Fungiciders påvirkning af mykorrhizasvampes diversitet og funktion
4 Fungiciders indflydelse på mykorrhiza i parvise kombinationer mellem planter og mykorrhizasvampe
- 4.1 Dosis-responsmålinger med forskellige arter af mykorrhizasvampe
- 4.2 Effekt af fungicider på forskellige genotyper (isolater) af samme mykorrhizasvampeart
Tidligere undersøgelser har vist, at effekten af et givet fungicid kan variere mellem forskellige mykorrhizasvampe (Dodd & Jeffries, 1989); (Schreiner & Bethlenfalvay, 1997). Det er ikke undersøgt, hvorledes sådanne forskelle afhænger af fungiciddosis, og om forskelle i fungicidfølsomhed også kan erkendes inden for forskellige genotyper af den samme mykorrhizasvampeart. Disse spørgsmål er belyst i dette kapitel med anvendelse af svampe isoleret fra forsøgsmarken i Tåstrup.
4.1 Dosis-responsmålinger med forskellige arter af mykorrhizasvampe
4.1.1 Formål
Det var formålet at eksponere forskellige mykorrhizasvampe fra forsøgsmarken for forskellige doser mancozeb og carbendazim med henblik på at teste følgende hypoteser: (i) forskellige mykorrhizasvampe reagerer forskelligt på en given fungicidbehandling, (ii) der er forskel på skadevirkningen af carbendazim og mancozeb og (iii) fungicideffekter på mykorrhizasvampe er ikke målbare ved doseringer lavere end No Observed Adverse Effect Level (NOAEL). De opnåede resultater skulle i øvrigt anvendes til at fastlægge fungicid-dosering i de øvrige projektaktiviteter.
4.1.2 Eksperimentelt
Eksperimentet blev gennemført som et potteforsøg med Solare-ært og fire mykorrhizasvampe fra forsøgsmarkens økologiske sædskifte: Glomus claroideum (Schenck & Smith) (BEG88), G. mosseae (Nicol. & Gerd.) Gerdemann & Trappe (BEG84), G. caledonium (Nicol. & Gerd.) Trappe & Gerd (H07-1) og G. geosporum (Nicol. & Gerd.) Walker (BEG90). BEG-numre refererer til svampens nummer i den europæiske kultursamling af mykorrhizasvampe, International Bank for the Glomeromycota. Svampene blev opformeret i potter med jordkløver (Trifolium subterraneum L.) og jord fra disse kulturer blev anvendt som inokulum.
Ærteplanter blev dyrket i potter med 1000g bestrålet jord (10 MeV elektronstråling, 15 kGy) som blev tilført alle nødvendige plantenæringsstoffer, bortset fra fosfat (Pearson & Jakobsen, 1993). Bestrålingen var nødvendig for eliminering af de naturligt forekommende mykorrhizasvampe og for dermed at kunne gennemføre kontrollerede studier af fungiciders indflydelse på specifikke svampe. Nogle mikroorganismer overlever bestrålingen, og andre vil kolonisere potterne i løbet af vækstperioden. Jorden er derfor ikke steril. Jorden til de enkelte potter blev blandet med 50 g inokulum af en af de fire mykorrhizasvampe og andre potter modtog ikke inokulum. Hver af de fem mykorrhizabehandlinger blev kombineret med syv fungicidbehandlinger: ubehandlet, mancozeb (1, 5 eller 25 x markdosis) og carbendazim (0,01; 0,1 eller 1 x markdosis). Den valgte dosering af mancozeb var baseret på indledende undersøgelser, der tydede på, at mykorrhiza ikke blev påvirket af mancozebdoser lavere end markdosis. Tidligere studier har derimod vist, at carbendazim-doser lavere end markdosis har en skadelig effekt på mykorrhizasvampe (Schweiger & Jakobsen, 1998). Doseringerne blev opnået ved iblanding af 3.3, 16.7 og 83,3 µg mancozeb g-1 jord samt 0,006; 0,060 og 0,602 µg carbendazim g-1 jord. Der var fire gentagelser for hver behandling. Det beskedne antal doserings-niveauer tillod ikke en optimal analyse af dosis-respons forløb fx ved hjælp af Gompertz regressionsmodellen. Resultaterne er derfor analyseret ved hjælp af variansanalyse.
