|
| Forside | | Indhold | | Forrige | | Næste |
Effekt af bekæmpelsesmidler på flora og fauna i vandløb
4 Effekter af pesticider på invertebrater i vandløb
Forsøgsdyrene blev udvalg således at de repræsenterede væsentlige elementer af den danske vandløbsinvertebratfauna. 70-80% af de taxa der findes i vandløb er insekter, og derfor er insekterne tilsvarende
overrepræsenteret som forsøgsorganismer. De udsøgte organismer er desuden udvalgt så de varierer med hensyn til taksomisk gruppe, livscyklus, fødefunktionalitet, trofisk niveau og tolerance over for
organisk forurening. Sidstnævnte kriterium betyder at de undersøgte organismer er indplaceret bredt i Dansk Vandløbsfaunaindeks (DVFI) (Miljøstyrelsen, 1998), der bruges som biologisk
bedømmelsesmetode i det nationale overvågningsprogram (NOVA2003) på 1050 stationer samt flere steder i det regionale tilsyn. Derudover dækker de udvalgte forsøgsorganismer et bredt spektrum med
hensyn til tolerance over pesticider. Wogram & Liess (2001) har fundet følgende rangorden med hensyn til de i vore undersøgelser anvendte invertebraters pesticidfølsomhed: Slørvinger > krebsdyr >
døgnfluer > tovinger > vårfluer > snegle. Tabel 8 viser forekomst og densitet af hovedparten af de anvendte forsøgsorganismer i de 1050 NOVA 2003 overvågningsvandløb.
Tabel 8. Fundfrekvens og tæthed på 1050 NOVA 2003 stationer i 2001 af hovedparten af de i projektet undersøgte invertebrater. Det taksonomiske niveau der anvendes i NOVA2003 er som minimum det i DVFI foreskrevne (Miljøstyrelsen, 1998), men enkelte amter udfører en fuld artsbestemmelse. Derfor indeholder tabellen både DVFI niveauet samt et højere taksonomisk niveau (slægt og art) hvor dette har forekommet. Hvad dette betyder for tolkningen af tabellen kan eksemplificeres for Gammarus pulex: Gammarus er identifikationsniveauet i DVFI, men en del amter har bestemt til artsniveau (G. pulex). Hovedparten af de Gammarus der er fundet på de 81% af stationerne vil formentlig være G. pulex da den anden art, G. lacustris, er langt mindre udbredt i vandløb. Imidlertid vides det kun med sikkerhed at der er G. pulex på 49% af stationerne.
Taxa |
Fundet på antal stationer
(Frekvens) |
Densitet
(middel antal individer pr.
station) |
Gammarus
Gammarus pulex
Asellus
Baetidae
Baetis rhodani
Leuctra
Isoperla
Isoperla grammatica
Hydropsyche
Glossosomatidae
Agapetus fuscipes
Sericostomatidae
Sericostoma personatum
Chironomus
Chironomus riparius |
848 (81%)
519 (49%)
658 (63%)
766 (73%)
317 (30%)
125 (12%)
73 (7%)
20 (2%)
356 (34%)
24 (2%)
9 (<1%)
145 (14%)
94 (9%)
63 (6%)
10 (1%) |
200
242
50
84
85
12
11
26
18
25
36
8
7
24
72 |
4.1 Anvendte invertebrater
Gammarus pulex, ferskvandstangloppen, er et krebsdyr (Malacostraca; Amphipoda). Denne organisme blev primært valgt på grund af dens gode laboratoriegenskaber, samt dens store udbredelse i
danske vandløb (Tabel 8). Ifølge undersøgelser foretaget af Fyns Amt (2000) udgør G. pulex ikke mindre end 25% af den samlede smådyrsfauna i vandløb og kan antage en tæthed på hen ved 8.000
individer pr. m². Dens hyppige forekomst betyder da også, at den udgør et meget vigtigt fødegrundlag for især fisk og fugle. Den udmærker sig desuden ved at være velbeskrevet i litteraturen, hvorfor
sammenligningsgrundlaget for forsøg med G. pulex udsat for andre miljøfremmede stoffer er stort. G. pulex er et væsentligt element i DVFI, hvorfor den tillægges stor økologisk betydning i forbindelse med
kvalitetsbedømmelse af danske vandløb. G. pulex vil findes både i rene og moderat forurenede vandløb. Den indgår i Nøglegruppe 3 og som positiv diversitetsgruppe i DVFI og er meget væsentlig for at
kunne at skelne de let og moderat forurenede vandløb fra de stærkere forurenede vandløb.
Fødefunktionelt er G. pulex meget plastisk men betegnes overvejende som en ituriver/samler, der primært ernærer sig ved nedfaldne blade og andet dødt organisk materiale. G. pulex opholder sig ved
bunden eller brinkerne, hvor den søger føde. G. pulex er meget mobil og findes hyppigt i drift. Desuden udviser den meget udpræget opstrøms migration (positiv rheotaxi) for at kompensere for driften. G.
pulex er derfor en god kolonisator, sålænge den ikke møder barrierer mod opstrømsmigration i form af f.eks. rørunderføringer ved veje. G. pulex udgør en nøglerolle i danske vandløb, da den kvantitativt er
den mest dominerende invertebrat. Den udgør fødegrundlag for højere trofiske niveauer og har desuden en nøglerolle som bio-ingeniører i omsætningen af groft organisk stof til fint. Det fine organiske stof
indgår som et væsentligt element i detritivorfødenettet, og har stigende betydning ned gennem vandløbet.
Asellus aquaticus er som Gammarus et krebsdyr (Malacostraca; Isopoda). Hovedincitamentet for anvendelse af A. aquaticus er, at den ligesom G. pulex er vidt udbredt i de fleste vandløb. Asellus
indtager en vigtig rolle i DVFI, blot med den forskel at dens forekomst anses at indikere en høj forureningsgrad. Fødefunktionelt betegnes den som ituriver/samler, hvorfor den i lighed med G. pulex udfylder
en vigtig post som omsætter af organisk materiale. Naturligt forekommer A. aquaticus blandt makrofyter i kantzonen i større vandløb. Imidlertid vil A. aquaticus findes i flere vandløbstyper, når disse er
påvirkede af organisk forurening. A. aquaticus findes på 63% af NOVA2003 stationerne (Tabel 8), hvilket bl.a. afspejler at halvdelen af disse vandløb ikke har en tilfredsstillende biologisk vandløbskvalitet
(faunaklasse 4 eller lavere). A. aquaticus opholder sig primært på bunden af et vandløb, hvor den kan forekomme i meget store individantal ( 10.000 m²) i stærkt belastede vandløb.
Isoperla er en slørvinge (Plecoptera) tilhørende familien Perlodidae. Isoperla tilhører gruppen Setipalpia, der er prædatorer. Isoperla er valgt fordi den befinder sig på et højere trofisk niveau i forhold til
andre invertebrater der er indgået i forsøgsserierne, og fordi det er en meget rentvandskrævende organisme, der indgår i DVFI som Nøglegruppe 1 taxsa og positiv diversitetsgruppe. Isoperla grammatica
findes i hele landet, mens den anden danske art, I. difformis, kun er fundet i Jylland. Isoperla er fundet i 7% af NOVA 2003 vandløbene (Tabel 8). De lever ikke i de mindste vandløb, men er ellers udbredt
i hele vandløbskontinuumet sålænge vandkvaliteten tillader det. Isoperla har en 1-årig livscyklus og emergerer i maj-juni.
Leuctra er slørvinger (Plecoptera; Leuctridae), der lever af at samle fint organisk stof (FPOM) og af at iturive blade og lign. (CPOM). Leuctra er primært knyttet til de mindre vandløb og de er forholdsvis
almindelige (Tabel 8). Der er 4 arter i danske vandløb og de kræver alle gode iltforhold. Leuctra indgår således i Nøglegruppe 1 og tæller som positiv diversitetsgruppe i DVFI. De fleste Leuctra, men især
Leuctra nigra, er imidlertid tolerante overfor lav pH i det omgivende vand. Alle Leuctra arterne har en 1-årig livscyklus med undtagelse af L. nigra, der har en 2-årig livscyklus. To arter emergerer i
april-maj (hippopus og nigra) og to arter september-oktober (fusca og digitata). Der vil derfor være store Leuctra nymfer i vandløbene det meste af året såfremt vandkvaliteten tillader det.
Baetis (Ephemeroptera; Baetidae) er en meget udbredt døgnflueslægt i danske vandløb (Tabel 8). Den vil forekomme fra de øvre kildebække til store vandløb. Baetis ernærer sig som skraber af alger og
biofilm på sten og makrofyter. Desuden ernærer Baetis sig som samler af FPOM. Der er 10 arter i Danmark, hvoraf B. rhodani og B. vernus er de mest udbredte. B. rhodani er forholdsvis tolerant overfor
forringede iltforhold sålænge vandet strømmer og kan derfor være hyppig i moderat forurenede vandløb. B. rhodani har to generationer om året, mens nogle af de øvrige Baetis-arter formentlig er 1-årige.
Sericostoma personatum (Trichoptera; Sericostomatidae) er en typisk ituriver i skovvandløb. Den vil kunne forekomme i mere åbne vandløb sålænge temperaturen ikke bliver for høj. S. personatum lever
i områder med lav strømhastighed, og er derfor særlig følsom for lavt iltindhold i det omgivende vand. Sericostomatidae indgår i Nøglegruppe 1 og tæller som en positiv diversitetsgruppe i DVFI.
Sericostomatidae er af de undersøgte organismer den mest hyppigt forekommende Nøglegruppe 1 organisme i NOVA 2003 vandløbene (Tabel 8). S. personatum har en 2-årig livscyklus og emergerer om
foråret.
Agapetus fuscipes er en vårflue (Trichoptera; Glossosomatidae), der lever af at skrabe alger og andet biofilm af sten. Findes i Danmark primært i mindre vandløb men er forholdsvis sjælden (Tabel 8). A.
fuscipes kræver gode iltforhold og familien Glossosomatidae indgår både i Nøglegruppe 1 og som positiv diversitetsgruppe i DVFI. A. fuscipes har en 1-årig livscyklus og kan emergere en stor del af året.
Hydropsyche (Trichoptera; Hydropsychidae) har netspindende og fritlevende larver, der meget udbredte i danske vandløb (Tabel 8). Den ernærer sig ved at spinde et silkenet hvormed den fanger forskellige
suspenderede partikler som tranporteres med vandet. Pga. denne fødestrategi er Hydropsyche afhængig af groft, stabilt substrat og en kontinuerlig vandstrøm. Desuden er tætheden af Hydropsyche
afhængig af den mængde føde, der føres med vandløbet. Hydropsyche vil typisk findes på de mellemste vandløbsstrækninger, og kan findes i masseforekomst i søafløb, hvor de lever af de planktoniske
organismer o. lign. der kommer fra søen. Hydropsyche er generelt tolerante overfor dårlig vandkvalitet og vil kunne findes i moderat forurenede vandløb. Der 7 arter af slægten Hydropsyche i Danmark,
hvoraf en art (H. augustipennis) er udbredt og almindelig. Hydropsyche-arterne er formentlig alle 1-årige og flere har en lang flyveperiode, der strækker sig fra maj til august.
Chironomus riparius er en dansemyg (Chironomidae) tilhørende underfamilien Chironomini. Som repræsentant for organismer med en fuldstændig forvandling og dermed også et terrestrisk livsstadie blev
C. riparius valgt. Arten er naturligt forekommende i danske vandløb, men udmærker sig specielt ved at være egnet til at holde i laboratoriekultur, bl.a. fordi den har en meget kort generationstid. Sidstnævnte
forhold betyder også at arten er særligt velegnet til gennemførsel af komplette livscyklus tests. Efter forgæves at have forsøgt at indsamle Chironomus fra felten blev vi i stedet nødt til at importere C.
riparius fra en allerede eksisterende laboratoriekultur (se senere beskrivelse). Ifølge DVFI anses forekomst af Chironomus, i lighed med Asellus, at indikere en høj organisk forureningsgrad. Der er således
tale om en meget forureningstolerant art, der tåler endog meget lave iltkoncentrationer. Vi fandt det imidlertid relevant at undersøge om denne tilsyneladende høje tolerancetærskel ligeledes gør sig gældende
overfor forurening med pesticider. C. riparius er et udpræget bunddyr, der lever både i og af sediment, hvorfor den fødefunktionelt betegnes som en samler. Der er i visse tilfælde observeret forekomster på
op til 50.000 individer pr. m². Imidlertid har de sidste årtiers spildevandsrensning bevirket, at Chironomus ikke længere er ret udbredt i danske vandløb og de blev således kun fundet på 6% af NOVA
2003 stationerne i 2001.
Prodiamesa olivacea hører til underfamilien Prodiamesinae (familie: Chironomidae), der er en lillle underfamilie med få slægter og arter i Danmark. P. olivacea lever i sedimentet i vandløb. Den kræver
forholdsvis gode iltforhold og sedimenter med stort indhold af organisk stof, som den ernærer sig af (sedimentæder). Den kan være meget talrig i moderat forurenede vandløb med store forekomster af fint
organisk stof, men vil ikke findes i stærkt forurenede vandløb som f.eks. Chironomus.
Bithynia tentaculata er en forgællesnegl tilhørende familien Hydrobiidae. Da vi ikke kunne få indsamlet et tilfredsstillende stort antal individer af huesneglen Ancylus fluviatilis blev denne almindeligt
forekommende sumpsnegl anvendt som repræsentant for de græssende organismer. Den blev primært valgt udfra praktiske hensyn, idet den optrådte i stort antal på indsamlingslokaliteten. Den var således
en meget hyppig bifangst ved indsamling af de øvrige organismer. B. tentaculata er ikke omfattet af DVFI, men det betyder ikke at der er tale om en økologisk ubetydelig art, idet den, med dens store
individtæthed taget i betragtning, kan opfylde en vigtig rolle som græsser og skraber af diverse algetyper. B. tentaculata skraber biofilm af makrofyter og var i indsamlingsvandløbet især knyttet til Elodea
(vandpest). Vi fandt det desuden relevant at vælge en art med en meget begrænset og langsom bevægelsesadfærd, der bevirker at den ikke umiddelbart kan forlade et forurenet område.
4.2 Standardtests til bestemmelse af akut toksicitet (LC50)
Alle de arter som blev udvalgt til screeningtests er almindeligt forekommende vandløbsinvertebrater i Danmark, og de fleste spiller en vigtig rolle i DVFI til bedømmelse af et vandløbs forureningsgrad.
Kriterierne for udvælgelsen af de respektive pesticider, samt ved hvilket koncentrationsniveau forsøgene skulle gennemføres er opsummeret i nedenstående:
- Pesticiderne blev udvalgt i samarbejde med Miljøstyrelsen på baggrund af deres anvendelse, virkemekanisme, samt deres effekt på standard testorganismer.
- Eksponeringskoncentrationer blev valgt på baggrund af en kombination af følgende: worst-case PECs fra Tier 1 eksponeringsscenarier (f.eks., højeste applikationsrate ved direkte oversprøjtning på
en 30 cm dyb vandoverflade); toksicitet overfor standard testorganismer (bl.a. Daphnia), værdier tidligere fundet i danske vandløb, samt i flere tilfælde på baggrund af udførte pilotforsøg.
Selvom de tilgængelige målinger over pesticidforekomster i danske vandløb sandsynligvis ikke repræsenterer den maksimale koncentration som kan forekomme i pågældende vandløb, er det værd at notere,
at koncentrationer på over 2000 ng l-1 er yderst sjældne for noget pesticid (DMU 2002). Ikke desto mindre inkluderede alle screeningtests koncentrationer på minimum op til 2000 ng l-1, og i flere tilfælde
endda en faktor 1000 højere. For mange af arterne var det imidlertid ikke muligt at bestemme en LC50-værdi på grund af lav eller slet ingen dødelighed ved den højeste koncentration. Hvis der ikke blev
observeret dødelighed ved 2000 ng l-1 eller derover i løbet af en kontinuerlig eksponeringsperiode på 24 timer (eller længere) blev det antaget, at det pågældende pesticid ikke at havde en målbar effekt ved
kortere pulseksponeringer med lignende koncentrationer. Derfor blev de pågældende art-pesticid kombinationer nedprioriteret med henblik på yderligere forsøg.
Som det vil fremgå af resultaterne fra de udførte screeningstests viste Gammarus pulex sig konsekvent at være den mest sensitive af alle de testede arter. Af alle de testede pesticider var esfenvalerat klart
det mest toksiske overfor ferskvandsinvertebrater efterfulgt af azoxystrobin. Ingen af de øvrige pesticider, der blev testet, havde målbare effekter på arter testet ved miljørealistiske koncentrationer.
I bestræbelsen på at bestemme effekterne af miljørealistiske pulsekponeringer på andre endpoints end dødelighed, blev G. pulex herefter valgt som nøgleorganisme for videre studier til bestemmelse af
følsomheden af forskellige livsstadier.
Af de screenede arter var det udelukkende muligt (indenfor dette projekts begrænsede tidsramme) at udføre fulde livscyklusstudier med Chironomus riparius. En yderligere argumentation for at fokusere på
C. riparius er, at det er den eneste sedimentlevende invertebrat som regelmæssigt (under kronisk eksponering) bliver anvendt til risikovurdering af pesticider.
4.2.1 Metode
Der blev udført screeningstests til bestemmelse af LC50-værdier med 5 forskellige arter udsat for 5 forskellige pesticider. De anvendte arter og pesticider fremgår af oversigtsskemaet under resultatafsnittet.
Som nævnt indledningsvis viste den mest følsomme organisme sig at være G. pulex, hvorfor fokus under den følgende metode- og resultatgennemgang er lagt på denne art.
