Ukrudtsstriglingens effekter på dyr, planter og ressourceforbrug
Bilag A
1 Beskrivelse af ALMaSS-modelsystemet
I det følgende gives en beskrivelse af ALMaSS-modelsystemet som anvendes i forbindelse med modelscenarierne i projekt ”Ukrudtsstriglingens effekter på dyr, planter og ressourceforbrug”.
1.1 ALMaSS-modelsystemet
Navnet ’ALMaSS’ er en forkortelse af ´Animal, Land and Man Simulation System´og refererer til et samlet dynamisk modelsystem, der er i stand til at tage højde for det dynamiske samspil af forskellige biologiske og abiotiske faktorer i tid og rum, der har afgørende betydning for, hvordan, og med hvilken effekt, en given samling af individer (lokalbestand) i et nærmere defineret landskab bliver påvirket af forskellige driftsbetingelser.
Modelsystemet består i princippet af to hovedkomponenter – en landskabsmodel og en (eller flere) individbaserede dyremodeller samt en række undermodeller (vejrmodel, dyrkningsmodeller, vækstmodeller for afgrøde og vegetation, sædskifter og ’driftsleder’; se nedenfor). Landskabsmodellen kan bestå af et konstrueret landskab eller en kopi (digitalisering)af et virkeligt landskab som i nærværende projekt. Landskaberne kan være simple eller meget detaljerede, og udgør det miljø eller den fysiske ramme de enkelte individer i dyremodellerne lever i.
De individbaserede modeller adskiller sig fra de fleste andre modeltyper (eks. matematiske) i, at de har det enkelte individ som udgangspunkt og ikke en samling eller population af individer, der opfører sig ens på alle udefra kommende stimuli. I ALMaSS har de enkelte individer deres egne agenda: på et hvilket som helst tidspunkt og sted at agere efter, hvad der er bedst mht. overlevelse og reproduktion (denne modelegenskab har giver navn til modeltypen ’agent-baserede modeller’ forkortet ABM) . Dyrene kan kommunikere og erkende tilstande og ændringer, i det miljø de lever. Samlet betyder dette, at det enkelte individ hele tiden reagerer på det dynamiske samspil af (eller interaktioner mellem) forskellige faktorer, der påvirker det. De enkelte dyremodeller er konstrueret således, at det løbende under modelsimuleringerne er muligt at indhente oplysninger om forskellige variabler vedr. reproduktion, population mm.
1.2 Integration af enkeltfaktorer gennem modellering
Bestandsudvikling af de enkelte arter er bestemt af en lang række faktorer. Ved ændret driftspraksis ændres mange af disse faktorer. Modellering af effekten af forskellige dyrkningsscenarier er en måde at integrere den foreliggende viden om samspillet mellem de modellerede arter og det landskab, de befinder sig i og dermed bidrage til at forstå enkeltfaktorernes betydning.
ALMaSS er derfor et godt redskab til at undersøge, hvilke konsekvenser eks. en bestemt ændring i dyrkningspraksis vil have for en given population - eller dele heraf – i et bestemt område. Da en given ændring i dyrkningspraksis oftest påvirker forskellige arter i forskellig retning, er det som i indeværende projekt en fordel at undersøge de samme ændringer for en række arter med meget forskellig levevis.
I tidligere undersøgelser har ALMaSS eks. været anvendt til at undersøge pesticiders effekter på sanglærken set i relation til markstørrelse, og afgrødetype (Topping & Odderskær 2004). Modelleringerne, der foretages i nærværende projekt, har til formål gennem ’tekniske scenarier’ kvantitativt at beskrive konsekvenserne af forskellige striglings intensitet og timingen for striglingernes udførelse for tre almindeligt forekommende agerlandsarter, som repræsenterer forskellige trofiske niveauer og levevis (sanglærke samt edderkop og løbebille). Effekter måles på forskellige variabler for reproduktion og bestandsforhold i et konkret landskab, hvor alle bedrift og dyrkningsforhold kan manipuleres (scenarier), men som udgangspunkt er baseret på konkrete forhold i ’værkstedsområdet’ (for nærmere beskrivelse af dette se: Dalgaard et al. 2004). I princippet vil antallet af outputvariabler være så godt som ubegrænset, men i praksis har hovedparten af outputvariabler været inden for årlig baseret populationstørrelse (inkl. mål for forsk. livsstadier) herunder vækst-, ekstensions- reproduktions- og mortalitetsrater, antal individer der påvirkes af en bestemt ændring (eks. fra strigling), og for en enkelt art (markmus) er det også muligt at få output i form af genetiske variabler. I nærværende projekt anvendes følgende modeloutput: alle tre arter: årlige total pop. pr 31/12, sanglærke: total antal årsunger, klægningssucces, udflyvningssucces, ynglesucces, redemortalitet, ægkuldmortalitet (for nærmere forklaring se afsnit 3.4 i rapport).
