske processer, evolution og succession.
Ligegyldigt hvilke herbicider, der anvendes, så vil afdriften dog under alle omstændigheder have en negativ indflydelse på hegnets flora. Sammenlignet med floraen i hegn omkring usprøjtede marker, vil artsantallet være mindre. Artsammensætningen i hegnet vil afspejle de herbicider, der har været anvendt i marken (i GT-afrgrøder, altså særligt glyphosat), idet de arter, som er mest tolerante over for de anvendte herbicider, vil dominere. En reduktion af herbicidbelastningen vil derfor være gavnlig for vegetationen. Der mangler imidlertid viden om retableringen af planter ved reduktion af belastningen. Dels hvor stor en reduktion der skal til for, at arterne reetablerer sig, dels hvor lang tid reetableringsprocessen tager.
For at øge agerlandets biodiversitet ville der være ønskeligt at forøge mængden af ukrudt i markerne om foråret. Dette kunne opnås ved at udskyde herbicidbehandlingen i forhold til gængs praksis. Ved en udskydelse af behandlingen kan ukrudtet nå at vokse sig større inden behandling og kan derved, både i sig selv og som føde og levested for faunaen, bidrage til biodiversiteten. Imidlertid kræver større ukrudt også større herbiciddosis. En forøgelse af biodiversiteten gennem en udskydning af behandlingen vil altså forøge markens behandlingsindeks. Hvis man sammenligner den forøgelse af dosis, som er nødvendig med glyphosat i forhold til andre midler, så vil glyphosat føre til en mindre stigning i behandlingsindeks end de andre midler. Derfor vil det i GT-afgrøder være muligt at udskyde behandlingen med en mindre forøgelse af behandlingsindeks til følge end i konventionelle afgrøder.
I konkurrencesvage afgrøder som majs og roer kræves et større forsøgsarbejde for at kortlægge de præcise muligheder for at udsætte sprøjtningen i GT-sorter uden, at det går ud over dyrkningssikkerheden. Man taler om et ”sprøjtevindue”, det vil sige den periode, inden for hvilken bekæmpelsen skal udføres for at undgå udbyttetab. Sprøjtevinduet afhænger af afgrøde, ukrudt og herbicid samt jordbundsforhold of vejr. Det er disse mange faktorer, som gør at forsøgarbejdet bliver meget omfattende, samt det at vi ønsker at undersøge mulighederne for at udskyde sprøjtningen længst muligt af hensyn til biodiversiteten..
Som det fremgår af resultaterne af dettes projekts sociologiske del1, er landmændene skeptiske over for ideen om at opgive de traditionelle pesticider. Med deres nuværende viden og holdninger vil de i forbindelse med dyrkning af GT-afgrøder vælge at kombinere glyphosat med andre sprøjtemidler. Denne holdning skyldes ikke blot konservatisme, men ren fornuft. Således mindskes nemlig risikoen for udviklingen af glyphosattolerant ukrudt.
Herbiciders effektivitet vurderes normalt (blandt andet i forbindelse med godkendelsen af nye herbicider) ud fra tests på enkeltarter, men denne vurdering kan ikke anvendes til præcist at forudse effekten i et plantesamfund, da konkurrencen mellem arterne ændrer deres respons på herbicidet. Endvidere testes normalt korttidseffekten på plantens biomasse, men vi fandt, at langtidseffekten, opgjort som frøproduktionen hos det overlevende ukrudt, er mindre end korttidseffekten på biomassen. Effekten på biomasse og frøproduktion blev sammenlignet ved den dosis af herbicidet, der skulle til for at forårsage en 90% reduktion af enten biomasse eller frøprodsuktion. Forskellen i kort- og langtidseffekten varierer mellem herbiciderne. Ved indførelse af GT-majs vil forskellen mellem MaisTer og glyphosat føre til, at ukrudtets frøproduktion i GT-majs bliver mindre end i konventionel majs. Dette vil på længere sigt føre til at frøpuljen i GT-majs bliver mindre end i konventionel majs.
For at vurdere et herbicids langtidseffekt på ukrudtsbestanden er det altså nødvendigt at kende effekten på planternes reproduktionsevne. Da det er ret tids- og arbejdskrævende at måle frøproduktion, er disse data ofte ikke tilstede. Vores resultater viser, at biomasse er en mindst lige så følsom parameter at måle effekter på som frøproduktion, og der er derfor ikke grund til at antage, at en herbicidbelastning vil have større effekt på den reproduktive evne end på den vegetative vækst. Andre undersøgelser har imidlertid fundet negative virkninger af herbicider på planters blomstring,. pollenproduktion og frugtsætning. Det bør derfor undersøges nærmere, hvor udbredt disse giftvirkninger er blandt plantearterne i og omkring markerne.
Til vurdering af langtidseffekter på enårige arter er korttidseffekten (f.eks. biomasse 3 uger efter sprøjtning) anvendelig, mens langtidseffekten på flerårige arter kan være vanskelig at forudsige udfra korttidssymptomerne. På trods af flere ugers vækststandsning kan planterne stadig genoptage væksten, og dette kan kun afdækkes gennem længerevarende forsøg. Vi prøvede at finde metoder til at bestemme langtidseffekten af den midlertidige væksthæmning ud fra målinger på korttidseffekten, men det lykkedes ikke.
Mere ukrudt i marken kan give flere insekter, hvilket potentielt også kan have en positiv effekt længere op i fødekæden. Vi kan imidlertid ikke forudsige mængden af insekter blot ud fra ukrudtsmængden.
To simuleringsmodeller blev videreudviklet i dette projekt: FieldWeeds til simulering af ukrudtet i marken og FieldEco til simulering af planterne i hegnet. De vil også i fremtiden kunne indgå som værktøjer i forskningen i planters vækst og populationsdynamik. Ligeledes er den nye statistiske model for to arters konkurrence under herbicidpåvirkning en væsentlig nyskabelse i forhold til den gængse dosisresponsmodel for én art. Den baner vejen for mere realistiske forudsigelser af, hvorledes herbicider påvirker hele plantesamfund.
I forbindelse med modellering af herbiciders effekt på planters vækst giver vore resultater følgende fingerpeg. Ved en forventet sluteffekt på 15-50% skal der i modellen indbygges en vækststandsning på minimum 3 uger efter herbicidbehandling. Forventes sluteffekten mindre end 15% vil vækststandsningen være så kort, at den ikke skal indgå i modeller, mens der ved forventede sluteffekter på mere end 50% er tale om en vækststandsning af længere varighed end 3 uger .
6 Perspektiver
6.1 Forskningsmæssige perspektiver
I projektet undersøgtes konsekvenserne af de ændringer i sprøjtestrategien, som kan forventes ved skifte til GT-afgrøder, både på den landbrugsmæssige drift og den omkringliggende natur. GT-afgrøder er dog blot ét eksempel på, at en ny dyrkningspraksis kan medføre ændringer i sprøjtestrategien med deraf afledte positive og negative bivirkninger. Pløjefri dyrkning er f.eks. en aktuel ny dyrkningspraksis, som er tilknyttet en særlig sprøjtestrategi (hvori glyphosat også indgår). De metoder, der er blevet anvendt i dette projekt, og de værktøjer, der er blevet udviklet, vil med fordel kunne anvendes, hvis konsekvenserne af sådanne nye dyrkningspraktikker skal bedømmes. Blandt de nye metoder og værktøjer skal fremhæves
- flerårige forsøg med plantesamfunds succession under påvirkning af herbicider og gødning,
- dosisresponsforsøg udført ikke blot på enkeltarter, men på arter voksende i konkurrence med hinanden,
- statistisk model til analyse af sådanne kombinerede konkurrence- og dosisresponsforsøg
- simuleringsmodel (FieldEco) til analyse af, hvorledes herbicider påvirker flerårige plantearters vækst og formering,
- simuleringsmodel (FieldWeeds) til analyse af, hvorledes herbicider påvirker enårige plantearters populationsdynamik.
Selv om de anvendte modeller har givet os en ide om, hvilke effekter introduktionen af GT-afgrøder vil have på ukrudtsfloraens sammensætning, så er der behov for at følge disse simuleringer op med langtidsforsøg. Det gælder især for GT-majs, der som den eneste af de for Danmark relevante GT-afgrøder dyrkes ensidigt (det vil sige år efter år). Dyrkning af GT-majs kræver derfor, at der udvikles bekæmpelsesstrategier, som forebygger selektion af glyphosatresistente biotyper.
Det faktum, at ukrudtets frøproduktion i marken er mindre påvirket af herbicider end biomassen, betyder, at det specielt ved sene glyphosatsprøjtninger burde være muligt at bestemme doseringer, hvor ukrudtet bekæmpes tilfredsstillende, og der samtidigt er planter tilbage, som udvikler sig til frøbærere, således at dyrkning af GT-afgrøder ikke nødvendigvis føre til en udtømning af jordens frøreserver og deraf nedgang i biodiversiteten. At fastsætte en sådan ”økologisk” optimal dosering forudsætter, at fremtidige forsøg, der iværksættes for at fastlægge sprøjtevinduet for glyphosat (det vil sige det tidsrum, hvor der kan herbicidsprøjtes med sikkerhed for udbyttet), ikke blot skal fokusere på majsudbyttet, men også inddrage ukrudtets frøproduktion.
Der er stadig uklarhed omkring herbicidafdriftens effekt på reproduktionen hos planterne i hegnet omring marken. Reproduktionen har betydning for plantearternes overlevelse i agerlandet på længere sigt, for bestøvernes fødegrundlag og for den umiddelbare oplevelse af biodiveristet i form af blomsterflor.
Vi kunne ikke påvise en simpel sammenhæng mellem markens ukrudtsbiomasse og insektbiomassen i majs men har dog tidligere set, at mere ukrudt følges af flere insekter. Nærmere undersøgelser af sammenhængen mellem ukrudtet og højere niveauer i fødekæden bør formentlig foretages på artsniveau, hvorved man vil kunne afdække, om der er tale om en fødekædesammenhæng eller/og en effekt af ændret mikroklima.
6.2 Administrative perspektiver
Der er ikke fastsat en standarddosering for glyphosat i GT-majs, men da glyphosat vil erstatte anvendelsen af tankblandinger, kan det forventes, at med samme sprøjtepraksis (terminer) i GT-afgrøder som i konventionelle afgrøder vil pesticidforbruget blive reduceret. Vore undersøgelser viser, at en senere sprøjtning med glyphosat vil kræve højere doseringer, hvis effektkravet opretholdes, og dermed kan den øgede biodiversitet have en omkostning i form af en højere behandlingshyppighed.
På længere sigt udgør udviklingen af glyphosattolerante eller ligefrem glyphosatresistente ukrudtsarter den største risiko ved GT-afgrøder. Hvis sådanne biotyper udvikles (som følge af den ensidige brug af glyphosat, som GT-afgrøder muliggør) vil dette forøge herbicidforbruget og dermed behandlingshyppigheden.
I forhold til hegnsfloraen synes der med baggrund i simuleringerne af langtidseffekterne af glyphosatafdrift ikke at være behov for at regulere sprøjtning med glyphosat i GT-afgrøder anderledes end konventionelle sprøjtemidler. Ensidig dyrkning af GT-afgrøder bør dog forhindres for at undgå en ensidig påvirkning af miljøet i og omkring marken.
For at forebygge ovenstående risici bør indførelse af GT-afgrøder følges af regler, som sikrer, at glyphosat ikke anvendes ensidigt. Dette kan for eksempel sikres ved, at GT-afgrøder kræves dyrket i sædskite med ikke-GT-afgrøder.
