Effekter af cypermethrin, azoxystrobin og bentazon på limniske invertebrater2 Metoder2.1 Pesticidvalg2.1.1 Cypermethrin 2.1.2 Azoxystrobin 2.1.3 Bentazon 2.1.4 Stofkoncentrationer 2.2 Valg af forsøgsdyr 2.2.1 Cladoceer: Daphnia magna, Daphnia galeata, Diaphanosoma brachyurum og Chydorus sphaericus 2.2.2 Vandlopper: Cyclops vicinus og Eudiaptomus graciloides 2.2.3 Makroinvertebrater: Chaoborus, Chironomus, Cloeon og Hydropsyche 2.3 Forsøgsbetingelser 2.3.1 Dyrkning af algekultur 2.3.2 Zooplankton dyrkningsmedium 2.3.3 Kultivering af forsøgsdyr 2.3.4 Indsamling af forsøgsdyr i felten 2.4 Koncentration-respons forsøg 2.4.1 Formål og forsøgsprincip 2.4.2 Forsøgsdyr 2.4.3 Forsøgsopstilling 2.4.4 Databehandling 2.5 Reproduktionsforsøg 2.5.1 Formål og forsøgsprincip 2.5.2 Forsøgsdyr 2.5.3 Forsøgsopstilling 2.5.4 Databehandling 2.6 Videoregistrering af fysiologiske parametre 2.6.1 Formål og forsøgsprincip 2.6.2 Forsøgsdyr 2.6.3 Forsøgsopstilling 2.6.4 Databehandling 2.7 Svømmeadfærd 2.7.1 Formål 2.7.2 Forsøgsdyr 2.7.3 Forsøgsopstilling 2.7.4 Databehandling 2.1 PesticidvalgI samråd med Miljøstyrelsen blev det fastlagt at anvende hhv. et insekticid, et fungicid og et herbicid i undersøgelserne. Valget af pesticider blev foretaget ud fra Miljøstyrelsens pesticidliste til overfladevandsprojekter og gjort på baggrund af pesticidernes anvendte mængde i Danmark, deres biologiske aktivitet og forekomst i vandmiljøet og den nuværende viden om stoffernes effekt i vandmiljøet. Følgende 3 pesticider, som alle anvendes i dansk landbrug, blev udvalgt: cypermethrin, azoxystrobin og bentazon. 2.1.1 CypermethrinCypermethrin er et pyrethroid-insekticid.
I en ”worst case” situation (sprøjtebom passerer hen over et 0,3 m dybt vandhul) vil der ved anvendelse af normal markdosis (Friis et al., 2002) skabes en cypermethrin koncentration på 4,1 – 6,6 µg l-l, afhængig af afgrødetype (korn og majs). 2.1.2 AzoxystrobinAzoxystrobin er et strobilurin-fungicid.
I en ”worst case” situation (sprøjtebom passerer hen over et 0,3 m dybt vandhul) vil der ved anvendelse af normal markdosis (Friis et al., 2002) skabes en azoxystrobin koncentration på 33 – 83 µg l-l, afhængig af afgrødetype. 2.1.3 BentazonBentazon er et ”triazin”-herbicid.
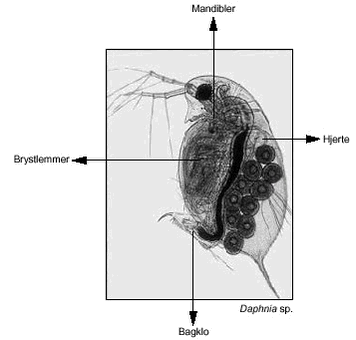
I en ”worst case” situation (sprøjtebom passerer hen over et 0,3 m dybt vandhul) vil der ved anvendelse af normal markdosis (Friis et al., 2002) skabes en bentazon koncentration på 0,16 – 0,48 mg l-l, afhængig af afgrødetype. 2.1.4 StofkoncentrationerDer blev i hele projektet arbejdet med nominelle koncentrationer, som alle er beskrevet i tabel 3 og tabel 4. Til kontrol af de nominelle koncentrationer blev der i de to forsøgsår udtaget hhv. 4 og 6 prøver til analysering. Prøverne blev udtaget i udglødede glasflasker umiddelbart før forsøgsstart. Hvis ikke prøverne blev sendt til analysering umiddelbart efter udtagelsen, blev de nedfrosset indtil analysering. Under transport blev de opbevaret ved < 4 oC. Prøverne blev analyseret hos Miljøkemi a/s (nu EUROFINS) ved anvendelse af gas- og væskekromatografi koblet til massespektrometri (GC-MS og LC-MS). 2.2 Valg af forsøgsdyr2.2.1 Cladoceer: Daphnia magna, Daphnia galeata, Diaphanosoma brachyurum og Chydorus sphaericusDaphnia magna og D. galeata består af et tveklappet kropsskjold, der omslutter krop og kropfødder, men ikke hovedet (figur 2). På hovedet sidder et stort sammensat øje. Foruden dette øje findes som regel det lille uparrede pandeøje. Dafnierne har to par antenner. Det forreste par er normalt korte og besat med sansehår. Det andet par antenner er kraftige svømmeredskaber. Kroppen er forsynet med 4-6 par kropslemmer, som er i stadig bevægelse og bidrager til respiration og fødeoptagelse ved at frembringe en vandstrøm ned mellem de to kropsskjolde. Halen er bøjet om mod bugsiden, og på halen sidder en bagklo. Hjertet sidder dorsalt over rugehulen (Røen, 1995). Daphnia magna tilhører underslægten Ctenodaphnia og bliver op til 6 mm lang; hannen bliver maksimalt 3 mm. På grund af dens størrelse og bevægelsesmønster er den et eftertragtet fødeemne for fisk. Dens levesteder er derfor vandhuller og småsøer med ingen eller meget lille fiskebestand (Røen, 1995). D. galeata tilhører underslægten Daphnia og bliver maksimalt 2,5 mm lang; hannen maksimalt 1,5 mm. Den ringere størrelse betyder, at den er mindre eftertragtet som føde for fisk. Derfor kan den forekomme i stort set alle danske søtyper, men vil, sammen med de øvrige dafniearter, blive kraftigt reduceret eller elimineret i eutrofe søer med stor fisketæthed (Jeppesen et al., 2000). Diaphanosoma brachyurum tilhører slægten Diaphanosoma. I Danmark findes kun én art. Den er karakteriseret ved et langt og smalt hoved uden rostrum. Hovedet er tydeligt afgrænset fra kroppen med en indskæring. 