|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 93, 2004 Bæredygtig produktion af småplanter i forstplanteskolerIndholdsfortegnelse
ForordDenne rapport udgør slutrapporten for projektet "Bæredygtig produktion af småplanter i forstplanteskoler" i perioden 2001-2004 under Miljøstyrelsens forskningsprogrammer vedrørende bekæmpelsesmiddelafgifter. Planteskoleejerforeningen støttede ligeledes projektet med midler fra gartneriernes afsætningsudvalg i samarbejde med DEG konsulent Bent Leonhard. Projektet blev startet af Lars Bødker, som var projektleder i 2001-2002, hvorefter John Larsen overtog projektledelsen da Lars Bødker tog orlov. Sabine Ravnskov har gennem hele projektperioden fungeret som daglig leder. Kaare Møller har stået for identifikation af patogener. Keld Mansfeld-Giese forestod de indledende undersøgelser af flere danske forstplanteskoler, hvorfra to blev udvalgt som forsøgsværter. Tina Tønnersen har gennem hele projektperioden udført alle laboratoriemæssige opgaver. Al det praktiske arbejde vedrørende markforsøgene blev udført af Peter Hartvig, Lis Madsen, Jan Schou, Jakob Sørensen og Karen Margrethe Madsen. Særligt takkes alle på lugeholdene, som brugte mange timer på knæ for at fjerne enårig rapgræs på Brøndlundgård, der udover "markgruppen" bestod af Steen Meier, Lone Fink, Karin Thygesen og Tina Tønnersen. Det var hårdt for ryggen, men hyggeligt. Projektet var baseret på markforsøg, som blev udført på to forstplanteskoler: 1) Hedeselskabets planteskole Brøndlundgård ved Vojens i Jylland og 2) Akkerup Planteskole på Fyn. Vi retter en stor tak til planteskolerne for godt samarbejde og forsøgsværtskab. Igennem projektperioden har vi hentet en del hjælp udenfor projektgruppen. Det drejer sig især om Barbara Kilieszewska-Rokicka (Institute of Dendrology, Polish Academy of Sciences) for vejledning og hjælp med analyser vedrørende ektomykorrhiza, Monica Welc (Institute of Dendrology, Polish Academy of Sciences) for arbejde med arbuskulær mykorrhiza i hvidel og Susanne Elmholt (Afdeling for Jordbrugsproduktion og Miljø, DJF) for hjælp med ergosterol analyser. Endelig rettes en stor tak til medlemmer af skiftende følgegrupper for skovprojekter i MST regi, hvor især Inge Knudsen (Institut for Plantebiologi, KVL) og Kirsten Thomsen (Statsskovenes Planteavlsstation, Skov og Naturstyrelsen) takkes for værdifuld sparring og samarbejde. Sammenfatning og konklusionerDer er store problemer med rodsygdomme og ukrudt i danske forstplanteskoler. Disse skadevoldere bekæmpes traditionelt med kemiske jorddesinfektionsmidler (Basamid) og herbicider. Brugen af Basamid kan give utilsigtede sundheds- og miljøpåvirkninger, hvilket giver anledning til, at myndigheder og planteskoleerhvervet efterlyser alternativer. Hovedformålet med nærværende projekt var derfor at udvikle et bæredygtigt alternativt dyrkningssystem i forstplanteskoler uden brug af kemiske jorddesinfektionsmidler og således på baggrund af den indsamlede viden at fremlægge forebyggelses- og bekæmpelsesstrategier for rodsygdomme og ukrudt i danske forstplanteskoler. Som alternative metoder til sanering af ukrudt og rodsygdomme, blev der fokuseret på dampning og brug af grønafgrøder alene og i kombination. Der blev opstillet identiske markforsøg hos to danske forstplanteskoler i udvalgte problemarealer med fire forskellige forstplanter; nordmannsgran (Abies nordmanniana), sitkagran (Picea sitchénsis), nobilis (Abies procera) og hvidel (Alnus incana), som alle har problemer med store udfald ved etablering i frøbede. Forsøgene havde 11 behandlinger: 1) ubehandlet, 2) damp forår, 3) damp efterår, 4) Basamid efterår, 5) Basamid forår, 6) majroe (Brassica rapa var rapa ) som efterafgrøde, 7) havre (Avena sativa) som efterafgrøde, 8) kombination af damp og majroe, 9) kombination af damp og havre, 10) kombination af Basamid og majroe og 11) kombination af Basamid og havre. Hver behandling havde fire gentagelser opstillet med et randomiseret blokdesign. Hver parcel var 4 meter lang og 1 meter bred og opdelt i fire 1 m2 underparceller med hver af de fire forsøgsplanter. Forsøgsarealerne blev etableret i juni 2002 med efterfølgende etablering af efterafgrøder august 2002. Damp- og Basamidbehandlingerne blev foretaget juli 2002 og april 2003. Efterafgrøderne blev nedmuldet april 2003, og en måned efter nedmuldning blev frøene sået og forsøget startet. Forsøget blev afsluttet oktober 2003. Virkningen af omtalte behandlinger på fremspiring af afgrøder og plantevækst, forekomst af rodpatogner og ukrudt, dannelse af ekto- og arbuskulær mykorrhiza og sammensætning af den generelle mikroflora blev undersøgt i dette projekt. Effekten af de forskellige behandlinger på rodsygdomme (målt indirekte med planteparametrene: fremspiring, dødelighed af fremspirede og antal planter høstet) afhang af plantearten. Der var ingen signifikante behandlingseffekter i nordmannsgran, sitkagran og hvidel, men i nobilisgran var der i alle behandlinger på nær efterafgrøder alene en bedre fremspiring og en mindre dødelighed blandt fremspirede planter frem til høst, hvilket resulterede i markant flere høstede planter, især i kombinationsbehandlingerne. Isolering af rodsvampe med efterfølgende patogenitettest viste, at især Fusarium oxysporum, Fusarium avenacium og Pythium ultimum forvolder store skader i de pågældende forstplanteskoler. Sammensætningen af populationer af rodsvampe fra planter med og uden symptomer var dog ikke nævneværdig forskellig. Som forventet var der en markant hæmning af fremspiring af ukrudt med damp og Basamid behandlingerne, mens brug af majroe som efterafgrøde gav en uventet tilsvarende markant hæmning af fremspring af enårig rapgræs, der var den dominerende ukrudtsart på begge planteskoler. Årsagen til den sanerende virkning af majroe som efterafgrøde overfor enårig rapgræs skyldes sandsynligvis udskillelse af allelopatiske forbindelser i forbindelse med nedmuldningen af majroer før såning. Majroe som efterafgrøde hæmmede også fremspiringen af hvidel og sitkagran, men havde ingen virkning på fremspiringen af nobilis og nordmannsgran. Havre som efterafgrøde alene havde ingen virkning på ukrudt og rodsygdomme. Der kunne ikke måles utilsigtede langtidsvirkninger af damp og Basamid på den generelle mikroflora målt med fedtsyrerprofiler, mens nedmuldningen af efterafgrøderne øgede den mikrobielle biomasse i jorden på en af planteskolerne. Der kunne dog måles langtidsvirkninger af damp og Basamid på forekomsten af jordsvampe, idet mængden af svampebiomarkøren 18:2ù6,9 var markant lavere 10 måneder efter damp og Basamid behandling i forhold til ubehandlet kontrol. Derimod var der 14 måneder efter behandling med damp og Basamid ingen effekter på dannelsen af ektomykorrhiza i nordmannsgran og arbuskulær mykorrhiza i hvidel. Ud fra de opnåede resultater er det ikke muligt at anvise en generel strategi til forebyggelse/bekæmpelse af rodsygdomme og ukrudt i forstplanteskoler, idet virkningen af de forskellige saneringsmetoder synes at være planteafhængig. Resultaterne fra dette projekt kan bruges som et første skridt i retningen af udvikling af alternativer til kemisk jorddesinfektion. Dampning ser ud til at kunne erstatte anvendelsen af Basamid til en generel sanering af ukrudt og rodsygdomme. Angående ukrudtsbekæmpelse ser brug af majroer meget lovende ud, men der er behov for yderligere forskning til at klarlægge mulighederne for at anvende majroer til biosanering af ukrudt og rodsygdomme. Summary and conclusionsRoot diseases and weeds are causing severe problems in forest nurseries. Control of these plagues is traditionally achieved by using chemical soil disinfectants (Basamid) and herbicides. However, due to heath issues and environmental concern there is a demand for alternatives. Therefore, the main objective of the present project was to develop a sustainable, alternative growth system in forest nurseries without use of chemical soil disinfectants and, based on this knowledge, to put forward prevention and control strategies for root diseases and weeds in Danish forest nurseries. As alternative methods to root diseases and weeds, we focused on seedbed steaming and green manure crops alone and in combination. Field experiments were conducted at two Danish forest nurseries in selected "problem areas" with four different forest plants; Caucasian Fir (Abies nordmanniana), Noble Fir (Abies procera), Sitka Spruce (Picea sitchénsis) and Grey Alder (Alnus incana), which are all having problems with pre and post emergence damping-off. The experiments had 11 treatments: 1) untreated, 2) steam autumn, 3) Steam spring, 4) Basamid autumn, 5) Basamid spring, 6) Turnip (Brassica rapa var rapa) as green manure, 7) Oat (Avena stiva) as green manure, 8) combination of steam and Turnip, 9) combination of steam and Oat, 10) combination of Basamid and Turnip and 11) combination of Basamid and Oat. Each treatment had four replicates and the experiments had a complete randomised block design. Each plot was four metres long and 1 metre wide with four 1m2-subplots with each of the four experimental plants. The experimental areas were established in June 2002, with subsequent establishment of green manure crops in August 2002. Steam and Basamid treatments were performed in July 2002 and April 2003. The green manure crops were incorporated into the soil in April 2003, and one month later the seeds were sown and the experiment initiated. The experiments were completed in October 2003. In this project we examined the influence of the above-mentioned treatments on plant parameters (emergence, mortality, total number of plants harvested and plant biomass at harvest), presence of weeds and root pathogens, formation of ecto-mycorrhizas and arbuscular mycorrhizas and the composition of soil microbial communities. The effects of the different treatments on root diseases (measured indirectly using the plant parameters: seedling emergence, mortality of emerged seedlings and number of plants harvested) were crop dependent. No significant treatment effects were observed in Caucasian Fir, Sitka Spruce and Grey Alder, but all treatments except green manure alone increased emergence and survival of Noble Fir resulting in significantly more plants harvested, especially in the combination treatments. Isolation of root fungi other than mycorrhizal fungi with subsequent test of pathogenicity showed that especially Fusarium oxysporum, Fusarium avenacium and Pythium ultimum are causing major damage in forest nurseries. Surprisingly, the composition of populations of root fungi other than mycorrhizal fungi did not differ between roots of healthy plants and plants with root rot symptoms. As expected, steam and Basamid markedly suppressed emergence of weeds, while Turnip unexpectedly gave a similar, marked suppression of emergence of Annual Bluegrass, which was the dominating weed in both nurseries. The reason for the suppression of Annual Bluegrass with Turnip as green manure is most likely due to allelopathic compounds released during the decomposition of Turnip prior to sowing. Turnip as a green manure crop also suppressed emergence of Grey Alder and Sitka Spruce but had no effects on Noble Fir and Caucasian Fir. In contrast to Turnip, Oat as a green manure had no effect on weeds and root pathogens. No long-term effects of steam and Basamid on the general soil microbial communities were measured, while incorporation of green manure crops increased the microbial biomass in the soil in one of the forest nurseries with low initial biomass compared with the other nursery where green manure incorporation did not result in an increased microbial biomass. However, 10 months after steam and Basamid treatments the biomass of soil fungi as measured using the signature fatty acid 18:2ù6,9 was still markedly lower than in the untreated control. On the other hand, 14 months after steam and Basamid treatments, formation of arbuscular mycorrhiza in Grey Alder and ecto-mycorrhiza in Caucasian Fir was unaffected by the various treatments. The results from the present project do not allow a general strategy for prevention/control of root diseases and weeds in forest nurseries, as the effects of the tested methods were plant dependent. However, the results can be used as a first step towards development of alternatives to chemical soil disinfection, showing that steaming of seedbeds seems to be as good as Basamid at preventing weeds and fungal root diseases. Turnip as a green manure looks very promising for control of Annual Bluegrass. This should be investigated further. 1 FormålProjektets mål var at medvirke til udvikling af et bæredygtigt dyrkningssystem i forstplanteskoler uden brug af kemiske jorddesinfektionsmidler og således på baggrund af den indsamlede viden at fremlægge forebyggelses- og bekæmpelsesstrategier mod ukrudt og sygdomme til produktion af småplanter i forstplanteskoler. Projektets formål var at bestemme indflydelsen af kemisk, termisk og biologisk jorddesinfektion på:
Den opnåede viden skulle anvendes til at fastlægge behovet for anvendelse af kemiske jorddesinfektionsmidler overfor rodpatogene svampe i forstplanteskoler og konsekvenserne af en reduktion i deres anvendelse. Termisk og kemisk desinfektion har en kraftig reducerende effekt på jordens mikrobielle samfund, herunder også på eventuelle nyttige antagonister og mykorrhizasvampe. Det ønskes undersøgt, om man ved at anvende nedpløjede grønafgrøder kan opnå en tilfredsstillende reduktion i patogenpopulationen uden disse sideeffekter. 2 BaggrundProduktion af småplanter i forstplanteskoler er forbundet med et stort forbrug af pesticider til forebyggelse/bekæmpelse af ukrudt, sygdomme og skadedyr. På grund af uønskede effekter på miljø og gartneriarbejdernes sundhed er der behov for at udvikle alternative forebyggelses- og bekæmpelsesstrategier, der bygger på pesticidfrie metoder. Ved Danmarks JordbrugsForskning er der i 1994 og i 2000 foretaget undersøgelser i fire forstplanteskoler vedrørende rodpatogene svampe i Alnus, Abies og Picea (Bødker & Noyé, 1994; Bødker & Larsen, 2000). Resultatet viste klart, at der er store problemer med høje smittetryk af rodsygdomme i danske planteskoler, som primært synes forårsaget af Fusarium og Pythium. Disse problemer afhjælpes for størstedelens vedkommende ved brug af jorddesinfektionsmidlet Basamid (handelsnavn Basamid granulat 980), som dog primært rettes mod bekæmpelse af ukrudt. En kemisk jorddesinfektion påvirker ikke alene ukrudt og de rodpatogene svampe, som ønskes nedbragt, men kan også have uønskede effekter på andre organismer, herunder antagonister mod rodpatogene svampe og andre organismer med gavnlige funktioner. Således har jorddesinfektionsmidlet Basamid en kraftig toksisk effekt overfor både ukrudt, mikroflora og mikrofauna, og forskydninger i den biologiske ligevægt i jorden kan måles i op til 105 dage efter en kemisk jorddesinfektion (Toyota et al., 1999). Dampning af frøbede er et oplagt alternativ til kemisk jorddesinfektion og vinder mere og mere indpas indenfor frilandsgartneri og er effektiv til bekæmpelse af ukrudt og rodsygdomme (Pinel et al., 1999; Melander et al., 2002). Denne metode er dog ligesom kemisk sanering også forbundet med risiko for hæmning af mikroorganismer, som gavner planters vækst og sundhed som f.eks. mykorrhizasvampe (Soulas et al., 1997) og den generelle mikrobielle aktivitet i jord (Elsgaard, 2003). Anvendelse af biosanering er et andet oplagt alternativ til kemisk jord desinfektion. Der er flere eksempler på biologisk sanering af rodpatogener med nedmuldning af efterafgrøder (Williams Woodward et al., 1997; Bødker & Thorup-Kristensen, 1999; Lazzeri & Manici, 2001). Den sanerende virkning af efterafgrøder som f.eks. Brassica arter beror sandsynligvis på en kombination af udskillelse af glycosinolater og opformering af sygdomshæmmende mikroflora ved omsætningen af grønafgrøden. Glycosinolater er kemiske forbindelser fra Brassica planter, som er toksiske overfor rodpatogene svampe (Sawar et al., 1998; Smolinska et al., 2003). Endvidere kan glycosinolater hæmme frøspiring af f.eks. salatfrø (Brown & Morra, 1996). Glycosinolater er ikke alene toksiske overfor rodpatogener, men også overfor mykorrhizasvampe (Screiner & Koide, 1993) og nitrificerende bakterier (Bending & Lincoln, 2000). Der er ingen publicerede undersøgelser, der sammenligner kemisk og termisk sanering med anvendelse af den biologiske sanering, der sker ved aerob omsætning af organisk materiale fra en grønafgrøde. Ligeledes mangler der viden om kombination af "kemisk/termisk" jord desinfektion med grønafgrøder, som bygger på naturlige reguleringsmekanismer. En sådan kombinationsbehandling kan tænkes anvendt i jord med højt sygdomstryk, hvor jorddesinfektion er tvingende nødvendig. Her kan den naturlige mikroflora reetableres gennem tilførsel af organisk materiale for derved hurtigst muligt at genskabe jordens frugtbarhed. Iværksættelsen af en dyrkningsstrategi, der udnytter biologiske ressourcer kræver derfor indgående kendskab til de sygdomsforårsagende svampe i de enkelte dyrkningssystemer samt til specificiteten i sygdomshæmning af de forskellige organiske materialer. Ved anvendelse af biologiske ressourcer er en totalsanering ud fra en kemisk synsvinkel meget vanskelig at opnå, og selv et halveret smittetryk kan forårsage store skader på planternes rødder. Endvidere skal udplantningsplanter være helt sunde, idet en tilsyneladende sygdomsfri plante med et lille angreb af rodpatogene svampe først vil udvise vækstdepression efter udplantning i skov eller læbælte. Dette er af større betydning end de planter, der kasseres allerede i planteskolen på grund af tydelige symptomer. Derfor bør der ved en eventuel praktisk anvendelse af de biologiske ressourcer tages udgangspunkt i de sygdomsfrie jorder, hvor en forebyggende dyrkningsstrategi fortsat kan bevare sygdomstrykket under skadetærskelen (Bødker & Thorup-Kristensen, 1999). Dette gælder for alle planteproduktionssystemer. Diskussionen vedrørende pesticiders effekt på jordøkosystemers mikrobielle samfund bygger i høj grad på studier i mikrokosmer, hvor det ikke er muligt at vurdere pesticideffekter i forhold til andre miljøpåvirkninger. I studiet af pesticiders sideeffekter er det vigtigt, at den naturlige fluktuation i diversitet og aktivitet af de mikrobielle samfund indgår som reference for, at tolkninger ikke kommer ud af proportion. Det er derfor vigtigt, at resultater opnået gennem laboratorieforsøg valideres med markforsøg, hvor det er muligt, at give et realistisk mål for indvirkningen af både kemisk, termisk og biologisk desinfektion på de mikrobielle samfund under naturlige planteskoleforhold. Det nærværende projekt havde derfor både et praktisk og erkendelsesmæssigt sigte. Sygdomstrykket er meget højt og homogent fordelt i danske planteskoler (Bødker & Larsen, 2000), men der er dog stadig en del uklarhed vedrørende hvilke patogener, der er den primære årsag til udfaldet af småplanter. Projektet startede derfor med en diagnosticering og identifikation af disse patogener. Derefter blev fokus rettet mod forskydninger i jordøkosystemets mikrobielle samfund og den hastighed, hvormed diversiteten og aktiviteten kan genskabes efter brug af forskellige organiske materialer med og uden en forudgående kraftig termisk eller kemisk jorddesinfektion. Ektomykorrhizasvampe findes naturligt i skovøkosystemer, hvor de danner symbiose med trærødder. Desuden danner nogle løvfældende træer som hvidel arbuskulær mykorrhiza. Disse svampe tillægges stor betydning for rodsystemets optagelse af næringsstoffer som f.eks. kvælstof og fosfor og modstandsdygtighed overfor sygdomme (Smith & Read, 1997). Kemisk og termisk jorddesinfektion vil givetvis ikke alene hæmme rodpatogener, men også disse gavnlige svampe. Indenfor de sidste 10 år er nye velegnede metoder til beskrivelse af mikrobielle samfund i jorden blevet tilgængelige. Diversiteten af mikroorganismer kan undersøges på isolatniveau gennem isolering af svampe og bakterier med efterfølgende fænotypisk (BIOLOG, fedtsyrer) og genotypisk (UP-PCR, RAPD, AFLP) karakterisering eller på samfundsniveau gennem analyse af materiale fra mikroorganismerne (DNA, phospholipider, enzymer) ekstraheret fra jorden samt gennem analyse af funktionel diversitet af den ekstraherede bakteriefraktion (BIOLOG-profiler). I dette projekt studerede vi effekten af jorddesinfektion på samfundsniveau, og til det formål valgte vi at anvende fedtsyreprofiler baseret på FAME (Cavigelli et al., 1995). Ved FAME-profilering ekstraheres jordens totale indhold af fedtsyrer og analyseres kvalitativt og kvantitativt ved gaskromatografi. Da en del af fedtsyrerne er specifikke for bestemte grupper af svampe og bakterier, kan resultatet give et mål for den totale svampe- og bakteriebiomasse samt for forholdet mellem Gram negative og Gram positive bakterier. I nærværende projekt blev der anlagt identiske forsøg på to danske forstplanteskoler med forsøgsarealer med høje smittetryk af rodsygdomme med brug af efterafgrøder i kombination med en forudgående damp- eller Basamidbehandling. Majroe, der indeholder fungitoksiske glycosinolater og havre, der ligeledes indeholder fungitoksiske avenaciner, blev anvendt som efterafgrøde. Valget af majroe og havre skyldes, at begge afgrøder i tidligere undersøgelser har vist effekt overfor rodpatogene svampe. For at kunne sammenligne med konventionel planteskolepraksis etableredes endvidere to behandlinger med henholdsvis damp- og Basamidbehandling umiddelbart før såning af kulturafgrøden i foråret. Dampning, Basamid og efterafgrøder blev testet alene og i kombination for deres effekt overfor angreb af rodpatogener, fremspiring af ukrudt og forekomst af mykorrhiza og forskydninger i jordens mikroflora i fire forstplantekulturer. 3 Forsøg3.1 Etablering af markforsøg3.1.1 LokaliseringForsøgene blev anlagt på følgende lokaliteter: Akkerup Planteskole (Akkerupvej 50, 5683 Haarby, UTM koordinater forsøgsmarken, Akkerup Planteskole, mark K7: UTM zone 32, ED50. Northing 6121232, Easting 572906) og Brøndlundgård (Hedeselskabet, Brøndlundsvej 2, 6500 Vojens, UTM koordinater forsøgsmarken, Brøndlundgård, mark 40,1: UTM zone 32, ED50. Northing 6123005, Easting 205979, UTM koordinater ekstra forsøgsmark, Brøndlundgård, mark2:UTM zone 32, ED50. Northing 6122912, Easting 506663. 3.1.2 DesignMarkforsøgene på begge planteskoler blev anlagt med parceller på 4 x 1 m med et randomiseret blok design med følgende behandlinger: 1. Ubehandlet, 2. Dampbehandling efterår, 3. Dampbehandling forår, 4. Basamid efterår 5. Basamid forår, 6. Majroer udlagt som efterafgrøde, 7. Havre udlagt som efterafgrøde, 8. Dampbehandling efterår + majroe udlagt som efterafgrøde, 9. Dampbehandling efterår + havre udlagt som efterafgrøde, 10. Basamid efterår + majroe udlagt som efterafgrøde, 11. Basamid efterår + havre udlagt som efterafgrøde. Figur 1. Design af markforsøgene på Brøndlundgård og Akkerup. Hver parcel er angivet med behandlingsnummer og tilhørende farvekode. Sideløbende blev der anlagt et ekstra forsøg på Hedeselskabets Planteskole mark 2, der havde til formål at undersøge flere forskellige efterafgrøders virkning på planternes sundhed. Forsøget blev ligeledes anlagt som et randomiseret blok design med fire blokke af hver af følgende behandlinger: 1. Ubehandlet, 2. Basamid forår, 3. Majroer udlagt som efterafgrøder, 4. Havre udlagt som efterafgrøder, 5. Gul sennep udlagt som efterafgrøder, 6. Olieræddiker udlagt som efterafgrøder. 3.1.3 Damp og Basamid behandlingerBasamid blev afvejet (80 g) pr. parcel i laboratoriet og udbragt manuelt efter tilladelse fra Miljøstyrelsen. Doseringen af Basamid var baseret på anbefalet dosering fra producenten til jorddesinfektion og almindelig gartnerpraksis. Basamid blev opblandet i det øverste 5 cm jordlag med 3 gange harvning med traktorrive. Efter udbringning, blev parcellerne dækket med plast i 10 dage. Dampbehandlede parceller blev dampet i 5 min., og jordtemperaturen nåede op på 80-85C i 5 cm dybde. 3.1.4 Efterafgrøde behandlingerEfterafgrøderne blev ligeledes afvejet pr. parcel (majroer 9 g, havre 80 g, gul sennep 3,2 g og olieræddike 3 g). Tilvæksten af efterafgrøderne blev visuelt bedømt ca. 4 og 8 uger efter såning på en skala fra 0 til 100; 0=ingen tilvækst, 100=100 % dækning. Majroerne på Akkerup Planteskole visnede pludselig ned før optimal vækst af en uvis grund, måske som følge af insektangreb. Vi prøvede at redde det med en eftersåning i september, men desværre uden held. Ved nedmuldning af efterafgrøderne blev alle parceller kørt igennem med bedopsætter i 3-5 cm jorddybde. 3.1.5 Gødskning og vandingGødskning og vanding er udført af planteskolerne efter almindelig praksis. Forsøgene blev renholdt manuelt hver 3-4 uge gennem hele vækstsæsonen. 3.1.6 Frømateriale af testplanterHver parcel i alle tre forsøg var 4 m2 med 1 m2 af hver af plantearterne: nordmannsgran (2000 frø/m2), nobilisgran (800 frø/m2), sitkagran (900 frø/m2) og hvidel (800 frø/m2). Nobilis og nordmannsgran blev leveret stratificeret efter almindelig praksis i 6 uger ved passende luftfugtighed og ventilation. I nordmannsgran blev der foretaget undersøgelse af frøbårne svampe ved Inge Knudsen (KVL), som kun fandt 1 % forekomst af potentielle patogener. I de tre øvrige plantearter blev frøene ikke testet for frøbårne sygdomme. Ved såning blev frøene bredsået og dækket med kalkfrit sand.