Jorden i det rodfri hyfekammer blev blandet med 5 kBq carrier-fri 33PO4 g-1 og kammeret blev placeret midt i potten. Der var 3 gentagelser for hver behandling, og de 95 potter blev placeret i vækstkammer med 16 t lys ved 21°C og 8 t mørke ved 16°C. Forsøget blev høstet 39 dage efter udplantning af spirede ærtefrø, og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, længde af hyfer i 33P-mærket jord samt planternes indhold af 33P og totalt P. Kolonisering af rødder med G. geosporum var 7 % eller mindre og dermed for lav til at en troværdig måling af fungicideffekter kunne opnås. G. geosporum data er derfor udeladt i det følgende.
4.1.3 Resultater og diskussion
Uden fungicidtilførsel havde 28 - 42 % af rodlængden udviklet mykorrhiza, og de to laveste doser af begge fungicider forøgede markant koloniseringen hos alle tre svampe (P < 0,001), medens den højeste dosis af begge fungicider førte til en reduceret mykorrhizadannelse i forhold til ubehandlet (Figur 4.1).
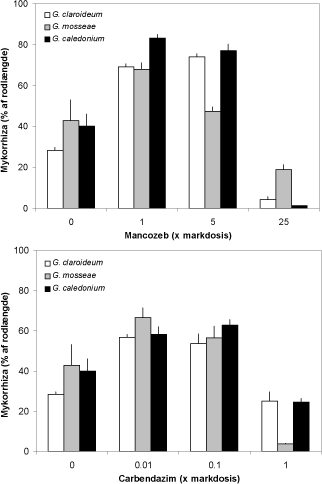
Figur 4.1. Effekt af forskellige doser af mancozeb og carbendazim på rodkolonisering hos ært dyrket med tre forskellige mykorrhizasvampe. Middelværdier og standardfejl er angivet.
Dosis-respons kurven havde således et omvendt U- eller J-forløb hos alle tre svampe, og kurvens form afhang af både svamp og fungicid. Mancozeb førte generelt til den kraftigste stimulering, og effekten af mancozeb på G. mosseae var anderledes end effekten på de to øvrige svampe. Dosering med 5x mancozeb førte til en reduceret G. mosseae-kolonisering, men ved den højeste dosering blev de to andre svampe hæmmet mest. I modsætning hertil, havde den højeste carbendazim-dosis en kraftigere negativ effekt på G. mosseae end på de to øvrige svampe.
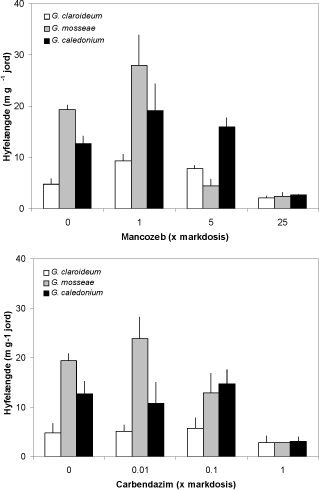
Figur 4.2. Effekt af stigende dosis af mancozeb og carbendazim på længden af rod-eksterne hyfer produceret af tre forskellige mykorrhizasvampe dyrket i symbiose med ært. Hyfelængden blev målt i 0-1 cm jordlaget tættest på nylonnettet, der adskilte den 33P-mærkede jord fra resten af pottevolumen. Middelværdier og standardfejl er angivet.