1. benpar hos hannen er forsynet med gribehage. Arten er karakteristisk ved at have 3 dorsalt siddende grove torne på furcakløerne (Røen, 1995). Hunnen bliver 0,75 – 1,2 mm og hannen ca. 0,75 mm. Pga. størrelsen er den mindre udsat for fiskepredation end de større Daphnia-arter.Chydorus sphaericus tilhører underfamilien Chydorinae. Chydorus består af et hoved- og et kropsskjold, som til sammen dækker hele kroppen. Der er ingen tydelig adskillelse mellem de to skjolde. Arterne er afrundede, kugleformede til aflange. Forrest på hovedskjoldet sidder et kort og spidst rostrum (”snabel”). Fremme på hovedskjoldet sidder et hoved- og biporearrangement. C. sphaericus har to antennepar ligesom Daphnia, 1. par er meget korte, 2. par lidt længere og bruges som svømmeredskaber. Hunnen bliver maksimalt 0,5 mm og hannen 0,4 mm lang. Hos dafnierne forekommer heterogoni. Størstedelen af individerne er hunner, som forplanter sig parthenogenetisk. De ubefrugtede æg gennemgår en hurtig udvikling i rugehulen under kropsskjoldet. Ungerne, der forlader rugehulen, ligner de voksne dafnier. Normalt følger et antal parthenogenetiske generationer efter hinanden. I stress-situationer (fx. ved lave temperatur eller dårlige fødeforhold) udvikles der hanner fra de ubefrugtede æg. Samtidig danner hunnerne mørkfarvede hvileæg, som befrugtes. Efter en hvileperiode klækkes disse og danner hunner, der igen kan formere sig parthenogenetisk. D. magna og D. galeata er fritsvømmende arter, der findes i de åbne vandmasser. Ved tilstedeværelse af prædatorer udviser de i dagtimerne flugtadfærd ved i dybe søer at flygte ud af epilimnion (overfladevandet i temperatur-lagdelte søer) mod metalimnion (springlaget i temperatur-lagdelte søer), hvor prædationsrisikoen er mindre (von Elert & Loose, 1996; Dodson et al., 1997). I lavvandede søer viser dafnier en alternativ flugtadfærd ved at flygte horisontalt mod vegetationsdækkede områder (se fx. review af Burks et al., 2002). Er der ikke prædatorer til stede, vil både D. magna og D. galeata svømme omkring i de åbne vandmasser døgnet rundt. Dafniernes føde består af fytoplankton, flagellater, bakterier og detritus. De har således flere fødekilder. Det betyder, at de ikke kun er afhængige af, hvad der findes i det åbne vand, men også kan finde føde på plante- eller sedimentoverflader. Hvad angår fytoplankton, er der størrelsesbegrænsninger på, hvad dafnierne kan håndtere, typisk algearter med GALD-værdier (største længde) på op til 40 µm (fx. Kasprzak & Lathrop, 1997). Diaphanosoma brachyurum er en planktonisk sommerform, dvs. den findes fritsvømmende i det åbne vand, men kan også forekommer i littoralen. I modsætning til Daphnia svømmer den med ”hoppende” bevægelser. Den findes i både større og mindre søer (Røen, 1995). C. sphaericus findes i alle søtyper og lever normalt i bredzonen mellem vegetationen, men kan i klarvandede søer også forekomme planktonisk. Den lever af fytoplankton og kan i søer med blågrønalger fasthæfte sig på større kolonier (Røen, 1995). C. sphaericus findes overalt i Danmark og er vores almindeligste dafnieart. 2.2.2 Vandlopper: Cyclops vicinus og Eudiaptomus graciloidesCopepoderne mangler et egentligt skjold (figur 2), men hele kroppen er omgivet af et kitinskelet. Hovedet er forsynet med et stort pandeøje, sideøjne mangler. De har to par antenner, hvoraf det første par er de længste. Hos de calanoide copepoder (E. graciloides) er første antennepar lige så lange som dyrets forkrop og ca. dobbelt så lange som hos de cyclopoide copepoder (C. vicinus). Forkroppen består af fem frie kropsled, som hvert bærer et benpar. Herefter følger fem lemmeløse led, som udgør bagkroppen. Herpå sidder et par uleddede halenokker (furca). Æggene bæres af hunnen i én (E. graciloides) eller to (C. vicinus) ægsække, som er fastgjort ved grunden af bagkroppen. E. Graciloides hunnen bliver 1 – 1,5 mm og C. vicinus hunnen 1,5 - 2,3 mm. Hannerne er noget mindre. Copepoderne forekommer i alle typer af danske søer og vandhuller. Calanoide og cyclopoide copepoder vil også ofte forekomme sammen, men typisk vil en af typerne dominere over den anden. I eutrofe søer er det normalt cyclopoide copepoder, der dominerer. Dette er fx. kendt fra Søbygaard Sø, hvor C. vicinus i lange perioder fuldstændigt dominerer zooplanktonsamfundet (Hansen & Jeppesen, 1992). I mesotrofe og oligotrofe søer vil de calanoide copepoder normalt dominere copepodsamfundet. I dybe, lagdelte søer forekommer den calanoide type næsten altid uanset søens trofigrad. Reproduktion hos copepoderne sker ved kønnet formering. Hannen fanger hunnen med den ene antenne og afsætter en spermatofor på hunnens genital-segment. Hunnen kan herefter blive befrugtet med spermatozoer fra denne spermatofor. De befrugtede æg opbevares i ægsække, som bæres på hunnens bagkrop (se ovenfor). Æggene klækkes, og nauplier forekommer nu fritsvømmende i søen. Nauplierne gennemgår 6 forskellige stadier, hvorefter der følger 5 copepoditstadier med hudskifte mellem hvert stadie. Hvilket copepoditstadie, dyret er på, afgøres af antal svømmeben og antal segmenter på bagkroppen. Herefter når de voksenstadiet. Undervejs i udviklingen går copepoderne i diapause (hvilestadie), hvor de i en periode, typisk ved ugunstige forhold, ligger inaktive i sedimentet. Dette sker typisk ved copepoditstadie 4 eller 5. Copepodernes livscyklus varierer mellem få uger og tre år. I danske søer forekommer normalt 3-6 generationer pr år.