Figur 2. forbehandlinger i markforsøgene med Basamid (A og B), dampning C og D) og efterafgrøder (E). Billede F er en oversigt fra forsøg på Brøndlundgård maj 2003.
Figur 3. Overdækning af frøbede med sand på Akkerup Planteskole maj 2003. Forløb
4 Analyser
4.1 Planteparametre4.1.1 Fremspiring, dødelighed, antal høstede planter og udbytteI hver parcel indenfor hver planteart blev en cirkel (0,25 m2) lagt i midten af afgrøden, og alle plantemålinger blev foretaget i dette fastlagte areal. Seks uger efter såning, blev der målt fremspiring af frø. Herefter blev planter med symptomer på rodråd registreret 10 og 14 uger efter såning og endelig 28 uger efter såning blev alle planter indenfor cirklen nænsomt gravet op, og planterne blev bragt til laboratoriet, rødderne vasket fri for jord og planterne talt og vejet. Grundet problemer med overslæb af frø mellem parceller ved etablering af forsøget på Akkerup Planteskole er plantedata for Akkerup ikke pålidelige og derfor ikke medtaget i denne rapport. 4.2 Registrering af ukrudtFremspiring af ukrudt blev registreret ved at kaste en cirkel (50 cm i diameter) tre tilfældige steder i hver parcel og indenfor cirklen blev art, antal og vægt af alle fremspirede planter registreret ca. en måned efter forsøgene blev anlagt. 4.3 Forekomst af rodpatogener4.3.1 JordtestTesten havde til formål at undersøge sygdomstrykket i jorden på nordmannsgran, sitkagran, nobilisgran og hvidel. Der blev anvendt frø fra samme frøparti som i markforsøgene. Testen blev udført som et fuldfaktor randomiseret blokdesign med tolv gentagelser i drivhus. Planterne blev dyrket i 2 kg potter i den rå jord indhentet fra planteskolerne umiddelbart før start af markforsøg. Inden parcelafsætning i markforsøgene blev 10 spadestik jord tilfældigt fra hver af de 4 blokke udtaget. Jorden fra hver enkelt blok blev blandet og anvendt til jordtest. Femten frø blev lagt på overfladen af den rå jord i hver potte og derefter dækket med 100 g kalkfrit sand. Potterne blev dækket med plastik indtil fremspiring. Antallet af spirede og døde planter blev optalt løbende. 4.3.2 Isolering og identifikation af svampe fra markforsøgeneFra hver parcel i begge planteskoler blev der indhentet 10 syge og 5 sunde planter af hver planteart. Rødderne fra den enkelte plante blev vasket og klippet i 1 cm stykker. Fire rodstykker fra hver plante blev overfladesteriliseret i 70 % ethanol, skyllet 3x i dobbeltdestilleret autoklaveret vand og lagt på Potato Dextrose Agar (PDA) med novobiocin (25 mg l-1) for detektion af svampe. Efter fremvækst blev svampene inddelt og optalt i grupper efter visuel bedømmelse (vækst, struktur og farve) og repræsentanter fra de enkelte svampe rendyrket på PDA. Pythium lignende svampe dog på P10VP (Corn meal agar med brassicol 100 mg l-1, ampicillin 250 mg l-1, pimaricin mg l-1og rifampicin 10 mg l-1). Derefter blev svampene identificeret i mikroskopet ved hjælp af følgende værker: Barron, 1968; Booth, 1971; Dick, 1990; Domsch et al., 1980a; Domsch et al., 1980b; Ellis, 1971; Sutton, 1980; von Arx, 1970. 4.3.3 PatogenitetstestDe isolerede svampe fra de to planteskoler blev dyrket op på PDA (Pythium på P10VP) og to tæt bevoksede plader af hver svamp blev anvendt i patogenitetstesten. Indholdet på de to plader blev blendet med 100 ml destilleret sterilt vand og blandet i 2 kg steril (autoklaveret) jord:sand (1:2) blanding. Fem forspirede frø af nordmannsgran (samme frøparti som blev anvendt i markforsøgene) blev lagt på 200 g af den inokulerede jord:sand blanding i en potte (n=8) og dækket med 20 g kalkfrit sand. En kontrolbehandling uden svamp blev inkluderet i forsøget. Planterne blev dyrket i væksthus med dag/nat på 16/8 timer og temperatur 20/18°C. Fremspiring af planter blev registreret efter 3 uger, og antallet af fremspirede men døde planter blev registreret efter 3, 4 og 6 uger. Fire uger efter såning blev fire syge planter fra hvert svampeisolat udtaget og rodstykker overfladesteriliseret og lagt på PDA (P10VP), som beskrevet under isolering af svampe for reisolering og identifikation af svampene. 4.4 Undersøgelse af mykorrhiza4.4.1 EktomykorrhizaFor at få indblik i mykorrhizafloraen på de to planteskoler blev der før forsøgsstart indhentet og analyseret planter (Picea abies (20 stk.), Abies alba (20 stk.), Fagus sylvatica (2 stk.)) fra Akkerup Planteskole. Rødderne blev vasket og undersøgt for tilstedeværelsen af ektomykorrhiza ved hjælp af lysmikroskopi. De blev inddelt på basis af morfotyper (farve og tekstur, samt tilstedeværelse af eksternt mycelie på svampekappen). Fra udvalgte rodspidser med mykorrhiza blev tynde snit mikroskoperet med henblik på at artsbestemme svampene mikroskopisk. Identifikation ved hjælp af Ingleby et al. (1990). Ved høst blev rødder af nordmannsgran fra alle 11 behandlinger klippet i 1-2 cm stykker og undersøgt for forekomst af ektomykorrhiza ved hjælp af lysmikroskopi. Procent rodspidser med mykorrhiza blev målt ved at mikroskopere 200 tilfældige rodspidser og beregne andelen af rodspidser med mykorrhiza. Desuden blev intensiteten af mykorrhiza koloniseringen målt med svampemarkøren ergosterol. Til dette formål blev der udtaget 5 g delprøve, som blev frysetørret og formalet med kuglekværn. 25 mg prøve blev udtaget og ekstraheret og målt for ergosterol som ifølge Nylund & Wallander (1991). 4.4.2 Arbuskulær mykorrhizaTi tilfældige hvidel planter blev udtaget fra de ubehandlede parceller i begge planteskoler og i alle parceller i Akkerup Planteskole. Rødderne blev vasket under rindende vand og klippet i 1 cm stykker. Rodstykkerne blev klaret først i 3 timer i 10 % KOH i vandbad ved 90 °C. KOH 10 % blev skiftet efter 1 time. Dernæst blev rødderne bleget i en 25 % NH4OH opløsning og 10 % H2O2 i vand i 2 timer ved stuetemperatur. NH4OH og H2O2 opløsningen blev skiftet efter en time. Derefter blev rodprøverne yderligere klaret i 10 % H2O2 i 1½ time, hvorefter de blevet farvet med 5 % trypanblå i 5 min. ved 90 °C. Rodstykker fra samme parcel blev grundigt blandet, tre delprøver udtaget og lagt på objektglas til mikroskopi. Præparaterne blev mikroskoperet ved 100x forstørrelse, og ± mykorrhiza blev registreret i 100 tilfældige punkter i roden. Koloniseringsprocenten for den enkelte parcel er et gennemsnit af registreringer af de tre præparater og for den enkelte behandling gennemsnittet af de fire parceller. 4.5 Fedtsyreanalyser til karakterisering af jordmikrofloraenMetoden som blev anvendt er baseret på ekstraktion af den totale mængde af fedtsyrer fra jordprøven. Disse fedtsyrer findes i alle levende organismer; primært som delkomponenter i membranlipider og i lipider til oplagsnæring. Der findes mange forskellige fedtsyrer, der varierer i længde (antal C atomer), forgreninger i C kæden, mætning (med og uden dobbeltbindinger) og tilknytning af hydroxy- og methylgrupper. På baggrund af viden om fedtsyresammensætninger hos forskellige mikroorganismer (se tabel 1) giver sammensætningen og koncentrationen af fedtsyrer i jorden både kvalitativ og kvantitativ information om mikrobielle samfund i jord. Metoden anvendes primært til identifikation af bakterier, men kan også anvendes som i dette projekt til beskrivelse af mikrobielle samfund i jord (Cavigelli et al., 1995). Metoden er baseret på frigørelse af fedtsyrer fra jordprøven med en stærk base kombineret med varme, hvor fedtsyrerne bliver spaltet fra deres lipider og omdannes til deres natrium-salt. Herefter methyleres fedtsyrerne til fedtsyremethylester, som forøger flygtigheden af fedtsyrerne til GC-analyse. Efter ekstraktion og basevask er prøverne klar til GC-kørsel. Gaskromatografen er fuldautomatisk og kan opbevare op til 99 prøver ad gangen. Hver prøve tager ca. 25 min. at analysere. 2 μl prøve injiceres, den fordampes, og dampen bæres gennem en 0,2 mm tyk og 25 m lang silica belagt kolonne med et flow af hydrogen gas. Kortkædede fedtsyrer vil passere kolonnen hurtigere end langkædede fedtsyrer. Hydroxylerede fedtsyrer er langsommere end cyclopropylerede eller forgrenede fedtsyrer osv. Så en specifik fedtsyre vil nå gennem kolonnen til detektoren på en specifik retentionstid afhængig af de fysiske egenskaber, og computeren vil via softwareprogrammet MIDI navngive den specifikke fedtsyre og måle intensiteten af signalet for kvantificering. Til kvantificering af de enkelte fedtsyrer er der indlagt en intern standard, som ikke forekommer naturligt. Tabel 1. Oversigt over udvalgte fedtsyresignaturer.
4.6 Statistiske analyserAlle forsøgsresultater blev underkastet envejs variansanalyse med behandlinger som faktor. Forskelle mellem behandlinger blev testet med LSD0.05 , såfremt variansanalysen gav signifikant behandlingseffekt. Inden variansanalyse blev data testet for normalfordeling og varianshomogenitet. Programmet StatGraphics Plus blev anvendt til de statistiske analyser. 5 Resultater
5.1 Plantedata5.1.1 FremspiringPå Brøndlundgård (Hedeselskabets planteskole) var der en signifikant højere fremspiring af nobilisgran i behandlinger med damp efterår og forår samt i kombinationsbehandlingen damp/havre. Basamid forår og begge kombinationsbehandlinger med Basamid fremmede ligeledes spiringsprocenten af nobilisgran (tabel 2). Der var ingen signifikante effekter af behandlingerne på nordmannsgran og sitkagran, men der var dog en tendens til, at majroer hæmmede fremspiringen af disse to plantearter (tabel 2). I ekstraforsøget med flere efterafgrøder var fremspiringen i nobilisgran, nordmannsgran, sitkagran og hvidel henholdsvis 44 %, 31 %, 72 % og 42 % i de ubehandlede parceller, og der var ingen behandlingseffekter. Tabel 2. Fremspiring af småplanter på Brøndlundgård i procent af antal frø sået. Forskellige bogstaver efter tal i samme kolonne betyder, der er signifikant forskel mellem behandlingerne LSD0.05 .
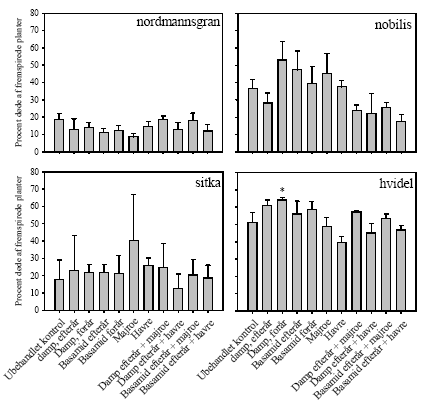
5.1.2 Udfald (andel døde planter)Damp forår forårsagede flere døde hvidel end i de ubehandlede parceller (figur 4). Ellers var der ingen signifikante behandlingseffekter i forhold til ubehandlet kontrol i de øvrige tre plantearter.
