Omfang og effekt af herbicidafdrift til læhegn
6 Scenarieberegninger: Effekter af afdriftsreducerende tiltag
- 6.1 Metode
- 6.2 Scenariebetingelser
- 6.3 Meteorologi: Timing af sprøjtning
- 6.4 Sprøjtefrie randzoner: Betydning af bredde
- 6.5 Betydning af antal sprøjtespor
- 6.6 Dyser: Typer og sprøjtetryk
- 6.7 Effekten af reduceret dosis
- 6.8 Betydning for tilgængelig fugleføde
6.1 Metode
Beregningerne af konsekvenserne af forskellige tiltag, der skal reducere afdriften af herbicider til danske læhegn (bufferzoner, alternative dysetyper og optimalt valg af sprøjtevejr) baserer sig som beskrevet i kapitel 1 på en kombination af modellens beregninger af afdriften under forskellige omstændigheder (kapitel 3), estimaterne for forekomsten af forskellige bærbærende hegnsarter (kapitel 4), data for effekten af en given metsulfuronafsætning på tjørn, hyld og seljerøn (kapitel 5; Kjær et al. 2004, Kjær et al. 2006) og data for den højdemæssige fordeling af bærproduktionen (kapitel 5 samt Kjær et al 2004). Kombinationen af disse data er brugt til at give et skøn over effekten af de forskellige afdriftreducerende tiltag på produktionen af bær i danske læhegn. Derudover har vi ud fra data om stand- og trækfugles konsumption af bær i læhegn (Snow & Snow 1988, Herrera 1998) samt bærrenes næringsværdi (Eriksson & Ehrlén 1991, Snow & Snow 1988) skønnet betydningen af reduceret herbicidafdrift for overvintrende drosselfugle i forhold til deres fødebehov.
I scenarieberegningerne indgår også omregning mellem tørvægt og friskvægt for tjørn på basis af eksisterende data (Kjær et al. 2004 og 2006, Snow & Snow 1988, Eriksson & Ehrlén 1991). Da output fra modellen er relateret til forventet afsætning på curlere, er der desuden anvendt en omregningsfaktor fra curlere til blade (se Figur 2.4).
6.2 Scenariebetingelser
Som Figur 3.16 viser, bidrager sprøjtesporene 2-10 væsentligt til den samlede herbicidafdrift til hegn, og da den gennemsnitlige markstørrelse i Danmark er ca. 16 ha, vil der typisk være i hvert fald 10 sprøjtespor af 12 m bredde i hver mark. Den relative betydning af bidraget fra spor 2-10 stiger med afstanden mellem sprøjte og hegn og afhænger også i nogen grad af sprøjtetidspunktet. De følgende scenarieberegninger har vi valgt primært at basere på afdriften fra de 10 sprøjtespor nærmest hegnet. Tilsvarende har vi anvendt estimaterne for sprøjtning kl. 11 i de scenarier, hvor sprøjtetidspunktet ikke varierer, da afdrift ved sprøjtning kl. 11 repræsenterer en mellemting mellem afdriften fra sprøjtning kl. 6 og kl. 16. Vi har ydermere antaget, at der på bladene og curlerne ligesom på papir kun afsættes dråber med en diameter større end 20 µm, og derfor kun regnet på variationen i afsætning af disse dråber.
6.3 Meteorologi: Timing af sprøjtning
Figur 3.15 viser, at det har stor betydning for afdriften af herbicid til hegn, hvornår der sprøjtes. I overensstemmelse med gældende anbefalinger giver sprøjtning tidligt på dagen mindst afdrift til læhegnene. Denne variation i afdriften afspejles også i de beregnede effekter på bærsætningen (Figur 6.1). Forskellen i effekt afhænger dog af arten, idet sprøjtetidspunktet har større betydning for tjørn og hyld end for seljerøn. Dette hænger sammen med højden af de forskellige arter samt fordelingen af bærproduktionen over højden (se Figur 5.5).