Figur 2 E. graciloides og C. vicinus er begge fritsvømmende arter, hvis adfærd er afhængig af dels deres føde og dels af prædatorer. Ved tilstedeværelse af prædatorer flygter de tilsvarende dafnierne, hvilket vil sige, at de i dagtimerne søger mod bunden, hvor prædationsrisikoen er mindre, mens de om natten søger mod optimale fødebetingelser. Vigtige prædatorer er planktivore fisk (skaller og brasen), men også Chaoborus kan være en betydelig prædator og kan inducere migration. Copepodernes føde varierer med arten. De cyclopoide copepoder er ofte omnivorer. Eksempelvis er C. vicinus en meget effektiv prædator, som både tager hjuldyr, nauplier, copepoditer og små dafnier (Einsle, 1996). Derudover kan den også tage flagellater og alger, hvilke dog er en mindre vigtig fødekilde. På nauplie-stadiet er algerne en vigtig fødekilde. 2.2.3 Makroinvertebrater: Chaoborus, Chironomus, Cloeon og HydropsycheChaoborus flavicans tilhører en lille familie af Chaoboridae (glasmyg) i insektklassen, hvor larverne er rovdyr og lever i stillestående vand (figur 2). Larverne klækkes typisk fra æg i løbet af sommeren/efteråret og gennemfører en række larvestadier (I-IV). Larverne er 10-20 mm lange og ernærer sig primært af små zooplankton (fx. Bosmina og mindre Daphnia-arter) fra vandfasen. Chaoborus-larverne kan være særdeles hyppige i eutrofe søer og damme, hvor de opholder sig i det dybeste vand og gerne i sedimentoverfladen i dagtimerne for at undgå prædation fra planktivore fisk og stiger op i de åbne vandmasser i nattetimerne for at jage zooplankton. Larverne har to luftfyldte sække i kroppen og kan ved hjælp af disse foretage de horisontale vandringer i vandsøjlen. Larverne må betragtes som ganske hårdføre, idet de kan tolerere meget lave iltkoncentrationer gennem længere tid. Interaktionerne mellem zooplankton og Chaoborus er bl.a. beskrevet i Christoffersen (1990), og en generel beskrivelse af Chaoboridae findes i Dall & Lindegaard (1995). Glasmyggelarver i 3. og 4. stadie kan ofte købes i akvarieforretninger, og alle stadier kan i vækstsæsonen indsamles i søer og damme med planktonet om natten eller med bundprøvetagere om dagen. Det er normalt ikke vanskeligt at fremskaffe materiale til eksperimenter. Chironomus plumosus tilhører en meget stor og divers underorden (Chironominae) af ordenen Chironomidae (dansemyg) i insektklassen. Der er mange slægter repræsenteret i Danmark, men taksonomien er dog vanskelig, og en egentlig artsbestemmelse kan kun foretages af eksperter. C. plumosus-larver (indtil 20 mm lange) lever i bunden af søer og rindende vand, hvor larverne sidder i U-formede rør dannet af mudderet. C. plumosus-larver findes typisk i store tætheder og udgør et vigtigt element i fødekæden som bl.a. føde for fisk. Larverne lever af organiske partikler (detritus, alger og bakterier), som synker ned på bunden. Partiklerne fanges med et klæbrigt fangstnet, som etableres inde i dyndrøret. Larverne æder det opfangede materiale fra nettets inderside, men kan også æde direkte fra sedimentoverfladen ved at strække forenden ud af røret. De har et højt indhold af hæmoglobin og har derfor en høj tolerance over for lave iltkoncentrationer. En generel beskrivelse af Chironomidae findes i Dall & Lindegaard (1995), hvor der også er henvisninger til speciallitteraturen. Cloeon dipterum/inscriptum (herefter kaldet C. dipterum-gr.) tilhører familien Baetidae i ordenen Ephemeroptera (døgnfluer) og er meget almindelig i damme og vandhuller. Det vandlevende nymfestadie er op til 20 mm lang og har en karakteristisk kropsform med tre haletråde med et tydeligt mørkt bånd og store afrundede gælleblade fastsiddende på bagkroppen. Gællernes placering og form anvendes ved artsidentifikation. Arten er almindelig og kan forekomme hele året i stillestående vand (damme og vandhuller). Det formodes, at dyrene kan klare en betydelig sænkning i iltkoncentrationen p.g.a. de veludviklede gæller. Dyret bevæger sig normalt rundt ved at kravle,men kan svømme ved at bugte kroppen op og ned og ved at bruge haletrådene som vifte. Nymferne lever af alger, krebsdyr og små insekter, som jages visuelt. En generel beskrivelse af Ephemeroptera samt henvisning til speciallitteraturen findes i Dall & Lindegaard (1995). Hydropsyche angustipennis tilhører familien Hydropsychidae i ordenen Trihoptera (Vårfluer) og er meget almindelig i rindende vand. Det vandlevende larvestadie er op til 30 mm lang med en delvist pansret krop med gællevedhæng på bagkroppen og krogformede fødder på bageste led. Hydropsycke angustipennis bygger fastsiddende huse på større sten og spinder et pladeformet net og placerer dette vinkelret på strømretningen således det opfanger fødepartikler fra vandet. Dyret æder dette materiale. Arten findes i eutrofierede vandløb og i afløb fra søer, hvor den bl.a. fanger zooplankton og er medtaget i dette projekt, da den er en repræsentant for faunaen knyttet til søafløb. En generel beskrivelse af Hydropsychidae findes i Dall & Lindegaard (1995) og Wiberg-Larsen (1980), som også henviser til mere specialiseret litteratur. 2.3 Forsøgsbetingelser2.3.1 Dyrkning af algekulturTil fodring af zooplanktonkulturer blev der dyrket kulturer af grønalgen Scenedesmus acutus. Algekulturerne blev dyrket som semi-kontinuerlige kulturer i Z8-vækstmedium (Kotai, 1972) i klimarum ved 20° C og en lystintensitet omkring 28 µmol s-1m-2. Kulturene blev gennemluftet med atmosfærisk luft forfiltreret med et 0,2 µm sterilfilter. Hver anden dag blev algekulturerne fortyndet med Z8-vækstmedium til en celletæthed på 50 mm-3 l-l. Algekultur anvendt til fodring blev centrifugeret ved 150 G i 10 minutter, hvorefter supernatanten blev hældt bort og erstattet af et zooplankton-medium (se nedenfor) før måling af absorbans i spektrofotometer ved 800 nm. Denne procedure sikrede, at der ikke blev overført næringsstoffer mm. til zooplanktonkulturene. Fodermængden til zooplanktonkulturerne blev beregnet ud fra en tidligere bestemt relation mellem cellevolumen af algen udtrykt i absorbans-enheder (800 nm) og kulstofindholdet per cellevolumen-enhed. 