Sprøjtepraksis i sædskifter med og uden glyphosattolerante afgrøder
3 Resultater
3.1 Herbicideffekter på markens biodiversitet
3.1.1 Ukrudtsbiomasse og frøproduktion
Både sort natskygge og hanespore udviste det typiske, S-formede dosis-respons på biomasse efter behandling med både glyphosat og MaisTer. Generelt var de anvendte doseringers effekt på frøproduktion lavere end på biomasse, men også denne parameter kunne beskrives ved hjælp af S-formede doseringskurver. Udfra doseringskurverne var det muligt at beregne hvilken dosering, der var nødvendig for at opnå en given effekt.
I relation til ukrudtsbekæmpelse er 90% effekt et ønsket mål og tabel 3.1 viser de beregnede ED90-doseringer af glyphosat og MaisTer på henholdsvis sort natskygge og hanespore med biomasse og frøproduktion som måleparameter. For begge ukrudtsarter krævede en sen sprøjtning (større udviklingstrin) for begge herbicider en højere dosering for at opnå 90% reduktion af biomassen. Doseringsbehovet øgedes mere for MaisTer end for glyphosat, hvilket sandsynligvis kan tilskrives, at glyphosat er mere systemisk end MaisTer.
Tabel 3.1. Reduktion af overjordisk plantebiomasse og frøsætning som respons på herbiciddosering. ED90 er den beregnede dosis for 90% reduktion. 95%-konfidensintervallet er vist i parentes. aEkstrapolerede doseringer. bMaksimalt anvendte doseringer (ekstrapolerede værdier var langt større).
| Ukrudtsart og herbicid | Udviklingstrin | Biomasse ED90 |
Frøsætning ED90 |
|||||
| Sort natskygge Glyphosat |
4 blade | 79,2 | (72,0...101) | 180a | ||||
| (g as./ha) | 8-10 blade | 263 | (169...360) | 443a | ||||
| MaisTer | 4 blade | 0,601 | (0,499...0,701) | >1,00b | ||||
| (g as./ha) | 8-10 blade | 4,10 | (2,40...5,80) | >2,00b | ||||
| Hanespore Glyphosat |
4 blade | 130 | (119...137) | 245 | (180...305) | |||
| (g as./ha) | 6-8 blade | 248 | (173...324) | 115 | (7,20...180) | |||
| MaisTer | 4 blade | 0,800 | (0,499...1,20) | 2,10 | (1,60...2,70) | |||
| (g as./ha) | 6-8 blade | 1,60 | (0,400...3.69) | 2,79 | (2,29...3,29) | |||
I forsøgene blev biomassen målt 3-4 uger efter sprøjtning og afspejler således korttidseffekten af behandlingerne, mens frøproduktionen blev målt flere måneder efter behandling og således afspejler langtidseffekten. Generelt skulle der højere doseringer til at reducere frøproduktionen end til at reducere biomassen (tabel 3.1). Det betyder, at plantens reproduktive evne er mindre følsom over for herbiciderne end dens vegetative vækst. Dvs. at selvom man opnår en vækstreduktion på kort sigt, så er planterne ofte i stand til på langt sigt at komme i vækst igen og producere frø.
Forskellen på ED90-doseringen for biomasse og frøproduktion var større, jo tidligere sprøjtningen blev udført, og var desuden generelt større med MaisTer end med glyphosat.
Det skal bemærkes, at forsøgene er udført som potteforsøg med planter i renbestand. Betydningen af konkurrence med afgrøden om lys, vand og næring er således ikke inddraget. Vi formoder at denne konkurrence vil forøge effekten af sprøjtningen, men at de relative forskelle fremhævet ovenfor vil gælde også i marken.
3.1.2 Glyphosatsprøjtning i majs
Forsøgene med forskellige sprøjtestrategier i majs faldt mere tydeligt ud i 2006 end i 2005 (figur 3.1-3.2). Når der ses på udbyttet, kan det konkluderes, at splitsprøjtning med glyphosat fungerede lige så godt som splitsprøjtning med MaisTer. Med blot én glyphosatsprøjtning var høstresultatet lige så godt, men kun med en tidlig sprøjtning (20. eller 29. juni), ikke med en sen (11. eller 12. juli).
Som ventet var ukrudtets tørvægt faldet ved prøvetagningen, som fulgte efter sprøjtningen. Den eneste undtagelse (agersennep ved tidlig glyphosatsprøjtning i 2006) skyldtes et enkelt afvigende plot – måske med blot en enkelt overlevende stor plante; dette fremgår af den store usikkerhed på gennemsnittet vist i figur 3.2.
3.1.3 Ukrudtets vækstforhold i majs
Den naturlige forekomst af hvidmelet gåsefod og lugtløs kamille i marken gjorde det svært at opnå de lave tæthedsniveauer i 2005 (tabel 3.2), men i 2006 lykkedes det (tabel 3.3). Som ventet for en afgrøde som majs, der i lang tid står åben, begrænsede den kun ukrudtets vækst lidt. Samtidigt led den selv betydeligt i konkurrencen (figur 3.3-3.4).
Tabel 3.2. Fremspiring af ukrudt i majsforsøg 2005 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
71,0 | ±31,22 | 49,5 | ±27,29 | 65,0 | ±82,31 | 99,5 | ±16,76 | ||||
| Med afgrøde Lav tæthed |
14,5 | ±14,18 | 6,00 | ±5,889 | 64,0 | ±88,18 | 34,0 | ±11,78 | ||||
| Mellem tæthed | 32,5 | ±19,49 | 24,5 | ±3,416 | 70,0 | ±106,9 | 34,0 | ±4,320 | ||||
| Høj tæthed | 65,5 | ±22,65 | 60,0 | ±16,73 | 36,0 | ±50,20 | 74,0 | ±36,11 | ||||
Tabel 3.3. Fremspiring af ukrudt i majsforsøg 2006 (gns.±s.d. kimplanter pr.
m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
31,5 | ±11,00 | 34,5 | ±19,69 | 28,5 | ±6,61 | 30,0 | ±12,96 | ||||
| Med afgrøde Lav tæthed |
10,5 | ±7,90 | 10,5 | ±3,79 | 13,5 | ±9,15 | 5,5 | ±1,00 | ||||
| Mellem tæthed | 8,5 | ±6,81 | 17,0 | ±6,63 | 22,5 | ±9,57 | 18,0 | ±7,12 | ||||
| Høj tæthed | 30,0 | ±7,48 | 32,5 | ±15,18 | 37,5 | ±11,00 | 24,5 | ±5,74 | ||||
3.1.4 Ukrudtets vækstforhold i vårbyg
Det lykkedes ikke helt at styre fremspiringstætheden af hvidmelet gåsefod, som i 2005 forsvandt helt fra to af tæthedsniveauerne (tabel 3.4), og af lugtløs kamille, som forsvandt i ét af niveauerne i 2006 (tabel 3.5).
Vårbyg var i alle tilfælde i stand til at trykke ukrudtet til betydeligt lavere tæthed, end når ukrudtet voksede i renbestand (figur 3.5-3.6). Det kan undre, at vårbyggen selv led så betydeligt i konkurrencen (figur 3.5-3.6 øverst), når man betænker, at ukrudtet aldrig (bortset fra burresnerre i 2005) blev særlig stort. Udbyttetabet er sandsynligvis et artefakt, idet lugearbejdet for at opretholde de ønskede ukrudtsbestande var ganske omfattende, og dette kunne ikke undgå at skade afgrøden.
Tabel 3.4. Fremspiring af ukrudt i vårbygforsøg 2005 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
658 | ±234,6 | 65,0 | ±94,34 | 67,5 | ±18,93 | 128 | ±40,31 | ||||
| Med afgrøde Lav tæthed |
87,5 | ±25,00 | 12,5 | ±9,574 | 0,0 | ±0,00 | 57,5 | ±37,75 | ||||
| Mellem tæthed | 123 | ±43,49 | 57,5 | ±22,17 | 0,0 | ±0,00 | 90,0 | ±24,49 | ||||
| Høj tæthed | 573 | ±131,0 | 205 | ±93,99 | 77,5 | ±17,08 | 220 | ±57,74 | ||||
Tabel 3.5. Fremspiring af ukrudt i vårbygforsøg 2006 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Hvidmelet gåsefod |
Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
65,0 | ±19,15 | 30,0 | ±42,43 | 60,0 | ±28,28 | 65,0 | ±25,17 | ||||
| Med afgrøde Lav tæthed |
12,5 | ±5,00 | 22,5 | ±20,62 | 42,5 | ±37,75 | 0,0 | ±0,00 | ||||
| Mellem tæthed | 45,0 | ±30,00 | 57,5 | ±9,57 | 12,5 | ±12,58 | 20,0 | ±33,67 | ||||
| Høj tæthed | 72,5 | ±53,15 | 62,5 | ±12,58 | 37,5 | ±25,00 | 35,0 | ±44,35 | ||||
3.1.5 Ukrudtets vækstforhold i vinterraps
Det var svært at styre fremspiringstætheden af ukrudtet især i efteråret 2005, hvor 5 ud af de 12 kombinationer mellem ukrudt og vinterraps endte uden ukrudt (tabel 3.6). For kornvalmue kom i alle tilfælde kun ganske få planter op (tabel 3.6-3.7).
Fremspiringen var stort set tilendebragt ved den første optælling af kimplanter i november i begge forsøg. Planterne fik imidlertid i de fleste tilfælde først rigtig gang i deres vækst i juni (figur 3.7-3.8). I 2006-2007 havde burresnerre og især fuglegræs dog opnået en betydelig størrelse allerede i november. De aftog i størrelse henover vinteren, hvorefter burresnerre atter tog fat fra juni af, mens fuglegræs helt uddøde. Denne uddøen må have skyldtes afgrødens konkurrence, da fuglegræs voksende alene genoptog sin vækst i juni akkurat som burresnerre (figur 3.8).
Tabel 3.6. Fremspiring af ukrudt i vinterrapsforsøg 2005-2006 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Kornvalmue | Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
194 | ±78,55 | 133 | ±115,7 | 35,0 | ±32,23 | 17,0 | ±21,26 | ||||
| Med afgrøde Lav tæthed |
35,0 | ±20,75 | 0 | ±0 | 0 | ±0 | 5,00 | ±5,030 | ||||
| Mellem tæthed | 83,0 | ±26,81 | 0 | ±0 | 0 | ±0 | 0 | ±0 | ||||
| Høj tæthed | 167 | ±54,59 | 77,0 | ±135,5 | 1,00 | ±2,000 | 16,0 | ±24,66 | ||||
Tabel 3.7. Fremspiring af ukrudt i vinterrapsforsøg 2006-2007 (gns.±s.d. kimplanter pr. m²).
| Parceltype | Burresnerre | Fuglegræs | Kornvalmue | Lugtløs kamille | ||||||||
| Uden afgrøde Høj tæthed |
229 | ±132,1 | 350 | ±100,0 | 19,0 | ±22,72 | 91,0 | ±6,83 | ||||
| Med afgrøde Lav tæthed |
21,0 | ±16,45 | 33,0 | ±31,56 | 1,00 | ±2,000 | 6,00 | ±5,16 | ||||
| Mellem tæthed | 63,0 | ±17,09 | 64,0 | ±41,70 | 0 | ±0 | 15,0 | ±12,38 | ||||
| Høj tæthed | 26,0 | ±14,79 | 30,0 | ±25,82 | 4,00 | ±8,000 | 43,0 | ±17,70 | ||||
Figur 3.1. Sprøjtescenarier i majs 2005.