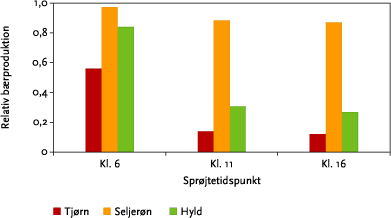
Figur 6.1. Bærproduktion ved sprøjtning på forskellige tidspunkter i forhold til bærproduktionen uden sprøjtning. For hver art er der summeret over bærproduktionen i hele hegnets højde. Værdierne er beregnet ud fra den modellerede afdrift af dråber > 20 µm fra 10 sprøjtespor for dysetypen XR11002 ved 3,1 atm og uden sprøjtefri bufferzone.
6.4 Sprøjtefrie randzoner: Betydning af bredde
Som vist i Figur 3.21 falder afdriften til den nederste del af hegnet med stigende bredde af den sprøjtefrie bufferzone, mens reduktionen i afdriften falder med højden, så der stort set ingen effekter er i den øverste del af hegnet. Den beregnede betydning af bredden af en sprøjtefri randzone på bærsætningen i tjørn, seljerøn og hyld fremgår af Figur 6.2. Bredden af bufferzonen har kun ubetydelig effekt på bærsætningen i tjørn og seljerøn, mens der for hyld er en stigende bærproduktion med stigende bufferzonebredde. Forskellen mellem arterne skyldes den højdemæssige fordeling af bærsætningen og forskelle i følsomhed over for sprøjtemidlet.
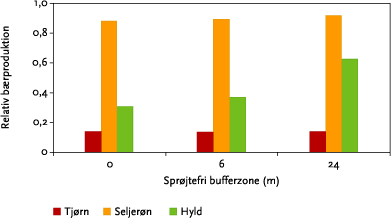
Figur 6.2. Estimeret bærproduktion som funktion af bredden af sprøjtefri randzone i forhold til bærproduktionen uden sprøjtning. For hver art er der summeret over bærproduktionen i hele hegnets højde for 10 sprøjtespor. Alle tal er for dysetypen XR11002 ved 3,1 atm og sprøjtetidspunkt kl. 11.
Til supplement er i Figur 6.3 vist interaktionen mellem bredden af en sprøjtefri randzone og sprøjtetidspunkt for bærsætningen i hyld, hvoraf ses, at randzonens bredde har størst betydning, når der sprøjtes på tidspunkter med forholdsvis høj vindhastighed og lav luftfugtighed (kl. 11 og 16 i forhold til kl. 6). For tjørn og seljerøn er der ingen forskel i betydningen af bufferzonen for de forskellige sprøjtetidspunkter.
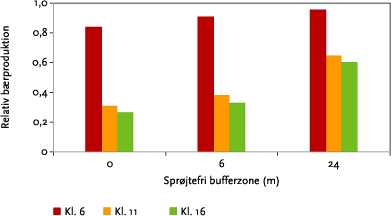
Figur 6.3. Betydning af bufferzonebredde ved forskellige sprøjtetidspunkter for bærsætningen i hyld i forhold til bærsætningen uden sprøjtning. Data er for dysetypen XR11002 ved 3,1 atm summeret over 10 sprøjtespor.
6.5 Betydning af antal sprøjtespor
Modelberegningerne viste, at det har stor betydning for estimeringen af afdriften, hvor mange sprøjtespor der medregnes (Figur 3.20), og at betydningen øges med højden i læhegnet.
Figur 6.4 viser, at det også for effektvurderinger af nogle arter er væsentligt at medregne flere sprøjtespor.
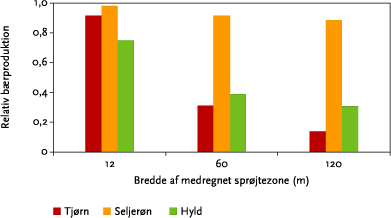
Figur 6.4. Estimeret bærproduktion som funktion af antal medregenede sprøjtespor i forhold til bærproduktionen uden sprøjtning. For hver art er der summeret over bærproduktionen i hele hegnets højde. Alle tal er for dysetypen XR11002 ved 3,1 atm og sprøjtetidspunkt kl. 11 uden sprøtjefri bufferzone.