2.3.2 Zooplankton dyrkningsmediumEt standardiseret zooplankton dyrkningsmedium, ADaM-zooplankton-medium (Klüttgen et al., 1994), blev anvendt til dyrkning af zooplanktonkulturer samt som medie i alle udførte forsøg. ADaM-zooplanktonmedium fremstilles ud fra stamopløsninger af syntetisk havsalt, CaCl2, NaHCO3 og SeO2 blandet med MilliQ-vand. Før anvendelse gennemluftes mediet med atmosfærisk luft forfiltreret med et 0,2 µm sterilfilter i mindst 1/2 time, hvorefter det filtreres gennem et 0,7 µm filter og henstår i ca. 1/2 time. 2.3.3 Kultivering af forsøgsdyrLaboratoriekulturer af Daphnia magna, Daphnia galeata og Chydorus sphaericus blev dyrket i ADaM-zooplanktonmedium i 750 ml glas med et stort overflade-volumen forhold. D. magna og D. galeata blev dyrket med en tæthed på 20 individer l-l, mens antallet af C. sphaericus var variabelt og lå på over 100 individer l-l. Kulturene blev overført med plasticpipette (dafnier) eller filtreret på 50 µm net (C. sphaericus) til frisk zooplanktonmedium hver anden dag og samtidig fodret med et frisk koncentrat af grønalgen Scenedesmus acutus svarende til en slutkoncentration på 1 mg C l-l. Laboratoriekulturene blev opbevaret overdækket med stanniol i klimarum med en temperatur på 20° C og konstant svagt lys ( 1,5 µmol s-1 m-2). Visse dafniearter kan ved kultivering have tendens til at blive fanget i vandhinden i vandoverfladen. Specielt neonate (unger der blev eksponeret fra de var 24-48 timer gamle) individer af D. galeata viste sig ved overførsel til rent zooplanktonmedium ofte at blive fanget i vandhinden uden evne til at komme fri. Det resulterede i, at dyrene ikke voksede op til det adulte stadie. For at forhindre dette blev der i kulturglas med D. galeata lagt et stykke tyndt gennemsigtigt plastic oven på vandoverfladen. Generelt var D. galeata følsom over for håndtering, og individer anvendt til forsøg blev derfor dyrket i det samme testmedium indtil forsøgstidspunktet. Oprindelsen af de anvendte laboratoriekulturer kan ses i tabel 2. TABEL 2. 2.3.4 Indsamling af forsøgsdyr i feltenZooplanktonarterne D. galeata, Diaphanosoma sp., E. graciloides og C. vicinus blev indsamlet ved planktontræk med et 140 eller 200 µm net. 2.4 Koncentration-respons forsøg2.4.1 Formål og forsøgsprincipFormålet er, ud fra en akut eksponering af forsøgsorganismer, at bestemme pesticiders toksicitet som EC50- og NOEC-værdier på baggrund af effektkriteriet immobilitet. Forsøgsprincippet er, ud fra standardiserede procedurer og betingelser (International Standard, 1996), at eksponere et antal forsøgsdyr for forskellige koncentrationer af pesticid og registrere antallet af immobile individer efter hver 24. time. Koncentrationsgradienten fastlægges på baggrund af forudgående pilotforsøg, således at der registreres: ingen, middel og maksimal effekt af stoffet. 2.4.2 ForsøgsdyrDen akutte toksicitet af cypermethrin og azoxystrobin blev undersøgt på 5 arter af zooplankton og 4 arter af insektlarver. For to zooplanktonarter, D. magna og C. sphaericus, blev der udelukkende udført test med individer fra laboratoriekulturer. For D. galeata blev udført test med individer fra både en laboratoriekultur og på individer indsamlet i felten. For alle øvrige anvendte arter blev individer indsamlet i felten. For herbicidet bentazon blev den akutte toksicitet testet på en laboratoriekultur af D. magna og på individer af arten E. graciloides indsamlet i felten. 2.4.3 ForsøgsopstillingAlle forsøg blev udført under dyrkningsforholdene beskrevet i afsnit 2.3.3 og uden tilsætning af føde. Testmediet var ADaM-zooplanktonmedium tilsat forskellige mængder af pesticid fra en stamopløsning af stoffet i acetone (99,5 %) og justeret til pH 7,8 ± 0,2. Alle testopløsninger inklusiv kontrollen, som ikke indeholdt pesticid, blev tilsat den samme mængde acetone. Det relative volumetriske indhold af acetone i hver forsøgsrække fremgår af tabel 3. I forsøg, hvor indholdet af acetone var 0,3 ml l-l, blev der ligeledes lavet en kontrol, som ikke var tilsat acetone. Indledningsvist var det i pilotforsøg konstateret, at koncentrationer på 1ml acetone l-l ikke havde nogen effekt på adfærden af neonate D. magna. TABEL 3. I forsøgene indgik 7 - 11 forskellige pesticidkoncentrationer inklusiv kontrol i en eksponentiel gradient bestemt på baggrund af indledende pilotforsøg (se bilag 6.1). De anvendte koncentrationer for alle forsøg fremgår af tabel 3. I hvert forsøg blev den samlede mængde forsøgsdyr poolet i en bakke. Herfra blev 20 individer pr. pesticidkoncentration udtaget tilfældigt og overført til et antal 20 ml steriliserede glasvials (5 ml glas i forsøg med C. sphaericus), som blev påført låg. Antallet af replikater pr. testopløsning fremgår af tabel 3. Dyrene blev overført med plasticpipette, ske eller spatel. Efter hver overførsel blev det efterset, at dyrene bevægede sig normalt. Efter 24 og 48 timers (evt. også 72 og 96 timers) eksponering blev antallet af immobile individer registreret. Et individ blev karakteriseret som immobilt, hvis det ikke svømmede eller bevægede sig efter en let omrystning af glasset (zooplankton) eller en let berøring med en podenål (C. sphaericus og makroinvertebrater). Ved start og efter 48 timer (evt. 72 og 96 timer) blev iltkoncentration og pH målt med henholdsvis mini-iltelektrode og pH-meter i et enkelt glas fra samtlige testopløsninger. Til bestemmelse af forsøgsdyrenes gennemsnitslængde blev 20 individer tilfældigt udtaget og konserveret i lugol ved forsøgets start. Længden af dyrene blev enten målt i stereolup ved 10 - 30 × forstørrelse, i omvendt mikroskop ved 400 × forstørrelse (C. sphaericus) eller med digital skydelære (C. plumosus). Målskitser fremgår af figur 2. I forsøg med ægbærende D. magna blev der anvendt individer, der havde dannet deres første kuld æg i rugehulen inden for de sidste 48 timer. Alle andre forsøg blev udført med individer, der ikke bar æg. I forsøg med neonate D. magna stammede individerne fra moderdyrenes 3. kuld unger. I forsøg med H. angustipennis blev forsøgsglassene opbevaret på rystebord med lav hastighed, da arten bedst trives, i rindende vand (jf. 2.2.3). 