Projektet har vist, at det er herbicidanvendelsen i forbindelse med de enkelte afgrøder, der har betydning for effekten, og ikke om der dyrkes GMO eller ikke-GMO afgrøder i sædskifterne. Dette var også en af hovedkonklusionerne fra de britiske Farm Scale Evaluations, hvor man undersøgte effekten af GM-forårsafgrøder på biodiversiteten i og udenfor marken139. For at reducere belastningen af de marknære habitater er det derfor vigtigt med tiltag, der reducerer eksponeringen (for eksempel sprøjtefri randzoner, udtagning af marginale jorder eller jorder nær særligt værdifulde naturområder, eller forøgelse af det økologisk dyrkede areal). I en samlet vurdering af dyrkningens indvirkning på de marknære habitater bør ud over belastningen med herbicider også indgå kvælstof, idet kvælstof ligesom herbicider forårsager en forarmning af floraen.
Litteratur
Nummerorden
1. Lassen,J., Nielsen,D.E., Vestergaard,L. & Sandøe,P. Miljøvenlige genmodificerede afgrøder? Vil landmændene have dem, og vil de blive brugt til gavn for naturen? Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 112. (2007).
2. Castle,L.A. et al. Discovery and directed evolution of a glyphosate tolerance gene. Science 304, 1151-1154 (2004).
3. Lassen,J., Holm,L. & Sandøe,P. Vedr. bioteknologi og offentlighed. Tveit,G. (ed.), pp. 10-14 (Center for Bioetik og Risikovurdering, 2003).
4. Lassen,J. & Jamison,A. Genetic technologies meet the public: the discourses of concern. Science Technology & Human Values 31, 8-28 (2006).
5. Moll,S. Commercial experience and benefits from glyphosate tolerant crops. Brighton Crop Protection Conference - Weeds , 931-940. (1997).
6. Jensen,P.E. Doseringsbehov ved kemisk ukrudtsbekæmpelse i almindelige og glyphosatresistente bederoer. 15. Danske Planteværnskonference.Ukrudt. 115-123. (1998).
7. Kudsk,P. & Mathiassen,S.K. Ukrudtsbekæmpelse i glyphosatresistente bederoer - betydningen af ukrudtets størrelse. 15. Danske Planteværnskonference.Ukrudt , 125-132. (1998).
8. Buckmann,H., Petersen,J., Schlinker,G. & Marlander,B. Weed control in genetically modified sugar beet - two year experiences of a field trial series in Germany. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz Sonderheft S7, 353-362 (2000).
9. Tharp,B.E. & Kells,J.J. Influence of herbicide application rate, timing, and inter-row cultivation on weed control in corn (Zea mays) yield in glufosinate-resistant and glyphosat-resistant corn. Weed Technology 13, 807-813 (1999).
10. Elmegaard,N. & Bruus Pedersen,M. Flora and fauna in roundup tolerant fodder beet fields. NERI Technical Report No.349 . (2001).
11. Strandberg,B. & Bruus Pedersen,M. Biodiversity on glyphosate tolerant fodder beet fields. NERI Technical Report No.410. (2002).
12. Strandberg,B., Bruus Pedersen,M. & Elmegaard,N. Weed and arthropod populations in conventional and genetically modified herbicide tolerant fodder beet fields. Agriculture, Ecosystems and Environment 105, 243-253 (2005).
13. Squire,G.R. et al. On the rationale and interpretation of the Farm Scale Evaluations of genetically modified herbicide-tolerant crops . Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1779-1799 (2003).
14. Heard,M.S. et al. Weeds in fields with contrasting conventional and genetically modified herbicide-tolerant crops. I. Effects on abundance and diversity. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1819-1832 (2003).
15. Heard,M.S. et al. Weeds in fields with contrasting conventional and genetically modified herb icide-tolerant crops. II. Effects on individual species. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1833-1846 (2003).
16. Brooks,D.R. et al. Invertebrate responses to the management of genetically modified herbicide-tolerant and conventional spring crops.I. Soil-surface-active invertebrates. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1847-1862 (2003).
17. Hawes,C. et al. Responses of plants and invertebrate trophic groups to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1899-1913 (2003).
18. Roy,D.B. et al. Invertebrates and vegetation of field margins adjacent to crops subject to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1879-1898 (2003).
19. Haughton,A.J. et al. Invertebrate responses to the management of genetically modified herbicide-tolerant and conventional spring crops. II. Within-field epigeal and aerial arthropods. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1863-1877 (2003).
20. Miljøstyrelsen. Bichel-udvalget. Rapport fra underudvalget for miljø og sundhed. (1999).
21. Collings,L.V., Cussans,J.W. & Oswin,A.L. Testing a simple approach to estimating the seed production of arable weeds within contrasting crops. Proceedings of the 13th EWRS Symposium, 19-23 June 2005, Bari, Italy. (2005).
22. Rasmussen,I.A. Will weed seed production become a problem by use of reduced doses of herbicides in cereal crops? Tidsskrift for Planteavls Specialserie 71-81 (1993).
23. Kleijn,D. & Snoeijing,G.I.J. Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34, 1413-1425 (1997).
24. Bruus Pedersen,M., Aude,E. & Tybirk,K. Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetation. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 87, 1-103. (2004).
25. ISAAA. International Service for the Acquisition of Agri-biotech Applications. (2007).
26. Madsen,K.H., Thomsen,J.N., Strandberg,B. & Sandøe,P. GM sugar beet – What are the concerns? Advances in Sugar Beet Research 25-44 (2005).
27. Melander,B. Ukrudtsbekæmpelse i Landbruget, pp. 41-190. DJF , Flakkebjerg. (1998).
28. Jensen,H.A. & Kjellson,G. Frøpuljens størrelse og dynamik i moderne landbrug 1, 1-141. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr.13. (1995)
29. Kjellson,G. & Rasmussen,K. Frøpuljens størrelse og dynamik i moderne landbrug 2. 1-98. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 14. (1995).
30. Jordan,N. Weed demography and population dynamics: implications for threshold management. Weed Technology 6, 184-190 (1992).
31. Mortimer,A.M. & Putwain,P.D. The prediction of weed infestations: concepts and approaches. Bulletin OEPP 14, 439-446 (1984).
32. Cousens,R.D. Can we determine the intrinsic dynamics of real plant populations? Functional Ecology 9, 15-20 (1995).
33. Kropff,M.J., Wallinga,J. & Lotz,L.A.P. Weed population dynamics. Brown, H. et al. eds.. Proceedings of the second international weed control congress, Copenhagen, Denmark, 25-28 June 1996, 3-14. (1996).
34. Oreskes,N., Shrader-Frechete,K. & Belitz,K. Verification, validation, and confirmation of numerical models in the early sciences. Science 263 , 641-646 (1994).
35. Holst,N., Rasmussen,I.A. & Bastiaans,L. Field weed population dynamics: a review of model approaches and applications. Weed Research 47, 1-14 (2007).
36. Siegel,S. & Castellan,N.J., Jr. Nonparametric Statistics for the Behavioral Sciences, 2.ed. McGraw-Hill Book Co. (1988).
37. Cousens,R.D. & Moss,S.R. A model of the effects of cultivation on the vertical distribution of weed seeds within the soil. Weed Research 30, 61-70 (1990).
38. Lutman,P.J.W. et al. The persistence of seeds of 16 weed species over six years in two arable fields. Weed Research 42, 231-241 (2002).
39. Roberts,H.A. & Boddrell,J.E. Seed survival and periodicity of seedling emergence in eight species of Cruciferae. Annals of Applied Biology 103, 301-304 (1983).
40. Górski,T., Górska,K. & Nowicki,J. Germination of seeds of various herbaceous species under a leaf canopy. Flora Bd. 166, 249-259 (1977).
41. Paolini,R., Principi,M., Froud Williams,R.J., Del Puglia,S. & Biancardi,E. Competition between sugarbeet and Sinapis arvensis and Chenopodium album, as affected by timing of nitrogen fertilization. Weed Research 39, 425-440 (1999).
42. Chepil,W.S. Germination of seeds. I. Longevity, periodicity of germination, and vitality of seeds in cultivated soil. Scientific Agriculture 26, 307-346 (1946).
43. Aarts,H.F.M. A computerized model for predicting changes in a population of Galium aparine. Proceedings of the 5th EWRS Symposium on Economic Weed Control, Stuttgart-Hohenheim 1986 , 277-284. 1986.
44. Benjamin,L.R. & Cussans,J.W. Impact of spring germinating weed seedlings in autumn-sown crops on weed seed bank dynamics. Aspects of Applied Biology 69 , 269-276 (2003).
45. Zwerger,P. & Hurle,K. Untersuchungen zur relativen Bedeutung einzelner populationsdynamischer Parameter fur die Entwicklung der Verunkrautung. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 96, 346-352 (1989).
46. Zwerger,P. & Hurle,K. Untersuchungen zur Abbildungsgute simulierter Befallsverlaufe bei Unkrautern. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 97, 133-141 (1990).
47. Niemann,P. Zur Variabilität der Klettenlabktautes (Galium aparine). Gesunde Pflanzen 40, 368-373 (1988).
48. Mennan,H. & Ngouajio,M. Seasonal cycles in germination and seedling emergence of summer and winter populations of catchweed bedstraw (Galium aparine) and wild mustard (Brassica kaber). Weed Science 54, 114-120 (2006).
49. Chancellor,R.J. Decline in arable weed seeds during 20 years in soil under grass and the periodicity of seedling emergence after cultivation. Journal of Applied Ecology 23, 631-637 (1986).
50. Conn,J.S. & Deck,R.E. Seed viability and dormancy of 17 weed species after 9.7 years of burial in Alaska. Weed Science 43, 583-585 (1995).
51. Defelice,M.S. Common chickweed, Stellaria media (L.) Vill. - "Mere chicken feed?". Weed Technology 18, 193-200 (2004).
52. Noronha,A., Andersson,L. & Milberg,P. Rate of change in dormancy level and light requirement in weed seeds during stratification. Annals of Botany 80, 795-801 (1997).
53. Briggs,D., Hodkinson,H. & Block,M. Precociously developing individuals in populations of chickweed [Stellaria media (L) Vill] from different habitat types, with special reference to the effects of weed control measures. New Phytologist 117, 153-164 (1991).
54. Grundy,A.C. et al. Emergence of Chenopodium album and Stellaria media of different origins under different climatic conditions. Weed Research 43, 163-176 (2003).
55. Grundy,A.C., Mead,A. & Burston,S. Modelling the emergence response of weed seeds to burial depth: interactions with seed density, weight and shape. Journal of Applied Ecology 40, 757-770 (2003).
56. Mulugeta,D. & Stoltenberg,D.E. Influence of cohorts on Chenopodium album demography. Weed Science 46, 65-70 (1998).
57. Williams,J.T. Biological flora of the British Isles. Chenopodium album L. Journal of Ecology 51, 711-725 (1963).
58. Benvenuti,S., Macchia,M. & Miele,S. Quantitative analysis of emergence of seedlings from buried seeds with increasing soil depth. Weed Science 49, 528-535 (2001).
59. Chancellor,R.J. The seasonal emergence of dicotyledonous weed seedlings with changing temperature. Proceedings of the 3rd EWRS Symposium, Mainz, 1979, 65-71. (1979).
60. Roberts,H.A. Emergence and longevity in cultivated soil of seeds of some annual weeds. Weed Research 4, 296-307 (1964).