6.6 Dyser: Typer og sprøjtetryk
Som vist i Figur 3.15, betyder dysetype og –tryk meget for afdrift af sprøjtemiddel til hegnet. Effekten af at anvende alternative dysetyper eller –tryk er illustreret i Figur 6.5. Anvendelse af dysetypen AI 11004 giver en betydelig større bærproduktion i tjørn og hyld end standarddysen XR 11002 ved 3,1 atm dysetryk, mens anvendelse af et lavere dysetryk i XR-dysen giver en lidt mindre stigning i bærproduktionen i tjørn og hyld. For seljerøns vedkommende har dysetype og -tryk ikke den store betydning for bærsætningen. For hyld aftager effekten af dysevalget med stigende bufferzonebredde, mens der for tjørn og seljerøn ikke er nogen interaktion mellem effekt af bufferzonebredde og valget af dysetype/-tryk (ikke illustreret).
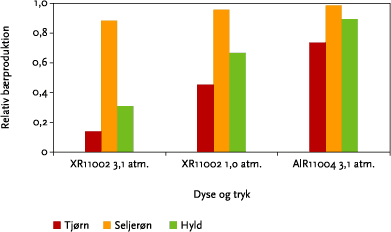
Figur 6.5. Bærproduktion ved sprøjtning med forskellige dysetyper og -tryk som funktion af bredden af sprøjtefri randzone i forhold til bærproduktionen uden sprøjtning. For hver art er der summeret over bærproduktionen i hele hegnets højde. Alle datapunkter er beregnet ud fra den modellerede afdrift fra 10 sprøjtespor ved sprøjtning kl. 11.
6.7 Effekten af reduceret dosis
Som omtalt i kapitel 3 vil afdriften til hegnet falde proportionalt med en reduktion i den anvendte herbiciddosering. Idet fx en halvering af doseringen vil resultere i en halveret afsætning i hele hegnets højde, vil effekten heraf på en given træarts bærsætning primært afhænge af artens følsomhed over for sprøjtemidlet og ikke af den vertikale fordeling af bærrene. Som følge deraf vil en reduceret metsulfurondosering have større effekt på tjørn og hyld end på den mindre følsomme seljerøn (Figur 6.6).
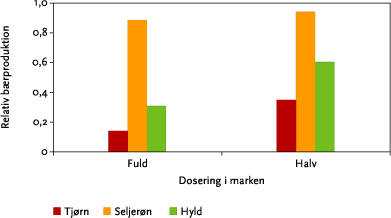
Figur 6.6. Bærproduktion ved sprøjtning med fuld dosis ( 4 g/ha) og halv dosis i forhold til bærproduktionen uden sprøjtning. For hver art er der summeret over bærproduktionen i hele hegnets højde. Alle datapunkter er beregnet ud fra den modellerede afdrift fra 10 sprøjtespor ved sprøjtning kl. 11 uden sprøjtefri bufferzone.
6.8 Betydning for tilgængelig fugleføde
Det er kendt, at bær fra tjørn, seljerøn og hyld er vigtige fødeemner for drosselfugle på træk og for standfugle gennem vinteren. Formålet med dette afsnit er at evaluere, om de estimerede effekter af herbicidafdrift på bærsætningen i hegn kan være biologisk betydende for bærspisende fugle, og i givet fald om fuglene vil påvirkes nævneværdigt ved anvendelse af afdriftreducerende tiltag. Dette gør vi ved hjælp af viden og antagelser om danske tjørne-, hylde- og seljerønhegns produktion af bær, bærrenes betydning for fugles vinterfouragering, og herbiciders påvirkning af frugtsætning hos bærbærende buske og træer. Afsnittet er opbygget, således at det først gennemgår de forskellige delforudsætninger, der indgår i ovennævnte vurdering. Dette gøres dels argumenterende, dels ved hjælp af den viden der findes rapporteret fra området. I tilfælde, hvor den eksisterende viden ikke er afklaret, har vi måttet træffe et argumenteret valg mellem de foreliggende resultater. Argumentet kan være, at undersøgelsen er lavet under betingelser, der er sammenlignelige med de danske forhold, eller en vurdering af resultaternes pålidelighed. Da beregningerne i dette afsnit således vil basere sig på en række mere eller mindre realistiske antagelser, må de ikke betragtes som præcise svar, men som overslag over mulige ændringer i de bærspisende fugles tilgængelige fødemængde i områder, hvor de primært finder deres føde i hegnene.