Gammarus pulex
Dette forsøg med G. pulex blev udført med henblik på bestemmelse af LC50-værdier af esfenvalerat (24h, 48h og 96h) for både voksne (kropslængde = 10-14 mm) og juvenile (kropslængde = 7-8 mm)
individer ved hjælp af et statisk testdesign. De nominelle testkoncentrationer var 0, 0,01, 0,05, 0,1, 0,5, 1,0 og 2,0 µl-1 og således dækkende for et miljørealistisk koncentrationsniveau, idet de højeste
koncentrationer målt i danske åer er 0,2 og 0,66 µl-1 (Fyns Amt 1999 og Wiggers 1999). Det ønskede antal testorganismer blev indsamlet fra Værebro Å og opbevaret i laboratoriekultur (se detaljeret
beskrivelse i senere afsnit) minimum 14 dage inden forsøgsstart. Der blev anvendt 3 replikater af hver 10 individer pr. testkoncentration. Testkamrene bestod af glaspetriskåle, der forudgående var blevet
mættet med den respektive testkoncentration, indeholdende 120 ml vand. Dyrene blev fodret dagligt med små bladskiver (diameter = 18 mm), der normalt blev konsumeret indenfor ét døgn. En afmålt
fødemængde blev anvendt at for at minimere overfladeabsorption af pesticidet. Både immobilitet og dødelighed blev observeret efter 1, 24, 48 og 96 timers eksponering. Voksne individer blev eksponeret
som par (5 hanner/5 hunner pr replikat), og der blev ligeledes gjort observationer over antallet af par der blev forstyrret af eksponeringen. Herudover noteredes eventuelle frigivelser af unger fra hunner som
følge af eksponeringen. De øvrige 4 pesticider blev testet efter samme forsøgsmetode.
Asellus aquaticus
Det ønskede antal testorganismer af denne art blev indsamlet i Værebro Å og efterfølgende opbevaret i laboratoriekultur og pesticideksponeret til de forskellige stoffer som beskrevet under G. pulex.
Kropslængden af de anvendte individer var som følger: voksne = 8-12 mm, juvenile = 2-4 mm.
Bithynia tentaculata
Det ønskede antal testorganismer af denne art blev ligeledes indsamlet i Værebro Å og efterfølgende opbevaret i laboratoriekultur og pesticideksponeret til de forskellige stoffer som beskrevet under G.
pulex. Skallængden af de anvendte individer var som følger: voksne = 10-12 mm, juvenile = 3-5 mm.
Chironomus riparius
Testorganismer af denne art blev indsamlet fra egen laboratoriekultur og efterfølgende pesticideksponeret til de forskellige stoffer som beskrevet under G. pulex. Kropslængden af de anvendte individer var
som følger: store = 10-12 mm (4. instar), små (2. instar) = 1-3 mm.
Prodiamesa olivacea
Testorganismer af denne art blev med assistance og anvisning fra C. Lindegaard (Ferskvandsbiologisk Laboratorium, Hillerød) indsamlet i Gadevang-renden og efterfølgende opbevaret i laboratoriekultur
som beskrevet under C. riparius. Pesticideksponeringen til de forskellige stoffer fulgte metoden som er beskrevet under G. pulex. Da det ikke lykkedes at kultivere denne art blev der udelukkende udført
forsøg med fuldvoksne larver med en kropslængde på 10-12 mm.
Statistik
LC50-værdier ( 95% konfidensintervaller) for 24, 48 og 96 timers eksponering blev estimeret ved logistisk regression ved hjælp af SYSTAT ver. 10 (SPSS, Chicago, IL).
4.2.2 Resultater
Som det fremgår af oversigtstabellerne (Tabel 9 og 10) for alle de testede arter og pesticider var både voksne og juvenile individer af G. pulex mest følsomme, specielt overfor esfenvalerat, hvorfor der
primært fokuseres på beskrivelse af data fra disse screeningstests. En lang række af de øvrige screeninger gav ikke anledning til forøget mortalitet hos de testede organismer, selv ikke ved urealistisk høje
koncentrationsniveauer, hvorfor disse ikke kommenteres yderligere.
Gammarus pulex eksponeret til esfenvalerat
Alle 30 voksne individer i alle eksponeringsgrupper (0, 0,01, 0,05, 0,1, 0,5, 1,0 og 2,0 µg l-1) var stadigt levende efter 1 times eksponering til esfenvalerat. I kontrolgruppen var 15 ud af 15 par stadigt
intakte, mens kun 4 ud af 15 par stadig var intakte i gruppen eksponeret til 0,01 µg l-1. Der var ingen intakte par tilbage i koncentrationer højere end 0,1 µg l-1 efter 1 times eksponering. Alle voksne individer
var stadig mobile i kontrolgruppen samt grupperne henholdsvis eksponeret til 0,01 og 0,05 µg l-1, mens 6 individer var immobile i gruppen eksponeret til 0,1 µg l-1 og alle individer i de resterende
eksponeringsgrupper var immobile. Unger blev frigivet fra hunner eksponeret til 0,1 µg l-1 og opefter.
Overlevelsen af voksne var faldende med eksponeringstid mellem 24 og 48 timer, men ikke mellem 48 og 96 timer. LC50-værdier blev estimeret til ca. 0,34 µg l-1 efter 24 timers eksponering og 0,14 µg l-1
efter 48 og 0,13 µg l-1 efter 96 timers eksponering. Hunnerne så ud til at være mere sensitive end hanner og forskellen var specielt synlig efter 24 timers eksponering (Tabel 11).
Tabel 9. Estimerede LC50-værdier (µg l-1) for voksne individer af krebsdyr og snegle. Testene med dansemyg blev gennemført med store individer (10-12 mm). Tallene i parenteserne angiver 95% konfidensintervaller.
| Forsøgsorganisme |
Esfen-
valerat |
Azoxy-strobin |
Diflubenz-
uron |
Pirimi-
carb |
Dime-
thoat |
Gammarus pulex
 |
LC50(24h) |
0,34
(0,31 – 0,38) |
244
(232 - 258) |
>200 |
>5000 |
>3000 |
| LC50(48h) |
0,14
(0,13 – 0,16) |
148
(139 - 159) |
>200 |
>5000 |
>3000 |
| LC50(96h) |
0,13
(0,12 – 0,14) |
72
(66 - 77) |
>200 |
>5000 |
>3000 |
Asellus aquaticus
 |
LC50(24h) |
6,89
(4,72-11,63) |
>1000 |
>3000 |
>5000 |
>3000 |
| LC50(48h) |
0,69
(0,58 – 0,85) |
>1000 |
>3000 |
>5000 |
>3000 |
| LC50(96h) |
0,52
(0,44 – 0,63) |
>1000 |
>3000 |
>5000 |
>3000 |
Chironomus ripar.
 |
LC50(24h) |
>2 |
435
(411 - 460) |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>2 |
395
(372 - 418) |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>2 |
306
(285 - 328) |
>5000 |
>5000 |
>5000 |
Prodiamesa oliva.
 |
LC50(24h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
Bithynia tentacul.
 |
LC50(24h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
Selvom antallet af unger der blev frigivet i løbet af den første time under eksponeringen ikke blev talt, blev disse observeret i live i grupperne op til 0,1 µg l-1, mens ungerne i grupperne 0,5, 1,0 og 2,0 µg l-1 alle var døde. I kontrollen var antallet af intakte par lavere efter 24 timer og resten af forsøgsperioden ud sammenlignet med 1 times eksponering. I modsætning hertil var der flere af de forstyrrede par i grupperne eksponeret til henholdsvis 0,01 og 0,05 µg l-1 der blev gendannet med tiden. Dette var imidlertid ikke tilfældet i de højere
eksponeringsgrupper.
Tabel 10. Estimerede LC50-værdier (µg l-1) for juvenile individer af krebsdyr og snegle. Testene med dansemyg blev gennemført med små individer (1-3 mm). Tallene i parenteserne angiver 95% konfidensintervaller.
| Forsøgsorganisme |
Esfen-
valerat |
Azoxy-strobin |
Diflubenz-
uron |
Pirimi-
carb |
Dime-
thoat |
Gammarus pulex
|
LC50(24h) |
0,24
(0,22 – 0,26) |
72
(67-78) |
>200 |
>5000 |
>3000 |
| LC50(48h) |
0,14
(0,13 – 0,15) |
10
(7-14) |
>200 |
>5000 |
>3000 |
| LC50(96h) |
0,14
(0,13 – 0,15) |
3
(1-5) |
>200 |
>5000 |
>3000 |
Asellus aquaticus
|
LC50(24h) |
6,16
(2,90 –5,19) |
>1000 |
>3000 |
>5000 |
>3000 |
| LC50(48h) |
0,75
(0,65 – 0,88) |
>1000 |
>3000 |
>5000 |
>3000 |
| LC50(96h) |
0,46
(0,39 – 0,53) |
>1000 |
>3000 |
>5000 |
>3000 |
Chironomus ripar.
|
LC50(24h) |
>2 |
681
(649-716) |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>2 |
546
(524-569) |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>2 |
395
(375-417) |
>5000 |
>5000 |
>5000 |
Prodiamesa oliva.
|
LC50(24h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>2 |
>1000 |
>5000 |
>5000 |
>5000 |
Bithynia tentacul.
|
LC50(24h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
| LC50(48h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
| LC50(96h) |
>10 |
>5000 |
>5000 |
>5000 |
>5000 |
Som det fremgår af resultatet af LC50-forsøg med eksponering af sammenhængende par viste den prækopulative fase sig at være yderst sensitiv overfor esfenvalerat. Derfor blev netop denne
forsøgsparameter underkastet yderligere opmærksomhed i de efterfølgende undersøgelser til bestemmelse af følsomheder i forskellige livsstadier hos Gammarus pulex.
Ligesom det var tilfældet for voksne individer blev der hos de juvenile observeret faldende overlevelse i tiden mellem 24 og 48 timer efter eksponeringen, men ikke mellem 48, og 72 eller 96 timer efter
eksponeringen. De juvenile individer var mere sensitive end de voksne individer med estimerede LC50-værdier på 0,24 µg l-1 efter 24 timer, 0,14 µg l-1 efter 48 og 96 timer. Ingen af de overlevende individer i gruppen eksponeret til 0,5 µg l-1 var aktive efter 24 timers eksponering i modsætning til gruppen eksponeret til 0,1 µg l-1, hvor ca. 50 % var aktive og gruppen eksponeret til 0,05 µg l-1 hvor ca. 75 % af de overlevende var aktive.
Tabel 11. Akut toksicitet af esfenvalerat på voksne Gammarus pulex. Der var 5 par voksne og 3 replikater per eksponeringsgruppe (n=30 pr gruppe).
Tid
(timer) |
Koncentra-
tion (µg l-1) |
Overlevende (hanner;
hunner) |
Levende men
immobile |
Intakte par |
| 1 |
0 |
15;15 |
0 |
15 |
| 0,01 |
15;15 |
0 |
4 |
| 0,05 |
15;15 |
0 |
0 |
| 0,1 |
15;15 |
6 |
0 |
| 0,5 |
15;15 |
30 |
0 |
| 1,0 |
15;15 |
30 |
0 |
| 2,0 |
15;15 |
30 |
0 |
| 24 |
0 |
15;15 |
0 |
9 |
| 0,01 |
15;15 |
0 |
3 |
| 0,05 |
15;13 |
2 |
0 |
| 0,1 |
13;12 |
19 |
0 |
| 0,5 |
10;1 |
16 |
0 |
| 1,0 |
6;2 |
8 |
0 |
| 2,0 |
0;0 |
- |
0 |
| 48 |
0 |
15;15 |
0 |
5 |
| 0,01 |
15;15 |
0 |
8 |
| 0,05 |
14;12 |
0 |
6 |
| 0,1 |
12;10 |
0 |
2 |
| 0,5 |
2;0 |
2 |
0 |
| 1,0 |
0;0 |
- |
0 |
| 2,0 |
0;0 |
- |
0 |
| 96 |
0 |
15;15 |
0 |
5 |
| 0,01 |
15;15 |
0 |
8 |
| 0,05 |
14;11 |
0 |
7 |
| 0,1 |
12;9 |
0 |
6 |
| 0,5 |
2;0 |
0 |
0 |
| 1,0 |
0;0 |
- |
0 |
| 2,0 |
0;0 |
- |
0 |
Mortalitet i strømrender
I driftundersøgelserne gennemført i recirkulerende strømrender (se afsnit 4.7) registreredes antal døde og overlevende individer ved forsøgenes afslutning svarende til 96 timers eksponering. For de
pesticider, der er testet overfor både krebsdyr (Gammarus, Asellus) og insektlarver (Baetis rhodani, Leuctra hippopus, Hydrosyche siltalai) og hvor der kunne beregnes en lethal koncentration var
krebsdyrene og især Gammarus de mest følsomme (Tabel 12).
Tabel 12. Sammenligning af LC50,96 timer, (ng l-1) hos krebsbyr og insektlarver i strømakvarier. * Pirimicarb udløste ingen dødelighed ved den højest anvendte koncentration (1 mg l-1).
| Pesticid |
Gammarus |
Asellus |
Baetis |
Leuctra |
Hydropsyche |
| Esfenvalerat |
21 |
29 |
120 |
90 |
440 |
| Sumi-alfa |
1,6 |
5 |
30 |
26 |
160 |
| Pirimicarb* |
>106 |
>106 |
>106 |
>106 |
>106 |
4.3 Effekt af eksponeringsvarighed
For at undersøge letale effekter af bekæmpelsesmidler i vandløb som funktion af eksponeringsvarigheden blev der udført laboratorieforsøg med invertebrater, primært Gammarus pulex, eksponeret for
forskellige pesticider. Den eksperimentelle tilgang har inkluderet undersøgelser af sammenhængen mellem eksponeringsvarigheden og toksicitet samt undersøgelser af eventuelle forsinkede effekter af
pesticider efter ophørt eksponering. Med stigende eksponeringsvarighed vil der ske et stigende optag af pesticid i organismen hvorfor effektkoncentrationen vil være faldende over tid. For let vandopløselige
stoffer vil en ligevægt mellem optag og effekt indstille sig relativt hurtigt, og herefter vil effektkoncentrationen teoretisk set være stabil.
Organismerne vil dog være udsat for et øget fysiologisk stress som funktion af eksponeringsvarigheden, hvorfor en forøget toksicitet kan forekomme efter at ligevægt mellem optag og effekt er indtruffet.
Lipofile stoffer optages langsommere end vandopløselige stoffer, og en ligevægt vil først indtræffe efter længere tids eksponering. Lipofile stoffer vil have tendens til at bioakkumulere i organismernes fedtvæv,
hvor koncentrationen vil overstige den i det omgivende miljø. Derfor kan toksiciteten af lipofile stoffer forventes at falde over en længere periode sammenlignet med vandopløselige stoffer. Ligeledes kan det
forventes at bioakkumulerbare stoffer i højere grad end vandopløselige stoffer vil kunne medføre forsinkede effekter efter endt eksponering som følge af det akkumulerede pesticid.
4.3.1 Metode
De gennemførte tests har i videst muligt omfang fulgt en standardiseret protokol for undersøgelse af akut toksicitet i det akvatiske miljø; OECD 202; Daphnia sp; Acute Immobilisation Test (OECD 1984).
Der foreligger ikke egentlige guidelines til gennemførelse af test med de her anvendte arter, men principperne i OECD guideline nr. 202 er fulgt i overordnede træk. De gennemførte forsøg har på nogle
områder afviget fra standardtesten; dels er relationen mellem toksicitet og eksponeringstid undersøgt over en længere serie af eksponeringstider, og dels er testorganismerne efter endt eksponering blevet
overflyttet til rent vand og holdt under observation i op til 14 dage.
Testorganismer blev indsamlet i vandløb af DMU og bragt til DHI, hvor de blev akklimatiseret under betingelser svarende til dem i forsøget i mindst 3 dage inden forsøgets start. Inden forsøget blev dyrene
sorteret efter størrelse og det blev tilstræbt at anvende mindre individer af samme størrelse og dermed nogenlunde samme alder, idet optag og dermed effekt af kemiske stoffer bl.a. antages at afhænge af
overflade-volumen forholdet samt udviklingsstadie af forsøgsorganismerne. Eksponeringskoncentrationerne i de enkelte forsøg blev fastlagt på baggrund af data for akut toksicitet for akvatiske organismer
fundet i litteraturen og/eller databaser. Pesticiderne blev maksimalt testet i koncentrationer op til 5-10 mg l-1. Højere koncentrationer end dette niveau vurderedes ikke at være miljørealistisk.
Eksponeringsforsøgene blev udført i åvand fra Brådebæk (Hørsholm). Testorganismerne blev eksponeret for en fortyndingsrække af det relevante pesticid i variable tidsforløb, hvorefter de blev overført til
rent vand. Herefter er udviklingen af toksiciteten fulgt i en periode op til 14 døgn. Ca. 150 dyr blev eksponeret for hver testkoncentration i 20 L akvarier med svag gennembobling. Eksponeringstiderne var:
½; 1; 2; 4; 6; 24; 48; 72; 96; 168, 240 og 336 timer. Til hver eksponeringstid og –koncentration blev der overført 10 dyr til rent vand i 250 ml krystallisationsskåle forsynet med svag gennembobling gennem
kanylespids. Forsøgene blev gennemført semi-statisk med to ugentlige vandskift i både eksponeringskar og krystallisationsskåle således at testkoncentrationerne i videst muligt omfang blev opretholdt i
testperioden.
Dødelighed samt synlige ændringer i adfærd blev bestemt, dels til de enkelte tider i løbet af eksponeringsperioden og dels hver 24. time efter at dyrene blev overført til rent vand. Testorganismerne blev
fodret med Tetramin under eksponeringen såvel som efter overførsel til rent vand. Ilt og pH blev målt løbende både under eksponering og i rent vand. Eksponeringerne blev udført i klimarum ved ca. 15 °C,
dog blev forsøg med vårfluelarver og juvenile slørvinger udført ved ca. 5 °C for at begrænse udviklingshastigheden og således sikre at dyrene ikke metamorfoserede i løbet af forsøget.
