Miljøprojekt nr. 1172, 2007
Randzoner og andre pesticidfrie beskyttelsesstriber i dyrkede arealer - en udredning
Indholdsfortegnelse
3 Status for udlagte randzoner, nationalt og internationalt
- 4.1 Mikroorganismer
- 4.2 Planter
- 4.3 Nyttedyr
- 4.4 Sommerfugle
- 4.5 Fugle
- 4.6 Betydning af et diverst landskab
5 Viden om randzoner og andre pesticidfrie beskyttelsesstriber
- 5.1 Formål med randzoner og andre pesticidfrie beskyttelsesstriber
- 5.2 Markrande
- 5.3 Bufferzone
- 5.4 Sammenfattende viden om markrand og bufferzone
- 5.5 Insektvolde, græsbælter og blomsterbræmmer inde i markerne
- 5.6 Fugle og andre højerestående dyr
- 5.6.1 Sammenfatning om fugle og andre højerestående dyr
6 Økonomiske aspekter af randzoner
- 6.1 Markrandes effekt på udbyttet
- 6.2 Ukrudt, sygdomme og skadedyr relateret til markrand og bufferzone
- 6.3 Sprøjtefri randzoner som pesticidreducerende virkemiddel
- 6.4 Incitamenter til udlægning af bufferzoner
- 6.5 Økosystemtjenester og eksternaliteter
- 6.6 Sammenfatning af økonomiske aspekter
8 Konkluderende og sammendragende bemærkninger
9 Forslag til forskningsindsats
Bilag A: Skematiske oversigter
Bilag B: Igangværende relevante projekter i Danmark
Bilag C: Igangværende relevante projekter i udlandet
Bilag D: Navne på planter og dyr
Forord
Dette projekt er igangsat på foranledning af Miljøstyrelsens Rådgivende Udvalg for Bekæmpelsesmiddelforskning, som har været stillet overfor forslag om forskning vedrørende brugen af ekstensive randzoner og markstriber som pesticidreducerende foranstaltninger. Imidlertid er de faglige problemstillinger, der knytter sig til randzoner og markstriber, meget omfattende, og det kan derfor være vanskeligt at prioritere en indsats. I denne situation har Det Rådgivende Udvalg fundet det hensigsmæssigt at få tilvejebragt en udredning over eksisterende viden, som kan danne baggrund for forskningsforslag i den kommende udbudsrunde af pesticidforskningsmidlerne, som administreres af Miljøstyrelsen.
Projektgruppen har derfor fået i opdrag af Miljøstyrelsen at sammenskrive en rapport, der med udgangspunkt i pesticidrelaterede problemer:
- Skaber overblik over igangværende indsatser og tilskudsordninger på området.
- Syntetiserer eksisterende viden om ekstensive randzoner.
- Peger på mangler i denne viden og fremkommer med forslag til områder, der bør underkastes videre forskning.
Det skal noteres, at udredningen har været koncentreret om betydningen af pesticider, men at eksisterende viden om ekstensive randzoner ofte er forbundet til anden ekstensiveret drift som f.eks. braklægning, der således også er inddraget indirekte i udredningen.
Det skal også noteres at arbejdet som udgangspunkt har omfattet alle relevante problemstillinger i relation til ekstensive randzoner, men at projektgruppen af praktiske grunde kun i mindre omfang har forholdt sig til grundvandsrelaterede og samfundsøkonomiske spørgsmål.
Projektgruppen har bestået af:
Prof. Peter Esbjerg, Institut for Økologi, Københavns Universitet
Post Doc. Søren Navntoft, Institut for Økologi, Københavns Universitet
Lektor Lene Sigsgaard, Institut for Økologi, Københavns Universitet
Projektgruppen ønsker at takke en række personer for forslag, synspunkter og bidrag. Det gælder primært styringsgruppen bestående af:
Jens Abildtrup, Fødevareøkonomisk Institut, Københavns Universitet
Lars Dinesen, Skov- og Naturstyrelsen
Lene Gravesen, Miljøstyrelsen
Jørgen Jespersen, Institut for Plantebeskyttelse og Skadedyr, DJF, Aarhus Universitet
Jørn Kirkegaard, Miljøstyrelsen
Per Kudsk, Institut for Plantebeskyttelse og Skadedyr, DJF, Aarhus Universitet
Ghita C. Nielsen, Dansk Landbrugsrådgivning, Landscenteret
Marianne Bruus Petersen, Danmarks Miljøundersøgelser, Aarhus Universitet
Desuden vil vi gerne takke for bistand fra Nina Karner og Irene Wiborg.
Sammenfatning
Randzoner, insektvolde, blomsterbræmmer etc. er strukturer, som er blevet etableret med en række formål under fællesnævneren: At begrænse negativ indflydelse af intensiv, industrialiseret landbrugsdrift. Et af de negative fællestræk, som er søgt opblødt, er påvirkningen fra pesticider, især de arealmæssigt meget udbredte herbicider og de mht. giftighed problematiske insekticider. En tilkoblet problemstilling er den strukturelle situation i landbruget i form af en stærk nedgang i små- og liniebiotoper (som f.eks. våde lavninger og skel). De negative følger heraf kan mindskes ved etablering af randzoner og markstriber.
Det kan konstateres, at ekstensive randzoner og marklinier etableres for at opnå naturforbedring, beskyttelse af tilgrænsende arealer (f.eks. vand og hegn) og for at øge naturlig regulering af skadevoldere. Der forefindes en omfattende litteratur om zoner og striber og tilgrænsende emner, men mål, metoder og målte effekter varierer meget. Denne variation giver sig også udslag i manglende terminologisk orden, og der er derfor etableret en oversigt, der også indeholder nye definitioner. ”Markstriber” bruges f.eks. som samlebegreb for alle aflange strukturer, der er placeret i marken. På tværs af alle forskellighederne fremstår, at randzoner og markstriber generelt virker forbedrende på biodiversiteten belyst ved planter, leddyr og fugle. På trods af forskellige dimensioner og tilstande, kan det udledes, at rande kan blive for smalle (1,5 m), og at enkelte fugle får det bedre jo bredere randzoner bliver. Dog kan det ikke udledes hvilke dimensioner af randzoner og randstriber, der giver optimalt resultat i forhold det forbrugte areal.
Mht. nyttevirknig, som især har været fokus for markstriber, er de foreliggende resultater spinkle. Der kan derfor ikke konkluderes noget om, ”hvor meget nytte der opnås i forhold til en given indsats”. Omvendt betyder dette ikke, at nyttevirkning kan afvises snarere, at den varierer fra væsentlig til fraværende og er vanskelig at ”måle”.
For diverse ekstensive randzoner udlagt i relation til tilskudsforordninger indenfor EU foreligger ikke klare begrundelser. Desuden er målene uklare eller fraværende, og der mangler generelt opfølgende undersøgelser af virkninger af sådanne randzoner incl. tvungne sprøjtefrie randzoner langs vandforekomster.
Set i et større perspektiv viser udenlandske undersøgelser, at randzoner og markstriber kan fremme heterogeniteten (habitat- og økosystem-diversitetet) i landskabet og dermed bidrage væsentlig til at forbedre biodiversiteten især i mere monotone landbrugsområder. Overordnet kan det konkluderes, at beskyttelse og forbedring af biodiversiteten er det mest oplagte mål for etablering af randzoner og markstriber. At dette også vil have landbrugsmæssig nyttevirkning kan dog på ingen måde afvises.
Der mangler især viden og dermed forskningsindsats om biodiversitetseffekter relateret til dimensioner og tilstande samt herunder etablering af markører/indikatorer for bestemte biodiversitetstilstande. Det gælder både for randzoner og for markstriber, og for begge typer strukturer mangler også viden om udbytteeffekter, ligesom der savnes sammenligning af randzoner og markstribers effekt mht. både biodiversitetsforbedring og indflydelse på markdrift og udbytter. Mht. især biodiversitet savnes også klarlæggelse af, hvilke effekter der kan opnås ved at skabe bedre sammenhæng vha. linieformede strukturer mellem de ikke-dyrkede småarealer, og hvilke virkninger der kan opnås dels i et større landskab og dels over en lidt længere tidshorisont. Endelig mangler der viden om opfattelse hos befolkning og jordbrugere, viden om konsekvenser for jordbrugerne og operarationel viden for samfundet om mulige virkemidlers effekt og pris af sådanne tiltag.
Summary
Unsprayed field margins, beetle banks and flower strips etc. are different structures, though with a major aim in common: the diminishing of the negative influence of agricultural intensification. This includes pesticide effects and in particular effects of the very widely used herbicides and effects of the very toxic insecticides. The tackeling of these problems also link up to the structural problem of the modern field landscape in which the small non-cropped habitats (e.g. water logged spots and ditches in between fields) are very few. Establishment of field margins and banks/strips may counteract losses of such habitats.
The reasons for establishing field margins and banks/strips are improvement of biodiversity, protection of neighboring habitats (water, hedgerows etc.) and natural regulation of insect pests. The literature on field margins and “strips” is voluminous but the aims, the methodologies and the effects found are highly variable. That variation also includes the lack of a coherent terminology, why this report includes a terminological overview, including some new definitions. “Field strips” for example, are used as the common denomination for all elongate structures placed within the field. Despite all the variation it can be concluded that field margins and strips generally improve the biodiversity assessed using plants, arthropods and birds. Despite the variation in dimension and status it is clear, that buffer margins may be too narrow (1.5 m) and that at least some bird species improve with increasing dimensions of buffer zones.
The question of increasing beneficial effect and hence yield improvement is much related to field strips but coherent results are few. Thus it cannot be concluded how much benefit that can be obtained by a particular quality of field strip in terms of reduced pest attacks and maybe also increased yield.
In a number of countries in EU, economical compensation can be obtained by establishing particular types of buffer zones but published reasons for specific initiatives are missing. Furthermore the targets of different member states are either obscure or missing and generally the effects of buffer zones are not assessed.
In a broader perspective some investigations in other countries demonstrate that field margins and field strips increase the heterogeneity of the landscape (habitat and ecosystem diversity), particularly in uniform landscapes with intensive agriculture. It can therefore be concluded, that a general improvement of biodiversity is the most obvious goal for the establishing of extensive field margins and field strips. Still it cannot be neglected, that this may also imply an improved natural regulation with some reflection in yields.
Overall there is a need for more coordinated research due to lacking knowledge on biodiversity effects related to dimensions and status of margins and strips as well a need for developing markers /indicators for specific levels of biodiversity. This includes buffer margins as well as field strips which also need to be compared in terms of biodiversity benefits and as a potential tool for improving natural regulation of pests. In relation to the biodiversity aspects it would de desirable, on a longer time scale, to investigate the potential of margins and strip as means of coherence in larger fragmented landscapes.
A last under-researched area is the perception / acceptance and economic consequences of such structures: the perception / acceptance of people in general, farmers in particular, and for the society the cost effectiveness of different types and approaches.
1 Indledning
1.1 Baggrund
Randzoner friholdt for sprøjtning med pesticider og siden insektvolde og blomsterbræmmer placeret inde i marker er elementer, der er skabt de seneste ca. 25 år. Bag disse tiltag ligger forskellige ideer til begrænsning af negative effekter af den intensive, industrialiserede landbrugsdrift – med hovedvægt på pesticidpåvirkninger. Med i baggrunden hører også tanker om at modvirke effekter af store ensartede dyrkningsflader, som har betydet at mange populationer af planter og dyr er blevet fordelt på adskilte habitater. For en del planters og dyrs vedkommende danner delpopulationerne i disse habitater tilsammen metapopulationer, som (per definition) er betinget af delpopulationerne og udvekslingen af individer mellem dem. Nogle af de mindre delpopulationer kan uddø i perioder og afhængig af den aktuelle arts spredningsformåen og graden af barrierevirkning i landskabet (f.eks. pga. store marker med korn) kan den pågældende art komme til at mangle over betydelige arealer. Det betyder tab af biodiversitet, og i nogle tilfælde tab eller mindskelse af naturlig regulering af skadegørere. Fragmentering og delpopulationernes negative skæbne påvirkes stærkt af en række driftmæssige operationer, ikke mindst pesticidanvendelse.
De første tiltag for at modvirke tab af biodiversitet og medvirke til bedre forhold for naturlige fjender af skadedyr var de sprøjtefrie randzoner i 1980’erne. Siden er der opstået større opmærksomhed på forringelse af plante- og dyreliv, og især de seneste 10 år har en betydelig europæisk forskning beskæftiget sig med landbrugsdriftens, herunder især pesticidanvendelsens, indflydelse på naturbetingelserne i marker og i det samlede landskab. Overordnede begreber og mål er funktionel biodiversitet, økologiske ydelser og biodiversitet generelt, der indbefatter natursikring og forbedring samt nyttevirkning overfor skadegørere. En del af forskningen peger på den stadigt stigende biologiske monotoni og netop fragmentering af naturelementerne som et gennemgribende problem ved stærkt industrialiseret landbrug. For eksempel peger en britisk stor skala undersøgelse på, at landbrugets tiltag rækker langt ud over de egentlige landbrugsarealer (Baillie et al. 2000) og en anden ny engelsk undersøgelse peger bl.a på vigtigheden af at undgå samme driftstiltag (f.eks. kemisk bekæmpelse eller jordbehandling) på alt for store sammenhængende arealer (Holland et al. 2005). Imidlertid beskæftiger en langt større del af forskningen sig med brydende og ”opblødende” tiltag på mark og til dels landskabsplan. Mest kendt er oprettelse af pesticidfrie randzoner, insektvolde og blomsterbræmmer. En ny tysk publikation peger på landskabsheterogenitet som opblødende faktor overfor virkninger af intensivt landbrug (Tscharntke et al. 2005).
På trods af en del forskning i pesticidfrie randzoner helt fra 1980’erne savnes en sammenhængende, operationel analyse af biologiske og driftsmæssige effekter af randzoners udformning, placering og drift, således at ønsket om øget naturlig regulering og biodiversitet i agerlandet ved anvendelse af randzoner bedst muligt kan forenes med landbrugsproduktionen. Udlægning af pesticidfrie randzoner kan, jfr. analyser fra Det Økonomiske Råd (Det Økonomiske Råd 2004), være et samfundsøkonomisk effektivt middel til at øge biodiversiteten i landbrugsarealer. Det danske landdistriktsprogram giver mulighed for tilskud til ekstensivering af randzoner, f.eks. til 12 m brede sprøjtefrie randzoner langs levende hegn og til braklagte randzoner langs vandløb og søer. Tilskudsordningerne til sprøjtefrie randzoner administreres dog meget forskelligt i EU. I Skotland gives der fx også tilskud til usprøjtede zoner på 1,5 til 6 meter, som placeres i randen af marken, svarende til de danske randzoner, men uden gødskning og uden krav om nærhed til fx levende hegn. Der gives tilskud til insektvolde (som er smalle, græsklædte volde usprøjtede bufferzoner inde i marken) i England. Der er imidlertid begrænset viden om hvorledes disse meget forskellige bufferzoner og de tilknyttede dyrkningskrav påvirker den vilde flora og fauna i marker og hegn. Der er ligeledes begrænset viden om bufferzonernes driftsøkonomiske omkostninger og deres omkostningseffektivitet for samfundet. Den biologiske betydning af at skåne markrande for sprøjtning blev på et vist niveau belyst fra midten af 1980’erne og i 1990’erne. Flere undersøgelser har vist at tætheden af vilde planter i marken aftager fra markkanten og ind mod midten (Marshall 1989; Wilson & Aebischer 1995). Derfor ville det største potentiale for at øge den botaniske diversitet i marker netop ligge fra markkanten og et stykke ind i marken. Tætheden af vilde planter tilknyttet den dyrkede mark er i Danmark som i andre Vesteuropæiske lande faldet markant på trods af en fremgang de senere år (Andreasen & Stryhn upubliceret). Dermed er diversiteten også faldet og nogle vilde plantearter tilknyttet agerlandets dyrkningsflade er i fare for at forsvinde (Stoltze & Pihl 1998). I England havde 80’ernes undersøgelser af sprøjtefrie rande især fokus på jagtbare fugle som agerhøne og fasan (Sotherton 1987; Sotherton et al. 1989; Potts 1986). I Danmark finansierede Miljøstyrelsen den såkaldte randzoneundersøgelse (Hald et al. 1988; Hald & Lund 1994) som belyste usprøjtede 6 m brede randzoners betydning for planter og insekter, herunder nyttefauna med betydning for naturlig regulering af bladlus i korn. Nyligt har en hollandsk undersøgelse af 3 og 6 meter sprøjtefrie rande langs grøfter (de Snoo 1999) påvist, at væsentlige forbedringer for planter og insekter kan opnås med en kun 3 meter bred bufferzone.
Til ovenstående knytter sig en række problemstillinger af sproglig og ikke mindst faglig og effektmæssig karakter. Således er det ofte umiddelbart vanskeligt at være sikker på, præcis hvilken form for ”hjælpestruktur” der er tale om, og ikke mindst hvor sammenlignelige de forskellige tiltag er. F.eks. er der pga. det mere vidtrækkende samspil mellem et hegn og den nærmeste markrand ikke basis for at konkludere direkte fra rande ved grøfter til rande ved hegn mht. bredde. Effekter af dette samspil er dokumenteret af Sparkes et al. (1998), der påviser at kornudbyttet fra markranden og ud i marken afhænger af forekomsten af træer i hegnet. Også muligheden for at fremme regulering af bladlus og andre skadedyr i marker ved etablering af insektvolde og blomsterbræmmer (nektar til svirrefluer og snyltehvepse) er belyst i flere undersøgelser (bl.a. Thomas, Wratten & Sotherton 1992; Sutherland, Sullivan & Poppy 2001; Collins et al. 2002).
I hidtidige danske analyser af pesticidfrie randzoners effekter (bl.a. Hald et al. 1988, Hald & Lund 1994) er der ikke foretaget analyser af forskellige typer af zoner bredere eller smallere end 6 m og effekten af sådanne båndformede zoners effekt i den tilstødende mark. En omkostningseffektiv placering og udformning af bufferzoner kræver viden om de miljømæssige og økonomiske konsekvenser ved forskellige typer af bufferzoner. En sådan viden vil kunne forbedre grundlaget for miljøøkonomiske analyser og udformning af tilskudsordninger, som understøtter en miljøøkonomisk optimal løsning. Flere uafklarede centrale spørgsmål vedrørende randzoner står derfor tilbage:
- Vil f.eks. en yderligere forøgelse af randzonebredden fra de 6 m medføre yderligere og væsentlige forbedringer for flora og fauna?
- Hvis relativet brede ekstensive zoner medfører væsentlige forbedringer ville det være mindre heldigt at holde sig til de 6 m’s bredde, og omvendt ville det være overordentligt nyttigt for beslutningstagere at få påvist, om de 6 m er tilstrækkeligt, eller måske bredere end nødvendigt, til at give både en markant forøgelse af markens naturindhold, og en følelig beskyttelse af hegn og vandløb langs markens rand.
- Ligeledes må man spørge hvordan en kortere- eller længerevarende braklægning af sådanne rande, samt tiltag til fremme af sammenhænge mellem hegn, rande, insektvolde og blomsterbræmmer (konnektivitet) både operationelt og landskabeligt vil virke. Sådanne tiltag kan i henhold til al økologisk teori modvirke negative effekter på biodiversitet i det markdominerede danske landskab.
Sådanne spørgsmål vil knytte sig til overordnede mål om at modvirke biodiversitetsforringelse både i terrestriske og aquatiske miljøer og forbedre naturlig regulering af skadegøre, således at en vis andel af pesticidbehandlingerne i bedste fald kan undværes.
1.2 Formål
Projektets formål er at skabe et overblik over og en syntese af den mangesidede og meget spredte viden, der foreligger om indflydelse på naturbetingelserne i marker og i det samlede landskab ved udlægning af forskellige ”økologisk beskyttende og kompenserende” linjestrukturer i og langs marker. Denne syntese skal munde ud i en udpegning af muligheder for at komme nærmere mod en operationalisering gennem videre forskning som dels kan være sammenfattende og dels fokuseret på enkelte særligt manglende delområder.
Projektet skal ud fra en gennemgang af den eksisterende litteratur, igangværende forskningsprojekter og udredningsprojekter m.m. resultere i en rapport vedrørende eksisterende viden om effekter på natur og landbrugsproduktion ved etablering af usprøjtede/udyrkede zoner i det åbne land på arealer, der hidtil har været dyrkningspligtige. Projektet skal desuden lægge vægt på viden om de nuværende udyrkede arealers påvirkning af de dyrkede arealer med hensyn til dels at fremme biodiversitet og dels fremme af landbrugsmæssig nytte gennem naturlig regulering af skadegørere.
2 Terminologi
En række ord og begreber knytter sig til undersøgelser af hegn og markrande. Især er der i den internationale engelsksprogede litteratur mange relevante og nyttige termer. Nedenfor præsenteres de danske fagtermer som indgår i rapporten (tabel 2.1). Da den nuværende danske terminologi ikke er fuldstændig, indgår forslag til nye danske fagtermer (mærket med *). Disse er så vidt muligt hentet/direkte oversat fra norsk, svensk, tysk eller engelsk. De danske termer anføres her sammen med det/de tilsvarende engelske. Til termerne knytter vi en kort ordforklaring samt, hvor relevant, en litteraturhenvisning til kilde. Dette kan være den originale kilde, eller den kilde, som bedst definerer termen som den anvendes i dag (state-of-the-art).
Begreberne samler sig typisk hierarkisk med et centralt begreb, hvorunder der samler sig begreber, der er delmængder heraf –f.eks biologisk bekæmpelse, herunder klassisk biologisk bekæmpelse, inundativ biologisk bekæmpelse, inokulativ biologisk bekæmpelse og funktionel eller biologisk bekæmpelse. Til underbegreber er, hvor det skønnes nødvendigt, anført det overordnede begreb i ordforklaringen de knytter sig til. Listen er ordnet alfabetisk, for at give den bedste opslagskvalitet. Efter tabellen følger en figur over mark og markrand og de termer der knytter sig hertil (figur 2.1).
Tabel 2.1. Definitioner. Danske fagtermer, alfabetisk ordnet, deres engelske oversættelse, definition samt kildehenvisning – normalt til nyere kilde hvor ordet indgår og defineres eller til første anvendelse (hvor relevant). Da den nuværende danske terminologi ikke er fuldstændig, indgår forslag til nye danske fagtermer (mærket med *).
| Dansk | Engelsk | Definition | Kilde |
| Afgrødebundne arter* | Crop-dwelling species | Arter som kun forekommer i visse afgrøder f.eks kornbladbille i korn. | |
| Afgrødekant | Crop edge | Afgrødens kant. Afgrænser afgrøden mod den øvrige markrand. Typisk smal strimmel på måske 10cm hvor afgrøden står lidt tyndere. | (Marshall & Moonen 2002) |
| Artsdiversitet | Species diversity | Mål for artsantal kombineret med arternes indbyrdes fordeling mht. antal eller masse. | (Wenger K.F. 1984) |
| Barjordsstribe | Sterile strip | En stribe (typisk 1-2m) renholdt jord som adskiller afgrøde fra markrand for at beskytte mod rodukrudt. I barjordsstriber sikres opvækst af 1-årigt ukrudt og f.eks agerhøns kan støvbade. | Bl.a. anvendt af DFFE (www.dffe.dk) |
| Biodiversitet | Biodiversity | Et fælles mål for artsdiversitet, økosystem kompleksitet og genetisk variation. | |
| Biologisk mangfoldighed | Mangfoldigheden af levende organismer fra alle kilder, herunder terrestriske, marine og andre akvatiske økosystemer og de økologiske strukturer, de indgår i; mangfoldigheden omfatter såvel variationen indenfor og mellem arterne som mangfoldigheden af økosystemer. | (Forenede Nationer 1992) | |
| Blomsterbræmme* | Wildflower strip | Stribe på 1-flere m’s bredde af en eller flere blomstrende urter. Kan være placeret i forbindelse med markrand ofte lige indenfor hegnet, men kan også være placeret inde i afgrøden, som f.eks i frugtplantager. | |
| Braklagte randzoner | Fallow strip | Udyrket randzone. | Anv. af Min. f. fødevarer, landbrug og fiskeri (www.dffe.dk) |
| Bufferzone | Buffer zone | Yderste typisk 1-6 m eller mere af afgrøden som ikke behandles med pesticider og kunstgødning eller anden gødning. | www.dffe.dk |
| Bæredygtig udvikling | Udvikling som imødekommer de øjeblikkelige behov uden at gå på kompromis med fremtidige generationers mulighed for at sikre deres behov. | (Brundtlandkommissionen, 1987) | |
| Det multifunktionelle landbrug | Landbrug hvor produktionsmålet er bredere end hidtil, og hvor output i form af traditionelle produkter som fx korn, kød og mælk suppleres med output i form af rent drikkevand, smukke landskaber, alsidig natur, rekreative og kulturelle værdier. |
(Miljø- og Energiministeriet 2001) | |
| Dyrkningsfladen | Arealer i omdrift. | ||
| Ekstensivt dyrket randzone | Buffer zone | Bufferzone dyrket uden pesticider og gødning. | |
| Eksternaliteter (fra landbruget) | Externalities | Landbrugets påvirkninger af eksempelvis vand, jord, biodiversitet, landskab og sundhed, som har betydning for menneskers velfærd, men som ikke har en markedspris, og som der følgelig ikke batales for. | (Pretty et al. 2000) |
| Fodpose | Hedge bottom | Urte- og græsvegetation umiddelbart under hegn. | |
| Forager | Farm track | Område på marken som anvendes til at vende traktoren på. Forageren kan helt eller delvis være sammenfaldende med sprøjtefri randzone, braklagt randzone eller ekstensiv randzone. | |
| Forbindelseslinier i landskabet | Linkages in the landscape | Linieformede strukturer i landskabet, f.eks hegn. Modvirker fragmentering ved at skabe korridorer for migration af flora og fauna. | |
| Fragmentering | Fragmentation | Opsplitning af et habitat i mindre adskilte dele. | |
| Funktionel biodiversitet | Functional biodiversity | Den biodiversitet som fremmer naturlig regulering af skadegørere. | |
| Funktionel biologisk bekæmpelse* | Conservation biological control | Biologisk bekæmpelse, opnået ved at sikre og skabe levesteder for nytteorganismer i og omkring marken. D.v.s genoprette/ fremme funktionel biodiversitet. | Engelske ord defineret af (Eilenberg, Hajek & Lomer 2001) |
| Gennemtrængelighed (når anvendt om dyr)* | Permeability | Andelen af individer fra en given art, som passerer en fysisk barriere som f.eks et hegn. | |
| Hegn | Hedge/ hedgerow | Et (levende) hegn kan defineres som et smalt bælte af vegetation, domineret af en blanding af buske og træer som adskiller et stykke jord fra et andet. | (Dowdeswell 1987) |
| Hegnsbundne arter* | Interstitial species | Arter som er afhængige af hegnet på alle stadier af livscyklus | |
| Inokulativ biologisk bekæmpelse | Inoculative biological control | Udsætning af mindre mængde af nytteorganismer. De udsatte organismer kan opformere sig, og den væsentlige biologiske bekæmpelse forventes fra deres afkom. | (Eilenberg, Hajek & Lomer 2001) (dir overs.) |
| Insektvold | Beetle bank | En plantebræmme, ofte tilsået med tuedannenede græsser, anlagt tværs gennem en mark, typisk på en jordvold for at skabe overvintrings- og levesteder for løbebiller og andre nyttedyr. | Først anvendt af Game Concervancy trust og Univercity of Southhampton |
| Inundativ biologisk bekæmpelse | Inundative biological control | Udsætning af større mængde af nytteorganismer. Den væsentligste biolgiske bekæmpelse forventes fra de udsatte nytteorganismer selv. | (Eilenberg, Hajek & Lomer 2001) (dir overs.) |
| Konnektivitet | Connectivity | Sammenhængen mellem markrande og hegn i et store område (fravær af huller). | (Fry & Robson 1994) |
| Korridor | Corridor | Forbindelse mellem to habitater, f.eks i form af et hegn. | (Forman & Godron 1986) |
| Lineære strukturer | Linear features | Lineære strukturer i landskabet (markrande, hjulspor, læhegn, vandløb m.v.) | |
| Læhegn | Windbreak/ shelter belt | Se hegn. Læhegn betoner hegns betydning som lægiver for afgrøden. | |
| Markgrænse | Barrier | Markens grænse til omgivende landskab. Grænsen markeres af f.eks levende hegn, trådhegn, dige, vandløb. | |
| Markrand | Field margin | Omfatter afgrøderanden og ud til og med markgrænse. Indbefatter derfor også eventuel sprøjtefri randzone, randstribe og forager. | (Dowdeswell 1987) |
| Markstribe* | Samlebetegnelse for insektvolde, græsbælter og blomsterbræmmer inde i marken. | ||
| Matrix | Matrix | Det naturelement i et landskab som dækker den største sammenhængende flade. | (Forman & Godron 1986) |
| Metapopulation | Metapopulation | En gruppe lokale populationer mellem hvilke der kan ske genflow, udryddelser og koloniseringer. | (Levins 1969) |
| Mosaik arter* | Mosaic species | Arter, der anvender flere habitater for at gennemføre livscyklus. Obligate mosaikarter, er afhængige af flere habitater for eksempel hegn og mark, for at gennemføre livscyklus. | |
| Naturkompensationsareal | Nature compensation area | Randstribe med selv- eller udsåede planter anlagt for at fremme naturlig flora og fauna. | |
| Naturlig regulering | Natural control | Biotiske såvel som abiotiske faktorer som regulerer populationer af skadedyr. | |
| Smålevested* | Patch | Små levesteder eller pletter der adskiller sig fra det omgivende landskab enten i kraft af deres ressource (=environmental ressource-patch) eller ved at de er forstyrrede (=Disturbance patches), eller som rester af en tidligere tilstand (=remnant patches). Set fra den enkelte organismes ’synspunkt’ er alt ikke-patch matrix. | (Forman & Godron 1986) |
| Randhabitat* | Non-crop habitat | Overgangszone mellem to habitater. Se også økoton. | Direkte oversat fra tysk |
| Randstribe* | Field margin (strip)/ Boundary strip/ Conservation headland/ Arable (field) margin/ wildlife strip |
En stribe (udyrket) jord mellem markgrænse, f.eks hegn og fodpose, og markkant. En field margin kaldes ”Wildlife strip” (6 m) hvis den er udyrket og kun plejes en gang årligt og ”Conservation headland” hvis der dyrkes (korn) men med reduceret anvendelse af pesticider. | UK Biodiversity Action Plan (http://www.ukbap.org.uk/) |
| Randzone | Buffer zone (ecotone) | Randzoner er dyrkningsfrie områder, der kan fungere som en bufferzone mellem landmandens mark og vandløb eller søer. På den måde beskytter de vandmiljøet mod forurening fra pesticider og næringsstoffer. Randzonerne giver bedre vilkår for de mange dyr og planter, der lever i overgangen mellem mark og vandkant. | Anvendt af Ministeriet for fødevarer, landbrug og fiskeri (www.dffe.dk) |
| Refugium | Refuge habitat | Et område hvor planter eller dyr typiske for et område kan overleve. | |
| Resiliens | Resilience | Hvor hutigt et forstyrret økosystem vender tilbage til dets oprindelige tilstand. | (Holling, 1973) |
| Småbiotoper | Hegn, diger, skel, grøfter, vandløb, rabatter, mergelgrave, mosehuller, småsøer, gravhøje, småbevoksninger, vildtremiser, enkeltstående træer m.m. |
(Agger et al. 1986) | |
| Sprøjtefri randzone | Buffer margin/ buffer zone | Usprøjtet rand af den dyrkede mark. | |
| Stabilitet | Stability | I et økosystem bestemmer enten inerti (modstandsdygtighed mod forandringer) eller resilience systemets stabilitet. | |
| Såede markrande* | Conservation headland | Såede markrande som hverken behandles med pesticider eller med gødning. Disse rande bevares i mindst to år på samme sted. | (Marshall & Moonen 2002) |
| Tilpasningsevne | Adaptive capacity | Den proces som styrer den økologiske resiliens. Beskriver økosystemers kapacitet til ’fornyelse’ af økologisk buffer. | (Gunderson 2000) |
| Trædesten | Stepping-stones | Habitater eller småstumper deraf som muliggør en arts spredning. | |
| Vedvarende habitat | Permanent habitat | Et habitat som ikke udsættes for menneskelig påvirkning | |
| Økologisk fodspor | Ecological footprint | Hvor meget produktivt landareal der skal til at tilfredstille en given befolknings efterspørgsel efter fødevarer, skovprodukter og fossil energi. | |
| Økologisk kompensationsareal | Ecological compensation area | Andel af bedrift allokeret til fremme af diversitet af plante- og dyreliv. Den Internationale Organisation for Biologisk bekæmpelse, IOBC har sat et minimum for en bedrift på 5 %. | (Boller, Häni & Poehling 2004) |
| Økologiske afstande* | Ecological distances | Afstande som har betydning for givne arter eller samspil mellem arter. For eksempel virker arter med lav mobilitet inden for korte økologiske afstande. | |
| Økologiske infrastrukturer | Ecological infrastructures | Ikke-dyrkede stumper, striber, hegn (hjørner, som åbner mulighed for flora og fauna som ikke findes på selve dyrkningsfladen. | Begrebet er introduceret af IOBC-WPRS (Boller, Häni & Poehling 2004) |
| Økosystem | Ecosystem | Et system bestående af mikroorganismer, planter og dyr deres fysisk-kemiske omgivelser om samspillet mellem disse komponenter. | (Tansley, 1935) |
| Økosystem funktion | Ecosystem function | Naturlige processers og komponenters evne til at tilvejebringe varer og tjenester som tilfredsstiller menneskers behov direkte eller indirekte. | De Groot, 1992 |
| Økosystem styring* | Ecosystem management | Aktiv og målrettet manipulation af økosystem for at udnytte dets produktivitet eller fremme dets biodiversitets- og naturbevaringsværdi. | |
| Økosystem tjenester | Ecosystem services | De goder som mennesket får fra et økosystem, dvs fra landbruget bl.a. afgrøder, foder, brændsel samt i større eller mindre grad bidrag til rent vand, ren luft, biodiversitet, naturoplevelser etc. | (Daily et al. 2000) |
| Økoton | Ecotone | En smal overlappende zone mellem forskelligartede samfund (plante- og dyresamfund) ofte forbundne med denne kant. Hegn/mark må betragtes som en kunstigt brat økoton. | |
| Ø-strukturer* | Insular/island features | I agerlandet f.eks. småpletter af vild vegetation, mindre grupper af træer, vandhuller og brakmarker. |
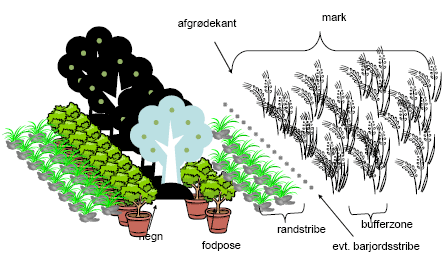
Figur 2.1. Markrand og bufferzone. Markranden består af hegn og hegnets fodpose, eller anden afgrænsning af marken, som et vandløb, af en evt. randstribe, som kan være græsstribe, brakstribe, blomsterbræmme eller ukrudsstribe på 1-6 m eller mere, efterfulgt at en evt barjordsstribe på typisk 1-2m. Den yderste del af afgrøden benævnes afgrødekant. Det er oftest blot nogle få cm, hvor afgrøden står lidt mere tyndt og regnes med til markranden. Hvis en stribe af afgrøden, ikke behandles med pesticider eller med gødning for at skåne miljøet benævnes denne bufferzone (typisk 6 m svarende til at jordbrugeren kører med den yderste sprøjtebom løftet).
3 Status for udlagte randzoner, nationalt og internationalt
3.1 Baggrund
Fra slutningen af 60’erne og op gennem 70’erne ændredes arealanvendelsen i det danske landbrug. Sædskifterne blev væsentligt mere ensidige, og korn, især vårbyg fik en mere dominerende rolle. Større markflader var nødvendige af hensyn til større maskiner. Samtidig steg bedriftsstørrelsen. I 2004 var der godt 45.000 bedrifter over 5 ha i Danmark, hvilket er en halvering på 20 år. Gennemsnitsstørrelsen på en bedrift er nu 58 hektar, langt over gennemsnittet i EU (Danmarks Statistik 2006). Landbruget har også måttet afgive areal til bl.a. byer og veje. I perioden 1990 til 2005 er det samlede dyrkede areal faldet fra 2.788.276 ha til 2.645.304 ha (Danmarks statistik 2006).
Omfanget af biotopnedlæggelsen på landet blev ikke fulgt, men kan i nogen grad følges på topografiske kort. En kortlægning med 5 års mellemrum af småbiotoper i et antal områder fra 1981-1996 sammen med interviews med landmændene i de undersøgte områder, giver et billede af udviklingen i den periode. Som det fremgår af tabel 3.1, som dækker fem test-områder i Vest-Danmark er nedgangen i småbiotoper voldsom, men aftaget i 80erne, med fremgang i 1990erne (Brandt 2004).