2.4.4 DatabehandlingAntallet af immobile individer ved hver pesticidkoncentration blev anvendt til at estimere effektkoncentrationen for 50 % af testpopulationen (EC50) hver 24. time i alle forsøg. EC50 med tilhørende 95 konfidensgrænser blev bestemt ved probit analyse metoden vha. programmet Probit Analysis 2.3 (Probit Analysis 2.3, 90). Her findes der på baggrund af immobilitetsdata en passende normalfordeling til forudsigelse af sandsynligheden for, at et individ vil reagere på et toksisk stof. EC50 blev beregnet i forsøg, hvor kontrolimmobiliteten var ≤ 25 % Bestemmelse af signifikante forskelle i sensitivitet (EC50-værdier) over tid, mellem arter, mellem laboratoriekulturer og feltpopulationer og mellem neonate og ægbærende dafnier blev gjort ud fra kriteriet om ikke overlappende 95 % konfidensgrænser. EC50 blev valgt som sammenligningsgrundlag, fordi denne værdi er karakteriseret ved at besidde den laveste variation på koncentration-respons kurven (Rand et al., 1995). Bestemmelse af NOEC-værdier blev gjort vha. en-vejs ANOVA (P < 0,05) på arcsin-transformerede data efterfulgt af post-hoc test med Dunnett’s test. Disse analyser blev gjort i programmet Statistica. På baggrund af ligningerne for de vægtede regressioner bestemt ved probitanalyse blev den forventede grad af effekt i form af % immobile individer til de fundne NOEC beregnet. 2.5 Reproduktionsforsøg2.5.1 Formål og forsøgsprincipFormålet er, ud fra en eksponering af udvalgte zooplanktonorganismer, at bestemme pesticiders toksicitet som NOEC og LOEC-værdier på baggrund af effektkriteriet reproduktion. Forsøgsprincippet er, ud fra standardiserede procedurer og betingelser, at eksponere forsøgsdyr for forskellige koncentrationer af pesticid og registrere, hvornår der dannes afkom i en periode på op til 3 uger efter eksponeringsstart. Undersøgelsen er et supplement til den traditionelle akutte toksicitetstest med det formål at vurdere, hvorvidt reproduktion er en anvendelig effektparameter til vurdering af effekter af subletale pesticidkoncentrationer. 2.5.2 ForsøgsdyrI reproduktionsforsøgene blev anvendt 4 forskellige zooplanktonarter. To dafniearter: Daphnia magna og D. galeata, og to copepodarter: Eudiaptomus graciloides og Cyclops vicinus. D. magna og D. galeata var to laboratoriekulturer dyrket under standardiserede forhold som beskrevet i afsnit 2.3.3. De to copepodarter blev begge indsamlet i felten som voksne ægbærende individer umiddelbart inden gennemførelsen af forsøgene (tabel 2). TABEL 4.
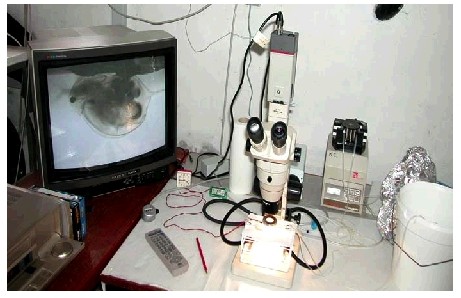
2.5.3 ForsøgsopstillingAlle forsøg blev udført under betingelser som dyrkningsforholdene for laboratoriekulturerne af zooplankton og med tilsætning af føde. Testmediet var ADaM-zooplanktonmedium tilsat forskellige mængder af pesticid fra en stamopløsning i acetone. Der blev anvendt 7 – 10 koncentrationer inklusiv kontrol i en gradient bestemt på baggrund af koncentration-respons forsøgene og indledende pilotforsøg (se bilag 6.2). De anvendte koncentrationer fremgår af tabel 4. Som udgangspunkt blev der anvendt en koncentrationsgradient, hvor de fleste koncentrationer lå under den letale dosis bestemt ved koncentration-respons forsøgene, afsnit 2.4. I reproduktionsforsøgene blev forsøgsdyrene eksponeret for pesticid igennem hele forsøgsperioden (se nedenfor), dvs. igennem en længere periode end i koncentration-respons forsøgene. Derfor forekom der letale effekter ved koncentrationer, som ikke var letale i koncentration-respons forsøgene. I hvert forsøg blev det samlede antal forsøgsdyr poolet i en bakke, hvorfra 10 tilfældigt udvalgte dyr pr. pesticidkoncentration blev overført med plasticpipette til det samme antal glasvials (et dyr pr. vial). Alle forsøg blev gennemført med 10 stk. replikater. Dyrene blev herefter tilset, at de bevægede sig normalt. Glasvials blev herefter monteret i et planktonhjul, som gennem hele forsøgsperioden roterede med en hastighed af ca. én omdr./min. Herved forblev den tilsatte føde suspenderet i vandfasen. Efter 24 og 48 timer og herefter hver anden dag blev de enkelte dyr tilset, og det blev registreret, hvorvidt de var døde (immobile efter let omrystning), og hvorvidt der var dannet æg og/eller unger. I forbindelse med tilsynet blev dyrene overført til en ny vial med frisk ADaM zooplanktonmedium, føde og pesticid. Første gang der blev observeret unger i en vial, blev indholdet i pågældende vial konserveret. Forsøgene er således ikke gennemført efter International Standard (1997). Udviklingsstadiet for de forskellige zooplanktonarter varierede. I forsøg med D. magna blev der anvendt neonate dyr (48 – 72 timer), hvor individerne stammede fra moderdyrenes 3. eller senere kuld unger. Ved forsøgsafslutning blev antal unger og æg registreret, og længden af moderdyrene blev målt vha. billedbehandlingsudstyr monteret på en Wild stereolup. I D. galeata forsøg blev anvendt 6 – 8 dage gamle dyr. Årsagen var, at juvenile D. galeata ikke tåler håndtering med plasticpipette, som D. magna gør det. Derfor blev de holdt i kultur og samtidig fodret i det ADaM medie, hvori de var klækket, indtil de havde opnået en tilstrækkelig størrelse til, at håndtering var