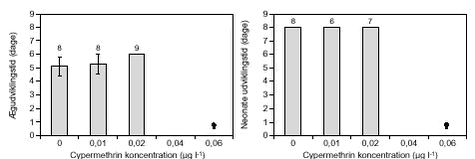
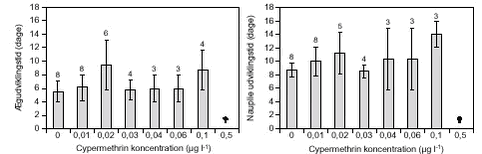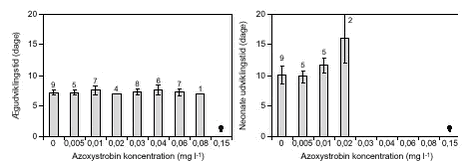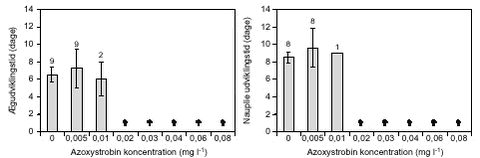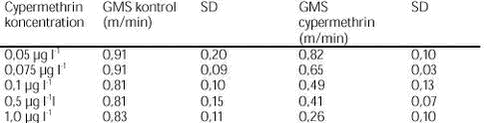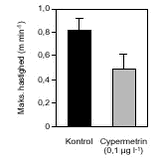Effekter af cypermethrin, azoxystrobin og bentazon på limniske invertebrater3 Resultater3.1 Koncentration-respons forsøg3.1.1 Cypermethrin 3.1.2 Azoxystrobin 3.1.3 Bentazon 3.2 Reproduktionsforsøg 3.2.1 Cypermethrin 3.2.2 Azoxystrobin 3.3 Videoregistrering af fysiologiske parametre 3.3.1 Cypermethrin 3.3.2 Azoxystrobin 3.4 Svømmeadfærd 3.4.1 Cypermethrin 3.4.2 Azoxystrobin 3.5 Pesticidkoncentrationer 3.1 Koncentration-respons forsøg3.1.1 CypermethrinSammenligning af EC50-værdier efter 24 og 48 timers eksponering viste, at sensitiviteten for alle arter undtaget makroinvertebraterne C. dipterum-gr. og H. angustipennis steg med eksponeringstiden (tabel 5). For 5 af arterne steg sensitiviteten signifikant svarende til et signifikant fald i EC50-værdien fra 24 – 48 timers eksponering. Forskellen i sensitivitet med tiden var størst for ægbærende D. magna, hvor EC50-værdien til eksponeringstiden 48 timer var en faktor 32 mindre end EC50 til eksponeringstiden 24 timer. E. graciloides havde den mindste forskel i sensitivitet med tiden. Her var EC50 til eksponeringstiden 48 timer 1,4 gange lavere end EC50 til eksponeringstiden 24 timer (tabel 5). Sammenligning af EC50-værdier for de testede arter viste, at D. magna var signifikant mindre sensitiv end alle andre arter efter 24 timers eksponering, mens D. magna’s sensitivitet lå på samme niveau som både C. sphaericus og H. angustipennis efter 48 timers eksponering (tabel 5). E. graciloides og laboratoriekulturen af D. galeata var de mest sensitive zooplanktonorganismer med EC50 på henholdsvis 0,14 (0,08 – 0,21) µg cypermethrin l-l og 0,08 (0,05 - 0,12) µg cypermethrin l-l efter 48 timers eksponering. Makroinvertebraterne C. plumosus, C. flavicans og C. dipterum-gr. havde alle samme sensitivitetsniveau på 0,02 - 0,06 µg cypermethrin l-l efter 48 timers eksponering. C. dipterum-gr. var dog signifikant mere sensitiv end de øvrige makroinvertebrater efter 24 timers eksponering (tabel 5). H. angustipennis var den mindst sensitive makroinvertebrat. Overordnet var døgnfluenymfen C. dipterum-gr. den mest sensitive af de testede invertebrater efter 24 timers eksponering (EC50 = 0,05 (0,04 – 0,07) µg cypermethrin l-l) og var ligeledes sidestillet med myggelarven C. flavicans de mest sensitive testede arter efter 48 timers eksponering med EC50 på henholdsvis 0,05 (0,04 – 0,06) µg cypermethrin l-l og 0,02 (0,01 – 0,04) µg cypermethrin l-l (tabel 5). C. flavicans havde også den laveste NOEC på 0,005 µg cypermethrin l-l efter 48 timers eksponering, mens ægbærende D. magna havde den højeste NOEC på 0,25 µg cypermethrin l-l (tabel 5). TABEL 5. Sammenligning af EC50-værdier for neonate og ægbærende D. magna viste, at neonate dafnier var mere sensitive over for cypermethrin end ægbærende dafnier efter både 24 og 48 timers eksponering. Forskellene var dog kun signifikante efter 24 timers eksponering. Forskellen i EC50-værdier mellem neonate og ægbærende dafnier var ca. en faktor 8 efter 24 timers eksponering (tabel 5). De brede konfidensgrænser omkring EC50-værdien for ægbærende D. magna til tiden 24 timer skal bemærkes og er resultatet af et manglende maksimalt respons (for få dyr dør) ved de anvendte koncentrationer. Sammenligning af EC50-værdier for felt- og laboratoriekulturer af D. galeata viste ingen signifikante forskelle i sensitivitet over for cypermethrin (tabel 5). På baggrund af koncentration-respons sammenhængen blev det forventede respons i form af immobilitet ved de fastlagte NOEC fundet til at variere mellem EC1,4 og EC42 (tabel 5). Det betyder med andre ord, at 1,4 – 42 % af forsøgspopulationen kunne forventes at være påvirket ved de fastlagte NOEC. For 50 % af NOEC-værdierne kunne det forventes, at ≤ 10 % af forsøgspopulationen blev immobiliseret, mens kun 20 % af NOEC-værdierne ville kunne forventes at give en immobilisering på ≤ 5 % af forsøgspopulationen. Ud over effekten af cypermethrin på mobiliteten af de anvendte invertebrater