Det høje niveau af begge fungicider havde en stærkt hæmmede effekt på hyfevæksten af alle tre svampe (P < 0,001), og de lave værdier i højre side af Figur 4.2svarede til den målte hyfelængde i jord med mykorrhiza-fri kontrolplanter. Svampemyceliet kolonisering af den 33P-mærkede jord varierede i øvrigt, således at G. mosseae opnåede de største hyfelængder, men G. mosseae var samtidig den eneste af de tre svampe der reagerede negativt på de næsthøjeste doser af begge fungicider. De to lave fungiciddoseringer havde ikke nogen effekt på hyfelængden, bortset fra en stimulering af G. claroideum ved 1 x mancozeb. Data i Figur 4.2 stammer fra 0-1 cm jordlaget tættest på rødderne, men samme tendens blev observeret i 1-4 cm jordlaget (data ikke vist), hvor G. caledonium producerede den største hyfelængde uden fungicidbehandling (~50 m g-1 jord). Til gengæld blev G. caledonium signifikant hæmmet af ikke blot det højeste fungicidniveau men også af de to laveste niveauer af begge fungicider. Spredning af G. mosseae i dette lag blev også hæmmet af den midterste dosering af begge fungicider.
De lave niveauer af fungicidtilførsel stimulerede ikke blot svampenes vækst i rødder og i jord, men også svampenes optagelse af fosfat. Uden fungicid tilførsel var G. caledonium den eneste svamp, der optog signifikant mere 33P end niveauet i den mykorrhizafri kontrol, hvor 33P optagelsen var 0,26 kBq (Fig. 4.3). Markdosis
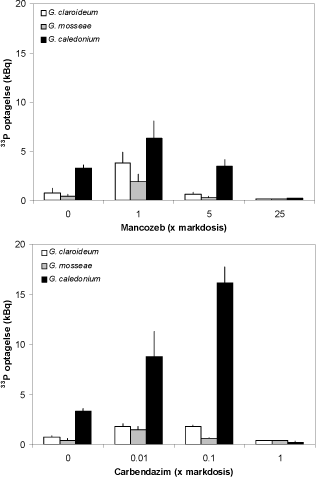
Figur 4.3. Effekt af stigende dosis af mancozeb og carbendazim på 33P-indhold hos ærteplanter, som blev dyrket i symbiose med tre forskellige mykorrhizasvampe. 33P er transporteret til planten via mycelium der vokser gennem nylonnettet og ind i den radioaktive jord. Middelværdier og SE er angivet.
af mancozeb resulterede i en general forøgelse af 33P optagelsen (P < 0,001 for alle svampe), og for G. claroideum var der tale om en 4x forøgelse (P < 0,01). En 5 x markdosis af mancozeb havde ikke nogen effekt på 33P optagelsen, medens en 25x dosis fuldstændig hæmmede svampenes 33P optagelse (P < 0,001). De laveste carbendazim doser forøgede ligeledes optagelsen af 33P (P < 0,001) og denne effekt var særlig udtalt hos G. caledonium, hvor optagelsen ved 0,1 x markdosis var 4,5 x så høj som i fraværet af fungicider. Markdosis af carbendazim hæmmede fuldstændigt optagelsen af 33P uptake (P = 0,02) i overensstemmelse med hæmningen af hyfevækst (P < 0,001).
Planternes vækst og totale P optagelse blev ikke signifikant påvirket af mykorrhiza, hvorimod 25 x mancozeb markdosis havde en fytotoksisk effekt (data ikke vist). Det kan ikke udelukkes, at den reducerede plantevækst ved 25 x mancozeb har påvirket etablering og funktion af mykorrhiza. Mancozeb i 5 x dosis havde en negativ effekt på P indholdet i symbiosen med G. mosseae, medens markdosis af carbendazim havde en generelt negativ indflydelse på planternes totale P indhold (data ikke vist).
En stimulerende effekt af en lav fungiciddosis blev bekræftet af resultater opnået i efterfølgende eksperimenter, hvor 1,0 x markdosis carbendazim blev anvendt som referencebehandling (se afsnit 4.2 og kapitel 5).