mulig. Herefter blev de overført til glasvials som nævnt ovenfor. Ved forsøgsafslutning blev antal æg og unger registreret, samt moderdyrenes størrelse målt. I C. vicinus forsøg blev anvendt voksne ægbærende individer indsamlet i felten (se tabel 2). De indsamlede dyr blev akklimatiseret i laboratoriet først ved 10oC i 24 timer og herefter ved 20oC. Forsøgsdyr med en synlig spermatofor blev bedøvet vha. kuldioxid-holdigt vand (danskvand). Under en stereolup blev ægsække pillet fra den enkelte hun vha. skalpel og dissektionsnål. Hunnen blev herefter anbragt i en glasvial og behandlet som nævnt ovenfor. Når hunnen havde dannet nye ægsække, blev hunnen igen bedøvet, ægsækkene pillet fra, disse blev fortsat eksponeret for pesticid, mens hunnen blev fikseret. Når der fremkom nauplier fra ægsækkene, blev forsøget stoppet, og antal nauplier og æg talt op. Ægsækkene blev isoleret fra moderdyret fordi de voksne dyr udviser kannibalisme, altså spiser nauplierne. I E. graciloides forsøg blev ligeledes anvendt voksne ægbærende individer indsamlet i felten (se tabel 2). Forsøgsdyrene blev akklimatiseret tilsvarende C. vicinus. Ægsækken hos Eudiaptomus har en helt anden konsistens, den er mere geléagtig end ægsækken hos C. vicinus. Under de givne forhold kunne Eudiaptomus’ ægsæk derfor ikke håndteres uden samtidig at beskadige den. Derfor blev de indsamlede individer med ægsække eksponeret for pesticiderne. Når der første gang blev observeret nauplier, blev forsøget stoppet, og antal nauplier og æg blev optalt. 2.5.4 DatabehandlingI forbindelse med alle gennemførte forsøg, det være sig både pilot- og endelige forsøg, blev der udarbejdet en protokol, hvori alle typiske og atypiske observationer blev noteret. Dette gjorde det muligt umiddelbart efter et forsøg at sammenligne observationer og data med tidligere gennemførte forsøg. Eksempelvis kunne der forekomme pludselige og uforklarlige dødsfald blandt kontroldyrene, eller der kunne forekomme dødsfald blandt dyr behandlet ved lave koncentrationer, mens dyr i højere koncentrationer overlevede. I sådanne tilfælde blev forsøget kasseret, da vi måtte antage, at enten var der sket en fejl i håndteringen, eller også var dyrene svækkede allerede ved forsøgsstart, hvilket formentlig ville gøre dyrene mere følsomme end dyr i optimal kondition. I reproduktionsforsøgene blev udviklingstiden for æg- eller ægsækdannelsen samt udviklingstiden for neonate dafnier og nauplier ved de forskellige behandlinger testet i forhold til kontrolbehandlingerne. Hertil blev anvendt en-vejs ANOVA. Data blev logaritme-transformeret for at opfylde normalitetskravet og varianshomogenitet. Der blev ligeledes testet for forskelle i antal afkom (levende unger plus æg), her blev også anvendt en-vejs ANOVA på logaritme transformerede data. Forskelle i dyrenes størrelser blev testet på ikke-transformerede data under antagelse af, at disse data var normalfordelte. 2.6 Videoregistrering af fysiologiske parametre2.6.1 Formål og forsøgsprincipFormålet med disse studier er at undersøge pesticid-inducerede ændringer på essentielle fysiologiske parametre hos krebsdyret D. magna, således at den traditionelle akutte toksicitetstest med D. magna kan udbygges gennem identifikation af parametre, som er sensitive over for subletale pesticidkoncentrationer. Dette kan anvendes til at vurdere, hvilke parametre der kan bruges som effekt-kriterier for subletale effekter i standardiserede økotoksikologiske tests baseret på subletale pesticidkoncentrationer. Teknikken består i at iagttage aktiviteten af hjerte, brystlemmer, mandibler og bagklo hos ægbærende D. magna eksponeret for pesticider vha. videooptagelse. I hvert forsøg er et enkelt individ af D. magna fasthæftet i et forsøgskammer, der gennemstrømmes af testmedium. Forsøgskammeret placeres under et mikroskop, og det fasthæftede dyr overvåges kontinuerligt gennem 26 timer med et kamera. 2.6.2 ForsøgsdyrDe anvendte forsøgsdyr var hunner af en laboratoriekultur af D. magna (jf. tabel 2) dyrket under standardiserede forhold som beskrevet i afsnit 2.3.3. Individer, som inden for 24 timer havde dannet deres første kuld af parthenogentiske æg i rugehulen, blev udtaget til forsøg. Dafnierne var på dette tidspunkt 8 ± 1 dage gamle og havde en længde på 2,43 – 2,96 mm (målt fra hovedspids til basis af haletornen, figur 2). Dette udviklingsstadie blev anvendt for at sikre, at dafnierne ikke havde hudskifte inden for de følgende 26 timer. 2.6.3 ForsøgsopstillingAlle forsøg blev udført i et 5 ml polycarbonatkammer, der blev gennemstrømmet (1 ml min-1) af ADaM-zooplanktonmedium (jf. 2.3.2) tilsat forskellige koncentrationer af pesticid. Forsøgskammeret var forbundet med siliconeslanger til en peristaltisk pumpe (Gilson M312), der opsugede testmediet fra en bluecap-glasflaske. Testmediet blev gennemboblet af sterilfiltreret (0,2 µm) luft og havde en temperatur og pH på henholdsvis 20 ± 2° C og 7,8 ± 0,2. I hvert forsøg blev et enkelt individ af D. magna fæstnet ventralt med vaseline til et stykke glas. Glasset blev derefter påmonteret forsøgskammeret, således at dyret kunne observeres lateralt under mikroskop (Olympus SZ-CTV, 40 × forstørrelse). Kammeret blev oplyst nedenfra med en koldtlyslampe (Olympus highlight 2100). Et kamera (SONY DXC-101P) påmonteret mikroskopet registrerede dyret kontinuert over de følgende 26 timer, og alle optagelser blev lagret på videobånd (Sanyo VHR-776G recorder). Hele forsøgsopstillingen ses på figur 3. De første 2 timer af forsøgsperioden var en akklimatiseringsperiode, hvor dafnien blev eksponeret for pesticidfrit medium, mens de efterfølgende 24 timer var med pesticideksponering. I kontrolforsøg blev individerne eksponeret for rent ADaM-zooplanktonmedium i hele forsøgsperioden. Der blev ikke tilsat føde under forsøgene.