I øverste række vises tørvægten (kg) ved høst af 4 m af parcellen: Kolber (venstre søjler) og stængler (højre søjler). Søjler med samme bogstav er ikke signifikant forskellige (P=0,05). Nedenunder vises overjordisk tørvægt (g pr. ½ m²) for tre ukrudtsarter, samt andet ukrudt totalt. Sprøjtetidspunkter er angivet ved stiplede linier. Data vist som gns.±s.e.
Figur 3.2. Sprøjtescenarier i majs 2006.
I øverste række vises tørvægten (kg) ved høst af 4 m af parcellen: Kolber (venstre søjler) og stængler (højre søjler). Søjler med samme bogstav er ikke signifikant forskellige (P=0,05). Nedenunder vises overjordisk tørvægt (g pr. ½ m²) for tre ukrudtsarter, samt andet ukrudt totalt. Sprøjtetidspunkter er angivet ved stiplede linier. Data vist som gns.±s.e.
Figur 3.3. Vækst af afgrøde og ukrudt i majs 2005.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,5 m²) vist som gns.±s.e.
Figur 3.4. Vækst af afgrøde og ukrudt i majs 2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,5 m²) vist som gns.±s.e.
Figur 3.5. Vækst af afgrøde og ukrudt i vårbyg 2005.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,1 m²) vist som gns.±s.e.
Figur 3.6. Vækst af afgrøde og ukrudt i vårbyg 2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,1 m²) vist som gns.±s.e.
Figur 3.7. Vækst af afgrøde og ukrudt i vinterraps 2005-2006.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,25 m²) vist som gns.±s.
Figur 3.8. Vækst af afgrøde og ukrudt i vinterraps 2006-2007.
I rækkerne vises hver plantearts vækst gennem sæsonen enten alene (diagonalt) eller i konkurrence med afgrøden. Ukrudt i høj (?·), mellem (o) eller lav tæthed (r) jf. tabel 3.4. Overjordisk tørvægt (g pr. 0,25 m²) vist som gns.±s.e.
3.1.6 Faunarespons på plantebiomasse
Væksten af afgrøde og ukrudt samt tætheden af leddyr fulgtes ad gennem sæsonen på begge de undersøgte marker i 2005 (figur 3.9; Spearmans r gav P<0,0001).
På en given dag var der imidlertid generelt ringe sammenhæng mellem biomassen af ukrudt (eller afgrøde) og insekter i de to majsmarker (figur 3.10). Det var således ikke muligt ud fra ukrudtsbiomassen at forudsige insektbiomassen en given dag.
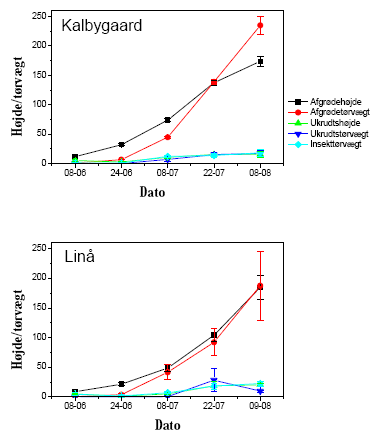
Figur 3.9. Afgrøde- og ukrudtbiomasse (g pr. 0,25 m²), afgrøde- og ukrudtshøjde (cm), samt leddyrbiomasse (mg pr. prøve à 0,81 m²). Punkter viser gns.±s.e. for 12 prøver.

Figur 3.10. Ukrudt og insekter i to majsmarker 2005.
Sammenhængen mellem biomassen af ukrudt og insekter er vist på fem datoer for begge marker (Kalbybaard og Linaa). Y-skala er den samme i alle figurer. Målingerne med nulværdier er udeladt.
3.1.7 Modellering af ukrudtets populationsdynamik
3.1.7.1 Ukrudtets frøproduktion
De supplerede data om ukrudtets frøproduktion, som blev indsamlet for at understøtte estimeringen af modellens parametre, gav robuste sammenhænge mellem biomasse og frøproduktion (figur 3.11).
![Figur 3.11. Ukrudtets frøproduktion. Tørvægt og frøproduktion af ukrudt høstet i plots á 0,1-0,5 m². Ukrudt enten uden konkurrence (o) eller i konkurrence med majs (?·) eller vårbyg (r). Lineære regressionslinier beregnet på log-transformerede data, punktet [·] ikke medtaget.](images/fig3_11.gif)
Figur 3.11. Ukrudtets frøproduktion.
Tørvægt og frøproduktion af ukrudt høstet i plots á 0,1-0,5 m². Ukrudt enten uden konkurrence (o) eller i konkurrence med majs (?·) eller vårbyg (r). Lineære regressionslinier beregnet på log-transformerede data, punktet [·] ikke medtaget.
3.1.7.2 Modelvalidering
Modellens parametre blev estimeret fra litteraturen samt figur 3.11, som beskrevet i afsnit 2.1.7. Det blev valgt ikke at tilpasse parameterværdierne, således at modellen bedst muligt beskrev resultatet af de seks valideringsforsøg (tre afgrøder gentaget over to år, se figur 3.3-3.8) – en såkaldt ”kalibrering” af modellen. Datagrundlaget i litteraturen blev anset for større og bredere, end det som kunne fremskaffes i dette projekt, og modellen baseret på litteraturen skulle således dække bredere, hvad angår forskelle i dyrkningsforhold og vejrlig.
Som testvariabel for valideringen anvendtes ukrudtsbiomassen inden høst, det vil sige på det tidspunkt, hvor den normalt topper, og hvor frøkastet sker i forhold til den opnåede biomasse. Derved testes modellens evne til at simulere resultatet af konkurrencen mellem afgrøde og ukrudt. Inden hver simulering indstilledes frøbanken til en tæthed, som gav den fremspiringstæthed for hver art, som var observeret i forsøget i afgrøden i det pågældende år (tabel 3.2-3.7). Valideringen checkede altså ikke, om modellen gav den rette fremspiring i forhold til vejr og jordbearbejdning, men kun slutresultatet af konkurrenceforløbet fra fremspiring og frem til høst.
Ser vi først på det generelle mønster, så ramte modellen godt i majs, mens der var en klar tendens til overestimering af ukrudtets vækst i vårbyg og vinterraps, især ved lave tætheder – under ca. 10 g/m², hvor modellen forudsagde en ukrudtsvægt på 10-100 g/m² (figur 3.12). Planteværn Online angiver for de fire anvendte ukrudtsarter i disse to afgrøder et effektkrav for behandling på 80-95%. Det vil sige, at man uden bekæmpelse ville forvente en betydelig vækst af ukrudtet gennem sæsonen og dermed et uacceptabelt udbyttetab, men ser man på vækstkurverne for ukrudtet i vårbyg (figur 3.5 og 3.6) og vinterraps (figur 3.7 og 3.8), fremgår det, at ukrudtet stort set blev udkonkurreret af afgrøden (forsøgene blev jo ikke sprøjtet). Det modellen forudsiger er altså, at ukrudtet i disse situationer ville opnå en vægt på 10-100 g/m², hvilket ville have været tabsgivende, men i de konkrete forsøg i Flakkebjerg de to år stod ukrudtet i stampe. Således opstår den konkrete overestimering.
Sammenligner man ukrudtets vækst i majs (figur 3.3 og 3.4) og vårbyg (figur 3.5 og 3.6), ser man, at ukrudtet både i 2005 og 2006 groede langt bedre i majs (konkurrencesvag) end i vårbyg (konkurrencestærk). Når ukrudtet trivedes dårligt i vårbyg, skyldtes det altså afgrødens vækst, som igen var styret af vejret. Man må forestille sig, at under andre vejr- og dyrkningsforhold ville ukrudtet have vokset sig stort og tabsgivende i vårbyggen. Modellen er imidlertid så simpel, at vejret ikke medregnes, og den kan derfor umuligt forudsige forskelle mellem år betinget af forskelle i vejret.
Modellen havde et særligt problem med vinterraps, hvor ukrudtets vækst afbrydes af vinteren. Overvintringen gør det mere kompliceret at simulere planters vækst korrekt. Det ses af valideringen, at særligt fuglegræs i 2006-2007 drillede (de tre punkter længst mod venstre i figur 3.12). Fuglegræs opnåede da sin maksimale vægt inden vinteren og ikke om sommeren før høst, som modellen antager. Dette er et væsentligt problem for modellen, som udbygges på dette punkt, hvis den skal anvendes til at modellere ukrudt i vintersæd i fremtiden. En yderligere komplikation opstår, når frøproduktionen skal modelleres. Det er ukendt, hvorvidt fuglegræs i 2006-2007 nåede at producere frø.
En sammenligning af de to år viste ingen forskel i modellens evne til at ramme observationerne (figur 3.12). Sammenfattende kan man sige, at modellen i disse tre afgrøder over to år havde en generel misvisning henimod en overestimering af ukrudtets vækst og dermed af dets frøsætning. Hvis denne misvisning gælder generelt, vil modellen have tendens til at overvurdere ukrudtsproblemet både inden for sæsonen og på længere sigt ved en overestimering af henholdsvis ukrudtets konkurrenceevne og populationsvækstrate. Imidlertid skal modellen her anvendes til at sammenligne forskellige dyrkningsstrategier og ikke til at forudsige absolutte ændringer i frøbanken. Til dette brug mener vi, at modellen er et pålideligt værktøj, selvom den med henblik på modellering af ukrudt i vintersæd bør udbygges.
3.1.7.3 Modellens antagelser
Konsekvensen af de fire dyrkningsscenarier (tabel 1.1 til 1.4) på frøbankdynamikken af de fem udvalgte ukrudstarter (afsnit 1.6) blev simuleret over 20 år, som vist i figur 3.13 til 3.16, begyndende med en moderat infestation på 100 frø pr. m² af hver art. Modellen har én fri parameter, frøpredation på jordoverfladen, som blev sat til 5% pr. dag. Frøpredationen er i virkeligheden meget variabel fra år til år og fra sted til sted, og den anvendte 5% pr. dag er den øvre grænse for det observerede106-108. Mens frøpredationen således blev sat temmelig højt, så blev herbicideffekten sat på den nedre grænse for det forventede. Modellen anvendte de målte effekter tre uger efter sprøjtning i væksthus, men effekten må for disse enårige arter forventes som regel at være større, da den overlevende biomasse vil være svækket og nemt vil kunne bukke under for konkurrence med afgrøden. For glyphosat anvendtes dosisresponskurver fra litteraturen7, mens Planteværn Onlines anvendtes for de øvrige herbicider.
Imidlertid er formålet med simuleringerne at forudsige forskelle i udviklingen af ukrudtsfloraen som følge af scenariernes forskellige sprøjtestrategier, snarere end at forudsige den absolutte størrelse af ukrudtspopulationerne. Ændringer af disse antagelser, frøpredationens størrelse og herbicidernes endelige effekt, vil ikke ændre på ukrudtsarternes relative position i konkurrencen, men blot flytte populationerne i samlet flok (og uændret rækkefølge) op eller ned ad y-aksen (figur 3.13-3.16).
3.1.7.4 Modellens forudsigelser
Dyrkning af GT-raps udløste ikke nogen ændring i ukrudtsfloraens sammensætning (figur 3.13). Hyppigheden af de fem ukrudtsarter efter 20 år var den samme som i sædskiftet med konventionel raps. Hvis sædskiftet havde været mindre anstrengt, og der kun havde været dyrket raps hvert fjerde år og ikke hvert tredje, så ville det blot have gjort ændringen om muligt endnu mindre.