Tabel 3.1. Procentvis ændringer pr. år af lineære og areelle biotoper i 5 testområder i Vest-Danmark (20 km²) 1954-1996 (efter Brandt 2004). Brandt (2004) anfører, at der er øgede regionale forskelle i både fremgang og tilbagegang.
| 1954-68 | 1968-81 | 1981-86 | 1986-91 | 1991-96 | |
| Antal år i hver periode | 14 | 13 | 5 | 5 | 5 |
| Lineære biotoper | |||||
| % ændring i længde, pr. år | - 0,6 | - 2,3 | - 1,3 | -1,3 | 0,9 |
| % ændring i areal, pr. år | mangler | mangler | -2,9 | 2,5 | |
| Areelle biotoper | |||||
| % ændring i antal, pr. år | -0,5 | -0,8 | -0,8 | -0,8 | 0,3 |
| % ændring i areal, pr. år | 3,0 | 1,7 | |||
Arealet med dyrkede marker faldt især fra 1991-1996. Det skyldtes primært braklægning. Markstørrelsen steg fra gennemsnitligt 3,8 ha i 1991 til 4,1 ha i 2001, svarende til 1 % årlig stigning. Primært som resultat af marksammenlægninger. Arealer med trædække og med søer og vandhuller steg mellem 1991 og 2001. Især tørre arealer med et vist trædække (f.eks. vildtremiser), øgedes. På landsplan faldt arealet med moser svagt frem til 1996. Op mod halvdelen af de arealer der i 2001 var registreret med trædække og med søer og vandhuller havde samme klassificering i 1981, resten var nye. Også for lineære biotoper er der stor dynamik (Levin & Brandt 2006).
Nedenstående tabel 3.2 (Brandt et al. 2002) viser, hvordan arealbeskyttelsen under naturbeskyttelsesloven har udviklet sig.
Tabel 3.2 Udviklingen i arealbeskyttelse omfattet af Naturbeskyttelseslovens §3 (Efter (Brandt et al. 2002).
| 1937 | 1972 | 1978 | 1984 | 1992 | 2002 | |
| Gravhøje | alle | alle | alle | alle | alle + 2m buffer | alle + 2m buffer |
| Andre fortidsminder | De fleste typer + 2m buffer | De fleste typer + 2m buffer | ||||
| Vandløb | > 1.5 m. bredde | > 1.5 m bredde + udpegede | > 1.5 m bredde + udpegede | +2m dyrknings-frie bræmmer | +2m dyrkningsfrie bræmmer | |
| Søer og vandhuller | Alle naturlige søer | Alle > 1000 m² | Alle > 500 m² | Alle > 100 m² | Alle> 100 m² | |
| Moser | > 5000 m² | > 5000 m² | > 2500 m² | Alle, der ligger i forbindelse med sø | ||
| Heder | > 50000 m² | > 2500 m² | > 2500 m², alene eller sammen med andre naturarealer eller sø | |||
| Strandenge | > 30000 m² | do. | ||||
| Ferske enge | > 2500 m² | do. | ||||
| Overdrev | > 2500 m² | do. | ||||
| Sten og jorddiger | Alle diger + 2 m buffer | (overgået til mu-seumsloven 2004) |
3.1.1 Landbrugsmiljøprogrammer
Forskellige landbrugsmiljøprogrammer sigter mod at beskytte miljøet mod bekæmpelsesmidler og mod næringsstoffer. Uanset fokus bidrager ordningerne til at skabe nye eller forbedrede levesteder for planter og dyr. Det er derfor interessant at kende den samlede tilslutning til ordningerne. I 2006 indgik 7.197 producenter, eller ca. 100.000 ha under MVJ ordningen inkluderet vandmiljøplan I og vandmiljøplan II. Disse 100.000 ha er meget andet end randzoner. Desuden sigter MVJ især på at nedbringe N (og P) i forhold til målene i vandmiljøplanen. Under den gamle økologi-ordning resterede pr. 2006 130 producenter, og under den nye ordning ”Miljøbetinget tilskud” indgik omkring 3.500 producenter eller ca. 155.000 ha (pers comm Henrik Skoven, DFFE). Hermed dækker de to ordninger omkring 8,5 % af det samlede landbrugsareal og over 10.000 producenter. I alt er der i dag 45.267 bedrifter i Danmark, heraf 21.357 defineret som heltidsbedrifter (Danmarks Statistik 2006).
Miljøvenlig forvaltning af landbrugsarealer, støtte til økologisk produktion, og randzoner langs vandløb indgår i landdistriktsprogram 2007-2013. Her integreres støtte til natur og miljø, herunder vandmiljø og biodiversitet med støtte til konkurrenceevne i fødevaresektoren og levevilkår og arbejdspladser i landdistrikterne. Herudover vil der blive tilskud til lokale/regionale natur- og miljøprojekter.
3.1.2 Læhegn og markrande
I 2001 var der i gennemsnit 27 m vej/ha, 20m markskel/ha og 15-20m hegn/ha (Levin & Brandt 2006). I alt er der 96.871 km levende hegn. Ud af det dyrkede areal på 26.453 km² er der 33 m levende hegn/ha (Bruus Pedersen et al. 2006). I et repræsentativt dansk område var der 108 m² mark /m markkant (inklusive levende hegn), eller omregnet 92,5 m markkant/ha (Bruus Pedersen et al. 2006). Ud fra disse tal, udgør levende hegn 1/3 af markkanterne, mens 2/3 er andre typer randzoner, nogenlunde svarende til fordelingen i denne landskabstype for de 15 gamle EU lande (EU-15) som sådan (Eurostat 2003) (se tabel 3.4).
Svagt stigende hegnstæthed i perioden 1991-2001, dækker over både plantning af nye hegn og nedlæggelse af ældre, og da de nyplantede hegn typisk er bredere end de ældre, ville stigningen have været større, om den havde været målt i areal og ikke i længde (Levin & Brandt 2006) Der var en vis generel stigning for både grøfter og vandløb i perioden 1991-1996. Efter en nedgang frem til 1991 i markgrænser ses markant fremgang fra 1991 til 1996, dernæst igen en mindre nedgang, som formentlig knytter sig til de øgede markstørrelser (Levin & Brandt 2006).
Ifølge LUCAS transect data (Eurostat 2003) kan hovedparten af Danmark defineres som ”tidligere åben mark” (Meeus 1995), dog i Nordvestjylland områder med høj tæthed af hække (”Atlantic bocage”), og i Sydvestjylland marsk. Disse landskabstyper har i gennemsnit på tværs af EU-15 hhv 70-80 m randzoner pr. ha, 80-110 m randzoner pr. ha og 50-70 m randzoner pr. ha (Eurostat 2003). Sammenlignet med de danske data er EU data udført på en mere grov skala (satellitfoto), men bekræfter i store træk de danske tal.
Ifølge DFFE er der givet støtte til plantning af over 950 km i 2002, over 600 km i 2003, knapt 500 km i 2004 og forventet plantning af 450 km/år herefter (www.dffe.dk). De reelle tal er formentlig højere, da der også plantes hegn uden ansøgning om støtte. Udviklingen i etablering af læhegn er svagt stigende (Levin & Brandt 2006), det vil sige, at der plantes mere end der fældes.
3.1.2.1 Braklagte, ekstensive og sprøjtefrie randzoner
Samlet var der i Danmark pr. 31. december 2005 under MVJ-ordningen givet tilsagn om 92 ha braklagte randzoner fordelt på 52 tilsagnshavere (Kilde: Direktoratet for FødevareErhverv -DFFE- pers. comm. Henrik Skoven). Herudover er der givet tilsagn til 71 ha ekstensive randzoner og 18 ha sprøjtefrie randzoner pr. 31. december 2005. Det samlede ansøgte beløb var på 300.000 kr.
3.1.3 Vandløb og søer
Der er ca. 64.000 km vandløb i Danmark. Vandløbene og deres randzoner er et vigtigt levested for mange dyr og planter. De målsatte vandløb udgør ifølge Miljøstyrelsens sammenfatning over miljøtilsyn i 2001 25.555 km vandløb.
Som led i evalueringen af pesticidhandlingsplan II i 2002 blev det iflg. Jacobsen (2006) estimeret, at der i randzonerne langs målsatte vandløb og søer var 6.800 ha med brak under hektarstøtteordningen, 2.300 ha med økologisk dyrkning og 140 ha sprøjtefri randzone under MVJ-ordningen - i alt 9.240 ha.
For vandmiljøplan III er det aftalt at der i perioden 2005-09 skal etableres 3-4000 ha vådområder og 30.000 ha dyrkningsfrie randzoner langs vandløb og søer (Jensen 2006).
I pesticidhandlingsplanen 2004-2009 er målet 25.000 ha sprøjtefri randzoner langs målsatte vandløb og søer inden 2009. I udgangspunktet i 2004 blev det antaget, at der allerede var udlagt 8.000 ha randzone som ikke blev sprøjtet fordi de var braklagte/udyrkede eller økologiske. De 8.000 ha lå alle langs målsatte vandløb og søer. Antages randzoner at være 10 m brede, vil den målsatte forøgelse svare til 25.000-30.000 km randzone.
Tabel 3.3. Estimerede randzonearealer langs vandløb og søer i Danmark, med forbehold for lille undersøgelsesstørrelse (29 landmænd) (Efter (Jacobsen 2006)).
| Totalt randzoneareal ved 10 m zone (ha) | Dyrket randzoneareal (ha) (50 % af totalarealet) | Dyrket randzoneareal i alm. omdrift (ha) | |
| Målsatte vandløb | 50.000 | 25.000 | 10.000 |
| Ikke-målsatte vandløb | 78.000 | 39.000 | 20.000 |
| Søer over 100 m² | 20.000 | 10.000 | 5.000 |
| Total | 148.000 | 74.000 | 35.000 |
Det potentielle omfang af randzoner på arealer i alm. omdrift langs vandløb og søer er mindst 35.000 ha (tabel 3.3) og korrigeret for en større andel lavbundsarealer i analyseområdet er det ca. 40.000 ha (Jacobsen 2006).
3.1.4 Nuværende regulering
3.1.4.1 Lovgivning om beskyttelseszoner mod bekæmpelsesmidler (afstandskrav)
I landzone skal der være en sikkerhedsbræmme på 2 m langs naturlige eller højt målsatte vandløb og søer. Sikkerhedsbræmmerne beskytter blandt andet mod pesticider.
En række pesticider må ikke benyttes indenfor en afstand på 10 eller 20 m fra vandløb (Dansk Landbrugsrådgivning 2004). Midler omfattet af afstandskrav omfatter både insekticider, herbicider og fungicider. Afstandskrav fastsættes ud fra en risikovurdering af toxicitet overfor akvatiske organismer (fisk, krebsdyr og alger) (Miljøstyrelsen 1999).
Sprøjtefri randzoner skal i henhold til MVJ ordningerne være 10 m brede, når de ligger langs vandløb og søer, hvor der er krav om en 2 m dyrkningsfri bræmme. Når randzonen ligger langs landskabselementer, hvor der ikke en krav om en dyrkningsfri bræmme på 2 meter skal randzonen være 12 m brede. Sprøjtefrie randzoner må ikke tilføres plantebeskyttelsesmidler, mens sprøjte- og gødningsfrie randzoner ikke må tilføres plantebeskyttelsesmidler og anden gødning, end den der afsættes af græssende husdyr.
3.1.4.2 Tilskud til ekstensive og braklagte randzoner
Via MVJ-ordningerne kan man ud over støtte til randzoner langs vandløb og søer søge tilskud til: a) Ekstensive randzoner, b) Braklagte randzoner. Administration af MVJ ordningerne overgik pr. 1/1 2007 fra amter til Direktoratet for FødevareErhverv, hvor også nærmere information om ordningerne kan findes. I 2006 kunne man kun søge tilskud til braklagte og ekstensive randzoner langs søer og vandløb. Da alle målsatte vandløb er udpeget til SFL-områder, kan man søge til alle målsatte vandløb (Anon. 2006).
Ekstensive randzoner (5 årige tilsagn) er fortsat dyrkede. Den ekstensive randzone skal være 10 meter bred, når randzonen er beliggende langs en 2 meter dyrkningsfri bræmme (vandløbsloven §69) eller 12 meter bred, når randzonen er beliggende langs andre landskabselementer. Også arealer der er beliggende langs særlige landskabselementer f.eks. vandløb, søer, levende hegn, diger, naturarealer m.m. kan indgå i ordningen. Der må ikke tilføres plantebeskyttelsesmidler i ekstensive randzoner, med undtagelse af bejdset udsæd og rodhalsbehandlet udplantningsmateriale. Langs søer, vandløb og kystlinier gælder, at udnyttelsen skal ske under størst mulig hensyn til at undgå jorderosion.
Braklagte randzoner skal være 10 meter brede og være udlagt med tæt, lavt plantedække, som skal afpudses minimum en gang årligt. Her er tilsagnsperioden fem år. Arealer der er placeret umiddelbart op til søer og vandhuller kan indgå i ordningen. Ordningen kan både benyttes på arealer, der ligger udyrket hen, og på arealer, der skal bruges til at opfylde udtagningspligten og kan så opnå både enhedsstøtte og MVJ-tilskud til arealet.
For begge typer randzoner gælder at arealerne forud skal have været anvendt til dyrkning af et eller flerårige afgrøder, til græsning, slæt eller braklagt indenfor omdrift. Tilskud til ekstensive randzoner er 690 kr/ha om året. For braklagte randzoner er det 750 kr./ha (www.dffe.dk). Desuden kan arealerne opnå enhedsstøtte. Det er tilladt ind mod den dyrkede mark at lægge en barjordsstribe på højest 2 m, som kan holdes fri for plantedække gennem hele vækstsæsonen.
Den 2 meter dyrkningsfrie bræmme kan indgå i MVJ-ordningen og kan via MVJ-ordningen medregnes ved beregning af udtagningspligten. For randzoneordningerne gælder min. størrelsen for et areal på 0,30 ha ikke.
Ifølge hektarstøttereglerne er det tilladt, at braklægge helt ned til en 5 meter zone langs søer, vandhuller og vandløb som altid er permanent vanddækkede.
Jordbrugere, der dyrker arealer uden brug af plantebeskyttelsesmidler og med begrænset anvendelse af kvælstof, kan søge tilsagn om miljøbetinget tilskud, som trådte i kraft 1. november 2003, som sammen med ordningen for tilskud til omlægning til økologi med miljøbetinget tilskud erstatter den hidtidige ordning med tilskud til økologisk jordbrugsproduktion (DFFE, 2006).
3.1.4.3 Tilskud til plantning af læhegn
”Bekendtgørelse om tilskud til lægivende og biotopforbedrende beplantninger” (læplantningsordningen) giver 40 % støtte til plantning af læhegn, men en højere sats på 60 % når krav til et "særligt miljøindhold" opfyldes. Man binder sig for 15 år og må ikke anvende andre plantebeskyttelsesmidler, end de der er tilladt i økologisk jordbrugsproduktion. Herudover skal man opfylde mindst tre af følgende betingelser: a) En mindst 1 m bred udyrket stribe jord (en randstribe) langs hegnet som mindst en gang hvert andet år pløjes, harves, fræses eller lignende, b) En insektvold i tilknytning til beplantningen og på de tilstødende marker, c) Beplantningen i mindst tre planterækker med åbninger for hver 50 m, d) En trampesti, hvorpå offentligheden kan færdes, og hvortil der er lovlig adgang, e) I beplantningen må der udelukkende anvendes hjemmehørende plantearter og provenienser.
De nævnte tiltage og viden om deres betydning for flora og fauna og begrænsning af pesticidanvendelsen gennemgåes nærmere i følgende kapitler. Tilskudsordningen til læhegn er midlertidig sat i bero, men forventes udbudt i 2007 (Pers comm. Irene Wiborg, jan 2007).
3.2 Internationalt
Ifølge LUCAS transect data findes der 8 millioner km linieformede strukturer i EU-15 (hegn, rækker af træer, græsstriber, stendiger, insektvolde), heraf adskiller 3 mio km marker, og 5 mio km afgrænser marker mod andre landskabstyper (f.eks. veje, byer) (Eurostat 2003) (tabel 3.4). Trods strukturernes vigtighed findes der kun begrænset data vedrørende deres status og udvikling. Ovenstående data over markrande og randzoner i EU-15 giver indblik i den nuværende status, men ikke i udviklingen, eller hvor stor en andel der er udlagte.
Tabel 3.4. Markrande og randzoner i EU-15. (Eurostat 2003).
| Type | Mellem marker | Mellem mark og anden landskabstype | |||
| Bredde | Estimeret længde i km | % | Estimeret længde i km | % | |
| Græs | <3m | 449 419 | 15 | 1 835 507 | 36 |
| >3m | 109 058 | 4 | 382 342 | 7 | |
| Buske/hegn | <3m | 946 490 | 31 | 1 063 789 | 21 |
| >3m | 672 881 | 23 | 887 293 | 18 | |
| Stendiger, trærækker o.lign | 694 737 | 22 | 454 581 | 8 | |
| Kombinerede linieformede strukturer | 156 288 | 5 | 524 424 | 10 | |
| Total | 3 028 873 | 5 147 935 | |||
3.2.1 Landbrugsmiljøprogrammer
Randzoner er et integreret element i mange EU-landes landbrugsmiljøprogrammer. I 1997 blev det opgjort at der samlet i EU blev anvendt 1,3 milliarder euro til landbrugsmiljøprogrammer. Fra 1992-2003 er der brugt 24 milliarder Euro på landbrugsmiljøprogrammer (Wittingham 2007). I alt gives der i dag EU og national støtte i EU-15 svarende til 3,5 milliarder euro om året (European Commision, 2005). På EU niveau omfatter landbrugsmiljøprogrammernes ordninger i alt ¼ af landbrugsarealet, svarende til hele Tysklands areal, men vil formentlig reduceres med 1/5 i 2007 (Whitfield 2006). I Danmark lå arealet i 2002 på ca. 12 %, i Holland under 10 %, i Storbritannien på 15 %, i Tyskland på 25 % og i Sverige på over 80 % (European Commision, 2005).
Landbrugsmiljøprogrammerne samfinancieres af medlemslandene og EU gennem EAGGF (= European Agricultural Guidance and Guarantee Fund). Den nationale implementering sker indenfor ret vide rammer. Overordnet er der to miljøhensyn i landbrugsmiljøprogrammer: a) At reducere den negative indflydelse fra landbrug på miljøet, især mht vandkvalitet, jord og biodiversitet b) at fremme en jordbrugspraksis der sikrer en bevaring af biodiversitet og landskab, herunder at undgå nedbrydning eller problemer fra underudnyttelse. De hovedelementer der kendetegner landbrugsmiljøprogrammer er:
- Landbrugere leverer en miljøtjeneste
- Aftalerne er frivillige for landbrugerne
- Tiltagene gælder kun landbrugsjord
- Betalinger dækker mistet indtægt, ekstraomkostninger og nødvendig yderligere økonomisk tilskyndelse
Tiltagene kendetegnes desuden ved at være mere vidtrækkende end god jordbrugspraksis (Good Agricultural Practice – GAP). I nogle tilfælde er tiltagene nationale, i andre regionale eller lokale (European Commision. 2005).
Mens for eksempel Sverige anvender over 80 % af EU støtten til landbruget under EAGGF ligger Tyskland, Storbritannien og Danmark på omkring 50 % og Holland på knapt 20 % i gennemsnit for perioden 2002-2003 (European Commision. 2005).
Opgjort i %-del areal under forskellige landbrugsstøtteordninger ligger Danmark højt i forhold til økologisk jordbrug, som modtager mere end 45 % af den samlede støtte mod 7-8 % i snit for EU-15. Til gengæld ligger landet lavt med hensyn til støtte til input-reduktion (< 25 % mod 35 %). Andre væsentlige ordninger i EU-15 vedrørede landskab og natur (25 %), ekstensivering (>5 %) og sædskifte (2-3 %) anvendes ikke i Danmark (European Commision. 2005).
3.2.2 Randzoner
Støtte til etablering af randzoner er udbredt. Ordningerne for en række lande blev opsummeret af det engelske fødevareministerium ”DEFRA” i 2001 (www.defra.gov.uk). Nedenstående gennemgang støtter sig til denne men med opdateringer, hvor det har været muligt. Der er væsentlige forskelle i fokus og betydning af disse ordninger landene imellem. For eksempel fokuserer finske ordninger på vandløbsbeskyttelse (Heikkilä 2001;Wallenius & Sihvonen 2000). I nogle lande, som Schweiz er kompensationer ifm med støtte til randzoner og andre naturarealer blevet af stor betydning i den enkelte landbrugers økonomi (Duelli & Obrist 2003). Her er det nu ikke muligt for landmænd at modtage landbrugsstøtte hvis de ikke allokerer mindst 7 % af deres areal til økologisk kompensationsareal.
Hovedvægten i gennemgangen af udenlandske ordninger er lagt på de lande der geografisk og landbrugsmæssigt ligger Danmark nærmest; Sverige, Tyskland, Storbritannien og Holland.
De forskellige ordninger i Storbritannien baseres alle på individuelle kontrakter med avlere, og for de fleste er kunstgødning og sprøjtemidler ikke tilladt. Avlerne modtager kompensation baseret på areal af omkranset mark for randstribe, græsstribe og billevold og per løbende meter for wildlife strips. Fra 2005/6 kan nye avlere ikke deltage i Countryside Stewardship Scheme. I stedet er oprettet Environmental Stewardship Scheme, (http://www.defra.gov.uk/erdp/schemes/css/default.htm). Især et punkt adskiller den fra tidligere ordninger, nemlig opdelingen mellem ”entry level stewardship” og det såkaldte ”higher level stewardship”, hvor de enkelte avlere kan vælge mellem at deltage i bevarelse, genopretning eller etablering af naturarealer.
I Tyskland er der programmer for beskyttelse af markrande i 7 forskellige delstater. Ordningerne omfatter bufferzoner (Schonstreifen), randstriber (Acker-randstreifen), markstriber (Saum- und Bandstrukturen) samt blomsterbræmmer og blomsterflader (Blühstreifen, Blühflächen). Som i de britiske støtteordninger er bredden på markranden og de specifikke dyrkningskrav variable, men som hovedregel tillades kunstgødning og pesticider ikke, eller ikke uden særlig tilladelse. Ved den ekstensive drift i markrand ønskes at fremme og beskytte hjemmehørende flora. De tyske landbrugsmiljøordninger frem til 2005 er gennemgået i Hartmann et al. (2006). I 2005 havde alle tyske forbundsstater landbrugsmiljøprogrammer undtagen Berlin og Mecklenburg-Vorpommern (MV). Hovedvægten ligger på jordskånende og erosionsbeskyttende tiltag hhv. regler vedr. økologisk jordbrug (Hartmann et al. 2006). For økologisk jordbrug har også MV et program, som bl.a fordrer at 3 % af arealet på et økolgisk jordbrug udlægges som blomsterbræmmer/ -flader og græsarealer ikke slås før 1. juli for at udløse støtte. Den tyske stat medfinancierer landbrugsmiljøprogrammer i forbundsstaterne under følgende ordninger: a) Mindst 5 landbrugsafgrøder i sædskifte, b) Dyrkning af efterafgrøder og udlæg, c) Anvendelse af jorddække eller direkte udsåning eller af jorddækkeplanter, d) Udbringning af flydende gødning med særligt miljøvenligt udbringningsudstyr, e) Udeladelse af herbicider i flerårige kulturer og træplanteskoler, f) Anlægning af blomsterflader eller blomsterbræmmer på op til 15 % af det dyrkede areal og g) Anvendelse af biologiske eller biotekniske bekæmpelsesmetoder i plantebeskyttelsen (Hartmann et al. 2006).
Nye regler gældende fra 2007 er pt. under udarbejdelse i Tyskland, hvorfor et samlet overblik er næsten umuligt. Nyeste informationer findes på hjemmesider fra de enkelte forbundsstater. En væsentlig ændring bliver tilsyneladende at kun tiltag vedr. markrande der ligger langs vandløb fremover vil kunne få støtte (Horst Steinmann, Pers. com.).
For at beskytte plante- og dyreliv findes regler om afstandskrav til biotoper (hegn, beplantninger og lignende) ved sprøjtning. Kravene er både afhængige af forekomsten af biotoper i området i forvejen (et "biotopindex" er udregnet for hele Tyskland) og afhængige af pesticidforbruget (afgrøder) i området (BMVEL 2005). Kun ukrudts- og insektmidler er mærket, mens svampe- og vækstreguleringsmidler kun undtagelsesvis er mærket (G. Cordsen, pers. komm.).
I Sverige har man siden 1996 udarbejdet naturplaner på bedriftsniveau. Under de nye svenske arealstøtteprogram indgår bufferzoner som arealer der kan modtage støtte, ligesom det gjorde under det tidligere program (kilde: Jordbrugsverket, www.sjv.se).
I Holland har man hidtil fokuseret på naturbeskyttelse i særlige områder (Ecological Main Structure), adskilt fra landbrugsområder. I landbrugsområder har man fulgt Nature Policy Plan NPP fra 1990 som sætter minimum for miljø og naturkvalitet her. Der er i Holland en holdningsudvikling mod integrering af naturhensyn i landbruget. Man satser især på udvikling og anvendelse af støtteordninger (Wossink & Wenum 2003). På engarealer og græsmarker fokuseres på fuglebeskyttelse og på at bevare digebredder, på dyrkede marker fokuseres på markrande og alternativ pasning af brakmarker. Samlet deltog i 2000 3 % af de hollandske planteavlere og 8 % af husdyrbrugere i diverse nationale og regionale ordninger vedr. bevarelse af biodiversitet (Wossink & Wenum 2003).
En række ordninger fra europæiske lande findes samlet i skemaform i bilag A (tabel 1). Skemaet er ikke komplet, da der ikke altid har været komplette oplysninger. Nogle af ordningerne der indgår, er ved at blive udfaset, andre er nye. Arealerne, når de kendes, er variable. Aftalenes varighed varierer og ligeledes aftalernes indhold.
3.3 Reguleringens vidensgrundlag
Der mangler stort set forskning til at påvise de positive effekter af landbrugsmiljøprogrammer på europæisk niveau (Whitfield 2006). En meta-analyse kaldet EASY, som gennemgik fem landbrugsmiljøprogrammer, har givet anledning til en del diskussion. Alle fem programmer byggede på parvise sammenligninger i marker som var med i en landbrugsmiljøordning eller udenfor. De undersøgte programmer var a) et schweizisk program, ”Ùkoausgleichsflächen”, med ekstensiv græs uden kunstgødning og pesticider sammenlignet med konventionelt græs, b) et tysk program for økologisk jordbrug, hvor økologiske hvedemarker blev sammenlignet med konventionelle, c) et spansk program for bevaring af en sjælden steppefugle gennem ekstensiv drift af kornmarker, d) et hollandsk program for bevarelse af engfugle med forsinket slæt på græsmarker og e) et britisk program med 6 m brede græsrande vs. marker med konventionel brug af markrande. Meta-analysen viste positive effekter af landbrugsmiljøprogrammerne på 12 ud af 25 kategorier af organismer. Højere plantediversitet kunne påvises i alle lande undtagen Schweiz. I Tyskland og Schweiz kunne påvises fordele for bier. I England påvistes fordele for græshopper og fårekyllinger og i Spanien for edderkopper. I de tilfælde hvor biodiversiteten forøgedes var det næsten altid almindelige arter, bortset fra i det spanske program, hvor en truet fugleart blev fremmet (Kleijn et al. 2006). På denne baggrund var EASY-rapporten meget kritisk overfor, om effekten af disse ordninger på biodiversitet stod mål med indsatsen. Problemet med EASY-rapporten er imidlertid, at biodiversitet ikke var et planlagt mål i alle de undersøgelser, der blev udvalgt, og at de derfor heller ikke kan evalueres på den baggrund. For eksempel er græsstriber i markranden, som er meget udbredt i England og udgjorde en del af undersøgelsen, aldrig blev gennemført med henblik på at beskytte sjældne arter.
En undersøgelse af effekten af det irske jordbrugmiljøprogram sammenlignede, i mangel af et baseline-studie, bedrifter som fik støtte med en tilsvarende gruppe, som ikke fik støtte. En af forskellene var, at de støttede brug, i kraft af støttereglerne, havde randstriber, der var lidt bredere (omkring 2 m brede) end kontrolbrugenes randstriber (omkring 1,5 m brede). Hverken plantediversiteten eller diversiteten af løbebiller var signifikant påvirket af randstribebredden. Dog var der højere tæthed af løbebiller i de bredere randstriber (Feehan, Gillmor & Culleton 2005). Den irske undersøgelse viste desuden, at visse elementer af støtteordninger var mindre heldige. For eksempel var støtte til faste hegn som beskyttelse af vandløb problematisk, da få høje arter, i fravær af græsning, kunne skygge andre plantearter ud (Feehan, Gillmor & Culleton 2005). Undersøgelsen satte desuden spørgsmålstegn ved støtte til at lave huller i hegn, da hullerne påvirkede løbebilletætheden negativt (Feehan, Gillmor & Culleton 2005). Endelig pegede undersøgelsen på, at langlivede variable som hegn bedre forklarede forskelle mellem individuelle bedrifters biodiversitet end om bedrifter indgik i en jordbrugsmiljøordning.
Når jordbrugsmiljøprogrammerne ofte har et begrænset resultat på biodiversiteten, er det formentlig fordi tiltagene sker på for små og isolerede arealer f.eks. på hegnsstumper a 50 m (Wittingham 2007). Wittingham fremhæver, at tiltagene skal ske på hele bedrifter og helst på flere tilstødende bedrifter samtidigt for at maksimere effekten på biodiversiteten.
En analyse af de studier der indgår i review fra 2003 (Kleijn & Sutherland 2003) viser at blot 26 % var blevet publiceret internationalt og kun 34 % havde baseline data (Sutherland 2004).
Der er ikke tvivl om, at det er problematisk at mange undersøgelser af landbrugsmiljøprogrammer er gennemført uden baseline studier (dvs. fastlæggelse af situationen før gennemførelsen) og at for få er videnskabeligt publiceret.
3.4 Sammenfatning
I Danmark modtager omkring en femtedel eller over 10.000 producenter enten MVJ eller MB støtte. Ordningerne dækker 8-9 % af landbrugsarealet. For en del af ordningerne er fokus primært på næringsstoffer.
Tilskud til plantning af læhegn og etablering af randzoner og bufferzoner har bidraget til øget etablering af disse strukturer. Tilskuddets størrelse er medvirkende faktor for hvor stort areal, der henlægges. Tilskuddet er i visse tilfælde lille i forhold til ansøgningsbyrden, således at der mangler økonomisk incitament til at søge (se kapitel 6).
Der er gavnlige effekter af landbrugsmiljøprogrammer og af randzoner, men yderligere dokumentation savnes. En sådan dokumentation kræves for at kvalificere vurderingen af værdien af forskellige ordninger på forskellige parametre.
Der er væsentlige forskelle i de nationale støtteordninger, men med en bevægelse mod større ensartethed indenfor EU landene. For eksempel er der tegn på en udvikling i støtte til bufferzoner hvor disse indgår i en bredere miljøkontekst. I Danmark gennem landdistriktsprogrammet. I Sverige tilsvarende gennem landsbygdsprogrammet. Et andet eksempel er den nye engelske ordning Environmental Stewardship Scheme, hvori der indgår støtte til både bevarelse, genopretning og etablering af naturarealer.
Endelig er der tendenser til nedskæringer i støtten. Dette ses blandt andet i Tyskland. Manglende dokumentation af effekter kan være en væsentlig hæmsko for udviklingen af bufferzoner og kan direkte bidrage til mindre økonomisk støtte i fremtiden.
4 Biodiversitet i agerlandet
- 4.1 Mikroorganismer
- 4.2 Planter
- 4.3 Nyttedyr
- 4.4 Sommerfugle
- 4.5 Fugle
- 4.6 Betydning af et diverst landskab
Selv om naturreservater og beskyttede områder altid vil være vigtige, er det nødvendigt i stigende grad at rette opmærksomheden mod bevarelse af biodiversitet i dyrkede systemer (Edwards & Abivardi 1998).
Effekter på flora og fauna afhænger af småbiotopens habitatkvalitet, størrelse, alder (stabilitet), form samt afstand til andre tilsvarende habitater. Disse effekter udfolder sig forskelligt på forskellige organismer afhængigt af disse organismers karakteristika og med hensyn til skala særligt deres mobilitet.
I kapitlet gennemgår vi væsentlige organismegrupper med forskellige karakteristika, og hvordan disse påvirkes af ændringer i linieformede habitater. Betydningen af skala er beskrevet i afsnit 4.3 om nyttedyr.
Mikroorganismer og planter har stor betydning for et velfungerende økosystem. Nyttedyr som svirrefluer og løbebiller har man ofte søgt at fremme ved etablering eller ændringer af linieformede strukturer for at øge deres gavnlige effekter. En anden vigtig gruppe, som har fået en del opmærksomhed bl.a. på grund af deres oplevelsesmæssige værdi, er sommerfugle. Endelig er fuglene en velundersøgt gruppe, der ofte bruges som bioindikatorer i agerlandet (Ormerod & Watkinson 2000). Fuglene har interesse både af hensyn til jagt og som naturelement.
4.1 Mikroorganismer
Mikroorganismer omfatter bl.a. svampe, bakterier og vira som er afgørende for naturens stofkredsløb. Dette afsnit fokuserer på svampe, som påvirkes direkte af fungicider, og derved er en interessant gruppe i relation til sprøjtefrie randzoner. Svampe er en vigtig del af økosystemet specielt for nedbrydningen af organisk materiale. Svampe optræder både som nytteorganismer, der bl.a. bruges ved bekæmpelse af skadedyr (insektpatogene svampe), og som sygdomsfremkaldende organismer som f.eks. meldug og knækkefodsyge, der angriber afgrøderne. Fungiciderne bekæmper naturligvis de skadelige svampe, men det er sandsynligt, at de samtidig også påvirker de nyttige svampe. Der findes dog meget lidt viden på området. En undersøgelse af fungiciders effekt på nyttige mykorrhizasvampe, der lever i samliv med rødder, viste ikke umiddelbart negative effekter på svampene (Jakobsen & Rosendahl 2006). En af de vigtigste insektpatogene svampe Beauveria bassiana forekom hvor som helst i et marksystem med levende hegn (Meyling 2005). Imidlertid rummede læhegn en meget større genetisk variation af insektpatogene svampe end marken (Meyling 2005). Det er derved muligt, at ekstensivt drevne randzoner kan øge den naturlige pulje af nyttesvampe i marken.
Logisk set må fungicider også ramme fungivore mikro- og makroarthropoder, idet svampbelægningerne de ”græsser” på fjernes. Nedgang i forekomsten af disse mindre ”svampegræssere” kan påvirke forekomsten af bl.a. løbebiller, som præderer på disse, og derved kan effekten af fungiciderne forplante sig gennem fødenettet. Disse forhold mangler at blive belyst både generelt og i relation til ekstensive randzoner.
4.2 Planter
Vilde planter i agerlandet kan opdeles i planter, der lever på ikke-dyrkede habitater f.eks. i levende hegn, og planter på selve marken som ofte kategoriseres som ukrudt. Ukrudt er problematisk for landbrugsdriften, men er samtidig en vigtig del af markens økosystem, idet det giver levesteder og føde for insekter og andre leddyr samt fugle og pattedyr. Der findes ca. 200 vilde plantearter på de danske marker, som kan opdeles i én- og tokimbladede arter, én-, to-, og flerårige arter samt frøformerende og vegetativt formerende arter, hvor specielt sidstnævnte kan være problematiske for landbruget.
Andreasen et al. (1996) og Andreasen & Stryhn (upubliceret) undersøgte 67 almindelige arter i sammenlignelige undersøgelser på konventielle brug i 1969-70, 1987-89 og i 2001-04 i forsøg, hvor der ikke blev udbragt herbicider i undersøgelsesperioderne. Fra slutningen af 60’erne til slutningen af 80’erne viste resultatet et fald i ukrudtshyppigheden på 60 %, og specielt arterne rød arve, alm. markarve, hønsetarm, burre-snerre, glat vejbred og nat-limurt var gået tilbage i antal (Andreasen et al. 1996). Andreasen & Stryhn (upubliceret) sammenlignede herefter perioden fra slutningen af 80’erne med perioden 2001-04 og fandt på tværs af afgrøder en fremgang på 45-75 % for flere af de 67 undersøgte arter, men især enårig rapgræs, vindaks og agertidsel gik frem. Alm. kvik, glat vejbred og enårig knavel gik tilbage mellem de to undersøgelsesperioder. Alm. markarve, burre-snerre, liden storkenæb, haremad, nat-limurt og alm. spergel gik tilbage mellem de to første undersøgelser og frem mellem de to sidste.
De dominerende arter, som hver især udgjorde over 10 % af ukrudtsfloraen i mindst én afgrøde, var i alle tre undersøgelser hyrdetaske, hvidmelet gåsefod, alm. kvik, mark-forglemmigej, enårig rapgræs, vejpileurt, snerlepileurt, fersken-pileurt, alm. fuglegræs, mælkebøtte, mark-ærenpris, storkronet ærenpris og ager-stedmoderblomst. I 2001-04 udgjorde vindaks, skive-kamille og lugtfri kamille ligeledes over 10 % af ukrudtsfloraen. Mellem de to sidste undersøgelser steg det totale antal ukrudtsarter fra 108-124 i kornafgrøder (Andreasen et al. 1996 og Andreasen & Stryhn upubliceret).
Andreasen & Stryhn (upubliceret) påpeger, at de tre undersøgelser ikke fortæller noget om udviklingen af sjældne arter. Undersøgelsen blev gennemført i konventionelle, der var usprøjtede i forsøgsperioderne, og hvorledes billedet ville have været, hvis markerne havde været herbicidbehandlet i forsøget, vides derfor ikke. 10 arter udgør typisk mere end 50 % af den samlede ukrudtsmængde i en mark i dag, hvor det før 1980 var 15 arter, der udgjorde halvdelen af biomassen (Rasmussen et al. 2004). Endvidere er ukrudtsbiomassen i danske kornmarker i dag meget lavt og oftest mindre end 1 g tørvægt pr. m² i juni sammenlignet med 1987-1988, hvor niveauet var 5-10 g tørvægt (Sønderskov et al. 2006).