61. van der Weide,R.Y. Phenology of arable and hedgerow populations of Galium aparine L. in relation to climate and soil conditions. Weed Research 32, 249-258 (1992).
62. Auge,H. & Mahn,E.G. Experimentelle Untersuchungen zur Populationsökologie von Gallium aparine L. (Rubiaceae). Verhandlungen der Gesellschaft für Ökologie 20, 917-926 (1991).
63. Miura,R. & Kusanagi,T. Variation in the factors determining flowering time in the Stellaria media complex. Weed Research 41, 69-81 (2001).
64. Sobey,D.G. Stellaria media (L) Vill. Journal of Ecology 69, 311-335 (1981).
65. Huang,J.Z. et al. Effect of temperature and photoperiod on the phenological development of common lambsquarters. Weed Science 49, 500-508 (2001).
66. Kropff,M.J., Spitters,C.J.T., Schnieders,B.J., Joenje,W. & de Groot,W. An ecophysiological model for interspecific competition, applied to the influence of Chenopodium album L. on sugar-beet. 2. Model evaluation. Weed Research 32, 451-463 (1992).
67. Kay,Q.O.N. Tripleurospermum inodorum (L) Schultz Bip. Journal of Ecology 82, 681-697 (1994).
68. Cousens,R.D. A simple model relating yield loss to weed density. Annals of Applied Biology 107, 239-252 (1985).
69. Storkey,J. A functional group approach to the management of UK arable weeds to support biological diversity. Weed Research 46, 513-522 (2006).
70. Blackshaw,R.E., Anderson,G.W. & Dekker,J. Interference of Sinapis arvensis L. and Chenopodium album L. in spring rapeseed (Brassica napus L.). Weed Research 27, 207-213 (1987).
71. Wilson,B.J. & Wright,K.J. Predicting the growth and competitive effects of annual weeds in wheat. Weed Research 30, 201-211 (1990).
72. Storkey,J., Cussans,J.W., Lutman,P.J.W. & Blair,A.M. The importance of mortality in weed populations between autumn and spring on the reliability of yield loss predictions in winter wheat. 1997 Brighton Crop Protection Conference - Weeds , 1025-1030. (1997).
73. Seavers,G.P. & Wright,K.J. Crop canopy development and structure influence weed suppression. Weed Research 39, 319-328 (1999).
74. Mennan,H. & Zandstra,B.H. Effect of wheat (Triticum aestivum) cultivars and seeding rate on yield loss from Galium aparine (cleavers). Crop Protection 24, 1061-1067 (2005).
75. Dicke,D., Fries,A. & Gerhards,R. Determination of weed thresholds for site-specific weed control in malting barley. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz-Journal of Plant Diseases and Protection 413-421 (2004).
76. Lutman,P.J.W., Bowerman,P., Palmer,G.M. & Whytock,G.P. Prediction of competition between oilseed rape and Stellaria media. Weed Research 40, 255-269 (2000).
77. Conn,J.S. & Thomas,D.L. Common lambsquarters (Chenopodium album) interference in spring barley. Weed Technology 1, 312-313 (1987).
78. Swinton,S.M., Buhler,D.D., Forcella,F., Gunsolus,J.L. & King,R.P. Estimation of crop yield loss due to interference by multiple weed species. Weed Science 42, 103-109 (1994).
79. Weaver,S.E. Impact of lamb's-quarters, common ragweed and green foxtail on yield of corn and soybean in Ontario. Canadian Journal of Plant Science 81, 821-828 (2001).
80. Moechnig,M.J., Boerboom,C.M., Stoltenberg,D.E. & Binning,L.K. Growth interactions in communities of common lambsquarters (Chenopodium album), giant foxtail (Setaria faberi), and corn. Weed Science 51, 363-370 (2003).
81. Schweizer,E.E. Common lambsquarters (Chenopodium album) interference in sugarbeets (Beta vulgaris). Weed Science 31, 5-8 (1983).
82. Jensen,P.K. Comparison of different weed-rankings as predictors of yield loss. Bekæmpelsesmiddelforskning fra Miljøstyrelsen 22, (1996).
83. Peters,R.H. The ecological implications of body size. Cambridge University Press, (1983).
84. Lutman,P.J.W. Estimation of seed production by Stellaria media, Sinapis arvensis and Tripleurospermum inodorum in arable crops. Weed Research 42, 359-369 (2002).
85. Franz,K., Kaiser,F. & Gerowitt,B. Wirkung unterschiedlich hoher Stickstoffdüngung auf Entwicklung und Samenproduktion ausgewählter Unkrautarten im Winterweizen. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz, Sonderheft 13, 127-135 (1990).
86. Wright,K.J. Weed seed production as affected by crop density and nitrogen application. 1993 Brighton Crop Protection Conference - Weeds , 275-280. (1993).
87. Mertens,S.K. & Jansen,J. Weed seed production, crop planting pattern, and mechanical weeding in wheat. Weed Science 50, 748-756 (2002).
88. Freckleton,R.P. & Watkinson,A.R. Predicting the determinants of weed abundance: a model for the population dynamics of Chenopodium album in sugar beet. Journal of Applied Ecology 35, 904-920 (1998).
89. Grundy,A.C., Mead,A., Burston,S. & Overs,T. Seed production of Chenopodium album in competition with field vegetables. Weed Research 44, 271-281 (2004).
90. Pedersen,J.O. & Rasmussen,I.A. The influence of herbicides on weed seed production. 7. Danske Planteværnskonference - Ukrudt 1990 , 73-83. (1990).
91. Shinozaki,K. & Kira,T. Intraspecific competition among higher plants. VII. Logistic theory of the C-D effect. J. Inst. Polytech, Osaka City University 7, 35-72 (1956).
92. Firbank,L.G. & Watkinson,A.R. On the analysis of competition within two-species mixtures of plants. Journal of Applied Ecology 22, 503-517 (1985).
93. Law,R. & Watkinson,A.R. Response-surface analysis of 2-species competition - an experiment on Phleum arenarium and Vulpia fasciculata. Journal of Ecology 75, 871-886 (1987).
94. Cousens,R.D. Aspects of the design and interpretation of competition experiments. Weed Technology 5, 664-673 (1991).
95. Damgaard,C. Plant competition experiments: Testing hypotheses and estimating the probability of coexistence. Ecology 79, 1760-1767 (1998).
96. Damgaard,C. Modeling plant competition along an environmental gradient. Ecological Modelling 170, 45-53 (2003).
97. Damgaard,C. Evolutionary ecology of plant-plant interactions - an empirical modelling approach. Aarhus University Press, Aarhus (2004).
98. Seber,G.A.F. & Wild,C.J. Nonlinear regression. John Wiley & Sons, New York (1989).
99. Carlin,B.P. & Louis,T.A. Bayes and empirical Bayes methods for data analysis. Chapman & Hall, London (1996).
100. Hansen,K. Dansk Feltflora. Gyldendal, (1991).
101. Levy,E.B. & Madden,E.A. The point method of pasture analysis. New Zealand Journal of Agriculture 18, 267-279 (1933).
102. Hays,R.L., Summers,C. & Seitz,W. Estimating Wildlife Habitat variables. Biological Services Program, Fish and Wildlife Service, US Department of the Interior (1981).
103. Goodall,D.W. Some considerations in the use of point quadrats for analysis of vegetation. Australian Journal of Science Research 5, 1-41(1952).
104. Jonasson,S. Evaluation of the point intercept metod for the estimation of plant biomass. Oikos 52, 101-106 (1988).
105. Jonasson,S. The point intercept method for non-destructive estimation of biomass. Phytocoenologia 11, 385-388 (1983).
106. Cromar,H.E., Murphy,S.D. & Swanton,C.J. Influence of tillage and crop residue on postdispersal predation of weed seeds. Weed Science 47, 184-194 (1999).
107. Holmes,R.J. & Froud-Williams,R.J. Post-dispersal weed seed predation by avian and non-avian predators. Agriculture Ecosystems & Environment 105, 23-27 (2005).
108. Westerman,P.R., Wes,J.S., Kropff,M.J. & van der Werf,W. Annual losses of weed seeds due to predation in organic cereal fields. Journal of Applied Ecology 40, 824-836 (2003).
109. Thompson,K., Bakker,J. & Bekker,R. The soil seed banks of North West Europe: methodology, density and longevity. (1997).
110. de Snoo,G.R. & de Wit,P.J. Buffer zones for reducing pesticide drift to ditches and risks to aquatic organisms. Ecotoxicology and Environmental Safety 41, 112-118 (1998).
111. de Snoo,G.R. Unsprayed field margins: effect on environment biodiversity and agricultural practice. Landscape and Urban Planning 46, 151-160 (1999).
112. Boutin,C., Jobin,B., Belanger,L. & Choiniere,L. Comparing weed composition in natural and planted hedgerows and in herbaceous field margins adjecent to crop fields. Canadian Journal of Plant Science 81, 313-324 (2001).
113. Damgaard,C., Mathiassen,S.K. & Kudsk,P. Modelling effects of herbicide drift on the competitive interactions between weeds. Enviromental Toxicology and Chemistry 27 (in press), (2008).
114. Pline,W.A., Edmisten,K.L., Wilcut,J.W., Wells,R. & Thomas,J. Glyphosate-induced reductions in pollen viability and seed set in glyphosate-resistant cotton and attempted remediation by gibberellic acid (GA(3)). Weed Science 51, 19-27 (2003).
115. Thomas,W.E., Pline-Srnic,W.A., Viator,R.P. & Wilcut,J.W. Effects of glyphosate application timing and rate on sicklepod (Senna obtusifolia) fecundity. Weed Technology 19, 55-61 (2005).
116. Kudsk,P. Weed Management Handbook. Naylor,R.E.L. (ed.), pp. 323-344 (Blackwell Publishing, Oxford, UK,2002).
117. Owen,M.D.K. Genetically modified crops: successes and problems in the Midwest USA. Proc.XVI International Plant Protection Congress 2007,Glasgow, UK , 506-507. (2007).
118. Graglia,E., Melander,B. & Jensen,R.K. Mechanical and cultural strategies to control Cirsium arvense in organic arable cropping systems. Weed Research 46, 304-312 (2006).
119. Aude,E., Tybirk,K. & Bruus Pedersen,M. Vegetation diversity of conventional and organic hedgerows in Denmark. Agriculture, Ecosystems & Environment 99, 135-147 (2003).
120. Aude,E. et al. Conservation value of the herbaceous vegetation in hedgerows - does organic farming make a difference? Biological Conservation 118, 467-478 (2004).
121. Bruus,M. et al. Omfang og effekt af herbicidafdrift til danske læhegn. Bekæmpelsesmiddelforskning fra Miljøstyrelsen. (2008).
122. Marrs,R.H., Williams,C.T., Frost,A.J. & Plant,R.A. Assessment of the effects of herbicide spray drift on a range of plant species of conservation interest. Environmental Pollution 59, 71-86 (1989).
123. Marrs,R.H., Frost,A.J. & Plant,R.A. Effect of herbicide spray drift on selected species of nature conservation interest: effects of plant age and surrounding vegetation structure. Environmental Pollution 69, 223-235 (1991).
124. Marrs,R.H. & Frost,A.J. A microcosm approach to the detection of the effects of herbicide spray drift in plant communities. Journal of Environmental Management 50, 369-388 (1997).