Ved dyrkning af majs i sædskifte med vårbyg havde sprøjtestrategien indflydelse på ukrudtsfloraens udvikling (figur 3.14). Splitsprøjtning var mere effektiv med glyphosat end med MaisTer. Én sprøjtning med glyphosat den 8. juni havde mindre effekt, mens udsættelse til den 20. juni tillige forårsagede en voldsom udvikling af hvidmelet gåsefod. I praksis ville landmanden ikke acceptere en sådan udvikling men ændre sin sprøjtestrategi i majs. Det ses tydeligt, at frøbanken stiger efter år med majsdyrkning efterfulgt af et mindre fald i året med vårbyg.
Ensidig majsdyrkning viste sig at være en problematisk strategi i konventionel majs, da den fremmede lugtløs kamille som et problemukrudt (figur 3.15). Dette kan skyldes kamilles biologi. Dens frøpulje er mere persistent end de fem arters (mortalitetsrate på blot 0,1 år–1, tabel 2.7). Samtidigt har den den anden laveste fremspiringsrate (0,3 år–1), kun hvidmelet gåsefods er lavere (0,25 år–1) (tabel 2.7). Dyrkning af GT-majs med anvendelse af glyphosat som splitsprøjtning gav tilfredsstillende kontrol med ukrudtet, men med blot én sprøjtning fik hvidmelet gåsefod dog et voldsomt udbrud akkurat som i sædskiftet, hvor majs vekslede med vårbyg (figur 3.14).
I sædskifterne med roer gjorde anvendelse af glyphosat i GT-roerne ingen forskel på udviklingen af ukrudtsfloraen i forhold til den traditionelle sprøjtestrategi (figur 3.16).
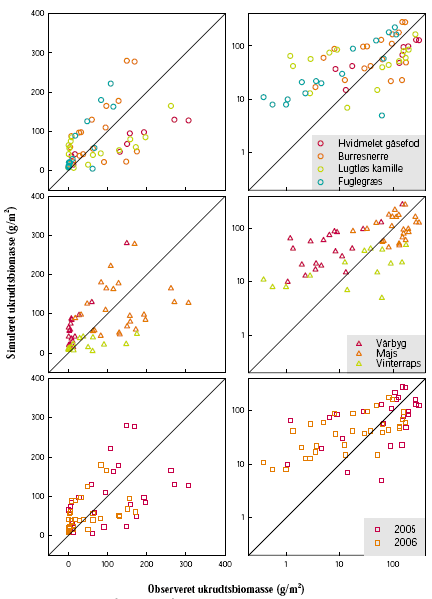
Figur 3.12. Validering af modellen for markukrudt. Ukrudtsbiomassen ved høst.
Sammenligning mellem modellens forudsigelser og biomassen observeret i markforsøgene i Flakkebjerg (figur 3.3-3.8). Punkter over/under diagonalen angiver over-/underestimering af modellen. De samme data er vist seks gange: På lineær og logaritmisk skala (venstre og højre), for ukrudtsarterne (øverst), afgrøderne (midt) og forsøgsårene (nederst).
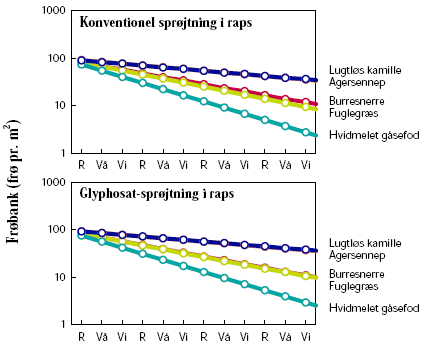
Figur 3.13. Scenarie 1: Sædskifte med konventionel vs. GT-vinterraps.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”R”=Vinteraps, ”Vå”=Vårbyg, ”Vi”=Vinterhvede.
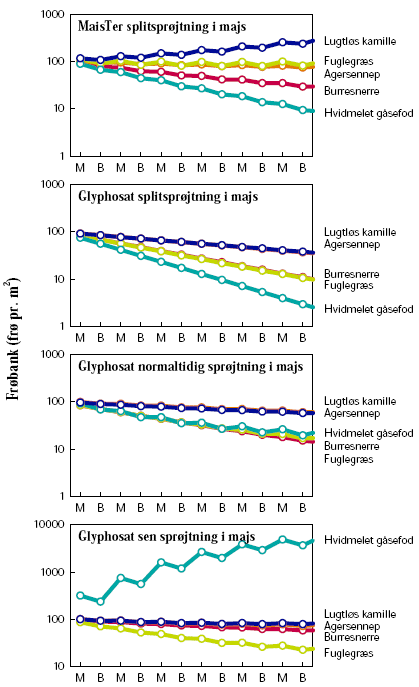
Figur 3.14. Scenarie 2: Sædskifte med konventionel vs. GT-majs.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”M”=Majs, ”B”=Vårbyg.
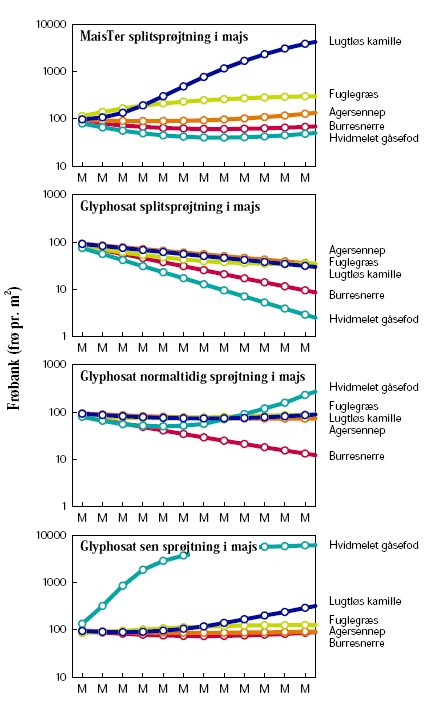
Figur 3.15. Scenarie 3: Ensidigt sædskifte, konventionel vs. GT-majs.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”M”=Majs.
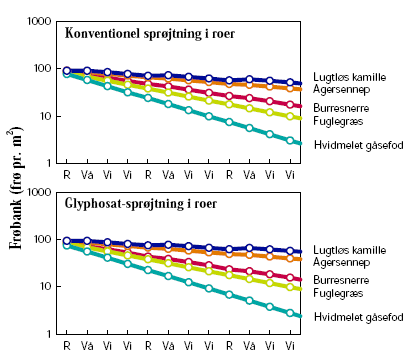
Figur 3.16. Scenarie 4: Sædskifte, med konventionelle vs. GT-roer.
Størrelsen af den simulerede frøbank er vist for hvert år den 1. januar. Tidsaksen angiver på samme tidspunkt den senest høstede afgrøde: ”R”=Roer, ”Vå”=Vårbyg, ”Vi”=Vinterhvede.
3.2 Herbicideffekter på hegnets flora
3.2.1 Ukrudtsbiomasse og frøproduktion
Effekten af glyphosat og MaisTer på biomasseproduktionen af alm. hvene, fåresvingel og mælkebøtte kunne beskrives ved hjælp af S-formede doseringskurver (tabel 3.8). Derimod var det kun muligt at beskrive frøproduktionen af mælkebøtte og alm. hvene (kun MaisTer) med S-formede kurver. På alm. hvene var effekten af glyphosat på frøproduktionen for høj, mens effekten af MaisTer var meget variabel. Fåresvingel satte slet ikke frø sandsynligvis på grund af utilstrækkelig vernalisering.
På mælkebøtte øgedes ED50-doseringen af begge herbicider med stigende udviklingstrin både for biomasse og frøproduktion. Effekten var mere udtalt på den overjordiske end den underjordiske (resultater ikke vist) biomasse.
For mælkebøtte var der ikke nogen forskel på ED90-doseringen af glyphosat for biomasse og frøsætning, hvilket indikerer, at planterne ikke kunne kompensere for skaden på lang sigt. Det modsatte var tilfældet for MaisTer, hvilket tyder på, at for dette herbicid var vækststandsningen kun midlertidig. Senere kunne planterne genoptage væksten og sætte frø. Det betyder, at der skal opnås en større effekt af MaisTer sammenlignet med glyphosat på biomassen for at hindre frøproduktion.
Resultaterne viser, at for græsserne havde udviklingstrinnet langt mindre betydning for effekten end for de tokimbladede arter (tabel 3.1 og 3.8). Undtagelsen var fåresvingel, hvor ED90-doseringerne af begge herbicider øgedes stærkt i strækningsfasen. En af årsagerne til, at der ikke var så stor forskel på doseringsbehovet på de to første udviklingstrin af græsserne, kan være, at græsserne på de tidlige udviklingstrin har meget oprette blade, hvor der kun sker en begrænset afsætning.
Tabel 3.8. Reduktion af overjordisk plantebiomasse og frøsætning som respons på herbiciddosering. ED90 er den beregnede dosis for 90% reduktion. 95%-konfidensintervallet er vist i parentes.
| Ukrudtsart og herbicid | Udviklingstrin | Biomasse ED90 |
Frøsætning ED90 |
|||||
| Alm. hvene Glyphosat |
1 sideskud | 50,4 | (39,6...61,2) | |||||
| (g as./ha) | 6 sideskud | 43,2 | (28,8...64,8) | |||||
| strækning | 25,2 | (18,0...36,0) | ||||||
| MaisTer | 1 sideskud | 1,30 | (0,992...1,61) | 2,08 | (0,310...4,50) | |||
| (g as./ha) | 6 sideskud | 2,08 | (1,71...2,39) | 0,900 | (0...5,80) | |||
| strækning | 2,08 | (1,71...2,48) | ||||||
| Fåresvingel Glyphosat |
6 sideskud | 115 | (93,6...137) | |||||
| (g as./ha) | 20 cm høj | 122 | (101...162) | |||||
| strækning | 227 | (180...317) | ||||||
| MaisTer | 6 sideskud | 6,60 | (5,49...7,69) | |||||
| (g as./ha) | 20 cm høj | 7,91 | (5,49...10,4) | |||||
| strækning | >124 | |||||||
| Mælkebøtte Glyphosat |
6 blade | 32,4 | (25,2...39,6) | 64,8 | (54,0...79,2) | |||
| (g as./ha) | 9 blade | 61,2 | (43,2...101) | 101 | (82,8...130) | |||
| 12 blade | 82,8 | (54,0..220) | 122 | (101...155) | ||||
| MaisTer | 6 blade | 0,279 | (0,217...0,775) | 0,868 | (0,713...0,992) | |||
| (g as./ha) | 9 blade | 0,589 | 1,18 | (1,18...1,61) | ||||
| 12 blade | 2,11 | (2,11...2,79) | ||||||
Alm. hvene og mælkebøtte var væsentlig mere følsomme over for begge herbicider end fåresvingel. Følsomheden af alm. hvene og mælkebøtte (tabel 3.8) var ikke væsentlig forskellig fra følsomheden af ukrudtsarter som sort natskygge og hanespore (tabel 3.1). Af de undersøgte arter vil fåresvingel således være den art, der vil blive påvirket mindst af sprøjtedrift fra disse to herbicider.
3.2.2 Vækststandsning
På flerårige arter fremkalder sublethale doseringer ofte en vækststandsning i en periode efter sprøjtning, hvorefter planten genoptager væksten ved at translokere reserver fra de underjordiske forrådsorganer. En sådan midlertidig væksthæmning kan have betydning for plantens konkurrenceevne.
Generelt var der stor variation i resultaterne af dosisresponsforsøgene på mælkebøtte og stor nælde. Det er et velkendt fænomen, at variationen i herbicidforsøg med flerårige arter er større end variationen i forsøg med enårige arter. Normalt vil man derfor øge antallet af gentagelser i forsøget for at opnå en større statistisk sikkerhed, men i dette forsøg, hvor der var inkluderet 5 forskellige høsttider (4 forskellige tidspunkter efter sprøjtning samt et hold planter til biomarkørtest), ville et øget antal gentagelser betyde en voldsom stor forøgelse af antallet af potter. Resultaterne må således tages med forbehold og kan udelukkende indikere, om det er muligt at afgøre, om der er tale om midlertidig væksthæmning og i givet fald anslå varigheden af en sådan væksthæmning.