Sammenfattende ser det ud til, at antallet af almindelige ukrudtsarter faldt frem til slutningen af 80érne, mens der i den efterfølgende periode har været fremgang. Der er samtidig sket en generel nedgang i ukrudtsbiomassen og en forskydning i fordelingen mod færre arter der dominerer ukrudtsbiomassen.
En del af den forskydning der er sket i ukrudtsfloraen skyldes ændret sædskifte, herbicidforbrug og næringsstoftilførelse i landbruget. Vinterafgrøder favoriserer efterårsspirende ukrudt, hvorimod forårsafgrøder fremmer andre arter (Marshall, West & Kleijn 2006). Arealet med vinterkorn er steget fra 300.000 ha fra 1982 til 900.000 ha i 2004 (Danmarks statistik 2007) mens arealet med vårkorn er faldet fra 1.500.000 ha til ca. 600.000 ha. Dette kan være en af forklaringerne på den lavere ukrudtsbiomasse, idet vinterkorn i højere grad reducerer ukrudtsbiomassen end vårkorn (Kauppila 1990). Når markjorden tilføres næringsstoffer ændres vilkårene for plantevækst, hvilket resulterer i en ændret artssammensætning. Kornafgrøder og visse ukrudtsarter trives optimalt ved høj næringstilførsel, mens andre ukrudtsarter har specialiseret sig i at overleve ved lavt næringsindhold. Hovedparten af de ukrudtsarter, der findes i dyrkede marker i dag er tilpasset høje kvælstofniveauer (Rasmussen et al. 2004). Kvælstoftilførslen er dog faldet markant siden starten af 1990’erne, hvor tilførslen toppede (Danmarks Statistik 2007). Et lavere herbicidtryk i dag end i midten af 1990érne, hvor forbruget toppede (Danmarks Statistik 2007), er formentlig også en del af forklaringen på den stigende artsdiversitet, der er set de senere år. Således fandt Esbjerg & Petersen (2002), at nedsatte herbiciddoser medførte flere ukrudtsarter, højere tæthed og øget blomstring.
Sprøjtefri zoner vil alene og i kombination med fritagelse for gødning medføre en forøgelse af ukrudtsindholdet i zonerne og dermed i sig selv øge biodiversiteten, men også øge levevilkårene for dyrelivet (se kapitel 5). F.eks. fandt Powell et al. (1985) et højere antal nyttige rovlevende insekter i herbicidfri marker. Moreby & Sotherton (1997) viste at større ukrudtsdiversitet øgede forekomsten af vigtig fugleføde og Navntoft et al. (2003) at øget blomstring øger antallet af sommerfugle.
Landbrugsdriften kan også påvirke plantelivet på de ikke-dyrkede habitater. Der findes flere plantearter i levende hegn på økologiske bedrifter sammenholdt med konventionelle hegn og artssammensætningen i økologiske hegn har større ligheder med halv-kultur habitater end med andre plantesamfund. (Aude et al. 2004; Aude, Tybirk & Bruus Pedersen 2003; Petersen et al. 2006). De mener det formentlig skyldes fravær af pesticidafdrift og lavere næringsstofindhold i de økologiske hegn.
4.3 Nyttedyr
4.3.1 Naturlige fjender
Naturlige fjender kan overordnet opdeles i specialister og generalister. I virkeligheden vil grænsen naturligvis aldrig være helt skarp. Specialister angriber kun en enkelt eller nogle få nærtbeslægtede arter. Mange snyltehvepse er specialister for eksempel bladlussnyltehvepse i Aphidius slægten. Blandt rovinsekterne skelner man ofte mellem de som primært angriber bladlus og generalister, som angriber mange forskellige arter af byttedyr. Til bladlusspecialisterne hører de fleste guldøjelarver, mariehøns og svirrefluelarver. Af generalister kan nævnes edderkopper, løbebiller og rovbiller.
Habitatkvalitet herunder fødetilgængelighed er en vigtig parameter for at øge reproduktionen af naturlige fjender, hvilket kan medvirke til at forbedre deres forekomst og indvirkning mod skadedyr. Interessen er stigende for funktionel biologisk bekæmpelse, som søger at øge antallet og aktiviteten af eksisterende naturlige fjender, ofte generalister, og derved begrænse populationer af skadedyr. Dette sker ved at pleje habitaten og gøre den mere divers (Symondson et al. 2002).
For at forøge nyttedyrenes forekomst, diversitet og kvalitet gennem at forbedre reproduktionen er det nødvendigt at skabe forhold, som giver et bredt udbud af fødemuligheder. Fødeudbud fører til øget frugtbarhed- altså numerisk respons hos løbebiller (Wallin et al. 1992). Numerisk respons er sammen med funktionel respons, -at nyttedyrene søger hen hvor skadedyrene er-, de to mekanismer der bestemmer tætheden af prædatorer. Også specialister har brug for andre fødekilder end det primære bytte, bl.a. pollen- og nektarressourcer (Altieri og Whitcomb 1979).
Et habitats værdi for naturlige fjender er ikke nødvendigvis kun bestemt af byttetætheden. Fysiske forhold som mikroklima, gemmesteder og overvintringssteder er også af stor betydning. En strategi, som skaber varierende habitater indenfor og imellem marker er derfor nødvendig for at maksimere reproduktionen af naturlige fjender og øge den naturlige regulering (Sunderland et al. 1994).
I forsøg, hvor både diversitet af nyttedyr, skadedyr og planter indgår, ser man oftest positive effekter af øget nyttedyrs diversitet (Aquilino et al., 2005; Snyder et al. 2006).
Afgørende er nyttedyrenes mobilitet. Overordnet kan nyttedyrene opdeles i to væsentlige hovedkategorier a) arter der spreder sig ved at gå, b) arter der spreder sig ved at flyve (aktivt eller med vinden, som f.eks. edderkopper med flyvetråde), og dermed spredes væsentligt hurtigere og lettere (Coombes og Sotherton, 1986). En mere detaljeret opdeling er foreslået af Wratten og Thomas (1990), som opererer med a) kolonisering af afgrøder i foråret fra markrand og bevægelser tilbage til markrand i efteråret, b) bevægelser mellem afgrøder indenfor dyrkningssæsonen, bestemt af at afgrøderne er på forskellige udviklinstrin, c) kolonisering af nye habitater efterhånden som det dyrkede areal udvides eller indskrænkes, d) rekolonisering af arealer efterhånden som virkningen af bredspekterede pesticider aftager, e) aggregerende og reproduktionsbundne bevægelser som reaktion på lokalt høje byttedyrsforekomster og f) stor-skala migrationer ind i dyrkede arealer.
Lineære strukturer kan påvirke nyttedyrenes mobilitet. For gående arter kan et vandløb for eksempel være en effektiv barriere, mens hegn typisk vil have varierende grader af gennemtrængelighed. Ordet permeabilitet dækker på engelsk over den andel af individer af en given art som ved møde med en barriere krydser denne (New 2005). Permeabiliteten kan påvirkes af mikroklimatiske faktorer herunder læ-effekten af hegn/markrand, af byttetætheden, typisk relativt høj byttetæthed i hegn/markrand, fysiske strukturer i hegn/ markrand f.eks tilstedeværelsen af en jordvold som i insektvolde.
Afhængig af markrandens permeabilitet kan man skelne mellem en ”hård” rand, som få eller ingen individer af en given art trænger igennem, og en ”blød” rand, samt mellemformer (Duelli et al. 1990). Hegn fremhæves ofte som korridorer, men der mangler dokumentation. Ligeledes savnes grundige eksperimentelle studier på artsniveau af, hvad der bestemmer, om og hvordan enkeltarter vælger, om de vil krydse en markrand/hegn eller ej (New, 2005).
For visse arter kan noget som er en barriere for et livsstadie være let at passere for et andet. For eksempel spreder voksne jagtedderkopper og store unger sig ved at gå, mens yngre individer kan sprede sig med flyvetråde. New (2005) fremhæver, at de populationsbiologiske konsekvenser af forskellige spredningsevne hos forskellige livsstadier ikke er afklaret.
Fragmentering, dvs opsplitning af habitater i mindre og adskilte dele fører typisk til færre arter, mindre poulationer og mindre genetisk udveksling mellem opsplittede populationer. Effekten påvirkes af areal, rand-effekter, fragmentets form, graden af isolation og landskabets konnektivitet (New 2005). Hegn og markrande kan afhjælpe fragmentering ved at fungere som forbindelse mellem adskilte habitater. Særlig vigtigt i denne sammenhæng er ’nodier’, det vil sige de punkter hvor markrande mødes, som kan opfattes som en slags vejkryds (Joyce 1999), men der mangler analyser af betydningen af blandt andet markrandes bredde. Man kan gruppere invertebrater efter hvordan de vil påvirkes af markranden som a) indre arter som er knyttet til randens kerne og hvis forkomst derfor afhænger af dens bredde, b) randarter hvis forekomst og tilstedeværelse ofte er nært knyttet til randen, c) andre arter som mest er knyttet til marken, og ikke påvirkes væsentligt af randen (Laurance & Yensen 1991).
Specialister
Specialister forekommer typisk når et skadedyrsangreb allerede er begyndt. En væsentlig del af specialiseringen er typisk varmekrav der er højere end for byttedyr/vært. I biologiske forstand er det logisk. Fremkomst væsentligt før værtsdyret/ byttedyret kan betyde sultedøden for en specialist.
Svirrefluer (Diptera: Syrphidae) er væsentlige bladlusfjender. Som voksne ernærer de sig af pollen og nektar. Tidligere studier viser betydningen af blomstrende planter for deres bidrag til bladlusbekæmpelse i hvede (Cowgill, Wratten & Sotherton 1993; Klinger 1987). Det kan være vanskeligt at afpasse blomstringsperioden, så den passer med perioden, hvor nyttedyrene ønskes i marken, ligeledes kan forsøg i lille skala give usikre resultater pga svirrefluers høje mobilitet (Pascual-Villalobos et al. 2006). Under danske forhold er svirrefluer ofte af mindre betydning, da de optræder for sent i forhold til bladlusangreb.
Mariehøns (Coleoptera: Coccinellidae) er væsentlige bladlusfjender. Både voksne og larver er bladlusprædatorer. Deres betydning under danske forhold er variererende. Mariehøns overvintrer som voksne bl.a. i hegn. I Danmark optræder mariehøns ofte ‘for sent’ i forhold til skadedyr, men i Sydeuropa kan de være væsentlige nyttedyr. En af de mest almindelige arter er den syvplettede mariehøne Coccinella septempunctata L. Dens stærke varmekrav med et udviklingsnulpunkt på 12.5 °C betyder, at der kan være forsomre hvor dens populationsvækst ”står stille”, mens nogle af de vigtige bladlusarter, som har udviklingsnulpunkter mellem 4 og 6 °C, når at opformere sig betydeligt under de samme forhold.
Snyltehvepse (Hymenoptera). De fleste dyrearter har mindst én parasit. Blandt markens leddyr er snyltehvepse en af de mest udbredte fjender. For de specialiserede snyltehvepse gælder, at de optræder med nogen forsinkelse i forhold til deres værtsdyr. Udviklingsnulpunktet for tre af de mest almindelige bladlussnyltehvepse Aphidius ervi, A. rhopalosiphi og Praon volucre ligger lidt højere end for bladlus hhv. til mumie 2,2 °C, 4,5 °C og 3,8°C og til voksen 6,6 °C, 7,2°C og 5,5°C (Sigsgaard 2000). Hvepsene optræder dog ikke i marken fra det tidspunkt man ud fra udviklingsnulpunkterne kan beregne sig frem til (Sigsgaard 2002). Det skyldes, at hvepsene først er aktive i markranden især hegnene (Langer 2001), hvor den første generation vil opformere sig på bladlus, der lever i hegnene.
Fælles for snyltehvepse er, at larvestadiet snylter på en vært, mens den voksne primært lever af nektar. Dog findes arter af snyltehvepse bl.a. Aphelinus sp. hvor hunnen kan prikke hul i værten med læggebrodden og i stedet for at lægge æg kan vælge at drikke af såret -’host feeding’ (Heimpel, Rosenheim & Kattari 1997).
For at opnå god parasitering er adgang til nektar for de voksne snyltehvepse af stor betydning. Et hollandsk studie kunne vise bedre ernærinstilstand hos Cotesia snyltehvepse i kålmarker med blomsterbræmmer i markranden end i kålmarker uden (Wackers & Steppuhn 2003). Dog kan nogle arter erstatte nektar helt eller delvist med bladlusenes honningdug.
Generalister
De hyppigste generalister i landbrugsmarker på danske breddegrader er edderkopper (Araneae), løbebiller (Carabidae) og rovbiller (Staphylinidae).
Generalisterne fungerer som naturlige fjender mod skadedyr (Sunderland 1975). De er specielt vigtige tidligt på sæsonen hvor specialist-nyttedyr endnu ikke er til stede, og hvor de bl.a. kan reducere populationsopbygninger af havrebladlus (Rhopalosipum padi L.) i vårafgrøder (Ekbom, Wiktelius & Chiverton 1992). En étårig undersøgelse har bl.a. vist, at tilstedeværelsen af generalister reducerede det udbyttetab der kunne tilskrives høj tæthed af havrebladlus med 50 % (303 kg/ha) (Ùstman, Ekbom & Bengtsson 2003).
Samtidig udgør generalisterne en vigtig fødekomponent for andre dyr i agerlandet, bl.a. fugle (Potts 1986; Sotherton & Moreby 2006; Wilson et al. 1999).
Edderkopper kan bidrage væsentligt til bekæmpelse af skadedyr. Der er flest jagtedderkopper i og nær markranden, mens det omvendte gør sig gældende for tæppespindere (Sunderland 1987, Malfait og de Keer 1990, Nyffeler og Breene 1992, Huusela-Veistola 1998). Marker er domineret af nogle få ’agrobionte’ edderkoppearter (Luczak 1979). I Ungarn tilhører 75 % af alle edderkopper i marken fem familier, hvoraf to Pardosa agrestis og Oedothorax apicatus udgjorde næsten 60 %, og var nært knyttet til kornmarker (Samu & Szinetár 2002). De mest almindelige arter i Europa (England, Schweiz) er Batyphantes gracilis (Blackwell), Leptyphantes tenuis (Blackwell), Erigone atra (Blackwell), Milleriana inerrans (O.P. Cambridge), Oedothorax sp., Meioneta rurestris (C.L. Koch) og Erigone dentipalpis (Wider) (Sunderland, Fraser & Dixon 1986).
Hegn og markrande har højere artsdiversitet af edderkopper end omgivende marker og højere samlet antal af edderkopper (Kajak & Lukasiewicz, 1994). Edderkopper kan overvintre i hegn og markrande, og der er mulighed for at fremme disse habitater for edderkopper med bl.a. tuedannende græsser (Bayram og Luff 1993), vilde blomster (Harwood et al., 1994, Thomas og Marshall 1999) og ved nedsat dyrkningsintensitet (Feber et al 1995). Der mangler en afklaring af hegns bidrag til edderkoppepopulationen i marken.
I juni udgør bladlus 10 % af tæppespinderes diæt. Resten af året er 99 % af deres føde springhaler. Men i en vinterhvedemark fjernede tæppespindere 31 kornbladlus pr m² pr dag svarende til en reduktion af bladlustrykket på 37 % (Sunderland, Fraser & Dixon 1986). Eller med andre ord: tæppespindernes overlevelse afhænger af nedbryderkæden, men de giver alligevel et væsentligt bidrag til bekæmpelse af skadedyr. De Barro (1992) viste, at der var 18 gange færre edderkopper (fra 3,6 ned til 0,2 pr cm²) i bure i græsmarker uden adgang for edderkopper, og angreb af havrebladlus havrebladlus var 16 gange større i burene.
Edderkopper er bedst studeret i naturlige habitater men faktorer, der kan påvirke forekomsten af edderkopper, er mikrohabitatets fysiske struktur, mikroklima, adgang til føde, forstyrrelse, konkurrence, biotiske samspil såsom steder at fæstne spind, og prædation på edderkopper. I et review af 24 studier gav 15 øget forekomst af edderkopper ved øget diversitet. Metoderne var reduceret jordbehandling, at lade noget ukrudt stå samt jorddække (Sunderland & Samu 2001). Øget jordzone heterogenitet (små fordybninger i jorden gav tæppespindere mulighed for at placere spind) gav 13 gange flere edderkopper (Alderweireldt 1994) og jorddække i i grønsager 7-14 gange flere edderkopper (Riechert & Bishop 1990).
Edderkopper kan sprede sig ved at gå og ved hjælp af flyvetråde. Snarere end at gå ind i markerne vil edderkopper, der har overvintret i hegn og markrande sprede sig ved flyvetråde (Greenstone et al, 1987; Sunderland 1991). Flyvende edderkopper spredes passivt. De fleste spredes formentlig over ret korte afstande (Halley et al 1996), men i passende vejr kan edderkopper godt flyve flere gange på en dag (Riechert og Gillespie 1986). På denne baggrund må det forventes at afgrøder tættere på reservoirs vil have flere edderkopper (Sunderland og Samu, 2001), en hypotese der understøttes af bl.a arbejde af Thomas (1996) og Topping (1997 og 1999). Toft (1995) analyserede fangster af edderkopper fra dels en 12.2 m høj sugefælde (Rothamsted fælde opereret på Højbakkegaard af H. Philipsen fra 1971-77) og indsamling på et 1m højt trådhegn. Hvor trådhegnet viste et forårs- og et efterårsmaksimum, viste sugefælden blot et maksimum mod slutningen af sommeren. Resultaterne kan fortolkes således, at mens edderkopperne spredes nær jorden i det kølige forår og efterår, tillader den varme sommer spredning i større højde og dermed i større afstande (Toft 1995).
En væsentlig mortalitetsrisiko for flyvende edderkopper er at lande i et ikke- habitat. Græsslåning og høst er andre væsentlige mortalitetsfaktorer. Græsslåning reducerede antallet af tæppespindere med 56-89 %. Fåregræsning tillod kun små populationer af tæppespindere. Pløjning og harvning nedsatte antallet med 89 % (Thomas og Jepson 1997).
Løbebiller er en artsrig familie som er meget almindelig på dyrkede habitater (reviewed af Kromp 1999). De er oftest polyphage og jager en række skadedyr (Sunderland 1975). Løbebiller inddeles i forårsarter og efterårsarter (Larsson 1939). Forårsarterne overvintrer som voksne - ofte i græskanterne langs markerne. Om foråret løber de ind i markerne, hvor de formerer sig. Efterårsarterne parrer sig sidst på sommeren og om efteråret. De overvintrer hovedsageligt som larver i jorden og de voksne biller kommer frem i løbet af sommeren. Forårsarterne (f.eks. spraglet kvikløber og hedeglansløber) er de vigtigste bladlusprædatorer, fordi de er aktive så tidligt på sæsonen, at de kan fouragere på de først ankomne bladlus og derved forsinke og reducere bladlusenes populationsopbygning (Edwards, Sunderland & George 1979; Ekbom, Wiktelius & Chiverton 1992). Det er også vigtigt at tætheden af prædatorer forbliver høj senere på sæsonen, hvor de kan have indflydelse på skadedyr som kornbladlus (Sitobion avenae (F.) (Holland & Thomas 1997). En typisk efterårsart er markjordløber (Wallin 1989) der ofte dominerer i marken om sommeren (Navntoft et al. 2006). Ud over at fungerer som et nyttedyr, kan markjordløbere dog også ved prædation reducere forekomsten af andre vigtige nyttedyr (Prasad & Snyder 2006).
Rovbiller er en af de største billefamilier og mange er polyphage prædatorer. Rovbiller er hovedsageligt aktive om dagen, men de foretrækker skyggefulde mikro-biotoper (reviewed af Bohach 1999). Blandt rovbillerne er agerrovbiller vigtige i landbrugsmarker, fordi de er aktive og almindelige bl.a. i bladlusenes etableringsfase (Petersen 1997).
4.3.2 Bestøvere
Humlebier, vilde bier og honningbier er de vigtigste bestøvere i landbruget. Andre bestøvere er sommerfugle og bl.a. fluer og glimmerbøsser. Mængden af honningbier afhænger af nærhed til bigårde eller placering af bistader i marken i frøafgrøder og visse andre afgrøder som frugt og bær. For humlebier og vilde bier er adgang til egnet redebygningssted og til blomstrende planter at trække på i hele vækstsæsonen afgørende. Feltobservation i marken af bi-besøg viste af flerårige planter samlet fik flest besøg (Fussell & Corbet 1991). Langtungede humlebier (agerhumle og havehumle) foretrak læbeblomstrede planter. Korttungede humlebier (jordhumle/lille jordhumle, markhumle, stenhumle) og honningbier besøgte brombær, horse-tidsel, lådden dueurt, alm. bjørneklo og raps (Fussell & Corbet 1991). Humlebier er gået væsentligt tilbage i det moderne landbrug. Såede markrande med en blomster-græs blanding, som sikrede en succession af blomster, fremmede humlebier allerede fra første år mens brakstriber kun gav flere humlebier to år efter etablering pga. uønsket fremvækst af tidsler (Carvell et al. 2004). Værdien af såede markrande for bier og humlebier er bekræftet i andre undersøgelser (Meek et al. 2002).
Tilstrækkelig bestøvning og egnede bestøvere kan være afgørende for sjældne plantearters overlevelse. Fragmentering fører til nedsat bestøvning fra humlebier og reducerer udveksling af pollen mellem de isolerede habitater (Goverde et al. 2002). Bestøvning af kulturplanter har stor økonomisk betydning. I dokumenterede tilfælde med mangelfuld bestøvning af afgrøder er forarmning af landbrugslandet den hyppigste årsag (Richards 2001).
4.3.3 Funktionel biodiversitet
Funktionel biodiversitet er den del af biodiversiteten som har en funktion i forhold til nyttedyrenes regulering af skadegørere, bestøvning af kulturplanter m.m. Biologisk bekæmpelse kan gennemføres ved målrettet at søge at fremme nytteorganismernes levevilkår i marken og dens omgivelser. For eksempel ved at etablere plantesamfund i og omkring marken som på en eller flere parametre bidrager til overlevelse og opformering af nytteorganismer. Det har vi valgt at kalde funktionel biologisk bekæmpelse (på engelsk conservation biological control) (se kapitel 2).
Effekten af funktionel biologisk bekæmpelse på grupper af nyttedyr vil afhænge af flere forskellige karaktertræk ved nyttedyrene. Væsentligt er, om nyttedyrene er afhængige af en enkelt eller få arter af byttedyr, eller om de er mere alsidige. Et andet væsentligt træk er nyttedyrenes mobilitet, f.eks. hvor langt de kan bevæge sig i deres søgen efter føde. Der findes kun få undersøgelser, der viser en udbyttemæssig gevinst af funktionel biologisk bekæmpelse, og en del undersøgelser som ikke har kunnet påvise effekter. På bedriftsniveau er den økonomisk værdi af bladlusprædatorer undersøgt på ti svenske fuldtidslandbrug i forhold til deres evne til at reducere tæthed af havrebladlus, og øge udbyttet i byg. Studiet blev gennemført i et år med stort bladlusangreb. Tilstedeværelsen af generalister reducerede det udbyttetab der kunne tilskrives høj tæthed af havrebladlus med 50 % (303 kg/ha), en udbyttegevisnt som er sammenlignelig med den fra pesticider. Det øgede udbytte var højest på konventionelle landbrug målt i absolutte tal, men den procentvise stigning var størst på økologiske brug (Ùstman, Ekbom & Bengtsson 2003). Cardinale et al. (2003) giver et andet eksempel på at øget biodiversitet medfører udbyttegevinster. De viste at en øget artsdiversitet af ærtebladlusens naturlige fjender og et øget udbud af andre byttedyr til disse naturlige fjender tilsammen øgede produktionen af lucerne.
Plantediversitet kan også give øget produktivitet. Øget plantediversitet i såede græsmarker til slæt viste sig at øge græsudbyttet med 60 % i en engelsk undersøgelse (Bullock et al. 2001). Tilsvarende er påvist i et større europæisk studie (Spehn et al. 2005). Det er uklart hvor stor en andel der kan tilskrives forbedret funtionel biologisk bekæmpelse.
4.4 Sommerfugle
Tidligere har man især beskæftiget sig med sjældnere sommerfugles tilbagegang i forbindelse med særlige habitater som f.eks. hede. Nyere forskning har imidlertid påvist, at almindelige sommerfugle fra det åbne land også har haft betragtelig tilbagegang som følge af ændret dyrkningspraksis, især tab af hegn og urtevegetation i hegnenes fodposer. Sidstnævnte skyldes både, at der bliver pløjet tættere ind under hegnene og påvirkning af hegnenes urtevegetation med pesticider og gødning. Gødning har ført til en stærkere dominans af enårigt ukrudt (Boatman 1992). Sommerfugle er nær knyttet til markrande i dyrkede marker. De er afhængige af adgang til nektar fra blomster i markranden. Mange arter af sommerfulge ses flyve langs hegn. Et hegn over 2 m passeres sjældent af sommerfugle.
I randzoner omkring vinterhvedemarkerne steg antallet af arter af de seks fremherskende sommerfuglearter: Græsrandøje, vejrandøje, okkergul randøje, lille kålsommerfugl, grønåret kålsommerfugl og stregbredpande signifikant (svarende til en faktor 2.3) og individantallet (en faktor 4,6-4,9) med usprøjtede randzoner på 3-6 meter (de Snoo, van der Poll & Bertels 1998). Effekten kunne også ses i afgrøderand og i kanten af vandløbene (digebredderne). Der var ikke samme tydelige effekt af usprøjtede randzoner omkring kartoffelmarker. Kun I 1992 steg artsantallet og individantallet statistisk signifikant (hhv. en faktor 3.6 og 6.9), og det var også kun dette år, der sås effekter i afgrøderand og markkant (de Snoo, van der Poll & Bertels 1998).
En dansk undersøgelse (Holbeck, Clausen & Reddersen 2000) af sammenhængen mellem sommerfugle og lineære vs ikke-lineære habitater (dvs udyrkede lidt større områder) viste den store betydning af nektarkilder for alle arter af sommerfugle. Især blåhat og i nogen grad rød-kløver i lineære habitater og i ikke-lineære især gule kurveblomster sammen med tidsler. Undersøgelsen viste at ikke blot lineære habitater var væsentlige for sommerfugle i det åbne land, men at de ikke-lineære habitater også havde væsentlig betydning. Inddrager man ikke de ikke-lineære habitater risikerer man at undervurdere både antal og artsrigdom (Feber & Hopkins 1997; Feber, Smith & MacDonald 1994; Feber, Smith & MacDonald 1999; Hopkins & Feber 1997; Snoo, Poll & Bertels 1998; Sparks et al. 2000).
4.5 Fugle
Fugle bruges ofte som bioindikatorer i agerlandet bl.a. fordi fuglene påvirkes af intensiteten af landbrugsdyrkningen (Ormerod & Watkinson 2000). Mellem 1970 og 1990 gik udbredelsen af agerlandets fugle i Storbritannien tilbage for 86 % af alle arter og 83 % af alle arter faldt i antal (n = 18), hvoraf syv arter havde populationsnedgange på mere end 50 % (Fuller et al. 1995).
Forklaringer på det faldende antal fugle i agerlandet i Storbritannien er nedgangen i mængden af vilde fødeplanter (Firbank & Smart 2002) og nedgangen i mængden af insektføde (Benton et al. 2002), men også mangel på egnede levesteder har betydning. Agerlandets fugle kan overordnet inddeles i to grupper: Flest arter er tilknyttet vegetation i haver, levende hegn og/eller andre småbiotoper (f.eks. gulspurv, tornsanger, solsort og bogfinke), og så er der de (færre) arter tilknyttet det åbne land (f.eks. sanglærke, vibe, agerhøne og bomlærke (Petersen 1998). Som nævnt et det dog ikke alle fuglearter i agerlandet, der er i tilbagegang. De hegnstilknyttede fugle i Danmark er generelt ikke længere i tilbagegang på nær enkelte arter som gulspurven, som søger føde ude i marken, hvorimod bestandsudviklingen er anderledes negativ for fugle tilknyttet markfladen som sanglærker og viber (Fox 2004; Heldbjerg 2005; Petersen 1998).
Gulspurven er en almindelig fugl tilknyttet højere vegetation i småbiotoper og dermed agerlandets randzoner. Den er stadig talrig i Europa, men begyndte bl.a. at gå tilbage i Storbritannien i slutningen af 1980érne (Bradbury et al. 2000). De senere år er bestanden gået tilbage med ca 10 % årligt og lå i 2000 på det laveste niveau i Storbritannien siden målingerne startede i 1961 (Bradbury et al. 2000). Tilbagegangen skyldes primært intensiveringen af landbruget (Bradbury et al. 2000). Danmark har oplevet en gradvis nedgang fra sidste halvdel af 80'erne med omkring en tredjedel og der er ingen tegn på at udviklingen er vendt (Heldbjerg 2005).
Sanglærken er et typisk eksempel på en art, som yngler på åben mark, og er således ikke direkte tilknyttet randzoner. Den er ligeledes i tilbagegang, og i perioden 1976-1985 gik bestanden af sanglærke tilbage med omkring 50 %, hvilket tilskrives intensiveret landbrugsproduktion og en stigning i pesticidforbruget. Fra midten af 1980erne er bestanden til gengæld gået lidt men jævnt frem til et niveau på ca. 70 % af niveauet i 1976. Denne positive tendens i sanglærkens bestandsudvikling skyldes primært de mange milde vintre. Sidst i 1990erne er bestanden begyndt at falde igen. Arten er i tilbagegang i hovedparten af Europa med størst tilbagegang i de vesteuropæiske lande (Heldbjerg 2005).
4.6 Betydning af et diverst landskab
De enkelte habitaters størrelse har afgørende betydning for såvel flora og fauna. For floraen har markrande og små habitatfragmenter nogen betydning for bevarelse af biodiversiteten, men sammenlignet med næringsfattige marker, med væsentligt større areal er diversiteten lavere i markrande og småhabitater (Smart et al. 2002; Smart et al. 2006).
4.6.1 Landskabsdiversitet (heterogenitet) i stor skala
Landbrugets effekter på plante- og dyresamfund vil optræde på forskellige skala fra mark til landskab afhængig af organismernes økologiske træk. Vigtige træk, der bestemmer dyrs og planters respons på forskellige grader af landskabsheterogenitet, er deres mobilitet og livscyklus (Burel & Baudry 2005).
En større grad af landskabsheterogenitet er knyttet sammen med en højere biodiversitet i agerlandet, både når man måler i lille og stor skala (Benton, Vickery & Wilson 2003). Heterogenitet er nødvendig for at sikre tilgængelige ressourcer året rundt for artsrige samfund (Benton et al. 2003).
Det betyder, at skala kan være afgørende for, om a) et givent tiltag vil påvirke en given gruppe organismer, og/eller b) om effekten er målelig. For eksempel kan effekten af pesticidanvendelse ikke måles på tæppespindere på mark- eller bedriftsskala, på grund af denne gruppes store evne til spredning, som vil jævne forskelle ud på lille skala, mens effekten er tydelig på landskabsskala, hvor pesticidbehandlinger kan nedsætte tæppespinderantallet med 50-90 % i grupper af marker som fik højt pesticid input (Vickerman 1992, Burn 1992). Halley et al (1996) udviklede en model for at kunne forudsige effekten af pesticider på tæppespindere over større skala end enkeltmark. Modellen opererer med uge og årscykler og landskabet som et bånd af firkantede marker, hvis størrelse er defineret af marksidens længde i meter. Der regnes alene med vindspredning og alene i én retning og kun en celle (mark) eller given radius ad gangen. I England er tre årlige sprøjtninger maksimum. I værste fald dræbes 90 % tæppespindere ved en sprøjtning. Mere end én sådan sprøjtning kan stort set udrydde tæppespindere. I modellen opnås ved 25 % græsmarker eller usprøjtede kornmarker ca. den tæthed som vi i dag finder i sprøjtede kornmarker. Risiko for at lande i et ikke-habitat med tilhørende høj mortalitetsrisiko gør det optimalt, at ca. 50 % af en population af tæppespindere spreder sig ved flyvetråde.
Diversiteten af edderkopper er påvirket af to rumlige skalaer (kant vs. midte af mark og simpelt vs. komplekst landskab), men ikke på to andre skalaer (dyrkningsmåde, region), hvilket understreger behovet for at undersøge forskellige rumlige skalaer (Clough et al. 2005). Desuden understreger studiet at heterogenitet på landskabsniveau er en af nøglerne til at fremme edderkopperdiversitet i agroøkosystemer.
Landskabsheterogenitets indflydelse på bestøvere er også påvist. For solitære bier kunne der påvises positive effekter af mere halvnaturligt habitat (som randstriber) landskabsheterogenitet på en skala op til 750 m, men for humlebier og honningbier var der ingen effekt på denne lille skala. For honningbier sås endda en positiv effekt af mindre andel af halvnaturligt habitat helt op til 3000 m (Steffan-Dewenter et al. 2002).
Arter interagerer og påvirkes samtidig på forskellig rumlig skala. Eksisterende data tyder på, at organismer med høj spredningskapacitet driver disse biodiversitetsmønstre og økosystemtjenester på grund af a) deres evne til rekolonisering b) deres adgang til flere ressourcer. Frem for at fokusere på få arter og lokale processer er der behov for at fokusere på diversitet af de nøglearter, der kan sikre resiliens (Tscharntke et al. 2005).
I de senere år er forskningen i det dyrkede land i relation til landskab for alvor sat i gang. Landskabseffekter på en større gruppe organismer blev gennemført på 14 lokaliteter i Bretagne, som adskilte sig med hensyn til jordbund (en gradient fra finsandet til grovsandet) og dyrkningsintensitet og andel af flerårige habitater som skov. Responsen ved et mere monotont landskab var enten et fald i antal arter (Tovingede: Chironomidae, Empididae), udskiftning af arter (replacement) (løbebiller) eller ingen forandring (små pattedyr). Årsagen til den forskellige respons var at finde i dyrenes livscyklus og deres mobilitet. Den afhang desuden af hvilke parametre i landskabsstrukturen, de var følsomme overfor (Burel & Baudry 2005). For tovingernes vedkommende bestemte tætheden af hegn og de enkelte hegns gennemtrængelighed den rumlige udbredelse af voksne tovinger afhængigt af disses flyveevner. Den gennemsnitlige løbebillestørrelse faldt med stigende dyrkningsintensitet, da de små arter er bedre tilpasset til forstyrrelser frem for de store arter, der hører til i et stabilt habitat. De samme arter af småpattedyr optrådte over begge gradienter, men artssammensætningen hang sammen med andelen af afgrøder i forhold til mere stabile landskabselementer (Burel & Baudry 2005).
Forholdet mellem økologisk produktivitet og artdiversitet ændres med ændret rumlig skala (Chase & Leibold 2002). For eksempel fandt man, at artsdiversiteten i en sø med gennemsnitlig algeproduktivitet var størst. Men på regional skala var det søer med høj algeproduktivitet, der samlet set gav højest artsdiversitet. Selv om hver enkelt sø havde lav artsdiversitet, var der nemlig kun lille overlap mellem diversiteten i de enkelte søer (Samways 2005).
Bevarelsen af et heterogent landskab med mange forskellige habitater er afgørende for en række arter som for eksempel humlebier (Goverde et al. 2002; Kells & Goulson 2003) og guldsmede (Steytler & Samways 1995).
På svenske landbrug var artsrigdommen i en undersøgelse af planter, rovbiller, løbebiller, sommerfugle og edderkopper størst på brug med et heterogent landskab, så som en blanding af dyrkede marker, enge og skov (Weibull, Ùstman & Granqvist 2003). Herved kommer det enkelte lille smålevested –eller patch til at spille en væsentlig rolle for at bevare en høj biodiversitet –og kunne opnå en høj funktionel biodiversitet.
Det er påvist at både økologisk dyrkning og landskabsheterogenitet fremmer sommerfugle. Ved høj landskabsheterogenitet er effekten af økologisk drift ikke signifikant (Rundlùf & Smith 2006). Med andre ord opnår man mest miljøgevinst ved indsatser i i forvejen fattige landskaber.
Et omfattende transekt studie gennem landbrugs- og naturområder i Schweiz bestående af vådområder, landbrug, græs, eng og skov konkluderede, at tilbageblevne rester af naturlige habitater eller halvnaturlige habitater var de væsentligste populationskilder i landbrugsmiljøprogrammer til fremme af biodiversitet i et ellers forarmet landskab. Således kunne de påvise, at 63 % af alle dyrearter i området (Limpach-dalen) var afhængige af tilstedeværelsen af halvnaturlige habitater, heraf afhang 12 % af økotoner, dvs. hegn og lignende lineære habitater (Duelli & Obrist 2003).
Insekter som bevæger sig fra marker til tilstødende naturlige habitater udgør en vigtig fødekilde for dyrelivet - specielt fugle (Rand, Tylianakis & Tscharntke 2006a). Hvis man forudsætter en øget vandring fra bufferzoner som f.eks. sprøjtefri randzoner til tilstødende naturområder kan dette have en positiv effekt på dyrelivet her (Rand, Tylianakis & Tscharntke 2006a). Omvendt kan markens nyttedyr også konkurrere med den naturlige habitats fauna, når de sidst på sommeren forlader marken og søger ud mod markrand og omgivende habitater kaldet ’spill-over effect’ (Rand, Tylianakis & Tscharntke 2006; Tscharntke, Rand & Bianchi 2005).