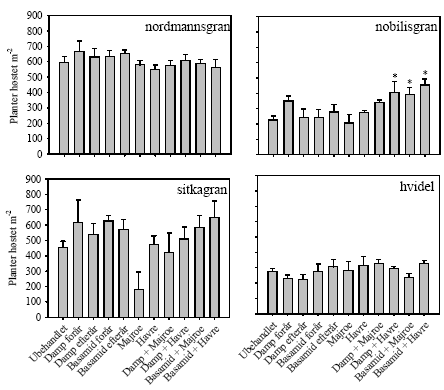
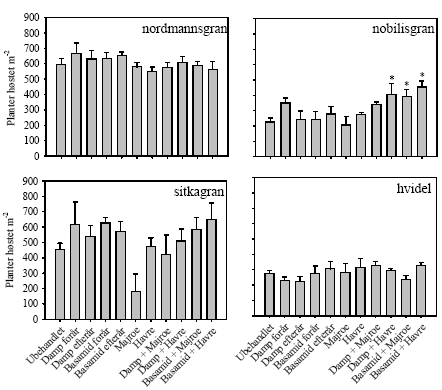
Figur 4. Procent døde planter på Brøndlundgård i parceller med fire forskellige planterarter (nordmannsgran, nobilisgran, sitkagran, hvidel) dyrket i ubehandlet jord og i jord med forskellige metoder til sanering af ukrudt og rodsygdomme. * angiver siginifikant forskel til ubehandlet kontrol baseret på LSD0.05 5.1.3 UdbytteAntallet af nobilisgran høstet i kombintionsbehandlingerne med damp/Basamid og grønafgrøder var højere end i ubehandlet kontrol på nær damp og majroe (figur 5), men biomassen af nobilisgran var upåvirket af de forskellige behandlinger (figur 6). Der var ingen signifikante behandlingseffekter på hverken antal høstede planter eller deres biomasse hos de tre øvrige plantearter (figur 5 og figur 6).
Figur 5. Antal planter høstet pr. m-2 af fire plantearter (nordmannsgran, nobilisgran, sitkagran, hvidel) på Brøndlundgård i ubehandlet jord og i jord med forskellige metoder til sanering af ukrudt og rod-sygdomme. * angiver signifikant forskel til ubehandlet kontrol baseret på LSD0.05.
Figur 6. Tørvægt af planter ved høst af fire forskellige plantearter (nordmannsgran, nobilisgran, sitkagran og hvidel) dyrket på Brøndlundgård i ubehandlet jord og i jord behandlet med forskellige metoder til sanering af ukrudt og rodsygdomme. Der var ingen signifikante forskelle baseret på envejs variansanalyse. 5.2 Forekomst af ukrudtDe to planteskoler var meget forskellige med hensyn til forekomst af ukrudt, der var mere ukrudt på Brøndlundgård (tabel 3) end i Akkerup Planteskole (tabel 4), og ukrudtsfloraen på Brøndlundgård var helt domineret af enårig rapgræs. Generelt hæmmede alle behandlinger enårig rapgræs, på nær behandlinger med havre, (tabel 3 og tabel 4). Endvidere var der betydeligt mere tokimbladet ukrudt i behandlingen med havre som efterafgrøde på Brøndlundgård. Tabel 3. Ukrudt antal og vægt (g) registreret på Brøndlundgård juni 2003. Forskellige bogstaver efter tal i samme kolonne betyder, der er signifikant forskel mellem behandlingerne LSD0.05 (n=4). I søjler uden bogstaver var der ingen signifikante forskelle mellem behandlingerne.
Tabel 4. Ukrudt antal og vægt (g) registreret i Akkerup Planteskole juni 2003. Forskellige bogstaver efter tal i samme søjle betyder, der er signifikant forskel mellem behandlingerne LSD0.05 (n=4). I søjler uden bogstaver var der ingen signifikante forskelle mellem behandlingerne.
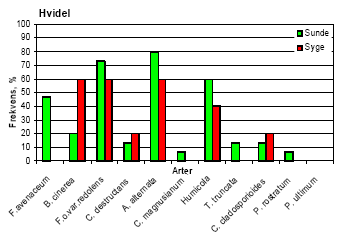
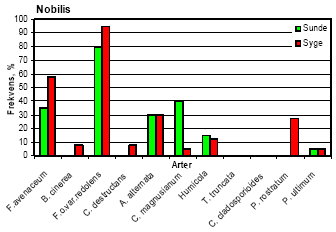
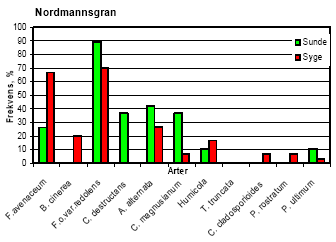
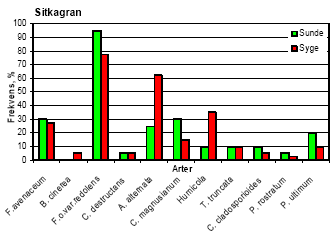
I Akkerup Planteskole var ukrudtstrykket meget lavt, og der kunne ikke måles signifikante behandlingseffekter på enårig rapgræs og hyrdetaske. I behandlingerne med havre var der signifikant mere tokimbladet ukrudt end i nogen anden behandling (tabel 4). 5.3 Forekomst af rodpatogener5.3.1 Jordtest af forsøgsjorder inden forsøgsstartResultaterne fra de indledende jordtest viste, at begge Abies arter både spirede og overlevede meget dårligt i begge planteskolejorde, især i Akkerup jord, hvor kun 24 % af de 42 % fremspirede nobilisgran overlevede (10% af de udsåede frø). Hvidel overlevede langt bedre, især i Akkerup jord, hvor kun få planter døde. Der blev i alt isoleret 110 svampe, som var fordelt på 43 Pythium og resten med repræsentanter fra primært Fusarium og Rhizoctonia. Phytophthora blev ikke fundet. 5.3.2 Isolering af svampe fra rødderUndersøgelse af identitet og frekvens af svampe på syge og sunde planter underbyggede resultatet af de indledende jordtests for forekomst af rodpatogener. Således var frekvensen af Fusarium oxysporum var. redolens særdeles høj - fra 60 % til 95 % på såvel syge som sunde planter af alle fire træarter i Akkerup. I Akkerup fandtes endvidere en relativt høj forekomst af Fusarium avenaceum (26-67 % uafhængigt af træart), mens denne art ikke forekom i Brøndlundgård. Også Pythium-arter blev fundet i planter fra begge planteskoler: P. ultimum (eventuel varietet ikke bestemt) og P. rostratum i Akkerup, og P. ultimum var. sporangiferum og P. periplocum i Brøndlundgård. Der blev fundet en kompleks sammensætning af rodassocierede svampearter, hvoraf flere kan findes beskrevet i litteraturen som medvirkende til problemer med før og efter plantefremspiring i forstplanteskolekulturer af både nåle- og løvtræer. Samlet blev der fra overfladesteriliserede rødder af småplanter af de fire træarter isoleret i alt 20 svampearter. Der fandtes således 15 arter på planter fra Akkerup Planteskole, men kun 10 på planter fra Brøndlundgård (se tabel 5). Fem arter forekom på planter fra begge planteskoler (Cylindrocarpon destructans, Fusarium oxysporum var. redolens, Humicola sp., Phoma glomerata og Pythium ultimum (Brøndlundgård: var. sporangiferum, varietet i Akkerup ikke afklaret)). I smitteforsøg blev der påvist yderligere 12 arter, der ikke var fundet i mykofloraen fra planteskolerne. Eftersom disse arter ikke indgik som inokulum i smitteforsøgene, og eftersom pladespredning fra det benyttede vækstmedie kun viste forekomst af to Penicillium spp. samt af Peziza ostrachoderma (data bringes ikke her), må det antages, at i hvert fald 9 af disse 12 arter var frøbårne (tabel 5), når der tages forbehold for Penicillium spp., Peziza ostrachoderma samt for fund af Alternaria alternata, der må forventes at kunne forekomme som almindelige luftbårne forureninger i et væksthusmiljø. Seks af de svampearter, der kunne isoleres fra Akkerup, regnes for patogene i forstplanteskolekulturer (Fusarium oxysporum var. redolens, F. avenaceum, Pythium ultimum, Botrytis cinerea, Cylindrocarpon destructans og Alternaria alternata). Af disse forekom kun F. oxysporum var. redolens, Pythium ultimum og Cylindrocarpon destructans i Brøndlundgård. Yderligere fandtes der formodet frøbårne forekomster af de nævnte Fusarium-arter, af Alternaria alternata samt af F. culmorum og F. oxysporum var. oxysporum, der ligeledes regnes som patogene i skovplantekulturer (tabel 5). Et overblik over frekvensfordelingen af svampearter på syge henholdsvis symptomfri planter fra Akkerup giver ikke et entydigt billede af de nævnte arter som konsekvent forbundne med syge planter. Således forekom eksempelvis Fusarium oxysporum var. redolens i nogle tilfælde hyppigere på symptomfri planter end på syge nordmannsgran, hvidel og sitkagran, men var hyppigst på syge planter af nobilisgran (figur 7). Frekvensen af F. avenaceum var markant højere i syge nobilis- og nordmannsgran end i symptomfri, mens arten forekom ret ligeligt fordelt i syge og symptomfri planter af sitkagran. I hvidel forekom denne Fusarium art kun i symptomfri planter. Botrytis cinerea forekom på 60 % af de syge planter af hvidel, men kun på 20 % af de symptomfri planter, og fandtes i øvrigt udelukkende på syge planter af de tre nåletræsarter, hyppigst på nordmannsgran (20 %, figur 7). Pythium ultimum forekom i lave frekvenser på både syge og symptomfri planter i alle kulturer. Tabel 5. Opgørelse over svampearter associeret med småplanter af hvidel, nobilisgran, nordmannsgran og sitkagran fra Akkerup og Brøndlundgård planteskoler samt med frø af nordmannsgran. Kode i parentes angiver isolatnavne.
*1 Der forekom flere Penicillium spp. i såvel vækstmediet (to arter) som i frøpartiet af Abies nordmanniana, hvorfor disse forekomster kan stamme fra såvel frøet som mediet. Alternaria alternata, Penicillium spp. og Peziza ostrachoderma kan desuden stamme fra kontaminering fra luften.
Figur 7. Frekvens af de vigtigste svampearter på rødder af syge og sunde småplanter af hvidel, nobilis, nordmannsgran og sitkagran, udtaget fra Akkerup Planteskole. 5.4 PatogenitetstestDa identifikation af de isolater, der var udvalgt til patogenitetstests ikke var afsluttet inden testene blev indledt, indgår samme svampearter som inokulum i mere end én test i flere tilfælde. Identifikationer viste endvidere, at der forekom blandings-inokulae i fire af testene. Tabel 6. Fremspiringsprocent samt procent overlevende planter (nordmannsgran) fire uger efter såning af forspirede frø i steril jord inokuleret med svampe isoleret fra planter dyrket på Akkerup Planteskole. Tallene i parentes angiver SE værdier n=8. Tal fremhævet med fed er signifikant forskellige fra kontrol LSD0.05.
Tabel 7. Fremspiringsprocent samt procent overlevende planter (nordmannsgran) fire uger efter såning af forspirede frø i jorde inokuleret med diverse svampe isoleret fra planter dyrket på Hedeselskabets Planteskole. Tallene i parentes angiver SE værdier n=8. Tal fremhævet med fed er signifikant forskellige fra kontrol LSD0.05.
Fusarium oxysporum var redolens
Cylindrocarpon destructans
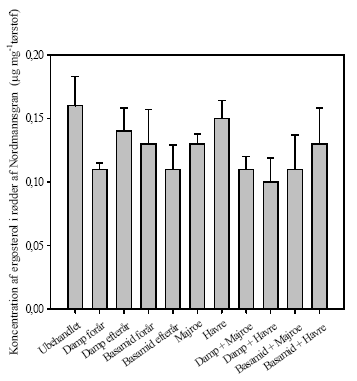
Sordaria fimicola Figur 8. Eksempler på svampe isoleret fra rødder i kulturer fra Brøndlundgård. 5.5 Mykorrhiza5.5.1 Ektomykorrhiza forundersøgelserResultaterne fra Akkerup Planteskole viste, at rødgran (Picea abies) (2/0) var 100 % koloniseret med ektomykorrhiza. Kun en morfotype (Humaria hemispherica (Ascomycotina)) i forskellige udviklingsstadier blev fundet. Denne morfotype er en af de mest almindelige ektomykorrhizasvampe på rødgran, skovfyr (Pinus sylvestris) og lærk (Larix decidua). Ædelgran (Abies alba) (2/0) var ligeledes koloniseret 100 %, dog også kun med en enkelt art i flere udviklingsstadier. Svampen er uidentificeret. Bøg (Fagus sylvatica) var også 100 % koloniseret, men med to forskellige uidentificerede morfotyper. I Hedeselskabet var nordmannsgran (2/1) 95 % koloniseret med en enkelt uidentificeret morfotype, sitkagran (2/1) var 80 % koloniseret med tre morphotyper ITE 1 (2 %), ITE 2 (1 %) og ITE 2 (77 %) og rødel (Alnus glutionosa) (1/1) 50 % med en uidentificeret morfotype. Rødel var også koloniseret med arbuskulær mykorrhiza og Frankia. 5.5.2 Forekomst af ektomykorrhiza i nordmannsgran ved høstRødder af nordmannsgran var 90-100 % koloniseret med en morfotype, og der var ingen synlige forskelle mellem behandlinger. Målinger af ergosterol i rødder af nordmannsgran gav heller ingen signifikante behandlingseffekter (figur 9).