3.2.2.1 Væksthusforsøg
En midlertidig vækststandsning er kendetegnet ved, at sluteffekten er mindre end effekten målt kort tid efter sprøjtning. Da forsøgene er opgjort som tørvægt relativt til ubehandlede planter (figur 3.17 og 3.18), er en midlertidig vækststandsning tilstede, når kurven for sluteffekt ligger over kurverne for relativ tørvægt 1, 2 og 3 uger efter sprøjtning.
3.2.2.2 Effekt af glyphosat i væksthus
For glyphosat på frøfremspiret mælkebøtte ses, at den relative biomasse 1, 2 og 3 uger efter sprøjtning var højere end eller lig med slutbiomassen (figur 3.17). Der er her tale om en stigende effekt med tid efter sprøjtning, hvilket også for de højeste doseringers vedkommende ses af, at biomassen 2 og 3 uger efter sprøjtning er lavere end 1 uge efter sprøjtning. På rodfremspiret mælkebøtte har de samme doseringer ikke givet nogen effekt på slutbiomassen. Her ses op til 60% effekt af flere doseringer ved høst kort tid efter sprøjtning. Væksthæmningens varighed afhænger af doseringen, og i de 4 laveste doseringer genoptog planterne væksten efter 3 uger.
På stor nælde blev der opnået høj sluteffekt på de frøfremspirede planter. Doseringerne blev reduceret på de rodfremspirede planter, og her blev der kun opnået ca. 20% effekt. Generelt er effekten af samme dosering væsentlig lavere på de rodfremspirede sammenlignet med frøfremspirede planter. I lighed med effekterne på mælkebøtte er der for de frøfremspirede planters vedkommende ikke tale om midlertidig vækststandsning, men om en fremadskridende og permanent vækststandsning. Det noget varierende forløb af kurverne for biomasse på rodfremspirede planter gør det vanskeligt at drage konklusioner. Sluteffekten er i flere tilfælde mindre end effekten kort tid efter behandling, hvilket indikerer, at der er tale om midlertidige væksthæmninger. Det parallelle forløb og placeringen af kurverne 1, 2 og 3 uger efter sprøjtning indikerer, at væksten er genoptaget 3 uger efter sprøjtning.
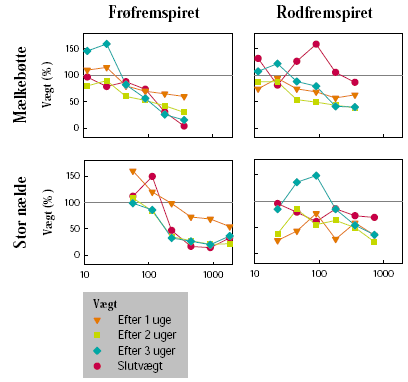
Figur 3.17. Glyph osateffekt på væksten af biomasse.
Biomassen blev målt 1, 2 og 3 uger efter sprøjtning på planter fremspiret fra frø eller rod. Slutvægten måltes efter 11 (fra frø) eller 24 (fra rod) uger for mælkebøtte og efter 13 (fra frø) eller 20 (fra rod) uger for stor nælde. Vægten er angivet som procent af vægten af ubehandlede planter.
3.2.2.3 Effekt af MaisTer i væksthus
På begge arter blev der anvendt højere doseringer af MaisTer på de rodfremspirede end på de frøfremspirede planter. Effekten over for mælkebøtte var væsentlig lavere på rodfremspirede end på frøfremspirede planter (figur 3.18). På de frøfremspirede planter var der kun i de to laveste doseringer tale om midlertidig vækststandsning, og væksten synes at være genoptaget 3 uger efter sprøjtning. På de rodfremspirede planter var der midlertidig vækststandsning i de 3 laveste doseringer, og her var væksten stadig hæmmet 3 uger efter sprøjtning. På stor nælde var sluteffekten på både frø- og rodfremspirede planter større end eller lig med effekten kort tid efter sprøjtning, og der var således ikke tale om midlertidig væksthæmning.
3.2.2.4 Effekter i væksthuset generelt
Generelt må det konkluderes, at frøfremspirede planter af flerårige arter er væsentligt mere følsomme end rodfremspirede planter. Midlertidige væksthæmninger blev i forsøgene observeret hos såvel frø- som rodfremspirede planter men er formentlig hyppigt forekommende hos planter, som kan translokere næring fra forrådsorganer til nye skud. I flere tilfælde synes væksten i forsøgene genoptaget 3 uger efter sprøjtning, men varigheden afhænger formodentlig af effekten af de anvendte doseringer, således at jo større effekt der opnås, jo længerevarende vækststandsning må forventes. Resultaterne viser, at der i modeller for flerårige arters vækst med herbicidpåvirkning bør indbygges en vækststandsning i minimum 3 uger, når planterne eksponeres for herbiciddoseringer, som forventes at give mellem 15 og 50% sluteffekt. Ved sluteffekter højere end 50% er der oftest tale om en permanent vækststandsning, og ved mindre end 15% effekt vil vækststandsningen være så kort, at den ikke skal indregnes.
En permanent langtidseffekt på flerårige arter forudsætter, at de underjordiske organer ødelægges. Translokationen af aktivstof ud i disse organer afhænger af, hvor systemisk herbicidet er, men flerårige arter med et udbredt rodsystem vil kunne overleve adskillige års eksponering af selv et meget systemisk herbicid som glyphosat.
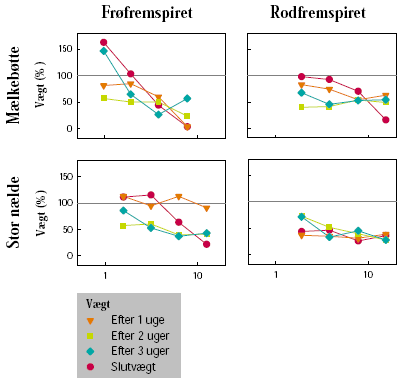
Figur 3.18. MaisTer-effekt på væksten af biomasse.
Biomassen blev målt 1, 2 og 3 uger efter sprøjtning på planter fremspiret fra frø eller rod. Slutvægten måltes efter 11 (fra frø) eller 24 (fra rod) uger for mælkebøtte og efter 13 (fra frø) eller 20 (fra rod) uger for stor nælde. Vægten er angivet som procent af vægten af ubehandlede planter.
3.2.2.5 Markforsøg
Den uforstyrrede vækst var for begge arter karakteriseret ved en tilvækst gennem hele sæsonen. I efteråret afsattes tørstof fra bladmassen ned i rødderne, som fungerer som reserveorgan vinteren over (figur 3.19).
De to arter reagerede modsat på herbiciderne: Mælkebøtte blev efter en kortvarig hæmning stimuleret til vækst og opnåede derved en større slutvægt end usprøjtede planter. For stor nælde derimod havde væksthæmningen blivende effekt, og for den art blev slutvægten mindre som følge af sprøjtningen (figur 3.19).
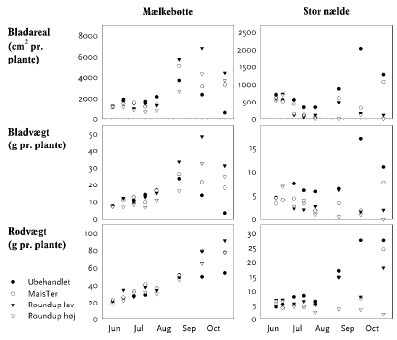
Figur 3.19. Herbicidforsøg i marken, Flakkebjerg 2006.
MaisTer: Mælkebøtte (25 g aktivstof/ha) og stor nælde (100 g aktivstof/ha).
Glyphosat: Mælkebøtte (lav 120 g aktivstof/ha, høj 360 g aktivstof/ha) og stor nælde (lav 540 g aktivstof/ha, høj 1080 g aktivstof/ha). Data vist som gns.±s.e.
3.2.3 Planters konkurrenceforhold
3.2.3.1 Simpelt respons uden konkurrence
Resultaterne af enkeltartsforsøg med alm. hvene og fåresvingel udført sideløbende med konkurrenceforsøget er vist i tabel 3.9. Fåresvingel var mere modstandsdygtig end alm. hvene og krævede en dosis, der var 2 til 9 gange større for at opnå samme effekt.
3.2.3.2 PCENV-estimering af respons på glyphosat
Efter en Box-Cox transformation (λ1 =-0,8λ2 =1) af både data og model viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 88% af den observerede variation.
Tabel 3.9. Reduktion af overjordisk plantebiomasse som respons på herbiciddosering. ED10, ED50 og ED90 er de beregnede doser for 10%, 50% og 90% reduktion. 95%-konfidensintervaller er vist i parentes.
| Herbicid og ukrudtsart |
ED10 | ED50 | ED90 | ||||||
| Glyphosat (g as./ha) Alm. hvene |
18,0 | (10,8...25,2) | 36,0 | (28,8...46,8) | 75,6 | (61,2...86,4) | |||
| Fåresvingel | 36,0 | (10,8...57,6) | 115 | (79,2...151) | 374 | (256...486) | |||
| MaisTer (g as./ha) Alm. hvene |
0,372 | (0,186...0,589) | 0,992 | (0,682...1,30) | 2,88 | (2,20...3.60) | |||
| Fåresvingel | 3,29 | (1,80...4,90) | 6,11 | (4,71...7,60) | 36,5 | (8,90...13,5) | |||
De estimerede parametre i konkurrencemodellen måler effekten af glyphosat på hele responskurven, og glyphosat havde en signifikant positiv effekt på biomassen af fåresvingel ved lav tæthed i de beskedne doser, som anvendtes i forsøget og en signifikant negativ effekt på biomassen af alm. hvene ved høje tætheder. Der var en signifikant negativ effekt af glyphosat på alm. hvenes konkurrenceevne overfor fåresvingel (tabel 3.10). Glyphosat skubbede altså til konkurrenceforholdet mellem fåresvingel og alm. hvene til fordel for fåresvingel.
Tabel 3.10. PCENV-parameterværdier for responset på glyphosat for fåresvingel (art 1) og alm. hvene (art 2). Værdier estimeret ved hjælp af maksimum likelihood og testet ved hjælp af likelihood ratio.
| Parameter | Værdi | Signifikans | ||
| a1 | –0,17 | - | ||
| a2 | –9,29 | - | ||
| b1 | –4,51 | - | ||
| b2 | –47,58 | - | ||
| c1 | –2,365 | - | ||
| c2 | 1,152 | - | ||
| d1 | –0,163 | Exp(1); P = 0,011 | ||
| d2 | 1,983 | |||
| f1 | 0,0459 | Exp(1); P = 0,016 | ||
| f2 | 2,663 | |||
| 1 | –0,00684 | ?0; P = 0,007 | ||
| 2 | –0,0616 | = 0; P = 0,17 | ||
| 0,00204 | = 0; P = 0,67 | |||
| 0,0674 | 0; P < 0,001 | |||
| 0,0211 | = 0; P = 0,072 | |||
| –0,00633 | 0; P < 0,001 |
ED10 af glyphosat på alm. hvene ved forskellige tætheder af fåresvingel og alm. hvene blev beregnet ud fra ligning 2 i afsnit 2.2.3. Medianværdien af ED10 for alm. hvene afhang af tætheden af fåresvingel og alm. hvene (figur 3.20), dog var der en betydelig usikkerhed på de beregnede ED10-værdier(figur 3.21 og 3.22). Den beregnede ED10 fra konkurrenceforsøget er sammenlignelig med den estimerede ED10 fra standardtesten (= 19.0 med et 95% konfidensinterval mellem 12.1 og 25.9; fra tabel 3.9).