125. Jobin,B., Boutin,C. & Desgranges,J.L. Effects of agricultural practices on the flora of hedgerows and woodland edges in southern Quebec. Canadian Journal of Plant Science 77, 293-299 (1997).
126. Kleijn,D. & Verbeek,M. Factors affecting the species composition of arable field boundary vegetation. Journal of Applied Ecology 37, 256-266 (2000).
127. OEPP/EPPO. OEPP/EPPO Standards PP1, vol. 4. Efficacy evaluation of herbicides and plant growth regulators. (2004).
128. Asman,W.A.H. et al. Pesticides in air and in precipitation and effects on plant communities. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr.57. (2001).
129. Wilson,R.G. et al. Glyphosate-induced weed shifts in glyphosate-resistant corn or a rotation of glyphosate-resistant corn, sugarbeet, and spring wheat. Weed Technology 21, 900-909 (2007).
130. Tuesca,D. & Puricelli,E. Effect of tillage systems and herbicide treatments on weed abundance and diversity in a glyphosate resistant crop rotation. Crop Protection 26, 1765-1770 (2007).
131. Cerdeira,A.L., Gazziero,D.L.P., Duke,S.O., Matallo,M.B. & Spadotto,C.A. Review of potential environmental impacts of transgenic glyphosate-resistant soybean in Brazil. Journal of Environmental Science and Health Part B-Pesticides Food Contaminants and Agricultural Wastes 42, 539-549 (2007).
132. Cerdeira,A.L. & Duke,S.O. The current status and environmental impacts of glyphosate-resistant crops: A review. Journal of Environmental Quality 35, 1633-1658 (2006).
133. Beckie,H.J. et al. A decade of herbicide-resistant crops in Canada. Canadian Journal of Plant Science 86, 1243-1264 (2006).
134. Culpepper,A.S. Glyphosate-induced weed shifts. Weed Technology 20, 277-281 (2006).
135. Owen,M.D.K. & Zelaya,I.A. Herbicide-resistant crops and weed resistance to herbicides. Pest Management Science 61, 301-311 (2005).
136. Sweet,J., Simpson,E. & Law,J. Botanical and rotational implications of genetically modified herbicide tolerance in winter oilseed rape and sugar beet (BRIGHT Project). HGCA, Project Report No.353 , 1-40. (2004).
137. Christensen,K. Herbicidet Starane 180S' effekt på pollenproduktionen hos mælkebøtte og rødkløver - påvirkninger af fødeudbudet i markhegn på konventionelle og økologiske brug. Specialerapport. KU-LIFE. (2008).
138. Yu,C. et al. Inducing male sterility in Brassica napus L. by a sulphonylurea herbicide, tribenuron-methyl. Plant Breeding 125, 61-64 (2006).
139. Firbank,L.G. et al. The implications of spring-sown genetically modified herbicide-tolerant crops for farmland biodiversity: A commentary on the Farm Scale Evaluations of spring sown crops. 1-37. London, UK, Defra Publications. (2003).
140. Ravn,H.W. et al. Har ukrudtsmidlet virket? Et nyt kit præsenteres. Plantekongres Herning 2006 , 63-65. (2006).
141. Cedergreen,N., Streibig,J., Kudsk,P. & Mathiassen,S.K. The occurence of hormesis in plants and algae. Dose-Reponse (International Hormesis Society) 5, 1-13 (2007).
Alfabetisk
43. Aarts,H.F.M. A computerized model for predicting changes in a population of Galium aparine. Proceedings of the 5th EWRS Symposium on Economic Weed Control, Stuttgart-Hohenheim 1986 , 277-284. 1986.
128. Asman,W.A.H. et al. Pesticides in air and in precipitation and effects on plant communities. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr.57. (2001).
120. Aude,E. et al. Conservation value of the herbaceous vegetation in hedgerows - does organic farming make a difference? Biological Conservation 118, 467-478 (2004).
119. Aude,E., Tybirk,K. & Bruus Pedersen,M. Vegetation diversity of conventional and organic hedgerows in Denmark. Agriculture, Ecosystems & Environment 99, 135-147 (2003).
62. Auge,H. & Mahn,E.G. Experimentelle Untersuchungen zur Populationsökologie von Gallium aparine L. (Rubiaceae). Verhandlungen der Gesellschaft für Ökologie 20, 917-926 (1991).
133. Beckie,H.J. et al. A decade of herbicide-resistant crops in Canada. Canadian Journal of Plant Science 86, 1243-1264 (2006).
44. Benjamin,L.R. & Cussans,J.W. Impact of spring germinating weed seedlings in autumn-sown crops on weed seed bank dynamics. Aspects of Applied Biology 69 , 269-276 (2003).
58. Benvenuti,S., Macchia,M. & Miele,S. Quantitative analysis of emergence of seedlings from buried seeds with increasing soil depth. Weed Science 49, 528-535 (2001).
70. Blackshaw,R.E., Anderson,G.W. & Dekker,J. Interference of Sinapis arvensis L. and Chenopodium album L. in spring rapeseed (Brassica napus L.). Weed Research 27, 207-213 (1987).
112. Boutin,C., Jobin,B., Belanger,L. & Choiniere,L. Comparing weed composition in natural and planted hedgerows and in herbaceous field margins adjecent to crop fields. Canadian Journal of Plant Science 81, 313-324 (2001).
53. Briggs,D., Hodkinson,H. & Block,M. Precociously developing individuals in populations of chickweed [Stellaria media (L) Vill] from different habitat types, with special reference to the effects of weed control measures. New Phytologist 117, 153-164 (1991).
16. Brooks,D.R. et al. Invertebrate responses to the management of genetically modified herbicide-tolerant and conventional spring crops.I. Soil-surface-active invertebrates. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1847-1862 (2003).
24. Bruus Pedersen,M., Aude,E. & Tybirk,K. Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetation. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 87, 1-103. (2004).
121. Bruus,M. et al. Omfang og effekt af herbicidafdrift til danske læhegn. Bekæmpelsesmiddelforskning fra Miljøstyrelsen. (2008).
8. Buckmann,H., Petersen,J., Schlinker,G. & Marlander,B. Weed control in genetically modified sugar beet - two year experiences of a field trial series in Germany. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz Sonderheft S7, 353-362 (2000).
99. Carlin,B.P. & Louis,T.A. Bayes and empirical Bayes methods for data analysis. Chapman & Hall, London (1996).
2. Castle,L.A. et al. Discovery and directed evolution of a glyphosate tolerance gene. Science 304, 1151-1154 (2004).
141. Cedergreen,N., Streibig,J., Kudsk,P. & Mathiassen,S.K. The occurence of hormesis in plants and algae. Dose-Reponse (International Hormesis Society) 5, 1-13 (2007).
132. Cerdeira,A.L. & Duke,S.O. The current status and environmental impacts of glyphosate-resistant crops: A review. Journal of Environmental Quality 35, 1633-1658 (2006).
131. Cerdeira,A.L., Gazziero,D.L.P., Duke,S.O., Matallo,M.B. & Spadotto,C.A. Review of potential environmental impacts of transgenic glyphosate-resistant soybean in Brazil. Journal of Environmental Science and Health Part B-Pesticides Food Contaminants and Agricultural Wastes 42, 539-549 (2007).
49. Chancellor,R.J. Decline in arable weed seeds during 20 years in soil under grass and the periodicity of seedling emergence after cultivation. Journal of Applied Ecology 23, 631-637 (1986).
59. Chancellor,R.J. The seasonal emergence of dicotyledonous weed seedlings with changing temperature. Proceedings of the 3rd EWRS Symposium, Mainz, 1979, 65-71. (1979).
42. Chepil,W.S. Germination of seeds. I. Longevity, periodicity of germination, and vitality of seeds in cultivated soil. Scientific Agriculture 26, 307-346 (1946).
137. Christensen,K. Herbicidet Starane 180S' effekt på pollenproduktionen hos mælkebøtte og rødkløver - påvirkninger af fødeudbudet i markhegn på konventionelle og økologiske brug. Specialerapport. KU-LIFE. (2008).
21. Collings,L.V., Cussans,J.W. & Oswin,A.L. Testing a simple approach to estimating the seed production of arable weeds within contrasting crops. Proceedings of the 13th EWRS Symposium, 19-23 June 2005, Bari, Italy. (2005).
50. Conn,J.S. & Deck,R.E. Seed viability and dormancy of 17 weed species after 9.7 years of burial in Alaska. Weed Science 43, 583-585 (1995).
77. Conn,J.S. & Thomas,D.L. Common lambsquarters (Chenopodium album) interference in spring barley. Weed Technology 1, 312-313 (1987).
37. Cousens,R.D. & Moss,S.R. A model of the effects of cultivation on the vertical distribution of weed seeds within the soil. Weed Research 30, 61-70 (1990).
68. Cousens,R.D. A simple model relating yield loss to weed density. Annals of Applied Biology 107, 239-252 (1985).
94. Cousens,R.D. Aspects of the design and interpretation of competition experiments. Weed Technology 5, 664-673 (1991).
32. Cousens,R.D. Can we determine the intrinsic dynamics of real plant populations? Functional Ecology 9, 15-20 (1995).
106. Cromar,H.E., Murphy,S.D. & Swanton,C.J. Influence of tillage and crop residue on postdispersal predation of weed seeds. Weed Science 47, 184-194 (1999).
134. Culpepper,A.S. Glyphosate-induced weed shifts. Weed Technology 20, 277-281 (2006).
97. Damgaard,C. Evolutionary ecology of plant-plant interactions - an empirical modelling approach. Aarhus University Press, Aarhus (2004).
96. Damgaard,C. Modeling plant competition along an environmental gradient. Ecological Modelling 170, 45-53 (2003).
95. Damgaard,C. Plant competition experiments: Testing hypotheses and estimating the probability of coexistence. Ecology 79, 1760-1767 (1998).
113. Damgaard,C., Mathiassen,S.K. & Kudsk,P. Modelling effects of herbicide drift on the competitive interactions between weeds. Enviromental Toxicology and Chemistry 27 (in press), (2008).
110. de Snoo,G.R. & de Wit,P.J. Buffer zones for reducing pesticide drift to ditches and risks to aquatic organisms. Ecotoxicology and Environmental Safety 41, 112-118 (1998).
111. de Snoo,G.R. Unsprayed field margins: effect on environment biodiversity and agricultural practice. Landscape and Urban Planning 46, 151-160 (1999).
51. Defelice,M.S. Common chickweed, Stellaria media (L.) Vill. - "Mere chicken feed?". Weed Technology 18, 193-200 (2004).
75. Dicke,D., Fries,A. & Gerhards,R. Determination of weed thresholds for site-specific weed control in malting barley. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz-Journal of Plant Diseases and Protection 413-421 (2004).
10. Elmegaard,N. & Bruus Pedersen,M. Flora and fauna in roundup tolerant fodder beet fields. NERI Technical Report No.349 . (2001).
92. Firbank,L.G. & Watkinson,A.R. On the analysis of competition within two-species mixtures of plants. Journal of Applied Ecology 22, 503-517 (1985).
139. Firbank,L.G. et al. The implications of spring-sown genetically modified herbicide-tolerant crops for farmland biodiversity: A commentary on the Farm Scale Evaluations of spring sown crops. 1-37. London, UK, Defra Publications. (2003).
85. Franz,K., Kaiser,F. & Gerowitt,B. Wirkung unterschiedlich hoher Stickstoffdüngung auf Entwicklung und Samenproduktion ausgewählter Unkrautarten im Winterweizen. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz, Sonderheft 13, 127-135 (1990).