Det blev tilstræbt at undersøge et bredt spektrum af pesticider for at afdække effekterne af pesticider med forskelligartede virkemekanismer inden for de forskellige grupper af insekticider, fungicider og
herbicider. Der blev testet 7 pesticider i forsøg med Gammarus pulex: esfenvalerat, pendimethalin, azoxystrobin, pirimicarb, prochloraz, dimethoat og ioxynil. Esfenvalerat blev yderligere testet på Isoperla
grammatica og Agapetus sp. I det omfang det har været muligt er eksperimenterne blevet udført med radioaktivt mærkede pesticider (esfenvalerat, pendimethalin og ioxynil). Ved brug af radioaktivt
mærkede stoffer kan man dels følge den aktuelle eksponeringskoncentration og dels opnå informationer om akkumulering og koncentration af stoffet i testorganismerne.
LC50 værdier for pesticidernes akutte letale effekt som funktion af eksponeringstid blev beregnet ved hjælp af probit analyse (Probit analyse, version 2.3, 1990).
4.3.2 Resultater
Toksicitet under eksponering
Resultaterne fra forsøg med Isoperla grammatica og Agapetus sp. eksponeret for esfenvalerat viste, at disse to arter ikke var velegnede til langtidsforsøg i laboratoriet i det anvendte testdesign. For
Isoperla grammatica blev der først observeret effekter efter 7 dages eksponering (0-0,5 µg l-1). Efter 10 dage indtraf en forhøjet dødelighed hos kontroldyrene og der blev således ikke opnået information
om effekter af eksponeringsvarighed. For Agapetus sp. kunne der heller ikke etableres en sammenhæng mellem effekter og eksponeringsvarighed indenfor koncentrationsintervallet 0-1 µg l-1 da der
ligeledes indtraf en forhøjet dødelighed hos kontroldyrene allerede efter 72 timers varighed. Der blev derfor udelukkende arbejdet videre med Gammarus pulex som i højere grad var anvendelige til
langtidsforsøg i den anvendte forsøgsopstilling.
Eksponering af Gammarus pulex for de valgte pesticider viste som forventet en stigende mortalitet ved øget eksponeringsvarighed. For esfenvalerat og azoxystrobin, som var de mest toksiske pesticider,
indtrådte en dødelig effekt i de højest testede koncentrationer allerede efter få timers eksponering (Fig. 19). For de øvrige pesticider sås letale effekter først senere i eksponeringsforløbet, hvorfor
effektkoncentrationer (LC50) først kunne estimeres efter flere døgns eksponering.
Idet eksponeringsforsøgene skulle tilvejebringe informationer om toksiciteten i et relativt langt tidsinterval (0-14 dage) var det vanskeligt at vælge koncentrationsserier som både medførte effekter på en
meget kort tidsskala (minutter-timer) uden samtidig at slå hele testpopulationen ihjel inden for få døgn. Esfenvalerat havde en markant højere toksicitet overfor Gammarus pulex sammenlignet med de øvrige
pesticider. Letale effekter af esfenvalerat gjorde sig gældende i koncentrationsområdet ng l-1 mens effekter af de øvrige pesticider sås i koncentrationsområdet mg l-1. Pirimicarb medførte ikke toksiske
effekter overfor Gammarus pulex i de testede koncentrationer (0-5 mg l-1).
Klik her for at se Figur 19.
LC50 blev bestemt som funktion af eksponeringstiden for de enkelte pesticider. I Figur 20 ses sammenhængen mellem eksponeringstid og LC50 for esfenvalerat, azoxystrobin, pendimethalin, prochloraz,
dimethoat og ioxynil. I Bilag B er de eksakte værdier for både LC50 og LC10 koncentrationer angivet.
Figur 20 illustrerer hvorledes pesticidernes toksicitet øges som funktion af eksponeringstiden. Toksicitetsforøgelsen sker typisk inden for de første 2 døgn efter begyndende toksicitet observeres, og efterfølgende flader toksicitetskurven ud. Dette skyldes bl.a. at der kræves en vis eksponeringsvarighed før der indtræder en ligevægt mellem pesticid koncentration i organismen og eksponerings-koncentration. Teoretisk set vil denne ligevægt indstille sig hurtigere for vandopløselige stoffer sammenlignet med lipofile stoffer. Lipofile stoffer kan akkumuleres over en længere periode, og effektkoncentrationen vil derfor falde over en længere tidsskala. De her undersøgte pesticider har vidt forskellige opløseligheder. Esfenvalerat, pendimethalin, prochloraz og ioxynil kan karakteriseres som lipofile stoffer (log Kow > 3, se Tabel 4) mens azoxystrobin og især dimethoat og pirimicarb er relativt vandopløselige. Effekten af eksponeringsvarighed synes dog ikke at variere for de hhv. opløselige og lipofile pesticider. LC50 stabiliseres ikke markant hurtigere for de vandopløselige pesticider azoxystrobin og dimethoat sammenlignet med de øvrige, mere lipofile pesticider. For alle 6 pesticider ses imidlertid en tendens til at toksiciteten vedbliver med at falde, hvilket kan skyldes det konstante fysiologiske stress dyrene er udsat for under eksponeringen.
Klik her for at se Figur 20.
De anvendte pesticider udgøres af både insekticider, herbicider og fungicider. For at sammenligne den akutte toksicitet for de forskellige typer af pesticider er de beregnede LC50 værdier efter endt
eksponering vist i Tabel 13. Således fås følgende rangordning af de 7 pesticiders toksicitet: esfenvalerat > azoxystrobin > pendimethalin > prochloras > demethoat > ioxynil > pirimicarb.
Tabel 13. Sammenlignende toksicitet af de anvendte pesticider efter 14 dages eksponering
| Pesticid |
Type |
Virkemekanisme |
LC50 (14 dage)
mg l-1 |
| Esfenvalerat* |
Insekticid |
Nervegift |
3,2·10-5 |
| Azoxystrobin |
Fungicid |
Mitichondriel respiration inhibitor |
0,01 |
| Pendimethalin* |
Herbicid |
Celledeling inhibitor |
0,26 |
| Prochloraz |
Fungicid |
Ergosterol biosyntese inhibitor |
0,56 |
| Dimethoat |
Insekticid |
Cholinesterase inhibitor |
1,29 |
| Ioxynil* |
Herbicid |
Inhibitor af fotosyntese og oxidativ phosphorylering |
5,73 |
| Pirimicarb |
Insekticid |
Cholinesterase inhibitor |
>5 |
| * LC50 baseret på målte koncentrationer af radioaktivt stof |
Det fremgår af Tabel 13 at de undersøgte insekticider ikke konsekvent var de mest toksiske af pesticiderne på trods af at insekticiderne i kraft af deres egenskaber kunne forventes havde en mere specifik
virkning på invertebrater end herbicider og fungicider.
For pendimethalin, prochloraz, dimethoat, ioxynil og pirimicarb indtraf der ikke akutte letale effekter inden for det første døgn i de testede koncentrationer. Da de testede koncentrationer generelt er meget
høje i forhold til hvad der er målt i danske vandløb kan der ikke forventes umiddelbare toksiske effekter ved realistiske koncentrationer.
Forsinket toksicitet
Observationerne af dyr der er blevet overført til rent vand efter eksponering gav ikke et entydigt billede af forsinkede effekter som følge af eksponering til pesticider. I forsøg med Isoperla grammatica samt
Agapetus sp. indtraf en forhøjet dødelighed både hos kontroldyr og eksponerede dyr umiddelbart efter overførsel til rent vand, hvorfor der ikke blev opnået information om eventuelle forsinkede effekter. I
forsøg med Gammarus pulex blev der i flere tilfælde observeret en tendens til at toksiciteten bibeholdes efter endt eksponering og i nogle tilfælde ligefrem forstærkes, idet dødeligheden øgedes efter endt
eksponering. Især i koncentrationer, hvor der ikke blev observeret mortalitet under eksponeringen, sås en tendens til forsinkede toksiske effekter i de efterfølgende ca. 14 dage i rent vand. Dette kunne tyde
på at korte pulseksponeringer, der ikke umiddelbart resulterer i akutte, toksiske effekter, kan medføre en forsinket langtidseffekt hos vandløbsdyr. I andre tilfælde sås en initiel ”recovery” effekt af
Gammarus efter overførsel til rent vand, som dog efterfølges at en øget mortalitet. Dette gør sig gældende for de mere toksiske eksponeringskoncentrationer, hvor mortaliteten falder umiddelbart efter at
dyrene overføres til rent vand, men senere øges til et niveau svarende til de dyr der er under eksponering.
Især to faktorer gør tolkningen af de forsinkede effekter hos Gammarus pulex vanskelige. I de glas hvor dyrene opholdt sig i rent vand blev der observeret en stigende grad af kannibalisme med tiden, trods
regelmæssig fodring igennem hele forsøget. De individer der blev spist af deres artsfæller blev udeladt fra beregningerne. Dertil medfører forsøgsdesignet at de individer der overføres til rent vand kan have en
anderledes tolerance overfor det anvendte pesticid i forhold til den samlede testpopulation. Ved koncentrationer der ikke medfører akutte toksiske effekter (lave koncentrationer, kort eksponeringsvarighed)
må individer der udtages til rent vand formodes at have en tolerance der er repræsentativ for hele populationen. Ved koncentrationer hvor der observeres en høj akut mortalitet (høje koncentrationer, lang
eksponeringsvarighed) må de individer der overføres til rent vand formodes at have en højere tolerance overfor det anvendte pesticid, idet de mest følsomme individer allerede er døde.
I relation til miljørealistiske eksponeringsscenarier er det mest relevant at se på de kortvarige eksponeringer hvor koncentrationen af det givne pesticid ikke medfører synlige toksiske effekter, og hvor de
individer der overføres til rent vand må formodes at have en følsomhed svarende til gennemsnittet for testpopulationen. Eksponeringer på op til 14 dage er væsentligt længere end den relevante tidsskala for
eksponering i vandløb (se Fig. 6). For at undersøge om kortvarige eksponeringer kan medføre forsinkede effekter er det således valgt at fokusere på sub-letale eksponeringskoncentrationer, dvs.
koncentrationer der ikke medførte mortalitet under eksponeringen. For de pesticider hvor der blev observeret en forsinket mortalitet i rent vand som følge af eksponering i op til 24 timer blev LT50rent vand
beregnet. LT50rent vand angiver det ”mediane letale tidspunkt”, hvor 50% af individerne i rent vand forventes at dø som følge af en kortvarig eksponering.
Forsinkede effekter af kortvarige, sub-letale eksponeringer gjorde sig gældende for esfenvalerat, azoxystrobin og prochloraz. For pendimethalin, dimethoat, ioxynil og pirimicarb blev der ikke observeret
forsinkede effekter som følge af korttidseksponering (0-24 timer). Esfenvalerat medførte ingen akutte effekter i op til 4 timers eksponering (0-1 µg l-1), men i den efterfølgende periode i rent vand indtraf en
stigende dødelighed hos dyr eksponerede for hhv. 0,25, 0,5 og 1 µg l-1. Figur 21 viser hvorledes mortaliteten øges hos individer der er overført til rent vand efter 0-4 timers eksponering for 1 µg l-1
esfenvalerat.
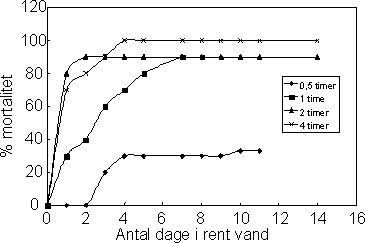
Figur 21. Forsinket mortalitet hos Gammarus pulex efter eksponering til 1 µg l-1 esfenvalerat i hhv. 0,5, 1, 2 og 4 timer
For Gammarus pulex eksponeret for 1 µg l-1 esfenvalerat blev LT50rent vand beregnet til 16,4 dage efter 0,5 timers eksponering, 2,1 dage efter 1 times eksponering og 0,76 dage efter 4 timers
eksponering (Tabel 13). Således falder LT50rent vand med stigende eksponeringsvarighed. Endelig kan det ses at den samme dødelighed (ca. 100%) opnås tidsforskudt som følge af både 1, 2 og 4 timers
eksponering til esfenvalerat.
Azoxystrobin (0-6,25 mg l-1) medførte akut mortalitet i de testede koncentrationer allerede efter 2 timers eksponering, hvorfor der kun er begrænsede data for sub-letale eksponeringer. Ved 0,5-1 times
eksponering for 2,5 mg l-1 sås en forsinket mortalitet i rent vand med en LT50rent vand på 2,8-5,4 dage (Tabel 14). Ved 0,5-1 times eksponering for 6,25 mg l-1 sås en forsinket mortalitet i rent vand med
en LT50rent vand på 1,4-1,9 dage.
Prochloraz medførte ikke akutte effekter inden for de første 6 timers eksponering (0-10 mg l-1), men i den efterfølgende periode i rent vand indtraf en stigende dødelighed hos dyr eksponeret for hhv. 2, 5
og 10 mg l-1. Figur 22 viser hvorledes mortaliteten øges hos individer der er overført til rent vand efter 0,5-6 timers eksponering for 10 mg l-1 prochloraz.
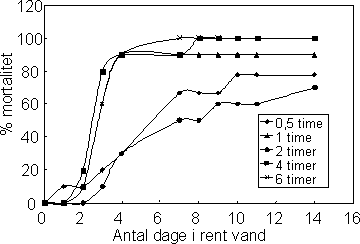
Figur 22. Forsinket mortalitet hos Gammarus pulex som følge af eksponering til 10 mg/l prochloraz i hhv. 0,5, 1, 2, 4 og 6 timer
For Gammarus pulex eksponeret til 10 mg l-1 prochloraz blev LT50rent vand beregnet til 5,7 dage efter 0,5 timers eksponering, 4,3 dage efter 1 times eksponering, 3,8 dage efter 4 timers eksponering og
2,8 dage efter 6 timers eksponering (Tabel 14).
Tabel 14. Median letale tid (LT50rent vand), hvor 50% af individerne dør efter en kortvarig eksponering.
| Pesticid |
Koncentration |
Eksponering
(timer) |
LT50rent vand
(dage) |
| Esfenvalerat |
1 µg l-1 |
0,5 |
16,4 |
| - |
1 |
2,1 |
| - |
4 |
0,76 |
| Azoxystrobin |
2,5 mg l-1 |
0,5 |
5,4 |
| - |
1 |
2,8 |
| 6,25 mg l-1 |
0,5 |
1,9 |
| - |
1 |
1,4 |
| Prochloraz |
10 mg l-1 |
0,5 |
5,7 |
| - |
1 |
4,3 |
| - |
4 |
3,8 |
| - |
6 |
2,8 |
For de mest toksiske pesticider ses således en tendens til at selv ganske kortvarige, ikke letale eksponeringer helt ned til 0,5 timer udløser et forsinket toksisk respons i form af en forøget dødelighed efter
eksponeringens ophør. Dette kan skyldes en tidsforskydning mellem eksponering og effekt ved korte pulseksponeringer, da pesticidet ikke når dets ”site of action” under eksponeringen. I sådanne tilfælde
kan tidsrummet mellem eksponering og effekt forventes at afhænge af pesticidernes virkemekanismer samt optagelses- og fordelingskinetik, idet transporthastigheden til de forskellige ”sites of action” vil
variere. Eksemplerne ovenfor illustrerer således at korte pulseksponeringer, der ikke medfører synlige akutte effekter, kan underestimere de toksiske effekter. Det ses også at eksponeringstiden influerer på
hvor hurtigt det efterfølgende toksiske respons indtræffer, idet LT50rent vand bliver fremskyndet med stigende eksponeringsvarighed.
Den forsinkede mortalitet, der blev observeret i forsøg med esfenvalerat, azoxystrobin og prochloraz er betinget af relativt høje eksponerings-koncentrationer. I vandløb vil koncentrationerne typisk være
langt lavere, og der kan ikke forventes forsinket mortalitet efter kortvarig eksponering. Forsinkede effekter kan dog også komme til udtryk i form af sub-letale effekter som f.eks. hæmmet udvikling, vækst
eller reproduktion, som på længere sigt kan påvirke populationerne (se 4.5). De udførte forsøg illustrerer dog vigtigheden af at vælge eksponeringsscenarier der tager højde for eksponeringsvarighed samt
semi-kroniske effekter, der indtræffer med forsinket virkning.
Resultaterne indikerer således, at de almindeligt anvendte parametre til bestemmelse af akut toksicitet (48-96 timers LC50) i flere tilfælde ikke kan anvendes til at forudsige korttidstoksiciteten af pesticider i
vandløb, da man ved ekstrapolering til effektkoncentrationer ved korte eksponeringstider ikke tager højde for eventuelle forsinkede effekter. For at kortlægge effekterne af pulseksponeringer for pesticider i
vandløb er det således relevant at inddrage en ”post-eksponerings fase” der også omfatter eventuelle forsinkede effekter.
4.4 Bestemmelse af interne effekt koncentrationer
Det normalt anvendte endpoint i toksicitets test, EC/LC50, angiver den testkoncentration i det omgivende miljø der medfører en effekt på 50% af testpopulationen. EC/LC50 kan derfor betragtes som den
eksterne effekt koncentration. Denne varierer som følge af almindelig biologisk variation testorganismer imellem, men også som følge af betingelserne i det omgivende miljø. F.eks. vil biotilgængeligheden af
lipofile stoffer være påvirket af mediets sammensætning, indhold af partikler og organisk materiale samt typen af testbeholdere, da disse parametre influerer på stoffets sorption.
Som supplement til eksterne effekt koncentrationer kan man undersøge kemiske stoffers interne effektkoncentration ved at relatere en given effekt (f.eks mortalitet) til den molære koncentration af stoffet i
organismen (mmol/kg vådvægt). Den interne effektkoncentration for et kemisk stof er teoretisk set konstant for forskellige organismer, idet det antages at en toksisk effekt udløses når vævskoncentrationen
når en effekt-specifik tærskelværdi. Det er dertil antaget at stoffer med forskellige virkemekanismer vil resultere i forskellige interne effektdoser.