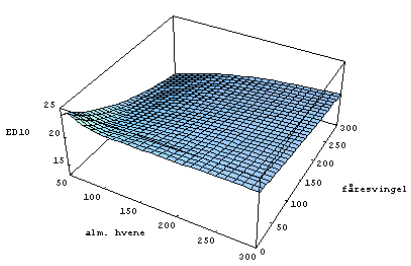
Figur 3.20. Effekten af glyphosat på alm. hvene i konkurrence med fåresvingel.
Den beregnede medianværdi af ED10 (10% biomassetab) for alm. hvene som funktion af tætheden af fåresvingel og alm. hvene (m–2).
Trods den betydelige usikkerhed på den beregnede ED10 var der en signifikant men lille effekt af den intra-specifikke konkurrence på den beregnede ED10 for alm. hvene, således at der ved højere tæthed af planten kræves en relativ lavere dosis glyphosat for at reducere biomassen med 10% (ED10(0,100)-ED10(0,200) > 0, P = 0.03). Denne lavere dosis er dog ubetydelig (median = 0.06) i forhold til usikkerheden på de beregnede ED10 værdier (figur 3.21).
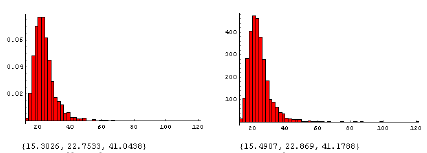
Figur 3.21. Effekt af glyphosat på alm. hvene i intraspecifik konkurrence.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 200 m-2 (til venstre) og 100 m-2 (til højre). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
Derimod var der en betydelig signifikant effekt af fåresvingel på ED10 for alm. hvene (ED10(0,100)-ED10(100,100) > 0, P = 0.03), således at glyphosat har en kraftigere effekt på alm. hvene, når alm. hvene konkurrerer med fåresvingel (figur 3.22). Den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10%, er ca. 16% lavere, hvis alm. hvene konkurrerer med fåresvingel i forholdet 1:1 i forhold til, hvis alm. hvene kun konkurrerer med sig selv. Hvis forholdet af fåresvingel er højere end 1:1, da reduceres den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10% yderligere (figur 3.20).
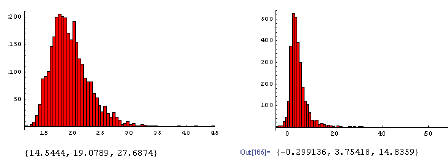
Figur 3.22. Effekt af glyphosat på alm. hvene i konkurrence med fåresvingel.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 100 m–2 for alm. hvene og 100 m–2 for fåresvingel (A) og fordelingen af ED10 (alm. hvene:100 m–2) – ED10 (alm. hvene:100 m–2, fåresvingel: 100 m–2). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
3.2.3.3 PCENV-estimering af respons på MaisTer
Efter en Box-Cox transformation ![]() af både data og model (2.4.2.2.1) viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 66% af den observerede variation.
af både data og model (2.4.2.2.1) viste forskellige residualplots, at residualvariationen kunne antages at være både homogen og normalfordelt, og modellen forklarede 66% af den observerede variation.
De estimerede parametre i konkurrencemodellen måler effekten af MaisTer på hele responskurven, og MaisTer havde en signifikant positiv effekt på biomassen af fåresvingel ved både lav og høj tæthed i de beskedne doser, som anvendtes i forsøget, og en signifikant negativ effekt på biomassen af alm. hvene ved både lave og høje tætheder. Der var en signifikant positiv effekt af MaisTer på fåresvingels konkurrenceevne overfor alm. hvene og omvendt en signifikant negativ effekt af MaisTer på alm. hvenes konkurrenceevne overfor fåresvingel (tabel 3.11).
Tabel 3.11. PCENV-parameterværdier for responset på MaisTer for fåresvingel (art 1) og alm. hvene (art 2). Værdier estimeret ved hjælp af maksimum likelihood og testet ved hjælp af likelihood ratio.
| Parameter | Værdi | Signifikans | ||
| a1 | 9.68327 | - | ||
| a2 | –21.5456 | - | ||
| b1 | –67.8936 | - | ||
| b2 | –50.5823 | - | ||
| c1 | –0.988756 | - | ||
| c2 | 1.81707 | - | ||
| d1 | 2.76147 | ≠Exp(1); P < 0,001 | ||
| d2 | 2.13329 | |||
| f1 | 4.39852 | ≠Exp(1); P < 0,001 | ||
| f2 | 2.77588 | |||
| 1 | –12.6327 | 0; P = P < 0,001 | ||
| 2 | 4.95714 | 0; P = P < 0,001 | ||
| –2.03603 | = 0; P = 0,69 | |||
| 3.11466 | 0; P < 0,001 | |||
| 0.431216 | 0; P < 0,001 | |||
| –0.237765 | 0; P < 0,001 |
Effekten af MaisTer på alm. hvene ved forskellige tætheder af fåresvingel og alm. hvene blev beregnet ved ED10 [ligning 2]. Medianværdien af ED10 for alm. hvene afhang af tætheden af fåresvingel og alm. hvene (figur 3.23), dog var der en betydelig usikkerhed på de beregnede ED10 (figur 3.24 og 3.25). Den beregnede ED10 fra konkurrenceforsøget er sammenlignelig med den estimerede ED10 fra standardtesten (= 0,4 med et 95% konfidensinterval mellem 0,2 og 0,4; fra tabel 3.9).
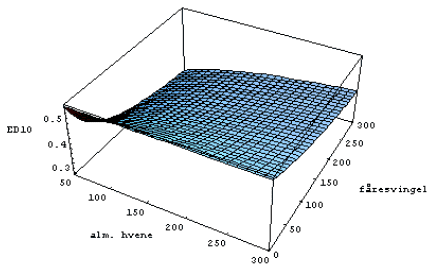
Figur 3.23. Effekten af MaisTer på alm. hvene i konkurrence med fåresvingel.
Den beregnede medianværdi af ED10 (10% biomassetab) for alm. hvene som funktion af tætheden af fåresvingel og alm. hvene (planter pr. m²).
I modsætning til glyphosat var der ingen signifikant effekt af den intraspecifikke konkurrence på den beregnede ED10 for MaisTer på alm. hvene (figur 3.24; P = 0,78). Men ligesom for glyphosat var der en betydelig signifikant effekt af fåresvingel på ED10 for MaisTer på alm. hvene (ED10(0,100)-ED10(100,100) > 0, P = 0,03), således at MaisTer havde en kraftigere effekt på alm. hvene, når den var i konkurrence med fåresvingel end uden (figur 3.25). Den dosis MaisTer, som kræves for at reducere biomassen af alm. hvene med 10%, er ca. 33% lavere, hvis alm. hvene konkurrerer med fåresvingel i forholdet 1:1 i forhold til, hvis alm. hvene kun konkurrerer med sig selv. Hvis forholdet af fåresvingel er højere end 1:1, da reduceres den dosis glyphosat, som kræves for at reducere biomassen af alm. hvene med 10% yderligere (figur 3.23).
ED10 for MaisTer på fåresvingel kunne ikke estimeres troværdigt ud fra ligning 2 på grund af for lave herbiciddoser i konkurrenceforsøget (resultater er ikke vist).
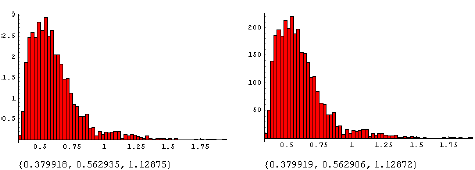
Figur 3.24. Effekt af MaisTer på alm. hvene i intraspecifik konkurrence.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 200 m-2 (til venstre) og 100 m-2 (til højre). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
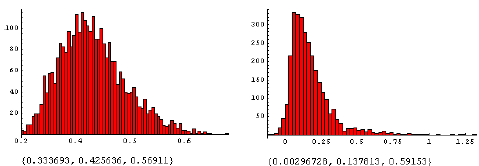
Figur 3.25. Effekt af MaisTer på alm. hvene i konkurrence med fåresvingel.
Den beregnede fordeling af ED10 (10% biomassetab) ved tætheder på 100 m–2 for alm. hvene og 100 m–2 for fåresvingel (A) og fordelingen af ED10 (alm. hvene:100 m–2) – ED10 (alm. hvene:100 m–2, fåresvingel: 100 m–2). Tallene under figuren er {2.5%, 50%, 97.5%} percentilerne.
3.2.4 Plantesamfundets succession
3.2.4.1 Frøbankens påvirkning af glyphosat og kvælstof
Det totale antal frø, der spirede og udviklede sig til identificerbare planter, var 7906. Fire spirer udviklede sig aldrig til planter, der kunne artsbestemmes. Tokimbladede arter var talrigest repræsenteret og udgjorde 70% af det samlede antal fremspirede planter. Knap 45% eller 3522 planter var en- eller toårige arter og tilhører således en gruppe, der for at være repræsenteret i floraen skal forekomme i frøbanken.

Figur 3.26. Fotoet viser 4 spiringsbakker udvalgt fra en tilfældig transekt. Prøverne er fra plot, der har modtaget 100 kg N/ha, og bakkerne er opstillet således, at de fra venstre mod højre indeholder prøver fra plot, der modtaget en stigende glyphosatdosering (0, 14, 4, 72 og 360 g as./ha).
Variansanalysen viste, at glyphosat reducerede frøbanken med stigende dosis (P=0,008, figur 3.26), mens gødskningen ingen effekt havde (P=0,12). Der var ingen interaktion mellem de to variable. Den samlede mængde af spiringsdygtige frø i frøbanken var afhængig af den behandling plottet havde modtaget (figur 3.27)
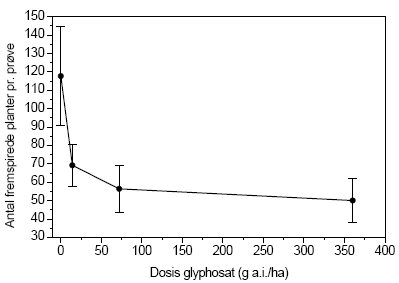
Figur 3.27. Total fremspiring af frø (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, som funktion af årlig glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
En opdeling af de fremspirede frø i en- og tokimbladede arter (figur 3.28) viste, at tokimbladede arter udgjorde langt den største del af planterne, og at antallet af disse faldt mest markant (godt 60 %) med stigende glyphosatdosis (P=0,03), mens antallet af enkimbladede planter var betydelig lavere og var relativt upåvirket af glyphosatdosis (P=0,07). Gødskning påvirkede frøbanken positivt for de tokimbladede (P=0,001), mens de enkimbladede var upåvirkede (P=0,18). Ved det højeste kvælstofniveau (100 kg N/ha) var der dog et tydeligt fald i antallet af spiringsdygtige frø med stigende glyphosatdosering.
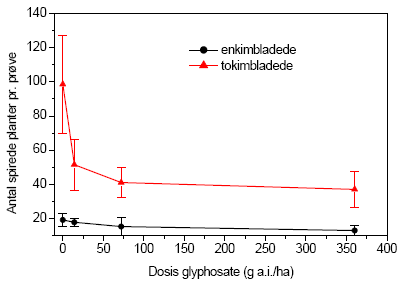
Figur 3.28. Fremspiring af planter (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, fordelt på en- og tokimbladede arter som funktion af glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
Den samlede fremspiring af en- og toårige arter (figur 3.29) viste en negativ påvirking af frøbanken med stigende glyphosatdosering (P=0,002) og en positiv påvirkning af gødskning (P=0,0002). Der var ingen interaktion mellem parametrene.