De opnåede resultater bekræfter, at mykorrhizasvampe er følsomme i forskellig grad for behandling af jorden med fungicider (Dodd & Jeffries, 1989); (Schreiner & Bethlenfalvay, 1997). Nærværende studie udvider denne erkendelse til at gælde hyfernes vækst i jorden og især hyfernes transport af P til planten. Resultaterne viser tydeligt, at skadetærskelen (NOAEL) er langt højere for behandling med mancozeb end med carbendazim. Det skal bemærkes, at denne rapport estimerer NOAEL vha. ekstrapolering af responskurver til nul-behandlings niveauet. Resultaterne kunne derimod ikke bekræfte hypotesen om, at fungiciddoser lavere en NOAEL ikke påvirker svampene. Små mængder fungicid gav således en markant stimulering af mykorrhizasvampes vækst og P optagelse. Dette er tidligere antydet for lave doser af carbendazim (Schweiger & Jakobsen, 1998) og for lave doser af en fenpropimorph-propiconazol blanding under markforhold (Schweiger et al., 2001). Der er flere mulige forklaringer på den uventede stimulering af mykorrhizasvampene ved de lave fungiciddoser. En mulighed er at mykorrhizasvampenes funktion påvirkes negativt af andre mikroorganismer (McAllister et al., 1997), og en relativt lavere skadetærskel (NOAEL) for eventuelle antagonister ville dermed kunne resultere i en positiv effekt af små fungiciddoser på mykorrhizasvampene. Denne forklaringsmodel forekommer dog mindre sandsynlig, idet lavdosis-stimuleringerne er blevet observeret med to fungicider i en række uafhængige eksperimenter og med anvendelse af forskellige mykorrhizasvampe. Forekomsten af eventuelle antagonistiske grupper må forventes at have varieret betydeligt imellem de nævnte eksperimenter.
En anden, og mere sandsynlig forklaring er, at de observerede stimuleringer skyldes hormesis, som er et velkendt økotoksikologisk fænomen (Calabrese, 2005). Hormesis (’excite’ eller stimulere) betegner den situation, hvor toksiner har en gunstig indflydelse på en organismes vækst, reproduktion eller levetid (Calabrese & Baldwin, 2003). Den biologiske baggrund for hormesis er ikke klarlagt. Der kunne være tale om en direkte effekt, men det er mere sandsynligt, at hormesis skyldes en over-kompensation som reaktion på en forstyrrelse af homeostasis (Trewavas & Stewart, 2003; Calabrese, 2005).
En sikker påvisning af hormesis kræver, at dosis-respons studier inddrager et tilstrækkeligt antal sub-NOAEL doser, samt at effekten af de lave doser studeres over tid. Et typisk hormesis-forløb viser en tidlig negativ effekt af lave toksin-doser, hvorefter organismens kompensering/overkompensering fører til at toksineffekten fremtræder som gunstig for organismen (Calabrese, 2005). Den klokkeformet ’sub-NOAEL’-responskurve, der i nærværende studie blev observeret for flere af de målte variable, svarer til resultater af dosis-respons studier af en lang række toksiner på stort set alle grupper organismer, herunder svampe (Calabrese, 2005). Hormesis-effekter på mykorrhizasvampe kan entydigt undersøges i dosis-respons studier på petriskål-baserede kulturer af rødder og mykorrhizasvampe (se (Nielsen et al., 2002). Disse kulturer indeholder ikke andre mikroorganismer end mykorrhizasvampe og er velegnede til direkte studier af tidsforløb.
Dosis-respons mønsteret var i nogle tilfælde væsentligt påvirket af arten af mykorrhizasvamp. Dette eksemplificeres tydeligst af den meget kraftige positive effekt af 0,01 og 0,1 x markdosis carbendazim på 33P optagelsen hos G. caledonium, hvor 1 x markdosis hæmmede P optagelsen fuldstændigt. Det er sandsynligt, at de doser, som mykorrhizasvampene eksponeres for under markforhold, er lavere end markdosis som følge af afgrødens opfangning af størstedelen af et sprøjtemiddel samt midlets binding til jorden i større eller mindre grad. Heraf følger, at, i det omfang mykorrhizasvampe i markjord eksponeres for fungicider, så er der sandsynligvis tale om sub-NOAEL doser, der har positive, snarere end negative effekter.