4.4.1 Metode
I denne undersøgelse blev den interne letale koncentration (lethal body burden, LBB) bestemt i forsøg med de radioaktivt mærkede pesticider esfenvalerat, pendimethalin samt ioxynil eksponeret til
Gammarus. LBB blev kvantificeret ved at bestemme den akkumulerede radioaktivitet i nyligt døde individer indsamlet løbende fra forsøgene med varierende eksponeringsvarighed. Således blev der
beregnet LBB værdier til forskellige eksponeringstider og eksponeringskoncentrationer.
4.4.2 Resultater
De beregnede LBB værdier syntes at være uafhængige af eksponeringstiden mens der var en tendens til LBB steg med eksponeringskoncentrationen (data ikke vist). Spektret af de beregnede LBB værdier
er vist i Figur 23 (bemærk logaritmisk skala).
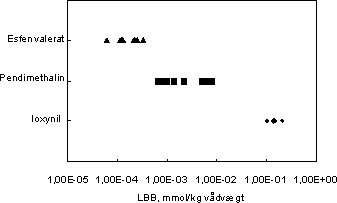
Figur 23. Lethal body burdens for esfenvalerat, pendimethalin og ioxynil.
Gennemsnittet af LBB værdierne for de tre pesticider er vist i Tabel 15. Ligeledes er LC50 efter endt eksponering (14 dage) angivet for at illustrere den relative toksicitet af pesticiderne.
Tabel 15. Lethal body burdens for esfenvalerat, pendimethalin og ioxynil
| Pesticid |
LBB, mmol/kg vådvægt |
LC50 (14 dage)
mg l-1 |
Esfenvalerat
(insekticid) |
1,7·10-4 (±8,3·10-5) |
3,2·10-5 |
Pendimethalin
(herbicid) |
2,8·10-3 (±2,6·10-3) |
0,26 |
Ioxynil
(herbicid) |
1,5·10-1(±5,4·10-2) |
5,73 |
Trods spredningen af LBB værdierne for de enkelte pesticider fremgår det at der er en simpel sammenhæng mellem LBB og pesticidernes akutte toksicitet, idet LBB falder med stigende toksicitet. Således
er den interne kritiske koncentration lavest for esfenvalerat som er det mest toksiske af de tre pesticider, mens den indre kritiske koncentration er væsentligt højere for ioxynil, som har en relativt lav akut
toksicitet. Med andre ord er den akutte toksicitet negativt korreleret med den interne kritiske koncentration: jo højere toksicitet, jo lavere er den indre kritiske koncentration der udløser det toksiske respons.
Dette kan understøtte teorien om, at forskellige virkemekanismer er associeret med forskellige spektre af body burdens.
Det var i denne undersøgelse ikke muligt at skaffe flere radioaktivt mærkede pesticider, hvorfor LBB ikke kunne beregnes for et bredt udsnit af pesticider med forskellige virkemekanismer. Både
esfenvalerat, pendimethalin og ioxynil er relativt lipofile stoffer hvorfor en akkumulering i organismerne over tid kan forventes. Det havde været relevant at undersøge om forsinkede effekter generelt var
betinget af et højt akkumuleringspotentiale, idet den akkumulerede body burden forventes at være højere for lipofile stoffer sammenlignet med vandopløselige stoffer. Dette var dog ikke muligt at gennemføre
i nærværende projekt.
4.5 Livscykluseffekter hos Gammarus pulex
Det var det primære mål med denne delundersøgelse at kvantificere den relative følsomhed hos forskellige livsstadier hos krebsdyret Gammarus pulex. Effekter på livscyklus hos insektet Chironomus riparius er omtalt i afsnit 4.6. Sammenholdt giver disse forsøg mulighed for at vurdere forskelle i følsomhed mellem arter, forskellige stadier og effektparametre.
I projektets sene fase blev der forsøgt at udføre livscyklusundersøgelser med dansemyggen Prodiamesa olivacea, der i modsætning til C. riparius lever udbredt i vandløb og med betydeligt højere krav til iltindhold og formodes at være mere følsom end C. riparius. Der var dog problemer med kultiveringen af P. olivacea og forsøgene kunne ikke gennemføres. I Bilag F er beskrevet forsøgene på kultivering.
4.5.1 Indsamling og forsøgsoversigt
Individer af G. pulex blev indsamlet fra Værebro Å, en sedimentrig og langsomtløbende å nær Roskilde Amts prøvestation nr. 1213. Indsamlingslokaliteten, der på den ene side var omgivet af brakmarker
og på den anden side af buske og træer, klassificeres af Roskilde Amt (1998) som tilhørende forureningsgrad II/III eller faunaklasse 4. Indsamlingen af organismer foregik ved brug af ketcher og
sparkeprøver. I laboratoriet blev organismerne forsigtigt sorteret i vandfyldte plastkar og kun ubeskadigede og ikke-parasitinficerede (Echinorrhyncus sp.) individer blev udvalgt til senere eksperimentelt
brug.
Opbevaring og kultivering af dyrene fulgte generelt McCahon & Pascoe's (1988a; 1988b) anvisninger herfor. Minimum 14 dage før start af et eksperiment blev dyrene opbevaret i 10 liters plastakvarier med
en tæthed på ca. 200-300 dyr pr. akvarium. Dyrene blev holdt i luftgennemboblet vand, der bestod af en blanding (50:50) af vand indsamlet på feltlokaliteten og kunstigt ferskvand. Det kunstige ferskvand
blev fremstillet efter OECD Guideline 202.
Både under kultivering og eksperimenter blev dyrene holdt i klimarum ved 13°C og kunstige lysforhold (12 timer lys:12 timer mørke). Foderet bestod af delvist opløste elleblade (Alnus sp.) indsamlet på
feltlokaliteten. Bladene, der alle var på samme forrådnelsesniveau, blev konditioneret i vand indsamlet på feltlokaliteten i en periode på minimum 14 dage inden brug.
Afhængig af eksperimentet varierede testkamrene i størrelse og volumen, men i de fleste tilfælde blev der anvendt plastbeholdere med et volumen på 200 ml vand. For at minimere turbulens i testkamrene og
stress af testorganismerne, blev der i visse eksperimenter tilført luft gennem kanylespidser som beskrevet af Welton & Clark (1980). I enkelte eksperimenter bestod testkamrene ligeledes af en sedimentfase.
I tilfælde af en sedimentfase blev der anvendt vasket akvariegrus med en partikelstørrelse på 2-3 mm.
Der blev udført 6 forskellige forsøg med Gammarus pulex varierende fra ”worst-case” eksponering af voksne individer til detaljerede forsøg med esfenvalerats effekt på parringsadfærd og effekter på
afkom.
Detaljer om dosering and analyse af esfenvalerat samt anvendt statestik fremgår af Bilag C.
4.5.2 Worst-case eksponering af G. pulex
Dette forsøg blev udført med henblik på bestemmelse af effekten af et ”worst-case scenarium”, hvor G. pulex blev eksponeret til en 1-times puls af esfenvalerat ved koncentrationerne: 0 og 4,65 µl-1, hvor
den høje koncentration svarer til den højeste mulige eksponering ved direkte oversprøjtning på en 2 meter dybt vandområde, beregnet udfra anvendelse af EU's maksimale dosis på 93 g a.i./ha. Herudover
blev der desuden anvendt en acetonekontrol. I både kontrolgruppen og acetonekontrolgruppen blev der anvendt 10 voksne (kropslængde = 10-14 mm) individer, mens der i eksponeringsgruppen blev
anvendt 20 voksne individer. Efter pulseksponering, der foregik i bægerglas, blev dyrene skyllet i rent vand og overført til testkamre indeholdende rent vand. Overlevelse og immobilitet blev herefter
observeret over en periode på 144 timer. Undervejs blev dyrene fodret med bladskiver (diameter = 18mm).
Figur 24 illustrerer effekter på overlevelsen af voksne G. pulex efter, at de har været udsat for et “worst-case pulseksponerings scenarium”. Allerede 1 time efter pulseksponeringen er kun 60% af de
eksponerede individer i live, mens der blev observeret 100 % dødelighed 144 timer efter eksponeringen. Til sammenligning er der henholdsvis 10% og 90% overlevelse i kontrol- og actonegruppen.
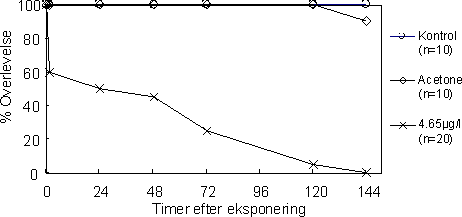
Figur 24. Overlevelse efter worst-case (4.56µg l-1) pulseksponering af G. pulex.
4.5.3 Effekt på juvenile og ikke-reproducerende voksne
Dette forsøg undersøgte overlevelsen af både juvenile (kropslængde = 3-7 mm) og ikke-reproducerende voksne (kropslængde = 9-15 mm) eksponeret til en 1-times puls af esfenvalerat. Foruden en kontrol
og en acetone-kontrolgruppe blev der anvendt testkoncentrationer på 0,1 og 0,3 µl-1. I forsøget blev testkamrene foruden vand tilført sediment (1 del sediment: 5 dele vand, på volumen basis). I halvdelen af
eksponeringsgrupperne blev testorganismerne efter pulseksponeringen overført til testkamre med rent vand og rent sediment, mens den anden halvdel af testorganismerne blev overført til testkamre hvor kun
vandet blev skiftet. Formålet med dette forsøgsdesign var at bestemme i hvor høj grad esfenvalerat bandt sig til sedimentpartiklerne under pulseksponeringen, og om det således efterfølgende kunne virke
som en kontinuerlig eksponeringskilde. Der blev anvendt 10 replikater af hver 10 individer pr koncentration for voksne og 10 replikater af 5 individer pr koncentration for juvenile. Umiddelbart efter
eksponering blev dyrene overført til deres respektive testkammer, hvor overlevelsen blev observeret hver anden dag over en periode på to uger.
Alle voksne individer overlevede den 1 times pulseksponering med esfenvalerat koncentrationer på op til 0,3 µg l-1. Overlevelsen var faldende i perioden efter eksponeringen, men variationer i median
overlevelsestid testgrupperne imellem blev først detekteret syv dage efter pulsen (Kruskal Wallis p-værdier var som følger: Dag 1: 0,746; Dag 2: 0,746; Dag 3: 0,141; Dag 4: 0,179; Dag 7: 0,033; Dag 9:
0,005; Dag 11: 0,046; Dag 14: 0,038). Gennemsnitlig og median overlevelsestid for hver gruppe, estimeret udfra overlevelseskurverne, fremgår af Tabel 16.
Table 16. Gennemsnitlig og median overlevelsestid (dage) hos juvenile og voksne efter 1-times pulseksponering til esfenvalerat. Værdierne i den sidste kolonne er fra sammenligninger i overlevelseskurver (Tarone-Ware Log-rank test) mellem juvenile og voksne indenfor hver eksponeringsgruppe.
| Gruppe |
Overlevelsestid
hos juvenile
Gennemsnitlig;
Median |
Overlevelsestid
hos voksne
Gennemsnitlig;
Median |
Tarone-Ware,
χ2 og
P-værdier |
| Kontrol |
11,44; >13 |
13,85; >14 |
19,5, 0.00001 |
| Acetone |
11,84; >13 |
13,84; >14 |
19,2, 0.00001 |
| 0.1, kontamineret sediment |
5,42; 3 |
13,43; >14 |
117,7, <0.00001 |
| 0.1, rent sediment |
6,46; 4 |
13,74; >14 |
103, <0.00001 |
| 0.3, kontamineret sediment |
1,88; <1 |
13,25; >14 |
163,6, <0.00001 |
| 0.3, rent sediment |
1,66;<1 |
13,1; >14 |
149,6, <0.00001 |
Betragtes overlevelseskurverne ses en tendens til at overlevelsen i de 14 dage efter pulseksponeringen faldt hurtigere i de eksponerede grupper end i kontrollerne og at overlevelsen i grupperne eksponeret til 0,3 µg l-1 faldt hurtigere end grupperne eksponeret til 0,1 µg l-1 (Figur 25). Selvom figuren indikerer en lille forskel i overlevelse mellem grupperne der modtog den samme pulskoncentration - men som blev overført til testkamre henholdsvis med eller uden kontamineret sediment - var forskellen mellem kurverne ikke signifikant (Tarone-Ware; 0,3 µg l-1: p=0.808; 0,1 µg l-1: p=0,601). Overlevelsen var generelt høj i alle grupper og oversteg 80 % i alle grupper (Figur 25).
Alle juvenile individer overlevede en times pulseksponering til esfenvalerat koncentrationer på op til 0,3 µg l-1. Overlevelsen faldt over tid i perioden efter pulsen, og i modsætning til de voksne individer kunne
der detekteres variationer i median overlevelsestid grupperne imellem allerede et døgn efter pulsen (Kruskal Wallis p-værdi 0,001 for alle dage). Overlevelseskurverne indikerer at overlevelsen faldt hurtigere
efter pulsen i de eksponerede grupper end i kontrol grupperne og at overlevelsen i grupperne eksponeret til 0,3 µg l-1 faldt hurtigere end i grupperne eksponeret til 0,1 µg l-1 (Figur 26). Selvom figuren
indikerer en lille forskel i overlevelse mellem grupperne der modtog den samme pulskoncentration - men som blev transporteret til testkamre henholdsvis med eller uden kontamineret sediment - var
forskellen mellem kurverne ikke signifikant (Tarone-Ware; 0,3 µg l-1: p=0,638; 0,1 µg l-1: p=0,184). Gennemsnitlig og median overlevelsestid for hver gruppe, estimeret udfra overlevelseskurverne, kan ses i
Tabel 16.
Ved slutningen af forsøget var overlevelsen i kontrollen og acetonekontrollen på henholdsvis 74 og 76 %, mens overlevelsen i grupperne eksponeret til 0,1 µg l-1 var 24% (rent sediment) og 14 %
(kontamineret sediment). I grupperne eksponeret til 0,3 µg l-1 var overlevelsen på henholdsvis 0 % (kontamineret sediment) og 2% (rent sediment).
Overlevelsen faldt hurtigere i alle eksponeringsgrupper hos de juvenile end hos de voksne individer, og trods små forskelle var der også signifikante forskelle i overlevelsen mellem juveline og voksne i
kontrol- og acetonekontrolgrupperne (Tabel 16).
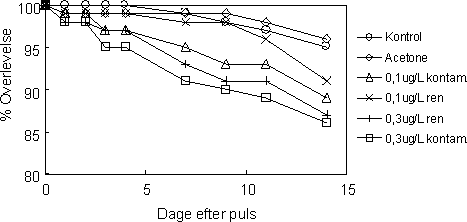
Figur 25. Overlevelse af voksne individer af G. pulex udsat for en 1-times puls af esfenvalerat.
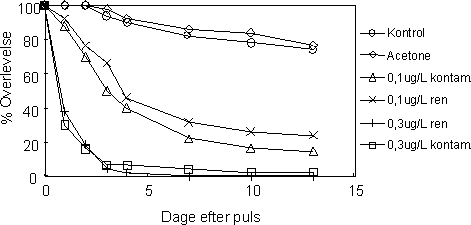
Figur 26. Overlevelse af juvenile individer af G. pulex udsat for en 1-times puls af esfenvalerat.
4.5.4 Effekt på reproducerende par og deres afkom
I dette forsøg undersøgtes effekten af en kort puls af esfenvalerat på reproduktionsadfærden hos voksne G. pulex. Fjorten sammenhængende par af G. pulex, alle med en ægbærende hun, blev eksponeret
til hver en 1-times puls af følgende testkoncentrationer: 0, 0,05, 0,1 og 0,3 µl-1. Eksponeringsproceduren var den samme som beskrevet under tidligere eksperimenter. Efter eksponering blev hvert par placeret individuelt i testkamre og overlevelsen af de voksne og deres afkom observeret over en periode på to uger. Juvenile der blev frigivet fra de gravide hunner under eksponeringen og den efterfølgende observationsperiode blev talt og opbevaret separat. Afkommet blev isoleret for at undgå prædation fra de voksne under forsøget.
Under den 1-times pulseksponering frigav hunnerne et stort antal befrugtet afkom (Tabel 17).
Tabel 17. Oversigt over antal unger frigivet under eksponering, procent unger observeret ved forsøgets slutning, antal overlevende voksne (hunner + hanner), samt antal gendannede par af 14 mulige.
Koncentration
(µgl-1) |
Antal unger
frigivet under
eksponering |
% levende unger
ved ekspone-
ringens ophør |
Antal levende voksne 15
dage efter eksponerin-
gen (hunner, hanner) |
Antal
gendan-
nede par |
| 0 |
12 |
100 |
6, 14 |
11 |
| 0,05 |
129 |
89 |
2, 7 |
0 |
| 0,1 |
142 |
89 |
3, 4 |
0 |
| 0,3 |
114 |
49 |
2, 0 |
0 |
Den procentvise overlevelse hos de unger der blev frigivet under pulseksponeringen var faldende med stigende koncentration (Figur 27). Overlevelsen af afkommet fortsatte med at falde over tid under den to uger lange observationsperiode i rent vand og var afhængig af pulskoncentrationen. Parrede voksne var relativt sensitive overfor esfenvalerat (sammenlignet med tidligere studier af ikke-parrede voksne) og overlevelsen ved den højeste koncentration (0,3 µg l-1) var meget lav for både hanner og hunner (Figur 28). Hertil kommer, at overlevelsen hos hunnerne var faldende over tid og ca. halvt så stor som hos hannerne ved slutningen af den to uger lange observationsperiode, og dette forhold gjorde sig ligeledes gældende i kontrollen (Tabel 17).