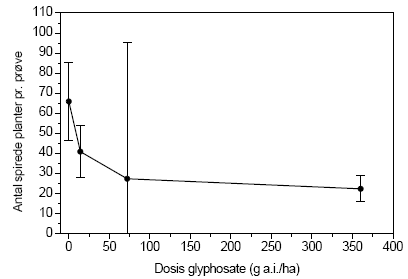
Figur 3.29. Fremspiring af en- og toårige arter (gns.±s.e.) fra frøbankprøver indsamlet i forsøgsplottene på Kalø, september 2004, som funktion af glyphosatdosis ved den højest anvendte gødskning på 100 kg N/ha/år.
Der blev fundet 40 plantearter i frøbanken, hvoraf 33 var tokimbladede og 7 enkimbladede. Fordelingen af spirede frø på de enkelte arter og behandlinger fremgår af Bilag C. Blandt de enkimbladede var en art en hybrid mellem græsserne alm. hvene (Agrostis capillaris) og stortoppet hvene (A. gigantea). Blandt de oprindeligt såede 33 arter, hvoraf 27 var etablerede og fortsat tilstede i plottene ved sidste forudgående registrering i 200324, fandtes spiringsdygtige frø af 17 af disse arter. De øvrige 23 arter var dels almindelige markukrudtsarter, en almindelig skovukrudtsart (alm. gederams, Epilobium angustifolium) og træet dunbirk (Betula pubescens).
Fire arter, rejnfan (Tanacetum vulgare), filtbladet kongelys (Verbascum thapsus), alm. stedmoderblomst (Viola tricolor) og stortoppet hvene (Agrostis gigangtea), udgjorde samlet 85% af de fremspirede planter og henholdsvis 19, 24, 14 og 28%. De øvrige arter var ofte ganske fåtallige, og kun 4 arter (mælkebøtte, prikbladet perikon, fåresvingel og hvene-hybriden) blandt de resterende bidrog med mere end 1%.

Figur 3.30. Forsøglokaliteten nær Kalø i august 2006.
3.2.4.2 Vegetationens påvirkning af glyphosat og kvælstof
Selv med det blotte øje kunne man se, at vegetationen var påvirket af behandlingerne (figur 3.30). Sprøjtningen med glyphosat forårsagede større områder med bar jord eller død vegetation to uger efter behandlingen i sammenligning med usprøjtede plots (figur 3.31), og der skete ikke noget fald i andelen af bar jord over den treårige projektperiode (dvs. år 4-6 i successionen)(figur 3.32). For hovedparten af arterne faldt dækningen (figur 3.32) og biomassen (figur 3.34) med stigende glyphosatdosering. Det gjaldt f.eks. alm. hvene, kvik, stortoppet hvene og rejnfan. Fåresvingel var den eneste art, der havde en stigende dækning og biomasse med stigende glyphosatdosering.
Kvælstof havde en positiv virkning på dækning og biomasse af de to græsser, kvik og stortoppet hvene (P<0,0001), men for de øvrige arter var der ingen (f.eks. rejnfan) eller en negativ effekt af kvælstof.
Der var enkelte signifikante tredjeordens interaktioner (år´art´plot), hvor udviklingen af en enkelt art var anderledes end de viste gennemsnit (figur 3.32 og 3.34), men det drejede sig højst om ét plot pr. behandling.
Græsser udgjorde ved alle behandlinger og tidspunkter omkring 50% eller mere af den samlede dækning. Gennem årene var det dog forskellige græsarter, der bidrog til dækningen. I 2005 dominerede alm. hvene, fåresvingel og stortoppet hvene, mens kvik havde en ubetydelig dækning. Dækningen af kvik øgedes imidlertid betydeligt gennem projektperioden navnlig ved det højeste kvælstofniveau, hvor kvik i 2007 nåede en dækning på 1/3 af arealet undtagen ved den højeste dosis af glyphosat, hvor dækningen af de øvrige græsarter navnlig fåresvingel var højere (P<0,0001)(figur 3.32).
Fåresvingel var den dominerende græs ved højeste dosis af glyphosat uanset kvælstofniveauet. Ved lavere glyphosatdosis var alm. hvene dominerende, medens kvik var dominerende ved højt kvælstofniveau (figur 3.32).
Artsantallet var generelt faldende med stigende glyphosatdosis (figur 3.33). Blandt de 33 arter, der blev sået i 2001 (Bilag C), spirede og etablerede alle arter sig med undtagelse af knoldet mjødurt (Filipendula vulgaris). Agertidsel (Cirsium arvense) blev ikke registreret i det tidligere projekt24 men blev fundet i hele denne projektperiode 2005-2007. Desuden blev få levedygtige frø registreret i frøbanken i 2004. Hvorvidt agertidsel er etableret fra de oprindeligt såede frø eller fra frø, der efterfølgende er spredt til området, vides ikke, men agertidsel har frø, der overlever længe i frøbanken109. På trods af at der blev fundet en stor mængde spiringsdygtige frø i frøbanken, udgjorde andre arter end de tre græsser alm. hvene, fåresvingel og kvik en faldende andel af den samlede dækning ved alle behandlinger (figur 3.32).
Seksogtyve arter af de 33 såede arter findes fortsat i forsøgsområdet. Blandt de seks forsvundne arter er tre arter alm. dværgløvefod (Aphanes arvensis), enårig rapgræs (Poa annua) og kornblomst (Centaurea cyanus) ikke registreret inden for plottene siden 2001, haremad (Lapsana communis) og alm. kællingetand (Lotus corniculatus) ikke siden 2003 og alm. pimpinelle (Pimpinella saxifraga) og alm. agermåne (Agrimonia eupatoria) ikke siden 2005. Kun 9 arter forekommer hyppigt, se Bilag C.
Blandt de nye arter, der er registreret, forekommer kun alm. røllike (Achillea millefolim), gul snerre (Galium verum), smalbladet rapgræs (Poa annua) og rødknæ (Rumex acetosella) ved gentagne registreringer i den samme blok og kan dermed anses for etablerede. Blandt de sporadisk forekommende arter er en del enårige ukrudtsarter f.eks. hvidmelet gåsefod (Chenopodium album), burresnerre (Galium aparine), vej pileurt (Polygonum aviculare) og snerlepileurt (Fallopia convolvulus).

Figur 3.31. Glyphosateffekt på etableret vegetation.
Usprøjtet plot (venstre) og plot sprøjtet med 360 g glyphosat pr. ha (højre) 2 uger efter sprøjtning.
Den mest markante forandring i forsøgsperioden (2005-2007) var den voldsomme vækst af kvik (figur 3.32 og 3.34). I 2003 udgjorde kvik <0,1% af den høstede biomasse. I 2007 var biomassen steget til 35-45 g/m² ved høj kvælstoftilførsel uanset glyphosatdoseringen. Stigningen modsvaredes af et tilsvarende fald i biomassen af alm. hvene (figur 3.34) Biomassen af fåresvingel var højest ved de høje glyphosatdoseringer og nogenlunde uforandret gennem forsøgsperioden (figur 3.34), selvom dækningen steg i perioden (figur 3.32).
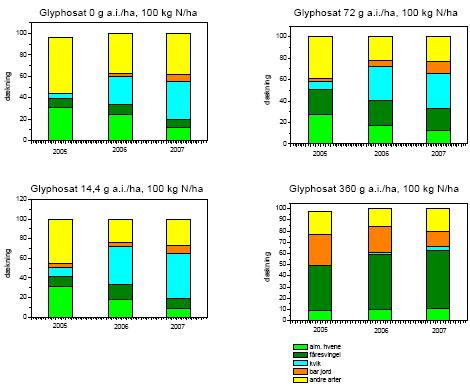
Figur 3.32. Udviklingen i dækningen fra 2005 til 2007 af udvalgte arter, andre arter samt af bar jord.
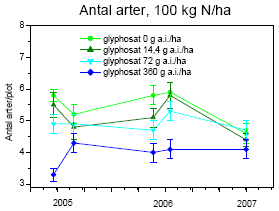
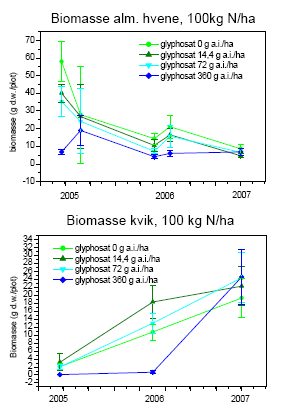
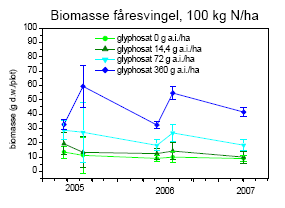
Figur 3.34. Udvikling i plantebiomasse fra 2005 til 2007. Data viser gns.±s.e. (n=10).
3.2.4.3 Vegetationens påvirkning af MaisTer
Undersøgelsen af effekter af MaisTer var ikke designet således, at vi kan adskille effekten af gødskning fra effekten af herbicidet. De effekter, vi ser, er kombinationseffekter. Ved sammenligning af udviklingen med og uden herbicidbehandling kan faldet i antal arter (figur 3.35) og i dækningen af fåresvingel og rejnfan (figur 3.36) umiddelbart efter første sprøjtning i 2005 bedst forklares som en herbicideffekt, mens alm. hvene i mindre grad påvirkedes af MaisTer. Gødskningen stimulerede tydeligvis både alm. hvene og rejnfan, hvorimod dækningen af fåresvingel reduceredes efter kvælstoftilførslen. Fåresvingel er en langsomt voksende græs, der er mest konkurrencedygtig ved lave kvælstofniveauer, hvilket kan forklare, at dækningen af fåresvingel reduceredes ved begge behandlinger.
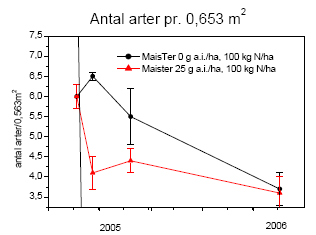
Figur 3.35. Artsantallet efter behandling med MaisTer (25 g as./ha) i forhold til ubehandlet; begge gødet med 100 kg N/ha. Den vertikale linie angiver behandlingstidspunktet. Punkter viser gns.±s.e.
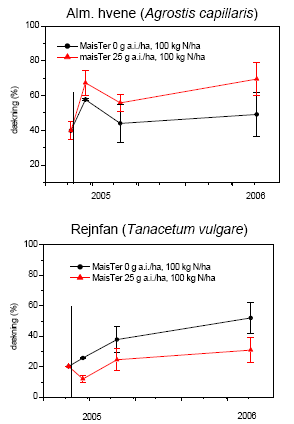
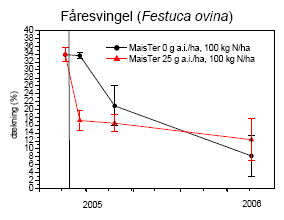
Figur 3.36. Dækningen af tre udvalgte arter efter behandling med MaisTer (25 g as./ha) i forhold til ubehandlet; begge gødet med 100 kg N/ha. Den vertikale linie angiver behandlingstidspunktet. Punkter viser gns.±s.e.