4.6.2 Landskabsheterogenitet over tid
I en plantesuccession fra pionerlandskab til marker i tidlig succession og i midt succession var der ikke forskel i sommerfugles artsdiversitet, men artssammensætningen ændrede sig grundlæggende mod mindre kropsstørrelse, mindre evne til migration, færre arter der overvintrede som voksne, færre generationer og længere larvestadie. Overraskende fandt man dog ikke ændring i graden af plantespecialicering (Steffan-Dewenter & Tscharntke 1997). Et landskab med plantesamfund i forskellige stadier af succession vil med andre ord kunne have størst samlet diversitet, forudsat at der er tilstrækkelig mulighed for migration mellem lignende habitater til at undgå lokal udryddelse (Samways 2005).
Konnektivitet er et nøglebegreb i landskabsøkologi, da ordet beskriver bevægelser af organismer styret af landskabsstrukturer. I måling af konnektivitet kan blandt andet indgå afgrødediversitet i det dyrkede land. I Bretagne fandt man, at over en 7-årig undersøgelsesperiode var 70 % af landskabet, som blev anvendt til majsdyrkning forbundet i store clustere, snarere end i adskilte smålevesteder, således at dyr kunne bevæge sig fra smålevested til smålevested fra år til år (Burel & Baudry 2005).
I Australien er op til 95 % af den naturlige vegetation ryddet i visse områder, og her er der forsket i betydningen af naturlige småbiotoper for krybdyr i landbrugslandet (Driscoll 2004;Jellinek, Driscoll & Kirkpatrick 2004). De fandt dog, at artsrigdom og forekomst af firben ikke blev signifikant påvirket af habitatfragmentering eller fragmenternes størrelse men derimod af småbiotopernes heterogenitet. Desuden fandt Driscoll (2004), at formen af småbiotopen havde en stor betydning med færre arter i linære end i kvadratiske biotoper.
4.6.3 Effekter af sammenhænge gennem landskabet
Et landskab kan betragtes som en mosaik af landskabselementer. Ikke-dyrkede habitater har en vigtig rolle i form af spredningskorridorer og øer i et fragmenteret landskab, da de muliggør individers spredning over større områder (Benton et al. 2003). Disse korridorer som f.eks. levende hegn kan i sig selv fungere som habitater, og de kan fungere som filtre ved kun at være korridorer for visse arter. Hvorvidt et landskab har tilstrækkeligt med spredningskorridorer og øer og herved migrationsmuligheder mellem adskilte smålevesteder (patches) kan bestemmes ved at monitere de arter, der har lavest kapacitet for spredning (Nordén & Appelqvist 2001). Smålevestedets kvalitet og størrelse har betydning for artsrigdommen, og de tilknyttede korridorers karakter, størrelse og pasning har betydning for spredningen imellem dem.
Betydningen af smålevesteders størrelse og hvor tæt de ligger er påvist i flere undersøgelser. Det påvist, at stigende habitatisolation medfører mindre bestøvning, lavere frøsætning, indavl og stigende risiko for udryddelse af mange plantearter (Matthies et al. 1995; Fischer & Matthies 1997; Goverde et al. 2002; Steffan-Dewenter & Tscharntke 1999).
Biedermann (2000) har påvist en klar sammenhæng mellem størrelsen af smålevesteder og hyppigheden af lokal udryddelse af visse insektarter (skumcikader) og Zschokke et al. (2000) m.fl. har vist at habitatstørrelse er positivt korreleret med både det samlede antal sommerfugle og artsantallet af sommerfugle, og dermed at antallet af sommerfuglearter falder jo mere isoleret habitaten er placeret.
Vigtigheden af hegn som korridorer for skovlevende løbebiller er påvist i Bretagne (Burel et al. 2004), og Fitzgibbon (1997) viste, at skov-smålevesteder som var forbundet af levende hegn, havde større populationer af både skovmus og rødmus.
Det er muligt at øge landskabsheterogenitet med nye habitater. Nyoprettede halvnaturlige habitater hhv. isoleret og forbundet til skov gennem nyoprettet læhegn opnåede overvintringstætheder som i markens gamle kantområde (150 biller/m²), men færre end i tilsvarende gammel biotop. Kantbidrag fra den nye isolerede biotop af forårsformerende arter var under 1 % (Serholt & Heller 1997).
5 Viden om randzoner og andre pesticidfrie beskyttelsesstriber
- 5.1 Formål med randzoner og andre pesticidfrie beskyttelsesstriber
- 5.2 Markrande
- 5.3 Bufferzone
- 5.4 Sammenfattende viden om markrand og bufferzone
- 5.5 Insektvolde, græsbælter og blomsterbræmmer inde i markerne
- 5.6 Fugle og andre højerestående dyr
- 5.6.1 Sammenfatning om fugle og andre højerestående dyr
Markrande og bufferzoner vil i praksis udgøre et sammenhængende areal i randzonen. Dette afspejles i, at mange undersøgelser behandler markrand og bufferzoner under et. I forhold til den teoretiske tegning i kapitel 2, er det ikke givet, at alle elementer vil indgå i markrand og bufferzoner. For eksempel kan en bredere randstribe, så som en braklagt bufferzone ligge direkte op til den dyrkede mark uden nogen pesticidfri bufferzone imellem.
Markrande og bufferzoner har stor betydning for plante- og dyreliv som permanente eller semipermanente habitater. Der er udført et stort antal undersøgelser af dette. Mest opmærksomhed samler sig om de arter, som er iøjnefaldende, populære eller har betydning for produktionen enten skadelige (ukrudt, skadedyr) eller nyttig (bestøvere og nyttedyr). Rapporten afspejler dette med særlig omfattende afsnit om fugle og om nyttedyr. Markstriber inde i marken er anlagt særligt med tanke på at fremme funktionel biodiversitet, hvilket gør at bredere biodiversitetsdata er sjældnere for markstriber.
5.1 Formål med randzoner og andre pesticidfrie beskyttelsesstriber
Randzoner anlægges med tre mål for øje
a) agronomiske formål bland andet med hensyn til udbytte, rationel drift, hensyn til nytte- og skadedyr samt ukrudtsbekæmpelse,
b) hensyn til biodiversitet og fremme af et alsidigt plante- og dyreliv samt
c) hensyn til landskabets rekreative værdier.
Ved anlæg af randzoner vil valget af plantedække have væsentlig indflydelse på plantesamfundet og dyrelivet i randzonen. Det gælder uanset om det er det naturlige plantedække, som man lader etablere sig selv, eller om det plantes eller sås ud, og senere hvordan det passes.
Mulige ulemper ved alle typer af randzoner er udbyttetab, risiko for opformering af vanskeligt ukrudt samt opformering af skadedyr.
5.2 Markrande
Markranden defineres her som i Greaves og Marshall (Greaves & Marshall 1987; Marshall & Moonen 2002) som bestående af:
a) Markgrænsen. For eksempel markeret af et hegn.
b) Det delvist naturlige habitat der er knyttet til markgrænsen. Ofte hegn og fodpose.
c) En stribe jord mellem markgrænsen og afgrødekanten ’randstribe’ (eng: margin strip), som vil hedde noget forskelligt efter, hvordan avleren har valgt at anvende den. Denne kan evt. være afgrænset mod marken af en barjordsstribe. Randstriben kan for eksempel være forager, græsstribe, brakstribe eller blomsterbræmme.
d) Afgrødekanten regnes som en del af markranden. Det er typisk en smal stribe af marken, som tydeligt adskiller sig i plantetæthed og eller højde.
Bufferzonen er den usprøjtede og evt. ugødede yderste 1-10 m af afgrøden, dvs at afgrødekanten indgår heri (se figur 1.1).
Markranden har en mere divers flora end marken. Til forskel fra marken kan man desuden finde vedagtige planter, buske og træer i markranden. Jordbunden i markranden, bearbejdes mindre intensivt end i marken, og har ofte også lavere gødningsniveau, ligesom pesticideksponering kan være lavere. Mikroklimaet i markranden vil ofte også adskille sig fra markens. Den større habitat- og plantediversitet betyder, at flere dyr og flere dyrearter kan leve i markranden. Hvilke arter og artsantal, der vil findes i markranden afhænger af, hvordan markranden opfylder deres forskellige behov, herunder adgang til føde og skjul. For fugle kan føde for eksempel være insekter eller frø, og skjul kan være egnet redebygningssted eller egnede hvilepladser. Tilsvarende har andre dyregrupper specifikke behov. For eksempel har bestøvere og flere nyttedyr behov for adgang til pollen og nektar. For mange arter af leddyr er overvintringsmuligheder i markranden væsentlige.
Markranden indgår ofte i undersøgelser af sprøjtefrie randzoner, både til belysning af effekter på flora og fauna og effekter på afdrift (Se bilag A, tabel 3). Ud over funktioner beskrevet under hegn (se nedenfor) kan markrandens ønskede/formulerede funktioner i det moderne landbrug omfatte:
- Fremme af økologisk stabilitet i afgrøder, gennem habitater til vilde planter og dyr
- Nedsat brug af insekticider gennem fremme af naturlige fjender af skadedyr
- Fremme af bestøvning
- Begrænsning af ukrudsindtrængen og herbicidforbrug
- Reduceret tab af gødning og pesticider til omgivelserne
- Fremme af biodiversitet og beskyttelse af vilde planter og dyr i agerlandet
- Bevarelse af landskabsdiversitet
- Fremme af vildt
5.2.1 Hegn
Hegn er det mest markante element i markrande og til forskel fra den øvrige mark rummer de vedagtige planter.
Formålene med hegn er blandt andet at:
- Beskytte mod vindens nedbrydende virkning
- Give læ til afgrøden og holde på jordfugtigheden
- Hensyn til jagt
- Høst af brænde og frugt
- Sikre gode levevilkår for dyr og planter
- Markere markens grænse i forhold til omgivelserne (ejerskab)
- Æstetetiske eller kulturelle værdier som bevarelse af historiske landskabstræk, kulturminder og 'stedfornemmelse'
I denne rapport lægges fokus på økologiske og miljømæssige aspekter, og der er derfor ikke inddraget historiske, æstetiske eller kulturelle aspekter.
Potentielle ulemper som skal minimeres omfatter
- Konkurrence med afgrøden om lys, vand eller næring
- Hegn som kilde til ukrudt, sygdomme og skadedyr
I et typisk læhegn kan man skelne mellem vindrækker, som er vestvendte rækker, højderækker i midten af hegnet og karakterrækker, som skal afrunde og stabilisere hegnet i læsiden. Her kan indplantes blomstrende buske og træer. Afhængig af klimatiske forhold kan hegnet bestå af 1-3 vindrækker, 1-3 højderækker og 1-2 karakterrækker, ifølge læplantningsloven dog sammenlagt normalt ikke over syv rækker (Bruus Pedersen, Aude & Tybirk 2004). Under og på begge sider af hegnet findes fodposen.
5.2.1.1 Hegnenes flora
Hegnenes flora er resultatet af plantevalg og –pleje parret med den naturlige floras etablering i hegnet. Hegnenes flora afhænger for de flerårige planters vedkommende af hvad der oprindelig er plantet. Hertil kommer selvsåede flerårige og enårige. Ved foryngelse, beskæring og slåning kan floraen 0løbende manipuleres. Det vil sige, at hegnenes flora er resultatet af en fortløbende proces, som påvirkes af forskellige tiltag, herunder brug af gødning og herbicider.
For en forståelse af effekten af herbicider på hegn er to undersøgelser som sammenligner to grupper af ensaldrende hegn, begge plantet af Hedeselskabet, og placeret dels i økologisk og dels i konventionel drift, særlig interessante, da de belyser effekter over mange år. Begge undersøgelser viste signifikant flere arter i hegn som ikke havde været påvirket af herbicidafdrift (Aude et al. 2004; Aude, Tybirk & Bruus Pedersen 2003). Der var en signifikant større lighed mellem floraen i de økologiske hegns fodposer og naturlig vegetation end mellem den konventionelle urtevegetation i hegnene og naturlig vegetation. De arter, der især var flere af i de økologiske hegn, var ukrudtsarter, arter hørende til halvnaturlige habitater og til åbne landskaber som græsenge og mosser (Aude et al. 2004).
Denne forskel i artssammensætning svarer til den, der er fundet i sammenligninger af markfloraen under økologisk og konventionel drift (Hald 1999).
En høj diversitet opnås ikke nødvendigvis ved at udelade pasning. Et hegn, der blev undersøgt med 27 års mellemrum, havde nedgang i fodposens floradiversitet både i dele, som lå op til dyrkede marker og blev passet årligt og i dele, som ikke blev passet (Garbutt & Sparks 2002). Omvendt fandt Petersen (2006) at diversiteten øgedes allerede få år efter omlægning.
Herbicidet glyphosat kan reducere antallet af plantearter med ca. ¼. En tilsvarende reduktion findes af kvælstofgødskning alene (Bruus Pedersen, Aude & Tybirk 2004). Glyphosat alene havde en tydelig effekt på plantebiomassen ved højt gødskningniveau, men næsten ingen ved 0 kg kvælstof. Ved højt glyphosat-niveau gik leddyrene tilbage i antal og diversitet (med maksimale reduktioner på henholdsvis 47% og 26%). Effekten på leddyr var specielt tydeligt ved højt kvælstofniveau, og afspejlede den store ændring af vegetationen under disse forhold (Bruus Pedersen, Aude & Tybirk 2004).
5.2.1.2 Hegnenes fauna
Hegnenes fauna består af arter, der er afhængige af hegnets særlige plantearter, dets særlige mikroklima eller både/og. Til de mikroklimaafhængige hører fugtelskende arter som springhaler, mejere, bænkebidere og snegle. Hegnene er væsentlige for mange arters overvintring, i bl.a. hegnenes bundlag. I foråret rummer hegnene føde før der er føde at finde i marken. Det gælder tidlige blomster at trække på for bestøvere, frø og insekter til fugle og bytte for rovinsekter og snyltere.
Faunaen kan opdeles efter habitat, med
a. En gruppe som er udelukkende knyttet til hegn/ markrand
b. En gruppe som er primært knyttet til hegn,
c. En gruppe som er primært knyttet til mark og
d. En gruppe som forekommer i såvel hegn/markrand som mark
Ændringer i dyrkningspraksis vil påvirke faunaen forskelligt afhængigt af habitatgruppe.
Især forår og forsommer er tætheden af leddyr størst i hegn. I en undersøgelse fandt man således op til 600 individer/m² i fodpose og en biomasse på 0,9 - 1,6 g/m² mod < 0,2g/m² i bufferzone og mark (Hald et al. 1988).
5.2.2 Randstribe
Randstriben, placeret mellem markgrænsen, et eventuelt hegn eller vandløb og selve den dyrkede mark, har ringere produktionspotentiale end den øvrige mark, fordi den ofte ligger delvis i skygge og ofte belastes af mere kørsel and den øvrige mark. Endvidere kan der være vanskelige hjørner eller våde pletter, som er vanskelige at håndtere med store markredskaber. Desuden kan der forekomme konkurrence med hegnets planter om bl.a. vand. Samtidig besidder randstriben et væsentligt biodiversitetspotentiale for flora og fauna såvel som en mulig ressource for nyttedyr til marken (Marshall, West & Winstone 1994). Muligheden for at anvende et stykke mindre produktivt jord til at fremme biodiversitet har givet anledning til en række undersøgelser.
5.2.2.1 Randstribens flora
Forskellige måder af forvaltning påvirker floraen på forskellig måde. Hald & Lund (1994) undersøgte fire sprøjtefri driftsformers betydning for markers randzoner henholdsvis vårbyg, vårbyg med udlæg af kløver-græsblanding med urter, kløver-græsblanding med urter og egentlig brak. Forsøget omfattede desuden tre niveauer af kvælstoftilførsel (0, 1/2 og 1/1). Tætheden af vilde planter var dobbelt så stor ved braklægning sammenlignet med de øvrige behandlinger efterfulgt af kløver-græsblanding med urter. Disse to behandlinger havde også en høj blomstring og ukrudtsbiomassen var ligeledes markant i disse to behandlinger sammenlignet med ”korn-behandlingerne”. Der var endvidere en tendens til lavere artsantal og mindre forskel mellem behandlingerne ved øget afstand fra hegnet (0-2,5 m fra hegn sammenlignet med 2,5 til 6 m fra hegn). Der var ikke effekt af kvælstofniveauet i relation til biomassen af vilde planter (Hald & Lund 1994).
Især ved etablering af randstriben kan der være risiko for besværligt ukrudt. Problemet kan begrænses meget. Et eksperiment af Asteraki et al. (2004) sammenlignede tre typer udsåede urteblandinger af hjemmehørende arter og naturligt regenererende randstribe. Efter to år havde alle typer udsåede rande færre uønskede ukrudtsarter (konkurrerende med afgrøden) end den naturligt regenererende randstribe. I den mest diverse udsåede randstribe med blandet græs og bredbladede urter blev dette opnået allerede efter et år. Efter to år var den oprindelige frøbank ikke til at spore i de udsåede plots. Udsåning havde størst positiv betydning efter herbicidanvendelse, hvor risiko for uønskede ukrudtsarter i hegn er størst (Asteraki et al. 2004). Marshall (2001) fandt ligeledes, at udsåning af græs eller blomsterblandinger i randstriben langs et hegn effektivt reducerede mængden af uønsket ukrudt, som også påvist af andre (Denys & Tscharntke 2002).
Ikke overraskende konvergerer forskellen mellem udsået og naturligt etablerede randstribers florasammensætning over tid (De-Cauwer et al. 2005).
Slåning fremmer artsdiversiteten af planter (Hovd & Skogen 2005). Desuden kan slåning med efterfølgende fjernelse af plantemateriale bruges til at fjerne næringsstoffer, så mindre N-krævende plantearter kan etablere sig.
5.2.2.2 Randstribens fauna
Hald & Lund’s (1994) undersøgelser af fire sprøjtefri driftsformers betydning for markers randzoner henholdsvis vårbyg, vårbyg med udlæg af kløver-græsblanding med urter, kløver-græsblanding med urter og egentlig brak viste, at insektfaunaen overordnet havde de bedste betingelser i vårbyg med udlæg og i kløver-græsblandingen med urter. Kløver-græsblandingen med urter gav tidligere blomstring til fordel for flere insekter, bl.a. fordi der var iblandet honningurt og boghvede, som tiltrækker mange insekter. De mindre mobile insekter, som mange er vigtige fuglefødeemner (f.eks. sommerfuglelarver, løbebiller og rovbiller), var talrigest, hvor der var udsået vårbyg med udlæg af kløver-græsblanding med urter. Vårbyg og specielt egentlig brak gav dermed de dårligste betingelser for insektfaunaen (Hald & Lund 1994).
Vegetationen har således stor betydning for dyrelivet og udsåning af bestemte arter kan i nogen grad bruges til at styre dette. Tuedannende græsser i randstriben er specielt velegnede til at fremme løbebillers overvintring (Woodcock et al. 2005), og udsåning af blomsterblandinger fremmer andre nyttedyr. Humlebier tiltrækkes i høj grad af udsåede blomstrende urter. De blomstrende urter bidrager med nektar- og pollenkilder til humlebier, som har mindre gode trækmuligheder på enårige ukrudtsarter (Pywell et al. 2005) (Carvell 2007).
I Sverige er værdien for dyrelivet af randstriber på 6 og 20 m undersøgt (Chiverton 1999). Randstriberne var dyrket med hhv. havre, sennep, boghvede, kløvergræsblanding og en blanding af havre, sennep, solsikke eller boghvede. I det første år overvintrede flest prædatorer i kløvergræsrandstriben. Snyltehvepse og insekter af værdi som føde for agerhøns var der flest af i havre-sennep-solsikke randstriben. Valg af 6 eller 12 m havde lille effekt på de flest leddyr-grupper i denne undersøgelse. Sidst på vinteren var der 6-10 gange så mange småvildt i randstriberne som i tilsvarende områder uden randstriber. I England er den positive betydning af markrand og sprøjtefrie bufferzoner for vildt velundersøgt (Boatman & Wilson 1988;Vickery, Carter & Fuller 2002) (se også afsnit 5.6).
Man fandt en højere tæthed af løbebiller i lidt bredere randstriber i en irsk undersøgelse, som også satte spørgsmålstegn ved støtte til at lave huller i hegn, da hullerne påvirkede løbebilletætheden negativt (Feehan, Gillmor & Culleton 2005) (se kapitel 3).
Ældre randstriber (6 år gamle) har større prædatorpopulation, især af edderkopper, og højere prædator:bytte andel end yngre randstriber. Ældre randstriber har dog stadig lavere prædator:bytte andel end store brakmarker (Denys & Tscharntke 2002). Artsrigdom af insekter er kun marginalt påvirket af arealstørrelse og ikke af randstribens alder, dvs der er ikke påvist sammenhæng mellem artsdiversitet og biologisk bekæmpelsesfunktion (Denys & Tscharntke 2002).
Plejning af randstriben har betydning for insektfaunaen. Woodcock et al. (2007) fandt i en sammenligning af græsbevoksede randstriber flest biller i uplejede græsstriber samt i striber, som fik taget ét slet i juli. Endvidere fandtes de største tætheder af blomster- og frøfouragerende biller i ikke-gødede græsstriber.
5.3 Bufferzone
En bufferzone eller sprøjtefri randzone udgøres af en 1-flere m bred rand af den dyrkede mark. Dvs. at den indadtil grænser op til marken og udadtil grænser op til markranden. En bufferzone er dyrket med samme afgrøde som den øvrige mark. Er der inddraget en del af marken i markranden som udyrket, eller sået med en anden afgrøde – typisk græs eller en frøblanding, regnes dette som (usprøjtet) randstribe.
Ved etablering af sprøjtefrie randzoner søges et eller flere af nedenstående mål opfyldt:
- Beskyttelse af nytteorganismer som kan bidrage til regulering af skadegørere i dyrkede afgrøder
- Beskyttelse af flora og fauna i mark, hegn, vandløb mm, mod pesticider
- Beskyttelse af sjældne arter eller af artsdiversitet generelt
En af fordelene ved sprøjtefrie randzoner er, at pesticidafdrift fra marken til nærliggende vandløb, hegn mm begrænses. Beskyttelse mod pesticidafdrift er væsentlig for flora og fauna i markranden og i omgivelser.
Reduktionen i afdrift afgøres af:
a. Bufferzonens bredde
b. Bufferzonens højde
c. Sprøjteudstyr
d. Vindhastighed
e. Afgrødetype (f.eks større afdrift fra frugtplantager)
Depositionsmålinger ved en sprøjtefri randzone på 6 m i marken (10 m fra hegnet), i den sprøjtefrie randzone (3 m fra hegnet) og i fodposen (0,5 m fra hegnet) viste normalt afsætning i den sprøjtefrie randzone. Dog aldrig over 4.8% af markdosis. I hegnskanten var depositionen gennemsnitligt endnu 2-10 gange lavere. I en forsøgsdel uden randzone var deposition i hegnskanten 0,5 m fra sprøjtet mark til 50-60 % af markdosis (Hald et al. 1988). Data bekræftes af andre undersøgelser (Cuthbertson & Jepson 1988). Tilsvarende resultater er fundet i Holland, hvor lave vindhastigheder førte til 2-7% afdrift til vandløb, og væsentligt mere på bredden –ca 25% ved lav vindhastighed. Med 3 m brede bufferzoner nedsættes afdriften til vandløb med 95%, med 6 m striber med 99% (de Snoo & de Wit 1998).
Afdrift afhænger også af højden på bufferzonen. Således var afdrift fra sprøjtning i sukkerroer ved en 1,25 m bred stribe elefantgræs halveret når elefantgræsset var klippet i afgrødehøjde sammenlignet med bar jord. En højde på 0,5 og 1 m reducerede afdrift i forhold til bar jord med hhv 80 og 90%. Sprøjteteknik (air assisted sprayer) kunne også nedsætte afdriften (Zande et al. 2000).
Ud over afdrift kan zoner og randstriber nedsætte afstrømning. Græsbræmmers mulighed for at reducere afstrømning er veldokumenteret, men variable, da mange faktorer påvirker effektiviteten (Lacas et al. 2005).
5.3.1 Flora i bufferzone
Afdrift af herbicider kan påvirke sammensætning af plantearter (Kleijn & Snoeijing 1997) som også indirekte påvist i danske undersøgelser (Aude et al. 2004a; Aude, Tybirk & Pedersen 2003b). Det er dokumenteret at bufferzoner kan nedsætte pesticidafdrift og –udvaskning. Således har Ravn & Friberg (2004) dokumenteret herbicideffekter på vilde planter op til 14 m fra marken med de kraftigste effekter indenfor de første 2 m fra marken, og de konkluderer på den baggrund, at sprøjtefri randzoner langs vandløb skal have en bredde på 10-20 m hvis zonen for at være effektiv.
Sprøjtefri randzone øger ikke besværlige ukrudtarters etablering (gråbynke, agertidsel), men forhindrer en bekæmpelse. Blomstrende ukrudt er mest almindelig i sprøjtefrie randzone og blomstrer tidligere (Hald et al. 1988). Ukrudtsniveauet øges i de sprøjtefrie randzoner, men ikke længere inde i marken. Dette er især udtalt i de første år, hvor pionerukrudt er betydende (Hald et al, 1988).
5.3.2 Fauna i bufferzone
Leddyr kan påvirkes direkte af sprøjtning med insekticider. For eksempel er edderkopper sårbare overfor insekticider. I en undersøgelse af bivirkninger af fenitrothion, bromophos-ethyl og deltamethrin var tæppespindere den mest alvorligt påvirkede leddyrsfamilie (Everts et al 1989). Indirekte påvirkes leddyrsfaunaen af herbicider pga. ændret plantesammensætning.
De enkelte arters årsrytme vil være med til at afgøre om og hvor meget de udsættes for pesticider. Desuden vil arters mobilitet have stor betydning for deres evne til at rekolonisere et område hvor de er blevet nedsat i antal efter sprøjtning.
Betydningen af bredden på sprøjtefri randzoner er undersøgt af flere forfattere. Flest undersøgelser har set på effekten af 6 eller 12 meter brede randzoner af den enkle grund, at dette passer med bredden på en spøjtebom – altså med indstillingen af sprøjteredskaber.
Hald et al (1988) opgjorde fire arter af løbebiller med faldgrubefælder placeret i hegn, sprøjtefri randzone og mark; spraglet kvikløber, hedeglansløber, markjordløber og markgrotteløber. Alle de fire undersøgte arter forplanter sig i marken. Det kan forklare at kun arten markgrotteløber, i den tre-årige forsøgsperiode blev signifikant mere talrig i sprøjtefri randzone. En opgørelse af andre løbebillearter i 1987 viste at især håret markløber og skovjordløber var mere talrige i den usprøjtede randzone.
Sprøjtefrie randzoner havde efter insekticidbehandling væsentligt højere individtæthed end tilsvarende sprøjtede. En forskel der stort set holdt sæsonen ud bortset fra de meget mobile grupper –snyltehvepse, cikader og rovbiller-samt edderkopper (Hald et al. 1988).
En kombination af depositions-målinger i stigende afstand fra sprøjtning og bioassay af disse depositioners giftighed overfor nyttedyr er et praktisk bud på, hvordan effekter på non-target organismer kan måles (Langhof, Gathmann & Poehling 2005).
5.4 Sammenfattende viden om markrand og bufferzone
Nedenstående tabel (5.1.) opsummerer værdien af henholdsvis randstribe med græs, blomster eller braklagt og bufferzone i henholdsvis korn, kartofler og sukkerroer med hensyn til nedsat afdrift og biodiversitet samt konsekvenser for markdriften.
Tabel 5.1. Randstriber og bufferzoner og deres værdi for nedsættelse af pesticidafdrift, fremme af biodiversitet, beskyttelse af nytteorganismer og konsekvenser for markdriften (Delvis efter De Snoo og Chaney, 1999).
| Bufferzone | Randstribe | |||||
| Korn | Kartofler | Sukkerroer | Græsstribe | Blomsterbræmme | Brakstribe | |
| Miljø | ||||||
| Nedsat afdrift | + | + | + | + | + | + |
| Biodiversitet | ||||||
| Flora | + | ± | + | 0 | 0/+, hvis der udsåes hjemmehørende arter | + (med årlig jordbearbejdning) |
| Insekter på planter | + | ? | ? | 0 | + | + |
| Invertebrater på jorden | ± | ± | ± | + | + (vinterhabitat) | + (vinterhabitat) |
| Fugle | + | ? | ? | + (vinterhabitat) | ± | + |
| Pattedyr | + | 0 | 0 | + (vinterhabitat) | ± | ? |
| Markdriften | ||||||
| Omkostninger | lave | lave | høje | høje | høje | høje |
| Jordbrugerens accept | + | - | - | + | ? | - |
5.5 Insektvolde, græsbælter og blomsterbræmmer inde i markerne
I moderne landbrug er mange levende hegn fjernet for at gøre markerne større og derved effektivisere driften. Dette har mindsket biodiversiteten i agerlandet og øget den rumlige fragmentering af økosystemet (Potts & Vickerman 1974). Lineære flerårige markstriber er blevet udlagt inde i markerne i et forsøg på genopretning af økosystemer og biodiversitet. Flerårige habitater er vigtige bl.a. for forekomsten af polyfage prædatorer, da de kan opretholde en relativ konstant forsyning af fødeemner for prædatorerne, og fordi den flerårige vegetation skaber gunstige levesteder og overvintringsmiljøer sammenlignet med de egentlige markflader (Dennis, Thomas & Sotherton 1994; Riedel 1991; Thomas, Mitchell & Wratten 1992; Varchola & Dunn 1999). Formålet med insektvolde, græsbælter og blomsterbræmmer i marken er at bringe ”kanteffekterne” ind på dyrkningsarealet. De ”reducerer” markstørrelsen og bidrager derfor til en større spredning af prædatorer og andre nyttedyr, som overvintre i markkanter ind i marken om foråret (Collins et al. 2002). Det kan f.eks. være løbebillerne spraglet kvikløber og hedeglansløber, som fra markkanterne spreder sig så tidligt i marken, at de kan prædere på bladlusene i disses etableringsfase (Sunderland & Vickerman 1980). Blomsterbræmmer anlægges med henblik på at tilføre supplerende fødekilder til markens nyttedyr, til bestøvere og til sommerfugle.
Insektvolde skabes ved to modsatrettede pløjninger. De er oftest tilsået med græs og typisk placeret i midten af større marker (Thomas & Wratten 1988). Blomsterbræmmer og græsbælter etableres ved udsåning af frøblandinger med frø af udvalgte – typisk hjemmehørende arter. Blomsterbræmmer anvendes meget i Tyskland og Schweiz (Eggenschwiler et al. 2004). Græsstriber anvendes mere udbredt i Europa. Græsstriber og blomsterbræmmer kan også udsås i forbindelse med markranden.
Videnskabelige undersøgelser til belysning af effekten af insektvolde m.m. inde i marken er samlet i tabel 2 i bilag A. Der er listet 16 undersøgelser af markstriber udlagt i marken, og disse undersøgelser sammenfatter i bred forstand den forskning, der er foretaget på området. Enkelte af disse undersøgelser har været publiceret flere steder og indeholder derfor flere referencer. Markstriber har generelt en markant positiv indflydelse på antallet af polyfage prædatorer som løbebiller og rovbiller. Undersøgelserne viser bl.a., at volde og bræmmer hurtigt bliver koloniseret af nyttedyr efter etablering. Temperatur-bufferede og veldrænede forhold i insektvolde medfører f.eks. øget overlevelse for løbebiller, rovbiller og edderkopper (Thomas, Goulson & Holland 2000b). Volde, græsbælter og blomsterbræmmer bevirker samtidig et skift i artssammensætningen fra primært kortlivede arter med høj frugtbarhed og stor dødelighed over mod længerelevende arter med lavere frugtbarhed, hvilket kan øge stabiliteten i agro-økosystemer (Mader 1988). Zonerne i marken med blomstrende planter kan også skabe et rigere udbud af fødekilder som f.eks. pollen og nektar for dyr, der er afhængige af disse ressourcer, f.eks. bier, sommerfugle og svirrefluer, men også for mange mere upåagtede insekter, som er vigtige fødeemner for højere trofiske niveauer (bilag A, tabel 2). Insektvolde og græs- og blomsterbræmmer skaber fristeder og øgede fourageringsmuligheder for fugle og mindre pattedyr (bilag A, tabel 2 & 4) og kan sandsynligvis bidrage som spredningsveje for flora og fauna i markerne.
Tuegræsset hundegræs er den mest velegnede græsart til insektvolde og græsbælter fordi tætheden af overvintrende prædatorer er høj (specielt af de vigtige forårsformerende prædatorer som glansløbere og spraglet kvikløber). Frøene er relativt billige og græsset er nemt at vedligeholde (Collins et al. 2003ab). Spredning af tuegræsser ind i marken er endvidere meget begrænset. I undersøgelserne af blomsterbælter inde i markerne er der anvendt et bredt udvalg af hjemmehørende blomstrende urter og ikke fremhævet særligt velegnede arter (bilag A, tabel 2). Andre undersøgelser har dog dokumenteret positive effekter af honningurtbræmmer på svirrefluer i markrande (Cowgill, Wratten & Sotherton 1993; Hickman & Wratten 1996).
For at være gode levesteder for flora og fauna skal bredden af de linære zoner i marken være stor nok til at tillade en uafhængig vedligeholdelse af zonerne (f. eks. slåning) men samtidig så smalle, at der forekommer en interaktion mellem markstribe og afgrøde (Mensah 2004). Oftest er volde og bræmmer 2-2,5 m brede (bilag A, tabel 2), men f.eks. de største koncentrationer af overvintrende prædatorer findes i de yderste 20 cm af de græsbevoksede kanter ind mod marken (Riedel 1992). Det er derfor sandsynligt at volde, græs- og blomsterbælter kan være væsentlig smallere og stadig have den samme effekt.
Den optimale afstand mellem blomsterbælter på 24 m (Cowgill, Wratten & Sotherton 1993; Heitzmann, Lys & Nentwig 1992; Hickman & Wratten 1996). Dette gælder muligvis også insektvolde og græsbælter. En passende afstand mellem insektvolde afhænger bl.a. af spredningen af prædatorer ind i afgrøden og de driftsmæssige aspekter. Mængden af nyttedyr i marken falder med øget afstand til flerårig vegetation (Coombes & Sotherton 1986; Navntoft et al. unpubl.), og det må derved være muligt at finde en optimal afstand mellem de linære strukturer for at opnå den størst mulige effekt på faunaen.
Orienteringen har betydning for lysindstrålingen og derved for mikroklimaet, hvilket igen påvirker insekterne. Riedel (1992) rapporterede om flest overvintrende nyttedyr på syd-siden af en insektvold.
På trods af formålet med de linære strukturer om at øge den funktionelle biologiske bekæmpelse er der kun få undersøgelser der har målt nyttevirkningen i selve marken. Kun 3 ud af de 16 undersøgelser omtaler prædation af skadedyr i marken som et forskningselement. Der blev undersøgt prædation på døde bladlus udlagt i marken (Thomas & Wratten 1988), prædation på levende bladlus (Collins et al. 1997; Collins et al. 2002) og prædation på flueæg udlagt i marken (Prasad & Snyder 2006). I undersøgelsen med levende bladlus blev der påvist lavere bladlusangreb i marken nær en insektvold, men ikke om dette havde en positiv effekt på udbyttet. Det er derfor meget usikkert om strukturerne opfylder deres primære formål. Omvendt er der heller ikke rapporteret om øgede skadedyrsproblemer som følge af udlægning af volde og bræmmer.
Insektvolde og græs- og blomsterbælter øger biodiversiteten bl.a. ved at øge markernes indhold af plantearter, nyttedyr og egnet fugleføde samt ved at blive udnyttet af sanglærker og ved at skabe egnede ynglesteder for dværgmus (bilag A, tabel 2).
5.5.1 Sammenfatning af insektvolde, græsbælter og blomsterbræmmer
Det primære formål med at udlægge insektvolde og bræmmer med græs og blomstrende urter ude i marken og i en vis afstand fra randzonerne har været at øge den naturlige skadedyrsregulering i afgrøderne. Generelt har det medført markant højere tætheder af nyttedyr både i selve striben men også ude i marken, og strukturerne har også medført en øget biodiversitet. Det mangler dog stadigt at blive demonstreret, at tilstedeværelsen af egnede levesteder og overvintringshabitater i marken i form af insektvolde, græsbælter og blomsterbræmmer medfører generel øget prædation af skadedyr i et omfang, der udbyttemæssigt retfærdiggør oprettelsen af sådanne kunstige habitater. Tuegræsser (specielt hundegræs) er velegnede til insektvolde og græsbælter, fordi tætheden af overvintrende prædatorer er relativ høj under tuerne. Bælter med blomstrende urter medfører dog højere overvintringstætheder, men der er lidt viden om hvilke specifikke blomstrende urter, der er mest velegnede evt. i kombination med tuegræsser. En oplagt mulighed er derfor at græsset suppleres med flerårige planter, f.eks. hvid okseøje, stor knopurt, alm. røllike, rødkløver og kællingetand, og potentialet i dette er ikke tilstrækkeligt belyst. Der er ikke rapporteret om øgede ukrudtsproblemer i markerne som følge af udlægning af linjeformede strukturer, men potentielle problemer med rodukrudt er dog formentligt et vigtigt aspekt i udlægningen af markstriber i markerne. Fordelen ved etablering af volde sammenholdt med flade strukturer er formentlig dræning, hvilket sandsynligvis øger insekternes overvintringsmuligheder. Græsbælter og blomsterbræmmer øger også overvintringstætheder af polyphage prædatorer (Prasad & Snyder 2006a), og etablering og pasning af volde er mere omkostningsfuld for landmanden. Fordele/ulemper af at anlægge en vold sammenholdt med en flad markstribe er dermed ikke tilstrækkeligt belyst. Oplysninger om den bedste orientering af voldene (N-S, Ø-V) er ligeledes mangelfuld. Den optimale bredde og indbyrdes afstand af strukturerne er heller ikke ordentligt undersøgt. Endeligt kan pesticidsprøjtninger i marken muligvis være et problem for flora og fauna på insektvoldene, men der foreligge ikke oplysninger om dette.