blev der også observeret en subletal effekt på H. angustipennis’s netspindingsadfærd. Efter 48 timers eksponering blev det i alle forsøgsglas med koncentrationer ≥ 0,62 µg cypermethrin l-l observeret, at der ikke var spundet net i disse glas modsat glas med koncentrationer < 0,62 µg cypermethrin l-l, hvor der var net til stede. I alle forsøg var mængden af opløst oxygen ≥ 6,6 mg O2 l-l efter 48 timers eksponering,, undtagen i forsøg med ægbærende D. magna, C. plumosus og C. flavicans, hvor iltindholdet var ≥ 3,73, ≥ 2,42 og ≥ 3,19 mg O2 l-l efter 48 timers eksponering. For alle forsøg varierede pH mellem 6,23 – 7,84 målt efter 48 timer. I de enkelte forsøg lå pH i opløsninger med pesticid inden for ± 0,91 enheder i forhold til pH i kontrolopløsningerne. Immobilisering af 10 % og 15 % af kontrolindividerne blev observeret i forsøg med henholdsvis D. galeata (F) og Diaphanosoma sp. efter 24 timers eksponering. Kontrolimmobiliseringen var 5 % for E. graciloides, 10 % for C. flavicans, 65 % for D. galeata (F) og 75 % for Diaphanosoma sp. efter 48 timers eksponering. I alle øvrige forsøg var der ingen immobiliserede kontrolindivider. 3.1.2 AzoxystrobinFor alle testede makroinvertebrater undtagen myggelarven C. plumosus var sensitiviteten over for azoxystrobin meget lav. For C. flavicans kunne der ikke observeres nogen effekt ved den maksimale testkoncentration på 6 mg azoxystrobin l-l. For C. dipterum-gr. og H. angustipennis blev der observeret en koncentration-respons sammenhæng efter henholdsvis 48 og 24 timer, mens der på trods af immobile individer i de højeste testkoncentrationer ikke kunne observeres en koncentration-respons sammenhæng til de øvrige tidspunkter (tabel 6). Modsat makroinvertebraterne var alle testede zooplanktonorganismer følsomme over for azoxystrobin. Sammenligning af EC50-værdier efter 24 og 48 timers eksponering for de følsomme arter viste, at sensitiviteten over for azoxystrobin steg med eksponeringstiden for 6 af de 7 arter, hvor EC50-værdier lod sig beregne til begge tidspunkter (tabel 6). For 4 af disse arter faldt EC50-værdien signifikant med ca. en faktor 2 fra 24 til 48 timers eksponering. Sammenligning af EC50-værdier for de følsomme arter viste, at efter 24 timers eksponering var D. magna signifikant mindre sensitiv end alle andre arter undtagen vårfluelarven H. angustipennis. H. angustipennis blev beregnet på baggrund af kun 5 immobile individer, heraf 3 immobile i den højeste koncentration, og konfidensgrænserne omkring EC50-værdien er derfor brede. TABEL 6. Efter 48 timers eksponering var C. dipterum-gr. den mindst sensitive art med en EC50 = 3,56 (2,72 – 5,12) mg azoxystrobin l-l, mens E. graciloides var den signifikant mest sensitive art med en EC50 = 0,038 (0,03 – 0,04) mg azoxystrobin l-l (tabel 6). C. flavicans havde den højeste NOEC på > 6 mg azoxystrobin l-l, mens E. graciloides havde den laveste NOEC på 0,034 mg azoxystrobin l-l (tabel 6). Sammenligning af EC50-værdier for neonate og ægbærende D. magna viste, at neonate dafnier var signifikant mere sensitive over for azoxystrobin end ægbærende dafnier efter både 24 og 48 timers eksponering. Forskellen i EC50-værdier mellem neonate og ægbærende dafnier efter 24 og 48 timers eksponering var henholdsvis en faktor 3,6 og 3 (tabel 6). Sammenligning af EC50-værdier for laboratoriekulturen af D. galeata og D. galeata indhentet i felten viste, at laboratoriekulturen var signifikant mere sensitiv over for azoxystrobin end feltpopulationen efter både 24 og 48 timers eksponering (tabel 6). På baggrund af koncentration-respons sammenhængen blev det forventede respons i form af immobilitet ved de fastlagte NOEC-værdier fundet til at variere mellem EC2 og EC40 (tabel 6). For omtrent 45 % af NOEC-værdierne kunne det forventes, at < 10 % af forsøgspopulationen blev immobiliseret, mens 19 % af NOEC-værdierne ville kunne forventes at immobilisere ≤ 5 % af forsøgspopulationen. I alle forsøg var mængden af oxygen ≥ 6,5 mg O2 l-l efter 48 timers eksponering, undtagen i forsøg med ægbærende D. magna og C. plumosus, hvor mængden af opløst oxygen var henholdsvis ≥ 2,24 og ≥ 2,15 mg O2 l-l efter 48 timers eksponering. For alle forsøg varierede pH mellem 6,4 – 8,19 målt efter 48 timer. I de enkelte forsøg lå pH i opløsninger med pesticid inden for ± 0,8 enheder i forhold til pH i kontrolopløsningerne. Immobilisering af 5 % af kontrolindividerne blev observeret i forsøg med neonate D. magna og C. sphaericus efter henholdsvis 24 og 48 timer. Kontrolimmobiliseringen var 10 % og 20 % efter henholdsvis 24 og 48 timer i forsøg med E. graciloides og 25 % efter 48 timer i forsøg med D. galeata (F). I alle øvrige forsøg var der ingen immobiliserede kontrolindivider. 