I det følgende beskrives de enkelte elementer i ALMaSS modelsystemet. I princippet kan alle modelparametre ændres efter behov, men nogle ændringer er lette at ændre medens en ændring af andre rent tidsmæssigt er forbundet med et stort arbejde. I det følgende beskrives disse forhold under de enkelte delmodeller. For beskrivelsen af de individbaserede modeller er der udvalgt eksempler for sanglærken, men der vil ikke i det følgende blive givet en detaljeret oversigt over parametriseringen af de enkelte artsmodeller, da dette ligger udenfor rammerne for nærværende notat. Detaljerede oversigter kan findes i følgende publikationer: -(sanglærke: Topping & Odderskær 2004; edderkop: Thorbek & Topping, 2005; løbebille: Bilde & Topping, 2004; samlet for hele modelsystemet: www.almass.dk).
1.3 Landskabsmodellen
Landskabsmodellen – eller værkstedsområdet, som er en kopi af et virkeligt område, udgør den ”verden” de modellerede dyr skal leve og overleve i. Den indeholder en detaljeret gengivelse af alle vigtige landskabselementer (læhegn, markskel, marker, veje og bygninger mm.). Med mindre det er ønsket at undersøge forskellige udgaver af et landskab, holdes størrelse og placering af disse elementer konstant både i ’før-scenariet’, som repræsenterer de nuværende forhold, og i alle ’efter- scenarier’ der simuleres, og kun markafgrøder (sædskifte, eller som i denne undersøgelse, dyrkningspraksis) ændres mellem scenarierne.
Det udvalgte værkstedsområde, hvori de enkelte modelscenarier er udført, er et 10 x 10 km stort område beliggende sydøst for Bjerringbro (Figur 1).
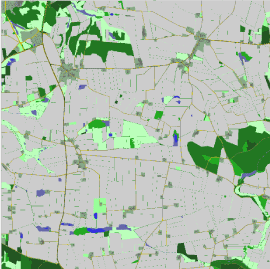
Figur 1. Udsnit (5x5 km, sydøst for Bjerringbro) af modellandskab, hvori modelkørsler foretages.
Området omfatter alle bedrifter i hele området. Dette valg er truffet ud fra modelmæssige hensyn, ud fra et ønske om, at det valgte landskab indeholder så stor andel dyrket land som muligt, hvilket for de pågældende modellerede arter er centralt. Ud fra flyfoto og markblok kort blev der foretaget en detaljeret digitalisering af området. Alle digitaliserede arealer (polygoner) blev defineret i forhold til en række udvalgte kategorier (marker, bygninger, haver, areelle småbiotoper, læhegn, skove, veje/vejrabatter, vandløb mm., Figur 2). Ved en efterfølgende inspektion af området blev alle arealinformationer vedrørende de forskellige polygontyper korrigeret for evt. fejl i forhold til placering, arealstørrelse og polygontype. For de individuelle læhegns vedkommende blev der desuden suppleret med oplysninger om højde, strukturel udformning mm. Alle oplysninger vedrørende bedriftstyper og ejerforhold (her: reelt oplysninger om arealer drevet af samme person) har været tilgængelige via GLR/CHR - register oplysninger.
Som udgangspunkt er de enkelte bedriftsenheder (i alt 123 bedrifter) og typer i modellandskabet defineret ud fra de konkrete forhold i 1998. Den enkelte bedriftsstørrelse og geografiske placering er defineret ud fra den person, der driver de pågældende marker (Figur 3). Alle ”beslutninger” om udførelsen af markoperationer på en given mark på et givet tidspunkt bliver foretaget i en ”Driftsleder” (se nedenfor), der samlet koordinerer oplysninger om bedriftstype, sædskifte, jordbunds- og vejrmæssige forhold.
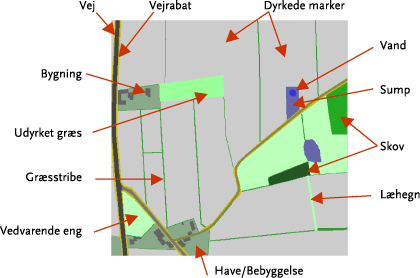
Figur 2. Udsnit af det digitaliserede landskabsmodel, med angivelse af selvstændigt definerede polygontyper (habitater).
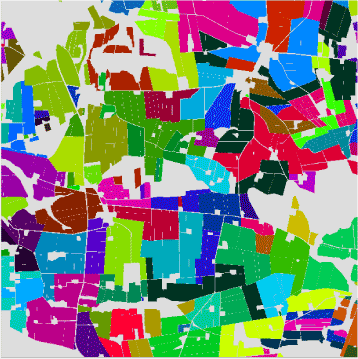
Figur 3. Angivelse af bedriftsforhold: marker med samme farve er drevet af samme landmand.
Ændring af landskabsparametre
Arbejdsmæssigt er udarbejdelsen (digitalisering) af en detaljeret kopi af et virkeligt landskab som her anvendte (10x10 km) en omfattende proces, der tager ca. tre måneder at generere. Derefter er en ændring i f.eks. typen af eksisterende polygontyper let at foretage, mens en ændring i den fysiske placering eller ’mængden af’ en eller flere strukturer i landskabet (polygontyper som f.eks. læhegn eller skove) forbundet med et større arbejde – afhængigt af ændringernes omfang. Ligeledes vil eks. en bestemt – og individuel - ændring af eksisterende markers størrelse være forbundet med et større digitaliseringsarbejde, medens en ens opdeling af alle marker vil være forholdsvis let at foretage.