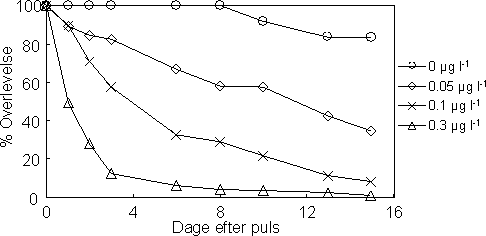
Figur 27. Overlevelse af unger frigivet efter en 1-times pulseksponering af forældrene ved følgende koncentrationsniveauer af esfenvalerat: 0, 0,05, 0,1 og 0,3 µg l-1.
Ingen par i de eksponerede grupper blev gendannet under den to ugers observationsperiode sammenlignet med kontrollen, hvor 11 ud af 14 par blev gendannet (Tabel 16). Ligeledes var overlevelsen blandt
de juvenile, der blev produceret under de to uger efter eksponeringen betydeligt lavere i forsøgsgrupperne end i kontrollen (Figur 29).
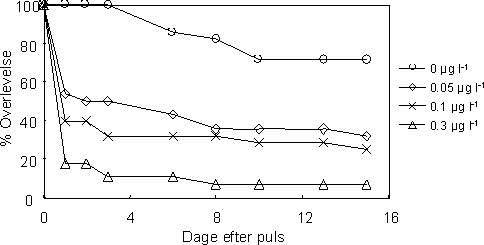
Figur 28. Overlevelse af reproducerende voksne G. pulex efter en 1-times pulseksponering ved følgende koncentrationsniveauer af esfenvalerat: 0, 0,05, 0,1 og 0,3 µg l-1.
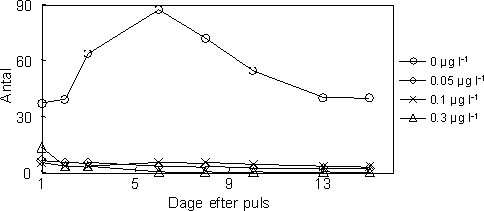
Figur 29. Antal overlevende unger produceret af voksne individer af G.
pulex over en periode på to uger efter en 1-times pulseksponering til esfenvalerat
ved følgende koncentrationsniveauer: 0, 0,05, 0,1 og 0,3 µg l-1.
4.5.5 Effekt på nyfødte
Da det forgående forsøgt afslørede en relativ høj dødelighed blandt de unger, der var blevet frigivet af den tidligere eksponerede moder, blev dette forsøg udført for at undersøge nyfødte individers sensitivitet overfor en kortvarig puls af esfenvalerat. Halvtreds nyligt frigivede juvenile (kropslængde = 1-2 mm) pr. testgruppe blev eksponeret til en 1-times puls af hver af følgende koncentrationer: 0, 0,05, 0,1 og 0,3 µl-1 og overlevelsen observeret over en periode på en uge. Der blev ikke anvendt replikater, idet alle individer fra hver koncentration blev holdt i et enkelt testkammer. Under observationsperioden blev de
juvenile fodret med fækalier produceret af voksne fra kulturakvarierne.
Overlevelsen blandt nyfødte eksponeret for en 1-times puls af esfenvalerat faldt med stigende koncentration i ugen efter eksponering (Tarone-Ware Log-rank Test, 2=138,2, P<0,00001; Figur 30). Den gennemsnitlige overlevelsestid var: 6,8 dage (kontrol), 5,1 dage (0,05g l-1 gruppe), 3,5 dage (0,1 µg l-1 gruppe) og 2,3 dage (0,3 µg l-1 gruppe).
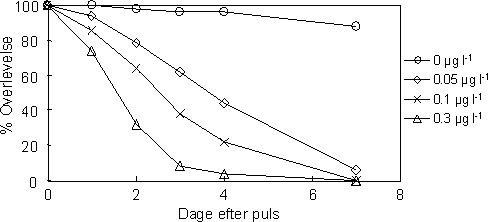
Figur 30. Overlevelse af nyligt producerede juvenile G pulex efter en 1-times pulseksponering til esfenvalerat ved følgende koncentrationsniveauer: 0, 0,05, 0,1 og 0,3 µg l-1.
4.5.6 Effekt på parringsadfærd og efterfølgende reproduktion
I modsætning til tidligere beskrevne eksperimenter relateret til G. pulex's reproduktive adfærd, blev dette forsøg udført med par af G. pulex hvor hunnerne endnu ikke var blevet befrugtet og derfor endnu
ikke bar juvenile på maven. Ti prækopulative par per testgruppe blev forsigtigt adskilt efter metoden beskrevet af Malbouisson (1995), idet det sammenhængende par blev lagt på et filterpapir indtil hannen
gav slip på hunnen. Hanner og hunner blev herefter individuelt eksponeret til en 1-times puls af hver af følgende koncentrationer: 0, 0,05, 0,1, 0,3 og 0,6 µl-1. Kropslængden hos hannerne varierede mellem
10-15 mm mens kropslængden hos hunnerne varierede mellem 9-12 mm. Efter eksponering blev dyrene skyllet i rent vand og placeret i testkamre med den samme partner som før adskillelsen. Overlevelse,
tid til evt. gendannelse af par, samt den procentvise gendannelse af par blev herefter observeret over en periode på to uger. Antallet af juvenile produceret per reproducerende hun blev ligeledes noteret.
Juvenile og døde individer blev straks fjernet fra testkamrene. Par hvor en af parterne døde i løbet af observationsperioden blev ikke anvendt i den endelige dataanalyse.
Under den 1-times eksponeringspuls blev de fleste af individerne i de tre højeste koncentrationsgrupper immobile og forblev liggende på bunden af eksponeringskamrene. Denne effekt fortsatte et par dage efter eksponeringen. Til trods for den relativt store størrelsesforskel kønnene imellem, blev der ikke observeret nogen statistisk signifikans i overlevelsen mellem hanner og hunner (Tarone-Ware Log-rank test χ2≤0,07 og P≥0,78 for alle koncentrationer). Tretten dage efter pulseksponeringen var overlevelsen på 95 og 100% i henholdsvis kontrollen og acetonekontrollen, hvorimod overlevelsen kun var på 60% i den højeste eksponeringsgruppe (0,6 µg l-1). For begge køn viste 0,1 µg l-1 gruppen en lavere overlevelse end 0,3 µg l-1gruppen. Den gennemsnitlige overlevelsestid var: 17,4 dage (kontrol), 13,0 dage (acetone), 11,8 dage (0,05 µg l-1), 12,7 dage (0,1 µg l-1), 12,2 dage (0,3 µg l-1), og 10,9 dage (0,6 µg l-1) (Figur 31).

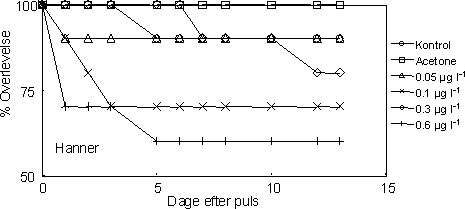
Figur 31. Overlevelse af reproducerende voksne G. pulex forstyrret fra præcopula ved separation og pulseksponering (1 time) til esfenvalerat ved følgende koncentrationer: 0, 0,05, 0,1, 0,3 og 0,6 g l-1. Der blev desuden anvendt en acetonekontrol. Øverst: hunner; Nederst: hanner.
Efter to dage var 60% af de overlevende par i både kontrollen og acetonekontrollen gendannet. Til sammenligning tog det ca. seks gange så lang tid (12 dage) før de forstyrrede par, der havde været
eksponeret til esfenvalerat, blev gendannet. Eksponeringskoncentrationen havde en signifikant effekt på den tid det tog for parrene at blive gendannet (Tarone-Ware Log-rank test χ2=16,29 og P=0,006).
Det bør dog noteres, at tiden til gendannelse af par i de højeste eksponeringsgrupper er baseret på færre individer, idet overlevelsen var faldende med stigende koncentrationer (Tabel 18).
Tabel 18. Varighed for genetablering af præcopula, procent hunner som reproducerede og gennemsnitligt antal afkom er hun efter pulseksponering til esfenvalerat.
| Grupper |
Gennemsnitlige tid til
gendannelse af par
(dage) |
% reproducerende
hunner 13 dage efter
eksponering |
Gennemsnitlig
(± SD) antal unger
pr hun |
| Kontrol |
4,5 |
80 |
16,1 (3,46) |
| Acetone |
7,4 |
80 |
13,4 (5,59) |
| 0,05 µg l-1 |
6,9 |
60 |
10 (3,03) |
| 0,1 µg l-1 |
14,9 |
16,7 |
10 (8,49) |
| 0,3 µg l-1 |
12,2 |
25 |
15 (7,21) |
| 0,6 µg l-1 |
12,3 |
33,3 |
8,5 (3,54) |
Eksponering til en kortvarig puls af esfenvalerat havde en signifikant effekt på antallet af juvenile, der blev produceret i perioden efter pulsen (Figur 32). Hos alle hunnerne havde esfenvalerat en signifikant effekt på antallet af juvenile produceret efter eksponering (Kruskal-Wallis Test, P=0,002). Kontrolgruppen producerede signifikant flere juvenile end 0,1 µg l-1 gruppen og 0,6 µg l-1 gruppen, men ikke 0,3 µg l-1 gruppen. Betragter man udelukkende de ynglende hunner vil esfenvalerats effekt på antallet af producerede juvenile derimod være marginal signifikant (Kruskal-Wallis Test, P=0,09). Dette indikerer at effekten på reproduktionen formentlig kan tilskrives det lavere antal reproducerende hunner.
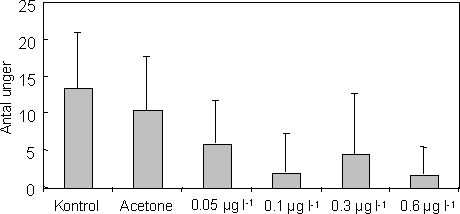
Figur 32. Gennemsnitlige antal unger produceret af gendannede par i de forskellige eksponeringsgrupper.
Det største antal juvenile (total = 133 individer produceret af 8 hunner) blev fundet i kontrolgruppen, hvilket kan forklares med en høj overlevelse samt hurtig genparring blandt de voksne. Til sammenligning blev der fundet færrest juvenile i 0,6 µg l-1 gruppen (total = 17 individer produceret af 2 hunner). Antallet af juvenile produceret pr. ynglende hun varierede fra 4 (0,1 µg l-1 gruppen) til 25 (acetone gruppen). Det bør noteres, at antallet af juvenile fundet i hver gruppe kan være en smule underestimeret grundet prædation fra de voksne.
4.5.7 Effekt af ugentlige eksponeringer til korte pulser
Dette forsøg blev udført for at undersøge effekten af gentagende pulser af 1 times varighed ved koncentrationerne: 0 og 0,1 µl-1 på parringsadfærden hos G. pulex. Der blev anvendt tre replikater af fem par voksne (kropslængde = 10-14 mm) individer af G. pulex per testgruppe. Testgruppe A blev udsat for tre pulseksponeringer (1 hver uge i tre uger), testgruppe B blev udsat for 2 pulseksponeringer (1 hver uge i de to første uger) mens testgruppe C blev udsat for en enkelt pulseksponering (1 i den første uge). Kontrolgruppen fik maksimal stressbehandling som testgruppe A, dvs. tre eksponeringer, blot med den forskel at eksponeringen foregik i rent vand. Eksponeringsproceduren var som tidligere beskrevet, idet parrene blev eksponeret i glasbeholdere og efterfølgende skyllet og overført til testkamre med rent vand. Dyrene var aldrig fødebegrænsede, idet de blev foderet regelmæssigt med bladskiver. Under den tre ugers forsøgsperiode blev overlevelse og gendannelse af parrene noteret.
Den højeste overlevelse efter tre uger blev observeret i kontrollen (77 %), mens den laveste overlevelse blev observeret i testgruppe A (60 %), der havde været udsat for tre pulseksponeringer (Figur 33). I
testgruppe B, der havde været udsat for to pulser, var overlevelsen 70 %, mens testgruppe C, der havde været udsat for en enkelt puls, sluttede med en overlevelse på 73 %.
Gendannelsen af par var generelt lav i alle testgrupper efter første eksponering, hvilket antyder, at de fleste par allerede havde reproduceret og derfor ikke behøvede at gendannes.
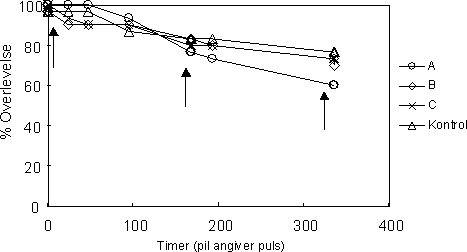
Figur 33. Overlevelse af G. pulex par udsat for en ugentlig 1-times pulseksponering af esfenvalerat (0,1µg l-1). Pilene angiver eksponeringstidspunkterne. A blev udsat for tre pulseksponeringer (1 hver uge i tre uger), B blev udsat for 2 pulseksponeringer (1 hver uge i de to første uger) mens C blev udsat for en enkelt pulseksponering (1 i den første uge).
4.6 Livscykluseffekter hos Chironomus riparius
400 larver fra henholdsvis en finsk og en engelsk kultur af Chironomus riparius blev anvendt til start af egen kultur. Kultiveringsteknikken fulgte generelt OECD Guidelines for the Testing of Chemicals
(Proposal for a New Guideline 219). Således blev larverne holdt i kulturbure med følgende dimensioner: højde = 40 cm, længde = 34 cm, bredde = 20 cm, hvor bunden blev forsynet med et ca. 3 cm tykt
lag kunstigt sediment fremstillet efter OECD's anvisning herfor. Vandet bestod af afboblet hanevand i en dybde på ca. 6 cm og der blev tilført luft og omrøring i vandsøjlen ved hjælp af en svag
gennemboblinge. Toppen af kulturburene bestod af en aftagelig sværmekasse fremstillet af myggenet med en maskestørrelse på 1mm. Dyrene blev holdt ved 20C og kunstige lysforhold (12 timers lys: 12
timers mørke) med en lysintensitet på ca. 1000 lux. Foderet bestod almindeligt fiskefoder (TetraMin) knust og opløst i vand ved forholdet: 1 gram fiskefoder/20 ml vand. I henhold til OECD's guideline
anbefales små larver en fodermængde på ca. 0,25-0,5mg/dag mens store larver kræver ca. 0,5-1 mg/dag for at opnå optimale vækstbetingelser. Producerede ægsække blev høstet dagligt og den eksakte
læggedato noteret, idet de herefter blev overført til petriskåle.
Anvendt statistik er anført i Bilag C.
4.6.1 Eksponering til en 1-times puls (uden sediment)
Forsøget havde til formål at undersøge effekten af en kortvarig puls af esfenvalerat ved et koncentrationsniveau på 0,2µg l-1. Seks ægsække med samme alder ( 1 døgn) fra egen laboratoriekultur blev
isoleret i en petriskål. En uge senere blev det ønskede antal larver udtaget og eksponeret i glasbeholdere med et vandvolumen på 0,5 liter. Der blev anvendt 4 replikater af 30 dyr pr. testkammer fordelt på
tre forsøgsgruppe. Forsøgsgrupperne var omfattede: en kontrolgruppe; en testgruppe eksponeret til en 1-times puls på 0,2µg l-1 og overført til testkamre med rent sediment; samt en testgruppe eksponeret til
en 1-times puls på 0,2µg l-1 og overført til testkamre med sediment der ligeledes havde været kontamineret til en 1-times puls på 0,2µg l-1.
Efter endt eksponering blev dyrene med automatpipette overført til et større kar med en stort vandvolumen, for at undgå overførsel af pesticidrester. Herfra blev dyrene atter ved hjælp af automatpipette
overført til de respektive forsøgskamre. Testkamrene målte 7 cm i diameter og 4 cm i højden og blev tilført 20 ml kunstigt sediment og 100 ml gennemboblet hanevand. 30 individer i hvert forsøgskammer
gav en tæthed på: 1 individ/ 1,3 cm². Hvert testkammer blev forsynet med et aftageligt låg, hvis top bestod af myggenet, til opsamling af klækkede individer. Der blev fodret svarende til et niveau på 0,5
mg/larve/dag.
Efterfølgende blev forsøgskamrene observeret i en periode på 39 dage med henblik på bestemmelse af følgende effektparametre:
- tid til klækning
- antal og kønsfordeling af færdigudviklede og sværmende individer
- reproduktion fra sværmende individer (antal producerede æg)
- klækningssucces af lagte æg
Antal og køn af klækkede individer i hvert testkammer blev dagligt noteret og de færdigudviklede individer blev straks overført til sværmebure med følgende dimensioner: højde = 20 cm, længde = 34 cm,
bredde = 20 cm. Sværmeburene indeholdt ligeledes en vandfyldt petriskål til opsamling af ægsække. Alle færdigudviklede individer tilhørende den samme forsøgsgruppe blev, uafhængigt af replikat, overført
til det samme sværmebur. Dette var for at sikre et tilstrækkeligt stort antal individer pr. sværmebur til at reproduktion kunne foregå. Manglen på replikater som følge af denne forsøgsopstilling betød, at det
ikke var muligt at udføre en statistisk analyse på de opnåede reproduktionsdata. Ved forsøgets afslutning blev sedimentet i samtlige testkamre sigtet for at bestemme antallet af evt. uklækkede larver og
pupper.
Figur 34 viser den procentvise kumulative andel af alle klækkede individer i forsøgsperioden, idet hver kurve repræsenter både hanner og hunner. Den højeste andel af klækkede individer over tid blev
observeret i kontrolgruppen (ca. 50%), mens den laveste andel blev fundet i gruppen med kontamineret sediment (ca. 12%).