Hormesis-effekterne på mykorrhizasvampenes vækst og P optagelse er i denne undersøgelse observeret hos planter høstet 39 dage efter såning. Det er derfor ikke muligt at udtale sig om den langsigtede konsekvens af hormesis for svampenes vækst, næringsstoftransport og produktion af sporer. Det er muligt, at dosis-responskurven ændrer form over tid og at den fungicid-inducerede stimulering er et tidsbegrænset fænomen.
4.2 Effekt af fungicider på forskellige genotyper (isolater) af samme mykorrhizasvampeart
4.2.1 Formål
Vi har tidligere påvist en betydelig variation i vækstmønster af 15 isolater af G. mosseae (Munkvold et al., 2004), men det er uvist, om der også eksisterer intraspecifik variation i mykorrhizasvampenes tolerance for fungicider. Dette blev undersøgt ved anvendelse af G. caledonium isolater fra Snubbekorsgård forsøgsmarken samt nogle enkelte reference-svampe. Undersøgelsen testede hypotesen, at effekten af mancozeb og carbendazim ikke blot varierer mellem arter af mykorrhizasvampe men også mellem genotyper af den samme art.
4.2.2 Eksperimentelt
Eksperimentet afveg fra de øvrige i projektet ved at anvende Medicago truncatula som værtplante for mykorrhizasvampene i stedet for ært. Medicago truncatula er en bælgplante ligesom ært, og de to arter er temmelig homologe mht. til de gener der er centrale for mykorrhizasvampes kolonisering af planteroden (Oldroyd et al., 2005). Væksten af M. truncatula er relativt stærkt afhængig af mykorrhiza, og dermed kan eventuelle negative effekter af fungicider monitoreres i løbet af vækstperioden. Dette er ikke muligt hos ært, hvor vækstrespons på mykorrhiza er meget mindre end hos M. truncatula.
Der blev anvendt 7 pottekulturer af G. caledonium: fire enkeltsporekulturer fra mark 29 (K209, K225, K219 og K230), en enkeltsporekultur fra mark 21 (M21), en multisporekultur fra mark 29 (H-07) og en multisporekultur fra UK (BEG20). Enkeltspore- og multisporekultur betyder, at pottekulturen er initieret fra en enkelt eller fra flere sporer. Kun når kulturen er startet fra en enkelt spore, kan man være sikker på, at den kun indeholder en genotype. DNA sekvensering af svampene tyder på at K209 og K225 er en og samme genotype (type 1) medens K219 og K230 repræsenterer en anden genotype (type 2). M21 kulturen er også en type 2 genotype. De to sidste G. caledonium svampe (H-07 og BEG 20) er hverken type 1 eller 2. For at muliggøre sammenligning af fungicideffekter på variation inden for en art med variation imellem arter, blev der inddraget yderligere to svampe: En enkeltsporekultur af G. mosseae fra mark 29 (K218) og en multisporekultur af G. intraradices isolat fra sjællandsk overdrev (BEG87). Svampene blev opformeret i potter med jordkløver (Trifolium subterraneum L.) og jord fra disse kulturer blev anvendt som inokulum.
Medicago truncatula (Jemalong) blev dyrket i potter med 1000 g bestrålet jord tilført næringsstoffer, bortset fra fosfat (Pearson & Jakobsen, 1993). Jorden til hver potte blev iblandet 50 g inokulum af en af de ni svampe eller forblev fri for mykorrhizainokulum. Hver af de 10 mykorrhizabehandlinger blev kombineret med tre fungicidbehandlinger: ubehandlet, mancozeb (0,5 x markdosis) og carbendazim (0,5 x markdosis). Doseringerne blev opnået ved iblanding af 1,67 µg mancozeb g-1 jord samt 0,301 µg carbendazim g-1 jord. Fuld markdosis af carbendazim til ekstra potter med G. intraradices blev anvendt som referencebehandling til kontrol af modelsystemets funktion. Tidligere studier har vist at denne dosis hæmmer mykorrhizasvampes 33P optagelse med mere end 95 % (Schweiger & Jakobsen, 1998). Der blev yderligere inddraget en 0,1 x carbendazim markdosis for at undersøge eventuelle hormesis effekter på G. intraradices.