1.4 Vejrdata og vækstmodeller for afgrøder
De valgte modelarters muligheder for overlevelse og reproduktion er i høj grad afhængig af vækstforholdene og strukturen (bladareal, biomasse, højde) af de enkelte afgrøder indenfor yngle- eller ’home-range’ området. For på en realistisk måde at ”gengive” virkelige forhold, er der udviklet specifikke vækstmodeller for de afgrøder, der er anvendt i forskellige typer af sædskifter (se efterfølgende). Vækstmodellerne er udarbejdet på grundlag af eksisterende modeller (Olesen og Heidmann, 1990; Plauborg og Olesen, 1991) samt data indsamlet ved DJF og DMU. Modellerne er baseret på ”graddage”, således at både bladareal og højdeudvikling er relateret til temperatursummer med en basistemperatur på 0°C (for nærmere beskrivelse, se Bilag B). Vejrdata, der er anvendt i plantevækstmodeller, dyremodeller og ”Driftslederen” (se efterfølgende) er baseret på data fra DJF-Foulum fra perioden 1988-1999. I modelleringerne er der anvendt daglige gennemsnit for temperatur (°C), nedbør (mm) og vindstyrke (m/sek.). De samme vejrdata indgår således i det modellerede tidsrum på 55 år i alt i perioder af 11 år.
Ændring af vejrdata og vækstmodeller
En ændring af vejrdata er let at foretage. Det vil dog til dels afhænge af, i hvor stor grad det er muligt at fremskaffe over et specifikt lokalt område, og over hvor lang en (evt. bestemt) årrække, det er ønskeligt at anvende data fra. De enkelte vækstmodeller (afgrøder og urtevegetation generelt) er forbundet med et stort arbejde at foretage ændringer i.
1.5 ’Driftsleder’ og dyrkningsmodeller
Dyrkningsmodeller, der beskriver vækstforhold og dyrkningstiltag er baseret på dyrkningsvejledninger (Dansk Landbrugsrådgivning) og konsulentrådgivning fra Djursland Landboforening - Landbocentret Følle, Godsforvaltningen ved Fussingø Statsskovdistrikt og DMU, Afd. for Vildtbiologi og Biodiversitet. For hver enkelt afgrødemodel er der i videst mulig omfang medtaget de variationer i dyrkningsmæssige tiltag, der er bestemt af f.eks. brugstype, geografiske forhold, jordbund, vejrforhold mm. Den indbyggede variation bevirker, at vækstforhold og dyrkningstiltag indenfor de enkelte afgrøder skifter mellem marker og bedrifter i de enkelte modelår. I alt er der udarbejdet dyrkningsmodeller for ca. 50 forskellige afgrøder (inkl. forskellige dyrkningsvarianter af disse), herunder de afgrøder der benyttes under nærværende projekt (Tabel 1). Et eks. på en dyrkningsmodel for vårbyg er angivet i projektbeskrivelsen (Appendiks 2 i Odderskær et al. 2003) for nærværende projekt.
Tabel 1. Oversigt over modelafgrøder, der indgår i sædskiftet på bedriftstypen kvægbrug i nærværende projekt.
| Kvægbrug |
| Vårbyg m. udlæg |
| 1. års kløver græs til slet/afgræsning |
| 2. års kløver græs til slet/afgræsning |
| Vinterhvede |
| Vårbyg |
| Vårbyg m. udlæg (til ensilering) |
| 1. års kløver græs til slet/afgræsning |
| Vårbyg m. udlæg(til ensilering) |
| Vårbyg |
I dyrkningsmodellen for de enkelte afgrøder antages, at markoperationerne i Tabel 2 kun udføres under bestemte vejrforhold. De fleste markoperationer vil således ikke blive udført før 3 – 5 dage efter forudgående nedbør, høst foretages tidligst 5 dage efter nedbør og marksprøjtninger udføres kun i tørvejr og under gunstige vindforhold (vindstyrker < 4, Boufort skala).
Tabel 2. Samlet oversigt over de markoperationer der er anvendt i de eksisterende dyrkningsmodeller.
| Markoperationer | |
| Efterårspløjning | Fungicidbehandling |
| Efterårsharvning | Insekticidbehandling |
| Efterårstromling | Radrensning |
| Efterårssåning | Strigling |
| Forårspløjning | Blindstrigling |
| Forårsharvning | Kunstvanding |
| Forårstromling | Skårlægning |
| Forårssåning | Høst |
| Gødskning | Køer på græs |
| - NPKS | |
| - NPK | Græsslet til hø |
| - PK | Græsslet til ensilage |
| - Flydende ammoniak | Græsslet til hø/ensilage |
| - Gylle | Afpudsning af ukrudt |
| - Mangansulfat | Halmsnitning |
| - Fastgødning | Halm/Høvending |
| - Slam | Halm/Høpresning |
| - N-ammoniumsulfat | |
| Herbicidbehandling | Stubharvning |
| Vækstregulering | |
Som en vigtig del mht. sanglærken specielt er mængden af føde, der er tilgængelig fra en given habitat, afhængig af den tilstedeværende insektbiomasse (total for alle insekt fødeemner), som igen er proportionelt relateret til vegetationsstrukturen (højde og biomasse). Disse relationer blev for almindelige landbrugsafgrøder genereret fra feltdata indhentet i modelsimuleringsområdet ved Bjerringbro (Topping, unpublished). Ud fra disse data er der udarbejdet nogle generelle funktioner, der beskriver bestandsudviklingen af insekter i relation til den tidsmæssige udvikling (plantebiomasse) af de enkelte afgrøder og ukrudt (generelt). Insekttætheder omregnes herefter til en gennemsnitlig tørvægt per arealenhed for de vigtigste grupper af insektfødeemner efter eksisterende datamateriale (Odderskær et al. 1997).