3.1.3 BentazonBentazon havde ingen effekt på E. graciloides ved koncentrationer op til 500 mg l-l, mens stoffet var meget svagt toksisk over for D. magna, med en EC50 efter 48 timer på 3797 mg l-l og en NOEC efter 48 timer på 125 mg l-l (tabel 7). EC50 for D. magna skal tages med forbehold, da bestemmelsen er gjort på baggrund af kun 2 og 4 immobile individer efter henholdsvis 24 og 48 timers eks-ponering registreret ved eksponeringskoncentrationer ≥ 250 mg bentazon l-l. NOEC-værdien til tiden 48 timer skal ligeledes tages med forbehold, idet eksponering for 250 mg l-1 gav et signifikant anderledes respons end kontrollen, mens responset ved eksponering for 500 mg l-1 ikke var signifikant forskellig fra kontrollen (jf. tabel 3). For mobile D. magna var der både efter 24 og 48 timers eksponering for ≥ 250 mg bentazon l-l flere individer, der lå i vandhinden. Effekten synes at opstå i forbindelse med hudskifte, hvor dafnierne havde svært ved at komme fri af den gamle hud. TABEL 7. I alle forsøg var mængden af opløst oxygen ≥ 7,6 mg O2 l-l efter 48 timers eksponering. pH varierede mellem 7,2 – 7,6, og i de enkelte forsøg lå pH i opløsninger med pesticid inden for ± 0,2 enheder i forhold til pH i kontrolopløsningerne. Ingen kontrolindivider var immobiliseret eller udførte unormal adfærd. 3.2 Reproduktionsforsøg3.2.1 CypermethrinHer er beskrevet to forsøg med D. magna. I første forsøg blev der fokuseret på koncentrationer op til 1 µg l-1. Mellem 80 – 90 % af forsøgsdyrene overlevede koncentrationer op til 0,02 µg l-1. Ved 0,04 µg l-1 (LOEC, tabel 8) døde 80 % af dyrene døde efter 4-6 dage og de overlevende udviklede ikke æg (figur 6). Ved højere koncentrationer døde alle dyr efter 4 døgn. Ægudviklingstiden lå på 5 – 6 dage og var ikke længere ved 0,02 µg l-1 end i kontrolbehandlingerne. I alle tre behandlinger med ægudvikling skete der også en videreudvikling til neonate indenfor 8 dage (figur 6). Pesticidpåvirkningen havde ikke effekt på moderdyrenes længde. I det andet forsøg blev der fokuseret på koncentrationer < 0,04 µg l-1 (figur 7). Der var ikke nogen effekt af cypermethrin på hverken ægudviklingstiden eller udviklingstiden af neonate inden for koncentrationsgradienten 0 – 0,08 µg l-1. Ved alle behandlinger overlevede 90 – 100 % af forsøgsdyrene. Forsøgsdyrenes størrelse varierede heller ikke som følge af pesticidpåvirkningen, kontroldyrene målte 3,03 ± 0,06 mm, mens dyrene ved 0,08 µg l-1 målte 2,95 ± 0,1 mm.
FIGUR 6.
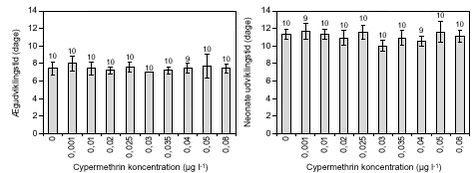
FIGUR 7. Reproduktionen hos D. galeata blev testet ved cypermethrin koncentrationer op til 0,08 µg l-1 (figur 8). Op til 0,04 µg l-1 var der ingen signifikant forskel i ægudviklingstiden (NOEC, tabel 8), men dog tendens til en svag stigning i udviklingstiden. Ved 0,05 µg l-1 (LOEC, tabel 8) var ægudviklingstiden signifikant (p = 0,039) længere, 11,6 dage mod 6,6 dage i kontrolbehandlingen. Samtidig reduceredes antallet af ægproducerende dafnier også fra 7 – 10 i kontrolbehandlingen og de lavere koncentrationer til 5 ved 0,05 µg l-1. Ved 0,08 µg l-1 døde alle dafnier indenfor 2 – 7 dage og dermed inden ægdannelsen. Havde D. galeata udviklet æg skete der en videreudvikling til neonate hos næsten alle dafnier (figur 8). Der var ikke forskel på udviklingstiden fra æg til neonate, den varierede i alle behandlinger mellem 2 – 3 dage. Imidlertid var antal neonate plus æg pr. D. galeata signifikant (p < 0,001) mindre ved 0,05 µg l-1 (3,3 neonate + æg) end i kontrolbehandlingen (21 neonate + æg), ligesom de eksponerede dafnier var mindre end i kontrolbehandlingen (p < 0,001) ved forsøgsafslutningen (figur 9).
FIGUR 8.
FIGUR 9. FIGUR 10.