Eksponering til esfenvalerat reducerede tiden til klækning i både gruppen med rent og kontamineret sediment (Tabel 19). Generelt var hunnerne længere tid om at klække end hannerne. Selvom der ikke var
signifikans i interaktion mellem esfenvalerat og køn, indikerede separate parvise sammenligninger af hver gruppe, at det kun var i kontrolgruppen at klækningstiden hos hunner var signifikant længere end
hanner (ANOVA; kontrol: P=0,001; rent sediment: P=0,88; esfenvalerat sediment: P=0,22). Median klækningstid er vist i Tabel 20. Klækningstiden blev reduceret for både hanner og hunner ved
pulseksponering til esfenvalerat, men effekten var større på hunner end på hanner. Eksponering til esfenvalerat havde også en markant effekt på det totale antal af voksne der klækkede (Tabel 20; Pearson
Chi-square = 49,44, df=2, P << 0,001). Dette forhold var gældende for både gruppen holdt i rent og kontamineret sediment efter pulseksponeringen.
Figur 34. Procentvise kumulative andel af alle klækkede individer over en forsøgsperiode på 39 dage.
Tabel 19. Effekt af esfenvalerat og køn på tid til klækning hos Chironomus riparius. Interaktionen mellem esfenvalerat og køn var ikke signifikant (P=0,50) og en reduceret model, uden interaktion, blev anvendt.
| Effekt |
df |
F |
P |
| Esfenvalerat |
2 |
7,22 |
0,001 |
| Køn |
1 |
9,94 |
0,002 |
| Error |
88 |
|
|
Det totale antal æg produceret af kontrolgruppen var 1180, 164 æg blev produceret af gruppen holdt i rent sediment efter eksponeringen, mens der blev produceret 472 æg af gruppen holdt i kontamineret
sediment efter eksponeringen. Af disse klækkede 835 succesfuldt i kontrolgruppen (70,1%), 122 i gruppen med rent sediment (74,4%) og 319 i gruppen med kontamineret sediment (67,6%).
Tabel 20. Median tid til klækning og total antal klækkede voksne individer af Chironomus riparius.
| |
Kontrol |
Rent Sediment |
Esfenvalerat
Sediment |
| |
Median klækningstid (dage) |
| Hanner |
33 |
29 |
29,5 |
| Hunner |
37 |
30,5 |
32 |
| |
Total antal klækkede individer |
| Hanner |
37 |
15 |
4 |
| Hunner |
21 |
4 |
11 |
| Total |
58 |
19 |
15 |
| Procent |
64,4 |
21,1 |
16,7 |
4.6.2 Eksponering til en 1-times puls (med sediment)
Da den mest realistiske eksponeringsvej for den sedimentlevende organisme C. riparius formodes at foregå gennem sedimentet blev testorganismerne i dette forsøg pulseksponeret mens de befandt sig i
sedimentet, modsat i ovenstående forsøg, hvor testorganismerne blev pulseksponeret udenfor sedimentet.
Foruden bestemmelse af effekten af en kortvarig puls til esfenvalerat ved et koncentrationsniveau på 0,2 µg l-1 havde dette forsøg til formål at undersøge betydningen af populationstætheden i testkamrene.
Fra egen laboratoriekultur blev udtaget 6 ægsække med samme alder (1 døgn). Disse blev isoleret i en petriskål og en uge senere blev 600 larver udtaget og fordelt i følgende forsøgsgrupper af hver 10
replikater: en kontrolgruppe med en tæthed på 1 larve pr. 5 cm², en testgruppe med en tæthed på 1 larve pr. 5 cm² eksponeret til en 1-times puls esfenvalerat (0,2 µg l-1); en kontrolgruppe med en tæthed
på 1 larve pr. 2,5 cm², en testgruppe med en tæthed på 1 larve pr. 2,5 cm² eksponeret til en 1-times puls esfenvalerat (0,2 µg l-1). Pulseksponeringen af testorganismerne foregik først et døgn efter, at
larverne var blevet introduceret til de respektive testkamre indeholdende kunstigt sediment og gennemboblet hanevand i volumenforholdet (1:5). Herved havde larverne fået lejlighed til at grave sig ned i
sedimentet inden selve eksponeringen. Testkamrene målte 8 cm i diameter og 4 cm i højden. Ligesom i det foregående forsøg blev hvert testkammer forsynet med et aftageligt låg til opsamling af klækkede
individer. Der blev foderet svarende til et niveau på 0,25 mg/larve/dag.
Herefter fulgte forsøget den samme procedure som beskrevet under forsøget med eksponering udenfor sedimentet, idet der blev testet for de samme endpoints, samt betydningen af individtætheden i
testkamrene. Dette forsøg blev kørt over en periode på 44 dage.
Figur 35 viser den procentvise kumulative andel af alle klækkede individer i forsøgsperioden, idet hver kurve repræsenter både hanner og hunner. Den højeste andel af klækkede C. riparius blev observeret
i kontrolgruppen med lav individtæthed, hvor hele 93% af de tilførte testorganismer var klækket ved forsøgets afslutning efter 44 dage. Den laveste andel (57%) af klækkede C. riparius blev fundet i
gruppen med høj individtæthed eksponeret til 0,2 µg l-1.
Figur 35. Procentvise kumulative andel af alle klækkede individer over en forsøgsperioden på 44 dage.
Hos hanner havde esfenvalerat eksponering en reducerende effekt på tid til klækning, mens stigende individtæthed havde en øgende effekt på tid til klækning (Tabel 21). Hos hunner var der ingen effekt af
esfenvalerateksponering på tid til klækning, mens stigende individtæthed havde en signifikant øgende effekt på tid til klækning. Både i kontrol og esfenvalerat eksponerede grupper, og ved både lave og høje
individtætheder, blev der hos hunnerne observeret en signifikant længere tid til klækning end hos hannerne (p0,001 for alle tests).
Table 21. Effekt af esfenvalerat og tæthed på tid til klækning hos Chironomus riparius. Hverken for hanner eller hunner var interaktionen mellem esfenvalerat og tætheden signifikant (Hanner; P=0,685, Hunner; P=0,241).
| Køn |
Effekt |
Df |
F |
P |
| Hanner |
Esfenvalerat |
1 |
8,69 |
0,004 |
| Tæthed |
1 |
102.50 |
<0,001 |
| Error |
215 |
|
|
| Hunner |
Esfenvalerat |
1 |
0,031 |
0,86 |
| Tæthed |
1 |
86,13 |
<0,001 |
| Error |
190 |
|
|
Tiden til 50% af individerne klækkede og det totale antal af klækkede individer i forsøgsperioden på 44 dage er vist i Tabel 22. Individtætheden havde en signifikant og negativ effekt på klækningssuccesen
af larver i både kontrollen (Pearson Chi-square=24,02; P < 0,001) og gruppen eksponeret til esfenvalerat (Pearson Chi-square=24,10; P < 0,001). Derimod havde esfenvalerat ingen signifikant effekt på
den procentvise andel af larver med klækningssucces i gruppen med lav individtæthed (Pearson Chi-square=1,45, P=0,23) eller i gruppen med høj individtæthed (Pearson Chi-square=1,22; P=0,27).
I Tabel 23 er vist det samlede reproduktive output i de forskellige grupper over den 44 dage lange forsøgsperiode. Hverken eksponering til esfenvalerat eller individtæthed havde en signifikant effekt på
ægproduktionen (Esfenvalerat: df=1,4; F=0,31; P=0,61; Tæthed: df=1,4; F=1,86; P=0,24; esfenvalerat x Tæthed: df=1,4; F=0,99; P=0,38). Ligeledes blev der heller ikke observeret nogen signifikant
effekt på antallet af unger der klækkede (esfenvalerat: df=1,4; F=0,03; P=0,87; Tæthed: df=1,4; F=0,42; P=0,55; esfenvalerat x Tæthed: df=1,4; F=0,96; P=0,38). Den gennemsnitlige klækningssucces af
æggene var for alle grupper 77% (±11,25=SD).
Tabel 22. Median klækningstid og antal klækkede voksne individer af Chironomus riparius ved 1 larve pr. 5 cm² og 1 larve pr. 2.5 cm².
| |
Median klækningstid (dage) |
| Kontrol |
Esfenvalerat |
| Tæthed |
1 larve
pr. 5 cm² |
1 larve pr.
2,5 cm² |
1 larve pr.
5 cm² |
1 larve pr.
2,5 cm² |
| Hanner |
29 |
35 |
28 |
32 |
| Hunner |
34 |
42 |
35 |
40 |
| |
Total antal klækkede individer |
| Kontrol |
Esfenvalerat |
| |
10 |
20 |
10 |
20 |
| Hanner |
49 |
64 |
37 |
68 |
| Hunner |
44 |
52 |
51 |
46 |
| Total |
93 |
118 |
88 |
114 |
Procent
klækkesucces |
93 |
64,5 |
88 |
57 |
Tabel 23. Reproduktive output fra Chironomus riparius ved 1 larve pr. 5 cm² og 1 larve pr. 2.5 cm². Klækkede individer fra replikat 1-5 og replikat 6-10 blev puljet for at sikre tilstrækkeligt antal voksne individer til reproduktion.
| |
Total antal æg (overlevende afkom) |
| Kontrol |
Esfenvalerat |
1 larve pr.
5 cm² |
1 larve pr.
2,5 cm² |
1 larve pr.
5 cm² |
1 larve pr.
2,5 cm² |
| Rep. 1 – 5 |
1142 (932) |
801 (676) |
1678 (1368) |
1163 (956) |
| Rep. 6 - 10 |
1804 (1356) |
1011 (832) |
1056 (533) |
1395 (1105) |
4.7 Drift hos invertebrater efter pesticideksponering
En sammenligning af følsomheden af forskellige effektparametre i test med pesticider i kunstige vandløb har vist, at forøget drift (korttidsrespons) af insektlarver og gammarider generelt er mere følsom end
reduktion i abundans (langtidsresponse) (Møhlenberg et al. 2001). I vandløb er drift en naturlig adfærd hos bundlevende krebsdyr og insektlarver, der knytter sig til spredning, fødesøgning og forplantning.
Ved ugunstige forhold f.eks. som følge af tørke og forekomst af toksiske stoffer så som pesticider kan der udløses katastrofedrift i invertebratbestandene. Denne drift kan dels være udtryk for en aktiv
response for ugunstige forhold eller den kan skyldes en narkotisk virkning af et pesticid og en følgende passiv transport med strømmen.
Under normale forhold kan 1-5 % af en population være i drift på et givet tidspunkt. Hvis bunddyrene udsættes for insekticider som lindan, permetrin oa. øges driftraten, i enkelte tilfælde med op til 10000 %
(Møhlenberg et al. 2001). Generelt kan drift og ændring i abundans opfattes som reciprokke processer, idet øget drift fra et område vil føre til reduktion i abundans.
I projektet er der undersøgt for pesticiders effekt på drift hos krebsdyr og insektlarver, dels under laboratorieforhold i små systemer (recirkulerende strømrender) dels under under ”semi-naturlige” forhold i
kunstige vandløb uden recirkulering
4.7.1 Drift i recirkulerende strømrender
I eksperimenterne anvendtes 16 strømakvarier placeret i termorum (Fig. 36). Akvarierne er fremstillet i en blanding af transparent, lys og mørk grå, 5mm tyk akryl, bukket i form under opvarmning og samlet
ved svejsning med akryltråd. Akvarierne er 1m lange, 20cm brede og adskilt i to parallelle render med en 80cm lang midtervæg. Vandstrømmen genereres af en cylinder i PVC med en diameter på 11cm.
Ydervæggen er 7,5cm høj og under forsøgene er vanddybden 5cm ved 10 L vand, hvorved cylinderen stikker 2,5-3 cm ned i vandet. Strømhastigheden i systemerne bestemmes af omdrejningshastigeheden
af cylinder, der styres af 2 frekvensregulerede motorer, som hver er koblet til 8 strømrender

Figur 36. To af de 16 strømakvarier. Under forsøg tilsættes kvartsgrus og konditionerede blade som substrat for invertebraterne i renden modsat den roterende cylinder
I forsøgene anvendtes minimum 10 individer per art per strømakvarium. Et døgn inden et forsøg blev strømakvarierne fyldt med filtreret åvand, der placeredes 200 g kvartsgrus (5 mm) i renden modsat det
roterende hjul over en strækning på 45 cm. Der placeredes yderlige 10 ”konditionerede” blade (elm, el, eg) fra en beholder med åvand. Bladene blev fastholdt med sten. Dyrene blev tilsat og vandstrømmen
indstillet til 9-10 cm s-1 (fri hastighed ved overfladen). Efter 24 timer vil hovedparten af dyrene sidde i eller på blade/grus. Dyrenes placering (indenfor og udenfor grus) og aktivitet noteredes kontinuerligt
indtil 40 min. Herefter tilsattes pesticidet (opløst i acetone for tungtopløselige pesticider) i koncentrationer fastlagt ved indledende forsøg og dyrenes placering og aktivitet noteres igen kontinuerligt indtil 90
min og derefter over en 15 min periode hver 24. time indtil 96 timer. Ved hver registrering (ca. 4-5 gange i minuttet) noteres dyrenes placering (på strækning udenfor sten). Aktiviteten udenfor afsnittet med
sten inddeles i:
- Opstrøms migration
- Drift (nedstrøms)
- Anden bevægelse
- Inaktivitet
Intensiteten i optællingen varierede mellem 2-6 gange per 5 min periode afhængig af antal strømakvarier i et forsøg og især af dyrenes art og størrelse. For at undgå eventuelle bias blev observationerne
midlet over 5 min intervaller.
Hver forsøgsserie gennemførtes med minimum 2 kontrolrender og et varierende antal eksponeringsrender (6-14). I de tilfælde hvor det var nødvendigt at opløse pesticidet i acetone (esfenvalerat,
deltamethrin, pendimethalin, azoxystrobin) blev der tilsat acetone i samme koncentration i alle strømrender. Temperaturen under forsøget blev afpasset til årstiden og in situ temperaturen (6-15 °C). Der
anvendtes temperaturer indenfor intervallet 7 – 12 °C.
Driftraten blev udtrykt i procent af aktive (og observerbare) individer:
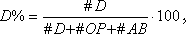
hvor #D = antal levende individer i drift, #OP = antal individer under optrøms migration, #AB = antal individer der viste andre aktivitetsmønstre eller kunne observeres som levende.
I stedet for at basere beregningerne på det totale antal tilsatte dyr i strømrenderne sikrede denne beregning, at døde og inaktive (evt. paralyserede) individer ikke indgik. Ulempen ved fremgangsmåden var
dog en større relativ variation fordi ”nævneren” (summen af #D, #OP og #AB) var variabel og ofte væsentlig lavere end det tilsatte antal dyr (10).
Bindingen af pesticider til overflader i strømakvarier kan mindske eksponeringen. Adsorptionen af esfenvalerat (som har den højeste Kd af de undersøgte pesticider) blev undersøgt ved hjælp af radioaktivt
mærket esfenvalerat. Tabet blev bestemt til at være mellem 20-30% over en time, som var den normale forsøgsvarighed (Figur 37). Vi antog derfor, at de nominelle koncentrationer gav et dækkende udtryk
for de reelle koncentrationer under forsøgene.
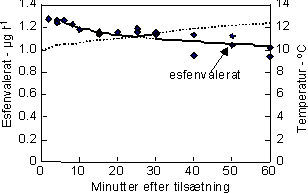
Figur 37. Tidslig variation i koncentrationen af radioaktivt esfenvalerat gennem 60 min i strømrende.
Driftrespons blev undersøgt hos Gammarus sp., Asellus, Baetis, Leuctra, Hydropsyche, efter eksponering til pesticiderne pendimethalin, pirimicarb, azoxystrobin, deltamethrin samt esfenvalerate (Tabel
24). Sidstnævnte blev undersøgt som rent kemisk stof og i handelsproduktet Sumi-alpha, som indeholder detergenter og hjælpestoffer i ikke offentliggjort koncentration og art.
Tabel 24. Oversigt over gennemførte forsøg med driftrespons (og toksicitet) i recirkulerende vandsløbsrender.
| Pesticid |
Organismer |
| Pendimentalin |
Gammarus, Asellus |
| Pirimicarb |
Gammarus, Baetis, Leuctra, Hydropsyche, Asellus |
| Azoxystrobin |
Gammarus, Asellus |
| Esfenvalerat |
Gammarus, Baetis, Hydropsyche, Leuctra, Asellus |
| Sumi-alfa (Esfenv.) |
Gammarus, Baetis, Hydropsyche, Leuctra, Asellus |
| Delta-methrin |
Gammarus |
Undersøgelserne viste, at kun pyrethroiderne (esfenvalerat og deltamethrin) udløste drift hos krebsdyrene og insektlarverne (Fig. 38 - 40), mens ingen af herbiciderne, fungicidet azoxystrobin eller insekticidet
pirimicarb udløste flugtrespons (Bilag D).
For esfenvalerat var der forøget drift hos Gammarus ved 0,6 ng l-1 45-90 min efter tilsætning, mens følsomheden hos Asellus, Baetis og Leuctra var ringere (typisk 6 ng l-1). Undersøgelserne viste ingen
drift hos Hydropsyche uanset koncentration eller art pesticid.
Klik her for at se Figur 38.
Klik her for at se Figur 39.
Undersøgelser med anvendelse af esfenvalerat i det formulerede produkt Sumi-alfa viste, at driften hos alle arter udløstes tidligere og ved lavere koncentrationer end det rene stof (Fig. 40 & 41).
Klik her for at se Figur 40.
Klik her for at se Figur 41.
”No effect concentrationer” (NEC) blev bestemt ved log-lineær regression (se også Fig. 7) som skæringen mellem den øverste konfidensgrænse for kontrollen og den nederste konfidensgrænse for
pesticideksponeringerne (Fig. 42). Ved høje koncentrationer af pyrethroider skete der ingen yderligere stigning i driftraten og ved længerevarende eksponering skete et fald i driftraten ved stigende koncentration fordi dyrene blev inaktive (se Fig. 40). Disse værdier indgik ikke i dosis-respons regressionen. EC50 koncentrationen blev beregnet udfra dosis-respons regressionen, hvis nedre grænse defineredes af driftraten i kontrolforsøgene og den øvre grænse af den maksimale driftrate i det enkelte forsøg (se Fig. 42). I eksemplet med esfenvalerat var kontroldriften 11% og den maksimale driftrate 53% ved 63,5 ng l-1.