Der er stor lighed mellem markskel og insektvolde, og det understøtter vigtigheden af at bevare de allerede eksisterende skel. Den viden, der eksisterer om vegetationens sammensætning og pleje, kan sandsynligvis også overføres direkte til pleje af de allerede eksisterende markskel.
5.6 Fugle og andre højerestående dyr
Fugle og andre højestående dyr er beskrevet selvstændigt som overordnede bioindikatorer i agerlandet. Petersen (1998) undersøgte fordelingen af fugle i det danske agerland og fandt, at artsantallet var negativt korrelleret med afstand til permanent græs, vådområder og skov og positivt korrelleret med antallet af tørre småhabitater, våde småhabitater med vedagtig bevoksning, haver og landbrugsbygninger. Bl.a. levende hegn er nødvendige habitater for mange fuglearter i agerlandet og understøtter et diverst fugleliv ofte med høje tætheder sammenlignet med andre habitater som skov og åbne marker (Moore, Hooper & Davis 1967). Kendskab til at forvalte levende hegn i relation til fugle og andre pattedyr i bevarelsesøjemed er derved vigtig (Fuller et al. 2001; Whittingham et al. 2005). Overordnet er højde og bredde de to vigtigste hegnsfaktorer, som er positivt relateret til fuglenes artsrigdom og antal, selvom store hegn ikke er en fordel for alle fuglearter (Hinsley & Bellamy 2000). En hegnsbredde og højde på ca 5 m tilgodeser mange af agerlandets hegnstilknyttede fugle (Green, Osborne & Sears 1994). Det skal bemærkes ved sammenligning til danske forhold, at der er forskel på engelske og danske hegn, hvor engelske hegn typisk minder om store velpassede hække (B.S. Petersen pers. com.). Hegnenes botaniske og strukturelle kompleksitet er dog også vigtig f.eks. indehold af høje træer, der tjener som sangposter (Hinsley & Bellamy 2000). Udover at være hjemsted for vigtige fødeemner om sommeren, producerer levende hegn desuden fugleføde til vinteren i form af bær og andre frugter (Hinsley & Bellamy 2000).
Levende hegns værdi for fugle kan generelt øges ved at kombinere med andre faktorer bl.a. ekstensive/braklagte randzoner, bælter med udsåede vildfugleblandinger og tilknyttede volde og grøfter (Hinsley & Bellamy 2000). Et af de bedst kendte eksempler på værdien af sprøjtefri randzoner for fugle blev leveret af Potts (1986), som viste at 6 m sprøjtefri randzoner markant reducerede dødeligheden af agerhønse- og fasankyllinger som følge at et øget udbud af ukrudt- og insektfødeemner. Specielt viste Potts (1986) en stærk sammenhæng mellem forekomsten af pileurter, pileurtbladbille og agerhøns. Sidenhen er gulspurven blevet et andet velstuderet eksempel på en randzonetilknyttet fugl, som drager fordel af ekstensive randzoner. Arten kræver et territorium med et egnet redested f.eks. i levende hegn (helst brede men samtidig korte til mellemlange) og bevoksede grøfter, et egnet siddested til sang (f.eks. højt træ) og umiddelbar nærhed af føderessourcer, da fouragereingen oftest finder sted indenfor 100 m fra reden (Whittingham et al. 2005). Lille (1996) opgiver en fødesøgningsradius på 250 m, som kan udvides til 350 m. Lille (1996) har vist, at brakstriber på 25 m har stor positiv indflydelse på gulspurve, som bl.a. andet giver udslag i form af øget fouragering på disse arealer og i en højere ungevægt, der er positiv korreleret til ungeoverlevelse. Stubmarker og brakmarker i umiddelbar nærhed af levende hegn er siden vist at øge forekomsten af gulspurve i hegnene (Hancock & Wilson 2003), men også græsbevoksede (både slåede og ikke-slåede) randzoner er foretrukne fourageringssteder for ynglende gulspurve (Perkins et al. 2002). Bradbury et al. (2000) fandt ved undersøgelse af flere faktorers indflydelse på antallet af gulspurvens yngleterritorier, at bredden af ukultiverede græsbevoksede randzoner havde den stærkeste og mest konstante positive relation til antallet af territorier (figur 5.1). I modsætning til Lille (1996) viste Bradbury et al. (2000) dog ikke sammenhæng mellem udnyttelsen af tilstødende arealer og redesucces. Hart et al. (2006) viste, at gulspurveungers vægt (som er relateret til deres overlevelseschancer) er positivt korreleret med mængden af vigtige insektfødeemner i yngleterritorierne.
Figur 5.1. Tætheden af gulspurveterritorier som funktion af bredden af græsbevoksede randzoner. * indikerer signifikant effekt af zonebredde (Efter Bradbury 2000).
Tilsyneladende har gulspuvene fordel af flere forskellig slags ekstensiv randzonvegetation. Vickery, Carter & Fuller (2002) sammenlignede udbuddet af fugleføde i flere forskellige slags randzonevegetation. Forfatterne fandt generelt, at det største fødeudbud for fugle om vinteren i arealer tilsået med ”vildfugleblandinger” og egentlig brak i omdrift. Om sommeren var fødeudbuddet størst i græs iblandet vilde blomster, vildfugleblandinger og egentlig brak i omdrift efterfulgt af usprøjtet afgrøde (korn). Rene græsbælter havde den laveste fødeværdi, men værdien af rent græs var stadig større end konventionelt korn (Vickery, Carter & Fuller 2002). Ekstensive rand- og bufferstribers vegetation og bredde/areal er dermed vigtig for gulspurve.
Fugle tilknyttet markfladen som sanglærke og vibe kræver åbne arealer med lav vegetation uden hegn og vildtremiser i nærheden (Hinsley & Bellamy 2000), bl.a. fordi dette mindsker risikoen for prædation fra f.eks. krager (Donald et al. 2001, B.S. Petersen pers. comm.). Ekstensive zoner som insektvolde eller lignende midt i markerne kan imidlertid tilbyde vigtige ressourcer i form af føde og skjul til lærker og andre fugle tilknyttet åbent land (Firbank & Smart 2002; Thomas, Goulson & Holland 2000b). F.eks. foreslår ”The Royal Society for the Protection of Birds” - UK bl.a. 20 m brede græsbælter i permanente brakmarker og 2 småfelter med lav vegetation a ca. 20 m² pr. ha i vintersæd for at øge lærkernes ynglesuccess (som hæmmes af høj tæt vegetation som f.eks. vinterhvede). Småfelterne kan øge lærkeproduktionen mere end 50% i større marker (> 5 ha) med vinterkorn (http://www.rspb.org.uk/). Timingen af evt. slåning af zonerne er imidlertid vigtig, da den ødelægger reder og unger, hvis den udføres for tidligt (skal helst udføres efter 1. juli - B.S. Petersen pers.com.).
At sanglærker har gavn af sådanne mikrohabitater kan primært tilskrives deres præference for lav vegetation, som muliggør en relativ uhindret fødesøgning på jordoverfladen, snarere end at der er mere føde (Odderskær et al. 1997). Boatman & Bence (2000) har endvidere vist, at ”braklagte” striber tilsået med vildfugleblandinger (20 m brede) er foretrukne yngleområder for sanglærker. Ialt fandt Boatman & Bence (2000) 67 % af alle lærkereder disse striber, som udgjorde 8-10 % af undersøgelsesarealet (se tabel 4, bilag A). Boatman & Bence (2000) fandt endvidere at brak tilsået med ”vildfugleblanding” blev foretrukket frem for egentlig brak.
Sprøjtefri randzoner om alle marker er ved modellering vist at kunne fremme lærkebestanden i områder med få levende hegn (Det Økonomiske Råd 2004).
Bomlærken, som også er tilknyttet markfladen, har ligeledes præference for at fouragere for ekstensive græsarealer og vårbyg, men småhabitater som randstriber, bufferzoner med græs og brak, billevolde og selektivt sprøjtet forager blev også foretrukket til fouragering, og kan derfor udgøre vigtige småhabitater for bomlærken (Brickle et al. 2000). Brickle et al. (2000) fandt endvidere, at bomlærkernes ungevægt var positivt korreleret med insektfødemængden, som var negativt korreleret til antallet af insekticidbehandlinger.
Den gunstige effekt af markstriber for markfladens fugle kan imidlertid også udgøre et problem for markfladens jordrugende fugle, da de kan lede prædatorer ind i marken (Donald et al. 2001).
Ekstensive randzoner medfører flere mindre pattedyr som f.eks. skovmus (Tew, Macdonald & Rands 1992), og dværgmus har også stor fordel af græsbevoksede randstriber og markstriber som ynglesteder både i markranden og på insektvolde (Bence, Stander & Griffiths 2003). Derved øges jagtmulighederne for rovfugle som tårnfalke og slørugler (Vickery, Carter & Fuller 2002). Andre mindre pattedyr kan også have gavn af beskyttede randstriber og markstriber. Boatman & Stoate (2000) viste en markant fremgang for harer (se tabel 4, bilag A), men da udlagte beskyttede linjer kun var en del af flere tiltag, kan den specifikke effekt ikke adskilles fra andre dyrkningsmæssige tiltag.
Af den skematiske gennemgang af hegn, bufferzoner og andre småhabitaters betydning for agerlandets fugle og pattedyr (bilag A tabel 4) fremgår en klar tendens til markant fremgang for de enkelte arter som respons på de forskellige tiltag der skal fremme bestandene. Ofte er der dog foretaget flere ændringer samtidig som f.eks. opsplitning af marker samtidig med en generel reduceret pesticidanvendelse (Boatman & Bence 2000; Boatman & Stoate 2000), hvilket vanskeliggør en nærmere vurdering af de specifikke bufferzonetiltags betydning for bestandsfremgangen af eksempelvis harer.
Et andet dyr, som er trængt af ændringer i landskabet, er markfirbenet. Dette krybdyr har en relativ lille bevågenhed men er trængt bl.a. som følge af rydning af hegn og stengærde og øget næringsstoftilførsel til hegn og græsrabatter, hvorved vegetationen bliver tættere om mindre egnet til krybdyr (Fog 1997). Det er muligt at ekstensive randzoner, evt. med barjordsstriber, vil fremme bestanden af firben og andre krybdyr.
5.6.1 Sammenfatning om fugle og andre højerestående dyr
Forskelligheder i markskel, f.eks. om der er tale om et tæt levende hegn, et relativt åbent hegn eller bare en græsstribe og kvaliteten heraf, er afgørende for hvilke fugle, der findes. Levende hegn og andre småbiotoper med vedagtig vegetation er en forudsætning for en række af agerlandets fugle, men generelt klarer fuglene i hegnet sig godt. Ekstensive randzoner er dog vist at fremme den hegnstilknyttede gulspurv, og som et vigtigt element er det vist, at bredden af ekstensive randzoner er positivt relateret til gulspurve, og derfor sandsynligvis også for andre fugle, som har yngleterritorier, der omfatter randzoner tilknyttet hegn. Det er dog stadig ikke vist, at bredden/arealet af ekstensive randzoner er direkte relateret til øget ungeproduktion af gulspurve. Ligeledes vides det heller ikke hvilken randzonevegetation, der giver den største ungeproduktion (f.eks. énårig brak vs. ekstensivt dyrket korn med udlæg) og sikrer ungerne de største overlevelseschancer.
Markfladens fugle er i tilbagegang (bl.a. lærker og viber), men ekstensive zoner i marken er vist at fremme fuglebestande. Der vides, at markfladens fugle undgår høj vegetation, og der er også en vis viden om hvilken vegetation, der er bedst egnet til i de ekstensive zoner. Det vides dog ikke, hvor stort et areal af ekstensive zoner (herunder udformningen af disse), som er nødvendige i markerne for at mindske/stoppe tilbagegangen af markfladens fugle. Det vides heller ikke i hvilket omfang ekstensive zoner i markerne kan virke som ”fælder” for ynglefuglene ved at naturlige fjender kan øge prædationen uhensigtsmæssigt som følge af nemmere lokalisering af rederne.
Også for mindre pattedyr som mus er der observeret fremgang som følge af beskyttede randzoner.
6 Økonomiske aspekter af randzoner
- 6.1 Markrandes effekt på udbyttet
- 6.2 Ukrudt, sygdomme og skadedyr relateret til markrand og bufferzone
- 6.3 Sprøjtefri randzoner som pesticidreducerende virkemiddel
- 6.4 Incitamenter til udlægning af bufferzoner
- 6.5 Økosystemtjenester og eksternaliteter
- 6.6 Sammenfatning af økonomiske aspekter
De levende hegn har de sidste årtier fået stigende betydning for jagt, natur, landskab og ejendommens herlighedsværdi. Men det er stadig vigtigt at være opmærksom på muligheden for at opnå et merudbytte i forbindelse med etablering af nye hegn.
6.1 Markrandes effekt på udbyttet
Forskellige undersøgelser viser et gennemsnitligt merudbytte for alle afgrøder på 5-15 % i hegnenes læzone (20 gange hegnshøjden), i forhold til udbyttet udenfor læzonen (>20 gange højden). Tæt ved hegnet (0-3 gange højden) er udbyttet dog lavere. Udbyttet påvirkes ligeledes positivt af markrandens bidrag med nyttedyr. Bedst dokumenteret er effekten på bladlus.
I markranden er der altid lavere udbytte end længere inde i marken med et trend mod højere udbytte fra kanten mod midten af marken, og lavere eller tydelig lavere udbytte i kørespor og foragre. Desuden påvirkes randzoner af hegn og andre beplantninger, som konkurrerer om lys, vand og næringsstoffer, og er vandlidende langs vandløb og søer. Markrandens betydning på udbytte afhænger af afgrøde med store forskelle fra brug til brug.
Effekten af markrand på udbytte i sukkerroer, hvede og byg blev undersøgt i England 1992-97. Der var altid højere udbytte væk fra markrand, og særligt lavt udbytte i forager og sprøjtespor. Udbyttetab i markrand varierede stærkt mellem landbrug. I sukkerroe ydede markkanten 19-41% mindre end markmidten, med gennemsnitlige tab på 26%. I korn var de tilsvarende tal 3-19%, og et gennemsnit på 7%, med mindre tab i markforsøg end på kommercielle landbrug. Såning af græsstriber rykkede ikke disse markrandseffekter længere ind i marken, og påvirkede ikke afgrøden negativt. Skygge fra træer i markranden havde størst effekt på udbyttet. I de yderste 9 m af marken blev der høstet 4.4 t/ha hvede mens uskyggede dele af marken gav 8.1 t/ha. Vending af maskiner nedsatte også udbytte signifikant, mens harers og kaniners græsning ikke påvirkede udbyttet (Sparkes et al. 1998). Landsforsøg med usprøjtede randzoner i perioden 1987-1992 viste udbyttenedgange på 15-26%.
Med hensyn til sprøjtefrie randzoner er udbyttetabet i dækkende afgrøder begrænset. Udelades kunstgødning øges udbyttetabet. I mindre dækkende afgrøder som ærter kan ukrudt nedsætte udbyttet i usprøjtede randzoner.
Hald et al (1988) fandt udbyttereduktioner i randzonen af kornmarker på 15-51% sammenlignet med åben mark. Gennemsnitstabet blev opgjort til 3 hkg korn pr ha sprøjtefri randzone. De høje tab i dette forsøg skyldtes problemer med svampesygdomme. Udenlandske forsøg med usprøjtede randzoner i korn viste udbyttetab på 0-13%, men væsentligt højere, når kunstgødning også blev udeladt –helt op til 70-75% efter nogle år (Grub, Perritaz & Contat 1996; Snoo & Chaney 1999). Udbyttetabet i tyske usprøjtede og ugødede markrande var ca 30% (Raskin 1994). En hollandsk undersøgelse viste tab i sukkerroer og ærter på 24-30% ved usprøjtede randzoner, svarende til de engelske data for sukkerroer i randzoner uden særlig behandling. Samme hollandske undersøgelse fandt udbyttetab i kartofler på gennemsnitligt 2% (Snoo 1994).
Udbyttereduktioner i 12 m brede randzoner må forventes at være lavere end i 6 m brede zoner. Beregninger fra Fødevareøkonomisk institut har forsigtigt fastsat 10 % lavere udbytte end i standardkalkulerne. Endvidere har de regnet med et lavere udbytte som følge af restriktioner på pesticidanvendelsen langs målsatte vandløb og søer (Abildtrup 2004). Afstandskrav på 2 m fra vandløb og søer antages ikke at have dyrkningsmæssige konsekvenser, da de første 2 meter fra vandløb og søer udgøres af dyrkningsfri bræmmer. Der vil dog i nogle få tilfælde, f.eks. ved bekæmpelse af bjørneklo, kunne opstå en meromkostning ved ophør med sprøjtning af dyrkningsfri bræmmer. Ingen insekticider er godkendt til anvendelse i de 10 første meter fra målsatte vandløb og søer, og kun et begrænset antal fungicider er tilladt. Det vurderes, at denne restriktion begrænser en optimal beskyttelse mod svampesygdomme med 25%. (Jørgensen 2004). Det vurderes, at for standard-sædskifterne vil der ikke være problemer med ukrudtsbekæmpelsen med de tilladte midler langs vandløb og søer. Dette gælder dog ikke for sukkerroer, hvor det normalt ikke vil være muligt at gennemføre en effektiv ukrudtsbekæmpelse med de tilladte midler (Jensen 2004). Ved massive forekomster af flyvehavre kan der endvidere opstå en mindre meromkostning til lugning, idet sprøjtemidlerne mod flyvehavre har afstandskrav på 10 m. Kun et begrænset antal pesticider er omfattet af afstandskravet på 20 m fra vandløb og søer. Derfor vurderes afstandskravene ikke at have økonomiske konsekvenser for dyrkningen i 10-20 meters afstand fra vandløb og søer. Abildtrup (2004) fandt endvidere, at omkostningerne ved braklægning af randzonerne generelt ikke var højere end ved sprøjtefri drift. Dette skal ses i relation til, at de største miljø- og naturgevinster sandsynligvis opnås ved en kombination af sprøjte- og gødningsfri randzoner, som reelt vil medføre en udtagning af randzonen.
Sammenfattende kan det konkluderes at markrande i kraft af lævirkning og bidrag til funktionel biologisk bekæmpelse øger udbyttet, selvom sidstnævnte kræver yderligere dokumentation. Beregninger af udbyttetab må korrigere for det normalt lavere udbytte i den yderste del af marken, som i model af Kuemmel (2003).
6.2 Ukrudt, sygdomme og skadedyr relateret til markrand og bufferzone
Ukrudt findes i de højeste tætheder i markranden, og der er risiko for opformering af problematisk ukrudt som rodukrudt og ”høstbesværlige” og tabsvoldende arter som burresnerre ved udlægning af sprøjtefri og ekstensive randzoner. Ukrudtsstrigling er en mulighed for behandling af sprøjtefri randzoner, men metoden har ofte ringe effekt på rodukrudt og stor effekt på de mindre skadelige enårige ukrudtsarter, som har stor værdi biodiversitetsmæssigt. Risikoen for svampeangreb kan ligeledes være større nær markranden.
En række skadedyr er associeret til hegn og markrande. F.eks. overvintrer gulerodsfluer og de fleste snudebillearter som voksne i hegn, og flere bladlusarter overvintrer ligeledes i hegn og markrand. Hald et al. (1988) fandt, at skadegørende snudebiller, primært stribet bladrandbille var mest hyppige i hegn ved usprøjtet randzone, men fangsterne var lave. For kornbladbillen var der ingen signifikant effekt på tætheder i sprøjtefri vs sprøjtet randzone. Det vurderes ikke, at det øgede antal rapsskadedyr i usprøjtede randzoner ville have nogen effekt på disse arters populationsniveau. Højere tæthed af tæger bl.a. toplettet bederoetæge kan dog give problemer i bederoer og kartofler, hvor de kan optræde som skadedyr (Hald et al. 1988).
6.3 Sprøjtefri randzoner som pesticidreducerende virkemiddel
Der er foretaget analyser for at vurdere hvilken form for pesticidreduktion, der er mest hensigtsmæssig, når der tages højde for de økonomiske omkostninger og for konsekvenserne for vandressourcer og biodiversitet (Det Økonomiske Råd 2004; Jacobsen 2005). Som indikator for biodiversitet anvendtes bestanden af sanglærker, som forudsættes meget afhængig af fødegrundlaget. Fødegrundlaget forudsættes igen at have en nær sammenhæng med det generelle niveau for biodiversitet. Beregninger baseres på tre scenarier:
1. Ensartede pesticidafgifter, der giver en reduktion i pesticidforbruget på 25%.
2. Sprøjtefri randzoner omkring marker og eksisterende landindvindinger.
3. Delvis omlægning til økologisk jordbrug.
Ifølge de to kilders cost-benefit analyser vil Danmark drage størst fordel ved udlægning af sprøjtefri randzoner. Det Økonomiske Råd (2004) når frem til en randzonebredde på 5 m for de sprøjtefri randzoner, hvis den samfundsøkonomiske omkostning skal svare til omkostningen ved ensartede pesticidafgifter. Dette skulle betyde en fremgang i biodiversiteten på 10% (målt på markfugle). Jacobsen (2005) når frem til, at sprøjtefri randzoner skal omfatte mere end 20% af det totale landbrugsareal. Begge kilder anfører dog, at resultaterne skal fortolkes varsomt.
6.4 Incitamenter til udlægning af bufferzoner
På baggrund af en interview-undersøgelse udført af Jacobsen (2006) af 29 jydske landmænd udvalgt blandt 150 landmænd med jord til målsatte vandløb og søer vurderedes det, at ca. halvdelen af disse var interesseret i at etablere randzoner. Med de få interviewede er undersøgelsen naturligvis ikke bredt repræsentativ. Imidlertid gav undersøgelsen anledning til følgende anbefalinger for at øge mængden af randzoner:
1. Der skal være en klar udmelding om, at ekstensive randzoner kan etableres med støtte i hele landet, selvom den generelle støtte måske er begrænset til Natura 2000 arealer.
2. At der samlet skal være en økonomisk gevinst ved at deltage i ordningerne, hvis mange flere skal deltage i ordningerne. Den økonomiske gevinst må ikke blive ædt op af administrationsomkostninger. På nuværende tidspunkt vurderes det, at kun en mindre del af de etablerede randzoner vil blive tildelt MVJ-tilskud da tilskuddet pr. bedrift er lavt.
3. Mere information til lodsejere om de miljømæssige gevinster ved randzoner (primært fosfor og pesticider).
4. Ansøgning om MVJ-støtte kunne kombineres med enkeltbetalingsstøtten og derved give en administrativ lettelse.
5. På mindre bedrifter er der ikke krav om braklægning, og der synes på disse bedrifter at være et potentiale for at øge etableringen af randzonearealet.
(Jacobsen 2006)
Negativ omtale angående manglende gentegning af MVJ-aftaler, få midler og usikkerhed om situationen efter udløb af MVJ-aftaler i Natura 2000 kan begrænse indgåelsen af yderligere aftaler (Jacobsen 2006). Ifølge undersøgelse af Jacobsen 2006 er der kun ca. 1 ha støtteberettiget areal pr. bedrift. Rådgiveren skal typisk have 1.500 – 2.000 kr. for at lave en ansøgning, hvilket betyder, at landmanden ikke får økonomisk glæde af de to års tilskud.
Hvorvidt undersøgelsen af Jacobsen (2006) er repræsentativ for udlægning af sprøjtefri randzoner langs hegn og markskel vides ikke. Det er imidlertid dokumenteret at villigheden til at etablere bufferzoner er afhængig af støtte.
En hollandsk undersøgelse påviser stigende villighed til at etablere bufferzoner med 20% villige med en støtte på 1000 Fl./ha stigende til 60% ved 3000 Fl/ha, hvorefter andel villige ikke øges med øget støtte (Wossink & Wenum 2003). Tilsvarende viser tyske erfaringer at ved ophør at tilskud til sprøjtefrie bufferzoner gik arealet hurtigt ned (Albrecht & Mattheis 1998). En engelsk undersøgelse viste at Countryside Stewardship Programmet (se tabel 1 i bilag A) faktisk dækkede de reelle udgifter til at etablere randzoner og bufferzoner (Wilson 1997).
6.5 Økosystemtjenester og eksternaliteter
Biologisk mangfoldighed er defineret med De Forenede Nationers Konvention om den biologiske mangfoldighed fra 1992: Med biologisk mangfoldighed forstås mangfoldigheden af levende organismer fra alle kilder, herunder terrestriske, marine og andre akvatiske økosystemer og de økologiske strukturer, de indgår i; mangfoldigheden omfatter såvel variationen indenfor og mellem arterne som mangfoldigheden af økosystemer (FN, 1992).
Et økosystem er et dynamisk system af plante-, dyre- mikroorganismesamfund og disses abiotiske miljø, der vekselvirker som en funktionel enhed. Definitionen af biologisk mangfoldighed skal således forstås på tre niveauer; a) mangfoldigheden i samspillet mellem arter og deres omgivelser i økosystemer, b) mangfoldigheden af forskellige arter af planter og dyr, herunder også mikroorganismer, c) den genetiske mangfoldighed eller arvelige variation, der findes blandt forskellige individer og bestande af samme art (Det Økonomiske Råd 2000).
Beskyttelse af natur og biologisk mangfoldighed er med andre ord ikke alene et spørgsmål om at beskytte plante- og dyrearter. Beskyttelse af natur er også et spørgsmål om at sikre velfungerende økosystemer, som “gratis” yder en række økosystemtjenester (ecosystem services) som for eksempel recirkulering af næringsstoffer, nedbrydning af giftige forbindelser, omsætning af affaldsprodukter, bestøvning af kulturplanter og klimaregulering (Miljø- og Energiministeriet, Wilhjelmudvalget 2001).
Økosystemtjenester er væsentlige for jordbrugsproduktion, men arbejdet med at værdisætte disse er relativt nyt. Det sammen gælder i nogen grad for beregninger af eksternaliteter i landbruget. Constanza et al. (1997) beregnede/ skønnede verdens samlede økosystem tjenester til 33 · 109 USD/år.
De fleste potentielle skadedyr (95%, 100,000 arter af leddyr) reguleres naturligt; det vil sige at al bekæmpelse er rettet mod de tilbageværende 5.000 ater af leddyr (van Lenteren 2006). Denne økosystemfunktion af naturlig biologisk bekæmpelse vurderes at have en samlet værdi af 400 milliarder US$ årligt på verdensplan (Costanza et al., 1997), hvilket er et meget højt beløb sammenlignet med hvad der årligt anvendes på insekticider (8.5 milliarder US$/år). Omregnet til arealbasis satte man værdien af biologisk bekæmpelse i afgrøder til 24 USD/ha årligt, bestøvning til 14 USD/ha og fødevareproduktion til 54 USD/ha dyrket mark. Disse første forsøg på opgørelser af værdien af økosystemtjenester er i sagens natur usikre og kontroversielle. Efterfølgende har flere studier forsøgt at belyse værdien af økosystemtjenester, og mere sikre tal må forventes i fremtiden.
Ligesom værdien af biologisk bekæmpelse formetlig er underestimeret er værdien af biodiversitet og biodiversitetstab ved en fortsat forarmning af naturen det (Dobson 2005; Kremen & Ostfeld 2005). Et eksempel på at øget afgrødediversitet kan give øget udbytte kommer fra England, hvor man påviste større udbytte med blandinger af stigende antal udsåede arter i græs til hø (Bullock et al. 2001).
Eksempler på undersøgelser, der påviser randzoners potentielle øgning af funktionel biologisk bekæmpelse er få og nogle er ikke entydige. For eksempel fandt Powell et al. (2004) fandt stærk indikation for at blomsterrige randstriber havde reducerende effekt på bladlusangreb i korn som følge af højere tætheder af bladlusspecifikke prædatorer som svirrefluer. Dog reducerede randstriber tilsået med vildt-blandinger (kål, hirse, honningurt, triticale, solsikke, stenkløver samt diverse naturligt forekommende ukrudt) ikke ærtebladlus i ærtemarker og øgede antallet af bladlus i korn. Sådanne resultater understreger behovet for undersøgelser, der konkret vurderer den anvendte plantesammensætning i forhold til skadegørere og nyttedyr, som i påvist kål (Winkler, 2005) (se i øvrigt kap. 5 for yderligere information).
Hvis det antages, at ekstensive og diverse randstrukturer har en positiv effekt på afgrødeudbytter, hvor meget diversitet eller markrand er da nødvendigt for at sikre den nødvendige diversitet? Der er også meget lidt viden på dette område. Et eksempel er en undersøgelse omfattende det sydlige Europa, hvor mariehøns er vigtige i bladlusbekæmpelsen. Her viste en modelberegning, at landskaber med 9 -16% ikke-afgrøde habitat havde tilstrækkeligt med syvplettede mariehøner til at bekæmpe bladlus, mens landskaber med 1-4% ikke-afgrøde habitat ikke var tilstrækkeligt. I landskaber med tilstrækkeligt ikke-afgrøde habitat var bladlusbekæmpelsen bedst med jævnt fordelte læhegn frem for større sammenhængende stykker af ikke-afgrøde habitat (Bianchi & Van Der Werf 2003).
Eksternaliteter, det vil sige omkostninger, som ikke betales af den individuelle landbruger men bæres af samfundet, herunder f.eks vandløbsforurening, er beregnet til 208£/ha inklusive græsmarker eller 2343 mill £ for hele Storbritanien. Af disse er de væsentligste i forhold til denne rapport pesticidforurening af drikkevand, som er opgjort til120 mill £, eller ca. 5% af de samlede eksternaliteter, svarende til 105 kr/ha, skader på vildt, hegn, diger m.m. (125 mill £, eller ca 5% af de samlede eksternaliteter) og jorderosion og kulstoftab (106 mill £, eller ca. 4,5% af de samlede eksternaliteter). Disse eksternaliteter (i alt ca 300 kr/ha) kan randzoner og bufferzoner anvendes til at nedbringe (Pretty et al. 2000).
Disse forhold har ført til overvejelser om, at værdisætning af landbrugsvarer i fremtiden vil skulle afkobles fra produktionsomkostningerne og overflyttes til værdien af økologiske goder og tjenester (Gerowitt, Isselstein & Marggraf 2003), blandt andet drevet af fødevareindustriens krav til produktion med mindre pesticider (Levidow & Bijman 2002). Flytning af EU tilskud til landbruget over til arealtilskud kan betragtes som et skridt i denne retning.
For at forstå de negative og positive sider ved landbrugsdrift for bevarelse af biodiversitet behøves et landskabsperspektiv. Komplekse landskaber fremmer lokal biodiversitet, som kan kompensere for høj-intensiv drift. Det betyder, at landbrugsmiljøprogrammer vil have større effekt i simple end i komplekse landskaber (Tscharntke et al. 2005). Der mangler dog væsentlig viden om den relative betydning af lokale effekter versus landskabseffekter på biodiversitet, og hvordan disse forholder sig til økosystemtjenester, før der kan gives konkrete anbefalinger (Tscharntke et al. 2005).
6.6 Sammenfatning af økonomiske aspekter
Læeffekten fra hegn øger udbyttet, ligesom markrande kan bidrage med nytteorganismer og øget biodiversitet. Der er mindre udbytte nær hegn. Tabet reduceres med afstand til hegn og samlet giver læhegn øget udbytte. Tabet ved at etablere bufferzoner langs hegn er derved begrænset afhængigt af hvor meget udbyttet er lavere, og arbejdsmæssigt kan ’besværlige hjørner’ spares.
Oprettelse af randzoner og bufferzoner øger den funktionelle biodiversitet og er hermed kilder til funktionel biologisk bekæmpelse, men der er en udtalt mangel på viden om udbyttemæssige effekter af øget funktionel bekæmpese som følge af oprettelse af ekstensive randzoner.
Randzoner og bufferzoner bidrager til reduceret pesticidafdrift.
Udenlandske undersøgelser peger på at tilskuddets størrelse er vigtigt for, hvor store arealer der kommer ind under landbrugsmiljøordningerne.
Økosystemtjenester som biologisk bekæmpelse og eksternaliteter som pesticidafdrift er værktøjer til at dokumentere den samfundsmæssige værdi af mere miljøvenligt landbrug.
7 Diskussion
Som nævnt tidligere i denne udredning kan randzoner og bufferzoner udlægges med tre indgange: 1) En naturmæssig, hvor formålet vil være at opretholde eller berige biodiversiteten, 2) En kombineret landbrugs-og naturmæssig, hvor formålet vil være at beskytte tilgrænsende habitater (vand el. hegn) mod påvirkninger af landbrugsdriften, især pesticidpåvirkninger, og 3) En primært landbrugsmæssig, hvor formålet er at øge den naturlige regulering af skadevoldere for derved at mindske udgifter til bekæmpelse.
Selv om denne opdeling er hensigtsmæssig for at holde klarhed i begreber og formål, kan der i praksis være en sammenhæng mellem alle tre ikke mindst set ud fra et eventuelt overordnet ønske om at modvirke den forarmning af biodiversitet i det danske landskab, som er en følge af landbrugsarealets andel af Danmarks areal og driftens intensitivitet. I denne sammenhæng kan den primært landbrugsmæssige indgang (3) ende med at have tre formål: A) at begrænse planteavlerens bekæmpelsesudgifter, B) at hjælpe planteavleren til en begrænsing af behandlingsindekset af pesticider (BI) og C) at forbedre landbrugsarealernes naturindhold. 1), 2) og 3) samt A), B) og C) anvendes som pejlemærker i denne diskussion.
De ganske omfattende undersøgelser af sprøjtefrie bufferzoner (randzoner) må siges at have givet rimelig solide demonstrationer af, at der bliver et bedre planteudbud i randen (bl.a. Hald et al. 1988, afsnit 5.2.2.1 & 5.3.1), og i sammenhæng med planterne bliver der også rigere forekomster af leddyr ikke mindst insekter. Blandt disse leddyr findes der en blanding af nyttedyr, skadedyr, og ”seværdige dyr” som dagsommerfugle (se tabellerne 2 & 3, bilag A). Det er påvist, hvorledes randen virker som overvintrinshabitat og leveringskilde af nyttedyr som løbebiller, rovbiller og edderkopper (bl.a. Duelli et al. 1990; Hausammann 1996, afsnit 5.2.2.2), og man må formode, at denne funktion yderligere gælder for mange flere arter end de beskrevne. Går man skridtet videre i fødenettet til fugle og andre højestående dyr, giver undersøgelserne en klar påvisning af forbedrede fourageringsmuligheder og yngelproduktion (afsnit 5.6). I den forbindelse er sammenhængen mellem mere pileurt, flere pileurtbladbiller og bedre overlevelse af agerhønsekyllinger et klassisk eksempel (Potts 1986). Således viser de samlede resultater henover planter, insekter og fugle rimelig entydigt, at beskyttede ekstensive randzoner er værdifulde tiltag, der kan forbedre biodiversiteten i agerlandet.
Biodiversitetsmæssigt er der grund til at tillægge randzoner langs hegn en særlig høj værdi med støtte i ovenstående forskningsresultater og almindelig økologisk teori vedrørende økotoner. Her spares ved placering af en beskyttende randzone netop den biologiske rigdom, som er et produkt af økotonen, og som i vid udstrækning reduceres ved brug af pesticidsprøjtning. Også randzoner langs f.eks. skel eller grøfter vil have en positiv effekt på biodiversiteten, men der mangler konkret viden om denne type randzone i forhold til randzone langs hegn. Udover den lokale nærvirkning af en randzone, kan den forbedre den landskabelige heterogenitet og derved forbedre biodiversiteten. Dette er dog vanskeligt at måle sikkert, hvis ikke man undersøger i en geografisk stor skala med flere rumlige niveauer (Tscharntke et al. 2005). Denne form for biodiversitetsforbedringer angives bl.a. at gælde en række bestøvere, f.eks humlebier og solitære bier. Positiv virkning må også formodes at gælde fugle, som ofte kan afspejle forhold i et større landskab (Baillie et al. 2000).
Endelig må man f.eks. med tilbageblik på tidligere udbredelse af visse af de nu trængte dyr som firben (Fog 1997) formode, at der kan ske en delvis reetablering en række steder i agerlandet med udgangspunkt i velvalgt placerede randstriber. En række hegnstilknyttede fuglearter vil i sig selv kunne fremmes med flere levende hegn og der er dokumentation for bedre ungeproduktion, hvis nærforekomster af føde (som frø og insekter) forbedres gennem etablering af ekstensive randzoner langs sådanne hegn.