Figur 9. Indhold af ergosterol i rødder af nordmannsgran fra planter dyrket i ubehandlet jord og i jord med forskellige metoder til sanering af ukrudt og rodsygdomme. Ingen signifikante forskelle baseret på envejs variansanalyse.
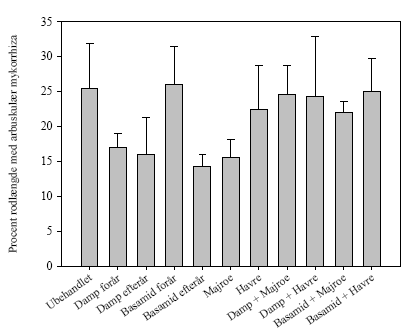
Figur 10. Ektomykorrhiza i nordmannsgran fra Brøndlundgård. 5.5.3 Arbuskulær mykorrhizaI Akkerup Planteskole var der arbuskulær mykorrhiza i hvidel i alle behandlinger (figur 11). Der var ingen signifikante behandlingseffekter (figur 11). Der var ikke arbuskulær mykorrhiza i hvidel i Brøndlundgård.
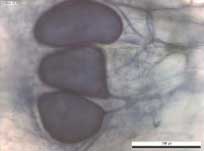
Figur 11. Arbuskulær mykorrhiza i hvidel rødder fra Akkerup Planteskole af planter dyrket i ubehandlet jord og i jord behandlet med forskellige typer af desinfektion. Ingen signifikante forskelle baseret på envejs variansanalyse. A B C Figur 12. Arbuskulær mykorrhiza i hvidel a). arbuskler og b og c) vesikler. 5.6 Den generelle mikroflora analyseret med fedtsyrer5.6.1 Forundersøgelser af mikrofloraen i forsøgsarealerne med fedtsyreanalyserDe indledende analyser af fedtsyresammensætningen i de enkelte parceller i de to planteskoler viste, at der ikke var signifikant forskel på forekomsten af de dominerende fedtsyrer mellem behandlingerne i den enkelte planteskole, hvilket betyder, at det mikrobielle miljø er ensartet i forsøgsmarkerne ved forsøgsstart (data ikke vist). 5.6.2 Fedtsyremarkører i forsøgsjord forår 2003Effekten af behandlingerne på kompositionen af det mikrobielle miljø målt ved hjælp af fedtsyresignaturer var forskellig i de to planteskoler (tabel 8 og tabel 9). Forekomsten af fedtsyrer var generelt lavere i Akkerup Planteskole (tabel 8) i forhold til i Brøndlundgård (tabel 9), mens der var flere forskellige fedtsyresignaturer til stede i Akkerup Planteskole sammenlignet med i Brøndlundgård. I Akkerup Planteskole havde damp- og Basamidbehandlinger alene ingen effekt på sammensætningen af det mikrobielle miljø (tabel 8), mens tilsætningen af organisk materiale i form af majroer og især havre overordnet stimulerede væksten af mikroorganismerne især gram positive bakterier (tabel 8). Størst effekt af behandlingerne var i begge kombinationsbehandlinger med efterafgrøder og damp samt i kombinationsbehandlingen af Basamid og havre, hvor forekomsten af fedtsyresignaturer for gram positive bakterier var signifikant højere end i kontrolbehandlingen. Endvidere var fedtsyremarkøren for arbuskulære mykorrhizasvampe (16:15) tilstede i jord fra alle behandlinger og i kombinationsbehandlingen damp/havre signifikant i højere grad end i den ubehandlede kontrol (tabel 8). I Brøndlundgård var der en signifikant lavere forekomst af markøren for svampe (18:26,9) i alle behandlinger undtagen havre og Basamid/havre i sammenligning med kontrol (tabel 10). Der var signifikant højere værdier for enkelte markører for gram positive bakterier (15:0i Basamid/majroe og 17:0i Basamid/havre), men overordnet var der ikke andre forskelle mellem ubehandlet kontrol og de forskellige behandlinger. Indikatoren 16:15 for arbuskulære mykorrhizasvampe var ikke til stede. Tabel 8. Relative værdier (relativ i forhold til intern standard) for flere fedtsyresignaturer i jord fra Akkerup Planteskole med forskellige forbehandlinger. Jordprøver udtaget primo april 2003. Forskellige bogstaver indenfor samme række indikerer signifikant forskel på forekomst af den pågældende fedtsyre i jord med forskellig behandling LSD 0.05 (n=4). Felter fremhævet med gråt indikerer signifikant effekt i forhold til ubehandlet. Tabel 9. Relative værdier (relativ i forhold til intern standard) for flere fedtsyresignaturer i jord fra Brøndlundgård med forskellige forbehandlinger. Jordprøver udtaget primo april 2003. Forskellige bogstaver indenfor samme række indikerer signifikant forskel på forekomst af den pågældende fedtsyre i jord med forskellig behandling LSD 0.05 (n=4). Felter fremhævet med gråt indikerer signifikant effekt i forhold til ubehandlet. 6 Diskussion
Det overordnede formål med dette projekt var at udvikle strategier til forebyggelse og bekæmpelse af rodsygdomme og ukrudt uden anvendelse af kemiske jorddesinfektionsmidler. På den baggrund har vi udført forsøg, som sammenlignede kemisk, termisk og biologisk jorddesinfektion af frøbede i forstplanteskoler i relation til sanering af rodsygdomme og ukrudt og eventuelle uønskede langtidseffekter på plantegavnlige mykorrhizasvampe og den generelle jordmikroflora. 6.1 Virkning af Basamid, damp og grønafgrøder på rodsygdommeDer var ingen signifikante behandlingseffekter i nordmannsgran, sitkagran og hvidel, men i nobilisgran var der i alle behandlinger på nær efterafgrøder en bedre fremspiring og en mindre dødelighed blandt fremspirede planter frem til høst, hvilket resulterede i markant flere høstede planter, især i kombinationsbehandlingerne. Således kunne der høstes næsten dobbelt så mange planter fra kombinationsbehandlingerne, mens ingen af de tre saneringsmetoder alene gav flere planter end i den ubehandlede kontrol. Disse resultater tyder på, at den primære årsag til dødelighed i småplanter af nobilisgran skyldes et patogen, der klarer sig dårligt i konkurrence med andre svampe som f.eks. Pythium, hvilket dog ikke bekræftes af analyserne af patogensamfund associeret til rødder af nobilisgran. De udeblevne behandlingseffekter i de tre øvrige plantearter kan skyldes, at sygdomsudviklingen i disse planter forårsages af andre patogener, der er mindre følsomme overfor de anvendte saneringsmetoder. I forsøgene i dette projekt blev der fokuseret på sanering af det øverste 5 cm jordlag, hvilket også kan være medvirkende årsag til de udeblevne behandlingseffekter, da der vil være levende smitstof af rodpatogener i det dybereliggende jordlag. Enebak et al. (1990) sammenlignede forskellige jorddesinfektionsmetoder til sanering af patogene rodsvampe i Weymourh-fyr, og deres resultater viste, at dazomet, som er det aktive stof i Basamid, ikke påvirkede fremspiringen af Weymourh-fyr, men reducerede dødeligheden af fremspirende planter fra 17 % dødelighed i ubehandlet kontrol til 5 % dødelighed med dazomet, men i samme undersøgelse var der ingen virkning af dazomet på forekomst af rodpatogene svampe i jordprøver. I modsætning hertil, viste Hansen et al. (1990), at populationer af Pythium og Fusarium blev markant reduceret af fumigering med dazomet i frøbede med Douglas fyr. Hansen et al. (1990) undersøgte virkningen af fumigering med dazomet alene og i kombination med forskellige grønafgrøder på tre forstplanteskoler. I en af planterskolerne var der en øget forekomst af Fusarium med grønafgrøderne sudangræs og bønne med og uden fumigering med dazomet, samtidig med et fald i antal høstede planter over en toårig periode. I nærværende projekt blev der ikke målt på forskydninger i patogen populationer som følge af de forskellige behandlinger, men der blev målt en øget generel svampebiomasse målt med svampebiomarkøren 18:2w6,9. Selvom vores forsøg ikke peger på, at grønafgrøderne giver anledning til opformering af saprotrofe rodpatogener som Fusarium, kan det ikke udelukkes, at de anvendte grønafgrøder kan medvirke til en opformering af rodpatogene svampe. Det kan også tænkes, at inokulumpotentialet af patogene rodsvampe i de anvendte forsøgsarealer i det nærværende projekt har været for højt til at ændringer i inokulumpotentialet kunne måles med planteessay. I fremtidige forsøg kan det være en fordel at udføre parallelle potteforsøg, hvor forsøgsjorden fortyndes med sygdomsfri jord for at muliggøre måling af forskydninger i inokulumpotentialet. Fremspiringen af nordmannsgran var meget lav, hvilket tyder på, at der var problemer med frøbårne patogener, hvilket også understøttes af de øvrige forsøg, hvor der blev anvendt samme frømateriale som i markforsøgene. F.eks. blev der i patogenitetstesten isoleret F. oxysporum var redolens fra behandlinger inokuleret med andre svampe. Disse resultater understreger vigtigheden af at anvende sygdomsfri frømateriale. Efterfølgende test af det anvendte frøparti viste dog kun 1 % forekomst af Fusarium oxysporum, og den lave fremspiring kan således skyldes, at inokulumpotentialet af rodsvampe, der er patogene på nordmannsgran, er specielt højt i de anvendte forsøgsarealer. Forsøg med damp har vist en markant sanerende effekt mod rodsygdomme som Pythium, Fusarium og Rhizoctonia (Bødker & Noyé, 1994; Pinel et al., 1999), men i dette forsøg giver damp ikke anledning til et større udbytte, hvilket kan skyldes, at en dampning giver lettere adgang for rodpatogener som Pythium, der i fravær af en saprotrof antagonistisk mikroflora lettere kan etablere sig. Dette vindue for patogener ser ud til at kunne lukkes ved at øge den mikrobielle aktivitet gennem tilførsel af organisk materiale umiddelbart efter jorddesinfektion, idet der er tendens til, at kombinationsbehandlingerne med damp/Basamid efterår og en efterfølgende etablering af efterafgrøder giver flere planter, dog ikke i nordmannsgran, hvor der generelt ikke var nogen behandlingseffekter. Den sanerende virkning af grønafgrøder som f.eks. Brassica arter beror sandsynligvis på en kombination af udskillelse af glycosinolater og opformering af en sygdomshæmmende mikroflora ved omsætningen af grønafgrøden. Det er veldokumenteret, at glycosinolater er toksiske overfor en bred vifte af rodpatogene svampe (Sawar et al., 1998; Smolinska et al., 2003). Bødker & Thorup-Kristensen (1999) undersøgte virkningen af grønafgrøder på udvikling af ærterodråd forårsaget af patogenet Aphanomyces euteiches under markforhold. I forsøget indgik to repræsentanter fra korsblomstrede, græsser og ærteblomstrede planter. De korsblomstrede grønafgrøder olieræddike og turnips reducerede hver især sygdomsudviklingen i et af to forsøg, mens de andre grønafgrøder ingen sanerende virkning havde. Disse resultater tyder på, at potentialet for brug af grønafgrøder til sanering af rodsygdomme er størst blandt Brassicaceae, hvilket støttes af Lazzeri & Manici (2001), som sammenlignede forskellige grønafgrøders sanerende virkning overfor rodpatogenet Pythium ultimum i potteforsøg. 6.2 Virkning af Basamid, damp og grønafgrøder på ukrudtPå begge forstplanteskoler var der, som forventet, problemer med ukrudt. Damp var effektiv til at bekæmpe ukrudt, hvilket stemmer overens med andre undersøgelser om effekter af damp på ukrudt (Bødker & Noyé, 1994; Pinel et al., 1999; Andersen & Hartvig, 2002; Melander et al., 2002). Vores resultater viser yderligere, at virkningen af damp og Basamid er langtidsholdbar, idet der ikke kunne måles signifikante forskelle på efterårs- og forårsdampningen, hvilket er i overensstemmelse med andre danske undersøgelser på området (Hartvig & Andersen, 2002; Andersen & Hartvig, 2003). Majroe som efterafgrøde havde meget overraskende en sanerende virkning overfor enårig rapsgræs, som var det dominerende ukrudtsproblem i forsøgene. Biosaneringen med majroer var lige så effektiv som damp og Basamid. Årsagen til den biosanerende virkning af majroer skyldes sandsynligvis allelopatiske forbindelser som isothiocyanater, der også hæmmer spiring af frø og kimplantevækst af hvede (Baily et al., 1990). Fremspiring af sitkagran og hvidel var dog også hæmmet af majroer, hvilket indebærer, at denne form for sanering sandsynligvis kun kan anvendes i specifikke situationer, hvor kulturen er mindre følsom overfor majroens allelopatiske forbindelser. Sammenfattende viser resultaterne fra dette projekt at damp fuldt ud kan erstatte Basamid, og at der er potentiale i biosanering af ukrudt med majroe. 