6.8.1 Bedømmelse af produktionen af bær i læhegn i Danmark
Vi har i kapitel 4 beregnet, at der er 17.339 km tjørnehegn, 13.518 km seljerønhegn og 11.235 km hyldehegn i Danmark, og ud fra kapitel 4 samt Kjær et al. (2002) har vi beregnet bærproduktionen for hyld, seljerøn og tjørn pr. km hegn til hhv. 1103, 1172 og 252 kg friskvægt. Det betyder, at der er en produktion på 15.843, 4.389 og 12.392 tons for hhv. seljerøn, tjørn og hyld i danske læhegn.
6.8.2 Hvilke fugle spiser bærrene i hegnene?
Det er primært drosselfugle, som spiser tjørnebær og seljerønbær (Hartley 1954; Snow og Snow 1988), men de spises også af andre fuglearter (Tabel 6.1). Hyld spises også af drosselfugle, men specielt stærene fortærer mange hyldebær.
Tabel 6.1 fugle der, på basis af engelske observation over 5 år, hyppigst spiser tjørnefrugter, rønnebær og hyld.
| Fugleart | Tjørn | Seljerøn | Hyld | Reference |
| Vindrossel Turdus iliacus |
× | × | Hartley (1954), Sorensen (1981), Guitian (1992) | |
| Solsort Turdus merula |
× | × | × | (Hartley, (1954), Sorensen (1981), Fuentes (1994) |
| Sjagger Turdus pilaris |
× | × | Hartley (1954), Sorensen (1981), Guitian (1992) | |
| Misteldrossel Turdus viscivorous |
× | × | Hartley (1954) | |
| Sangdrossel Turdus hilomelos |
× | × | × | Snow og Snow (1988) |
| Ringdue Columba palumbus |
× | × | × | Sorensen (1981), Snow og Snow (1988) |
| Stær Sturnus vulgaris |
× | × | × | Snow og Snow (1988) |
| Rødhals Erithacus rubecula |
× | × | × | Snow og Snow (1988) |
| Silkehale Bombycilla garrulus |
× | Snow og Snow (1988) | ||
| Blåmejse Parus caeruleus |
× | × | × | Snow og Snow (1988) |
| Kernebider Coccothraustes coccothraustes |
× | Snow og Snow (1988) |
Collinge (1941) angiver, at vild frugt udgør fra 26,5 til 30,5 % af føden hos solsort set over hele året, og det er derfor klart, at den vilde frugt må have stor betydning i efterår og vinter. Da solsort samtidig regnes som specialist i tjørnefrugter (Snow og Snow 1988), er det sandsynligt, at størstedelen af dens fødeindtag i perioden fra oktober til januar er tjørnebær.
Antagelse: Det er hovedsagelig drosselfugle, der spiser tjørnebær og seljerøn. Blandt drosselfuglene er solsorten den eneste, som ikke trækker væk fra Danmark. Vi antager, at den mængde bær, der er tilbage, når de øvrige drosler trækker væk, er af betydning for solsortens vinteroverlevelse.
6.8.3 Antallet af fugle der spiser bær i hegn
Det er hovedsageligt drosselfugle, som fouragerer i hegn. Derfor omfatter den følgende estimering af antallet af fugle, der søger deres føde i hegn, drosselfuglene og stære, der fortærer mange hyldebær.
Drosselfuglene i Danmark kan deles i to hovedgrupper, standfugle og trækgæster. Solsorten er en standfugl, medens de øvrige drosler altovervejende kommer igennem Danmark på deres træk fra Skandinavien til deres overvintringsområder. I følge Dansk Ornitologisk Forening er der ca. 2,25 mio. ynglepar af solsorte i Danmark. Med de unger, de får, og den dødelighed, der finder sted, er der formentlig mindst fem millioner solsorte i Danmark om vinteren. Det er dog langt fra alle solsortene, der lever i tjørnehegn. I dagens Danmark er solsorten i høj grad blevet til en havefugl, og det er kun en mindre del af populationen, som findes i mere naturlige habitater som hegn, krat og skov. Vi anslår, at halvdelen af solsortene energibehov i vinterperioden dækkes af bær fra hegn.