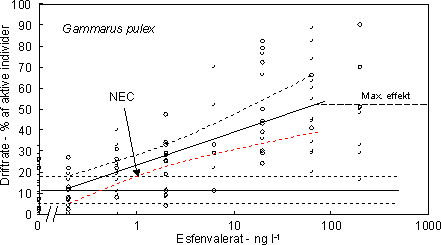
Figur 42. Dosis-respons relation for Gammarus pulex eksponeret til esfenvalerat. ”No effect concentration” (NEC) bestemmes ved skæringen af konfidensgrænserne for kontrolforsøg og eksponeringsforsøg (markeret med pil).
Tabel 25. Effekt af pyrethroider på drift hos Gammarus, Asellus, Baetis og Leuctra. ”No effect concentration” (NEC) og EC50 angivet i ng l-1 0-40 min og 45-90 min efter eksponeringsstart. - = kan ikke beregnes; nd = ingen data.
| Pesticid |
Tid |
Gammarus |
Asellus |
Baetis |
Leuctra |
| min |
NEC |
EC50 |
NEC |
EC50 |
NEC |
EC50 |
NEC |
EC50 |
| Esfenvalerat |
0-40 |
1,0 |
5,8 |
52 |
- |
9,1 |
17 |
8,1 |
51 |
| 45-90 |
0,55 |
3,2 |
9 |
68 |
6,4 |
14 |
- |
- |
Sumi-alfa
(esfenvalerat) |
0-40 |
0,19 |
0,29 |
1,1 |
8,0 |
0,77 |
2,8 |
0,46 |
3,6 |
| 45-90 |
0,16 |
0,36 |
0,90 |
2,9 |
0,60 |
1,5 |
0,80 |
3,3 |
| Delta-methrin |
0-40 |
3,5 |
12,2 |
nd |
nd |
nd |
nd |
nd |
nd |
| 45-90 |
1,6 |
7,8 |
nd |
nd |
nd |
nd |
nd |
nd |
De højeste koncentrationer af pyrethroider som ikke udløste forøget drift (NEC) og EC50 koncentrationerne varierede mellem arter og pesticider og reduceredes med varighed af eksponering. Gammarus
pulex var mest følsom med en NEC-værdi for esfenvalerat så lav som 0,55 ng l-1 (Tabel 25). Sammenlignet hermed var insektnymferne Baetis og Leuctra samt Asellus mindre følsomme med 10-20 gange
højere NEC- og EC50-værdier.
Testene med det formulerede produkt af esfenvalerat (Sumi-alfa) viste markant lavere effektværdier hos alle arter sammenlignet med det rene stof. Med undtagelse af EC50-værdien hos Asellus var
øgningen i følsomhed over tid begrænset, hvilket peger på en hurtigere effekt og optagelse end i det ikke-formulerede stof.
4.7.2 Drift under in-situ forhold
Driftrespons hos vandløbsinvertebrater er undersøgt før, under og efter pulseksponering med pyrethroidet esfenvalerat i semi-naturlige vandløbsrender. Undersøgelserne fokuserede hovedsageligt på
krebsdyret Gammarus pulex men også døgnfluen Baetis rhodani og vårfluen Sericostoma personatum indgik i undersøgelserne. Under forsøgene blev såvel koncentration som varighed varieret for at få
et indblik i driftresponset ved en række realistiske eksponeringsscenarier. Forsøgene blev først gennemført som enkeltarts test med G. pulex og derefter med samfund bestående af alle ovenstående arter.
Metode
Undersøgelserne blev udført på DMU's udendørs vandløbsrende-faciliteter i Lemming, 8 km nord for Silkeborg. Systemet består af 12 vandløbsrender som konstant fødes med friskt grundvand der hele
året har en temperatur på 7-8°C (Fig. 43). Inden vandet ledes i renderne føres det ned af en iltningstrappe og gennem et trykfilter, hvor bl.a. partikler og jern bindes (se Tabel 26). Fra trykfiltret føres vandet
til en fødekasse (50x70x60cm), hvorfra det, via et rørsystem, fordeles ligeligt til alle 12 render så de har en konstant vandføring på 3,2 l min-1. De 12 vandløbsrender er fire meter lange og 10 cm brede.
Faldet på vandløbsrenderne er indstillet til 1%, hvilket svarer til et naturligt fald på strygsekvenser i mindre danske vandløb. Substratet i vandløbsrenderne er udlagt således at det imiterer en strygsekvens i et
mindre dansk vandløb med grus og sten samt bladpakker i partierne med strømlæ (Figur 44). Ved den anvendte substratfordeling er vandets gennemsnitlige opholdstid i renderne 58,2 ± 1,3 sekunder og den
gennemsnitlige strømhastighed 0,06 m sek-1 (Tabel 26). Terminalt i hver vandløbsrende falder/løber hele vandføringen gennem et driftnet (maskevidde 1mm) som opfanger alle invertebrater større end 1
mm, der føres med strømmen.

Fig. 43. Foto af forsøgsopstillingen med de 12 vandløbsrender. Driftnettene er placeret i de lodrette nedløbsrør forrest i billedet.
Tabel 26. Fysiske og kemiske parametrer for vandet i renderne
| Kemiske parametrer |
Værdi/koncentration |
| NH4 |
0,009 mg l-1 |
| NO2 + NO3 |
2,720 mg l-1 |
| Total N |
2,830 mg l-1 |
| PO4 |
0,003 mg l-1 |
| Total P |
0,003 mg l-1 |
| Total Fe |
0,005 mg l-1 |
| pH |
7,0 |
| Fysiske parametrer |
| Vandføring per rende |
3,2 l min.-1 |
| Vandets gennemsnitlige opholdstid i renderne |
58,2 ± 1,3 sek. |
| Gennemsnitlig strømhastighed |
0,0637 ± 0,001 m sek.-1 |
| Vandtemperatur (range) |
6,50-8,19°C |
Såvel vandløbsrenderne som sedimentet i vandløbsrenderne blev udskiftet mellem alle forsøgsserier således at hver forsøgsserie indledtes med at etablere sedimentet. Først blev hver vandløbsrende tilført 4,5
kg grus (1-3 cm) fordelt jævnt på en 3 meter lang strækning. Gruset var forinden konditioneret 7 dage i vandløbsvand for at skabe en naturlig biofilm på substratet. Efter fordelingen af gruset blev der udlagt
knytnæve store sten for hver 30. centimeter. Disse sten blev indsamlet samme dag som etableringen af vandløbsrenderne i Hulbæk, et lille vandløb 5 km øst for Silkeborg. Inden stenene blev udlagt i
vandløbsrenderne blev de renset for dyr og skidt. Efter udlægningen af sten blev der i strømlæet bag hver sten placeret 2 elleblade (Alnus glutinósa (L)) som forinden var konditioneret 5 dage i kildevand
ved 10°C. Efter udlægningen af elleblade blev en Stowaway Tidbit temperatur logger placeret i nedstrømsenden af substratsstrækningerne. Efter etableringen af substratet blev et kendt antal invertebrater
introduceret umiddelbart opstrøms for den tre meter lange substratsstrækning i alle render. 1,5 time efter introduktionen af invertebraterne blev driftnettene udskiftet og de opfangede invertebrater blev
reintroducret opstrøms for substratsstrækningen.

Figur 44. Foto af de 3 meter lange substrat-strækninger bestående af grus, sten og elleblade.
Næste formiddag, ca. 20 timer efter introduktionen af invertebraterne, blev driftnettene skiftet igen og de fangede invertebrater kasseret og indgik dermed ikke i selve forsøget. Herefter blev driftnettene
udskiftet med 45 minutters intervaller resten af dagen og igen næste formiddag, ca. 20 timer efter pesticidtilførslen.
Under hver udskiftning blev de opfangede dyr skyllet ud i en hvid fotobakke, talt og konserveret i ethanol. Forsøgene blev afsluttet efter tømningen af driftnettene 20 timer efter pesticidtilførslen. Ved
forsøgenes afslutning blev substratet skyllet ud af renderne og tilbageværende invertebrater blev frafiltreret og talt. De ethanolkonserverede invertebrater blev i laboratoriet efterfølgende målt, således at deres
tørvægt kunne bestemmes på baggrund af tidligere beskrevne relationer mellem længden af en given morfologisk enhed (f.eks. hovedkapsel bredden) og dyrenes tørvægt (Zelinka & Marvan, 1976, Iversen
& Jessen, 1977, Friberg et al. 2002). Middel tørvægten af G. pulex og B. rhodani blev således bestemt til hhv. 3,85 ± 0,09 mg og 7,24 ± 0,63 mg mens den askefri tørvægt af S. personatum blev bestemt til 5,32 ± 0,17 mg.
Esfenvaleratopløsningerne blev fremstillet ved at opløse en given mængde stof i 10 ml acetone og derefter lave en fortyndingsserie med demineraliseret vand til de ønskede koncentrationer.
Koncentrationerne blev fremstillet således at tilsætning af 6 ml opløsning per minut gav den ønskede nominelle koncentration. Fremstillingsmetoden betød at acetonekoncentrationen i vandløbsrenderne
varierede mellem 1,9 x 10-8 – 7,4 x 10-5 %.
Efter 1,5 time, og dermed to driftnettømninger, blev pulstilførslen af pesticid indledt. Pesticidpulsen blev tilført fra 1,75 liters lavementposer forbundet med et infusionssæt indstillet til at afgive 6 ml
pesticidopløsning per minut til opstrømsenden af vandløbsrenderne (Fig. 45). Undersøgelserne omfattede scenarier med pulsvarighed på 30 og 150 minutter og koncentrationen varierede mellem 0,2 ng l-1
og 2 µg l-1 (Tabel 27). Undersøgelserne blev gennemført i seks serier, hvor der i hver serie blev gennemført forsøg med to pesticidkoncentrationer. Under serierne blev hver af de to pesticidkoncentrationer
tilført fire vandløbsrender mens de fire resterende vandløbsrender ikke blev tilsat pesticid og fungerede som kontrol. Under enkeltartsforsøgene med G. pulex blev der udført en forsøgsserie, hvor 4
vandløbsrender blev tilsat acetone svarende til koncentrationen under eksponeringen med 2 µg l-1 i 30 minutter og 4 vandløbsrender med acetone koncentrationer som under eksponeringen for 0,0002 µg
l-1 i 150 minutter. I modsætning hertil blev der under samfundsforsøgene tilsat acetone til kontrolrenderne under selve forsøget i en koncentration, der svarede til højeste koncentration anvendt i
eksperimentelrender.

Figur 45. Dosering af pesticidopløsning til vandløbsrender
Tabel 27 Undersøgte eksponeringsscenarier
| |
30 minutters
eksponering
(G. pulex) |
150 minutters
eksponering
(G. pulex) |
150 minutters
eksponering
(Samfund) |
Koncentrationer
(µg l-1) |
0, 0,002, 0,02, 0,2, 2,0 |
0, 0,0002, 0,002 |
0, 0,0002, 0,002, 0,02, 0,2 |
| Antal replikater |
4
kontrol(0): 2x4 |
4 |
4
kontrol (0): 2x4 |
Statistisk metode
Data fra driftundersøgelserne i de semi-naturlige vandløbsrender analyseres ved brug af logrank testen fra overlevelsesanalyse (Campbell & Machin, 1990). Denne test sammenligner driftmønstret til
forskellige tidspunkter for to grupper af observationer. Data for de fire render for hver eksponeringsscenarium er puljet under analysen.
Enkeltartsforsøg med G. pulex
Alle eksponeringsscenarierne under enkeltartsforsøgene med G. pulex medførte signifikant forøget drift (Tabel 28 og 29). Under forsøgene med 30 minutters eksponering driftede 50% ved en nominel pulskoncentration på 0,002 µg l-1 og 90 % ved 0,02 µg l-1, hvorimod alle G. pulex driftede ved koncentrationer på 0,2 og 2 µg l-1 (Figur 46). Størrelsen af driftresponset var således koncentrationsafhængigt men også responstiden var korreleret med pulskoncentrationen. Således var andelen af G. pulex i drift efter fra starten af pesticideksponeringen og 135 minutter frem signifikant mindre ved 0,002 µg l-1 end ved koncentrationerne 0,02, 0,2 og 2 µg l-1 under forsøgene med 30 minutters eksponering (Tabel 28). Samme forsinkelse af responset blev observeret ved 0,0002 µg l-1 under forsøget med 150 minutters pesticideksponering (Figur 47 og Tabel 28).
Figur 46. Akkumuleret drift af G. pulex under enkelt artsforsøgene med 30 minutters esfenvalerat eksponering. Den Akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten.
Tabel 28. Test af forskelle mellem de undersøgte koncentrationer ved 30 minutters eksponering. Værdierne angiver ?2(1) og de korresponderende P-værdier. n angiver antallet af individer tilstede til tiden –0,75 timer.
| Periode: 0 – 22 timer |
Koncentration
(µg l-1) |
0 |
0,002 |
0,02 |
0,2 |
n |
| 0 |
- |
- |
- |
- |
90/93 |
| 0,002 |
46,5 P<0,01% |
- |
- |
- |
92 |
| 0,02 |
115,3 P<0,01% |
37,6 P<0,01 |
- |
- |
96 |
| 0,2 |
135,8 P<0,01% |
53,7 P<0,01% |
2,4 P=12% |
- |
94 |
| 2 |
149,8 P<0,01% |
65,2 P<0,01% |
6,5 P=1,0% |
2,11 P=15% |
104 |
| Periode: 0 – 135 min |
Koncentration
(µg l-1) |
0,002 |
0,02 |
0,2 |
- |
- |
| 0,02 |
32,3 P<0,01% |
- |
- |
- |
- |
| 0,2 |
50,9 P<0,01% |
2,02 P=15% |
- |
- |
- |
| 2 |
78,9 P<0,01% |
11,3 P=0,08% |
1,41 P=23% |
- |
- |
Under forsøget med 150 minutters esfenvalerat eksponering driftede 50% af individerne ved eksponering for 0,0002 µg l-1 (0,2 ng l-1) og 95% af G. pulex driftede ud af vandløbsrenderne eksponeret for
0,002 µg l-1 (Fig. 47). Sammenholder man resultaterne fra forsøgene med 30 og 150 minutters eksponering ved 0,002 µg l-1 ses det, at driften øges fra 50 til 95% ved den lange eksponering. Desuden
svarede driftresponset, målt som akkumuleret drift ved forsøgets afslutning, under 150 minutters eksponeringen cirka til responset under 30 minutters eksponeringen efter eksponering for ti gange så høje
koncentrationer. Driftresponset er altså ikke kun afhængig af koncentration men også varighed af eksponeringen.
Figur 47 Akkumuleret drift af G. pulex under enkelt artsforsøget med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten.
Tabel 29, Test af forskelle mellem de undersøgte koncentrationer ved 150 minutters eksponering. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
| Periode: 0 – 22 timer |
| Koncentration (µg l-1) |
0 |
0,0002 |
n |
| 0 |
- |
- |
72 |
| 0,0002 |
35,8 P<0,01% |
- |
68 |
| 0,002 |
100,4 P<0,01% |
64,4 P<0,01 |
93 |
| Periode: 0 – 135 min |
| Koncentration (µg l-1) |
0 |
0,002 |
|
| 0 |
- |
- |
- |
| 0,0002 |
0,0013 P=97% |
- |
- |
| 0,002 |
73,0 P<0,01% |
70,0 P<0,01% |
- |
Acetoneeksponering af G. pulex påvirkede ikke driftraten selv ved den højeste acetonekoncentration benyttet under esfenvaleratforsøgene (Figur 48; Tabel 30). Den forhøjede drift under eksponeringen for esfenvaleratopløsninger kan altså udelukkende tilskrives effekt af pesticidet. Dog kan det ikke udelukkes at der findes synergistiske effekter af esfenvalerat og acetone, selvom litteraturen ikke beskriver
sådanne effekter.
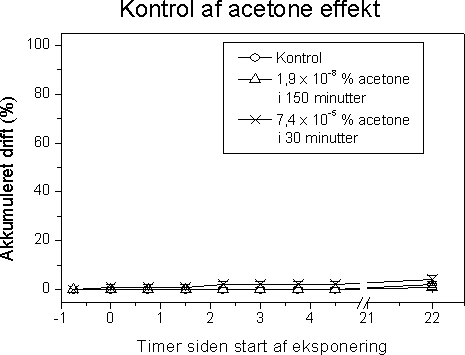
Figur 48. Akkumuleret drift af G. pulex under kontrol forsøgene med acetone eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af acetone eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten.
Tabel 30. Test af effekten af acetone eksponering på driften blandt G. pulex. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
| Periode: 0 – 22 timer |
| Koncentration (µg l-1) |
0 |
1,9 x 10-9 |
n |
| 0 |
- |
- |
88 |
| 1,9 x 10-9 |
0,33 P=56% |
- |
88 |
| 7,3 x 10-5 |
0,44 P=51% |
0,89 P=34% |
95 |
Samfundsforsøg
Under samfundsforsøgene blev der observeret store forskelle i arternes tendens til at indtræde i drift under esfenvalerateksponering. Andelen af individer i drift var under alle eksponeringer, G. pulex > B.
rhodani > S. personatum.
Driftraten hos vårfluen S. personatum blev hverken påvirket under eller efter nogen af pesticideksponeringerne (Figur 49). Natten var det eneste tidsrum, hvor der blev fanget S. personatum i driftnettene
men denne drift var ikke større efter pesticid eksponering. Den udprægede natdrift kan tilskrives at S. personatum primært søger føde om natten.
Figur 49 Akkumuleret drift af S. personatum under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten.