6.3 Non-target effekter af de forskellige metoder til sanering af rodsygdommeArbuskulær mykorrhiza. Rødder af hvidel kan danne symbiose med arbuskulær mykorrhiza (AM) såvel som ektomykorrhiza (EM) og kvælstoffikserende bakterier Frankia. AM forekommer primært i det første år, og herefter domineres rodsystemet af EM (Arveby & Granhall, 1998). I forsøget på Akkerup var rodkoloniseringen med AM i hvidel upåvirket af de forskellige saneringsmetoder. Der er dog en tendens til, at damp- og Basamidbehandlingerne hæmmede mykorrhizadannelsen. Dazomet kan anvendes til eliminering af naturlige populationer af mykorrhiza i jord som mykorrhizakontrol til at studere, hvordan mykorrhiza påvirker planters vækst (Thingstrup Olsson et al., 1999), og det var derfor forventet, at Basamid ville hæmme mykorrhizadannelsen. Årsagen til den udeblevne virkning af Basamid og damp kan skyldes, at behandlingen alene blev foretaget i de øverste 5 cm jordlag, og det må forventes, at der er inokulum af AM svampe i hele pløjelaget. Da majroe ikke danner mykorrhiza, sker der ingen opformering af mykorrhizasporer, mens havre danner mykorrhiza, og her vil der kunne forekomme en opformering. Spiring af mykorrhizasvampes sporer hæmmes af glycosinolater (Screiner & Koide, 1993), hvilket også kan være forklaringen på den hæmmende, men dog ikke signifikante, virkning af majroe på mykorrhizakoloniseringen i hvidel. Ektomykorrhiza. Forekomst af ektomykorrhiza i nordmannsgran var ikke påvirket af de anvendte saneringsmetoder. Ektomykorrhiza i Douglas fyr i amerikanske skovplanteskoler var ligeledes upåvirket af dazomet (Enebak et al., 1990). Det kan dog ikke udelukkes, at disse svampe på kort sigt påvirkes af de anvendte saneringsmetoder, hvilket også var tilfældet for ektomykorrhiza i sortfyr (Pinus nigra), hvor en hæmmende virkning af Basamid og damp holdt sig i mindst 3 måneder (Soulas et al., 1997). Forsøget blev dog udført i væksthus, så genindvandring af svampesporer ikke var mulig, hvilket er tilfældet under markforhold. Fedtsyreresultaterne indikerer dog, at en vedvarende hæmmende virkning af damp og Basamid på svampe og derunder også EM svampe er mulig (se afsnit om den generelle mikroflora). Den generelle mikroflora. I udgangspunktet var der en højere mikrobiel biomasse på Brøndlundgård end i Akkerup Planteskole, hvilket sandsynligvis skyldes, at der umiddelbart før forsøgsstart blev dyrket en afgrøde på Brøndlundgård, mens forsøgsarealet på Akkerup Planteskole lå brak. Overordnet var der på begge planteskoler ingen målbare effekter af damp og Basamid på forekomst af bakterier, mens forekomst af svampemarkøren 18:2w6,9 var reduceret, hvilket er i overensstemmelse med resultater fra Olsson et al. (1999), hvor mikrofloraen på nær svampe var reetableret allerede 3 måneder efter Basamid behandling. Svampemarkøren findes i både non-patogene og patogene svampe, som således samlet set reduceres med damp og Basamid. Konsekvenserne af dette kan altså både være et lavere smittetryk af patogener og lavere forekomst af saprotrofe svampe, der er centrale for jordens frugtbarhed i relation til planters ernæring gennem omsætning af organisk materiale. Grønafgrøderne derimod fremmede den generelle mikrobielle biomasse i Akkerup Planteskole, der ved forsøgsstart havde en lav mikrobiel biomasse. Det er imidlertid afgørende, om det er patogene rodsvampe og/eller saprotrofe non-patogene svampe, der opformeres af grønafgrøderne. 6.4 Beskrivelse af samfund af rodassocierede svampe (ikke mykorrhizasvampe) i forstplanteskolerI sammenfatning er der i dette projekt konstateret, at der i jordene i de to planteskoler forekommer et meget bredt udvalg af svampe med plantepatogene potentialer, spændende fra meget aggressive "før fremspiring patogener" som Pythium ultimum over de mindre aggressive så som Fusarium oxysporum var. redolens og Botrytis cinerea, der til gengæld forårsager både før og efter fremspirings problemer, det sidste gennem flere år under etablering af kulturerne, til de mindst aggressive arter så som Penicillium spp. og Cladosporium cladosporioides, der mest optræder som lejlighedsparasitter under særlige stressbetingelser. I tilgift må det konstateres, at mange af de fundne patogene arter ofte vil være frøbårne. I forhold til etablering af sunde kulturer hersker der derfor en temmelig kompleks problemstilling. Fusarium oxysporum. Arten Fusarium oxysporum omfatter adskillige formae speciales (f.sp.), hver med sit afgrænsede værtregister. Således er f.eks. Fusarium oxysporum (var. oxysporum) f. sp. pini tilpasset som patogen i nåletræsarter (f. eks. Booth, 1971). F. oxysporum er endvidere opdelt i varieteterne var. oxysporum og var. redolens, men det er sjældent, der i rapporter om F. oxysporum-angreb tages stilling til patogenets varietets- og f. sp.-tilhørsforhold. Det kan derfor ikke udelukkes, at angreb, der er rapporterede som F. oxysporum-angreb, kunne skyldes varieteten redolens. En lang række kilder rapporterer, at Fusarium oxysporum i bred forstand er udbredt årsag til sygdomsproblemer i forstplanteskolekulturer, og ofte i nåletræskulturer. I nærværende undersøgelse er det udelukkende varieteten redolens af Fusarium oxysporum, der er fundet på rødder af de fire træarter, i begge planteskoler. F. oxysporum var. oxysporum blev dog fundet som formodentligt frøbåren forekomst i forbindelse med smitteforsøg. F. oxysporum var. redolens var den hyppigst forekommende svamp i både syge og symptomfri planter i alle fire træarter i Akkerup. Ifølge Ocamb & Juzwik (1995) var F. oxysporum var. redolens den svampeart, der optrådte hyppigst på nekrotiske rødder af 2-årige, levende Pinus strobus-småplanter i to planteskoler (USA og Canada), fulgt af F. oxysporum og F. proliferatum samt yderligere mindst fire andre Fusarium-arter. Af samme undersøgelse fremgår det, at jordbehandling med Basamid reducerede Fusarium-rodråd, uden at eliminere det. Angreb af F. oxysporum i forstplanteskolekulturer forbindes generelt med "damping off" (rod- og kimbrand, væltesyge, spiringsfusarioser) samt rodråd, hvilket er rapporteret i adskillige nåletræarter (Stepniewska, 1999; Sinclair et al., 1987). I et oversigtsværk angiver Sinclair et al. (1987), at F. oxysporum-rodinfektion i nåletræer finder sted inden for de første 6 uger efter såning, hvorved hovedroden går til. Småplanterne kan overleve dette, om end i væksthæmmet form ved dannelse af adventivrødder ved rodhalsen, men varmestress, frost og invaderende mikroorganismer fører før eller siden til plantedød. Infektioner af F. oxysporum er mest betydende inden for småplanternes første år, ifølge Sinclair et al. (1987). Uroševic (1961) har undersøgt patogeniteten af en lang række svampe, isolerede fra frø af bl.a. rødgran og skovfyr, samt fra eg, bøg og ahorn, over for frø og småplanter af de to nævnte nåletræsarter. Artsspektret af frøbårne svampe omfattede blandt andet de fleste af de arter, der er isoleret i nærværende projekt. I smitteforsøg med disse frøbårne svampe i et kunstigt system (spiring og vækst af svampesporeinokuleret frø på fugtigt filtrerpapir) sås kun ubetydelig spirehæmning i P. abies, når F. oxysporum var. redolens anvendtes som inokulum, mens spiringen reduceredes med over 20 % i P. sylvestris med samme inokulum. I nærværende undersøgelse fandtes en signifikant reduktion af fremspiring af nordmannsgran, hvor rene eller blandede inokulae af Brøndlundgård-isolater af F. oxysporum var. redolens blev benyttet, mens det enlige Akkerup-isolat af samme art ikke gav nogen signifikant spiringsreduktion. Uroševic (1961) angav, at mens flere Fusarium-arter, herunder F. culmorum, er i stand til at forårsage spirehæmning ved at penetrere dybt i frøet, er andre Fusarium-arters, herunder F. oxysporum var. redolens, evne til dette mere begrænset, og disse arters patogene rolle manifesteres i højere grad efter frøets fremspiring. Uroševic (1961) fandt i sin undersøgelse, at F. oxysporum var. redolens kunne forårsage "damping-off" i 21 % kimplanter af P. sylvestris inden for 21 dage fra fremspiring og vækst i sand. Brøndlundgård-isolaterne synes at have været i stand til både at reducere fremspiring og til at angribe og dræbe planter efter fremspiring i nordmannsgran, mens Akkerup-isolatet ikke havde signifikant effekt på hverken fremspiring eller overlevelse inden for patogenitetstestenes fire ugers forløb. Selv om der i den afsøgte litteratur ikke er fundet undersøgelser, der specifikt dækker sammenhængen mellem F. oxysporum var. redolens og de her undersøgte træarter, støtter resultaterne af den nærværende undersøgelse den generelle påstand om artens patogenitet i nåletræskulturer i form af efterfremspiring "damping-off" problemer, og føjer til den eksisterende viden, at arten også har potentiale til at forårsage førfremspirings problemer, i hvert fald i nordmannsgran. Der er kun fundet en enkelt kilde, der angiver F. oxysporum som patogen i Alnus. Nyeko et al. (2002) angiver, at damping-off forårsaget af F. oxysporum udgjorde det alvorligste sygdomsproblem i ugandiske Alnus-planteskoler. Sammenfattende tolkes resultaterne af nærværende undersøgelse og litteraturens erfaringer sådan, at 1) der findes variabilitet indenfor arten F. oxysporum var. redolens med hensyn til patogent potentiale, jævnfør forskelle mellem Akkerup- og Brøndlundgård-isolater, 2) arten har et alment højt patogent potentiale inden for et bredt spektrum af nåletræsarter jævnfør litteraturen og 3) arten er i stand til at forårsage både pre- og post-emergence-problemer i kulturernes tidligste fase og til at svække etablerede planter så meget, at de går til inden for de første to år (Sinclair et al.,1987). Fusarium avenaceum kan forårsage damping-off i Pinus sylvestris (Hanso & Karoles, 1980; Karoles, 1980; Asiegbu et al. 1997) angiver, at Picea abies småplanterødder er modtagelige for angreb af F. avenaceum. Kacprzak et al. (2001) fandt ligeledes en vis F. avenaceum-patogenitet overfor Larix europaea, Picea abies og Pinus sylvestris. Der foreligger i litteraturen tilsyneladende ikke viden om F. avenaceums patogenitet specifikt i forhold til etablering af småplanter af de fire træarter, der indgår i dette projekt. Der fandtes imidlertid ret høje forekomster af dette patogen på planter i alle fire kulturer i Akkerup, og patogenitetstests viste signifikant reduktion af både fremspiring og overlevelse af nordmannsgran. Mens der i litteraturen er fundet en del eksempler på patogenitet af F. avenaceum over for diverse nåletræsarter, jvnf. ovenstående, er der ikke fundet eksempler på patogenitet overfor Alnus spp. Disse forhold afspejles muligvis i, at F. avenaceum udelukkende fandtes på symptomfri hvidel-planter fra Akkerup, mens arten forekom hyppigst i syge eksemplarer af nåletræsarterne nobilis og nordmannsgran og stort set var ligeligt fordelt mellem symptomfri og syge sitkagran. F. avenaceum forekom ikke i planter i Brøndlundgård, men var hyppig i Akkerup, og patogenitetstests viste det benyttede isolats evne til at forårsage en signifikant reduktion af såvel fremspiring som overlevelse af nordmannsgran. Pythium ultimum, Pythium rostratum og Pythium periplocum. Pythium ultimum har et særdeles vidtspændende værtsregister (e.g. Domsch et al., 1980a) og fremkalder generelt "damping–off" i forbindelse med de tidligste faser af fremspiring og etablering af planter. Der foreligger ingen kilder af nyere dato, der omtaler P. ultimums patogenitet i de fire kulturer, der indgår i dette projekt, men f.eks. Kozlowski & Metraux (1998), Lilja (1994) og Pradhan (1997) beskriver patogenitet i kulturer som Alnus nepalensis, Picea abies og Pinus sylvestris, og der er næppe grund til at tro, at de i projektet behandlede kulturer udmærker sig frem for andre Coniferae-arter ved at besidde en særlig resistens over for P. ultimum. Det er ikke her undersøgt, om der er grundlag for at skelne mellem patogenitet af P. ultimum var. ultimum kontra P. ultimum var. sporangiferum, og det konstateres blot, at sidstnævnte kunne påvises i Brøndlundgård. Set under den fælles artsbetegnelse blev P. ultimum isoleret i relativt lave frekvenser fra Akkerup, men dog fra alle fire træarter, og arten blev som nævnt også fundet i Brøndlundgård, hvor frekvenser blot ikke blev bestemt. I patogenitetstestene over for nordmannsgran forårsagede Brøndlundgård-isolatet (BPu) 100 % pre-emergence-død af alle spirende frø, mens Akkerup-isolatet (APu) signifikant reducerede fremspiring fra ca. 77 % til 50 % og bevirkede en ca. 50 % reduktion af overlevende planter fire uger efter såning, sammenlignet med uinokuleret kontrol. Pythium ultimum blev fundet i relativt lave frekvenser på planterødder i Akkerup. Dette forhold kan være udtryk for, at mange angrebne planter ikke blev genstand for udvælgelse til isolering af den simple grund, at angrebne planter ikke overlevede til fremspiring, jævnfør de resultater, der fandtes i patogenitetstest med isolat BPu. P. rostratum blev kun fundet på syge planter i nobilis og nordmannsgran, samt på symptomfri hvidel og på symptomfri og syge sitkagran i Akkerup, og blev ikke fundet i Brøndlundgård. Der er ikke fundet kilder i litteraturen, der antyder patogenitet af denne art i skovtræskulturer. Smitteforsøg med isolat APr viste imidlertid en signifikant reduktion af både fremspiring og overlevelse af nordmannsgran. P. periplocum er i litteraturen nævnt som plantepatogen i plænegræsser (Aoyagi et al., 1999) og er isoleret fra råd i vandmelon (Drechsler, 1939), men er ikke omtalt som patogen i skovtrækulturer. Arten er i øvrigt bedst kendt som mykoparasit og er flere gange søgt udnyttet til biologisk bekæmpelse (e.g. Hockenhull et al., 1992). I nærværende smitteforsøg med BPo havde isolatet ingen reducerende effekt på fremspiring, og havde tydeligvis en klart positiv effekt på overlevelse, der var bedre end den, der forekom i kontrolplanterne. Botrytis cinerea angives i Sinclair et al. (1987) oversigtsværk som årsag til "damping-off" og til blad- og skudbrand i planteskolekulturer af blandt andet arter af fyr og gran samt en række andre nåletræsslægter. Ministere des Ressources Naturelles (2000) angav, at B. cinerea var årsag til omfattende tab af småplanter i canadiske forstplanteskoler. Sinclair et al. (1987) omtaler desuden Botrytis-skimmel i mange arter forstplanteskolekulturer. Alnus spp. nævnes ikke specifikt, ligesom der ikke synes at foreligge nyere litteratur i denne sammenhæng. I nærværende undersøgelse forekom B. cinerea i 60 % af syge A. incana-planter, men kun i 20 % af de symptomfri fra Akkerup. Derudover forekom B. cinerea i lave frekvenser (op til 20 %) i nåletræsarterne fra Akkerup. I patogenitetstests forårsagede Akkerup-isolatet (A2) også signifikant reduktion af både fremspiring og overlevelse af nordmannsgran. Cylindrocarpon destructans og Truncatella truncata. Sutherland et al. (1972) fandt, at Cylindrocarpon destructans, der i British Columbia var kendt for at forårsage omfattende ødelæggelser i planteskolekulturer af Pseudotsuga menziesii (douglasfyr) som sårparasit, i et samspil med nematodearten Xiphinema bakeri, havde udviklet sig til et lignende problem i kulturer af Picea sitchensis, Picea glauca og Tsuga heterophylla, hvor planteskolejorden var infesteret med X. bakeri. C. destructans var i forsøg i stand til at dræbe småplanter af Picea abies inden for kort tid (Forbrig, 1989). Mosandl & Aas (1986) angav, at C. destructans var patogen på kimplanter af Picea abies, Abies alba og Fagus sylvatica, samt at den var den hyppigst forekommende "damping-off"-svamp i isoleringer fra en 120 år gammel træbestand af disse træarter. Dette patogen er i øvrigt almindeligt forekommende i rhizosfæren af skovtræer (Domsch et al., 1980a). Manka (1997) mente at konstatere, at C. destructans var årsag til omfattende tab i et år gamle, transplanterede småplanter af el, bøg, birk, eg m.fl. i Polen, og antog, at det var resultatet af sårinfektion i vinterskadede rødder. Mens C. destructans forekom hyppigere i syge end i symptomfri hvidel i Akkerup, var der ikke noget fælles mønster med hensyn til frekvensfordeling af svampen i syge og symptomfri planter i de tre nåletræsarter. I nordmannsgran fandtes C. destructans udelukkende i symptomfri planter i Akkerup, hvilket skal sammenholdes med, at Akkerup-isolatet A4 af denne svamp ikke gav anledning til signifikant reduktion af fremspiring eller overlevelse i nordmannsgran i smitteforsøget. Til gengæld fandtes signifikant reduktion af overlevelse i smitteforsøg med Brøndlundgård-isolatet B4 af C. destructans. Smitteforsøg med ren Truncatella truncata (isolat A8) forårsagede ikke signifikant reduktion af fremspiring, men reducerede overlevelsen af de fremspirede nordmannsgran signifikant. Isolatblandingen A11, der bestod af C. destructans (af anden oprindelse end A4) og Truncatella truncata (af anden oprindelse end A8), reducerede både fremspiring og overlevelse af nordmannsgran signifikant, og der var således en forstærket patogenitet af isolatblandingen i sammenligning med virkningen af de to rene isolater A4 og A8. T. truncata forekom i lave frekvenser i hvidel og sitkagran, men blev ikke fundet i nobilis og nordmannsgran i Akkerup, og der fandtes ingen forekomster fra Brøndlundgård-kulturer. Der er i litteraturen kun fundet et enkelt eksempel på patogenitet af T. truncata i skovtræer. Hanso & Karoles (1980) angav, at T. truncata forårsagede mere end 33 % pre-emergence-død, men ingen tilfælde af post-emergence-død i Pinus sylvestris i smitteforsøg. Cylindrocarpon magnusianum, Humicola sp, Phoma glomerata og Sordaria fimicola er almindeligt forekommende i forbindelse med skovplantekulturer og skovbundsmykoflora (e.g. Domsch et al., 1980a,b; Mittal et al., 1990). Der er ikke fundet kilder i litteraturen, der placerer disse arter som patogener i skovplantekulturer. Imidlertid var isolaterne A6 (C. magnusianum) og B30 (Sordaria fimicola) i smitteforsøgene i stand til at forårsage en signifikant og særdeles kraftig reduktion af fremspiring og overlevelse, og isolaterne A7 (Humicola sp.) og B100 (Phoma glomerata) reducerede overlevelsen signifikant i nordmannsgran. Humicola sp. forekom ret hyppigt i syge sitka- og nordmannsgran og i lavere frekvenser i nobilis og hvidel i Akkerup. C. magnusianum forekom hovedsageligt i symptomfri planter, i visse tilfælde i høje frekvenser. Frekvenser af Phoma glomerata og Sordaria fimicola, der stammede fra Brøndlundgård kendes ikke. Alternaria alternata, Cladosporium cladosporioides, Penicillium expansum, Penicillium sp. og Verticillium sp. er primært saprobe svampe (der forekommer dog en del stærkt plantepatogene Verticillium spp. i andre plantefamilier end de her omhandlede) og anses generelt kun som sekundære lejligheds-/sårparasitter i planter, men de kan dog under særlige omstændigheder af plantestress m.v. optræde patogent (e.g. Domsch et al., 1980a; Sinclair et al., 1987). Der foreligger imidlertid i litteraturen angivelser af, at A. alternata kan forårsage problemer under spiring og etablering af skovplanter. Mittal & Wang (1993) fandt således en svag patogenicitet af A. alternata, samt af Cladosporium cladosporioides, Penicillium aurantiogriseum og Rhizopus stolonifer, på frøspiring og tidlig kimplanteudvikling i Picea glauca og Pinus strobus. James & Woo (1987) fandt ligeledes, at A. alternata havde svag patogenicitet i kimplanter af Picea engelmannii og Pseudotsuga menziesii, samt at patogeniteten især syntes at optræde i forbindelse med stressning af planterne. Mittal & Wang (1986) angav, at A. alternata, F. oxysporum og Penicillium variabile forhindrede fremspiring eller gav henfald af skud ("damping-off"-symptomer) i Picea glauca. Uroševic (1961) angav, at Verticillium albo-atra hæmmede fremspiring i Picea abies, men ikke i Pinus sylvestris, og fandt i øvrigt spirehæmning i variabel grad i disse to træarter i smitteforsøg med fire forskellige Penicillium-arter. I Akkerup-kulturer forekom A. alternata hyppigst i symptomfri planter af hvidel (80 %), men der var dog også en høj forekomst i syge planter (60 %). Det samme gjaldt nordmannsgran, hvor begge frekvenser imidlertid var noget lavere. I nobilis fandtes en ligelig fordeling (30 %) på syge og symptomfri planter, mens der i sitkagran fandtes en markant højere forekomst i syge planter (62,5 %) end i symptomfri (25 %). Arten blev ikke isoleret fra Brøndlundgård-kulturer. I smitteforsøg med A. alternata (isolat A5) fandtes en signifikant reduktion af fremspiring, men ingen post-emergence-effekter, hvilket stemmer pænt med litteraturens angivelser af arten som særligt problem i forbindelse med fremspiring. C. cladosporioides havde ingen effekt på fremspiring og overlevelse i smitteforsøg, og forekom i øvrigt kun i lave frekvenser i Akkerup-kulturer og blev ikke fundet i Brøndlundgård. Penicillium sp.-Verticillium sp.-blandingen A9 havde i smitteforsøg en vis reducerende effekt på overlevelse efter fremspiring, men ingen effekt på fremspiringen af nordmannsgran. Forekomsten af disse to svampe var lav i Akkerup. Det er sandsynligt, men ikke bekræftet, at den omtalte Penicillium sp. i blanding A9 var af arten P. expansum, der fandtes i Brøndlundgård-kulturer (isolat B2). Isolat B2 reducerede i denne undersøgelses smitteforsøg både fremspiring og overlevelse signifikant. En screening af det benyttede frøparti af nordmannsgran viste i øvrigt særdeles høje forekomster af Penicillium spp. (data ikke vist). Rhizopus sp., Zygorrhyncus moelleri optrådte som små forureninger i inokulae benyttet til smitteforsøg og antages ikke at have haft indflydelse i denne sammenhæng. Disse arter, samt forekomster af andre skimmelsvampe samt Trichoderma spp., der er registreret som forekomster i Tabel 13 vil ikke blive omtalt nærmere her. Flere af disse arter er i øvrigt fundet som endog særdeles hyppige i det frøparti af nordmannsgran, der er benyttet i projektet (data ikke vist). Sammenligning af strategier til forebyggelse/bekæmpelse af rodsygdomme og ukrudt De opnåede resultater tyder på, at damp er lige så effektiv som Basamid til sanering af rodsygdomme og ukrudt, og at der ej heller er flere non-target effekter med dampning end med Basamid. Umiddelbart vil damp altså kunne erstatte brugen af Basamid, men der kan være andre økologiske problemer med at anvende dampning af frøbede, da dampning er meget energiforbrugende og dermed belaster miljøet i forhold til CO2 emission. Endelig er der økonomiske betragtninger for gartnerne, da dampning er bekosteligt. Biosanering med efterafgrøder (majroe og havre) er tilsyneladende ikke et egnet alternativ til sanering af rodsygdomme, men kombination af efterafgrøder og en forudgående jordesinfektion gav den bedste sanering af rodsygdomme i nobilis, hvilket tyder på, at der i denne kultur ikke er de samme sygdomsproblemer som i kulturerne af de øvrige tre plantearter. Samtidig giver den sanerende effekt af enårig rapgræs med nedmuldning af majroe anledning til at arbejde videre med efterafgrøder som et lovende biologisk alternativ til kemisk/termisk sanering af ukrudt. Ud fra de opnåede resultater er det ikke muligt at anvise en generel strategi til forebyggelse/bekæmpelse af rodsygdomme og ukrudt i forstplanteskoler, idet virkningen af de forskellige saneringsmetoder synes at være planteafhængig. For at kunne udvikle en strategi baseret på biologisk aktivering af frøbede er det nødvendigt at kende de forskellige rodpatogeners biologi. Blandt de patogener, som var fremherskende i de to forstplanteskoler, Fusarium og Pythium, er der store forskelle i deres evner til at konkurrere om organisk materiale, hvor Pythium hører til patogener med ringe konkurrenceevne, og Fusarium hører til patogener med gode konkurrenceevner. Denne grundlæggende forskel i patogenernes biologi gør det vanskeligt at forudsige virkningen af tilførsel af organisk materiale. Der er således behov for mere viden om, hvordan forskellige typer af organisk materiale, heriblandt grønafgrøder, påvirker rodpatogener og deres antagonister, før det er muligt at fremlægge strategier til forebyggelse af rodpatogener baseret på tilsætning af organisk materiale til frøbede. 7 Konklusion