Et helt naturligt spørgsmål er, hvilken bredde randzonen bør have af hensyn til biodiversitetsforbedring. Det savnes der en mere præcis viden om. Meget tyder på, at de tidlige engelske og danske undersøgelser benyttede 6 m’s bredde, fordi man derved blot kunne lukke den ene halvdel af datidens 12m brede sprøjtebom. Dog har en nyere hollandsk undersøgelse (de Snoo & de Wit 1998) påpeget en betydelig værdi af en blot 3m bred bufferzone langs vand, hvorfor også et hegn må opnå en beskyttelse af blot en så smal stribe. Dog peger også en enkelt undersøgelse (Feehan, Gillmor & Culleton 2005) på, at 1,5 m er væsentligt ringere for løbebiller end 2 m. Sammenholder man disse resultater, giver det anledning til at formode, at der for mange arter vil være en stigende forbedring med stigende randbredde, derpå et maksimum fulgt af enten plateau eller fald. Den ekstensive randzones værdi understøttes af Cuthberson & Jepsons (1998) resultater vedrørende 6 m bufferzone (sprøjtefri rand) langs hegn. Imidlertid mangler der viden om, hvilken randzonebredde, der tilgodeser flest arter af både planter og dyr set i forhold til etableringsomkostninger og drift. Ligeledes mangler der en metodisk tilgang, der i nogen grad kan binde tidligere og kommende undersøgelser sammen, f.eks. ved at man anvender 3m som udgangsmodul (3x1 (de Snoo 1999), 3x2=6 (Potts 1986; Hald et al. 1988; Hald & Reddersen 1994 m.fl.), 3x3 og 3x4).
Også kvaliteten af randzonernes vegetation (brak, ekstensivt dyrket korn, græs, blomsterblandinger og forskellige kombinationer heraf) og vegetationens alder og pleje (f.eks. slåning) er delvis belyst (afsnit 5.2.2.1 & 5.3.1), men der mangler en systematisk belysning i relation til biodiversitet. Endelig er der foretaget nogle undersøgelser, der ser på betydning af mark: rand forholdet (B. Ekbom pers. com.), men heller ikke her kan der siges at være nået en operationel størrelse i relation til rand og areal. Jacobsen et al. (2005) fremfører ud fra økonomiske forudsætninger og under sammenligning med delvis omlægning til økologisk drift eller generel dosisnedsættelse til 25%, at ”man får mest biodiversitet for pengene” ved at bruge 20% af landbrugsarealet på bufferzoner på 5m’s bredde. Der mangler dog yderligere undersøgelser af betydningen for biodiversiteten specielt i relation til varierende bredde og placering af randzoner. I en større sammenhæng ville det være ønskeligt at vide, om f.eks. 5.000 km randzone á 3 m’s bredde har samme værdi mht. biodiversitet, landskabelig fremtoning og rekreationsværdi som 2.500 km randzone á 6 m’s bredde. Hvis den smalle randzone er tilstrækkelig god, vil der være bedre mulighed for at udtage den udbyttemæssigt ringeste del, og der kunne spekuleres i skabelse af sammenhæng (connectivitet) til modvirkning af den betydelige fragmentering, som præger det danske agerland.
Randzoners driftsmæsige fordele er et stadigt diskuteret emne. At rande bidrager med nyttedyr i marken er sikkert. Der er også et relativt godt kendskab til hvilken bevoksning der er bedst egnet til forskellige slags nyttedyr, (f.eks. tuegræs til løbebiller og blomster til svirrefluer), men der mangler sammenstillende undersøgelser der viser hvilen vegetation der optimerer antallet af nyttedyr samlet set. Der savnes ligeledes en gennemgribende belysning af randes landbrugsmæssige værdi, indbefattende hvilke dimensioner og evt. pasning de bør have. At der kun er udført få og begrænsede undersøgelser kan forstås, når man forsøger at opstille en plan med det nødvendige antal led og kombinationer under opretholdelse af blot rimelige krav til videnskabelige standarder. Det vil være omkostningskrævende, og det vil ikke være let at finde egnede landbrug nok til en god undersøgelse. Det største af alle problemer vil nok være at behovet for, og dermed værdien af den nyttedyrseffekt man får overfor skadegørere, vil variere betragteligt mellem lokaliteter og år. Det kræver større undersøgelser at overkomme en sådan variation og derved få belyst de overordnene effekter af nyttedyr taget over tid og afgrøder. At en sådan tilgang kan være frugtbar, indiceres af de tidligere demonstrationsforsøg med integreret produktion (IP) på Lautenbach Hof 1978-1990 (Steiner, El Titi & Bosch 1986) som bl.a. viste, at over en 6-års-periode med 40 % nedsættelse af pesticianvendelse (og med samme økonomiske resultat) steg forekomsterne af en række nyttedyr kraftigt i IP-arealerne, og i anden halvdel af perioden faldt forekomsterne af bladlus i disse arealer trods en ensartet start-indvandring i IP- og konventionelle arealer (El Titi 1986; Zeddies & Jung, 1986). Forskellen i bladlusforekomst kunne med betydelig sandsynlighed tilskrives nyttedyrvirkning; men et nagelfast bevis forelå ikke og ville have krævet en meget, meget større indsats. For randstriber og bufferzoner kan det blive endnu vanskeligere at føre noget, der ligner bevis, men det kan overvejes, om en knapt så nagelfast tilgang ville være acceptabel. Det kunne f.eks. være deleksperimenter til belysning af givne komponenters nytte i relation til bestemte skadedyr med modellering som sammenbindende element.
Ulemper ved bufferzoner og randstriber er et aspekt, der også kræver opmærksomhed ved drift. I de tidlige forsøg, hvor man havde 6 m bufferzone i korn langs hegn opstod høstproblemer pga. ukrudtsmængderne. I dag betragter de fleste randstriben, som et ikke-dyrket areal, og det har været fremført, at langs hegn er det en udmærket anvendelse af et areal, som ellers leverer begrænset udbytte (kapitel 6). I nyere engelske retninglinier er man gået et skridt videre (se kapitel 3 samt www.defra.uk). Her anbefales, at en 1 m bred barjordsstribe holdes helt ren mekanisk, for at forhindre problemskabende rodukrudt som f.eks agertidsel i at sprede sig gennem jorden. Tilsvarende tillades i danske støtteordninger at have 2 m barjordsstribe. Denne problematik er værd at inddrage i alle videre tiltag, da det kan være vigtigt at balancere rigtigt mellem skabelse af ekstra plantediversitet og undgåelse af arter, som er udpræget problematisk for landbrugsdriften.
For markstriber inde i marken i form af insektvolde, blomsterbælter eller kombinationer heraf er den driftsmæssige nytteproblematik meget lig den for randstiber. For disse typer markstriber angives typiske dimensioner, f. eks. 2-2,5 m bredde og 0,5 m højde i engelske anvisninger for insektvolde. At disse volde giver en potentiel nyttevirkning er dokumenteret i en række undersøgelser, men kun i et tilfælde er der påvist en klar udbytteeffekt (Collins 2002, tabel 2, bilag A). Samtidig melder sig et uafklaret spørgsmål, om hvor megen nytte man måske kunne få af meget simplere volde; f.eks. blot 20-30 cm i højden og 50 cm i bredden. Det mangler der ganske enkelt viden om. Et sådant spørgsmål ville også skulle udbygges med en landbrugsøkonomisk analyse på den ene side og en analyse af biodiversitets-gevinster på den anden side. At der sker noget positivt biodiversitetsmæssigt både for planter og dyr fremgår af de foreliggende undersøgelser (tabel 2 bilag A). Imidlertid savnes en systematisk eksperimentel tilgang, der kan muliggøre sammenhængende svar. I en mere systematisk undersøgelse vil det være indlysende at efterspørge betydning af mængden af insektvold og/eller blomsterbælte i forhold til markfladen – dvs som meter (og kvadratmeter) markstribe per hektar mark. Endelig vil det også være naturligt at afklare en eventuel betydning af mere eller mindre god sammenhæng (connectivity) mellem lineære strukturer indbyrdes, og mellem markstriber og randstriber. At sådanne undersøgelser ikke foreligger, afspejler nok komplikationsgrad og ikke mindst omkostningsniveau. At også markstriber medvirker til den øgning af landskabelig heterogenitet, som anses for meget vigtig (Tscharntke et al. 2005), kan der ikke være tvivl om. Men der foreligger væsentlige, delvist ubesvarede spørgsmål om gavnligheden af kombinationer herunder ultimativt om randstribeinvesteringer er bedre (mere biodiversitet pr. krone?) end markstribeinvesteringer -specielt for store marker –, og hvor størrelsesgrænsen i givet fald ligger?
Randzoner, primært til beskyttelse af vandløb, er en kategori for sig. De er krævet for at beskytte livet i vandløb mod pesticidpåvirkninger, men man kan spørge hvilken beskyttelse der rent faktisk er blevet opnået. For nogle pesticider er der krav om 2, 10 eller 20 m brede usprøjtede markrande mod vandforekomster, og der er heller ikke tvivl, om at nogle pesticider er meget risikable for ferskvandsvandflora og især fauna. Blandt de mest undersøgte pesticider er pyrethroiderne, der kan give destruktive effekter på ferskvandskrebsdyr og fisk i endog meget små koncentrationer (Nørum & Bjerregaard 2003). Det er blandt andet mod sådanne pesticider, de bredere randstriber mod vand skal beskytte, og der foreligger en række depositionsstudier, som er brugt som grundlag for de krævede bredder (se f.eks. Langhof, Gathman & Poehling 2005). Imidlertid synes der ikke at foreligge nogen systematisk opfølgning af de faktisk opnåede effekter af disse beskyttende randstriber. Der er heller ikke fundet væsentlige oplysninger relateret til beskyttende randstriber om forholdene for det biodiversitetsmæssigt interessante plante- og dyreliv langs vandløb. Kun konstaterer en enkelt undersøgelse (Ravn & Friberg 2004) at floraen langs vandløb i det danske agerland generelt ikke er særlig artsrig.
Tilskudsordninger omkring rand- og bufferzoner er der mange forskellige af med forskellige nationale tilsnit. I EU er der brugt 24 milliarder Euro fra 1992-2003 på landbrugsmiljøprogrammer ofte med et begrænset resultat på biodiversiteten fordi tiltagene sker på for små og isolerede arealer f.eks. på hegnsstumper a 50 m (Wittingham 2007). Der er ikke grund til betvivle, disse ordninger på et eller andet plan tager afsæt i resultater af undersøgelser, men der indgår efter alt at dømme også en række andre lokale forhold i udformning af de enkelte ordninger. Man må derfor formode at vurderinger og forhandlinger udgør en væsentlig del af baggrunden for de enkelte ordninger, og et af problemerne for denne udredning er, at der ikke findes en samlende nedskrivning af baggrunden for hver enkelt ordning. Eksempelvis vil baggrunden for de danske tilskud kræve en meget omfattende gennemgang af udvalgsdokumenter, som måske ikke en gang altid er fyldestgørende. Hertil kan føjes, at der i stort set alle tilfælde kun forefindes nogle upræcise, overordnede målsætninger for disse ordninger, mens mål, der kan bruges i opfølgende monitering af resultater savnes.
8 Konkluderende og sammendragende bemærkninger
Det er påvist at randzoner og bufferzoner (markrande, insektvolde, græsstriber og blomsterstriber) kan anvendes til at:
- Fremme antallet af vilde plantearter i den samlede markstruktur. – Dette er strengt taget selvindlysende, men det vides ikke særlig nøjagtigt, hvor mange arter man får ved en given type tiltag.
- Fremme mængden af fugle gennem adgang til redesteder og især føde. Der foreligger gode resultater vedrørende gulspurven, men en bredere belysning ville være hensigtsmæssig.
- Fremme vildt og småpattedyr. – Der foreligger solide resultater bl.a. for agerhøns og fasaner i forhold til 6 m-zoner, men hvilke bredder og vegetationsforhold der er tilstrækkelige eller optimale vides ikke.
- Fremme antallet af nyttedyr inde i randstrukturen, bl.a. i kraft af forbedret overvintring. – Gode enkeltresultater foreligger, dog mangler en systematisk undersøgelse af dimensioners indflydelse, idet det ikke vides ”hvor lidt man kan nøjes med og alligevel få en væsentlig forbedring”.
- Reducere trykket af visse skadedyr i en vis afstand fra randstrukturen, aftagende med afstanden. Dog mangler mere eksakte undersøgelser med inddragelse af flere arter af skadedyr.
- Fremme artsantallet – dog primært af forholdsvis almindelige arter (påvist for en række ukrudtsarter og arthropoder). Samlet viden om biodiversitetseffekt (betydeligt antal arter) mangler.
- Fremme sammenhæng (connectivitet) i landskabet; ordentlige effektmålinger og nogle eksperimentelle dokumentationer mangler.
Det er påvist at biodiversiteten i randzoner og bufferzoner afhænger af:
- Plantevalget og sammensætningen ved udsåning. – Dette er nærmest selvindlysende; men der mangler viden om skæringsfladen mellem det ideelt ønskelige og det driftsmæssigt rimelige.
- Randzonens alder. Der mangler dog en systematisk opstilling af mål og etablering af eksperimentelt baseret viden, om hvilken plejebetinget successionstilstand, der giver hvilken biodiversitet.
- Randzonens areal. Som nævnt ovenfor mangler en systematisk undersøgelser af randzoners bredde; men der foreligger stort set ingen viden om betydningen af en given randzones samlede areal – nogle arter kan sikkert opnås på små arealer men arter, som f.eks. fugle må formodes at have behov for noget større areal, hvis randen er hovedleverandør af føde.
Det er påvist, at forekomsten af nyttedyr i randzoner og bufferzoner afhænger af:
- Habitatkvalitet, bl.a. i forhold til overvintring. Engelske undersøgelser har påvist værdien af tuedannende græsser; men erfaringer fra indsamlinger af forsøgsdyr tilsiger en stor værdi af blot en minimal jordvold/kant med græsser og anden flerårig vegetation og mindre stenbunker (som kan placeres inde mod foden af et hegn) – der mangler detaileksperimenter.
- Adgang til næring i form af pollen, nektar og alternativt bytte – som helt eller delvis afhænger af vegetationen og dens tilstand. Der er behov for afklaring af den optimale vegetationssammensætning, som gavner flest arter af nyttedyr samtidigt.
Det er påvist, at funktionel biologisk bekæmpelse afhænger af:
- Afstanden fra markranden, f.eks. relateret til indvandring af visse generalist prædatorer og visse snyltehvepse, men der foreligger ikke eksperimentel viden, så man kan sige, hvilken afstand der opfylder betingelserne for flest arter. Det vides heller ikke i hvilken grad negative effekter af store marker kan ”opblødes” ved placering af insektvolde, blomsterbælter eller andre hjælpestrukturer – evt. ved behersket ændring af plejespor.
- Strukturens alder, hvad enten der er tale om randzone, insektvold eller andet. Der foreligger begrænset og ikke sammenhængende viden om hvilket successionstrin efter etablering af randzone eller markstribe inde i mark, man med fordel kan standse ved vha. pleje (slåning, slet, strigling, stubharvning etc.).
Det er påvist, at rand- og bufferzoner:
- Reducerer pesticidafdrift afhængigt af bredde og højde, men der foreligger ikke systematisk, opfølgende viden om etablerede bufferzoners faktiske virkning på dels flora og fauna i vandløb og dels den terrestrisk fauna i selve bufferzonen og tilstødende natur. Der mangler svar på differentierede spørgsmål relateret til bufferzonens karakter.
- Reducerer næringsstofudvaskning til ferskvandsforekomster afhængig af bufferzonens bredde og plantesammensætning. Dog synes der at mangler mere præcis viden som kan omsættes i anbefalinger, der bør sikre både den ønskede beskyttelse, men også en forbedring af biodiversitet mere generelt.
- Bidrager til øget plantediversitet. Dette er indlysende, men der synes mange steder langs vandløb at være en relativt artsfattig vegetation, og der mangler viden om hvordan man bedst opnår en så rig vegetation som muligt, hvilket som sidegevinst må forventes at give en rigere insektfauna og dermed øget mængde og kvalitet af fugleføde.
Det er påvist at landskabsheterogenitet har en række effekter af særlig interesse i stærkt udnyttede landområder:
- Øget landskabsheterogenitet giver øget biodiversitet, men der mangler viden, om hvilket niveau af heterogenitet (strukturer, småhabitater per areal) der har hvilken effekt.
- Heterogenitet har effekter på organismer i både tidsmæssigt og rumlig skala. Der mangler viden om effekter på arter og på genetisk diversitet.
- Afhængig af spredningsevne og biologi påvirkes organismer af landskabsheterogenitet på et eller flere rumlige niveauer. Nogle resultater tyder på at der er effekter af det intensive landbrug som rækker væsentligt ud over marker og deres naboarealer, men der foreligger ikke konkret dansk viden.
- Øget landskabsheterogenitet kan øge sammenhængen (connectiviteten) gennem korridoreffekter, men der savnes eksperimentel evidens for ønsket spredning gennem etablering af korridorer (der samtidig tjener andre formål knyttet til heterogenitet).
I relation til etablering af diverse forordninger om indlagte ændringer i dyrkede områder mangler en række oplysninger samt viden om samfundsmæssige forhold.
- Der er en helt generel mangel på samlet nedskrivning af argumenterne for og de præcise formål med og forventninger til de enkelte ordninger, hvorom der gives påbud eller hvortil der gives støtte. Det ville være formålstjeneligt at dette blev gennemført på EU-niveau, således at man som minimum kunne læse om formål, naturvidenskabelige grundlag og helst også om samfundsmæssige og økonomiske betragtninger.
- Der mangler sociologisk viden om mulige samfundsmæssige gevinster af støttede tiltag i dyrkede arealer: I hvilken grad kommer sådanne tiltag den øvrige befolkning til gode og i hvilken grad erkendes og påskønnes forøgelse af biologiske værdier?
- Der mangler en undersøgelse af hvilke mulige tiltag for biodiversitet og nytte, der bedst kan forenes med en rationel landbrugsdrift.
- Det bør overvejes først at koncentrere indsats om karakterisering af bestemte biologiske resultater, biologisk værdi i form af biodiversitet, for siden at muliggøre anden værdisætning.
- Der kan for nærværende ikke gennemføres en lødig sammenligning af forskellige bufferzone-ordninger på tværs af lande, og der foreligger heller ikke gode opfølgninger mht opnåede resultater indenlands.
9 Forslag til forskningsindsats
Bufferzoner og randzoner
De eksisterende resultater viser, at sprøjtefrie og braklagte randzoner kan bidrage til at standse nedgangen i og sandsynligvis øge biodiversiteten i agerlandet, samtidig med at randzoner langs følsomme biotoper som fx vandløb og hegn kan være med til at beskytte disse habitater. Der er dog nogle metodiske problemer, som sætter begrænsninger for de eksisterende resultaters tolkning og anvendelse. Således stammer de fra undersøgelser af en meget blandet natur mht. formål og metode, og det er derfor ikke muligt at udmelde, hvilken forbedring af biodiversiteten man får ved en given indsats (f.eks. at så meget 6m bred rand med en given bevoksning og pleje pr. så og så mange hektar giver .....). I en dansk kontekst giver det anledning til at foreslå forskning, der kan afklare effekten på kort og langt sigt af forskellige typer randzoner og deres placering i landskabet mht. generel biodiversitet og beskyttelse af marknær natur samt hvilke tiltag, der indenfor et rimelig kort sigt giver en definerbar biodiversitetsforbedring, et bestemt niveau af beskyttelse af f.eks. vandløb, og efterfølgende hvilke konsekvenser, disse tiltag måtte medføre for jordbrugsdriften. Det vil være ønskeligt at inddrage en bred vifte af forskellige håndteringer af randzoner mht. bredde, areal, placering, varighed, vegetation og pleje. Som udgangspunkt kan det dog være hensigtsmæssigt at tage mest muligt afsæt i eksisterende viden og forordninger med en vis fokusering.
I undersøgelser af biodiversitetsforbedring vil det være nærliggende også at inddrage opstilling af indikatorarter for både planter, leddyr og fugle; nogle kan forudsiges, mens andre ville kunne ekstraheres af de opnåede biodiversitetsresultater. Indikatorarter skal først og fremmest relativt enkelt og sikkert kunne indicere en tilstand og eventuelle ændringer, således at der kan iværksættes opfølgning af givne tiltag ved hjælp af et mindre omfattende arbejde.
Med disse overvejelser som baggrund kan nedenstående forskningsområder foreslås:
1) Biodiversitetsforbedringer (blandt et udvalg af planter, leddyr og hvirveldyr) på kort og langt sigt ved placering af forskellige typer, bredder og længder af beskyttende randzoner langs eksisterende linjehabitater eller småbiotoper så som hegn, skel og grøfter, og vandløb.
2) Biodiversitetsændringer (planter, leddyr og hvirveldyr) af forskellige typer (f.eks. dimensioner og vegetation) bufferlinier inde i marker samt udbyttemæssige effekter af disse bufferlinier.
3) Sammenligning af 1) og 2) med henblik på målretning af en eventuel indsats for øgning af biodiversiteten vha. randzoner.
4) Undersøgelse af kort- og langsigtede biodiversitetsvirkninger af dels øget sammenhæng (connectivitet) opnået ved sammenkobling af randzoner og bufferlinier i marker over større områder, og dels øget heterogenitet i et ellers mere homogent og mindre diverst marklandskab.
5) Sociologisk og økonomisk undersøgelse af hhv. befolkningens og jordbrugernes opfattelse af tiltag for og værdi af biodiversitet, samt af jordbrugserhvervets adfærd og de driftsmæssige konsekvenser for dette erhverv.
6) Undersøgelse af hvilke virkemidler, der bedst og mest økonomisk kan fremme biodiversitet.
For alle de nævnte forslag bør gælde, at der tilstræbes en indbygget udvikling af metoder, der fra starten kan medvirke til at opstille mål for ønsket biodiversitet og efterfølgende kan muliggøre kontrol af den faktiske indsats og ”måling” af, om de ønskede virkninger er opnået. Endvidere bør det tilstræbes at inddrage resultater fra tidligere undersøgelser, hvor dette er relevant.
10 Referencer
Abildtrup, J. (1999) Økonomiske beregninger af kompensation ved beskyttelseszoner langs målsatte vandløb og søer. Statens Jordbrugs- og Fiskeriøkonomiske Institut. Udredning for Miljøstyrelsen. 11 pp.
Abildtrup, J. (2004) Indkomsttab ved miljøgræs, vådområder og ekstensive randzoner pp. 1-26. Fødevareøkonomisk Institut, Afdeling for Jordbrugets Driftsøkonomi, København.
Agger, A. (1986) Udviklingen i agerlandets småbiotoper i Danmark. Institut for Geografi, Samfundsanalyse og Datalogi, Roskilde Universitetscenter.
Alderweireldt, M. (1994) Habitat manipulation increasing spider densities in agroecosystems: possibilities for biological control. Journal of Applied Entomology 118, 10–16.
Aquilino, K.M., Cardinale, B.J. & Ives, A.R. (2005) reciprocal effects of host plant and natural enemy diversity on herbivore suppression: an empirical study of a model tritrophic system. Oikos 108, 275-282.
Albrecht, H. & Mattheis, A. (1998) The effects of organic and integrated farming on rare arable weeds on the Forschungsverbund Agrarokosysteme Munchen (FAM) research station in southern Bavaria. Biological Conservation 86, 347-356.
Altieri, M.A. & W.H. Whitcomb (1979) Manipulation of insect populations through seasonal disturbance of weed communities. Protection Ecology 1, 185–202.
Andreasen, C. & Stryhn, H. (unpubl.) Increasing weed flora in Danish arable fields and its importance for the biodiversity. Submitted to Weed Research.
Andreasen, C., Stryhn, H. & Streibig, J.C. (1996) Decline of the flora in Danish arable fields. Journal of Applied Ecology 33, 619-626.
Anon. (2006) Frederiksborg Amt. www.frederiksborgamt.dk.
Asteraki, E.J., Hart, B.J., Ings, T.C. & Manley, W.J. (2004) Factors influencing the plant and invertebrate diversity of arable field margins. Agriculture, Ecosystems and Environment 102, 219-231.
Aude, E., Tybirk, K. & Bruus Pedersen, M. (2003) Vegetation diversity of conventional and organic hedgerows in Denmark. Agriculture Ecosystems & Environment 99, 135-147.
Aude, E., Tybirk, K., Michelsen, A., Ejrnæs, R., Hald, A.B. & Mark, S. (2004) Conservation value of the herbaceous vegetation in hedgerows - does organic farming make a difference? Biological Conservation 118, 467-478.
Baillie, S.R., Sutherland, W.J., Freeman, S.N., Gregory, R.D. & Paradis, E. (2000) Journal of Applied Ecology 37, 88-102.
Bayram, A. & Luff, M.L. (1993) Winter abundance and diversity of lycosids (Lycosidae, Araneae) and other spiders in grass tussocks in a field margin. Pedobiologia 37, 357–364.
Bence, S., Stander, K. & Griffiths, M. (1999) Nest site selection by the harvest mouse (Micromys minutus) on arable farmland. Aspects of Applied Biology 54, 197-202.
Bence, S.L., Stander, K. & Griffiths, M. (2003) Habitat characteristics of harvest mouse nests on arable farmland. Agriculture Ecosystems & Environment 99, 179-186.
Benton, T.G., Bryant, D.M., Cole, L. & Crick, H. Q. P. (2002) Linking agricultural practice to insect and bird populations: a historical study over three decades. Journal of Applied Ecology 39, 673-687.
Benton, T.G., Vickery, J.A. & Wilson, J.D. (2003) Farmland biodiversity - is habitat heterogeneity the key? Trends in Ecological Evolution 18, 182-188.
Bianchi, F.J.J.A. & Van Der Werf, W. (2003) The effect of the area and configuration of hibernation sites on the control of aphids by Coccinella septempunctata (Coleoptera : Coccinellidae) in agricultural landscapes: a simulation study. Environmental entomology 32, 1290-1304.
Biedermann, R. (2000) Metapopulation Dynamics of the Froghopper Neophilaenus albipennis (F., 1798) (Homoptera, Cercopidae). What is the Minimum Viable Metapopulation Size? Journal of Insect Conservation 4, 99-107.
BMVEL (2005) Bundesministerium für Verbraucherschutz, Ernährung und Landwirtschaft (BMVEL) 11055 Berlin.
Bohac, J. (1999) Staphylinid beetles as bioindicators. Agriculture, Ecosystems and Environment 74, 357-372.
Boatman, N.D. (1992) Herbicides and the management of field boundary vegetation. Pesticide Outlook 3, 30-34.
Boatman, N.D. & Wilson, P.J. (1988) Field edge management for game and wildlife conservation. Aspects of Applied Biology 16, 53-61.
Boatman, N.D. & Bence, S.L. (2000) Management of set-aside to enhance biodiversity: the wild bird cover option. Aspects of Applied Biology 62, 73-79.
Boatman,N.D. & Stoate, C. (2000) Integrating biodiversity conservation into arable agriculture. Aspects of Applied Biology 62, 21-30.
Boller, E., Häni, F. & Poehling, H.-M. (2004) Ecological infrastructures. Ideabook on functional biodiversity at the farm level, 1 edn.
Bradbury, R.B., Kyrkos, A., Morris, A. J., Clark, S. C., Perkins, A. J. & Wilson, J. D. (2000) Habitat associations and breeding success of yellowhammers on lowland farmland. Journal of Applied Ecology 37, 789-805.
Brandt, J. (2004) Anvendelse af topografiske kort til monitering af biotoper. Geoforum Perspektiv 5, 53-61.
Brandt, J.J.E., Bunce, R.G.H., Howard, D.C. & Petit, S. (2002) General principles of monitoring land cover change based on two case studies in Britain and Denmark. Landscape and Urban Planning 62, 37-51.
Brickle, N.W., Harper, D.G.C., Aebischer, N.J. & Cockayne, S.H. (2000) Effects of agricultural intensification on the breeding success of corn buntings Miliaria calandra. Journal of Applied Ecology 37, 742-755.
Brundtlandkommissionen (1987) Vor fælles fremtid. FN-forbundet og Mellemfolkeligt Samvirke, København.
Bruus Pedersen, M.B., Aude, E. & Tybirk, K. (2004) Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetation pp. 1-103. Danmarks Miljøundersøgelser. Bekæmpelsesmiddelforskning for Miljøstyrelsen.
Bruus Pedersen, M., Andersen, H.V.G.M., Løfstrøm, P., Jensen, B., Strandberg, M., Bak, J. & Kjær, C. (2006) Vurdering af omfang og effekt af afdrift af ukrudtsmidler til danske læhegn eksemplificeret ved metsulfuron. Årsrapport 2006,
Miljøministeriet, Miljøstyrelsen.
Bullock, J.M., Pywell, R.F., Burke, M.J.W. & Walker, K.J. (2001) Restoration of biodiversity enhances agricultural production. Ecology Letters 4, 185-189.
Burel, F., Butet, A., Delettre, Y.R. & de la Pena, N.M. (2004) Differential response of selected taxa to landscape context and agricultural intensification. Landscape and Urban Planning 67, 195-204.
Burel, F. & Baudry, J. (2005) Habitat quality and connectivity in agricultural landscapes: The role of land use systems at various scales in time. Ecological Indicators 5, 305-313.
Burn A.J. (1992) Interactions between cereal pests and their predators and parasites. In Greig-Smith P., Frampton G. & Hardy T. (eds): Pesticides, Cereal Farming and the Environment. The Boxworth Project. HMSO, London, pp. 110–131.
Cardinale, B.J., Harvey, C.T., Gross, K. & Ives, A.R. (2003) Biodiversity and biocontrol: emergent impacts of a multi-enemy assemblage on pest suppression and crop yield in an agroecosystem. Ecology Letters 6, 857-865.
Carvell, C., Meek, W.R., Pywell, R.F. & Nowakowski, M. (2004) The response of foraging bumblebees to successional change in newly created arable field margins.
Biological Conservation 118, 327-339.
Chapple, D.G., Wade, D.R., Laverick, R.M. & Eldridge, P.J. (2002) Whole farm integrated management and farmland birds. Aspects of Applied Biology 67, 129-135.
Chase, J.M. & Leibold, M.A. (2002) Spatial scale dictates the productivity-biodiversity relationship. Nature 416, 427-430.
Chiverton, P.A. (1989) The creation of within-field overwintering sites for natural enemies of cereal aphids. Brighton Crop Protection Conference – Weeds, 1093-1096.
Chiverton, P.A. (1999) Buffer zones of different floral composition - effects on the beneficial arthropod fauna. Aspects of Applied Biology 54 307-314.
Clough, Y., Kruess, A., Kleijn, D. & Tscharntke, T. (2005) Spider diversity in cereal fields: comparing factors at local, landscape and regional scales. Journal of Biogeography 32, 2007-2014.
Collins, K.L., Wilcox, A., Chaney, K., Boatman, N.D. & Holland, J.M. (1997) The influence of beetle banks on aphid population predation in winter wheat. Aspects of Applied Biology 50, 341-346.
Collins, K.L., Boatman, N.D., Wilcox, A., Holland, J.M. & Chaney, K. (2002) Influence of beetle banks on cereal, aphid predation in winter wheat. Agriculture Ecosystems & Environment 93, 337-350.
Collins, K.L., Boatman, N.D., Wilcox, A. & Holland, J.M. (2003a) A 5-year comparison of overwintering polyphagous predator densities within a beetle bank and two conventional hedgebanks. Annals of Applied Biology 143, 63-71.
Collins, K.L., Boatman, N.D., Wilcox, A. & Holland, J.M. (2003b) Effects of different grass treatments used to create overwintering habitat for predatory arthropods on arable farmland. Agriculture Ecosystems & Environment 96, 59-67.
Coombes, S.D. and Sotherton, N.W. (1986) The dispersal and distribution of polyphagous predatory Coleoptera in cereals. Annals of Applied Biology 108, 461–474.
Cowgill, S.E., Wratten, S.D. & Sotherton, N.W. (1993) The selective use of floral resources by the hoverfly Episyrphus balteatus (Diptera: Syrphidae) on farmland. Annals of Applied Biology 122, 223-231.
Cuthbertson, P.S. & Jepson, P.C. (1988) Reducing pesticide drift into the hedgerow by the inclusion of an unsprayed field margin pp. 747-751. Thornton Heath, UK: British Crop Protection Council.
Daily, G.C., Sùderquist, T., Aniyar, S., Arrow, K., Dasgupta, P., Ehrlich, P.R., Folke, C., Jansson, A., Jansson, B.-O., Kautsky, N., Levin, S., Lubchenko, J., Mäler, K.-G., Simpson, D., Starrett, D., Tilman, D. & Walker, B. (2000) The value of nature and the nature of value. Science 289, 395-396.
Danmarks Statistik (2006) Landbrugsfamiliernes husstandsindkomst 2004. Nyt fra Danmarks Statistik.
Danmarks Statistik (2007) http://www.statistikbanken.dk/statbank.
Dansk Landbrugsrådgivning (2004) Vejledning i planteværn 2004. Danmarks JordbrugsForskning og Landskontoret for Planteavl., Skejby.
Dennis,P., Thomas, M.B. & Sotherton, N.W. (1994) Structural Features of Field Boundaries Which Influence the Overwintering Densities of Beneficial Arthropod Predators. Journal of Applied Ecology 31, 361-370.
Det Økonomiske råd (2004). Dansk økonomi efterår 2004. http://www.dors.dk/sw1149.asp.
De Barro, P.J. (1992) The impact of spiders and high temperatures on cereal aphid (Rhopalosiphum padi) numbers in an irrigated perennial grass pasture in South Australia. Annals of Applied Biology 121, 19–26.
de-Cauwer,B., Reheul, D., Nijs, I. & Milbau, A. (2005) Biodiversity and agro-ecology in field margins. Communications in Agricultural and Applied Biological Sciences 70, 17-49.
de Snoo, G.R. & de Wit, P.J. (1998) Buffer zones for reducing pesticide drift to ditches and risks to aquatic organisms. Ecotoxicology and Environmental Safety 41, 112-118.
de Snoo, G.R., van der Poll, R. J. & Bertels, J. (1998) Butterflies in sprayed and unsprayed field margins. Journal of Applied Entomology-Zeitschrift fur Angewandte Entomologie 122, 157-161.
de Snoo, G.R. (1999) Unsprayed field margins: effects on environment, biodiversity and agricultural practice. Landscape and Urban Planning 46, 151-160.
de Groot, R.S. (1992) Functions of nature. Evaluation of nature in
environment, planning management and decision making, Amsterdam.
Dennis, P., Thomas, M.B. & Sotherton, N.W. (1994) Structural Features of Field Boundaries Which Influence the Overwintering Densities of Beneficial Arthropod Predators. Journal of Applied Ecology 31, 361-370.
Denys, C. & Tscharntke, T. (2002) Plant-insect communities and predator-prey ratios in field margin strips, adjacent crop fields, and fallows. Oecologia 130, 315-324.
Dobson, A. (2005) Monitoring global rates of biodiversity change: challenges that arise in meeting the Convention on Biological Diversity (CBD) 2010 goals. Philosophical Transactions of the Royal Society B-Biological Sciences 360, 229-241.
Donald, P.F., Buckingham, D.L., Moorcroft, D., Muirhead, L.B., Evans, A.D. & Kirby, W.B. (2001) Habitat use and diet of skylarks Alauda arvensis wintering on lowland farmland in southern Britain. Journal of Applied Ecology 38, 536-547.
Dowdeswell, W.H. (1987) Hedgerows and verges. Allen and Unwin, London.
Duelli, P., Studer, M., Marchand, I. & Jakob, S (1990). Population movements of arthropods between natural and cultivated areas. Biological Conservation 54, 193-207.
Duelli, P. & Obrist, M. K. (2003) Regional biodiversity in an agricultural landscape: the contribution of seminatural habitat islands. Basic and Applied Ecology 4, 129-138.
Edwards, C.A., Sunderland, K.D. & George, K.S. (1979) Studies on Polyphagous Predators of Cereal Aphids. Journal of Applied Ecology 16, 811-823.
Edwards, P.J. & Abivardi, C. (1998) The value of biodiversity: Where ecology and economy blend. Biological Conservation 83, 239-246.
Edwards, C. A. & Lofty, J.R. (1979) The effects of straw residues and their disposal on the soil fauna In: E. Grossbard (ed), Straw Decay and its Effect on Disposal and Utilisation. John Wiley & Sons, Chichester, pp. 37–44.
Eggenschwiler, L., Jacot, K., Studer, S. & Edwards, P. (2004) Influence of seed mixtures on the development of the seed bank of wildflower strips and rotational fallows and the persistence of species in the subsequent crop. Botanica Helvetica 114, 49-66.
Eilenberg, J., Hajek, A. & Lomer, C. (2001) Suggestions for unifying the terminology in biological control. Biocontrol 46, 387-400.
Ekbom, B.S., Wiktelius, S. & Chiverton, P.A. (1992) Can Polyphagous Predators Control the Bird Cherry-Oat Aphid (Rhopalosiphum padi) in Spring Cereals - A Simulation Study. Entomologia Experimentalis et Applicata 65, 215-223.
El Titi, A. (1986) Managenent of cereal pests and diseases in integrated farming systems. British Crop Protection Conference – Pests and Diseases, 147-155.
Esbjerg, P. & Petersen, S. (eds) (2002) Effects of reduced pesticide use on flora and fauna in agricultural fields. Pesticides Research 58. Danish Environmental Protection Agency.
European Commision. (2005) European Commission, Directorate General for Agriculture and Rural Development, Unit G-4 - Evaluation of Measures applied to Agriculture, Studies.