Ændring af dyrkningsmodeller
En ændring i eksisterende dyrkningsmodeller er forholdsvis let (en til få dage) afhængig af, om det et er spørgsmål om f.eks. at ændre i den procentdel der udfører en bestemt markoperation, eller det f.eks. er en ændring i typen eller rækkefølgen af markoperationer). Derimod vil det arbejdsmæssigt tage fra to til tre uger i alt at lave og teste en ny afgrøde fra bunden afhængigt af, hvor kompliceret et dyrkningsmønster, der er tale om.
1.6 Effekter af markoperationer
Alle de tre modelarter i nærværende undersøgelse påvirkes af markoperationer i forskellig udstrækning. Som eks. på dette vises nedenstående - med udgangspunkt i sanglærken og insektfødeemner generelt (litteraturdata samt egne data) - en oversigt over de i lærkemodellen anvendte værdier for effekter af forskellige markoperationer (Tabel 3). Hvor data vedrørende de tilknyttede påvirkninger på æg/redeunger/insekter for en markoperation ikke har været tilgængelige i litteraturen, er der for æg og ungers vedkommende tilknyttet en dødelighed på 100 % (angivet med ** i Tabel 3) i de tilfælde, hvor den pågældende markoperation er af en umiddelbar radikal negativ karakter. I øvrige tilfælde er der som udgangspunkt tildelt en dødelighed på 0 %. De angivne dødeligheder på æg og unger som følge af markstrigling stammer fra de resultater (kunstige reder), der er fremkommet i nærværende undersøgelse (samlet gennemsnit for 2004 og 2005). Dødelighed på æg og unger som følge af pesticidsprøjtninger er ikke medtaget under de direkte følger af markoperationer, da disse indirekte påvirkninger (Odderskær et al. 1997) modelleres som påvirkninger gennem en fødereduktion.
Tabel 3. Estimerede effekter (angivet i % dødelighed) af forskellige markoperationer på sanglærkeæg og –unger anvendt i modellen (kilder: Busche 1989, Jenny 1990, Odderskær et al. 1997 b, Daunicht 1998, Odderskær 1998, Poulsen et al. 1998, Wakeham-Dawson et al. 1998, Odderskær et al. 2001), samt insekter (kilder: Sunderland & Topping 1993, Poulsen et al. 1998, Topping & Sunderland 1998, Wakeham-Dawson et al. 1998). For æg og unger angiver de opførte procenter den individuelle dødelighed i det tilfælde de udsættes for den pågældende markoperation. De anførte græsningseffekter på æg/unger er en daglig (tilfældig) dødelighed pr. rede m. æg/unger. For insekter er den angivne dødelighed en gennemsnitlig dødelighed (reduktion) for populationen af insekter i den pågældende mark, der udsættes for den givne markoperation. * : indirekte effekt modelleret gennem en reduktion af insekter (for æg/unger) og plantebiomasse (for insekter). **: angiver, at den pågældende begivenhed er skønnet til at medføre en dødelighed på 100 %, men at den pågældende begivenhed i øvrigt kun i sjældne tilfælde vil finde sted.