8 PerspektiveringProjektet har givet en værdifuld indsigt i fluktuationer i de mikrobielle samfund efter en kemisk, termisk eller biologisk påvirkning, som sammen med den indsamlede viden om rodsvampe associerede til rødder af skovsmåplanter, kan anvendes i det videre arbejde med at udvikle effektive og miljøvenlige metoder til forebyggelse/bekæmpelse af frøukrudt og jordbårne rodpatogener. I forhold til sygdomsbekæmpelse tyder vores resultater på, at en kombination af damp og en efterfølgende biologisk aktivering af såbedet gennem tilsætning af organisk materiale og eventuel podning med kendte mikrobielle antagonister vil kunne virke forebyggende mod rodsygdomme. Her er der også brug for yderligere forskning for at finde frem til en optimal metode til biologisk aktivering af såbedet. I dette projekt blev der taget udgangspunkt i sanering af frøukrudt og rodsygdomme i de øverste 5 cm jordlag. Det kan tænkes at desinfektion i større jorddybde f. eks 10 cm ville give bedre sanering af rodsygdomme, men sandsynligvis også tilsvarende større utilsigtet virkning på plantegavnlige mikroorganismer. Der er behov for yderligere forskning for at klarlægge dette forhold. Der er også behov for yderligere forskning vedrørende sanering/opformering af rodpatogene svampe og plantegavnlige mikroorganismer med grønafgrøder. På det anvendte plan peger resultaterne fra dette projekt på, at dampning kan erstatte brugen af Basamid til sanering af ukrudt i frøbede i forstplanteskoler. Det skal dog tages med forbehold, da resultaterne alene bygger på forsøg i to planteskoler, hvorfor der er behov for flere valideringsforsøg, før der kan drages mere generelle konklusioner 9 LitteraturAndersen L & Hartvig P (2002). Forebyggelse og kemisk ukrudtsbekæmpelse på containerpladsen. GartnerTidende 1: 12-13. Andersen L & Hartvig P (2003). Dampning kan ske om efteråret. GartnerTidende 119: 51-52 Aoyagi A, Kageyama K & Hyakumachi M (1999). Isolation of Pythium species from zoysia grass and their effect on severity of large patch disease. Plant Disease 83:171-175 Arveby AS & Granhall U (1998). Occurrence and succession of mycorrhizas in Alnus incana Swedish Journal of Agricultural Research 28 : 117-127 1998. Asiegbu FO, Johansson M & Daniel G (1997). Cytopathological responses of Picea abies seedling roots challenged in vitro by a necrotrophic Fusarium avenaceum (Fr.) Sacc. Journal of Phytopathology 145: 239-244. Baily Z, Oleszek W, Lewis J and Fenwick GR (1990). Alllelopatic potential of glucosinolates (mustard oil glycosides) and their degradation products against wheat. Plant and Soil 129:
277-281. Barron GL (1968). The genera of hyphomycetes from soil. Waeverly Press Inc. William and Wilkins Company, Baltimore, U.S.A. 364 pp. Bending GD & Lincoln SD (2000). Inhibition of soil nitrifying bacteria communities and their activities by glucosinolate hydrolysis products. Soil Biology and Biochemistry 32: 1261-1269. Booth C (1971). The genus Fusarium. Commonwealth Mycological Institute, Kew, Surrey, England. Eastern Press Limited, London and Reading, U.K. 237 pp. Brown PD & Morra MJ (1995). Glucosinolate-containing plant tissues as bioherbicides Journal of Agricultural and Food Chemistry. 43:3070-3074. Bødker L & Larsen J (2000). Rodsydomme i hvidel (Alnus incana) i dansk planteskoler. DJF-Rapport. Bødker L & Noyé G (1994). Effekten af varmebehandling af overfladejord i nåletræer over for ukrudt og rodpatogene svampe. 11. Danske Planteværnskonference. SP-rapport 7, 239-249. Bødker L & Thorup-Kristensen K (1999). Effect of green manure crops on root rot and arbuscular mycorrhizal fungi in pea roots. 337-344. Olesen, J.E., Eltun, R., Gooding, M.J., Jensen, E.S. & Köpke, U. (Eds) Designing and testing crop rotations for organic farming. DARCOF Report no. 1. Cavigelli MA, Robertson GP & Klug MJ (1995). Fatty acid methyl ester (FAME) profiles as measures of soil microbial community structure. Plant and Soil 170: 99-113. Dick MW (1990). Keys to Pythium. College of Estate Management, Whiteknights, Reading RG6 2AW, U.K. 64 pp. Domsch KH, Gams W & Anderson, TH (1980a). Compendium of soil fungi. Volume 1. Second printing 1986. Academic Press, London, U.K. 859 pp. Domsch KH, Gams W & Anderson, TH (1980b). Compendium of soil fungi. Volume 2. Second printing 1986. Academic Press, London, U.K. 405 pp. Drechsler C (1939). Several species of Pythium causing blossom-end rot of watermelons. Phytopathology 29: 391-422. Ellis MB (1971). Dematiaceous hyphomycetes. Commonwealth Mycological Institute, Kew, Surrey, England. Second reprint, 1980, Cambrian News, Aberyswyth, Dyfed, U.K. 608 pp. Elsgaard L, Elmholt S & Jørgensen MH (2003). Damp påvirker jordens mikroliv. Økologisk Jordbrug, 31. oktober. Enebak SA, Palmer MA & Blanchette RA (1990). Managing soilborne pathogens of white pine in a forest nursery. Plant Disease 74: 195-198. Forbrig R (1989). Anatomische und histologische Untersuchungen an pilzinfizierten Fichtenkeimlingen (Picea abies Karst.). II. [Anatomical and histological investigations on fungus-infected Norway spruce (Picea abies) seedlings. 2.] Allgemeine Forst und Jagdzeitung 160: 137-144. Hansen EM, Myrold MM & Hamm PB (1990). Effects of soil fumigation and cover crops on potential pathogens, microbial activity, nitrogen availability, and seedling quality in conifer nurseries. Phytopathology 80: 698-676. Hanso M & Karoles K (1980). Okaspuude infektsioonilise tousmepoletiku tekitajaid ja nendega kaasnevaid mikroseeni Eesti metsataimlatest. [Some agents of damping off in conifer seedlings in Estonian forest nurseries and associated microfungi]. Metsanduslikud-Uurimused, Estonian SSR 16: 120-134. Hartvig P & Andersen L (2002). Langtidsvirkende dampbehandling. GartnerTidende 50: 4-5. Hockenhull J, Jensen DF & Yudarti T (1992). The use of Pythium periplocum to control damping-off of cucumber seedlings caused by Pythium aphanidermatum. In: E.C. Tjamos, G.C. Papavizas & R.T. Cook (eds.) NATO Advanced Research Workshop on Biological Control of Plant Diseases: Progress and Challenges for the Future. Plenum Press: 203-206. Ingleby K, Mason PA, Last FT & Fleming LV (1990). Identification of ectomycorrhizas, ITE research publication no. 5, Edinburgh Research Station, Scotland; ISBN 0 11 701461 3. James RL & Woo JY (1987). Pathogenicity of Alternaria alternata on young Douglas-fir and Engelmann spruce germlings. Report Northern Region, USDA Forest Service No. 87-9, 4 pp. Kacprzak M, Asiegbu FO, Daniel G, Stenlid J, Manka M & Johansson M (2001). Resistance reaction of conifer species (European larch, Norway spruce, Scots pine) to infection by selected necrotrophic damping-off pathogens. European Journal of Plant Pathology. 107: 191-207. Karoles K (1980). Several aspects of the effects of fungicides on Fusarium avenaceum - an agent of damping off in Scots pine seedlings. Metsanduslikud Uurimused, Estonian SSR 16: 135-143. Lazzeri L & Manici LM (2001). Allelopathic effect of glucosinolate-containing plant green manure on Pythium sp and total fungal population in soil. Hort Science 36: 1283-1289. Kozlowski G & Metraux JP (1998). Infection of Norway spruce (Picea abies (L.) Karst.) seedlings with Pythium irregulare Buism. and Pythium ultimum Trow.: histological and biochemical responses. European Journal of Plant Pathology 104: 225-234. Lilja A (1994). The occurrence and pathogenicity of uni- and binucleate Rhizoctonia and Pythiaceae fungi among conifer seedlings in Finnish forest nurseries. European Journal of Forest pathology 24: 181-192. Manka M (1997). Broadleaved tree transplants dieback in spring '97 in Poland. Phytopathologia Polonica No. 13: 150-151. Melander B, Heisel T & Jørgensen MH (2002). Band-steaming for intra-row weed control. 5th Workshop of the EWRS working group: Physical and cultural weed control, Pisa Italy, 11-13 March 2002. Ministere des Ressources Naturelles (Quebec) Canada, Quebec (2000) Insectes, maladies et feux dans les forets quebecoises en 1999. [Forest pests, diseases and fire damage in Quebec in 1999]. 56 pp. Mittal RK & Wang BSP (1986). Emergence failure and top decay in white spruce germinants due to three fungi. Canadian Plant Disease Survey 66: 5-7. Mittal RK & Wang BSP (1993). Effects of some seed-borne fungi on Picea glauca and Pinus strobus seeds. European Journal of Forest Pathology 23: 138-146. Mosandl R & Aas G (1986). Vorkommen und Bedeutung von Keimlingspilzen im Bergmischwald der ostbayerischen Kalkalpen. [The occurrence and importance of fungi on germinating seedlings in montane mixed forest of the East Bavarian calcareous Alps]. Forst und Holzwirt 41: 471-475. Nyeko P, Edwards-Jones G & Day RK (2002). Insect herbivores and pathogens of Alnus species in Uganda. Agroforestry Systems 56: 117-127. Ocamb CM & Juzwik J (1995). Fusarium species associated with rhizosphere soil and diseased roots of eastern white pine seedlings and associated nursery soil. Canadian Journal of Plant Pathology 17: 325-330. Olsson PA, Thingstrup I, Jakobsen I & Bååthe E (1999). Estimation of the biomass of arbuscular mycorrhizal fungi in a linseed field. Soil Biology and Biochemistry 31: 1879-1887. Pinel MPC, Bond W, White JG & De Courcy Williams M (1999). Field vegetables: assessment of the potential for mobile soil steamning machinery to control diseases, weeds and mites of field salad and related crops. Final report on HDC Project FV229, East Malling: Horticultural Development Council, 55 pp. Pradhan M (1997). Occurrence of severe damping-off disease of Himalayan alder and its cultural management. Journal of Hill Research 10: 127-130. Sawar M, Kirkegaard JA, Wong PTW & Desmarchelier JM (1998). Biofumigation potential of brassicas. III. In vitro toxicity of isothiocyanates to soil-borne fungal pathogens. Plant and Soil 210: 103-112. Screiner RP & Koide RD (1993 ). Mustards, mustards oils and mycorrhizas. New Phytologist 123:107-113. Sinclair W A, Lyon HH & Johnson WT (1987). Diseases of trees and shrubs. Cornell University Press, Ithaca, New York, USA. 574 pp. Smith S & Read D. 1997. Mycorrhizal symbiosis. Academic Press. Smolinska U, Morra MJ, Knudsen GR & James RL (2003). Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum. Plant Disease 87: 407-412. Soulas ML, Le Bihan N, Camporota P, Jarosz C, Salerno MI and Perrin R (1997). Solarization in a forest nursery: effect on ectomycorrhizal soil infectivity and soil receptiveness to inoculation with Laccaria bicolor. Mycorrhiza 7: 95-100. Stepniewska H (1999). The vitality of forest tree seedlings grown on sawdust-peat substrates diversified in view of their utilisation time limit Sylwan 143: 43-49. Sutherland JR, Sluggett LJ & Lock W (1972). Corky root disease observed on two spruce species and western hemlock. Tree Planters' Notes 23: 18-20. Sutton BC (1980). The Coelomycetes. Fungi imperfecti with pycnidia, acervuli and stromata. Commonwealth Mycological Institute, Kew, Surrey. Robert MacLehose and Co., Ltd. Glasgow, U.K. 696 pp. Thingstrup I, Rubaek G, Sibbesen E, Jakobsen I (1998). Flax (Linum usitatissimum L.) depends on arbuscular mycorrhizal fungi for growth and P uptake at intermediate but not high soil P levels in the field. Plant and Soil 203: 37-46. Toyota K, Ritz K, Kuninaga S & Kimura M (1999). Impact of fumigation with metam sodium upon soil microbial community structure in two Japanese soils. Soil Science and Plant Nutrition 45: 207-223. Uroševic B (1961). The influence of saprophytic and semi-arasitic fungi on the germination of Norway spruce and Scots pine seeds. Proceedings of the International Seed Testing Association 26: 537-556. von Arx JA (1970). The genera of fungi sporulating in pure culture. 3rd edition, 1981. A. R. Gantner Verlag, K. G. Strauss & Cramer GmbH, Hirschberg, Germany. 424 pp. Nylund JE & Wallander H (1991). Ergosterol analysis as a means of quantifying mycorrhizal biomass. In: Methods in Microbiology vol 24. (ed by J.R. Norris, D.J. Read and A.K. Varma) Academic Press, London. Williams Woodward JL, Pfleger FL, Fritz VA and Allmaras RR (1997). Green manure of oat, rape and sweet corn for reducing coomon root rot in pea (Pisum sativum) caused by Aphanomyces euteiches Plant and Soil 188: 43-48.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||