88. Freckleton,R.P. & Watkinson,A.R. Predicting the determinants of weed abundance: a model for the population dynamics of Chenopodium album in sugar beet. Journal of Applied Ecology 35, 904-920 (1998).
103. Goodall,D.W. Some considerations in the use of point quadrats for analysis of vegetation. Australian Journal of Science Research 5, 1-41(1952).
40. Górski,T., Górska,K. & Nowicki,J. Germination of seeds of various herbaceous species under a leaf canopy. Flora Bd. 166, 249-259 (1977).
118. Graglia,E., Melander,B. & Jensen,R.K. Mechanical and cultural strategies to control Cirsium arvense in organic arable cropping systems. Weed Research 46, 304-312 (2006).
54. Grundy,A.C. et al. Emergence of Chenopodium album and Stellaria media of different origins under different climatic conditions. Weed Research 43, 163-176 (2003).
55. Grundy,A.C., Mead,A. & Burston,S. Modelling the emergence response of weed seeds to burial depth: interactions with seed density, weight and shape. Journal of Applied Ecology 40, 757-770 (2003).
89. Grundy,A.C., Mead,A., Burston,S. & Overs,T. Seed production of Chenopodium album in competition with field vegetables. Weed Research 44, 271-281 (2004).
100. Hansen,K. Dansk Feltflora. Gyldendal, (1991).
19. Haughton,A.J. et al. Invertebrate responses to the management of genetically modified herbicide-tolerant and conventional spring crops. II. Within-field epigeal and aerial arthropods. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1863-1877 (2003).
17. Hawes,C. et al. Responses of plants and invertebrate trophic groups to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1899-1913 (2003).
102. Hays,R.L., Summers,C. & Seitz,W. Estimating Wildlife Habitat variables. Biological Services Program, Fish and Wildlife Service, US Department of the Interior (1981).
15. Heard,M.S. et al. Weeds in fields with contrasting conventional and genetically modified herb icide-tolerant crops. II. Effects on individual species. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1833-1846 (2003).
14. Heard,M.S. et al. Weeds in fields with contrasting conventional and genetically modified herbicide-tolerant crops. I. Effects on abundance and diversity. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1819-1832 (2003).
107. Holmes,R.J. & Froud-Williams,R.J. Post-dispersal weed seed predation by avian and non-avian predators. Agriculture Ecosystems & Environment 105, 23-27 (2005).
35. Holst,N., Rasmussen,I.A. & Bastiaans,L. Field weed population dynamics: a review of model approaches and applications. Weed Research 47, 1-14 (2007).
65. Huang,J.Z. et al. Effect of temperature and photoperiod on the phenological development of common lambsquarters. Weed Science 49, 500-508 (2001).
25. ISAAA. International Service for the Acquisition of Agri-biotech Applications. (2007).
28. Jensen,H.A. & Kjellson,G. Frøpuljens størrelse og dynamik i moderne landbrug 1, 1-141. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr.13. (1995)
6. Jensen,P.E. Doseringsbehov ved kemisk ukrudtsbekæmpelse i almindelige og glyphosatresistente bederoer. 15. Danske Planteværnskonference.Ukrudt. 115-123. (1998).
82. Jensen,P.K. Comparison of different weed-rankings as predictors of yield loss. Bekæmpelsesmiddelforskning fra Miljøstyrelsen 22, (1996).
125. Jobin,B., Boutin,C. & Desgranges,J.L. Effects of agricultural practices on the flora of hedgerows and woodland edges in southern Quebec. Canadian Journal of Plant Science 77, 293-299 (1997).
104. Jonasson,S. Evaluation of the point intercept metod for the estimation of plant biomass. Oikos 52, 101-106 (1988).
105. Jonasson,S. The point intercept method for non-destructive estimation of biomass. Phytocoenologia 11, 385-388 (1983).
30. Jordan,N. Weed demography and population dynamics: implications for threshold management. Weed Technology 6, 184-190 (1992).
67. Kay,Q.O.N. Tripleurospermum inodorum (L) Schultz Bip. Journal of Ecology 82, 681-697 (1994).
29. Kjellson,G. & Rasmussen,K. Frøpuljens størrelse og dynamik i moderne landbrug 2. 1-98. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 14. (1995).
23. Kleijn,D. & Snoeijing,G.I.J. Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34, 1413-1425 (1997).
126. Kleijn,D. & Verbeek,M. Factors affecting the species composition of arable field boundary vegetation. Journal of Applied Ecology 37, 256-266 (2000).
66. Kropff,M.J., Spitters,C.J.T., Schnieders,B.J., Joenje,W. & de Groot,W. An ecophysiological model for interspecific competition, applied to the influence of Chenopodium album L. on sugar-beet. 2. Model evaluation. Weed Research 32, 451-463 (1992).
33. Kropff,M.J., Wallinga,J. & Lotz,L.A.P. Weed population dynamics. Brown, H. et al. eds.. Proceedings of the second international weed control congress, Copenhagen, Denmark, 25-28 June 1996, 3-14. (1996).
7. Kudsk,P. & Mathiassen,S.K. Ukrudtsbekæmpelse i glyphosatresistente bederoer - betydningen af ukrudtets størrelse. 15. Danske Planteværnskonference.Ukrudt , 125-132. (1998).
116. Kudsk,P. Weed Management Handbook. Naylor,R.E.L. (ed.), pp. 323-344 (Blackwell Publishing, Oxford, UK,2002).
4. Lassen,J. & Jamison,A. Genetic technologies meet the public: the discourses of concern. Science Technology & Human Values 31, 8-28 (2006).
3. Lassen,J., Holm,L. & Sandøe,P. Vedr. bioteknologi og offentlighed. Tveit,G. (ed.), pp. 10-14 (Center for Bioetik og Risikovurdering, 2003).
1. Lassen,J., Nielsen,D.E., Vestergaard,L. & Sandøe,P. Miljøvenlige genmodificerede afgrøder? Vil landmændene have dem, og vil de blive brugt til gavn for naturen? Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 112. (2007).
93. Law,R. & Watkinson,A.R. Response-surface analysis of 2-species competition - an experiment on Phleum arenarium and Vulpia fasciculata. Journal of Ecology 75, 871-886 (1987).
101. Levy,E.B. & Madden,E.A. The point method of pasture analysis. New Zealand Journal of Agriculture 18, 267-279 (1933).
84. Lutman,P.J.W. Estimation of seed production by Stellaria media, Sinapis arvensis and Tripleurospermum inodorum in arable crops. Weed Research 42, 359-369 (2002).
38. Lutman,P.J.W. et al. The persistence of seeds of 16 weed species over six years in two arable fields. Weed Research 42, 231-241 (2002).
76. Lutman,P.J.W., Bowerman,P., Palmer,G.M. & Whytock,G.P. Prediction of competition between oilseed rape and Stellaria media. Weed Research 40, 255-269 (2000).
26. Madsen,K.H., Thomsen,J.N., Strandberg,B. & Sandøe,P. GM sugar beet – What are the concerns? Advances in Sugar Beet Research 25-44 (2005).
124. Marrs,R.H. & Frost,A.J. A microcosm approach to the detection of the effects of herbicide spray drift in plant communities. Journal of Environmental Management 50, 369-388 (1997).
123. Marrs,R.H., Frost,A.J. & Plant,R.A. Effect of herbicide spray drift on selected species of nature conservation interest: effects of plant age and surrounding vegetation structure. Environmental Pollution 69, 223-235 (1991).
122. Marrs,R.H., Williams,C.T., Frost,A.J. & Plant,R.A. Assessment of the effects of herbicide spray drift on a range of plant species of conservation interest. Environmental Pollution 59, 71-86 (1989).
27. Melander,B. Ukrudtsbekæmpelse i Landbruget, pp. 41-190. DJF , Flakkebjerg. (1998).
48. Mennan,H. & Ngouajio,M. Seasonal cycles in germination and seedling emergence of summer and winter populations of catchweed bedstraw (Galium aparine) and wild mustard (Brassica kaber). Weed Science 54, 114-120 (2006).
74. Mennan,H. & Zandstra,B.H. Effect of wheat (Triticum aestivum) cultivars and seeding rate on yield loss from Galium aparine (cleavers). Crop Protection 24, 1061-1067 (2005).
87. Mertens,S.K. & Jansen,J. Weed seed production, crop planting pattern, and mechanical weeding in wheat. Weed Science 50, 748-756 (2002).
20. Miljøstyrelsen. Bichel-udvalget. Rapport fra underudvalget for miljø og sundhed. (1999).
63. Miura,R. & Kusanagi,T. Variation in the factors determining flowering time in the Stellaria media complex. Weed Research 41, 69-81 (2001).
80. Moechnig,M.J., Boerboom,C.M., Stoltenberg,D.E. & Binning,L.K. Growth interactions in communities of common lambsquarters (Chenopodium album), giant foxtail (Setaria faberi), and corn. Weed Science 51, 363-370 (2003).
5. Moll,S. Commercial experience and benefits from glyphosate tolerant crops. Brighton Crop Protection Conference - Weeds , 931-940. (1997).
31. Mortimer,A.M. & Putwain,P.D. The prediction of weed infestations: concepts and approaches. Bulletin OEPP 14, 439-446 (1984).
56. Mulugeta,D. & Stoltenberg,D.E. Influence of cohorts on Chenopodium album demography. Weed Science 46, 65-70 (1998).
47. Niemann,P. Zur Variabilität der Klettenlabktautes (Galium aparine). Gesunde Pflanzen 40, 368-373 (1988).
52. Noronha,A., Andersson,L. & Milberg,P. Rate of change in dormancy level and light requirement in weed seeds during stratification. Annals of Botany 80, 795-801 (1997).
127. OEPP/EPPO. OEPP/EPPO Standards PP1, vol. 4. Efficacy evaluation of herbicides and plant growth regulators. (2004).
34. Oreskes,N., Shrader-Frechete,K. & Belitz,K. Verification, validation, and confirmation of numerical models in the early sciences. Science 263 , 641-646 (1994).
135. Owen,M.D.K. & Zelaya,I.A. Herbicide-resistant crops and weed resistance to herbicides. Pest Management Science 61, 301-311 (2005).
117. Owen,M.D.K. Genetically modified crops: successes and problems in the Midwest USA. Proc.XVI International Plant Protection Congress 2007,Glasgow, UK , 506-507. (2007).
41. Paolini,R., Principi,M., Froud Williams,R.J., Del Puglia,S. & Biancardi,E. Competition between sugarbeet and Sinapis arvensis and Chenopodium album, as affected by timing of nitrogen fertilization. Weed Research 39, 425-440 (1999).
90. Pedersen,J.O. & Rasmussen,I.A. The influence of herbicides on weed seed production. 7. Danske Planteværnskonference - Ukrudt 1990 , 73-83. (1990).
83. Peters,R.H. The ecological implications of body size. Cambridge University Press, (1983).
114. Pline,W.A., Edmisten,K.L., Wilcut,J.W., Wells,R. & Thomas,J. Glyphosate-induced reductions in pollen viability and seed set in glyphosate-resistant cotton and attempted remediation by gibberellic acid (GA(3)). Weed Science 51, 19-27 (2003).
22. Rasmussen,I.A. Will weed seed production become a problem by use of reduced doses of herbicides in cereal crops? Tidsskrift for Planteavls Specialserie 71-81 (1993).