| Markoperation | Voksen | Æg | Redeunger | Udfløjne unger | Insekter (generelt) |
| Efterårspløjning | 0 | ** | ** | 0 | 90 |
| Efterårsharvning | 0 | ** | ** | 0 | 70 |
| Efterårstromling | 0 | ** | ** | 0 | 0 |
| Efterårssåning | 0 | ** | ** | 0 | 0 |
| Forårspløjning | 0 | 100 | 100 | 0 | 90 |
| Forårsharvning | 0 | ** | ** | 0 | 70 |
| Forårstromling | 0 | ** | ** | 0 | 0 |
| Forårssåning | 0 | ** | ** | 0 | 0 |
| NPKS | 0 | 0 | 0 | 0 | 0 |
| NPK | 0 | 0 | 0 | 0 | 0 |
| PK | 0 | 0 | 0 | 0 | 0 |
| Flydende ammoniak | 0 | 100 | 100 | 0 | 0 |
| Gylle | 0 | 2 | 2 | 0 | 0 |
| Mangansulfat | 0 | 0 | 0 | 0 | 0 |
| Fastgødning | 0 | 0 | 0 | 0 | 0 |
| Slam | 0 | 0 | 0 | 0 | 0 |
| N-ammoniumsulfat | 0 | 0 | 0 | 0 | 0 |
| Herbicid | 0 | 0 | * | * | * |
| Vækstregulator | 0 | 0 | 0 | 0 | 0 |
| Fungicid | 0 | 0 | 0 | 0 | 0 |
| Insekticid | 0 | 0 | * | * | 80 |
| Radrensning | 0 | 100 | 100 | 0 | 75 |
| Strigling, | 0 | 72 | 72 | 0 | 30 |
| Vanding | 0 | 0 | 0 | 0 | 0 |
| Skårlægning | 0 | 2 | 2 | 0 | 50 |
| Høst | 0 | ** | ** | 0 | 60 |
| Kvæg afgræsning | 0 | 2 | 2 | 0 | * |
| Græsslet til hø | 0 | 100 | 100 | 0 | 60 |
| Græsslet til ensilage | 0 | 100 | 100 | 0 | 60 |
| Halmsnitning | 0 | ** | ** | 0 | 60 |
| Høvending | 0 | 100 | 100 | 0 | 0 |
| Halmpresning | 0 | 0 | 0 | 0 | 0 |
| Afbrænding af stub | 0 | ** | ** | 0 | 60 |
| Stubharvning | 0 | ** | ** | 0 | 75 |
De fleste typer jordbehandling (pløjning, harvning, tromling, såning, stubharvning) er af en så radikal karakter, at de må antages at være forbundet med 100 % dødelighed for æg og redeunger, der bliver udsat for sådanne begivenheder. Mange af disse markoperationer udføres imidlertid på tidspunkter i lærkens yngleforløb, som bevirker, at ingen eller kun meget få reder bliver berørt. Derimod foregår mekanisk ukrudtsbekæmpelse ofte i starten af lærkernes yngleperiode. Af særlig interesse er ukrudtsstriglinger, hvis effekter på højere dyr ikke er beskrevet i litteraturen. Bedømt ud fra resultaterne fra forsøgene med kunstige reder i nærværende undersøgelse er der en stor risiko for, at en rede med æg eller unger vil gå tabt, hvis den fysisk bliver udsat for en eller flere striglinger.
Den samlede effekt af strigling i en markafgrøde vil afhænge af antallet af striglinger, der udføres, deres indbyrdes tidsmæssige placering og af antallet af reder m. æg/unger, der er i den pågældende afgrøde, på de tidspunkter striglingerne udføres. Striglingseffekten på æg og unger stiger med antallet af striglinger udført med tidsmæssige korte intervaller. For et succesfuldt yngleforsøg behøver sanglærken en tidsmæssig periode på mellem 25-28 dage fra det første æg er lagt, til ungerne har forladt reden. Bliver reden ødelagt i første eller andet yngleforsøg, vil en ny rede ofte blive påbegyndt indenfor 4-7 dage. Hvis der i en afgrøde udføres to eller flere striglinger med en indbyrdes tidsmæssig afstand på 8-10 dage, vil chancen for at et ynglepar får ødelagt to på hinanden følgende yngleforsøg indenfor en måned være markant. Hvis et yngleterritorium kun indeholder den ene (striglede) afgrøde, vil resultatet, som ovenfor nævnt ofte være, at et yngleterritorium opgives, da væksten af afgrøden i den mellemliggende periode gør den uegnet som ynglehabitat. Den samlede effekt på bedrifts- eller landskabsniveau vil derudover igen afhænge af yderligere en lang række biotiske og abiotiske faktorer.
Ændring i effektvariabler
Hvis der forligger data for effekten af en bestemt markoperation på f.eks. æg/unger/voksne af en modelart, er det meget let efterfølgende at ændre i disse, hvis det skulle være påkrævet. At fremskaffe sådanne data er som regel forbundet med et stort arbejde at fremskaffe, og i mange tilfælde eksisterer data ikke, og man må efter en ’rundspørge’ blandt specialister anvende kvalificerede gæt. I sådanne tilfælde kan det være nødvendigt at foretage en følsomhedsnalyse af en eller flere sådanne variabler.
1.7 Sædskifter og afgrøder
I modellen gennemløber hver enkelt mark det sædskifte, der er beskrevet og begrundet for forskellige bedriftstyper. I nærværende projekt er der kun opstillet ét konventionelt sædskifte (kvægbrug) baseret på de beslutninger, der blev taget og beskrevet i et tidligere notat til styregruppen (FØL.JORD 12/3/4). Udgangspunktet for det anvendte sædskifte er et sædskifte baseret på de faktiske forhold i området beskrevet under det tværfaglige forskningsprojekt ARLAS (Dalgaard et al. 2004). De modifikationer, der er foretaget i forhold til dette, er gjort for at kunne erstatte herbicidsprøjtningerne i de konventionelle afgrøder med striglinger (tidspunkter for striglingerne taget fra økologisk dyrkede ’søster afgrøder’). De enkelte marker på en bedrift har forskellige afgrøder som udgangspunkt i opstartsåret for modelleringerne. For at kunne simulere forekomsten af afgrøder i forhold til deres arealmæssige forekomst, er det muligt at opstille mangeårige model-sædskifter for den enkelte mark.