Jorden i de rodfri hyfeindvækstkamre blev blandet med 5 kBq carrier-fri 33PO4 g-1 samt med den relevante fungicidmængde, og kammeret blev placeret midt i potten. Der var 4 gentagelser for hver behandling, og de 128 potter blev placeret i vækstkammer med 16 t lys ved 21 °C og 8 t mørke ved 16 °C. Der blev tilført 100 mg NH4NO3-N i løbet af vækstperioden. Forsøget blev høstet 48 dage efter udplantning af spirede M. truncatula-frø, og følgende variable blev målt: Plantebiomasse, mykorrhizadannelse i rødder, længde af hyfer i 33P-mærket jord samt planternes indhold af 33P og totalt P.
4.2.3 Resultater og diskussion
Som forventet havde alle mykorrhizasvampe en betydelig effekt på plantevæksten, som i alle tilfælde blev mere end fordoblet (Figur 4.4). Mancozeb havde ikke nogen klar effekt på plantevæksten, hvorimod carbendazim påvirkede væksten negativt i alle tilfælde bortset fra G. intraradices symbiosen (BEG87, ingen effekt) og planter uden mykorrhiza (NM, en svag stimulering).
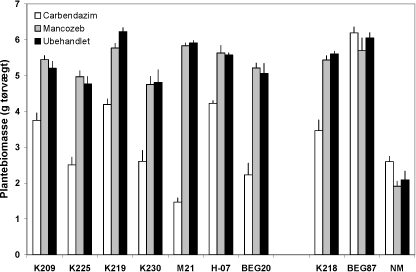
Figur 4.4. Effekt af mancozeb og carbendazim på biomasseproduktion hos M. truncatula dyrket i symbiose med 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218), G. intraradices (BEG87) eller uden mykorrhizapodning (NM). Middelværdier og standardfejl er vist. Effekten af svamp, af fungicidbehandling og interaktionen mellem de to faktorer var signifikant (P < 0,0001).
Resultaterne bekræfter således, at skadeeffekten af et fungicid afhænger af svampearten, og som noget nyt viser de, at effekten også varierer betydeligt inden for den samme art. Isolat M21 var fx betydelig mere følsomt for carbendazim end de to øvrige genotype 2 isolater (K219 og K230). Der var et nøje sammenfald mellem variationer i plantebiomasse (y) og planternes P indhold (x): y = 4,27Ln(x) - 2,36 (R² = 0,93), hvilket er i god overensstemmelse med, at P var eksperimentelt fastlagt som det vækstbegrænsende næringsstof. Det er et vigtigt resultat, at plantearter med en tydelig vækstrespons på mykorrhiza kan anvendes til direkte visualisering af skadeeffekter af fungicider og andre miljøfaktorer på mykorrhizasvampes funktion i næringsstofoptagelsen (Fig. 4.4).
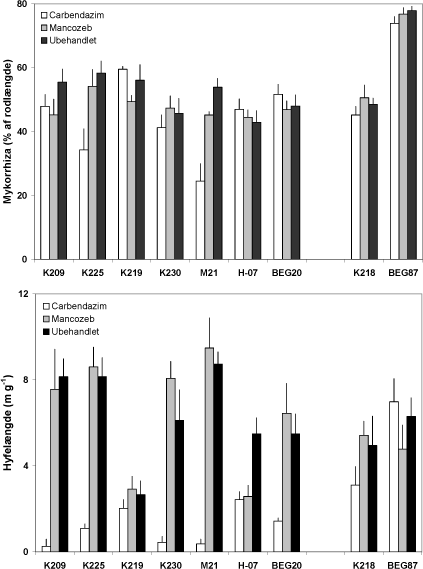
Figur 4.5. Effekt af mancozeb og carbendazim på mykorrhizaudvikling i rødder og i jord i symbioser mellem M. truncatula og 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218), G. intraradices (BEG87). Middelværdier og standardfejl er vist. Signifikansniveauer: mykorrhiza, P < 0,001 for fungicidbehandling og P< 0,0001for både svamp og interaktion; hyfelængde, P< 0,0001 for begge faktorer og for interaktionen.