140. Ravn,H.W. et al. Har ukrudtsmidlet virket? Et nyt kit præsenteres. Plantekongres Herning 2006 , 63-65. (2006).
39. Roberts,H.A. & Boddrell,J.E. Seed survival and periodicity of seedling emergence in eight species of Cruciferae. Annals of Applied Biology 103, 301-304 (1983).
60. Roberts,H.A. Emergence and longevity in cultivated soil of seeds of some annual weeds. Weed Research 4, 296-307 (1964).
18. Roy,D.B. et al. Invertebrates and vegetation of field margins adjacent to crops subject to contrasting herbicide regimes in the Farm Scale Evaluations of genetically modified herbicide-tolerant crops. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1879-1898 (2003).
81. Schweizer,E.E. Common lambsquarters (Chenopodium album) interference in sugarbeets (Beta vulgaris). Weed Science 31, 5-8 (1983).
73. Seavers,G.P. & Wright,K.J. Crop canopy development and structure influence weed suppression. Weed Research 39, 319-328 (1999).
98. Seber,G.A.F. & Wild,C.J. Nonlinear regression. John Wiley & Sons, New York (1989).
91. Shinozaki,K. & Kira,T. Intraspecific competition among higher plants. VII. Logistic theory of the C-D effect. J. Inst. Polytech, Osaka City University 7, 35-72 (1956).
36. Siegel,S. & Castellan,N.J., Jr. Nonparametric Statistics for the Behavioral Sciences, 2.ed. McGraw-Hill Book Co. (1988).
64. Sobey,D.G. Stellaria media (L) Vill. Journal of Ecology 69, 311-335 (1981).
13. Squire,G.R. et al. On the rationale and interpretation of the Farm Scale Evaluations of genetically modified herbicide-tolerant crops . Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 358, 1779-1799 (2003).
69. Storkey,J. A functional group approach to the management of UK arable weeds to support biological diversity. Weed Research 46, 513-522 (2006).
72. Storkey,J., Cussans,J.W., Lutman,P.J.W. & Blair,A.M. The importance of mortality in weed populations between autumn and spring on the reliability of yield loss predictions in winter wheat. 1997 Brighton Crop Protection Conference - Weeds , 1025-1030. (1997).
11. Strandberg,B. & Bruus Pedersen,M. Biodiversity on glyphosate tolerant fodder beet fields. NERI Technical Report No.410. (2002).
12. Strandberg,B., Bruus Pedersen,M. & Elmegaard,N. Weed and arthropod populations in conventional and genetically modified herbicide tolerant fodder beet fields. Agriculture, Ecosystems and Environment 105, 243-253 (2005).
136. Sweet,J., Simpson,E. & Law,J. Botanical and rotational implications of genetically modified herbicide tolerance in winter oilseed rape and sugar beet (BRIGHT Project). HGCA, Project Report No.353 , 1-40. (2004).
78. Swinton,S.M., Buhler,D.D., Forcella,F., Gunsolus,J.L. & King,R.P. Estimation of crop yield loss due to interference by multiple weed species. Weed Science 42, 103-109 (1994).
9. Tharp,B.E. & Kells,J.J. Influence of herbicide application rate, timing, and inter-row cultivation on weed control in corn (Zea mays) yield in glufosinate-resistant and glyphosat-resistant corn. Weed Technology 13, 807-813 (1999).
115. Thomas,W.E., Pline-Srnic,W.A., Viator,R.P. & Wilcut,J.W. Effects of glyphosate application timing and rate on sicklepod (Senna obtusifolia) fecundity. Weed Technology 19, 55-61 (2005).
109. Thompson,K., Bakker,J. & Bekker,R. The soil seed banks of North West Europe: methodology, density and longevity. (1997).
130. Tuesca,D. & Puricelli,E. Effect of tillage systems and herbicide treatments on weed abundance and diversity in a glyphosate resistant crop rotation. Crop Protection 26, 1765-1770 (2007).
61. van der Weide,R.Y. Phenology of arable and hedgerow populations of Galium aparine L. in relation to climate and soil conditions. Weed Research 32, 249-258 (1992).
79. Weaver,S.E. Impact of lamb's-quarters, common ragweed and green foxtail on yield of corn and soybean in Ontario. Canadian Journal of Plant Science 81, 821-828 (2001).
108. Westerman,P.R., Wes,J.S., Kropff,M.J. & van der Werf,W. Annual losses of weed seeds due to predation in organic cereal fields. Journal of Applied Ecology 40, 824-836 (2003).
57. Williams,J.T. Biological flora of the British Isles. Chenopodium album L. Journal of Ecology 51, 711-725 (1963).
71. Wilson,B.J. & Wright,K.J. Predicting the growth and competitive effects of annual weeds in wheat. Weed Research 30, 201-211 (1990).
129. Wilson,R.G. et al. Glyphosate-induced weed shifts in glyphosate-resistant corn or a rotation of glyphosate-resistant corn, sugarbeet, and spring wheat. Weed Technology 21, 900-909 (2007).
86. Wright,K.J. Weed seed production as affected by crop density and nitrogen application. 1993 Brighton Crop Protection Conference - Weeds , 275-280. (1993).
138. Yu,C. et al. Inducing male sterility in Brassica napus L. by a sulphonylurea herbicide, tribenuron-methyl. Plant Breeding 125, 61-64 (2006).
46. Zwerger,P. & Hurle,K. Untersuchungen zur Abbildungsgute simulierter Befallsverlaufe bei Unkrautern. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 97, 133-141 (1990).
45. Zwerger,P. & Hurle,K. Untersuchungen zur relativen Bedeutung einzelner populationsdynamischer Parameter fur die Entwicklung der Verunkrautung. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz 96, 346-352 (1989).
Plantenavne
| Dansk navn | Latinsk navn | |
| Agersennep | Sinapis arvensis L. | |
| Alm. hvene | Agrostis tenuis L. | |
| Burresnerre | Galium aparine L. | |
| Alm. kvik | Elymus repens (L.) Gould | |
| Fuglegræs | Stellaria media (L.) Vill. | |
| Fåresvingel | Festuca ovina L. | |
| Hanespore | Echinochloa crus-galli (L.) Beauv. | |
| Hvidmelet gåsefod | Chenopodium album L. | |
| Lugtløs kamille | Tripleurospermum inodorum (L.) Schultz Bip. | |
| Liden nælde | Urtica urens L. | |
| Majs | Zea mays L. | |
| Mælkebøtte | Taraxacum vulgare Weber | |
| Raps | Brassica napus L. | |
| Sort natskygge | Solanum nigrum L. | |
| Stor nælde | Urtica dioecá L. | |
| Strandbede | Beta vulgaris ssp. maritima (L.) Arcang. | |
| Strandsvingel | Festuca arundinacea Schreb. | |
| Vinterraps | Brassica napus L. | |
| Vårbyg | Hordeum vulgare L. | |
Herbicider
Glyphosat
- Aktivstoffet glyphosat findes i mange formuleringer. I denne rapport benævnes herbicidet ”glyphosat” frem for at fremhæve et af de mange produkter indeholdende glyphosat.
- Eksempelvis indeholder en liter af herbicidet Roundup Bio 360 g glyphosat som isopropylamin salt.
- En dosis Roundup Bio på 3,5 L/ha giver behandlingsindeks BI=1 til de anvendelser, hvor det er godkendt.
MaisTer
- Aktivstofferne i herbicidet MaisTer findes kun i dette produkt. Derfor benævnes herbicidet i denne rapport ”MaisTer”.
- Et kg af herbicidet MaisTer indeholder 300 g foramsulfuron + 10 g iodosulfuron, altså i alt 310 g aktivstof (as.) pr. kg produkt.
- En dosis MaisTer på 150 g/ha giver behandlingsindeks, BI=1.
- MaisTer anvendes altid sammen med additivet MaisOil, som tilsættes i forholdet 1,33 L MaisOil pr. 100 g MaisTer.
Kilde: www.middeldatabasen.dk.
Artsliste for successionsforsøg
| Udsået planteart | Frøbank | |
| Tokimbladede | ||
| filtbladet kongelys (Verbascum thapsus) | 1892 | |
| rejnfan (Tanacetum vulgare) | 1463 | |
| alm. stedmoderblomst (Viola tricolor) | 1134 | |
| toårig natlys (Oenothera biennis) | 296 | |
| mælkebøtte (Taraxacum sp.) | 111 | |
| prikbladet perikon (Hypericum perforatum) | 89 | |
| dunbirk (Betula pubescens) | 77 | |
| alm. torskemund (Linaria vulgaris) | 74 | |
| mark forglemmigej (Myosotis arvensis) | 58 | |
| alm. gåsemad (Arabidopsis thaliana) | 42 | |
| hvid okseøje (Leucanthemum vulgare) | 42 | |
| alm. rødknæ (Rumex acetosella) | 36 | |
| stor nælde (Urtica dioca) | 31 | |
| alm. spergel (Spergula arvensis) | 23 | |
| mark ærenpris (Veronica arvensis) | 22 | |
| liden klokke (Campanula rotundifolia) | 20 | |
| rank evighedsblomst (Gnaphalium sylvaticum) | 10 | |
| alm. kongepen (Hypochoeris radicata) | 9 | |
| ager tidsel (Cirsium arvense) | 6 | |
| hvidmelet gåsefod (Chenopodium album) | 4 | |
| kølle valmue (Papaver argemone) | 4 | |
| dueurt (Epilobium sp.) | 4 | |
| alm. gederams (Epilobium angustifolium) | 3 | |
| mark hindeknæ (Spergularia rubra) | 2 | |
| Salomons lysestage (Lepidium campestre) | 2 | |
| mørk kongelys (V. nigrum) | 2 | |
| snerlepileurt (Fallopia convolvulus) | 1 | |
| haremad (Lapsana communis) | 1 | |
| flipkrave (Teesdalia nudicaulis) | 1 | |
| gul reseda (Reseda lutea) | 1 | |
| alm. markarve (Arenaria serpullifolia) | 1 | |
| gråbynke (Artemisia vulgaris) | 1 | |
| håret høgeurt (Hierachium pilosella) | 1 | |
| Enkimbladede | ||
| stortoppet hvene (Agrostis gigantea) | 2192 | |
| fåre svingel (Festuca ovina) | 128 | |
| Agrostis hybrid | 100 | |
| eng rapgræs (Poa pratensis) | 18 | |
| alm. hvene (A. capillaris) | 2 | |
| tudse siv (Juncus bufonius) | 2 | |
| håret frytle (Luzula pilosa) | 1 |
Måling af væksthæmning
I mange forsøgssammenhænge er det interessant at bestemme varigheden af en væksthæmning samt at kunne forudsige en sluteffekt ved ikke-destruktive målinger. Oftest anvender man i disse sammenhænge visuelle vurderinger, hvor der gives en karakter for skade. Andre muligheder kunne være at måle plantens fotosyntese eller vandforbrug i en periode efter sprøjtning. Begge disse parametre er tæt relateret til vækst. Bladtilvækst er endnu en mulighed, og endelig arbejdes der i et andet projekt med udvikling af et test-kit, som kan forudsige effekten af en sprøjtning 1 uge efter dens udførelse (HWR test-kit140). I nærværende projekt sammenlignede vi anvendeligheden af disse forskellige metoder til vurdering af sluteffekt af glyphosat og MaisTer over for frø- og rodfremspirede planter af mælkebøtte og stor nælde.