Vedvarende græsarealer udgør i scenarierne for kvægbrug 10 %. Dette svarer godt til den procentuelle andel i værkstedsområdet (Dalgaard et al. 2004).
Ændring i sædskiftemodeller
Ændringer sædskiftemodeller kan foretages hurtigt, hvis der ikke er bestemte ønsker om, at disse sædskifter skal være baseret på statistikker for specifikke bedriftstyper i et område, eller at der skal foreligge et grundlag baseret på de enkelte afgrøders specifikke arealandele. Hvis sådanne ønsker derimod foreligger, indebærer det en arbejdsindsats på flere uger afhængig af, om sædskifterne også arealmæssigt skal forbindes til den person der driver de enkelte marker.
Som i foregående afsnit er det for anskuelighedens skyld valgt i de følgende to afsnit at anvende eksempler fra sanglærkemodellen.
1.8 De individbaserede modeller
Udviklingen af individbaserede modeller har givet nye muligheder for realistisk at simulere og modellere dyre- og plantearters (Huston et al. 1988; Hogeweg & Hesper 1990; Grimm 1999; McGlade 1999) økologiske respons på ændrede livsbetingelser i et modellandskab, der (som det er tilfældet i nærværende undersøgelse) kan være en detaljeret kopi af et eksisterende landskab. De enkelte dyrearters biologi er bestemmende for kravene mht. arealskala og detaljeringsgrad og type af arealinformationer der er behov for. Arealskalaen for modelleringen af et større rovdyr vil eksempelvis betyde, at der arbejdes på et landskabsniveau, men ikke nødvendigvis at der er behov for den samme detaljeringsgrad af den arealinformation, der for eksempel ville være nødvendig ved modelleringen af en insektart, hvor arealskalaen typisk vil være på mark- eller bedriftsniveau.
Udgangspunktet for individbaserede modeller er, som navnet antyder, det enkelte individ, for hvilket der er defineret et artsspecifikt sæt af regler, der er bestemmende for dyrets adfærd (eks. sociale interaktioner, energetik, reproduktion, bevægelse og spredning) under tidsmæssigt og rumligt skiftende forhold. Overgangen fra ét sæt af adfærdsregler, der beskriver og styrer begivenhederne i en bestemt periode (”stadie”) af et individs livsforløb, til et nyt stadie er beskrevet af et andet regelsæt (”overgangsstadie”), som er karakteristisk for den givne art (Figur 4). En del af et sådant stadie – overgangs diagram for en hunsanglærke er illustreret i Figur 5. Et komplet sæt af indbyrdes afhængige stadier og overgange mellem disse beskriver den specifikke individ-baserede biologiske model for en pågælden art.
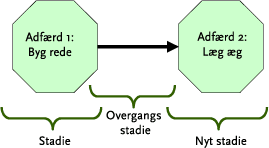
Figur 4. To stadier i en hunsanglærkes liv: hvert stadium og overgangen mellem disse er beskrevet af et bestemt sæt adfærdsregler, der bestemmer hunlærkens beslutninger under de givne omstændigheder.
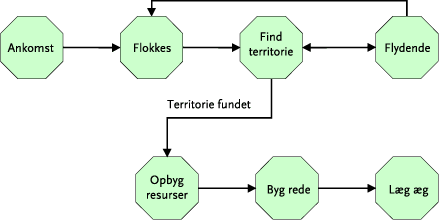
Figur 5. Brudstykke af et ”State/Transition-diagram” for hunsanglærken, der beskriver stadierne fra ankomst til ynglepladsen til påbegyndelse af æglægning. Pilene angiver overgangsstadier.
Modeltypen er karakteristisk ved, at alle individer til et givet tidspunkt i en given lokalitet på samme tid kan få alle relevante informationer om forskellige biotiske (f.eks. afgrødetype, -højde og dækningsgrad, tæthed af fødeemner, tilstedeværelsen af andre individer (f.eks. fra naboterritorier)) og abiotiske forhold (f.eks. pløjning, harvning, sprøjtning, temperatur og nedbør), der er betydende for de enkelte individers reaktioner og handlinger (Figur 6). På denne måde kan et antal individer modtage informationer om, og udnytte eksempelvis en lokal føderesource på en og samme tid. Den lokale population der modelleres, består således af indbyrdes ”kommunikerende” individer, der i tid og rum kan opfatte og reagere på forhold i det lokale miljø. For sanglærkens vedkommende afhænger eks. territoriestørrelsen dels af territoriets kvalitet, dels af den enkelte fugls styrkeforhold i forhold til andre territoriehævdende nabofugle, hvilket medfører en tæthedsafhængig modellering af populationen. De biologiske modeller består af en række undermodeller, der hver især beskriver de regelsæt, der karakteriserer de specielle forhold, der gør sig gældende for eks. hanlærker, hunlærker, ægudvikling, udvikling af redeunger og udvikling af ikke flyvefærdige unger.
En samlet oversigt over den komplekse struktur og sammenhæng mellem den overordnede models delelementer er vist for sanglærken på Figur 6.