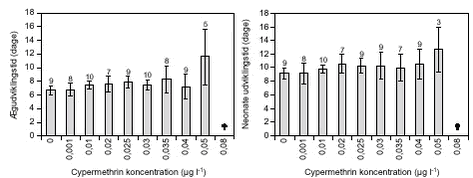
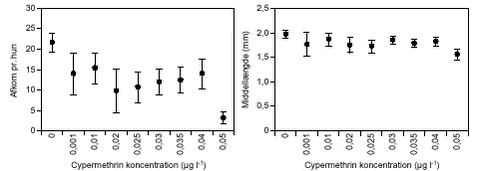
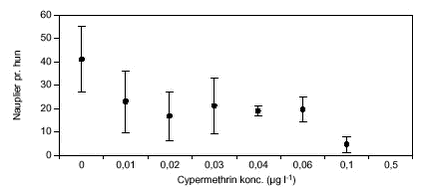
FIGUR 11. C. vicinus blev testet ved cypermethrin koncentrationer op til 0,5 µg l-1 (figur 10). Ved koncentrationer over 0,02 µg l-1 overlevede kun 30 – 40 % af forsøgsdyrene, de resterende dyr døde inden for 5 – 20 dage og ved 0,5 µg l-1 døde alle forsøgsdyr inden for 72 timer. Alle overlevende dyr udviklede ægsække inden for 5 – 9 dage, og der var ikke signifikante forskelle på ægudviklingstiden mellem behandlingerne. Hos næsten alle dyr, der havde udviklet ægsække, skete der en videreudvikling til nauplier (figur 10) inden for en periode på 1,5 – 6 dage i forhold til tidspunktet, hvor ægsækkene var dannet. Ved 0,1 µg l-1 (LOEC, tabel 8) var nauplie udviklingstiden signifikant (p = 0,008) længere end hos kontroldyrene, og ved koncentrationer > 0,03 udvikledes der signifikant færre nauplier pr. C. vicinus, helt ned til kun 4 nauplier pr. hun mod 42 nauplier pr. hun i kontrolbehandlingen (figur 11). TABEL 8. E. graciloides blev testet ved koncentrationer op til 1,0 µg l-1 (figur 12). Ægudviklingstiden blev ikke undersøgt for denne art p.g.a. metodevalget (se afsnit 2.5.3). Hos Eudiaptomus var det meget variabelt, hvor mange forsøgsdyr der overlevede. Der var ikke noget generelt mønster for, ved hvilke behandlinger dyrene overlevede. Dog var der færrest overlevende dyr (4) ved den højeste cypermethrinkoncentration. Udviklingstiden fra æg til nauplie varierede mellem 1 – 1,7 dage ligeledes uden et egentligt mønster i forhold til behandling. Antal dannede nauplier ved forsøgsafslutningen varierede mellem 16 og 23 nauplier pr. Eudiaptomus, men uden signifikante forskelle (figur 13).
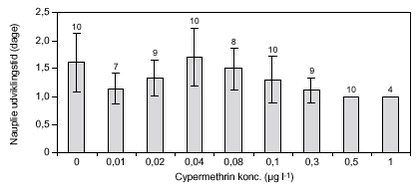
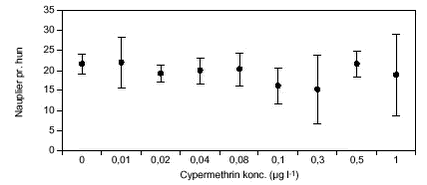
FIGUR 12.
FIGUR 13. 3.2.2 AzoxystrobinD. magna blev testet ved azoxystrobinkoncentrationer op til 0,28 mg l-1 (figur 14). Næsten alle forsøgsdyr overlevede eksponeringen, kun ved den højeste koncentration døde et større antal dyr (50 %). Disse døde indenfor en periode på 2 – 5 dage. Alle overlevende dyr udviklede æg inden for en periode på 6 – 7 dage, og der var ingen forskelle i udviklingstiden. I løbet af yderligere 2 – 3 dage blev der udviklet neonate, men kun ved koncentrationer på op til 0,04 mg l-1. Ved de højere koncentrationer skete der ingen videreudvikling af æggene, på trods af, at alle ægbærende dyr overlevede (figur 14). LOEC var således 0,08 azoxystrobin mg l-l (tabel 8). I de tre behandlinger, hvor der skete en udvikling af neonate, faldt antallet af individer, som udviklede neonate med øget pesticidkoncentration, fra 7 i kontrollen til 2 ved 0,04 mg l-1.
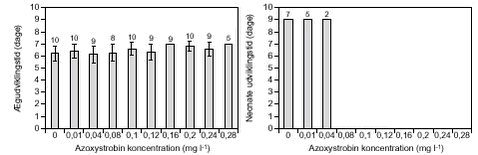
FIGUR 14.
FIGUR 15. Middellængden af D. galeata blev signifikant (p < 0,001) forøget ved koncentrationer ≥ 0,02 mg l-1 (figur 17) sammenlignet med kontrolbehandlingen. Evnen til at udvikle æg var, ligesom hos D. magna, ikke entydigt svækket som følge af azoxystrobineksponeringen, men der var meget stor variation i ægdannelsen hos de eksponerede dyr.
FIGUR 16.
FIGUR 17. C. vicinus blev testet ved koncentrationer op til 0,1 mg l-1 (figur 18). Ved koncentrationer ≥ 0,02 mg l-1 (LOEC, tabel 8) døde alle forsøgsdyr inden for 48 timer. Ved koncentrationerne < 0,02 mg l-1 udvikledes ægsække; ved 0,01 mg l-1 dog kun hos 25 % (2 dyr) af de overlevende dyr. Ægudviklingstiden var 6 – 7 dage uafhængig af azoxystrobinkoncentrationen. Efter yderligere 2 – 3 dage udvikledes der nauplier fra ægsækkene. Med undtagelse af et enkelt dyr pr. behandling udvikledes der nauplier fra alle ægbærende dyr (figur 18). Der var tendens til et reduceret antal nauplier pr. C. vicinus ved øget azoxystrobinkoncentration, men forskellen var ikke signifikant (figur 19).
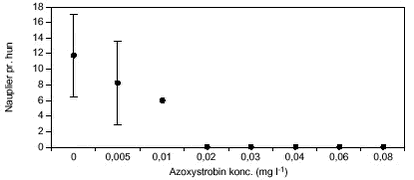
FIGUR 18. E. graciloides blev testet ved koncentrationer op til 0,01 mg l-1 (figur 20). Ved 0,01 mg l-1 døde alle forsøgsdyr inden for 48 timer. Ved de lavere koncentrationer udvikledes der nauplier hos 80 – 100 % af forsøgsdyrene inden for 1,5 – 3 dage. Der var ikke signifikante forskelle i udviklingstiden uanset eksponeringskoncentration. Fra kontrolbehandlingen op til en azoxystrobinkoncentration på 0,002 mg l-1 lå nauplie-antallet pr. E. graciloides mellem 14 – 15 nauplier. Ved 0,005 mg l-1 (LOEC, tabel 8) var nauplie-antallet signifikant (p = 0,032) lavere (9,5 nauplier pr. E. graciloides, figur 21).