Antallet af trækfugle er det endnu sværere at anslå; men der findes formentlig mindst 20 millioner andre drosler end solsorte i Sverige, Norge og Finland tilsammen (Asbirk 1997) (Tabel 6.2). Mange af disse passerer Danmark på deres trækrute. Derfor er det sandsynligt, at ca. 10 millioner trækkende drosler passerer Danmark. Det vides ikke, hvor længe de opholder sig her. Der er vel også tale om stor variation mellem år. På baggrund af antallet af potentielle trækfugle antager vi, at der i gennemsnit er en halv million trækdrosler til stede hver dag fra midten af oktober til midten af november, hvorefter antallet formodes at falde til nær nul omkring udgangen af november.
Tabel 6.2. Ynglende drosler og stære i Skandinavien (antal millioner par) efter (Asbirk et al., 1997).
| Sverige | Finland | Norge | Danmark | |
| Sjagger | 0,75 – 1,5 | 0,8 – 1,2 | 1,0 – 3,0 | |
| Sangdrossel | 1,5 – 3,0 | 1,0 – 1,5 | 0,5 – 1,0 | |
| Vindrossel | 1,0 – 2,0 | 1,5 – 3,0 | 1,0 – 1,5 | |
| Misteldrossel | 0,075 – 0,2 | 0,05 – 0,08 | 0,01 – 0,05 | |
| Solsort | 1,0 – 2,0 | 0,15 – 0,2 | 0,1 – 1,0 | 2,25 |
| Stær | 0,66 |
Antagelse: Der passerer 10 millioner drosselfugle gennem Danmark i løbet af efteråret. Dertil kommer at der findes ca. 5 millioner danske solsorte som normalt bliver i lande og 660.000 stære som forlader landet i løbet af september og oktober.
6.8.4 Drosselfugles energibehov om vinteren
En fugl på ca. 100 g skal ved 0 °C bruge ca. 50 kcal/døgn for at opretholde sin kropsvægt ved et lavt aktivitetsniveau (Kendeigh, 1970). Tilsvarende skal en fugl på 130 g bruge 55 kcal/døgn. Drosselfugle vejer typisk omkring 100 g i den del af sæsonen, hvor der er modne tjørnebær (Tabel 6.3).
Tabel 6.3. Nogle bærspisende fugles vægt (efter Snow & Snow 1988).
| Art | Vægt (g) | Sæson |
| Solsort | 113,9 | November – januar |
| Sangdrossel | 95,9 | November – februar |
| Misteldrossel | 144,0 | November – februar |
| Sjagger | 117,9 | September – januar |
| Vindrossel | 64,9 | September – november |
| Stær | 92,1 | November – februar |
Antagelse: Som gennemsnit skal drosselfugle bruge 50 kcal/døgn for at opretholde livet ved en lav aktivitet ved danske efterårstemperaturer.
Når man skal vurdere betydningen af, at bærbærende hegnsarters frugtsætning ændres, er det vigtigt at kende bærrenes værdi som fødekilde for fugle. Til belysning af dette foreligger der materiale først og fremmest fra England. Vi har ikke fundet danske undersøgelser, som kan supplere dette.
Ud fra kendskab til energiudnyttelse af føden og energiindhold i bærrene kan følgende fødebehov i form af bær beregnes (Tabel 6.4).
Det gennemsnitlige energibehov per døgn for en drossel udtrykt i gram tjørnebær er således 100 g tjørnebær, 74 g seljerønbær eller 179 g hyldebær. Denne mængde benævnes et drosseldøgn.
Antagelser: 100 g tjørnebær eller 74 g seljerønbær eller 179 g hyldebær kan forsyne en gennemsnitsdrosselfugl med den nødvendige energi per døgn. Denne mængde kaldes herefter drosseldøgnækvivalent eller blot drosseldøgn.