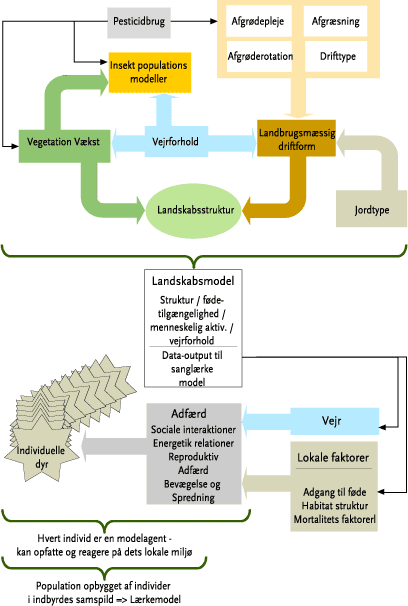
Figur 6. Diagrammatisk fremstilling af de faktorer, der alle påvirker de individuelle dyr, der modelleres i sanglærkemodellen.
1.9 Parameterisering
For at kunne vurdere modellens evne til at ”forudsige” lokale bestandstætheder under givne dyrkningsforhold, blev der for sanglærkens vedkommende i 16 delområder (af ca. 40 ha) indenfor et 10 x 10 km stort område ved Bjerringbro udført moniteringer af lærketætheder (lokalisering af territorier, udparrede hanner mm.) i 1998, 1999 og 2000 (Topping & Odderskær 2000). Efterfølgende er der ved at sammenholde modellens ”forudsigelser” af antal og placering af lærketerritorier og reder i de 16 delområder, hvori der gennem moniteringer forelå eksakte oplysninger om lokale bestandstætheder (herunder territorieplaceringer) i forhold til fysiske parametre (f.eks. tilstedeværelsen af læhegn, markskel, bygninger mm. – Figur 7), foretaget en kontrol og efterfølgende ”justering” af centrale biologiske modelparametre, indtil der blev opnået en generel god overensstemmelse mellem modelforudsigelser og reelle forhold. Modellens ’forudsigelser’ og de faktisk registrerede lærketerritorier i de 16 delområder viste en god overensstemmelse (Odderskær 2005).
Efter den foretagne justering af modeldelen der vedrører rugetid (notat fremsendt til styregruppemødet for nærværende projekt d. 14/11 2005: ’Notat vedrørende justering af modeldel for sanglærkens udrugningstid’) er der gennem de foretagne modelvalideringer af modelsystemet, konstateret en god overensstemmelse mellem faktiske og modellerede forhold.
Ændring i variabler, der indgår i dyremodeller
Genereringen af nye artsmodeller er den selvstændige del, der tager længst tid at udføre. I gennemsnit skal der regnes med ca. 1 års arbejde før en ny model er ’køreklar’. Efter en ny artsmodel er taget i anvendelse, vil der som regel med tiden blive behov for at justere på delelementer af modellerne, alene i kraft af at der med tiden dukker ny viden op på områder, hvor der tidligere kun har foreligget sparsomme eller ingen data. På dette punkt er forholdene ens med de betingelser, der eksistere for andre modelsystemer – der vil med tiden være behov for en løbende opdatering.
Der kan ikke siges noget konkret om, hvor vanskeligt det er at udføre specifikke ændringer i de enkelte modeller. Dette afhænger meget af, hvilken art (simpel/kompliceret model) der er tale om, og hvilke ændringer der er tale om, men det er erfaringen, at der fra det øjeblik det vides, hvordan en konkret ændring skal foretages, vil være behov for mellem en til to ugers arbejde, da der under alle omstændigheder efterfølgende skal foretages en række ’testsimuleringer’ for at sikre, at øvrige centrale variabler i modellen ikke har ændret sig.
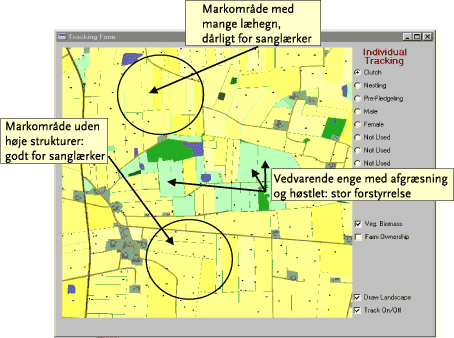
Figur 7. Eksempel på modellens indplacering af lærkepar og reder (blå og røde prikker) i et udsnit af værkstedsområdet. Forskellige tætheder i udvalgte delområder er angivet med pile.
2 Litteratturliste
Bilde, T. & C.J. Topping (2004). Life-history traits interact with landscape composition to influence population dynamics of a terrestrial arthropod: a simulation study. EcoScience 11(1): 64-73.
Busche, G. (1989). Drastische Bestandseinbußen der Feldlerche Alauda arvensis auf Grünlandflächen in Schlewig-Holstein. Die Vogelwelt, 2: 51-58.
Dalgaard, T., Kjeldsen, C., Rasmussen, B.M., Fredshavn, J.R., Munier, B., Schou, J.S., Dahl, M., Wiborg, I.A., Nørmark, P. & J.F. Hansen (2004). ARLAS’ scenariesystem – Et grundlag for helhedsorienterede konsekvensvurderinger af ændringer i arealanvendelsen og landbrugsproduktionen, pp. 97-119, i DJF rapport Markbrug nr. 110.