3.2.5 Modellering af fodposeplanternes populationsdynamik
3.2.5.1 Kalibrering af modellen
Modellen blev kalibreret til at simulere væksten af mælkebøtte, kvik, alm. hvene, fåresvingel, agertidsel, rejnfan, stor nælde og vild kørvel ved at tilpasse modellens parametre til beskrivelse af væksten til de indsamlede vækstdata på de otte arter (tabel 2.15). Tilpasningen blev foretaget ved at ændre på parametrene, indtil simuleringen af tørvægt og antal af plantens forskellige overjordiske organer (blade, stængler, knopper, blomster og frugter) stemte tilfredsstillende overens med observerede data. I denne proces blev benyttet både visuel sammenligning af feltdata og modeloutput samt optimering af modellens simuleringer ved hjælp af mindste kvadraters metode. Ved den sidste metode foretog modellen en lang række gennemregninger med tilfældigt valg af udvalgte parametre fra et interval, og efter hver beregning summerede den kvadraterne på forskellen mellem simulerede og observerede data. Den kombination af parametre, der gav det mindste kvadrat, blev så benyttet i den videre parameterisering. Da der er mange lokale minimumspunkter på sammenhængen imellem forskellige kombinationer af vækstparametre og kvadratet på afvigelsen, viste det sig hensigtsmæssigt at benytte en kombination af den visuelle bedømmelse af simuleringerne og mindste kvadraters metode.
Efter at modellen var blevet kalibreret til at simulere væksten af en planteart med hensyn til tørvægt og antal af de forskellige planteorganer, blev den kalibreret til at simulere højdefordelingen af disse organer ved hjælp af en tilsvarende kombination af den visuelle metode og mindste kvadraters metode.
Under kalibreringsprocessen blev der benyttet simuleringer over en seksårig periode, hvor der blev benyttet klimatisk input fra 2000 frem til 2006. Modellens output fra 2005 blev så benyttet i sammenligningerne med de observerede data, der stammede fra netop dette år. Modellen blev kalibreret ved hjælp af disse langtidssimuleringer, fordi de indsamlede data stammede fra flerårige planter, der havde vokset på de pågældende steder igennem flere år. FieldEco kan bringes til at simulere en plantes vækst ved hjælp af mange forskellige kombinationer af parametre, men ved at benytte tilpasning til både tørvægt og antal af alle overjordiske organer ved langtidssimuleringer er der sikret en realistisk parameterisering. Hvis parameteriseringen ikke lå indenfor realistiske rammer, opstod der ubalance imellem plantens forskellige organer i den simulerede plante, og simuleringerne kunne ikke forløbe over flere år.
Effekten af herbiciderne glyphosat og MaisTer blev kalibreret ved at simulere en sprøjtning på en given data og ved at justere effekten af herbicidet på plantens totale overjordiske biomasse til det observerede effektniveau (observeret i dette projekt eller fra andre kilder) fire uger senere.
Tilpasningsprocessen illustreres her med rejnfan som eksempel. Simuleringen af de observerede data (figur 3.37) viser, at modellen kunne simulere væksten igennem sæsonen ganske godt. Simuleringen af tørvægt af blade og stængler igennem sæsonen passer godt med data, mens det viste sig umuligt at få simuleringerne af antallet af blade sidst på sommeren til at passe med data. Dette skyldes, at rejnfan producerer en stor mængde relativt små blade i den sidste del af sommeren, og FieldEco er ikke programmeret til at simulere en variation i bladstørrelsen igennem sæsonen. Blev modellen kalibreret til at simulere antallet af blade korrekt, blev tørvægten af bladene for høj, og blev modellen kalibreret til at simulere tørvægten korrekt, blev antallet af blade midt på sommeren for lavt. Det var derfor nødvendigt at vælge enten det ene eller det andet, og der blev givet prioritet til tørvægten, da tørvægten er en drivende variabel i modellen. Dette problem gjorde sig også gældende for stor nælde og agertidsel, hvor der også blev givet prioritet til tørvægten. Simuleringen af tørvægten af blomsterstande passer heller ikke helt med data, da det ikke var muligt at få både antal og tørvægt af frugtstande på sidste dato til at passe godt. Det var ikke muligt at simulere et fald i tørvægt samtidig med en stigning i antal, hvilket må skyldes, at de sidst udviklede frugtstande bliver mindre end de først udviklede. Da FieldEco opererer med samme parameterisering af frugtudviklingen igennem hele sæsonen, var det ikke muligt at simulere dette. Der blev derfor også her givet prioritet til at simulere tørvægten godt.
FieldEco blev kalibreret til at simulere de øvrige syv plantearter på samme måde som rejnfan. Der var dog lidt forskel på, hvor mange planteorganer, der indgik i kalibreringen. Ved græsserne gav det ikke mening at skelne imellem stængler og blade, og ved kvik var blomstring og frøsætning så begrænset, at den blev ignoreret, det vil sige, at kun bladene indgik i parameteriseringen af kvik. Hos stor nælde var det meget svært at skelne imellem knopper, blomster (både han- og hunblomster) og frø under feltforhold, hvorfor de blev slået sammen som reproduktive organer.
3.2.5.2 Validering af modellen
Til validering af modellen anvendtes data, der var uafhængige af dem, der blev anvendt til kalibreringen. For mælkebøtte og stor nælde data fra Flakkebjerg 2006 (afsnit 2.2.2) og for alm. hvene, fåresvingel og kvik, anvendtes data fra successionsforsøget ved Kalø (afsnit 2.2.4). Valideringen blev foretaget uden ændring af modellens parameterværdier, hvor intet andet er anført.
For at simulere Flakkebjerg-data blev FieldEco blev sat op til at simulere et enkelt års vækst af enten mælkebøtte eller stor nælde med anvendelse af klimadata (temperatur og globalstråling) fra Forskningscenter Flakkebjergs klimastation
For at simulere Kalø-data blev FieldEco sat op til at simulere konkurrencen imellem fåresvingel, alm. hvene og kvik over en 6-årig periode, hvor der blev sprøjtet med RoundUp og gødsket, som angivet i tabel 2.14. Simuleringerne blev startet med 10 frø pr. ¼ m² af alm. hvene og fåresvingel den 11. april 2001, og med en enkelt kimplante af kvik den 25. juli 2004 ved høj kvælstoftilførsel (100 g N/ha/år). Kvik blev introduceret senere i simuleringen, da kvik indvandrede efter etableringen af forsøget. Der blev anvendt klimadata fra DMUs klimastation i Silkeborg. Simuleringerne blev foretaget med parameteriseringen af plantearterne, som beskrevet i afsnit 2.2.5.
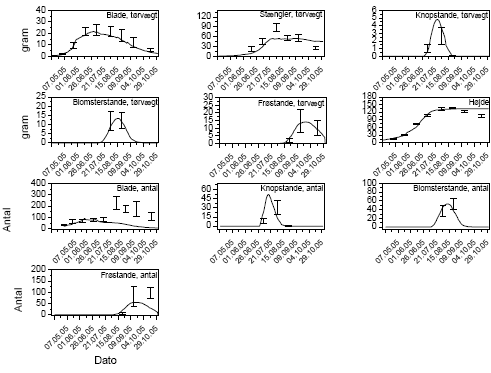
Figur 3.37. Tilpasning af modellen til at simulere parameteriseringsdata på rejnfan
(Tanacetum vulgare). Sammenligning imellem simulering (linien) og observerede værdier (gns.±s.e.).
Mælkebøtte
Feltdata viser en meget kraftig vækst af både rod og skud i august og september, hvilket modellen ikke er i stand til at simulere (figur 3.38). Der er dog foretaget en opjustering af kvælstofniveauet i simuleringen, da dette ikke kan antages at have været som i jorden, hvor planterne til parameterisering blev dyrket. Simuleringen uden anvendelse af herbicider viser stort set den observerede udviklingstendens, men når ikke det høje niveau fra observationerne, hvorimod simuleringen af rodbiomassen skyder klart under observationerne. Modellen er ikke i stand til at simulere faldet i skudbiomasse i juli-august, hvilket antagelig skyldes den tørkesituation, der forekom i denne periode, og som modellen ikke er programmeret til at tage i betragtning.
Modellen er blevet kalibreret til at simulere flerårige planter, mens der her er tale om planter, der er spiret fra frø det samme år. Der er søgt taget højde for dette ved at forhindre blomstring og udsætte allokering af ressourcer til oplagringsorganer til midt på sommeren, men modellen er faktisk ikke kalibreret til at håndtere førsteårsplanter. Tendenserne fra kontrollen gør sig i store træk også gældende for simuleringerne af situationerne med sprøjtninger med MaisTer og glyphosat, dog er modellen i højere grad i stand til at simulere skududviklingen i efterårsperioden. Simuleringen af sprøjteeffekterne fungerer tilfredsstillende ved at opfange tendenserne i udviklingen, det vil sige en reduktion af væksten efter sprøjtningen den 19. juni, hvorefter væksten genoptages. Tendenserne i simuleringerne af situationerne efter sprøjtning med glyphosat er i store træk sammenfaldende med observationerne, selv om niveauet i efteråret heller ikke her bliver fanget af simuleringen.
Det høje niveau i skudvægt i efteråret efter behandlingen med MaisTer oversteg niveauet fra kontrollen, hvilket måske skal tilskrives tilfældig variation af de indsamlede planter. Resultaterne skal i øvrigt ses i lyset af en meget stor variation imellem planterne i efterårsperioden og dermed følgende risiko for fluktuerende resultater.
Med hensyn til giftvirkningen i simuleringerne var det nødvendigt at reducere effekten af både glyphosat og MaisTer betydeligt i forhold til parameteriseringen ifølge resultaterne fra væksthus.
Stor nælde
Modellen simulerer kontrollen ganske godt men er ligesom ved mælkebøtte ikke i stand til at simulere vækststandsningen under tørken i juli-august, da den ikke er programmeret til det (figur 3.39). Simuleringen uden for denne periode er god. Med hensyn til simuleringen af effekten af glyphosat og MaisTer simuleres væksthæmningen ganske godt, men modellen overvurderer skudvæksten efter sprøjtningen. Dette skyldes antagelig igen tørken i juli-august, som sammen med giftvirkningen har nogle konsekvenser for planten, som FieldEco ikke tager med.
Simuleringerne underestimerer rodvægten i starten af sæsonen i alle simuleringer, hvilket hænger sammen med, at modellen ikke er blevet kalibreret til at simulere denne situation, hvor planterne er spiret fra frø det samme år. Ligesom for mælkebøtte er modellen kun blevet kalibreret til situationen med flerårige planter. Derfor er det faktisk ganske tilfredsstillende, at modellen alligevel fanger udviklingen i rodudvikling på disse førsteårsplanter, selv om den ikke rammer helt præcist fra starten, hvor spiringen er tættest på.
Almindelig hvene, fåresvingel og kvik
Der blev ikke foretaget ændringer i parameteriseringen af planternes vækst for at simulere de observerede data på fåresvingel, alm. hvene og kvik fra Kalø-eksperimenterne. Det var dog nødvendigt at foretage en justering af den tilgængelige mængde kvælstof i forhold til data. Dette blev udelukkende gjort i forhold til data uden sprøjtning med glyphosat (figur 3.40). De første simuleringer med glyphosatsprøjtninger viste, at planterne fra Kalø-felterne var klart mindre sensitive overfor dette stof, end data fra væksthuset viste. Det var derfor nødvendigt at reducere effekten af glyphosat for alle tre arter betydeligt.
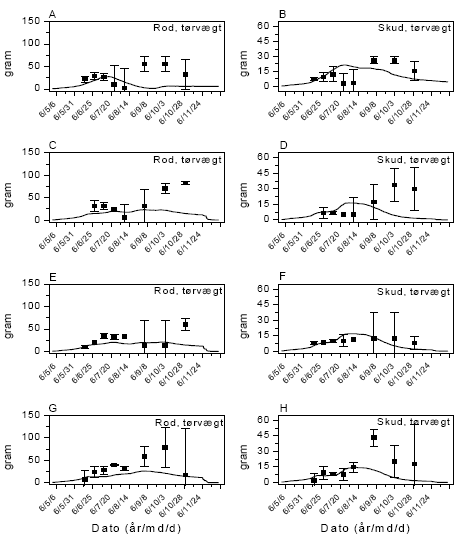
Figur 3.38. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af mælkebøtte. Kontrol (A,B), MaisTer (C, D), glyphosat lav (E,F) og glyphosat høj (G,H).