Eurostat (2003) LUCAS Technical documents 1-9. Tilgængelige på: http://forum.europa.eu.int/Public/irc/dsis/landstat/library?l=/lucas/reference_documentation&vm=detailed&sb=Title ).
Everts, J.W., Aukema, B., Hengeveld, R. & Koeman, J.H. (1989) Side-effects of pesticides on ground-dwelling predatory arthropods in arable ecosystems. Environmental Pollution 59, 203-225.
Feber, R.E., Smith, H. & MacDonald, D.W. (1994) The effects of field margin restoration on the meadow brown butterfly (Maniola jurtina). Farnham, UK: British Crop Protection Council Registered Office (BCPC).
Feber, R.E., Bell, P., Johnson, J., Smith, H., Baines, M. & Macdonald, D.W. (1995) The effects of arable field margin management on the abundance of beneficial arthropods. BCPC Symposium Proceedings 63, 163–170.
Feber, R.E. & Hopkins, A. (1997) Diversity of plant and butterfly species on organic farmland field margins in relation to management. British Grassland Society Fifth Research Conference, University of Plymouth, Newton Abbot, Devon, UK, 8 10 September 1997 63-64, -644.
Feber, R.E., Smith, H. & MacDonald, D.W. (1999) The importance of spatially variable field margin management for two butterfly species. Aspects of Applied Biology 54, 155-162.
Feehan, J., Gillmor, D.A. & Culleton, N. (2005) Effects of an agri-environment scheme on farmland biodiversity in Ireland. Agriculture, Ecosystems and Environment 107, 275-286.
Firbank, L. & Smart, S. (2002) The changing status of arable plants that are important food items for farmland birds. Aspects of Applied Biology 67, 165-170.
Fischer, M. & Matthies, D. (1997) Mating structure and inbreeding and outbreeding depression in the rare plant Gentianella germanica (Gentianaceae). American Journal of Botany 84, 1685-1692.
Fitzgibbon, C.D. (1997) Small mammals in farm woodlands: the effects of habitat, isolation and surrounding land-use patterns. Journal of Applied Ecology 34, 530-539.
Forenede Nationer (1992) Rio declaration on environment and development. Report of the united nations conference on environment and development. Rio de Janeiro, 3-14 June 1992.
Fog, K. (1997) Nordens padder og krybdyr. G.E.C. Gads forlag.
Forman, R.T.T. & Godron, M. (1986) Landscape ecology. Wiley NewYork.
Fox, A.D. (2004) Has Danish agriculture maintained farmland bird populations? Journal of Applied Ecology 41, 427-439.
Frampton, G.K. (2002) Long-term impacts of an organophosphate-based regime of pesticides on field and field-edge Collembola communities. Pest Management Science 58, 991-1001.
Frank, T. (1997) Species diversity of ground beetles (Carabidae) in sown weed strips and adjacent fields. Biological Agriculture and Horticulture 15, 297-307.
Frank, T. & Nentwig, W. (1995) Ground-Dwelling Spiders (Araneae) in Sown Weed Strips and Adjacent Fields. Acta Oecologica-International Journal of Ecology 16, 179-193.
Fry, G.L.A. & Robson, W. J. (1994) Field margins: integrating agriculture and conservation (ed. N.Boatman), pp. 111-116. British crop protection council, Croyton, UK.
Fuller, R.J., Gregory, R. D., Gibbons, D.W., Marchant, J.H., Wilson, J.D., Baillie, S.R. & Carter, N. (1995) Population declines and range contractions among lowland farmland birds in Britain. Conservation Biology 9, 1425-1441.
Fuller, R.J., Chamberlain, D.E., Burton, N.H.K. & Gough, S.J. (2001) Distributions of birds in lowland agricultural landscapes of England and Wales: How distinctive are bird communities of hedgerows and woodland? Agriculture Ecosystems & Environment 84, 79-92.
Fussell, M. & Corbet, S.A. (1991) Forage for bumble bees and honey bees in farmland: a case study. Journal of Apicultural Research 30, 87-97.
Garbutt, R.A. & Sparks, T.H. (2002) Changes in the botanical diversity of a species rich ancient hedgerow between two surveys (1971-1998). Biological Conservation 106, 273-278.
Gerowitt, B., Isselstein, J. & Marggraf, R. (2003) Rewards for ecological goods - requirements and perspectives for agricultural land use. Agriculture Ecosystems & Environment 98, 541-547.
Goverde, M., Schweizer, K., Baur, B. & Erhardt, A. (2002) Small-scale habitat fragmentation effects on pollinator behaviour: experimental evidence from the bumblebee Bombus veteranus on calcareous grasslands. Biological Conservation 104, 293-299.
Greaves, M.P. & Marshall, E.-J. P. (1987) Field margins: definitions and statistics. Monograph, British Crop Protection Council 3-10.
Green, R.E., Osborne, P.E. & Sears, E.J. (1994) The Distribution of Passerine Birds in Hedgerows During the Breeding-Season in Relation to Characteristics of the Hedgerow and Adjacent Farmland. Journal of Applied Ecology 31, 677-692.
Greenstone, M.H., Morgan, C.E., Hultsch, A.L., Farrow, R.A., & Dowse, J.E. (1987) Ballooning spiders in Missouri, USA, and New South Wales, Australia: family and mass distributions. Journal of Arachnology 15, 163–170.
Grub, A., Perritaz, J. & Contat, F. (1996) Promotion of the segetal flora by field margins on productive arable soil. Journal of Applied Botany-Angewandte Botanik 70, 101-112.
Gunderson, L.H. (2000) Ecological Resilience - In Theory and Application. Annual Review of Ecology and Systematics 31, 425-439.
Hald, A.B. (1999) Weed vegetation (wild flora) of a long established organic versus conventional cereal fields. Annals of Applied Biology 134, 307-314.
Hald, A.B., Nielsen, B.O., Samsø-Petersen, L., Hansen, K., Elmegaard, N. & Kjølholt, J. (1988) Sprøjtefri randzoner i kornmarker. Miljøministeriet, Miljøstyrelsen.
Hald, A.B. & Reddersen, J. (1990) Fugleføde i kornmarker- insekter og vilde planter. Miljøprojekt nr. 125. Miljøstyrelsen, Miljøministeriet.
Hald, A.B. & Lund, T. (1994) Firesprøjtefri driftsformer af markers randzoner. Konsekvenser for vilde planter, insekter og økonomi. Faglig rapport fra DMU, nr 103. Miljø- og Energiministeriet.
Hald, A.B., Redderesen, J. & Elbek-Pedersen, H. (1994) Sprøjtefri randzoner i sædskiftemarker, Plante- og insektliv samt udbytter: Landsforsøg 1987-92. Bekæmpelsesmiddelforskning fra Miljøstyrelsen 6. Miljøstyrelsen.
Hart, J.D., Milsom, T.P., Fisher, G., Wilkins, V., Moreby, S.J., Murray, A.W.A. & Robertson, P.A. (2006) The relationship between yellowhammer breeding performance, arthropod abundance and insecticide applications on arable farmland. Journal of Applied Ecology 43, 81-91.
Harwood, R.W. J., Wratten, S. D., Nowakowski, M. & Marshall, E. P. J. (1994) Wild flower strips and winter/summer populations of beneficial invertebrates in farmland. IOBC/WPRS Bulletin 17 211–219.
Halley, J.M., Thomas, C.F.G. & Jepson, P.C. (1996) A model for the spatial dynamics of linyphiid spiders in farmland. Journal of Applied Ecology 33, 471–492.
Hancock, M.H. & Wilson, J.D. (2003) Winter habitat associations of seed-eating passerines on Scottish farmland. Bird Study 50, 116-130.
Hartmann, E., Schekahn, A., Luick, R. & Thomas, F. (2006) Kurzfassungen der Agrarumwelt- und Naturschutzprogramme. Darstellung und Analyse von Maßnahmen der Agrarumwelt- und Naturschutzprogramme in der Bundesrepublik DeutschlandBundesamt für Naturschutz, Bonn. Link: http://www.bfn.de/fileadmin/MDB/documents/skript161.pdf
Hausammann, A. (1996) The effects of sown weed strips on pests and beneficials arthropods in winter wheat fields. Bulletin OILB/SROP 19, 106-109.
Heikkilä, M. (2001) Maatalouden ympäristùtuki ja luonnon monimuotoisuus (Environmental support for agriculture and biodiversity) pp. 1-41. Birdlife Suomi-Finland.
Heimbach, U. & Garbe, V. (1996) Effects of reduced tillage systemsin sugar beet on predatory and pest arthropods. Acta Jutlandica 71, 195–208.
Heimpel,G.E., Rosenheim, J.A. & Kattari, D. (1997) Adult feeding and lifetime reproductive success in the parasitoid Aphytis melinus. Entomologia Experimentalis et Applicata 83, 305-315.
Heitzmann, A., Lys, J.-A. & Nentwig, W. (1992) Nützlingsfùrderung am Rand - oder: Von Sinn des Unkrautes. Landwirtschaft Schweiz 5, 25-36.
Heldberg, H. (2005) De almindelige fugles bestandsudvikling i Danmark 1975-2004. Dansk Ornitologisk Forenings Tidsskrift 99, 182-195.
Hickman, J.M. & Wratten, S.D. (1996) Use of Phacelia tanacetifolia strips to enhance biological control of aphids by hoverfly larvae in cereal fields. Journal of economic entomology 89, 832-840.
Hinsley, S.A. & Bellamy, P.E. (2000) The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review. Journal of Environmental Management 60, 33-49.
Holbeck, H.B., Clausen, H.D. & Reddersen, J. (2000) Selection of nectar sources by butterflies and burnets in organic field boundary habitats (Papilionoidea, Hesperioidea, and Zygaenidae). Nordic Journal of Botany 20, 47-60.
Holland, J.M. & Thomas, S.R. (1997) Quantifying the impact of polyphagous invertebrate predators in controlling cereal aphids and in preventing wheat yield and quality reductions. Annals of Applied Biology 131, 375-397.
Holland, J.M., Thomas, C.-F. G., Birkett, T., Southway, S. & Oaten, H. (2005) Farm-scale spatiotemporal dynamics of predatory beetles in arable crops. Journal of Applied Ecology 42, 1140-1152.
Holland, J.M., Southway, S., Ewald, J. A., Birkett, T., Megbie, M., Hart, J., Parrott, D. & Allcock, J. (2006) Invertebrate chick food for farmland birds: spatial and temporal variation in different crops. Aspects of Applied Biology 67, 27-34.
Holling, C. S. (1973) Resilience and Stability of Ecological Systems. Annual Review of Ecological Systems 4, 1–23.
Hopkins, A. & Feber, R. E. (1997) Management for plant and butterfly species diversity on organically farmed grassland field margins Management for grassland biodiversity. Proceedings of the International Occasional Symposium of the European Grassland Federation, Warszawa omza, Poland, 19 23 May, 1997 pp. 69-73. Poznan, Poland: Organizing Committee of the International Occasional Symposium of the European Grassland Federation.
Hovd, H. & Skogen, A. (2005) Plant species in arable field margins and road verges of central Norway. Agriculture Ecosystems & Environment 110, 257-265.
Huusela-Veistola, E. (1998) Effects of perennial grass strips on spiders (Araneae) in cereal fields and impact on pesticide side-effects. Journal of Applied Entomology 122, 575-583.
Jacobsen, B.H. (2006) Randzoner langs vandløb og søer -potentiale, holdninger og barrierer. Fødevareøkonomisk Institut.
Jacobsen, L-B., Andersen, M., Jensen, D.J. (2004) Reducing the use of pesticides in Danish agriculture – macro and sector economic analyses. Baggrundsnotat, Vismandsrapporten Efterår 2004. Det Økonomiske Råds Sekretariat. www.dors.dk
Jacobsen, L.-B., Jensen, J.D., Andersen, M., Bjørner, T.B., Hauch, J. & Topping, C.J. (2005) Pesticide reducing instruments: an interdiciplinary analysis of effectiveness and optimality. 8. Annual Conference on Global Economic Analysis pp. 1-25.
Jacobsen, I. & Rosendahl, S. (2006) Fungiciders påvirkning af mykorrhizasvampes diversitet og funktion. Bekæmpelsesmiddelforskning fra Miljøstyrelsen 103. Miljøstyrelsen, Miljøministeriet.
Jensen, J. (2006) Landdistriktsprogrammet akse 2: Varienerende landskaber, rig natur og rent miljø. Momentum 4, 12-15.
Jepsen, J.U. (2004) Spatially explicit models in landscape and species management. PhD thesis. National Environmental Research Institute, Ministry of the Environment. Denmark.
Joyce, K.A., Holland, J.M. & Doncaster, C.P. (1999) Influences of hedgerow intersections and gaps on the movement of carabid beetles. Bulletin of Entomological Research 89, 523-531.
Kajak, A. & Lukasiewicz, J. (1994) Do semi-natural patches enrich crop fields with predatory epigean arthropods. Agriculture, Ecosystems and Environment 49, 149–161.
Kauppila, R. (1990) Conventional and Organic Cropping Systems at Suitia. 4. Weeds. Journal of Agricultural Science in Finland 62, 331-337.
Kells, A.R. & Goulson, D. (2003) Preferred nesting sites of bumblebee queens (Hymenoptera: Apidae) in agroecosystems in the UK. Biological Conservation 109, 165-174.
Kleijn, D., Baquero, R.A., Clough, Y., Diaz, M., De Esteban, J., Fernandez, F., Gabriel, D., Herzog, F., Holzschuh, A., Johl, R., Knop, E., Kruess, A., Marshall, E. J.P., Steffan-Dewenter, I., Tscharntke, T., Verhulst, J., West, T.M. & Yela, J.L. (2006) Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecology Letters 9, 243-254.
Kleijn, D. & Snoeijing, G.I.J. (1997) Field boundary vegetation and the effects of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34, 1413-1425.
Kleijn, D. & Sutherland, W.J. (2003) How effective are European agri-environment schemes in conserving and promoting biodiversity? Journal of Applied Ecology 40, 947-969.
Klinger, K. (1987) Effects of margin-strips along a winter wheat field on predatory arthropods and the infestation by cereal aphids. Journal of Applied Entomology 104, 47-58.
Kremen, C. & Ostfeld, R.S. (2005) A call to ecologists: measuring, analyzing, and managing ecosystem services. Frontiers in Ecology and the Environment 3, 540-548.
Kromp, B. (1999) Carabid beetles in sustainable agriculture: a review of pest control efficacy, cultivation impact and enhancement. Agriculture, Ecosystems and Environment 74, 187-288.
Kuemmel B. (2003) Theoretical investigation of the effects of field margin and hedges on crop yields. Agriculture, Ecosystems and Environment 95, 387-392.
Lacas, J.G., Voltz, M., Gouy, V., Carluer, N. & Gril, J.J. (2005) Using grassed strips to limit pesticide transfer to surface water: a review. Agronomy for Sustainable Development 25, 253-266.
Langer, V. (2001) The potential of leys and short rotation coppice hedges as reservoirs for parasitoids of cereal aphids in organic agriculture. Agriculture Ecosystems & Environment 87, 81-92.
Langhof, M., Gathmann, A. & Poehling, H.M. (2005) Insecticide drift deposition on noncrop plant surfaces and its impact on two beneficial nontarget arthropods, Aphidius colemani viereck (Hymenoptera, Braconidae) and Coccinella septempunctata L. (Coleoptera, Coccinellidae). Environmental Toxicology and Chemistry 24, 2045-2054.
Larsson, S.G. (1939) Entwicklungstypen und Entwicklungszeiten der Dänischen Carabidaen. Entomologiske meddelelser 20, 277-560.
Laurance, W. F., Yensen, E. (1991). Predicting the impacts of edge effects in fragmented habitats. Biological Conservation 55:11, 77-92.
Lee, J.C., Menalled, F.B. & Landis, D.A. (2001) Refuge habitats modify impact of insecticide disturbance on carabid beetle communities. Journal of Applied Ecology 38, 472-483.
Lethmayer, C., Nentwig, W. & Frank, T. (1997) Effects of weed strips on the occurrence of noxious coleopteran species (Nitidulidae, Chrysomelidae, Curculionidae). Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz-Journal of Plant Diseases and Protection 104, 75-92.
Levidow, L. & Bijman, J. (2002) Farm inputs under pressure from the European food industry. Food Policy 27, 31-45.
Levin, G. & Brandt, J. (2006) Dept. of Environmental, Social and Spatial Change (ENSPAC) Roskilde Universitetscenter/ Skov- og naturstyrelsen.
Levins, R. (1969) Some demographic and genetic consequences of environmental heterogeneity for biological control. Bulletin of the Entomological Society of America 15, 237-240.
Lille, R. (1996) Zur Bedeutung von Bracheflächen für die Avifauna der Agrarlandschaft: Eine nahrungsùkologische Studie an der Goldammer Emberiza citrinella. Agrarùkologie 21. Haupt, Bern.
Luczak, J. (1979) Spiders in agrocenoses. Polish Ecological Studies 5, 151–200.
Lys, J.-A. (1994) Carabid Beetles: Ecology and Evolution (eds K.Desender, M.Dufrêne, M.Loreau, M.L.Luff & J.-P.Maelfait), pp. 451-455. Kluwer Academic Publishers, Dordrecht / Boston / London.
Lys, J.A. & Nentwig, W. (1992) Augmentation of Beneficial Arthropods by Strip-Management .4. Surface-Activity, Movements and Activity Density of Abundant Carabid Beetles in A Cereal Field. Oecologia 92, 373-382.
Lys, J.A. & Nentwig, W. (1994) Improvement of the Overwintering Sites for Carabidae, Staphylinidae and Araneae by Strip-Management in A Cereal Field. Pedobiologia 38, 238-242.
Lys, J.A., Zimmermann, M. & Nentwig, W. (1994) Increase in Activity Density and Species Number of Carabid Beetles in Cereals As A Result of Strip-Management. Entomologia Experimentalis et Applicata 73, 1-9.
MacLeod, A., Wratten, S.D., Sotherton, N.W. & Thomas, M.B. (2004) 'Beetle banks' as refuges for beneficial arthropods in farmland: long-term changes in predator communities and habitat. Agricultural and Forest Entomology 6, 147-154.
Mader, H.J. (1988) Effects of increased spatial heterogeneity on the biocenosis in rural landscapes. Ecological Bulletins 39, 169-179.
Maelfait, J. P. & De Keer, R. (1990) The border zone of an intensively grazed pasture as a corridor for spiders (Araneae). Conservation 54, 223–238.
Marshall, E.J.P. (1989) Distribution patterns of plants associated with arable field edges. Journal of Applied Ecology 26, 247-258.
Marshall, E.J.P., West, T.M. & Kleijn, D. (2006) Impacts of an agri-environment field margin prescription on the flora and fauna of arable farmland in different landscapes. Agriculture Ecosystems & Environment 113, 36-44.
Marshall, E.-J.P. & Moonen, A.C. (2002) Field margins in northern Europe: their functions and interactions with agriculture. Agriculture, Ecosystems and Environment 89, 5-21.
Marshall, E.-J.P., West, T.M. & Winstone, L. (1994) Extending field boundary habitats to enhance farmland wildlife and improve crop and environmental protection. Aspects of Applied Biology 40, 387-391.
Matthies, D., Schmid, B. & Schmid-Hempel, P. (1995) The importance of population processes for the maintenance of biological diversity. Gaia 4, 199-209.
Meek, B., Loxton, D., Sparks, T., Pywell, R., Pickett, H. & Nowakowski, M. (2002) The effect of arable field margin composition on invertebrate biodiversity. Biological Conservation 106, 259-271.
Meeus, J.H.A. (1995) Pan-European landscapes. Landscape and Urban Planning 57-79.
Mensah, R.K. (2004) Ecological Engineering for Pest Management, Advances in Habitat Manipulation for Arthropods (eds. G.M.Gurr, S.D.Wratten & M.G.Altieri), pp. 187-197. CSIRO Publishing; CABI Publishing.
Meyling, N.V. (2005) Population ecology and genetic diversity of entomopathogenic fungi (Ascomycota: Hypocreales) in agroecosystems and field margins. Ph.D. thesis. Department of Ecology, Faculty of Lifesciences, University of Copenhagen.
Miljøstyrelsen (1999) Bichel-udvalget, Miljøstyrelsen.
Miljø- og Energiministeriet, Wilhjelmudvalget (2001) Et rigt natur i et rigt samfund. http://www.sns.dk/wilhjelm/endelig/
Moore, N.W., Hooper, M.D. & Davis, B.N.K. (1967) Hedges. Introduction and Reconnaissance Studies. Journal of Applied Ecology 4, 201-?
Moreby, S.J. & Sotherton, N. W. (1997) A comparison of some important chick-food insect groups found in organic and conventionaly-grown winter wheat fields in southern England. Biological Agriculture & Horticulture 15, 51-60.
Murray, K.A., Wilcox, A. & Stoate, C. (2002) A simultaneous assessment of farmland habitat use by breeding skylarks and yellowhammers. Aspects of Applied Biology 67, 121-127.
Navntoft, S., Petersen, B.S., Esbjerg, P., Jensen, A.-M., Johnsen, I., Kristensen, K., Petersen, P.H. & Ørum, J.E. (unpubl.) Effects of mechanical weed control in spring cereals – flora, fauna and economy. Draft version submittet to Pesticides Research. Danish Environmental Protection Agency. Danish Ministry of the Environment.
Navntoft, S., Esbjerg, P., Jensen, A.-M. M., Johnsen, I. & Petersen, B.S. (2003) Flora and Fauna Changes During Conversion from Conventional to Organic Farming. Pesticides Research 74. Danish Environmental Protection Agency, Danish Ministry of Environment, Denmark.
Navntoft, S., Esbjerg, P. & Riedel, W. (2006) Effects of reduced pesticide dosages on carabids (Coleoptera: Carabidae) in winter wheat. Agricultural and Forest Entomology 8, 57-62.
New, T.R. (2005). Invertebrate conservation and agricultural ecosystems. Cambridge University Press.
Nordén, B. & Appelqvist, T. (2001) Conceptual problems of Ecological Continuity and its bioindicators. Biodiversity and Conservation 10, 779-791.
Nyffeler, M. & Breene, R.G. (1992) Dominant insectivorous polyphagous predators in winter wheat: high colonization power, spatial dispersion patterns and probable importance of the soil surface spiders. Deutsche Entomologische Zeitschrift 39, 177–188.
Nørum, U. & Bjerregaard, P. (2003) Ferskvandsinvertebraters bevægelsesadfærd som biomarkør for pesticideksponering og –effekt. Bekæmpelsesmiddelforskning 75 Miljøstyrelsen, Miljøministeriet.
Odderskaer,P., Prang, A., Poulsen, J.G., Andersen, P.N. & Elmegaard, N. (1997) Skylark (Alauda arvensis) utilisation of micro-habitats in spring barley fields. Agriculture Ecosystems & Environment 62, 21-29.
Ormerod, S.J. & Watkins, A.R. (2000) Birds and agriculture. Journal of Applied Ecology 37, 699-705.
Ùstman, Ù., Ekbom, B. & Bengtsson, J. (2003) Yield increase attributable to aphid predation by ground-living polyphagous natural enemies in spring barley in Sweden. Ecological Economics 45, 149-158.
Perkins,A.J., Whittingham, M.J., Morris, A.J. & Bradbury, R.B. (2002) Use of field margins by foraging yellowhammers Emberiza citrinella. Agriculture Ecosystems & Environment 93, 413-420.
Pascual-Villalobos, M.J., Lacasa, A., Gonzalez, A., Varo, P. & Garcia, M.J. (2006) Effect of flowering plant strips on aphid and syrphid populations in lettuce. European Journal of Agronomy 24, 182-185.
Petersen, B.S. (1998) The distribution of Danish farmland birdsin relation to habitat characteristics. Ornis Fennica 75, 105-108.
Petersen, M.K. (1997) Life Histories of two Predaceous Beetles, Bembidion lampros og Tachyporous hypnorum in Agroecosystems. Agraria 35. Swedish University of Agricultural Scinces, Uppsala.
Petersen, S., Axelsen, J.A., Tybirk, K., Aude, E. & Vestergaard, P. (2006) Effects of organic farming on field boundary vegetation in Denmark. Agriculture Ecosystems & Environment 113, 302-306.
Pimentel, D., Wilson, C., McCullum, C., Huang, R., Dwen, P., Flack, J., Tran, Q., Saltman, T. & Cliff, B. (1997) Economic and Environmental Benefits of Biodiversity. BioScience 47, 747-757.
Potts, G.R. (1986) The Partridge. Pesticides, Predation and Conservation. Collins, London.
Potts, G.R. & Vickerman, G. P. (1974) Studies of the cereal ecosystem. Advances in Ecological Research 8, 107-197.
Powell, W., Dean, G.J. & Dewar, A. (1985) The influence of weeds on polyphagous arthropod predators in winter wheat. Crop Protection 4, 298-312.
Powell, W., Hara, S.A., Harling, R., Holland, J.M., Northing, P., Thomas, C.F.G. & Walters, K.F.A. (2004) Managing biodiversity in field margins to enhance integrated pest control in arable crops ('3-D Farming' Project) HGCA report 365, pp. 1-80.
Prasad, R.P. & Snyder, W.E. (2006) Polyphagy complicates conservation biological control that targets generalist predators. Journal of Applied Ecology 43, 343-352.
Pretty, J.N., Brett, C., Gee, D., Hine, R.E., Mason, C.F., Morison, J.I.L., Raven, H., Rayment, M.D. & van der Bijl, G. (2000) An assessment of the total external costs of UK agriculture. Agricultural Systems 65, 113-136.
Pywell, R.F., Warman, E.A., Carvell, C., Sparks, T.H., Dicks, L.V., Bennett, D., Wright, A., Critchley, C.-N.R. & Sherwood, A. (2005) Providing foraging resources for bumblebees in intensively farmed landscapes. Biological Conservation 121, 479-494.
Rand, T.A., Tylianakis, J.M. & Tscharntke, T. (2006a) Spillover edge effects: the dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecology Letters 9.
Raskin, R. (1994) Wirking pflanzenschutzmittelfreier ackerrandstreifen auf die entomofauna wintergetreidefeldern und angrenzenden saumbiotopen. PhD thesis. Aachen: Shaker.
Rasmussen, I.A., Holst, N., Graglia, E., Hansen, P.K., Melander, B., Mathiasen, S. K., Kudsk, P., Jensen, P.K., Boelt, B. & Madsen, K.H. (2004) Ukrudtsbogen (eds. K.H. Madsen & J. Kakobsen), pp. 19-42. Danmarks JordbrugsForskning.
Ravn, H.W. & Friberg, N. (2004) Effekt på akvatiske miljøer af randzoner langs målsatte vandløb - Pesticidhandlingsplan II. Faglig rapport fra DMU nr. 487.
Richards, A.J. (2001) Does low biodiversity resulting from modern agricultural practice affect crop pollination and yield? Annals of Botany 88, 165-172.
Riechert, S. E. & Gillespie R.G. (1986). Habitat choice and utilization in web-building spiders. In: W. A. Shear (ed.) Spiders; Webs Behaviour and Evolution. Stanford University Press, California, pp. 23–48.
Riechert, S.E. & Bishop, L. (1990) Pest control by an assemblage of generalist predators: Spiders in garden test systems. Ecology 71, 1441–1450.
Riedel, W. (1991) Overwintering and dispersal of Bembidion lampros (Coleoptera: Carabidae) from established hibernation sites in winter wheat fields in Denmark (eds L.Polgár, R.J.Chambers, A.F.G.Dixon & I.Hodek), pp. 235-241. Academic Publishing bv, The Hague.
Riedel, W. (1992) Hibernation and Spring Dispersal of polyphsgous predators in arable land. Ph.D. thesis in biology. Institute of Biology, Department of Zoology, University of Aarhus, Denmark.
Rundlùf, M. & Smith, H.G. (2006) The effect of organic farming on butterfly diversity depends on landscape context. Journal of Applied Ecology 43, 1121-1127.
Rypstra, A. L. & Carter, P.E. (1995) The web-spider community of soybean agroecosystems in southwestern Ohio. Journal of Arachnology 23, 135–144.
Samu, F. & Szinetár, C. (2002) On the nature of agrobiont spiders. Journal of Arachnology 30, 389–402.
Samways, M.J. (2005) Insect Diversity Conservation. Cambridge University Press.
Sigsgaard, L. (2000) The temperature-dependent duration of development and parasitism of three cereal aphid parasitoids, Aphidius ervi, A. rhopalosiphi, and Praon volucre. Entomologia experimentalis et applicata 95, 173-184.
Sigsgaard, L. (2002) A survey of aphids and aphid parasitoids in cereal fields in Denmark, and the parasitoids' role in biological control. Journal of Applied Entomology-Zeitschrift fur Angewandte Entomologie 126, 101-107.
Smart, S.M., Bunce, R.G.H., Firbank, L.G. & Coward, P. (2002) Do field boundaries act as refugia for grassland plant species diversity in intensively managed agricultural landscapes in Britain? Agriculture Ecosystems & Environment 91, 73-87.
Smart, S.M., Marrs, R.H., Le Duc, M.G., Thompson, K.E.N., Bunce, R.G.H., Firbank, L. & Rossall, M.J. (2006) Spatial relationships between intensive land cover and residual plant species diversity in temperate farmed landscapes. Journal of Applied Ecology 43, 1128-1137.
Snoo, G.-R. d. (1994) Field margins: integrating agriculture and conservation. Proceedings of a symposium held at Coventry, UK, 18-20 April 1994 (ed N.D.Boatman), Farnham, UK: British Crop Protection Council (BCPC).pp. 197-201.
Snoo, G.-R.d., Poll, R.-J. R.-J. & Bertels, J. (1998) Butterflies in sprayed and unsprayed field margins. Journal of Applied Entomology 122, 157-161.
Snoo, G.-R.d. & Chaney, K. (1999) Unsprayed field margins - what are we trying to achieve? Aspects of Applied Biology 54, 1-12.
Snyder, W.E., Snyder, G.B., Finke, D.L. & Straub, C.S. (2006) Predator biodiversity strengthens herbivore suppression. Ecology Letters 9, 789-796.
Sotherton, N.W. (1987) Farming and Wildlife. Naturopa 55E, 20-21.
Sotherton, N.W., Boatman, N.D. & Chiverton, P.A. (1989) The selective use of pesticides on cereal crop margins for game and wild life: The British experience. 30 swedish Crop Protection Conference 4, 18-29.
Sotherton, N.W. & Moreby, S.J. (2006) The importance of beneficial arthropods other than natural enemies in cereal fields. Aspects of Applied Biology 31, 11-18.
Sparkes, D.L., Jaggard, K.W., Ramsden, S.J. & Scott, R.K. (1998) The effect of field margins on the yield of sugar beet and cereal crops. Annals of Applied Biology 132, 129-142.
Sparks, T.H., Meek, W., Pywell, R.F. & Nowakowski, M. (2000) The influence of field margin management on the butterfly fauna of a Yorkshire arable farm. Aspects of Applied Biology 58, 407-410.
Spehn, E.M., Hector, A., Joshi, J., Scherer-Lorenzen, M., Schmid, B., Bazeley-White, E., Beierkuhnlein, C., Caldeira, M.C., Diemer, M., Dimitrakopoulos, P.G., Finn, J.A., Freitas, H., Giller, P.S., Good, J., Harris, R., Hogberg, P., Huss-Danell, K., Jumpponen, A., Koricheva, J., Leadley, P.W., Loreau, M., Minns, A., Mulder, C.P.H., O'Donovan, G., Otway, S. J., Palmborg, C., Pereira, J. S., Pfisterer, A.B.,
Prinz, A., Read, D.J., Schulze, E.D. & Siamantziouras, A.S.D. (2005) Ecosystem effects of biodiversity manipulations in European grasslands. Ecological Monographs 75, 37-63.
Steffan-Dewenter, I. & Tscharntke, T. (1997) Early succession of butterfly and plant communities on set-aside fields. Oecologia 109, 294-302.
Steffan-Dewenter, I. Münzenberg, U., Bürger, C., Thies, C. & Tscharntke, T (2002) Scale-dependent effects of landscape context on three pollinator guilds. Ecology 3, 1421–1432.
Steiner, H., El Titi, A. & Bosch, J. (1986) Integrierter Pflanzenschutz im Ackerbau: Das Lautenbach-Project I. Versuchsprogramm. Zeitschrift für Pflanzenkrankheiten end Pflanzenschutz 93, 1-18.
Steytler, N.S. & Samways, M.J. (1995) Biotope selection by adult male dragonflies (Odonata) at an artificial lake created for insect conservation in South Africa. Biological Conservation 72, 381-386.
Stoltze, M. & Phil, S. (eds.) (1998) Rødliste 1997 over planter og dyr i Danmark. Miljø- og Energiministeriet, Danmarks Miljøundersøgelser og Skov- og Naturstyrelsen, København.
Sunderland, K.D. (1975) Diet of Some Predatory Arthropods in Cereal Crops. Journal of Applied Ecology 12, 507-515.
Sunderland, K.D. (1987) Spiders and cereal aphids in Europe. Bulletin SROP/WPRS 1987/X/1: 82–102.
Sunderland, K.D. (1991) The ecology of spiders in cereals. Proceedings of the 6th International Symposium on Pests and Diseases of Small Grain Cereals. Board of Plant Protection Halle, Halle/Saale, Germany. 1: 269–280.
Sunderland, K.D. & Vickerman, G. P. (1980) Aphid Feeding by Some Polyphagous Predators in Relation to Aphid Density in Cereal Fields. Journal of Applied Ecology 17, 389-396.
Sunderland, K.D., Fraser, A.M. & Dixon, A.F.G. (1986) Field and laboratory studies on money spiders (Linyphiidae) as predators of cereal aphids. Journal of Applied Ecology 23, 433–447.
Sunderland, K.D., Bilde, T., Den Nijs, L.J.M.F., Dinter, A., Heimbach, U., Lys, J.A., Powell, W. & Toft, A. (1994) Reproduction of beneficial predators and parasitoids in agroecosystems in relation to habitat quality and food availability. I: Booij, K., Den Nijs, L. (eds.). Arthropod natural enemies in arable land II. Survival, Reproduction and Enhancement. Acta Jutlandica 71, 117-153.
Sunderland, K. & Samu, F. (2001) Effects of agricultural iversification on the abundance, distribution, and pest control potential of spiders: a review. Entomologia Experimentalis et Applicata 95, 1–13, 2000.
Sutherland, J.P., Sullivan, M.S. & Poppy, G.M. (2001) Distribution and abundance of aphidiophagous hoverflies (Diptera: Syrphidae) in wildflower patches and field margin habitats. Agricultural and Forest Entomology 3, 57-64.
Sutherland, W.J. (2004) A blueprint for the countryside. Ibis 146, 230-238.
Symondson, W.O.C., Sunderland, K. D. & Greenstone, M. H. (2002) Can generalist predators be effective biocontrol agents? Annual Review of Entomology 47, 561-594.
Sønderskov, M., Axelsen, J.A., Pedersen, M.B. & Tybirk, K. (2006) Assessment of the effects of reduced herbicide applications on selected arable weeds by a simulation model. Agriculture Ecosystems & Environment 116, 216-224.
Tansley, A.G. (1935) The use and abuse of vegetational concepts and terms. Ecology 16, 284-307.
Tew, T.E., Macdonald, D.W. & Rands, M.R.W. (1992) Herbicide Application Affects Microhabitat Use by Arable Wood Mice (Apodemus-Sylvaticus). Journal of Applied Ecology 29, 532-539.
Terry, A.C., Troumbis, A.Y., Woodward, F.I., Yachi, S. & Lawton, J.H. (2005) Ecosystem effects of biodiversity manipulations in European grasslands. Ecological Monographs 75, 37-63.
Thomas, C.F.G. (1996). Modelling aerial dispersal of linyphiid spiders. Aspects of Applied Biology 46, 217–222.
Thomas, C.F.G. & Jepson, P.C. (1997) Field-scale effects of farming practices on linyphiid spider populations in grass and cereals. Entomologia Experimentalis et Applicata 84, 59–69.
Thomas, C.F.G. & Marshall, E.J.P. (1999) Arthropod abundance and diversity in differently vegetated margins of arable fields. Agriculture, Ecosystems and Environment 72, 131–144.
Thomas, M.B. & Wratten, S.D. (1988) Manipulating the arable crop environment to enhance the activity of predatory insects. Aspects of Applied Biology 17, 57-66.
Thomas, M.B., Wratten, S.D. & Sotherton, N.W. (1991) Creation of Island Habitats in Farmland to Manipulate Populations of Beneficial Arthropods - Predator Densities and Emigration. Journal of Applied Ecology 28, 906-917.
Thomas, M.B., Mitchell, H.J. & Wratten, S.D. (1992) Abiotic and Biotic Factors Influencing the Winter Distribution of Predatory Insects. Oecologia 89, 78-84.
Thomas, M.B., Wratten, S.D. & Sotherton, N.W. (1992) Creation of Island Habitats in Farmland to Manipulate Populations of Beneficial Arthropods - Predator Densities and Species Composition. Journal of Applied Ecology 29, 524-531.
Thomas, S.R., Goulson, D. & Holland, J.M. (2000a) Spatial and temporal distributions of predatory Carabidae in a winter wheat field. Aspects of Applied Biology 62, 55-60.
Thomas, S.R., Goulson, D. & Holland, J.M. (2000b) The contribution of beetle banks to farmland biodiversity. Aspects of Applied Biology 62, 31-38.