Tabel 6.4. Bærspisende fugles energi- og fødebehov per døgn under forudsætning af at energien skaffes fra tjørnebær, seljerønbær eller hyldebær (baseret på opgivelser hos (Snow og Snow, 1988; Herrera, 1998 og Eriksson og Erhlen, 1991)).
| Art | Energi-behov | Frisk frugt | Energi udbytte |
Behov | Bær til rådighed | Drossel-døgn |
| Kcal/døgn | Kcal/g | % | g fr. frugt | ton | millioner | |
| Tjørn | 50 | 0,68 | 75 | 100 | 4.389 | 43 |
| Seljerøn | 50 | 0,925 (0,78-1,07) | 75 | 74 | 15.843 | 214 |
| Hyld | 50 | 0,38 | 75 | 179 | 12.392 | 69 |
6.8.5 Sammenligning af beregnet fødebehov og fødeproduktion
I beregningerne antages det, at hver mark i gennemsnit sprøjtes en gang med herbicid, samt at halvdelen af hegnene herved eksponeres i en eller anden grad. Det antages ligeledes, at alle herbicider har virkninger svarende til metsulfuron, eller at alle marker sprøjtes med metsulfuron. Det er selvfølgelig en simplificering, og de følgende beregninger kan derfor kun betragtes som overslag.
Fødemængde for fuglene beregnes ved at omsætte bærmægden til drosseldøgn for den enkelte hegnsart (Tabel 6.4). Reduktionen i fødemængden som følge af metsulfuron-afdrift beregnes ved at reducere i henhold til de effekter, der er beregnet tidligere i kapitel 6. I de præsenterede data er beregnet den bærmængde der vil være til rådighed ved den beregnede afdrift af sprøjtemiddel efter brug af 3 dysetyper, 2 dysetryk og med/uden bufferzone på 12 m (se Tabel 6.5 nedenfor). Alle beregninger er gennemført for en sprøjtning, der er gennemført klokken 11 med en metsulfuron-dosering på 4 g/ha.
Tabel 6.5: Oversigt over den forventede procentvise reduktion i tilgængelig fugleføde som følge af sprøjtemiddelafdrift ved tre dysetyper/-tryk og tilstedeværelse af 12 m bufferzone(jf. afsnit 6.4 og 6.5) sammenlignet med usprøjtede hegn.
| Dyse | AI11004 | AI11004 | XR11002 | XR11002 | XR11002 | XR11002 | |
| Dysetryk | 3,1 atm | 3,1 atm | 1 atm | 1 atm | 3,1 atm | 3,1 atm | |
| Hegnsart | Buffer-zone | 12 m | 0 m | 12 m | 0 m | 12 m | 0 m |
| Hyld | 0 | 4,7 | 11 | 16 | 34 | 56 | 69 |
| Seljerøn | 0 | 1,2 | 1,6 | 4,5 | 4,6 | 10 | 12 |
| Tjørn | 0 | 25 | 27,0 | 54 | 55 | 86 | 87 |
Dernæst anslås mængden af fugle, der fortærer bær, og denne mængde omsættes til behovet i drosseldøgn. Som tidligere nævnt er det vores antagelse, at 10.000.000 trækfugle kommer gennem Danmark over en måned. Det er antaget, at der er 500.000 drosler tilstede per dag i den periode. Derudover er der 5.000.000 solsorte, der få dækket 50 % af deres fødebehov ved at spise bær: Det betyder, at der er det, der svarer til 2.500.000 solsorte, fra september til november. I december udgør bær kun 25 % af solsortenes føde (1.250.000 solsorte), og i januar og februar falder andelen yderligere til 10 % (500.000 solsorte) (Snow og Snow 1988).
Beregningerne viser, som det fremgår af Tabel 6.4 og nedenstående Figur 6.7, at der er 326 millioner drosseldøgn til rådighed, ifald hegnene ikke påvirkes af herbicidafdrift. Samtidig har fuglene et fødebehov, der svarer til 326,5 millioner drosseldøgn.
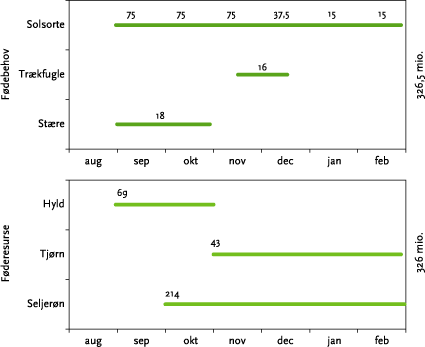
Figur 6.7. Oversigt over de antagelser der er gjort i beregningen vedrørende forekomsten af bær og bærspisende drosler og stære. Tallene præsenteret på den enkelte linje angiver drosseldøgn. Tallene ved de tre arter af bærbærende buske/træer er den skønnede mængde af drosseldøgn, der er til rådighed ved sæsonens start, ifald alle hegn er usprøjtede.