FIGUR 3. Pesticidkoncentrationer på 0,1, 1,0, 10 µg cypermethrin l-l og 0,5, 1,0, 2,0 mg azoxystrobin l-l blev fremstillet ud fra stamopløsninger af pesticiderne i acetone ved yderligere fortynding med ADaM-zooplanktonmedium. Den maksimale acetonekoncentration i forsøg med cypermethrin og azoxystrobin var henholdsvis 0,04 ml l-1 og 0,1 ml l-l. Kontrolforsøg blev ikke tilsat acetone. Til hver pesticidkoncentration blev der anvendt særskilte siliconeslanger, og forsøgskammeret blev rengjort i 99 % ethanol efter hvert forsøg. For hver koncentration af cypermethrin og azoxystrobin blev der udført forsøg på henholdsvis 5 og 3 individer, og yderligere 5 og 3 individer blev testet i pesticidfrit medium og fungerede som kontrol. Ud fra videooptagelserne blev der foretaget en manuel registrering over tiden af slagfrekvens af hjerte, brystlemmer, mandibler og bagklo samt tidspunkt for evt. varigt ophør i aktivitet af de undersøgte parametre, se figur 4. Aktiviteten blev registreret i 8 observationsperioder i løbet af forsøgsperioden på 24 timer. Den tidsmæssige placering af observationsperioderne blev bestemt på baggrund af flere forudgående pilotforsøg, hvor hele effektforløb blev gennemset for at vurdere de mest hensigtsmæssige observationstidspunkter til beskrivelse af forløbet. Den tidsmæssige placering af observationsperioderne for cypermethrin og azoxystrobin blev forskellig, på grund af stoffernes forskellige effekt på dafnierne. I forsøg med cypermethrin var en observationsperiode 5 minutter, hvor der for hver parameter (undtagen bagkloen) blev talt i 30 sekunder efterfulgt af 30 sekunders pause gentaget 5 gange. For bagkloen blev aktiviteten talt kontinuert i 5 minutter Observationsperioderne svarede til tidspunkterne 0, 15, 60, 180, 600, 960, 1320 og 1440 minutter, efter cypermethrin blev pumpet ind i forsøgskammeret. I forsøg med azoxystrobin var en observationsperiode 10 minutter, hvor der for hvert parameter (undtagen bagkloen) blev talt i 15 sekunder efterfulgt af 45 sekunders pause gentaget 10 gange. For bagkloen blev aktiviteten talt i 30 sekunder efterfulgt af 30 sekunders pause gentaget 10 gange. Observationsperioderne svarede til tidspunkterne 0, 60, 240, 420, 600, 780, 960 og 1440 minutter, efter azoxystrobin blev pumpet ind i forsøgskammeret.
FIGUR 4. Aktiviteten af bagkloen er ikke kontinuert i normalt fungerende dafnier under de givne betingelser. Derfor var observationsperioden for denne parameter af længere varighed end de øvrige undersøgte parametre. Til registrering af aktiviteten af brystlemmer, mandibler og bagklo blev der opstillet en række definitioner, mens tælling af hjerteslag var entydig. Et slag med brystlemmerne blev defineret som en koordineret bevægelse af alle brystlemmer, et slag med mandiblerne blev defineret som en bevægelse med samme amplitude, som under det respektive dyrs kontrolbetingelser (de første 2 timer af forsøgsperioden), og et slag med bagkloen blev defineret som en bevægelse, der rakte ud af dafniens skjolddele. 2.6.4 DatabehandlingMultivariat (MANOVA) og univariat (ANOVA) variansanalyse blev anvendt til statistisk behandling af data. De 4 responsvariable: slagfrekvens af hjerte, brystlemmer, mandibler og bagklo, blev målt samtidig på de samme individer. De uafhængige variable var Tid, Pesticidbehandling og Individer, som var parrede inden for pesticidbehandlingerne (skrevet Ind(Pest)), men ikke inden for tid, eftersom de samme individer (5 individer i cypermethrin forsøg og 3 individer i azoxystrobin forsøg) blev fulgt igennem hele forsøgsperioden. De gentagne observationer på hvert individ inden for en observationsperiode blev behandlet som replikerede observationer parret inden for individer. Replikaterne inden for en observationsperiode (5 i cypermethrin forsøg og 10 i azoxystrobin forsøg) blev alle udpeget til endepunktet i tidsintervallet. Variablen Tid blev behandlet som en kvalitativ variabel med 8 niveauer, svarende til et niveau for hver observationsperiode. Det første niveau (kaldet ”0”) henviser til observationer foretaget umiddelbart før, at pesticid blev tilsat mediet. Den kvalitative variabel Pesticidbehandling havde 4 niveauer (kontrol, 0,1, 1 og 10 µg l-l i cypermethrin forsøg og kontrol, 0,5, 1 og 2 mg l-l i azoxystrobin forsøg). Før de egentlige statistiske analyser blev påbegyndt, blev det undersøgt, hvorvidt data var normalfordelte med ens varianser. Nødvendige datatransformationer blev identificeret vha. Taylor’s power law (Elliott, 1971). Varianshomogenitet blev opnået ved at kvadratrodstransformere data for mandibler og bagklo i cypermethrin forsøg, mens kvadratrodstransformation og transformationen y* = y0.18 blev anvendt på data for henholdsvis mandibler og bagklo i forsøg med azoxystrobin. Eftersom de samme individer blev fulgt både før og efter pesticidtilsætning, var det vigtigt at bekræfte, at de 4 grupper af forsøgsdyr ikke var signifikant forskellige fra hinanden, allerede inden pesticid blev tilsat. Til at teste for overordnet homogenitet mellem behandlingsgrupper før pesticidtilsætning blev data for tid = ”0” analyseret vha. MANOVA. Den overordnede effekt var Pesticidbehandling (en fast faktor), mens den tilfældige faktor Ind(Pest) blev brugt som ”error term”, når der blev testet for forskelle mellem behandlingsgrupper. Forskelle mellem individer inden for behandlingsgrupper blev testet ved brug af ”mean squares” af Ind(Pest) i tælleren, og ”residual mean squares” i nævneren. Pillau’s trace (se fx. Quinn et al., 2002) blev brugt som testkriterium i de multivariate tests. Hvis MANOVA viste en overordnet forskel mellem behandlingsgrupperne før tilsætning af pesticid, blev univariate ANOVA anvendt til at identificere, hvilke af de 4 responsvariable, som udviste heterogenitet. Tukey’s multiple comparison test (Zar, 1984) blev anvendt til bestemmelse af, hvilke behandlingsgrupper der var signifikant forskellige. Forskelle i de 4 responsvariable efter tilsætning af pesticid blev analyseret vha. MANOVA, med Pesticidbehandling, Tid og interaktionen mellem Pesticidbehandling og Tid som faste faktorer og Ind(Pest) som en tilfældig faktor. På grund af manglende data til visse tidspunkter i forsøget med cypermethrin er test for behandlings- og tidsforskelle ubalancerede og derfor kun omtrentlige. Hvis MANOVA viste en overordnet effekt