Effekten af fungicidbehandlingerne blev undersøgt i større detalje ved analyse af svampenes vækst i rødder og jord samt af svampenes optagelse af 33P fra det rodfri mærkningskammer. Mykorrhizaen var veludviklet i rødderne af alle G. caledonium podede planter (44 - 58 % af rodlængden) og G. intraradices (BEG87) gav den kraftigste mykorrhizaudvikling (Figur 4.5). Mancozeb havde en mindre negativ effekt på rodkoloniseringen hos enkelte af G. caledonium isolaterne, medens carbendazim havde en tydelig negativ effekt hos K225 og M21 (forskellige genotyper). Hos de øvrige svampe kunne der ikke observeres nogen skadevirkning af carbendazim på rodkoloniseringen. Planter uden tilførsel af inokulum udviklede ikke mykorrhiza, og NM-behandlingen er derfor udeladt i både Fig. 4.5 og Fig. 4.6.
Hyfelængden i 33P mærket jord fra de mykorrhiza-fri planter var 1.01-1.21 m g-1, og de viste resultater i Figur 4.5 er alle justeret med baggrundsværdien for den respektive fungicidbehandling. Længden af mycelium i den 33P-mærkede jord varierede stærkt inden for G. caledonium isolaterne. Type 1 isolaterne (K209 og K225) var meget ens, hvorimod type 2 isolaterne varierede mellem 2.5 og 9 m hyfer g-1 jord (K219 og M21). Selv genotype 2 enkeltspore-isolaterne K219 og K230 fra samme mark var meget forskellige.
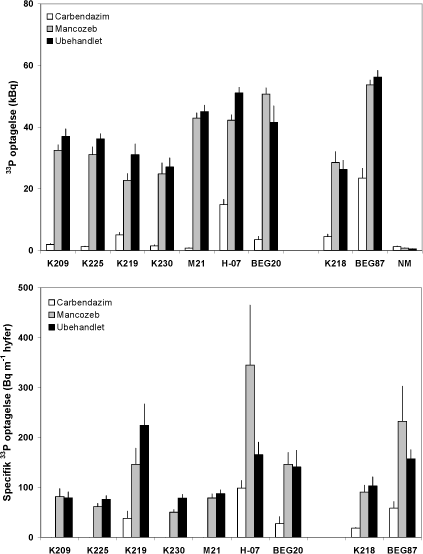
Figur 4.6. Effekt af mancozeb og carbendazim på mykorrhizasvampes totale og hyfelængde-specifikke optagelse og transport af 33P fra jord til M. truncatula, som blev dyrket i symbiose med 7 isolater af G. caledonium (K209-BEG20), G. mosseae (K218) eller G. intraradices (BEG87). Middelværdier og standardfejl er vist. Signifikansniveauer: 33P optagelse, P<0,0001 for begge faktorer og for interaktionen; specifik 33P optagelse, P<0,0001 for både svamp og fungicidbehandling og P<0,01 for interaktion.
Mancozeb gav en svag forøgelse i hyfelængden hos seks af symbioserne, men den eneste signifikante effekt var en reduktion i hyfelængde hos H-07. Carbendazim hæmmede hyfevæksten næsten 100 % hos K209, K230 og M21 symbioserne, men havde en ringe eller ingen effekt hos G. caledonium K219 og G. intraradices BEG87. Hos H-07 hæmmede begge fungicider hyfelængden med ~ 50 %. Der var en stor inter- og intraspecifik variation i fungiciders effekt på mykorrhizasvampenes vækst i jorden. Den mere markante effekt af fungicider på svampens vækst i jord end i rodkoloniseringen bekræfter tidligere resultater (Kjøller & Rosendahl, 2000b), og disse forskelle er formodentlig knyttet til en forskel i eksponeringsgrad over for fungicidet, således at svampen inde i roden i højere grad er beskyttet mod fungicidet end svampemyceliet i jorden.