Effekt af glyphosat
Effekten af de anvendte doseringer af glyphosat på frøfremspiret mælkebøtte kunne beskrives med S-formede doseringskurver (figur 1). Med undtagelse af to doseringer var den målte biomasse 1, 2 og 3 uger efter sprøjtning højere end slutbiomassen, og der var derfor ikke tale om midlertidig væksthæmning men om en øget effekt med tid efter sprøjtning. Beregnes ED50-doser ud fra data for vandforbrug og bladtilvækst henholdsvis 1, 2 og 3 uger efter sprøjtning fås generelt ED-doseringer, som er noget lavere end ved beregning udfra sluteffekt på biomasse (tabel 1). ED50-doseringen beregnet på bladtilvækst i den første uge efter sprøjtning er den værdi, som kommer tættest på ED50-doseringen beregnet udfra sluteffekt på biomasse, og disse er ikke signifikant forskellige. De bedste korrelationer til sluteffekt blev fundet for bladtilvækst 3 uger efter sprøjtning og visual score 3 uger efter sprøjtning (r²= 0,91 og 0,93).
På rodfremspiret mælkebøtte blev der ikke opnået nogen effekt på biomassen målt 24 uger efter behandling (tabel 1 og figur 1). De fleste doseringer stimulerede effekten (hormesis) – en reaktion som ofte ses i forbindelse med subletale doseringer141. På flerårige arter er det veldokumenteret, at stresspåvirkninger medfører en translokation af reserver fra forrådsorganer til nye skud. Måling af biomasse 1, 2 og 3 uger efter sprøjtning viste op til 60% effekt, og der var således tale om en midlertidig væksthæmning, som planten over et længere tidsrum var i stand til at kompensere for. De målte effekter på bladtilvækst og vandforbrug var moderate, men specielt vandforbrug og visuel score viste dog, at der var tale om en hæmning af plantens vækst i en periode. Ingen af værdierne korrelerede godt med slutbiomassen.
På frøfremspiret nælde var der ikke tale om midlertidige væksthæmninger, men om permanente effekter (tabel 2 og figur 2). ED50-doseringerne for tørvægt var højere end ED50-doseringerne beregnet ud fra vandforbrug (undtagen 1 uge efter sprøjtning) og bladtilvækst. På grund af de usikre estimater er der dog ikke tale om signifikante forskelle. På grund af for lav effekt var det ikke muligt at estimere doseringskurver for glyphosat på rodfremspiret stor nælde.
Effekt af MaisTer
Med MaisTer blev der på frøfremspiret mælkebøtte opnået en midlertidig væksthæmning i de to laveste doseringer, hvor den relative tørvægt 2 og 3 uger efter sprøjtning var reduceret, men sluteffekten 11 uger efter sprøjtning var stimuleret eller upåvirket af behandlingen (tabel 3 og figur 3). For de øvrige doseringer blev der opnået effekter fra 0 til 100%. Ved sammenligning af ED50-doseringerne beregnet ud fra vandforbrug og bladtilvækst 1, 2 og 3 uger efter behandling ses, at de parametre, som bedst beskriver sluteffekten, er vandforbrug 1 uge efter sprøjtning samt bladtilvækst 1 og 3 uger efter sprøjtning. De målte parametre var dårligere korreleret til sluteffekt end for behandlingerne med glyphosat, hvilket primært skyldes vækststimuleringen i laveste dosering. De bedste korrelationer til sluteffekten blev fundet med visuel score 1 uge efter sprøjtning og vandforbrug 1 uge efter sprøjtning (r²=0,81 og 0,79).
Effekten af MaisTer på rodfremspiret mælkebøtte var signifikant lavere end for frøfremspirede planter, men da doseringerne blev øget på de rodfremspirede planter, var sluteffekten høj. Effekten kunne derfor beskrives med S-formede doseringskurver, mens de destruktive målinger 1, 2 og 3 uger efter sprøjtning kun viste et svag respons på doseringerne. Som følge heraf er estimaterne usikre og konfidensintervallerne meget store, og der er ikke signifikant forskel på ED50-doseringerne. Korrelation mellem visuelle bedømmelser og sluteffekt var dårlige men stigende over tid, idet effekten af MaisTer først indtræder lang tid efter behandling. De bedste korrelationer blev fundet til bladtilvækst (r²= 0,95), som hæmmedes i samme takt som slutbiomassen.
På stor nælde var det kun muligt at estimere doseringskurver for tørvægt på de rodfremspirede planter (tabel 4 og figur 4). For de målte parametres vedkommende var der for lidt effekt til at kunne bestemme doseringskurverne.
Nye metoder
Resultater af biomarkørtesten (HWR testkit) indikerede, at man specielt med den ene stick-metode (A) fik tydelige farvegrupperinger i relation til den opnåede effekt på plantebiomassen (figur 5). På de rodfremspirede mælkebøtter var sluteffekten meget lav, og der var intet farverespons på sticksene (figur 6). Kittet er p.t. kun valideret for to herbicider og tre græsukrudtsarter, men der synes at være perspektiver i en målrettet videreudvikling af testkittet til vurdering af herbicideffekter på tokimbladede ukrudtsarter.
I forsøget fulgtes fotosynteseaktiviteten på et enkelt blad for hver plante (figur 7 og 8). Glyphosat forårsagede en kortvarig hæmning af fotosyntesen på de ramte blade, mens MaisTer havde en længerevarende negativ effekt. Begge herbicider bevirkede dog, at bladene på længere sigt var mere aktive end ubehandlede blade. Metoden kunne altså ikke anvendes som indikation for hele plantens vækst.
Konklusion
Det må konkluderes, at det er vanskeligt at forudsige effekten udfra ikke-destruktive målinger. Sammenhængen mellem visuelle bedømmelser af planternes udseende de første uger efter sprøjtning giver for frøfremspirede planter ligeså gode eller bedre forudsigelser af sluteffekt som måling af bladtilvækst og vandforbrug. For rodfremspirede planter er det derimod vanskeligt at vurdere sluteffekten udfra visuelle bedømmelser, idet der sker en kompensation af væksthæmning ved anvendelse af reserverne fra rodsystemet. For MaisTers vedkommende kan bladtilvæksten anvendes som et mål for sluteffekten. Det er ikke muligt at forudsige, om dette også vil være tilfældet for Roundup, hvis doseringerne havde været højere, og dermed givet højere effekt.
Tabel 1. Effekt af glyphosat på mælkebøtte vist som ED50 dosering (95% konfidensinterval i parentes) ved anvendelse af forskellige måleparametre på forskellige tidspunkter efter behandling. For mange parametre var det ikke muligt at estimere doseringskurven (i.e.) og derfor ikke muligt at bestemme ED50 doseringen.
Mælkebøtte, |
Frøfremspiret | Rodfremspiret | ||||||||||
| glyphosat-behandlet | b | ED50 (g as./ha) | b | ED50 (g as./ha) | ||||||||
| Tørvægt (g) | ||||||||||||
| efter 3 uger | –3,7 | (–5,7...–1,8) | 129,8 | (87,0...172,5) | i.e. | i.e. | ||||||
| Vandforbrug (ml/dag) | ||||||||||||
| efter 1 uge | –0,7 | (–1,0...–0,4) | 21,6 | (-5,6...48,8) | i.e. | i.e. | ||||||
| efter 2 uger | –1,0 | (–1,5...–0,6) | 27,6 | (3,0...52,2) | i.e. | i.e. | ||||||
| efter 3 uger | –1,1 | (–1,5...–0,8) | 25,6 | (7,6...43,5) | i.e. | i.e. | ||||||
| Bladtilvækst (cm/uge) | ||||||||||||
| efter 1 uge | –5,1 | (–9,9...–0,2) | 63,9 | (40,8...87,1) | i.e. | i.e. | ||||||
| efter 2 uger | –2,0 | (–4,2...–0,2) | 46,6 | (5,2...72,9) | i.e. | i.e. | ||||||
| efter 3 uger | –1,8 | (–3,8...–0,1) | 39,0 | (9,4...83,9) | i.e. | i.e. | ||||||
Tabel 2. Effekt af glyphosat på stor nælde vist som ED50 dosering (95% konfidensinterval i parentes) ved anvendelse af forskellige måleparametre på forskellige tidspunkter efter behandling. For mange parametre var det ikke muligt at estimere doseringskurven (i.e.) og derfor ikke muligt at bestemme ED50 doseringen.
Stor nælde, |
Frøfremspiret | Rodfremspiret | ||||||||||
| glyphosat-behandlet | b | ED50 (g as./ha) | b | ED50 (g as./ha) | ||||||||
| Tørvægt (g) | ||||||||||||
| efter 3 uger | –1,8 | (–3,1...–0,5) | 207,1 | (22,3...391,9) | i.e. | >710 | ||||||
| Vandforbrug (ml/dag) | ||||||||||||
| efter 1 uge | –1,1 | (–2,2...–0,1) | 552,2 | (79,6...1025) | i.e. | i.e. | ||||||
| efter 2 uger | –1,5 | (–2,3...–0,7) | 144,1 | (20,4...268,1) | i.e. | i.e. | ||||||
| efter 3 uger | –1,7 | (–2,7...–0,7) | 141,6 | (17,2...266,2) | i.e. | i.e. | ||||||
| Bladtilvækst (cm/uge) | ||||||||||||
| efter 1 uge | –1,9 | (–3,6...–0,2) | 141,9 | (7,3...276,6) | i.e. | i.e. | ||||||
| efter 2 uger | –3,1 | (–7,6...–1,4) | 150,2 | (0,2...300,4) | i.e. | i.e. | ||||||
| efter 3 uger | –3,1 | (–6,1...–0,02) | 125,6 | (24,0...227,3) | i.e. | i.e. | ||||||
Tabel 3. Effekt af MaisTer på mælkebøtte vist som ED50 dosering (95% konfidensinterval i parentes) ved anvendelse af forskellige måleparametre på forskellige tidspunkter efter behandling. For mange parametre var det ikke muligt at estimere doseringskurven (i.e.) og derfor ikke muligt at bestemme ED50 doseringen.
Mælkebøtte, |
Frøfremspiret | Rodfremspiret | ||||||||||
| MaisTer-behandlet | b | ED50 (g as./ha) | b | ED50 (g as./ha) | ||||||||
| Tørvægt (g) | ||||||||||||
| efter 3 uger | –4,9 | (–8,8...–1,1) | 3,3 | (2,1...4,4) | –4,3 | (–6,2... –2,5) |
9,9 | (7,9... 11.8) |
||||
| Vandforbrug (ml/dag) | ||||||||||||
| efter 1 uge | –5,8 | (–10,0...–1,7) | 2,3 | (1,4...3,3) | –1,0 | (–2,0.. .–0,01) |
19,1 | (3,2... 34,9) |
||||
| efter 2 uger | –4,0 | (–6,5...–1,6) | 1,6 | (0,9...2,4) | –1,2 | (–2,2... –0,2) |
16,8 | (7,2... 26,5) |
||||
| efter 3 uger | –4,1 | (–6,3...–1,9) | 1,4 | (0,9...2,0) | –1,7 | –2,9... –0,5) |
12,4 | (7,6... 17,1) |
||||
| Bladtilvækst (cm/uge) | ||||||||||||
| efter 1 uge | –1,5 | (–2,7...0,3) | 2,1 | (0,2...3,9) | –8,o | (–99,0... –7,9) |
14,6 | (3,9... 25,2) |
||||
| efter 2 uger | –1,4 | (–2,3...–0,4) | 1,8 | (0,3...3,2) | –4,8 | (–18,9.. .–9,2) |
15,2 | (10,3... 20,1) |
||||
| efter 3 uger | –1,8 | (–2,7...–0,9) | 2,3 | (1,1...3,4) | –1,7 | (–6,4... –3,0) |
10,9 | (–9,4... 31,2) |
||||
Tabel 4. Effekt af MaisTer på stor nælde vist som ED50 dosering (95% konfidensinterval i parentes) ved anvendelse af forskellige måleparametre på forskellige tidspunkter efter behandling. For mange parametre var det ikke muligt at estimere doseringskurven (i.e.) og derfor ikke muligt at bestemme ED50 doseringen.