af pesticidbehandlingen på responsparametrene, blev separate analyser udført for at teste for forskelle mellem behandlinger, mht. hvordan behandlingerne påvirkede responsparametrene over tid. Forskelle mellem behandlinger efter justering for tidseffekter blev analyseret vha. Tukey test, når forsøget var balanceret (azoxystrobin) eller ved least-squares means (Littell et al., 1991), når forsøget var ubalanceret (cypermethrin). Ved brug af den sidstnævnte metode blev P-værdier efterfølgende justeret vha. Bonferroni proceduren (Haccou & Meelis, 1992). Dunnett’s test (Zar, 1984) blev brugt til at teste gennemsnittet af kontrolniveauet mod de andre niveauer af en given responsparameter. Alle statistiske analyser blev udført i SAS version 8e. P-værdier mindre end 0.05 blev betegnet som signifikante, undtagen i test med flere samtidige sammenligninger. Her blev signifikansniveauet justeret i overensstemmelse med antallet af samtidige sammenligninger. 2.7 Svømmeadfærd2.7.1 FormålForsøgene har til formål at bestemme, i hvilken grad cypermethrin og azoxystrobin påvirker dafniers evne til at foretage en retningsbestemt svømmeaktivitet udløst af en stimulus. I naturen anvender dafnierne denne iboende egenskab til at placere sig optimalt i forhold til fødeindtagelse og som beskyttelse mod prædatorer. Lys blev anvendt som stimulus i dette projekt. 2.7.2 ForsøgsdyrD. magna blev indsamlet i felten (tabel 2) med planktonet i september og oktober og blev holdt i 3-4 uger under standardbetingelser som laboratoriekulturerne (se afsnit 2.3.3). Dog med undtagelse af mediet som var GF/C-filteret søvand. Til hvert forsøg udvalgtes 15 ensartede individer dvs. voksne hunner med æg. 2.7.3 ForsøgsopstillingDer anvendes et svømmekammer fremstillet af et transparent plexiglasrør med en indre diameter på 5,4 cm og en totallængde på 65 cm. Røret er forsynet med vandtætte polyethylenpropper i begge ender, og i hver prop er der boret et hul med en diameter på 0,25 cm, som kan lukkes med en gummiprop. Uden om dette rør er monteret et 10 cm kortere mørkfarvet (ikke-transparent) rør. Ved at forskyde det yderste rør kan den ene ende af det inderste blotlægges og resten holdes mørklagt. Udvendige markeringer ca. 5 cm fra hver ende i det inderste rør angiver start og slut af den afstand i svømmekammeret, som dafnierne kan svømme. Denne banes længde er 60 cm. I den faste forsøgsopstilling indgår det beskrevne dobbelte rør placeret vandret på et dertil indrettet bord, en lyskilde bestående af en koldtlyslampe med to bøjelige punktlys, et stopur samt en bærbar pc til indtastning af rådata. Se figur 5. Lysintensiteten i den mørklagte ende er ca. 1 µmol fotoner m-1 s-1 og 232 µmol fotoner m-1 s-1 i den belyste ende.
Figur 5. Umiddelbart før et forsøg placeres ca. 15 dafnier i 1 l ADaM-medium uden føde i 1 time. I slutningen af denne periode fyldes svømmekammeret med ADaM-medium og placeres i lodret position. Ti dyr overføres herefter til svømmekammeret vha. en pipette, som indføres gennem det lille hul i den ene prop. Proppen lukkes og røret bringes herefter i vandret position, således at lyskilderne findes i modsatte ende af dyrene, hvorefter stopuret startes, og det ydre rør skubbes hen over området med dyr. Tidspunktet for den første dafnies ankomst ved en markering ca. 5 cm fra rørets modsatte ende registreres manuelt og tilsynekomst af de øvrige dafnier sammesteds registres herpå efter 1, 1,5, 2, 3, 4 og 5 minutter. Når alle dafnier er samlet, vendes røret, således at lyskilden er placeret modsat dafnierne. Registrering af dafnierne foretages som omtalt. Disse procedurer gentages i alt fem gange umiddelbart efter hinanden og betegnes kontrolforsøg. Herefter overføres dafnierne og mediet til en plasticbeholder. Svømmekammeret fyldes herefter med nyt ADaM-medium tilsat cypermethrin i den ønskede koncentration (0,05; 0,075; 0,1; 0,5 eller 1 µg l-1). Dafnierne overføres atter til røret, deres svømmeaktivitet registreres efter samme princip som beskrevet ovenfor. Denne del af forsøget betegnes eksponeringsforsøget. Efter sidste registrering opsamles alle 10 dafnier i en glasvial og konserveres med sur lugol-opløsning (3%). Dafniers længde fra øje til basis af haletorn måles efterfølgende i et Wild stereomikroskop ved 30 gange forstørrelse. Der blev udført et hhv. kontrol- og eksponeringsforsøg med azoxystrobin efter samme principper som beskrevet ovenfor. Den anvendte stofkoncentration var 1 mg l-1. Alle eksperimenter er udført i et termosteret rum med dæmpet belysning (< 1 µmol fotoner m-1 s-1) og ved 20oC (±1oC) samt med samme megapopulation af D. magna. 2.7.4 DatabehandlingAlle tidsregistreringer omsættes til minut-decimaltal. Den maksimale svømmehastighed beregnes (m min-1) ved at dividere svømmetiden for den først registrerede dafnie med svømmebanens længde. Svømmehastigheden i eksponeringsforsøgene udtrykkes som procent af hastigheden i de respektive kontrolforsøg. I samtlige forsøg beregnes det geometriske gennemsnit og standardafvigelsen (SD) til dette. Svømmeaktiviteten for de øvrige registrerede dafnier er udtrykt som den andel af den samlede population i røret, der har tilbagelagt den samlede distance på en given tid. Den tilsvarende svømmehastighed udtrykker kun en minimumssvømmehastighed, idet det ikke kunne undgås (se bilag 6.3), at dafnierne efter en vis tid (ca. 2 min i kontrolforsøgene og 2-5 min i eksponeringsforsøgene) svømmede frem og tilbage i synsfeltet. Det skal noteres, at hastighedsudtrykkene kun angiver den målte en-dimen-sionelle (lineære) distance, som et givent dyr har svømmet, og ikke den faktiske tredimensionelle længde. Envejsfaktor ANOVA blev anvendt til at teste, hvorvidt: 1) den maksimale svømmehastighed i mellem kontrolforsøgene var signifikant forskellig, 2) der ved pesticideksponeringen kunne registreres en signifikant ændring i den maksimale svømmehastighed sammenlignet med det respektive kontrolforsøg, 3) der var signifikante forskelle i den maksimale svømmehastighed og i den procentvise reduktion af svømmehastigheden mellem de forskellige koncentrationer af pesticid (kun for cypermethrin). Sammenhængen mellem den procentvise reduktion af den maksimale svømmehastighed og koncentrationen af cypermethrin blev fastlagt ved regressionsanalyse, hvor EC50 for den maksimale svømmehastighed kunne bestemmes.
|