FIGUR 19.
FIGUR 20.
FIGUR 21. 3.3 Videoregistrering af fysiologiske parametre3.3.1 CypermethrinKvalitative effekter på D. magna De første symptomer på en effekt af cypermethrineksponeringen var en forøgelse i aktivitet af brystlemmer og mandibler, som udmøntede sig i sitrende bevægelser med lav amplitude (krampetilstand). De sitrende bevægelser blev hurtigt efterfulgt af en periode med helt eller delvist ophør i aktivitet, hvorfra der evt. kunne observeres en senere stigning i aktivitet. Samtidige effekter kunne observeres på aktiviteten af bagkloen og hjertet, som begge blev forøget. Effekterne opstod efter ca. 15 minutters eksponering. Homogenitet af forsøgsdyr før cypermethrin tilsætning Behandlings- og tidseffekter Med stigende cypermethrinkoncentration og stigende eksponeringstid viser aktiviteten af responsparametrene efter cypermethrintilsætning også en klar tendens til at være mere forskellige fra aktiviteten før cypermethrintilsætning (tabel 10, signifikante forskelle markeret med kursiverede tal) og fra kontrolaktiviteten (tabel 10, signifikante forskelle markeret med *). TABEL 9. Tabel 10. Dafnierne i kontrollen ændrede også adfærd i løbet af de 24 timer, forsøget varede (F24,616 = 4,75; P < 0,0001). Men for alle responsparametre var det kun til enkelte tidspunkter, at aktiviteten var signifikant forskellig fra startniveauet i kontrollen (tabel 10, signifikante forskelle markeret med *). For alle responsparametre (undtaget bagkloen ved eksponering for 10 µg l-l) indtrådte den maksimale effekt af stoffet inden for en eksponeringstid på 3 timer (figur 22 - 25). Men allerede efter 15 minutters eksponering for 1 µg cypermethrin l-l kunne der, sammenlignet med niveauet før cypermethrin-tilsætning, observeres effekter på aktiviteten af brystlemmer, mandibler og bagklo (signifikante forskelle markeret med * i tabel 10). Overordnet medførte cypermethrineksponering en signifikant forøgelse af aktiviteten af hjertet og bagklo og en signifikant reduktion i aktiviteten af brystlemmer og mandibler hos dafnierne (jf. tabel 10, figur 22 - 25). Efter 24 timers eksponering for 10 µg l-l var aktiviteten af hjertet, bagkloen, brystlemmerne og mandiblerne henholdsvis 131 %, 1650 %, 33 % og 53 % af aktiviteten før cypermethrin eksponering. Bagkloen var den mest følsomme responsparameter over for cypermethrin (F3,16 = 89,2; P < 0,0001), mens effekten af cypermethrin var mindst for mandiblerne (F3,16 = 5,98; P < 0,0062). Brystlemmeaktiviteten var den parameter, der ændrede sig mest over tiden (F6,631 = 22,1; P < 0,0001), mens hjerteaktiviteten ændrede sig mindst (F6,619 = 3,19; P < 0,0043). Aktiviteten af brystlemmerne udviste størst interaktion mellem cypermethrinbehandling og tid (F18,631 = 12,4; P < 0,0001), men interaktionen for de 3 andre responsparametre var også stærkt signifikant (tabel 10). Lineær kontrastanalyse mellem aktiviteten af responsparametrene før og efter cypermethrintilsætning viste, at bagklo og brystlemmer blev signifikant påvirket ved en cypermethrinkoncentration på 0,1 µg l-l. Mandibelaktiviteten blev signifikant påvirket ved en cypermethrinkoncentration på 1 µg l-l, mens hjerteaktiviteten var signifikant forskellig fra startaktiviteten ved eksponering for 10 µg cypermethrin l-l (tabel 10). Eftersom hjerteaktiviteten som udgangspunkt var signifikant forskellig mellem dafnier i kontrollen og dafnier eksponeret for 1 µg l-l, ses der signifikante behandlingseffekter ved 1 µg l-l (markeret ved kursiverede tal i tabel 10), men derimod ingen nævneværdige forskelle mellem aktiviteten før og efter cypermethrintilsætning (signifikante forskelle markeret med * i tabel 10). For mandiblerne var der en tendens til, at effekten af cypermethrin aftog med tiden, således at aktiviteten indstillede sig på et nyt niveau tættere på startaktiviteten efter 24 timers eksponering. Denne tendens blev observeret hos dafnier eksponeret for 1 µg l-l og 10 µg l-l efter henholdsvis 16 og 10 timers eksponering (figur 22 - 25). Selv om effekten på mandiblerne fortog sig med tiden, var aktiviteten dog stadig signifikant forskellig fra startniveauet (tabel 10). For de øvrige responsparametre indstillede aktiviteten sig efter ca. 6 timers eksponering omkring niveauet for den maksimale effekt ved den respektive koncentration (figur 22 - 25).
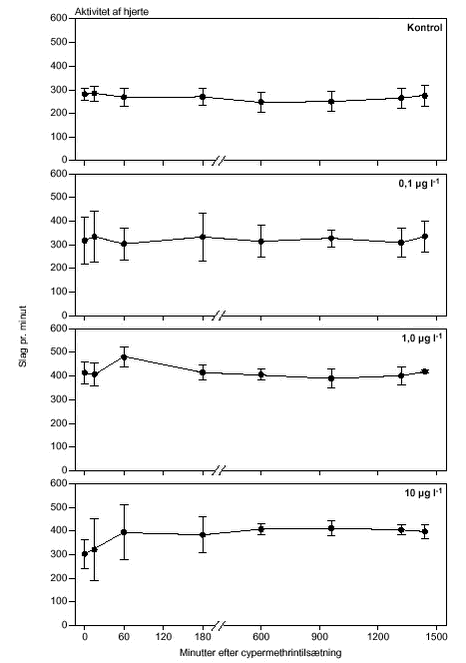
FIGUR 22.