Beregningerne for de forskellige drifthændelser indikerer, at valget af sprøjteudstyr og sprøjtetryk kan betyde, at mængden af fugleføde er mindre end behovet (Figur 6.8). Sprøjtning med AI11004 dyser giver således en meget begrænset påvirkning af den samlede fødemængde, hvorimod XR11002 brugt med et dysetryk på 3,1 atm resulterer i en reduktion på ca. 20 %. En bufferzone på fx 12 m har kun i meget begrænset grad indflydelse på fødemængden.
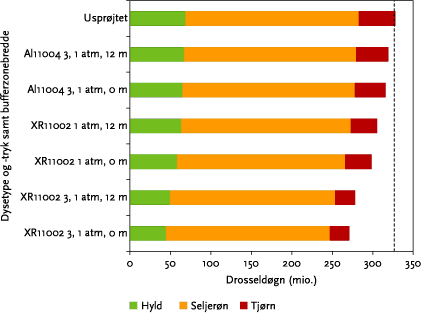
Figur 6.8. Sammenhængen mellem den tilgængelige fugleføde ved forskellige sprøjtevilkår brugen af 3 forskellige sprøjteindstillinger (dyser, dysetryk) samt +/- 12 m bufferzone. Den lodrette streg ved 308,5 millioner drosseldøgn markerer drosselfuglenes og stærenes skønnede behov for bær i drosseldøgn.
Det er i beregningerne antaget, at alle bærrene kan erstatte hinanden. Ved at sammenstille alle de tre hegnsarter ligevægtigt antager vi således, at fuglene ikke er specialiseret på en art. I det tilfælde, at der findes arter, der er specialiseret på bær af enten hyld eller tjørn, viser Figur 6.9, at de vil opleve en kraftig reduktion i fødemængden.
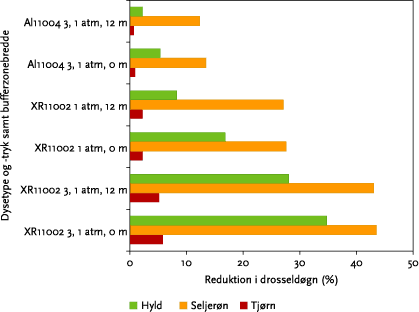
Figur 6.9. Procentvis reduktionen i tilgængelige fugleføde, udtrykt som drosseldøgn, for tre bærbærende hegnsarter ved forskellige sprøjtevilkår (3 forskellige sprøjteindstillinger (dyser, dysetryk) samt +/- 12 m bufferzone).
I det ovenstående regneeksempel er det ikke medtaget, at der også er andre fugle, der udnytter bærrene gennem efteråret og vinteren. Det betyder, at man må forvente, at disse vil opleve en fødemangel, eller at drosselfuglene hurtigere vil fjerne bærrene. Modsat er det heller ikke medtaget, at der findes flere bærbuske, som dog ikke har den samme store udbredelse (taks, kornel, slåen, kristtorn osv.).
Scenarieberegningerne tidligere i dette afsnit viste, at betydningen af sprøjtetidspunkt og betydningen af valget af sprøjtedyse var af samme størrelsesorden. Det betyder, at det er muligt at reducere effekten betydeligt ved at sprøjte tidligt om morgenen i forhold til at sprøjte i løbet af dagen (afsnit 6.3, Figur 6.1), hvilket også er den generelle anbefaling til sprøjteførere.
Ud fra de ovenstående beregninger kan man konkludere, at det er sandsynligt, at sprøjtemiddelafdrift kan have effekt på vinteroverlevelse af solsort, men at det er muligt at reducere denne effekt betydeligt ved at vælge afdriftreducerende dysetyper, det rette sprøjte tidspunkt og i et vist omfang etablering af bufferzoner.
Version 1.0 September 2008, © Miljøstyrelsen.