Daunicht, W.D. (1998). Zum einfluss der Feinstruktur in der Vegatation suf die Habitatwahl, Habitatnutzung, Siedlungsdichte und populationsdynamik von Feldlerchen (Alauda arvensis) in grossparzelligem Ackerland. PhD thesis, University of Bern.
Grimm, V. (1999) Ten years of individual based modelling in ecology: what have we learned and what could we learn in the future. Ecological Modelling 115, 129-148. Elsevier.
Hogeweg, P. & B. Hesper (1990). Individual-oriented Modelling in Ecology, 13 (6): 83-90.
Huston, M., DeAngellis, D, & W. Post(1988). New Computer models Unify Ecological Theory. BioScience, 38 (10).
Jenny, M. (1990). Territorialität und Brutbiologie der Feldlerche Aluda arvensis in einer intensiv genutzten Agrarlandschaft. Journal für Ornithologie, 131: 241-265.
McGlade, J.M. (1999). Induvidual-based models in ecology. In: Advanced Ecological Theory, Principles and Applications (ed. Jacqueline McGlade). Blackwell Science.
Olesen, J.E. & T. Heidmann (1990). EVACROP. Et program til tilberedning af aktuel fordampning og afstrømning fra rodzonen. Version 1.00. AJMET Arbejdsnotat nr. 9.
Odderskær P., Prang A, Poulsen J.G., Elmegaard, N. & P.N. Andersen (1997). Skylark Reproduction in Pesticide Treated and Untreated Fields. Pesticide Research No 32. Ministy Of Environment and Energy, Denmark. Miljø- og Energiministeriet Miljøstyrelsen.
Odderskær, P., Lilleør, O. & Dietz, H.D. (1997). Den vilde faunas forekomst i forbindelse med et udendørs svinehold. Slutrapport for delprojekt ”Den vilde fauna” under ”Nærmiljø og produktionssystemers indflydelse på sundhed i udendørs svineproduktion”, Strukturdirektoratet, Ministeriet for Fødevarer, Landbrug og Fiskeri.
Odderskær, P. (1998). Undersøgelser af sanglærkers levevilkår i agerlandet. Statusrapport for delprojekt Samfunds- og miljømæssige konsekvenser af forskellige strategier for udvikling og udbredelse af økologiske jordbrugssystemer under Det Strategiske Miljøforskningsprogram, Strukturdirektoratet.
Odderskær, P., Cracknell, G. & T. Kroier (2001). Sanglærker og økologisk jordbrug – undersøgelser på Ny Ryomgård 199-1998. Posters til besøgslandbrug Ny Ryomgård.
Odderskær, P. (2005). Notat vedrørende justering af modeldel for sanglærkens udrugningstid – ’Ukrudtsstriglingens effekter på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug’. Notat til styregruppen for ’Ukrudtsstriglingens effekter på markfladens naturlige plante- og dyreliv samt landbrugets ressourceforbrug’ – 31/10/2005.
Plauborg, F. & J.E. Olesen (1991). Udvikling og validering af modellen MarkVand til vandingsstyring i landbruget. Tidsskrift for Plantealvs Specialserie, S 2113.
Poulsen, J.G., Sotherton, N.W. & N.J. Aebischer (1998). Comparative nesting and feeding ecology of skylarks Alauda arvensis on arable farmland in southern England with special reference to set-aside. Journal of Applied Ecology, 35: 131-147.
Sunderland, K.D. & C.J. Topping (1993). The spatial dynamicks of linyphiid spiders in winterwheat. Memoires of the Queensland Museum, 33: 639-644.
Thorbek, P. & Topping, C.J. (2005). The influence of landscape diversity and heterogeneity on spatial dynamics of agrobiont linyphiid spiders: an individual-based model. BioControl 50: 1-33.
Topping, C.J. & K.D. Sunderland (1998). Population dynamics and dispersal of Lepthyphantes tenuis in an ephemeral habitat. Entomologia Experimentalis et Applicata, 87: 29-41.
Topping, C. & P. Odderskær (2000) Udvikling af faunamodeller og landskabsmodel. Statusrapport for delprojekt Arealanvendelse og landskabsudvikling belyst ved scenariestudier – vekselvirkninger mellem natur, jordbrug, miljø og arealforvaltning (ARLAS) under forskningsprogrammet Arealanvendelse – jordbrugeren som lansskabsforvalter, Direktoratet for FødevareErhverv – Forskningskontoret.
Topping C.J. & Odderskær, P. (2004). Modelling the influence of temporal and spatial factors on the assessment of impacts of pesticides on skylarks. Environmental Toxicology & Chemistry 23, 509-520.
Wakeham-Dawson, A., Scoszkiewitz K., Stern K. & N.J. Aebischer (1998). Breeding Skylarks Alauda arvensis on Environmentally Sensitive Area arable reversion grass in southern England: survey-based and experimental determination of density. Journal of Applied Ecology, 35, 635-648.
Version 1.0 Januar 2007, © Miljøstyrelsen.