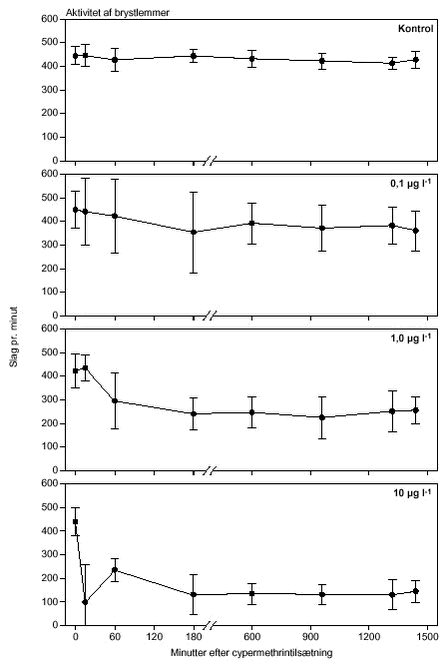
FIGUR 23.
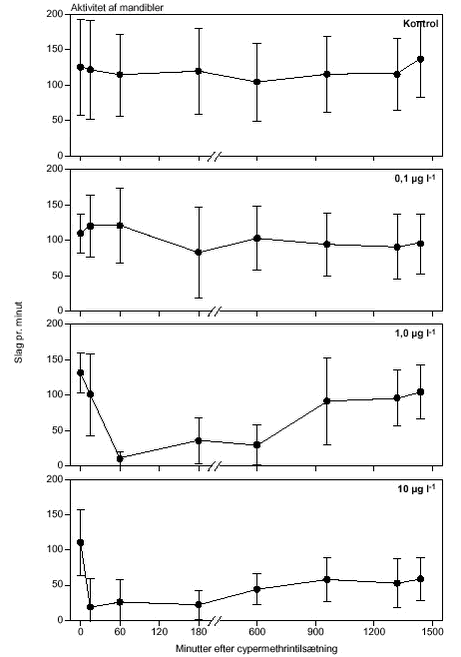
FIGUR 24.
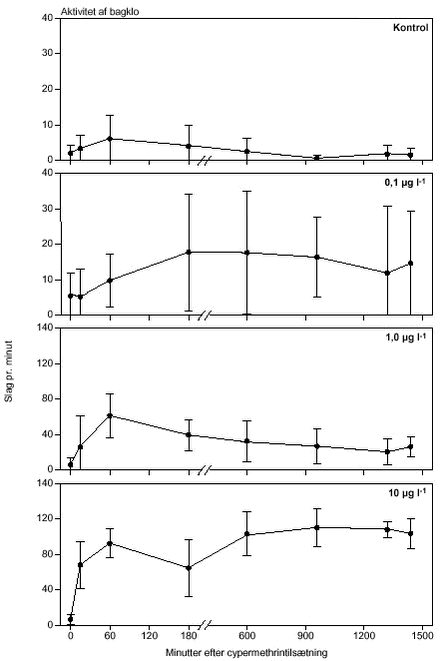
FIGUR 25. 3.3.2 AzoxystrobinKvalitative effekter på D. magna Homogenitet af forsøgsdyr før azoxystrobin tilsætning Univariate ANOVA viste, at brystlemmeaktiviteten, der varierede mellem 428 – 518 slag min-1 i de 4 behandlingsgrupper, var den mest heterogene responsparameter før azoxystrobintilsætning. Isoleret set var brystlemmeaktiviteten signifikant forskellig mellem behandlingsgrupperne (F3,8 = 6,18; P = 0,018), med en kontrolaktivitet signifikant (P < 0,05) lavere end grupperne eksponeret for 0,5 og 2 mg azoxystrobin l-l (Tukey’s test, tabel 11). Behandlings- og tidseffekter F24,804 = 12,64, 1 mg l-l: F24,804 = 17,22, 2 mg l-l: F24,804 = 17,56; P < 0,0001 for alle koncentrationer, tabel 11), hvor effekten steg jævnt med eksponeringstiden. For alle parametre sås den maksimale effekt af eksponeringen først til tiden 24 timer (figur 26 - 29). Dog var der en tendens til, at dafniernes hjerteaktivitet indstillede sig på et nyt signifikant lavere niveau efter 10 timers eksponering for 0,5 og 1,0 mg azoxystrobin l-l (figur 26). Dafnier i kontrollen ændrede også adfærd i løbet af de 24 timer, forsøget varede (0 mg l-l: F24,804 = 4,05; P < 0,0001), men det var særligt aktiviteten af hjerte og mandibler, der udviste signifikante forskelle til startniveauet (tabel 12, signifikante forskelle markeret med *). TABEL 11.