Mykorrhizasvampene optog forskellig mængde 33P fra den radioaktive jord, også på det intraspecifikke niveau. En af genotype 2 svampene (M21) optog således tydeligt mere end de to øvrige type 2 svampe (K219 og K230), som havde optaget næsten lige meget 33P på trods af en markant mindre hyfelængde hos K219 (Figur 4.5 og 4.6). De to type 1 svampe havde optaget samme mængde 33P. Manzoceb havde en svagt negativ effekt på 33P optagelsen hos de fleste svampe, hvorimod carbendazimn havde en stærkt hæmmende effekt, også hos de svampe, hvor hyfevæksten ikke var påvirket eller blev hæmmet i mindre omfang. Dette illustreres godt i Figur 4.6, der viser 33P optagelsen pr meter hyfe i den 33P mærkede jord. Dette udtryk tager ikke hensyn til eventuelle tidsbetingede forskelle i hyfevæksten. Det fremgår at type 2 svampene K219 og K230 har meget forskellig kapacitet for P optagelse pr. hyfelængdeenhed. Det fremgår også, at carbendazim ikke alene påvirker mykorrhizasvampes funktion via en effekt på svampenes vækst, men også via en effekt på myceliets kapacitet til at optage og transportere P til planten. Carbendazim havde således en ringe effekt på hyfelængden af K219 og BEG87, men hyfernes P optagelse var stærkt hæmmet. Manzoceb forøgede kapaciteten for P optagelse hos H-07 og BEG87, og disse svampe var samtidig mest tolerante over for carbendazim behandlingen.
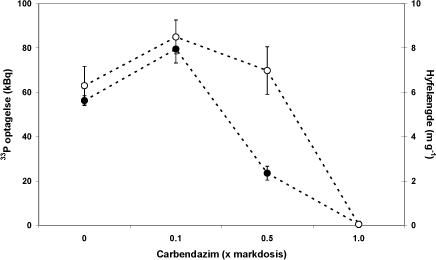
Figur 4.7. Dosis-respons for carbendazim effekter på hyfelængde (åbne cirkler) og 33P optagelse (fyldte cirkler) hos G. intraradices i symbiose med M truncatula.
Den negative kontrolbehandling (markdosis af carbendazim) medførte som forventet en >95 % hæmning i 33P optagelsen hos G. intraradices, hvilket bekræfter at modelsystemet fungerede optimalt (Fig. 4.7). Dosis-responskurver for carbendazimdoserne 0; 0,1; 0,5 og 1 x markdosis viser at den laveste dosis havde en stimulerende og hormesis-lignende effekt på både hyfelængden og 33P optagelsen hos G. intraradices, på samme måde som påvist for tre andre Glomus svampe i afsnit 5.1
Disse resultater fører til konklusionen, at tilførsel af 0,5 x markdosis mancozeb har en begrænset indflydelse på mykorrhizasvampes vækst og funktion i planters P optagelse. Derimod har 0,5 x markdosis carbendazim markante skadevirkninger, som dog afviger væsentligt imellem svampe, også imellem svampe af samme genotype. Det er bemærkelsesværdigt at en ensartet toksisk effekt på P optagelsen hos to svampe af samme genotype kunne være forårsaget af forskellige effekter af fungicidet: i det ene tilfælde blev hyfelængden reduceret kraftigt (K230) medens især P optagelseskapaciteten pr. hyfelængdeenhed blev påvirket hos den anden svamp (K219). Resultaterne har bekræftet den testede hypotese, at mykorrhizasvampe responderer forskelligt på fungicider, ikke blot imellem arter, men også blandt isolater eller genotyper inden for den samme art. Der konkluderes, at resultater opnået med en enkelt svamp ikke nødvendigvis gælder for andre svampe, heller ikke når der er tale om svampe inden for én enkelt art.
Version 1.0 September 2006, © Miljøstyrelsen.