Thomas, S.R., Goulson, D. & Holland, J.M. (2001) Ressource provision for farmland gamebirds: the value of beetle banks. Annals of Applied Biology 139, 111-118.
Thomas, S.R., Noordhuis, R., Holland, J.M. & Goulson, D. (2002) Botanical diversity of beetle banks effects of age and comparison with conventional arable field margins in southern UK. Agriculture Ecosystems & Environment 93, 403-412.
Toft, S. (1995) Two functions of gossamer dispersal in spiders? Acta Jutlandica 70, 257–268.
Topping, C.J. (1997) Predicting the effect of landscape heterogeneity on the distribution of spiders in agroecosystems using a population dynamics driven landscape-scale simulation model. Biological and Agricultural Horticulture 15, 325–336.
Topping, C.J. (1999) An individual-based model for dispersive spiders in agroecosystems: simulations of the effects of landscape structure. Journal of Arachnology, 378–386.
Tscharntke, T., Klein, A.M., Kruess, A., Steffan-Dewenter, I. & Thies, C. (2005) Landscape perspectives on agricultural intensification and biodiversity - ecosystem service management. Ecology Letters 8, 857-874.
Tscharntke, T., Rand, T.A. & Bianchi, F.J.J.A. (2005) The landscape context of trophic interactions: insect spillover across the crop-noncrop interface. Annales Zoologici Fennici 42, 421-432.
Van Lenteren, J.C. (2006). IOBC Internet Book of Biological Control. http://www.unipa.it/iobc/downlaod/IOBC%20InternetBookBiCoVersion4October2006.pdf
Varchola, J.M. & Dunn, J.P. (1999) Changes in ground beetle (Coleoptera : Carabidae) assemblages in farming systems bordered by complex or simple roadside vegetation. Agriculture Ecosystems & Environment 73, 41-49.
Vickerman, G.P. (1992) The effects of different pesticide regimes on the invertebrate fauna of winter wheat. In: P. Grieg-Smith, G. Frampton & T. Hardy (eds), Pesticides, Cereal Farming and the Environment. HMSO, London, pp. 82-110.
Vickery, J., Carter, N. & Fuller, R.J. (2002) The potential value of managed cereal field margins as foraging habitats for farmland birds in the UK. Agriculture Ecosystems & Environment 89, 41-52.
Wackers, F.L. & Steppuhn, A. (2003) Characterizing nutritional state and food source use of parasitoids collected in fields with high and low nectar availability. Bulletin OILB/SROP 26, 203-208.
Wallenius, S. & Sihvonen, T. (2000) Ympäristùtukiopas (Guide for environmental support, in Finnish).pp. 1-27. Ministry of agriculture and forestry.
Wallin, H. (1989) The influence of different age classes on the seasonal activity and reproduction of 4 medium-sized carabid species inhabiting cereal fields. Holarctic Ecology 12, 201-212.
Wallin, H., Chiverton, P.A., Ekbom, B.S. & Borg, A. (1992) Diet, fecundity and egg size in some polyphagous predatory carabid beetles. Entomologia Experimentalis et Applicata 65, 129-140.
Weibull, A.C., Ùstman, O. & Granqvist, A. (2003) Species richness in agroecosystems: the effect of landscape, habitat and farm management. Biodiversity and Conservation 12, 1335-1355.
Wenger, K.F. (1984) Forestry handbook. John Wiley & Sons, New York.
Winkler, K. (2006) Assessing the Risks and Benefits of Flowering Field Edges. Startegic Use and Nectar Sources to Boost Biological Control. Wageningen University.
White, A.J., Wratten, S.D., Berry, N.A. & Weigmann, U. (1995) Habitat manipulation to enhance biological control of Brassica pests by hover flies (Diptera: Syrphidae). Journal of Economic Entomology 88, 1171-1176.
Whitfield, J. (2006) Agriculture and environment: How green was my subsidy? Nature 439, 908-909.
Wilson, P.J. & Aebischer, N.J. (1995) The distribution of dicotyledonous arable weeds in relation to distance from the field edge. Journal of Applied Ecology 32, 295-310.
Wilson, J.D., Morris, A.J., Arroyo, B.E., Clark, S.C. & Bradbury, R.B. (1999) A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change. Agriculture Ecosystems & Environment 75, 13-30.
Wilson, P.J. (1997) Conserving the flora of arable field margins - how much does it cost? 1997 Brighton crop protection conference: weeds Proceedings of an international conference, Brighton, UK, 17 20 November 1997 991-996.
Winkler, K. (2005) Assessing the risk and benefits of flowering field edges. Strategic use of nectar sources to boost biological control. Thesis. Wageningen University.
Wise, D.H., Snyder, W.E. & Tuntibunpakul, P. 1999. Spiders in decomposition food webs of agroecosystems. Journal of Arachnology 27, 363–370.
Wittingham, M.J. (2007) Will agri-environment schemes deliver substantial biodiversity gain, and if not why not? Journal of Applied Ecology, 44, 1-5.
Whittingham, M.J., Swetnam, R.D., Wilson, J.D., Chamberlain, D.E. & Freckleton, R.P. (2005) Habitat selection by yellowhammers Emberiza citrinella on lowland farmland at two spatial scales: implications for conservation management. Journal of Applied Ecology 42, 270-280.
Woodcock, B.A., Potts, S.G., Pilgrim, E., Ramsay, A.J., Tscheulin, T., Parkinson, A., Smoth, R.E.N., Gundrey, A.L., Brown, V.K. & Tallowin, J.R. (2007) The potential of grass field margin management for enhancing beetle diversity in intensive livestock farms. Journal of Applied Ecology 44, 60-69.
Woodcock, B.A., Westbury, D.B., Potts, S.G., Harris, S.J. & Brown, V.K. (2005) Establishing field margins to promote beetle conservation in arable farms. Agriculture Ecosystems and Environment 107, 255-266.
Wossink,G.A.A. & Wenum, J.-H. v. (2003) Biodiversity conservation by farmers: analysis of actual and contingent participation. European Review of Agricultural Economics 30, 461-485.
Wratten, S.D. & Thomas, C.F.G. (1990) Farm-scale spatial dynamics of predators and parasitoids in agricultural landscapes. Species dispersal in agricultural habitats. Belhaven Press, London 219-237.
Zande, J.C.v.d., Michielsen, J.M.G.P., Stallinga, H. & Jong, A.d. (2000) The effect of windbreak height and air assistance on exposure of surface water via spray drift. Farnham, UK: British Crop Protection Council.
Zangger, A., Lys, J. A. & Nentwig, W. (1994) Increasing the Availability of Food and the Reproduction of Poecilus-Cupreus in A Cereal Field by Strip-Management. Entomologia Experimentalis et Applicata 71, 111-120.
Zeddies, J. & Jung, G. (1986) Integrierter Pflanzenschutz im Ackerbau: Das Lautenbach-Project II. Ùkonomische Auswirkungen. Zeitschrift für Pflanzenkrankheiten end Pflanzenschutz 93, 449-461.
Zschokke S., Dolt C., Rusterholz H.-P., Oggier P., Braschler B. & Thommen G.H. et al. (2000) Short-term responses of plants and invertebrates to experimental small-scale grassland fragmentation. Oecologia 125, 559–572.
Bilag A: Skematiske oversigter
Tabel 1. Bufferzoner. nogle Internationale støtteordninger.
Kilder: DEFRA, www.defra.uk.gov; (Hartmann et al. 2006), Landwirtschaftkammer Niedersachsen, 2006 (http://www.lwk-niedersachsen.de, (Heikkilä 2001;Wallenius & Sihvonen 2000) Det svenske Jordbrugsverket (www.sjv.se), (Wossink & Wenum 2003).
Bilag B: IGANGVÆRENDE RELEVANTE PROJEKTER I DANMARK
Projekter relateret til randzoneproblematikken:
Danmarks Jordbrugsforskning: Hvordan ser europæisk landbrug ud om 25 år?
Forskere ved DJF er med til at udarbejde modeller for, hvordan det europæiske landbrugsland kan komme til at se ud om 25 år. Modellerne skal bl.a. give politikerne et billede af, hvordan den fælles landbrugspolitik vil påvirke landdistrikterne.
Bruus et al.: Vurdering af omfang og effekt af afdrift af ukrudtsmidler til danske læhegn eksemplificeret ved metsulfuron. 2004-2007.
Pesticidforskningsprojekt
I projekterne ”Bærbærende buske og træer som indikatorer for biodiversitetsforandringer ved sprøjtemiddelafdrift. I og II” er det i kontrollerede felteksperimenter påvist, at minimidlet metsulfuron methyl påvirker produktionen af tjørnebær. Kontrollerede feltforsøg som disse kan imidlertid ikke umiddelbart beskrive konsekvenserne ved en afdrifthændelse. For det første er de gennemført som manipulerede forsøg, hvor sprøjtemidlet er sprøjtet direkte ind i hegnet med en rygsæksprøjte. Dernæst eksponeres hegnet ved afdrift fra mark fra overlappende sprøjtefaner og eventuelt fra flere sider. Endelig bruger mange landmænd reducerede doseringer i forhold til den af producenten angivne standarddose. Nærværende projekt har til formål at undersøge om og i hvilken grad bærbærende hegnstræer påvirkes af afdrift af metsulfuron methyl fra mark til hegn. Projektet vil endvidere vurdere konsekvensen af forskellige måder at forvalte sprøjtningen.
For at bestemme omfanget af afdrift må man kunne måle eller beregne, hvor stor en mængde sprøjtemiddel der rammer hegnet. Dette projekt vil måle sprøjtemiddelafsætningen i hegn efter en realistisk sprøjtning med flere overlappende sprøjtespor. Udfra disse data udvikles en model til at forudsige afsætningen under varierende betingelser. Efterfølgende skal denne model udnyttes til at beregne effekter af metsulfuronsprøjtning på flere hegnsarter. Effekten bestemmes på basis af den beregnede eksponering og den relation mellem sprøjtemiddelafsætning og effekt som er etableret gennem ”Bærbærende buske og træer som indikatorer for biodiversitetsforandringer ved sprøjtemiddelafdrift. II” samt i nærværende projekt for seljerøn og hyld.
Resultaterne af projektet vil kunne understøtte en vurdering af i hvilket omfang ukrudtsmiddelafdrift påvirker bærbærende hegnstræer. Hertil kommer en vurdering af om en ændring af sprøjtemiddelanvendelsen vil resultere i øget fødemængde for bærspisende organismer i hegn, specielt fugle, under danske vilkår. Vi vil fokusere på hvorledes variation af sprøjtedosering, dysetype, vejrtype og afstand fra markkant til hegn vil påvirke bærmængden. Projektet kan herigennem medvirke til at vurdere en eventuel gevinst ved administrativ regulering af sprøjtemiddelanvendelsen. Projektet består således af tre delforsøg: A: Vindafdrift af metsulfuron fra sprøjtespor hegn; B: Følsomheden af udvalgte hegnsarter overfor metsulfuron og C: Udbredelsen af bærbærende træer og buske i danske læhegn. Data fra disse forsøg kombineres i en række scenarieberegninger af effekten af metsulfuron på hegnsarter der producerer bær.
Strandberg et al.: Herbicidtolerante GM-afgrøder: Langtidseffekter af sprøjtestrategier på flora og fauna i mark og tilstødende biotoper. 2004-2007.
Pesticidforskningsprojekt
Dyrkning af genetisk modificerede afgrøder med tolerance over for glyphosat vil sandsynligvis ændre herbicidanvendelsen i dansk planteavl. Dermed vil forholdene for markens ukrudt og dyreliv kunne ændres, ligesom forholdene for flora og fauna i de omgivende markkanter vil kunne påvirkes som følge af sprøjteafdrift. I dette projekt vil vi derfor undersøge, hvorledes anvendelse af glyphosat i løbet af forsommeren i afgrøderne vinterraps og majs vil påvirke flora i mark og tilstødende hegn både på kort og langt sigt med deraf afledte effekter på faunaen. Foreløbige resultater fra bl.a. foderroer viser, at timingen af glyphosatsprøjtningerne er afgørende for ændringerne på kort sigt, idet sen glyphosatsprøjtning sammenlignet med konventionel sprøjtning kan gøre det muligt at forbedre forholdene for markens plante- og dyreliv i forsommeren i forhold til sprøjtestrategier med konventionelle herbicider. På længere sigt er det dog muligt, at ændringer i produktionen af ukrudtsfrø, som følge af glyphosat anvendt i GM-afgrøder, vil kunne påvirke ukrudtsfloraen både med hensyn til, hvilke arter der vil dominere og med hensyn til disses tæthed og biomasse med deraf afledte effekter på faunaen. For at belyse den samlede langsigtede effekt for flora og fauna i marken gennem hele sædskiftet og i markkanten i hegn benyttes modeller. Disse modeller integrerer den eksisterende viden om effekter som følge af glyphosatanvendelse i foråret, landbrugsmæssig erfaring og nye data fra dette projekt med henblik på at forudsige effekten på flora og fauna af forskellige sprøjtestrategier. Projektet vil således give viden om, hvorledes indførelsen af herbicidtolerante GM-afgrøder i sædskiftet, såvel på kort sigt (i sæsonen) som på langt sigt (over flere sædskifter), vil kunne påvirke antallet af plantearter og hvorledes disse vil udvikles over tid i såvel marken som tilstødende markkanter i hegn. Resultaterne vil blive sammenholdt med tilsvarende effekter som følge af dyrkning med konventionelle afgrøder. Projektet gennemføres således, at effekter på flora og fauna af forskellige strategier for glyphosatanvendelse kan vurderes i kombination med erfaringerne fra sociologiske undersøgelser af landmandens sprøjteadfærd. Et led i opstillingen af modelscenarier er deres sandsynlighed i forhold til den sociologiske undersøgelse. Hermed vil det være muligt at vurdere relevansen af de opstillede scenarier, der efterfølgende anvendes til at simulere langtidseffekterne. Med de samarbejdspartnere, der indgår i projektet, vil vi have mulighed for at kombinere landbrugsmæssig viden og forskningserfaring med erfaring i forskning i effekter på naturen i landbrugslandet. Samtidig vil vi sikre projektets samfundsmæssige relevans ved at undersøge, om der hos landmænd er den nødvendige opbakning til stede, for at det kan lykkes at realisere eventuelle naturmæssige gevinster i forbindelse med dyrkning af herbicidresistente afgrøder.
Strandberg et al.: Effekt af herbicider på planter i naturlige økosystemer: Hvordan kommer vi fra standardtest til naturlige habitater? 2006-2009. Pesticidforskningsprojekt
Det er velkendt, at herbicider kan påvirke de marknære omgivelser, dvs. hegn og markkanter. Der er imidlertid ikke mange undersøgelser af, om herbiciderne også har effekter længere væk fra marken. Dette aspekt er relevant bl.a. i relation til overholdelsen af EUs Habitatdirektiv og målet om at stoppe nedgangen i biodiversitet i 2010. De nuværende principper for godkendelse af herbicider bygger på simple, standardiserede test af afgrøde- og ukrudtsarter under væksthusforhold. Der er imidlertid mange forskelle mellem naturlige plantesamfund og enkeltplanter dyrket i væksthus.
Dette projekt undersøger betydningen af forskelle i eksponeringsform, arternes følsomhed over for herbicider, følsomheden af forskellige vækststadier, relevansen og følsomheden af forskellige mål for effekter af herbicider samt de komplekse interaktioner, der er i naturlige plantesamfund, for den enkelte arts følsomhed over for herbicider.
Målet er at kvantificere betydningen af de nævnte forhold, så vi bliver i stand til at vurdere, om naturlige plantesamfund i tilstrækkelig grad er beskyttet af de nuværende godkendelsesprocedurer, eller om nogle af de undersøgte forskelle mellem væksthusforhold og naturlige plantesamfund bør indgå i godkendelsesproceduren i fremtiden.
I projektet anvendes eksisterende data for effekter af herbicider på afgrøder, ukrudtsarter og vilde plantearter til at undersøge, om de plantearter, der forekommer i naturområder uden for markerne, er mere følsomme, end de arter, der for nuværende indgår i standardtests i forbindelse med godkendelse af sprøjtemidler. Analysen bruges også til at udvælge arter, der skal indgå i den øvrige del af projektet. For at belyse forskellene i eksponering mellem væksthus og naturlige plantesamfund, undersøger vi, om planter, der udsættes for herbicider via afdrift fra marksprøjtning, reagerer anderledes, end tilsvarende planter, der udsættes for de samme herbiciddoseringer ved direkte sprøjtning. Desuden sammenligner vi effekterne af sprøjtning på planter af forskellig størrelse og ser på, om vækst og reproduktion er lige følsomme mål for effekten. De nævnte undersøgelser foregår alle under kontrollerede forhold. For at belyse betydningen af konkurrence med andre planter, planteædende dyr, klima mv. for planters følsomhed over for herbicider følger vi desuden udviklingen af afmærkede enkeltplanter i et etableret plantesamfund, som udsættes for forskellige doseringer af herbicid og kvælstof.
Andersen et al. The role of Organic Farms as refugia for biodiversity (REFUGIA). 2006-2010. FØJO-projekt.
Økologisk jordbrug anses generelt for at have en positiv effekt på biodiversitet og andre landskabelige værdier. Gennem de sidste ti år er der imidlertid sket betydelige ændringer indenfor økologisk jordbrug og økologiske avlere omfatter i dag såvel den traditionelle holistiske økolog som økologen, der driver gården efter økonomisk optimering. I samme tidsrum er der også sket forbedringer af afgrøder og driftsmetoder, der har resulteret i effektive økologiske driftsformer som ofte kan sidestilles med konventionel landbrugsdrift. Økologisk jordbrug i dagens Danmark rummer derfor stor variation. I nærværende projekt undersøges effekten af økologisk jordbrug med varierende driftsintensitet på naturen med henblik på at belyse i hvilket omfang økologisk jordbrug fungerer som refugium for biodiversitet.
Det overordnede spørgsmål besvares ved at undersøge:
1) Strukturen, diversiteten og intensiteten af økologiske brug og de tre faktorers rolle for multi-funktionaliteten i Danmark. I forbindelse med disse undersøgelser udvikles nye teknikker til ud fra fly- og satellitfotos at undersøge forskellen i biodiversiteten på økologiske og konventionelle marker, og dermed økologisk jordbrugs rolle som refugium for biodiversitet (WP 2).
2) Ukrudt-insekt fødekæder i to afgrøder i økologiske marker, der dyrkes hhv. ekstensivt og intensivt. Resultaterne sammenholdes med eksisterende data fra tilsvarende konventionelle afgrøder. Undersøgelsen vil vise om der er mere ukrudt og mere ukrudts-baseret føde tilgængelig for markens dyreliv i en økologisk dyrket afgrøde sammenlignet med en konventionel. (WP 3)
3) Mængden af ukrudtsfrø, der produceres i hegn og markkanter på hhv. økologiske og konventionelle brug. Data vil belyse, i hvilket omfang allerede dokumenterede forskelle i plantediversitet mellem hegn på økologiske og konventionelle brug afspejles i blomstringsperiodens længde og i de ukrudtsfrø, der er tilgængelige for dyrelivet. (WP 4)
4) Den genetiske diversitet og populationsstrukturen hos en række vilde arter i agerlandet på økologiske brug for derigennem at belyse betydningen af økologiske brug som refugium for arter og som genetisk ressource. Det antages at habitater i hegn, markkanter og økologiske marker fungerer som ’øer’ og ’korridorer’ for den vilde flora og fauna og at brugen af pesticider i konventionelt jordbrug forårsager hyppige og gentagne lokale episoder af uddøen og rekolonisering for ukrudt og insekter, hvorved fugle og små pattedyr, der lever af disse indirekte påvirkes. Effekten på den genetiske struktur varierer sandsynligvis afhængig af arten og dens spredningspotentiale. Projektet fokusere på følgende tre arter: agerhøne (Perdix perdix), løbebillen (Pterostichus spp.) og markmus (Microtus agrestis), der varierer med hensyn til spredningspotentiale. (WP 5 og 6)
5) Indvirkningen af økologiske brug på indholdet af den vilde flora og fauna i landskabet ved at skabe et sæt model-landskabskonfigurationer. Dette udføres ved at benytte informationerne fra WP2 om graden af dyrknings-intensivitet i øko-brugene og deres fordeling samt fra WP3-6 om arternes økologi, genetiske mønster og arts-diversitet. Modellering vil benyttes til at syntetisere denne information og skabe et sæt index, der beskriver de økologiske brugs evne til at understøtte en række arter i den vilde flora og fauna. Resultatet bliver et index for den vilde flora og fauna for landskabet (landscape wildlife index, LWI), der vil have potentialet til at kunne benyttes af interessegrupper til at bestemme, hvilke scenarier der vil give et optimalt vildtpotentiale set fra deres synsvinkel. Fra en faunistisk synsvinkel vil indexet også være en effektiv måde til at måle ”naturkvalitet” ved at gøre det muligt at komme med et direkte estimat for hvor meget økologiske brug bidrager til værdien af landskabet for en række agerlands-arter. På denne måde kan en økologisk landmand vurdere hvilken påvirkning han som individ har på biodiversiteten via de valg han tager med hensyn til intensiteten hvormed han driver sin bedrift (WP7).
Bilag C: IGANGVÆRENDE RELEVANTE PROJEKTER I UDLANDET
Sustainable Arable Farming for an Improved Environment (SAFFIE). HGCA (Home-Grown Cereals Authority) Research and Development. Projectnumber HGCA 2617/LK0926.
Projektet startede 1. jan. 2002 og forventet varighed er 60 måneder.
Formålet med projektet er at øge agerlandets biodiversitet ved at integrere nye habitat-plejende tilgange til de dyrkede og udyrkede rande, for at udvikle mere bæredygtig landbrug. Øget forståelse af interaktionerne vil medføre stigninger i forekomster af leddyr og ukrudtsfrø og deres tilgængelighed vil være til fordel for agerlandets fugle.
Foreløbige resultater er bl.a:
1. Hvedemarker med ubevoksede refugier i marken medfører flere lærkeunger.
2. Udlægning ar randzoner øger biodiversiteten.
3. Ubevoksede refugier sammen med udlægning af randzoner medfører færre lærker, muligvis pga. øget prædation.
SEAMLESS, an EU FP6 Integrated Project, aims at generating an integrated framework of computer models. This framework can be used for assessment of how future alternative agricultural and environmental polices affect sustainable development in Europe.
SEAMLESS will develop an integrated and working framework called SEAMLESS-IF. This computer system will include models that simulate effects on the environment and on economic developments, next to procedures that enable assessment the social aspects of sustainability.
The SEAMLESS integrated project aims at developing a computerized, integrated and working framework (SEAMLESS-IF) to assess and compare, ex-ante, alternative agricultural and environmental policy options, allowing:
1. Analysis at the full range of scales (farm to EU and global), whilst focusing on the most important issues emerging at each scale;
2. Analysis of the environmental, economic and social contributions of a multifunctional agriculture towards sustainable rural development and rural viability;
3. Analysis of a broad range of issues and agents of change, such as climate change, environmental policies, rural development options, effects of an enlarging EU, international competition and effects on developing countries.
The project is coordinated by Wageningen University, The Netherlands.
Project duration 2005 – 2008. Project coordinator Dr. Martin van Ittersum, Wageningen University. Dansk deltager Skov- og Landskab v. Erling Andersen (link: http://www.seamless-ip.org/).
SENSOR is an Integrated Project within the 6th Framework Research Programme of the European Commission. 33 research partners from 15 European countries constitute the consortium, which develops science based ex-ante Sustainability Impact Assessment Tools (SIAT) to support decision making on policies related to multifunctional land use in European regions.
SENSOR directly responds to the European sustainability objectives as applied to land use and regional development. SENSOR’s main product is the Sustainability Impact Assessment Tool (SIAT), tailored to meet the needs of analysts and policy makers at the European level. SIAT will allow the assessment of land-use-related policy effects on sustainability by means of (1) European policy scenario analyses, (2) regional threshold assessments and (3) stakeholder targets identification. Making use of macro-econometric and sectoral land use models and impact indicators, European policy scenario analyses will forecast future land use changes, assess their multifunctional interrelations and economic, social and environmental impacts. To assess multifunctional land use effects at regional level, impact indicators will be verified on the basis of sustainability thresholds and targets derived from expert consultations and regional stakeholders, thus complementing the data-driven scenario analyses. Validation of assessment results will be conducted in case study areas of sensitive regions such as mountains, coastal zones, islands and post-industrialised areas across Europe. From the onset of the SIAT design, user requirements are explicitly taken into account. Danske deltagere: Danish Centre for Forest, Landscape and Planning; DK:: DIAS; Department of Agricultural Systems, Danish Inst. of Agricultural Sciences (link: http://www.sensor-ip.org/).
Bilag D: Navne på planter og dyr
Planter
| Videnskabeligt navn | Engelsk navn | Dansk navn |
| Acer pseudoplatanus L. | Great Maple | Ahorn |
| Achillea millefolium L. | Alm. Røllike | |
| Aethusa cynapium L. | Fool's Parsley | Hundepersille |
| Agrostis stolonifera L. | Krybende Hvene | |
| Anagallis arvensis L. | Scarlet Pimpernel | Rød Arve |
| Aphanes arvensis L. | Parslet Piert | Alm. Dværgløvefod |
| Apera spica-venti (L.) | Loose Silkybent | Alm. Vindaks |
| Arctium minus Bernh. | Liden Burre | |
| Arenaria serpyllifolia L. | Thyme-leaved Sandwort | Alm. Markarve |
| Arrhenatherum elatius (L.) | Draphavre | |
| Artemisia vulgaris L. | Grå Bynke | |
| Atriplex patula L. | Common Orache | Svine-Mælde |
| Beta vulgaris (L.) ssp. vulgaris | Beet | Roe |
| Betula sp. | Birch | Birk |
| Bilderdykia convolvulus (L.) Dumort. | Black Bindweed | Snerle-Pileurt |
| Brassica | Kål | |
| Brassica napus L. ssp. napus | Rape | Raps |
| Capsella bursa-pastoris (L.) Medicus | Shepherd's Purse | Hyrdetaske |
| Centaurea cyanus L. | Kornblomst | |
| Chaenorrhinum minus (L.) Lange | Small Toadflax | Liden Torskemund |
| Chamomilla suaveolens (Pursh) Rydb. | Rayless Mayweed | Skive-Kamille |
| Chenopodium album L. | Fat Hen | Hvidmelet Gåsefod |
| Chrysanthemum leucanthemum L. | Hvid Okseøje | |
| Cirsium arvense (L.) Scop. | Creeping Thistle | Ager-Tidsel |
| Cirsium vulgare | Bull Thistle | Horse-Tidsel |
| Cynosurus cristatus L. | Alm. Kamgræs | |
| Dactylis glomerata L. | Alm. Hundegræs | |
| Echium vulgare L. | Slangehoved | |
| Elymus repens (L.) Gould | Couch-grass | Alm. Kvik |
| Epilobium sp. | Willow-herb | Dueurt |
| Epilobium hirsutum | Lådden Dueurt | |
| Equisetum arvense L. | Common Horsetail | Ager-Padderok |
| Euphorbia exigua L. | Dwarf Spurge | Liden Vortemælk |
| Euphorbia helioscopia L. | Sun Spurge | Skærm-Vortemælk |
| Euphorbia peplus L. | Petty Spurge | Gaffel-Vortemælk |
| Festuca pratensis Hudson | Engsvingel | |
| Festuca rubra L. | Red Fescue | Rød Svingel |
| Fumaria officinalis L. | Common Fumitory | Læge-Jordrøg |
| Galeopsis tetrahit L. | Common Hemp-nettle | Alm. Hanekro |
| Galium aparine L. | Cleavers | Burre-Snerre |
| Geranium pusillum L. | Small-flowered Cranesbill | Liden Storkenæb |
| Heracleum sphondylium | Alm. Bjørneklo | |
| Holcus lanatus L. | Fløjlsgræs | |
| Hordeum vulgare L. | Barley | Byg |
| Kickxia elatine (L.) Dumort. | Fluellen | Spydbladet Torskemund |
| Continues |
| Planter | ||
| Videnskabeligt navn | Engelsk navn | Dansk navn |
| Knautia arvensis | Blåhat | |
| Lamium amplexicaule L. | Henbit | Liden Tvetand |
| Lamium hybridum Vill. | Cut-leaved Dead-nettle | Fliget Tvetand |
| Lamium purpureum L. | Red Dead-nettle | Rød Tvetand |
| Lapsana communis L. | Nipplewort | Haremad |
| Leucanthemum vulgare Lam. | Hvid Okseøje | |
| Lolium perenne L. | Rye-grass | Alm. Rajgræs |
| Matricaria perforata Merat | Scentless Mayweed | Lugtløs Kamille |
| Myosotis arvensis (L.) Hill | Common Forget-me-not | Mark-Forglemmigej |
| Papaver rhoeas L. | Field Poppy | Korn-Valmue |
| Phleum pratense L. | Engrottehale | |
| Plantago major L. | Rat-tail Plantain | Glat Vejbred |
| Poa annua L. | Annual Poa | Enårig Rapgræs |
| Poa trivialis L. ssp. trivialis/ Poa pratensis L. |
Rough-stalked/Smooth-stalked Meadow-grass | Alm./Eng-Rapgræs |
| Polygonum aviculare L. | Knotgrass | Vej-Pileurt |
| Polygonum convolvulus L. | Black bind-weed | Snerle-Pileurt |
| Polygonum lapathifolium L. | Pale Persicaria | Bleg Pileurt |
| Polygonum persicaria L. | Persicaria | Fersken-Pileurt |
| Ranunculus repens L. | Creeping Buttercup | Lav Ranunkel |
| Rubus fruticosus L. | Bramble | Brombær |
| Rumex crispus L. | Curled Dock | Kruset Skræppe |
| Sambucus nigra L. | Elder | Alm. Hyld |
| Scleranthus annuus L. | Enårig Knavel | |
| Senecio vulgaris L. | Groundsel | Alm. Brandbæger |
| Silene noctiflora L. | Night-flowering Campion | Nat-Limurt |
| Sinapis alba L. | Gul Sennep | |
| Sinapis arvensis L. | Wild Mustard | Ager-Sennep |
| Solanum nigrum L. ssp. nigrum | Black Nightshade | Sort Natskygge |
| Sonchus asper (L.) Hill | Spiny Milk-Thistle | Ru Svinemælk |
| Sonchus oleraceus L. | Milk-Thistle | Alm. Svinemælk |
| Stachys arvensis L. | Field Woundwort | Ager-Galtetand |
| Stellaria media (L.) Vill. | Chickweed | Fuglegræs |
| Tanacetum vulgare L. | Tansy | Rejnfan |
| Taraxacum sp. L. | Dandelion | Mælkebøtte |
| Trifolium | Kløver | |
| Trifolium repens L. | White Clover | Hvid-Kløver |
| Triticum aestivum L. | Wheat | Alm. Hvede |
| Trifolium pratense | Red Clover | Rød-Kløver |
| Urtica urens L. | Small Nettle | Liden Nælde |
| Veronica agrestis L. | Field Speedwell | Flerfarvet Ærenpris |
| Veronica persica Poiret | Large Field Speedwell | Storkronet Ærenpris |
| Veronica arvensis L. | Wall Speedwell | Mark-Ærenpris |
| Veronica hederifolia L. | Ivy-leaved Speedwell | Vedbend-Ærenpris |
| Viola arvensis Murray | Field Pansy | Ager-Stedmoderblomst |
Leddyr
| Arthropod klasse/orden | Lavere taksonomisk niveau | ||
| Videnskabeligt navn | Videnskabeligt navn | Engelsk navn | Dansk navn |
| Araneae | Spiders | Edderkopper | |
| Linyphiidae | Tæppespindere | ||
| Batyphantes gracilis | |||
| Erigone atra | |||
| Erigone dentipalpis | |||
| Leptyphantes | Småbaldakinspindere | ||
| Leptyphantes tenuis | |||
| Milleriana inerrans | |||
| Oedothorax | |||
| Oedothorax apicatus | |||
| Meioneta rurestris | |||
| Lycosidae | Jagtedderkopper | ||
| Pardosa agrestis | |||
| Opiliones | Harvestmen | Mejere | |
| Chilopoda | Centripedes | Skolopendre | |
| Diplopoda | Millipeds | Tusindben | |
| Collembola | Springtails | Springhaler | |
| Hemiptera | Næbmundede | ||
| Auchenorrhyncha | Cicada | Cikader | |
| Aphididae | Aphids | Bladlus | |
| Rhopalosiphum padi | Bird-cherry oat aphid | Havrebladlus | |
| Sitobion avenae | English grain aphid | Kornbladlus | |
| Heteroptera | Bugs | Tæger | |
| Calocoris norvegicus | Toplettet bederoetæge | ||
| Lepidoptera | Butterflies and moths | Sommerfugle | |
| Coenonympha pamphilus | Okkergul Randøje | ||
| Lasiommata megera | Vejrandøje | ||
| Maniola jurtina | Græsrandøje | ||
| Pieris napi | Grønåret Kålsommerfugl | ||
| Pieris rapae | Lille Kålsommerfugl | ||
| Thymelicus lineola | Stregbredpande | ||
| Diptera | Flies | Tovinger | |
| Tipulidae | Crane flies | Stankelben | |
| Syrphidae | Hover flies | Svirrefluer | |
| Neuroptera | Lacewings | Netvinger | |
| Chrysopidae | Green lacewing | Guldøjer | |
| Coleoptera | Beetles | Biller | |
| Carabidae | Ground beetles / Carabids | Løbebiller | |
| Anchomenus dorsalis | Spraglet Kvikløber | ||
| Bembidion | Brassy ground beetles | Glansløbere | |
| Bembidion lampros | Hedeglansløber | ||
| Harpalus rufipes | Håret Markløber | ||
| Pterostichus cupreus | Strawberry ground beetles | Bred metaljordløber | |
| Pterostichus melanarius | Strawberry ground beetles | Markjordløber | |
| Pterostichus niger | Skovjordløber | ||
| Trechus | A ground beetle genus | Grotteløbere | |
| Trechus quadristriatus | Markgrotteløber | ||
| Staphylinidae | Rove beetles | Rovbiller | |
| Tachyporus | Agerrovbiller | ||
| Silphidae | Carrion beetles | Ådselsbiller | |
| Scarabaeidae | Chafers and dung beetles | Gødningsbiller (torbister) | |
| Elateridae | Click beetles | Smældere | |
| Lathridiidae | Lathridiids | ||
| Cantharidae | Soldier beetles | Blødvinger | |
| Nitidulidae | Blossom beetles | Glansbiller | |
| Meligethes aeneus | Blossom beetle | Glimmerbøsse | |
| Cryptophagidae | Mould beetles | ||
| Coccinellidae | Ladybirds | Mariehøns | |
| Chrysomelidae | Leaf beetles | Bladbiller | |
| Gastrophysa polygoni | Knotgrass beetle | Pileurtbladbille | |
| Oulema | A leaf beetle genus | Bladbilleslægt | |
| Oulema melanopus | Cereal leaf beetle | Alm. kornbladbille | |
| Curculionidae | Weevils | Snudebiller | |
| Ceutorhynchus | A weevil genus | En snudebilleslægt | |
| Sitona lineatus | Bladrandbille | ||
| Hymenoptera | Årevingede | ||
| Apoidea | Bees | Bier | |
| Apis mellifera | Honey bee | Honnongbier | |
| Bombus | Bumble bees | Humlebier | |
| Bombus hortorum | Havehumle | ||
| Bombus lapidarius | Stenhumle | ||
| Bombus lucorum | Lille jordhumle | ||
| Bombus pascuorum | Agerhumle | ||
| Bombus pratorum | Markhumle | ||
| Bombus terrestris | Jordhumle | ||
| Symphyta | Savhvepse (bl.a. bladhvepse) |
||
| Tenthredinidae | Common sawflies | Egentlige bladhvepse | |
| Parasitica | Parasitic wasps | Snylte- og galhvepse | |
| Aphidius | Aphid parasitic wasps | Bladlussnyltehvepse | |
| Aphidius ervi | |||
| Aphidius rhopalosiphi | |||
| Aphelinus | |||
| Cotesia | |||
| Praon volucre |
Fugle
| Videnskabeligt navn | Engelsk navn | Dansk navn |
| Alauda arvensis | Skylark | Sanglærke |
| Alectoris rufa | Red-legged partridge | Rødhøne |
| Enberiza citrinella | Yellowhammer | Gulspurv |
| Falco tinnunculus | Kestrel | Tårnfalk |
| Fringilla coelebs | Chaffinch | Bogfinke |
| Passer montanus | Tree Sparrow | Skovspurv |
| Perdix perdix | Grey Partridge | Agerhøne |
| Phasianus colchicus | Phesant | Fasan |
| Turdus merula | Blackbird | Solsort |
| Turdus philomelos | Song Trush | Sangdrossel |
| Turdus viscivorus | Mistle Thrush | Misteldrossel |
| Tyto alba | Barn owl | Slørrugle |
Pattedyr
| Videnskabeligt navn | Engelsk navn | Dansk navn |
| Apodemus sylvaticus | Wood mouse | Skovmus |
| Micromys minutus | Harvest mouse | Dværgmus |
| Lepus eupopaeus | Brown hare | Brun hare |
| Clethrionomys glareolus | Bank Vole | Rødmus |
Krybdyr
| Videnskabeligt navn | Engelsk navn | Dansk navn |
| Lacerta agilis | Sand Lizard | Markfirben |
Version 1.0 Maj 2007 • © Miljøstyrelsen.