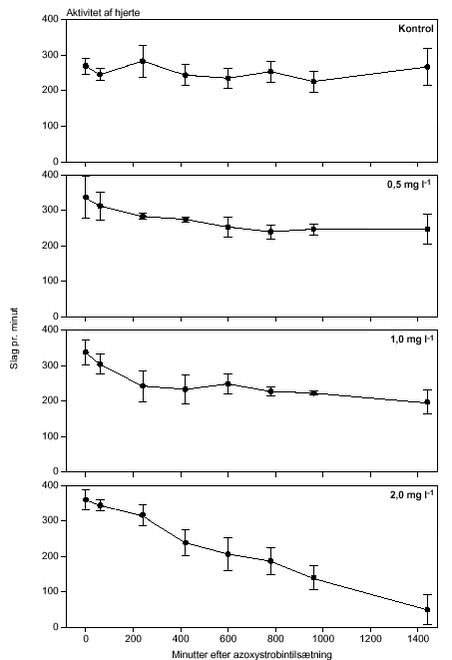
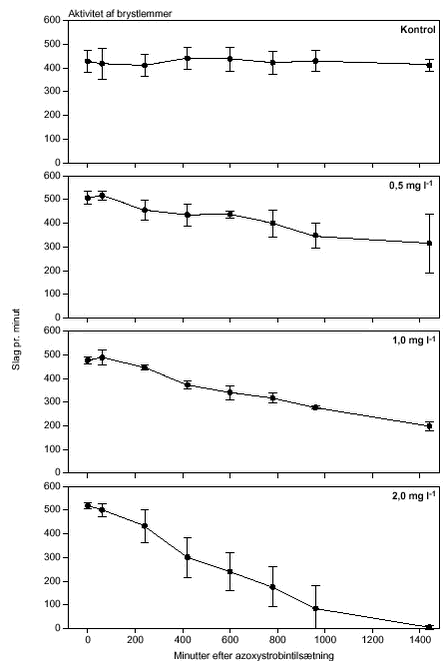
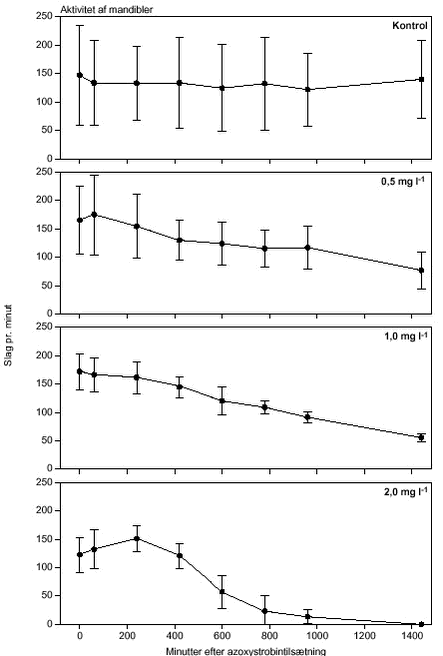
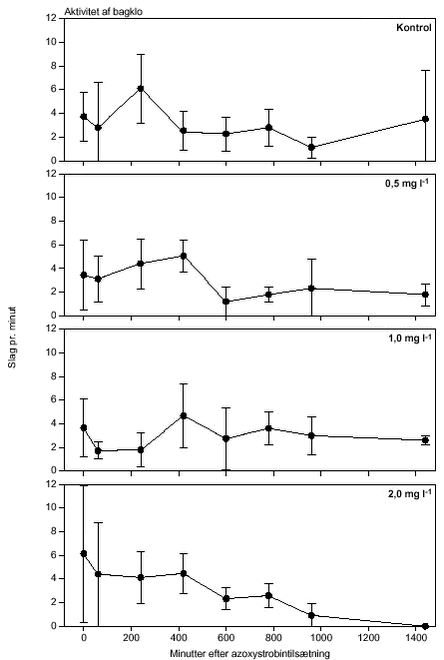
FIGUR 26.
FIGUR 27.
FIGUR 28.
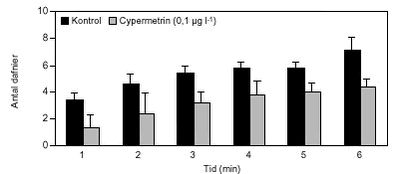
FIGUR 29. Overordnet medførte azoxystrobineksponering en reduktion i aktiviteten af alle 4 responsparametre (ANOVA, tabel 12 og figur 26-29). Efter 24 timers eksponering for 2 mg azoxystrobin l-l var aktiviteten af hjertet og brystlemmer henholdsvis 14 % og 0,8 % af aktiviteten før azoxystrobineksponering, mens aktiviteten af mandibler og bagklo var ophørt. Brystlemmeaktiviteten var den mest følsomme responsparameter over for azoxystrobin (F = 10,58; P < 0,0037), mens effekten af azoxystrobin var mindst og langt fra signifikant for bagkloen (F3,8 = 1,98; P < 0,195). Mandibelaktiviteten var den parameter, der ændrede sig mest over tiden (F6,804 = 244; P < 0,0001), mens aktiviteten af bagkloen ændrede sig mindst (F6,804 = 5,09; P < 0,0001). Hjerteaktiviteten udviste størst interaktion mellem azoxystrobinbehandling og tid (F18,804 = 91,6; P < 0,0001), men interaktionen for de 3 andre responsparametre var også signifikante (tabel 12). Lineær kontrastanalyse mellem aktiviteten af parametrene til tidspunkter svarende til før og efter azoxystrobintilsætning viste, at aktiviteten af hjertet og mandibler hos kontroldafnierne ikke var konstant over forsøgsperioden (hjerte: F1,203 = 10,6; P < 0,0013, mandibler: F1,203 = 13,2; P < 0,0003). Forskellene i aktivitet før og efter azoxystrobintilsætning var dog langt mere udtalte hos dafnier eksponeret for azoxystrobin med undtagelse af aktiviteten af bagkloen (tabel 12). Aktiviteten af hjertet, brystlemmer og mandibler blev signifikant (P < 0,0001) reduceret ved en azoxystrobinkoncentration på 0,5 mg l-l, hvorimod aktiviteten af bagkloen kun blev påvirket ved en koncentration på 2 mg azoxystrobin l-l, og effekten sås kun i slutningen af forsøget (tabel 12). Ved sammenligning til forsøget med cypermethrin var der i azoxystrobinforsøget færre signifikante forskelle mellem kontrollen og de 3 øvrige behandlinger, men flere signifikante forskelle mellem aktiviteten før og efter pesticidtilsætning. Det skal bemærkes, at teststyrken for forskelle mellem behandlinger er lavere i azoxystrobinforsøget p.g.a. en mindre prøvestørrelse (3 dafnier) i hver behandling. Det modsatte er gældende for tests vedrørende tidseffekter, fordi der i azoxystrobinforsøget blev udført flere observationer (10 replikater) på hver dafnie pr. observationsperiode. 3.4 Svømmeadfærd3.4.1 CypermethrinDe indledende forsøg (se bilag 6.3) viste tydeligt, at kun D. magna indsamlet fra en naturlig population umiddelbart før forsøgene blev udført, kunne foretage en reproducerbar retningsbestemt vandring med lys som stimulus. I samtlige forsøg blev der derfor anvendt en population af D. magna fra en mindre sø (Tabel 2). Til trods for en udtalt evne til at foretage vandringer viste der sig at være en vis individuel variation i responset. Dette blev imødegået ved dels at anvende samme forsøgsdyr til kontrol- og eksponeringsforsøg og dels ved at replikere alle forsøg 5 gange. Dafniernes gennemsnitsstørrelse var 2,1-2,5 mm. Den gennemsnitlige maksimale svømmehastighed i kontrolforsøg var 0,81-0,91 m