Døgnfluen B. rhodani responderede på esfenvalerat eksponeringen med forøget drift (Figur 50A). Dog var der intet driftrespons ved eksponering for 0,0002 µg l-1 og driften var først signifikant forhøjet ved
eksponering for 0,02 µg l-1 (Tabel 31). Driften var signifikant forhøjet efter eksponeringen for 0,02 og 0,2 µg l-1 men ingen af koncentrationerne medførte en drift på mere end 60% af individerne mod cirka
10% i kontrolrenderne.
Tabel 31 Test af forskelle driften af B. rhodani ved de undersøgte koncentrationer. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
| Periode: 0 – 20 timer |
| Koncentration (µg l-1) |
0 |
0,0002 |
0,002 |
0,02 |
0,2 |
| 0 |
- |
0,13 P=72% |
2,4 P=12% |
13,0 P=0,03% |
10,32 P=0,13% |
| n |
38/26 |
37 |
41 |
30 |
27 |
Figur 50A Akkumuleret drift af B. rhodani under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error. Bemærk bruddet på x-aksen om natten.
Driftresponset observeret blandt G. pulex under samfundsforsøgene var i god overensstemmelse med mønstret observeret under enkeltartsforsøgene (Figur 50B, Tabel 32). Eksponering for 0,2 ng l-1 medførte også under samfundsforsøgene, at ca. 50 % af individerne gik i drift og eksponering for koncentrationer på 0,002, 0,02 og 0,2 µg l-1 forårsagede 100 % drift blandt G. pulex. Ligeledes blev der i samfundsforsøgene observeret en klar negativ korrelation mellem eksponeringskoncentration og start på drift respons.
Figur 50B Akkumuleret drift af G. pulex under samfundsforsøgene med 150 minutters esfenvalerat eksponering. Den akkumulerede drift er afbilledet som procent af individerne tilstede til tiden ÷0,75 timer (n), hvor tiden 0 timer indikerer start af esfenvalerat eksponering. Punkterne angiver gennemsnittet af de fire replikater med standard error.
Tabel 32 Test af forskelle i driften af G. pulex ved de undersøgte koncentrationer efter 150 min eksponering. Værdierne angiver ?2(1) og P-værdien. n angiver antallet af individer tilstede til tiden –0,75 timer.
| Periode: 0 – 20 timer |
| Koncentration (µg l-1) |
0 |
0,0002 |
0,002 |
0,02 |
0,2 |
| 0 |
- |
43,5 P<0,01% |
103 P<0,01% |
91,9 P<0,01% |
84,3 P<0,01% |
| n |
85/83 |
84 |
87 |
67 |
71 |
Sammenligner man arternes driftrespons ved de enkelte koncentrationer fremkommer et klart billede med G. pulex som responderer med en signifikant større andel individer i drift end både B. rhodani og
S. personatum ved alle de testede koncentrationer (Tabel 33). Andelen af B. rhodani i drift var dog signifikant større end andelen af S. personatum ved samtlige esfenvalerateksponeringer (Tabel 33).
Tabel 33. Test af forskelle i driften hos arterne ved de enkelte koncentrationer fra start af eksponering og resten af forsøget (0 – 20 timer). Værdierne angiver ?2(1) og de korresponderende P-værdier.
| Koncentration |
S. personatum |
B. rhodani |
| 0,0002 |
B. rhodani |
5,08 P=2,4% |
- |
| G. pulex |
30,5 P<0,01% |
16,05 P<0,01% |
| 0,002 |
B. rhodani |
6,7 P=0,94% |
- |
| G. pulex |
57,3 P<0,01% |
56,0 P<0,01% |
| 0,02 |
B. rhodani |
5,6 P=1,8% |
- |
| G. pulex |
41,0 P<0,01% |
28,5 P<0,01% |
| 0,2 |
B. rhodani |
19,8 P<0,01% |
- |
| G. pulex |
39,4 P<0,01% |
23,8 P<0,01% |
Sammenligner man driftresponset hos G. pulex og B. rhodani fra starten af pesticideksponeringen og de følgende 135 minutter fremgår det klart at G. pulex responderer langt hurtigere end B. rhodani på esfenvalerat (Tabel 34).
Tabel 34. Test af forskelle i drift hos G. pulex og B. rhodani under de første 135 minutter efter start af pesticideksponering. Værdierne angiver ?2(1) og de korresponderende P-værdier.
Koncentration
|
|
G. pulex |
| 0,0002 |
B. rhodani |
3,12 P=7,8% |
| 0,002 |
B. rhodani |
49,0 P<0,01% |
| 0,02 |
B. rhodani |
34,2 P<0,01% |
| 0,2 |
B. rhodani |
31,7 P<0,01% |
4.8 Nedbrydning af detritus
Vandløbsinvertebraters nedbrydning af detritus under påvirkning af pesticider blev undersøgt under laboratorieforhold i klimarum. Forsøgene fokuserede på hastigheden hvormed krebsdyret Gammarus
pulex (L) nedbrød bladplader af rødel (Alnus glutinósa) efter eksponering for forskellige pesticider. Undersøgelserne omfattede studier, hvor G. pulex blev direkte eksponeret til insekticiderne esfenvalerat
og dimethoat samt fungicidet azoxystrobin i pulse af 30 og 150 minutters varighed inden de blev overført til forsøgsopstillingen. Desuden blev der gennemført undersøgelser af effekten af indirekte
eksponering af G. pulex for esfenvalerat og azoxystrobin gennem elleblade eksponeret for pesticiderne.
4.8.1 Metode
Forsøgsopstilling
Forsøgsopstillingen bestod af 75 nummererede 100 ml bægerglas. Bægerglassene var fyldt med 80 ml kildevand gennemluftet af en iltsten (Figur 51). Desuden indeholdte hvert bægerglas en G. pulex og
fødekilde i form af to bladplader af rødel med en diameter på 10 mm. Undersøgelserne foregik i klimarum hvor temperaturen blev holdt konstant på 10°C og lyset blev styret i 12 timers lys-/mørke-cykler.

Figur 51 Forsøgsopstillingen benyttet under forsøgene med nedbrydning af detritus. Hvert 100 ml bæger glas indeholder 80 ml vand, som bliver genemluftet af iltsten, 2 ellebladplader og en G. pulex.
Blade
Ellebladene var samlet i september direkte fra træerne lige inden løvfald og opbevaret ved 18°C. To til tre uger før brug blev der udstanset bladcirkler med en diameter på 10 mm. Bladcirklerne blev herefter
konditioneret i 4 liter iltet kildevand ved 10°C. Konditioneringen fandt sted for at kolonisere bladpladerne med mikroorganismer, hvilket øger deres fødeværdi for makroinvertebrater (Friberg & Jacobsen,
1999). Efter 2-3 uger i kildevand og umiddelbart inden forsøget blev bladpladerne afduppet parvis mellem to stykker filter papir ved standadiseret tryk i 5 sekunder og vejet. Det afvejede bladpladepar blev
herefter overført til et bægerglas i forsøgsopstillingen.
Forsøgsdyr
Alle G. pulex blev samlet i Hagenstrup Møllebæk (tilløb til Gudenåen) få hundrede meter nedstrøms for kildefeltet. Individerne til forsøgene blev udvalgt så størrelsesvariationen var så lille som mulig. Middel tørvægten af de anvendte G. pulex var 3,79 ± 0,04 mg. De benyttede G. pulex blev aklimatiseret til 10°C i minimum ti dage inden forsøgene og de sidste tre dage inden forsøgene gik de uden mad.
Forsøgene blev gennemført med to insekticider: dimethoat og esfenvalerat samt et fungicid: azoxystrobin. Pesticidopløsningerne til pulseksponering af dyrene blev fremstillet med demineraliseret vand. Dog
blev det aktive stof opløst i 10 ml acetone under fremstillingen af pesticidopløsningerne med de tungt opløselige stoffer esfenvalerat og azoxystrobin. Efterfølgende blev acetoneopløsningen fortyndet i
demineraliseret vand så der maksimalt var 0,1 µg acetone i opløsningen dyrene blev udsat for. Samtidig blev kontroldyrene/bladene uden pesticid også tilsat 0,1 µg acetone under eksponeringen.
Direkte eksponering
I forsøgene, hvor G. pulex blev direkte eksponeret for pesticidet blev de umiddelbart før overførslen til forsøgsopstillingen neddyppet i pesticidopløsningen i 30 eller 150 minutter. Under eksponeringen var
dyrene fanget i små akvarienet, hvor nettet hang ned i bægerglas fyldt med en liter pesticidopløsning gennemluftet af en iltsten. Under forsøgene med direkte eksponering blev G. pulex udsat en serie af
koncentrationer af de undersøgt pesticider (Tabel 35). Efter pesticideksponeringen blev nettet med dyr overført til et 1 liters bægerglas med rent vand gennemluftet af en iltsten. Dyrene opholdt sig 5-10
minutter i det rene vand for, at skylle eventuelle pesticidrester af inden de blev overført til forsøgsopstillingen.
Indirekte eksponering
Den indirekte eksponering af dyrene fandt sted ved at neddyppe bladene i pesticidopløsning inden de blev overført til forsøgsopstillingen. Under bladeksponeringen blev 50 bladplader efter to ugers
konditionering i kildevand overført til hvide fotobakker og overhældt med en liter pesticidopløsning. Bladpladerne forblev i pesticidopløsningen i syv dage inden de blev afduppet, vejet og overført til
forsøgsopstillingen
Tabel 35. Oversigt over de undersøgte eksponeringsscenarier
| |
esfenvalerat |
dimethoat |
azoxystrobin |
| Direkte eksponering |
Eksponeringsvarighed
(dyr) |
30 min. |
150 min. |
30 min. |
150 min. |
30 min. |
150 min. |
Koncentrationer
(µg l-1) |
0, 0,002, 0,02, 0,2, 2 |
0, 0,0002, 0,002, 0,02, 0,2 |
0, 0,002, 0,02, 0,2, 2, 20, 200, 2000 |
0, 0,02, 0,2, 2, 20, 200, 2000 |
0, 2, 20, 200, 2000 |
0, 2, 20, 200, 2000 |
| Antal replikater |
15 |
15 |
15 |
15 |
15 |
15 |
| Indirekte ekspon. |
Eksponeringsvarighed
(Blade) |
7 dage |
- |
7 dage |
Koncentrationer
(µg l-1) |
0, 0.,002, 0,02, 0,2, 2 µg l-1 |
- |
0, 2, 20, 200, 2000 µg l-1 |
| Antal replikater |
15 |
- |
15 |
Hvert kontamineringsscenarium blev gennemført med 15 replikater. Under forsøgene blev dyrenes tilstand observet dagligt. Forsøget blev afsluttet efter syv dage, ved afslutningen blev bladresterne fra hvert
bægerglas overført til forvejede foliedigler og tørret ved 60°C til konstant vægt. Længden af første thorakalled af G. pulex blev målt og tørvægt blev fundet ud fra en kendt relation mellem længde og tørvægt
(Iversen & Jessen, 1977).
Sideløbende med forsøgene blev 50 bladpladepar konditioneret og vejet som under de øvrige forsøg. Bægerglassene de blev overført til indeholdt dog ingen dyr, hvorfor forholdet mellem vådvægt før og
tørvægt efter forsøg uden påvirkning af dyr kunne etableres.
4.8.2 Resultater
Hastigheden hvormed krebsdyret G. pulex nedbrød groft detritus (CPOM) til fint detritus (FPOM) efter pulseksponering for pesticider var generelt nedsat.
Nedbrydningshastigheden efter eksponering for insekticidet esfenvalerat faldt med stigende eksponerings koncentration (Figur 52). Nedbrydningshastigheden faldt efter 30 minutters eksponering for 0,02 µg l-1 esfenvalerat og faldet var signifikant efter eksponering for 0,2 µg l-1 (Tabel 36). Det meget kraftige fald i nedbrydningshastighed efter eksponering for 2 µg l-1 skyldes primært, at 14 af 15 individer døde efter overførsel fra esfenvaleratopløsningen til forsøgsopstillingen. At bladvægttabet kan være negativt skyldes primært to ting: 1) Ikke to bladpar er ens og mængden af nervation i bladpladerne vil have betydning for det forhold vi etablerede mellem vådvægt før og tørvægt efter, 2) Mængden af mikroorganismer på bladene vil øges med tid og biomassen af disse mikroorganismer påvirkes af tilstedeværelsen af G. pulex.
Efter 150 minutters eksponering med esfenvalerat blev der også observeret et fald i nedbrydningshastigheden med stigende koncentration. Faldet var dog ikke signifikant på 5 % niveau efter eksponering for 0,2 µg l-1 (Tabel 36).
Faldet i nedbrydningshastighed var ikke entydigt blandt de testede insekticider da stoffet dimethoat ikke påvirkede nedbrydningshastigheden selv efter eksponering for 2000 µg l-1 (Bilag E).
Klik her for at se Figur 52.
Tabel 36. Test af forskel fra kontrol under forsøg med esfenvalerat samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de
korresponderende P-værdier.
| |
Test |
Antal replikater |
Dødelighed
Antal døde/forsvundne |
| 30 minutters ekspon. |
| Koncentration (µg l-1) |
Kontrol |
15 |
2 / 0 |
| 0,002 |
0,12; P=74% |
15 |
1 / 0 |
| 0,02 |
3,62; P=6,1% |
15 |
1 / 0 |
| 0,2 |
16,23; P=0,01% |
15 |
2 / 0 |
| 2 |
68,69; P<0,01% |
15 |
14 / 0 |
| 150 minutters ekspon. |
| Koncentration (µg l-1) |
Kontrol |
15 |
1 / 1 |
| 0,0002 |
1,32; P=25% |
15 |
0 / 0 |
| 0,002 |
0,95; P=33% |
15 |
1 / 0 |
| 0,02 |
0,00; P=97% |
15 |
2 / 0 |
| 0,2 |
1,87; P=18% |
15 |
1 / 0 |
Pulseksponering med fungicidet azoxystrobin forårsagede et fald i nedbrydningshastigheden hos G. pulex (Figur 53). Ved 30 minutters eksponering var falder signifikant efter eksponering for 20 µg l-1
(Tabel 37). Det kraftige fald i nedbrydningshastigheden efter eksponering for 2000 µg l-1 skyldes primært at 12 af 15 individer døde efter overførsel fra pesticidopløsningen til forsøgsopstillingen (Tabel 37).
Nedbrydningshastigheden efter 150 minutters eksponering var også negativt korreleret med azoxystrobin koncentrationen dog uden at nogen af koncentrationerne medførte et signifikant fald
nedbrydningshastigheden på 5 % niveau (Tabel 37).
Klik her for at se Figur 53.
Tabel 37. Test af forskel fra kontrol under forsøg med azoxystrobin eksponeret til G. pulex samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de korresponderende P-værdier.
| |
Test |
Antal replikater |
Dødelighed
Antal døde/forsvundne |
| 30 minutters ekspon. |
| Koncentration (µg l-1) |
Kontrol |
15 |
0 / 0 |
| 2 |
2,86; P=9,5% |
15 |
1 / 0 |
| 20 |
5,97; P=1,7% |
15 |
1 / 0 |
| 200 |
3,86; P=5,3% |
15 |
0 / 0 |
| 2000 |
12,42; P=0,08% |
15 |
12 / 0 |
| 150 minutters ekspon. |
| Koncentration (µg l-1) |
Kontrol |
15 |
0 / 1 |
| 2 |
1,56; P=22% |
15 |
1 / 0 |
| 20 |
2,70; P=11% |
15 |
0 / 0 |
| 200 |
3,88; P=5,4% |
15 |
3 / 0 |
Under forsøgene med indirekte eksponering var G. pulex ikke eksponeret for pesticiderne i vandfasen men derimod gennem deres fødeemne - elleblade. Denne form for eksponering medførte nedsat nedbrydningshastighed under forsøgene med blade eksponeret for esfenvalerat (Figur 54).
Nedbrydningshastigheden var signifikant nedsat ved eksponering af bladene for 0,002 µg l-1, hvorimod nedbrydningshastigheden ved blade eksponeret for 0,02 µg l-1 ikke var signifikant mindre end
kontrolforsøgene (Tabel 38). Nedbrydningshastigheden var yderligere kraftigt reduceret under forsøgene med blade eksponeret for 0,2 og 2 µg l-1 (Figur 54). I ingen af forsøgene døde mere end et individ
per eksponeringskoncentration og faldet i nedbrydningshastighed er altså ikke et artefakt af dødelighed blandt G. pulex som følge af pesticideksponering (Tabel 38).
Forsøgene med indirekte eksponering af G. pulex for azoxystrobin gennem føden medførte ingen signifikant ændring af nedbrydningshastigheden (Bilag E).
Figur 54 Nedbrydningen af detritus efter syv dages eksponering af elleblade i esfenvalerat er afbilledet som bladpladernes vægttab per mg dyr mod eksponeringskoncentration. Søjlediagrammerne angiver gennemsnittet af de 15 replikater med standard error. * indikerer at bladvægttabet er signifikant forskelligt fra kontrollerne (F-test, p<0,05)
Tabel 38 Test af forskel fra kontrol under forsøg med esfenvalerat eksponering af blade (7 dage) samt oversigt over dødelighed, hvor forsvundne er individer der er kravlet oven ud af glasset. Værdierne angiver F-værdien samt de korresponderende P-værdier.
| |
Test |
Antal replikater |
Dødelighed
Antal døde/forsvundne |
| Koncentration (µg l-1) |
Kontrol |
15 |
1 / 0 |
| 0,002 |
5,61; P=2,1% |
15 |
1 / 0 |
| 0,02 |
2,03; P=16% |
15 |
0 / 0 |
| 0,2 |
8,72; P=0,43% |
15 |
1 / 0 |
| 2 |
40,32; P<0,01% |
15 |
1 / 1 |
| Forside | | Indhold | | Forrige | | Næste | | Top |
Version 1.0 Januar 2004, © Miljøstyrelsen.
|