Stor nælde, |
Frøfremspiret | Rodfremspiret | ||||||||||
| MaisTer-behandlet | b | ED50 (g as./ha) | b | ED50 (g as./ha) | ||||||||
| Tørvægt (g) | ||||||||||||
| efter 3 uger | –3,2 | (–6,2...–0,4) | 7,5 | (4,1...11,0) | –0,7 | (–1,8...–0,5) | 6,0 | (–9,3...21,2) | ||||
| Vandforbrug (ml/dag) | ||||||||||||
| efter 1 uge | i.e. | i.e. | i.e. | i.e. | ||||||||
| efter 2 uger | i.e. | i.e. | i.e. | i.e. | ||||||||
| efter 3 uger | i.e. | i.e. | i.e. | i.e. | ||||||||
| Bladtilvækst (cm/uge) | ||||||||||||
| efter 1 uge | i.e. | i.e. | i.e. | i.e. | ||||||||
| efter 2 uger | i.e. | i.e. | i.e. | i.e. | ||||||||
| efter 3 uger | i.e. | i.e. | i.e. | i.e. | ||||||||
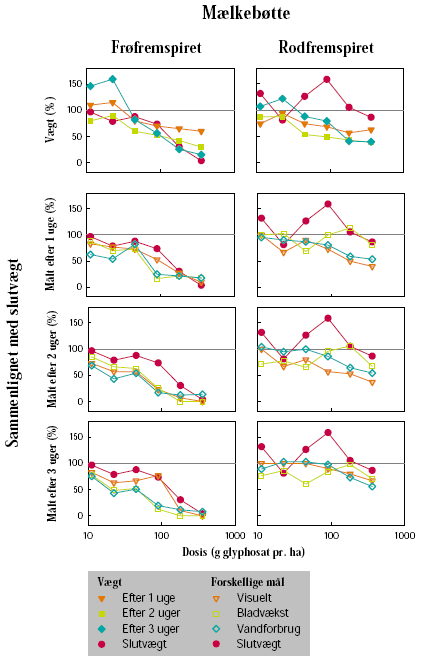
Figur 1. Mælkebøttes vækst efter behandling med glyphosat.
Responskurver vises som % i forhold til responskurven for ubehandlede planter. Responskurver for tre ikke-destruktive målemetoder sammenlignes med slutvægten, som måles destruktivt. Slutvægten bestemtes henholdsvis 11 og 24 uger efter fremspiring for frøfremspirede og rodfremspirede planter.
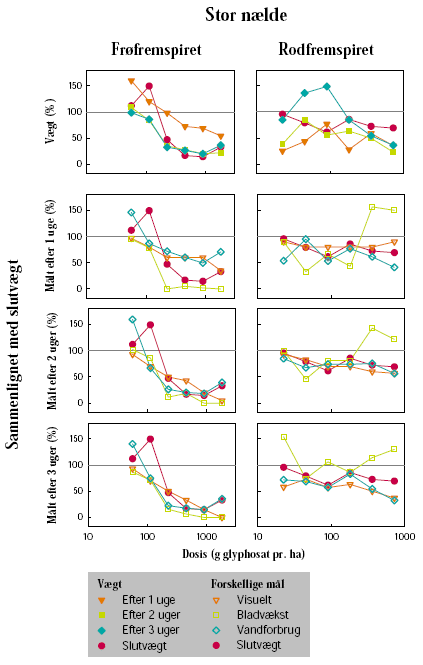
Figur 2. Stor nældes vækst efter behandling med glyphosat.
Responskurver vises som % i forhold til responskurven for ubehandlede planter. Responskurver for tre ikke-destruktive målemetoder sammenlignes med slutvægten, som måles destruktivt. Slutvægten bestemtes henholdsvis 13 og 20 uger efter fremspiring for frøfremspirede og rodfremspirede planter.
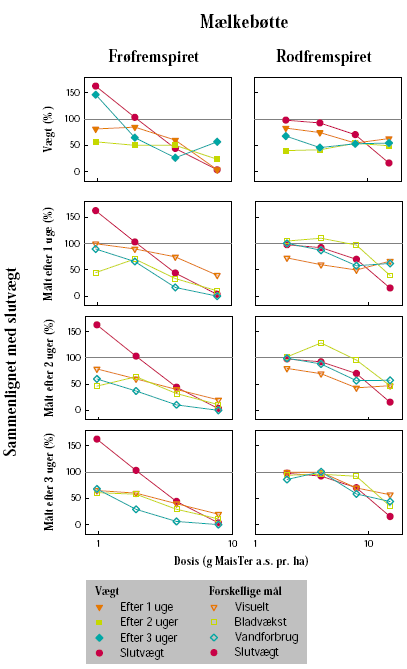
Figur 3. Mælkebøttes vækst efter behandling med MaisTer.
Responskurver vises som % i forhold til responskurven for ubehandlede planter. Responskurver for tre ikke-destruktive målemetoder sammenlignes med slutvægten, som måles destruktivt. Slutvægten bestemtes henholdsvis 11 og 24 uger efter fremspiring for frøfremspirede og rodfremspirede planter.
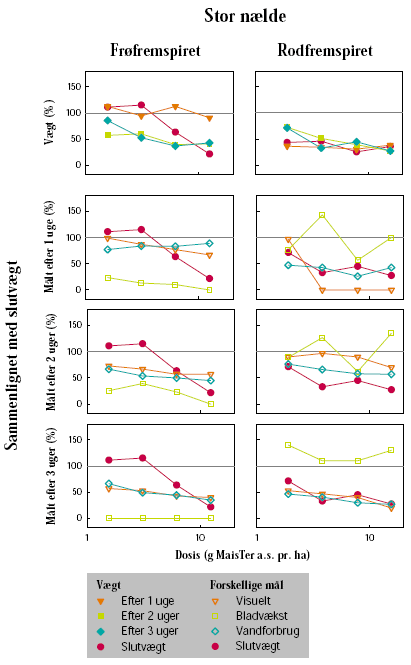
Figur 4. Stor nældes vækst efter behandling med MaisTer.
Responskurver vises som % i forhold til responskurven for ubehandlede planter. Responskurver for tre ikke-destruktive målemetoder sammenlignes med slutvægten, som måles destruktivt. Slutvægten bestemtes henholdsvis 13 og 20 uger efter fremspiring for frøfremspirede og rodfremspirede planter.
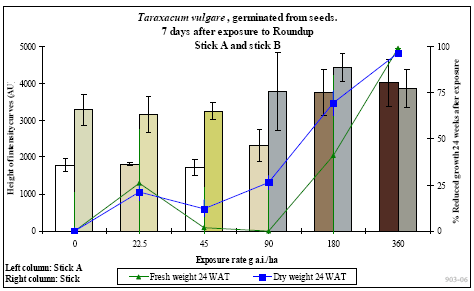
Figur 5. Resultater af HWR testkit på frøfremspiret mælkebøtte behandlet med glyphosat (360 g/ha). Procentuel reduktion i friskvægt (grøn kurve) og tørvægt (blå kurve) 24 uger efter behandling er vist sammen med farvereaktionen på stick A (venstre søjler) og stick B (højre søjler).
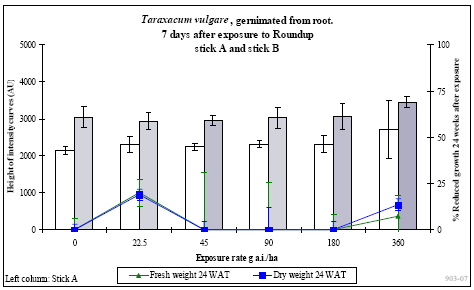
Figur 6. Resultater af HWR testkit på rodfremspiret mælkebøtte behandlet med glyphosat (360 g/ha). Procentuel reduktion i friskvægt (grøn kurve) og tørvægt (blå kurve) 24 uger efter behandling er vist sammen med farvereaktionen på stick A (venstre søjler) og stick B (højre søjler).
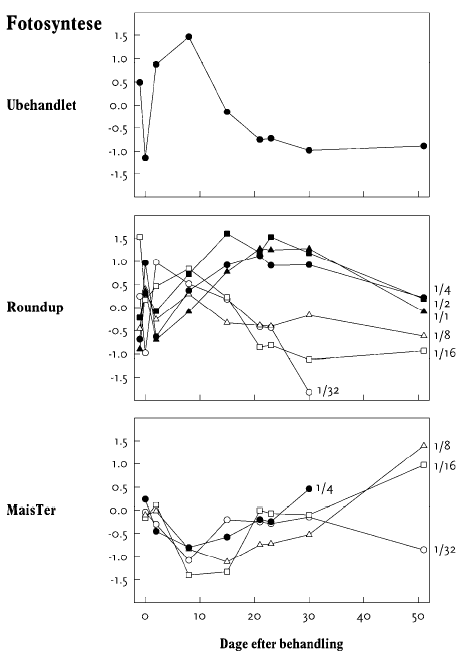
Figur 7. Fotosyntese i pottesprøjteforsøg med rodfremspiret mælkebøtte.
Fotosyntesen blev målt gentagne gange på samme blad. Den er angivet som et indeks, som inden for hver dato er standardiseret, så et gennemsnitsblad giver værdien 0. Brøker angiver andelen af en dosis på henholdsvis 1260 g/ha glyphosat og 46,5 g as./ha MaisTer. Kurver er afbrudte, hvis alle 3 blade (på hver sin plante) døde.
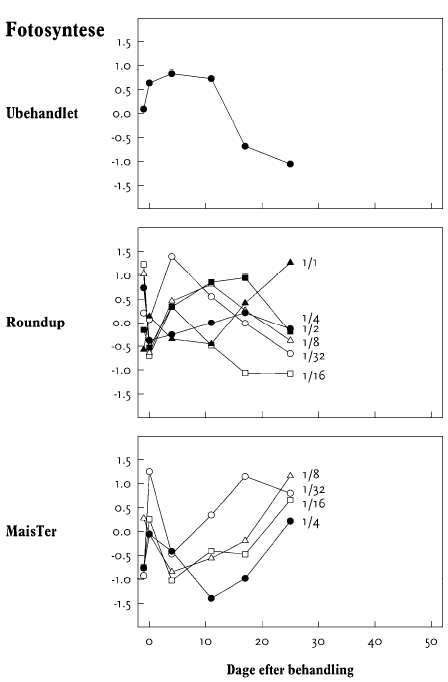
Figur 8. Fotosyntese i pottesprøjteforsøg med rodfremspiret stor nælde.
Fotosyntesen blev målt gentagne gange på samme blad. Den er angivet som et indeks, som inden for hver dato er standardiseret, således at et gennemsnitsblad giver værdien 0. Brøker angiver andelen af en dosis på henholdsvis 1260 g/ha glyphosat og 46,5 g as./ha MaisTer. Målingerne blev afbrudt, da mange blade pludseligt døde.
Version 1.0 November 2008 • © Miljøstyrelsen.