min-1 (tabel 13) med et gennemsnit på 0,85 m min-1, og der var ikke signifikante forskelle (ANOVA, p>0,05) mellem de enkelte forsøg. Ved de fem testede koncentrationer af stoffet (0,05 til 1,0 µg cypermethrin l-1) blev der observeret en reduktion i den maksimale svømmehastighed for D. magna (tabel 13). De gennemsnitlige værdier var alle indbyrdes signifikant forskellige, og alle på nær værdien for 0,05 µg cypermethrin l-1 var signifikant forskellige fra værdierne fra det respektive kontrolforsøg (ANOVA, p <0,05). TABEL 13. Et eksempel på resultaterne af et sammenhørende kontrol- og eksponeringsforsøg med en cypermethrinkoncentration på 0,1 µg l-1 er vist i figur 30. Den relative reduktion i den maksimale svømmehastighed er 40 % i eksemplet. Det kumulerede antal forsøgsdyr over tiden viste, at de eksponerede forsøgsdyr forblev langsommere end forsøgsdyrene i kontrolforsøgene (figur 31).
FIGUR 30.
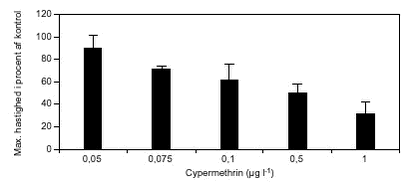
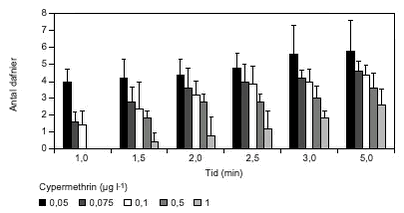
FIGUR 31. Den maksimale svømmehastighed i procent af kontrol faldt med stigende koncentrationer af cypermethrin (figur 32). Det kumulerede antal forsøgsdyr i hhv. kontrol- og eksponeringsforsøg afspejlede også denne sammenhæng (figur 33).
FIGUR 32.
FIGUR 33. Som omtalt i afsnit 2.7.4 og bilag 6.3 er det ikke muligt at beregne en egentlig gennemsnitlig svømmehastighed baseret på hele observationsperioden ud fra den anvendte forsøgsopstilling. Men, da ca. halvdelen af forsøgsdyrene i kontrolforsøgene (tabel 14) blev registreret entydigt (dvs. ingen af dyrene svømmede i den modsatte retning i svømmekammeret) ved registreringspunktet efter 1,5 min (= 0,38 m min-1), anvendes dette som en gennemsnitlig minimumssvømmehastighed. TABEL 14.
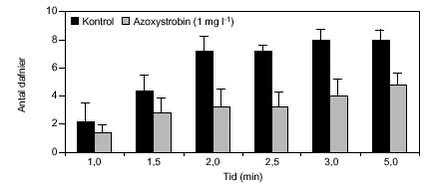
3.4.2 AzoxystrobinDer blev udført hhv. et pilot- og et fuldt eksperiment med azoxystrobin, for at undersøge, om D. magna også udviste en negativ respons i svømmeaktivitet over for dette pesticid. Den anvendte pesticidkoncentration var 1,0 mg l-1. Den gennemsnitlige maksimale svømmehastighed i kontrolforsøget var ikke signifikant forskellig fra værdierne opnået i forsøg med cypermethrin (ANOVA, p<0,05; tabel 15). Der kunne konstateres en reduktion i den maksimale svømmehastighed for D. magna (tabel 16), som var signifikant forskellig fra kontrolværdien, men på et lavt signifikansniveau (ANOVA, p=0,09). Reduktionen i den maksimale svømmehastighed var ca. 14%. TABEL 15. Det kumulerede antal forsøgsdyr over tiden viste, at de eksponerede forsøgsdyr forblev langsommere end forsøgsdyrene i kontrolforsøgene (figur 34). Andelen af eksponerede forsøgsdyr, som opnåede en gennemsnitlig min. svømmehastighed på 0,38 m min-1 var signifikant lavere (ANOVA, p<0,05) end den tilsvarende andel af kontroldyrene (tabel 16).
FIGUR 34. TABEL 16. 3.5 PesticidkoncentrationerI forsøgsperioden blev der udtaget 10 prøver, som blev analyseret for at sammenligne de nominelle pesticidkoncentrationer med de aktuelle. Resultaterne viste generelt god overensstemmelse med de nominelle koncentrationer (tabel 17). De aktuelle cypermethrinkoncentrationer lå lidt under de nominelle, mens de aktuelle azoxystrobinkoncentrationer lå svagt over i to tilfælde og ellers 20-30 % under de nominelle koncentrationer. TABEL 17.
|