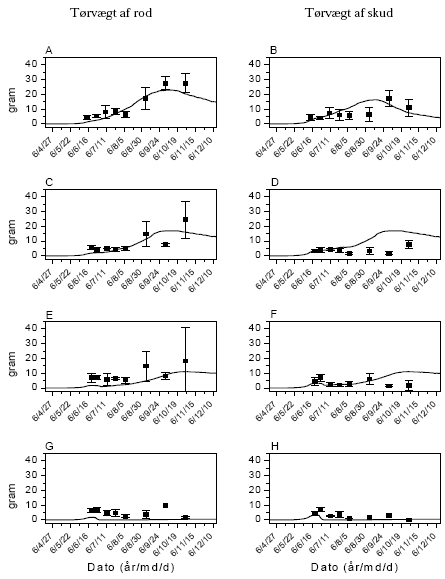
Figur 3.39. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af stor nælde. Kontrol (A,B), MaisTer (C, D), glyphosat lav (E,F) og glyphosat høj (G,H).
Modellen simulerer de observerede data ved lavt niveau af glyphosat for alle tre arter og viser, at FieldEco er i stand til at simulere plantekonkurrence over flere år (figur 3.41 og 3.42). Ved simulering af behandlingen med højeste dosis glyphosat opstår der imidlertid en underestimering af fåresvingels vækst, mens simuleringen af væksten af kvik og alm. hvene stadigvæk er god (figur 3.43). Problemerne med fåresvingel ved den høje belastning med glyphosat skyldes muligvis, at FieldEco ikke fanger fåresvingels tilvækst om vinteren. Dette bliver antagelig mere tydeligt ved høj belastning af glyphosat, hvor kvik og alm. hvene går tilbage (hvilket modellen fanger) og efterlader bedre plads til fåresvingel, som stort set ikke påvirkes af glyphosat. I simuleringen med høj glyphosatbelastning reagerer fåresvingel i overensstemmelse med observationerne med en højere biomasse i forhold til kontrollen men når ikke helt op på det observerede niveau. Denne mangel skyldes muligvis, at FieldEco ikke fanger den observerede vækst i efterårs- og vinterperioden, som derved mangles i simuleringen.
Sammenfaldet imellem simuleringer og observationer er dog generelt så godt, at det må konkluderes, at modellen er i stand til at simulere konkurrencen imellem de tre involverede arter over 6 år ved 4 forskellige påvirkninger af glyphosat (dog til dels med fåresvingel ved de højeste glyphosatniveauer som undtagelser).
Konklusion på validering
Simuleringerne af feltdata af fåresvingel, alm. hvene og kvik fra Kalø viser, at FieldEco er i stand til at simulere konkurrence imellem planter gennem flere år under påvirkning af glyphosat. Simuleringerne af feltdata fra Flakkebjerg faldt nogenlunde ud for stor nælde men mindre godt for mælkebøtte. Dette kan skyldes, at FieldEco ikke er blevet kalibreret til at simulere situationen med førsteårsplanter. Ydermere er mælkebøtte en meget variabel art, og mælkebøtterne fra Flakkebjerg kan meget vel være af en anden underart end den, som FieldEco er blevet kalibreret til.
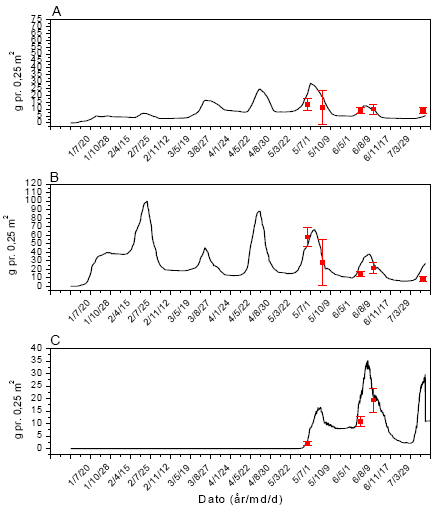
Figur 3.40. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) uden glyphosatpåvirkning.
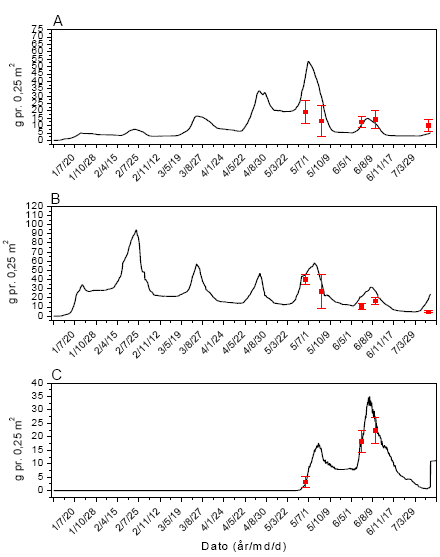
Figur 3.41. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 12 g glyphosat pr. ha.
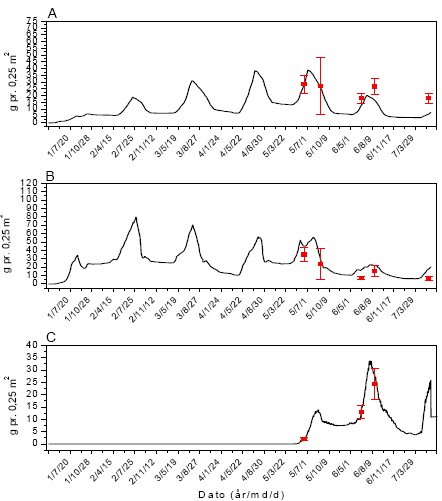
Figur 3.42. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 61 g glyphosat pr. ha.
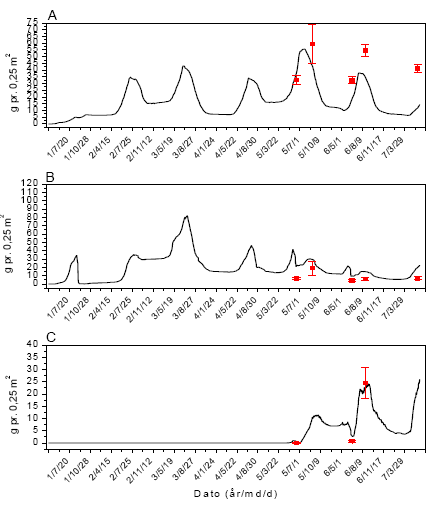
Figur 3.43. Sammenligning imellem simuleringer (linier) og punktobservationer (gns.±s.e.) af vækst af fåresvingel (A), alm. hvene (B) og kvik (C) ved årlig behandling med 306 g glyphosat pr. ha.
3.2.5.3 Simulering af scenarier
Alle scenarier blev sat op som 7 års simuleringer af effekterne på hegnsvegetationen af sprøjtestrategierne relateret til 4 forskellige sædskifter (afsnit 1.7). En stor del af de danske levende hegn og markskel består af græsser, hvor kvik er meget stærkt repræsenteret, og spredte flerårige urter. Derfor er scenarierne sat op som scenarier, hvor de flerårige urter agertidsel, mælkebøtte, rejnfan, vild kørvel og stor nælde udsættes for en herbicidpåvirkning, mens de vokser i konkurrence med kvik. Herved opnås en simulering af den kombinerede effekt af herbicidpåvirkningen og konkurrencen med kvik. Dette gjorde sig dog i simuleringerne kun gældende for mælkebøtte og vild kørvel, mens det viste sig umuligt at simulere sameksistens imellem kvik og henholdsvis rejnfan, agertidsel og stor nælde. I alle disse tilfælde blev kvik skygget væk i løbet af 2-3 år. Dette er imidlertid ikke specielt overraskende, da især stor nælde og rejnfan men også i nogen grad agertidsel er bestanddannende monopoliserende arter i naturen. I naturen forekommer de som regel i tætte bestande og ikke blandet med græsset. Dermed er simuleringerne med FieldEco i overensstemmelse med, hvad man kan iagttage i naturen. Dog ses i naturen i nogle tilfælde sameksistens imellem agertidsel og kvik, men med parameteriseringen af agertidsel fra dette projekt var det ikke muligt at simulere.
I alle scenarier blev de involverede arter startet fra frø, og planterne fik to år til at etablere sig, før herbicidpåvirkningerne blev introduceret i simuleringerne. Dog blev scenarier med en herbicidbehandling om efteråret startet med en behandling i efteråret år 2. Til simuleringerne anvendtes hvert år klimadata fra 2001 fra DMUs klimastation i Silkeborg. Der blev anvendt gentagelser af samme klima for at undgå sløring af herbicideffekter på grund af variationer i temperatur og globalstråling (sollys).
Det blev valgt, at begrænse simuleringerne til at have cirka samme tidsmæssige udstrækning, som simuleringerne af feltdata fra Kalø, det vil sige 7 år, da der ikke er nogen sikkerhed for pålideligheden af simuleringer over længere tidsrum.
Mælkebøtte i konkurrence med kvik
Det var muligt at simulere sameksistens mellem kvik og mælkebøtte igennem 7 år ved lavt kvælstofniveau, mens kvik udkonkurrerede mælkebøtte ved højere niveauer. Alle scenarier med mælkebøtte blev kørt med den årlige maksimale tørvægt af reproduktive organer (knopstande, blomsterstande, frugtstande) som måleparameter for herbicideffekten. Denne parameter blev valgt, fordi den har med hegnsplantens (i dette tilfælde mælkebøtte) reproduktive output at gøre, og fordi blomstring af hegnsplanter kan anses for en naturkvalitetsmæssig værdi.
Korttidseffekter
Scenarierne viste en lille reduktion i mængden af reproduktive organer ved afdrift af glyphosat fra vinterraps i sædskifte 1, hvor vinterraps forekommer hvert 3. år (figur 3.44 øverst), det vil sige søjlen for sædskifte 1b er lavere end søjlen for sædskifte 1a, hver gang vinterraps forekommer. Denne effekt modvirkes dog i årene med vårbyg og vinterbyg, hvor der ikke er forskel i sprøjtningen imellem sædskifte 1a og 1b. Stigningen i mælkebøttens frugtsætning her skyldes modellens dynamiske allokering af ressourcer. Mælkebøttens frugtsætning er i modellen (og antagelig også i virkeligheden) følsom overfor størrelsen af overvintrende planter. Jo større plante, jo bedre frugtsætning. Hvis der et år er en reduceret frugtsætning, allokerer planten flere ressourcer til vækst af øvrige organer, hvilket betyder, at den overvintrende plante bliver større og derfor har relativt flere ressourcer at sætte ind i frugtsætning det følgende år. Derfor ses en kompensation i årene efter den reducerede frugtsætning på grund af glyphosat, en kompensation, der giver en svagt bedre frugtsætning ved anvendelse af glyphosatresistent raps end ved konventionel raps. Den negative effekt af glyphosat ved vinterrapsdyrkningen skyldes overvejende, at den reducerer mælkebøttens vækst i efteråret, hvorved den overvintrende plante bliver mindre. Denne negative effekt modsvarer fordelen af, at kvik rammes hårdere end mælkebøtte af glyphosat. Ved scenarier (ikke vist) med glyphosatsprøjtninger sidst på foråret er effekten omvendt, da mælkebøttens konkurrencefordel opvejer sprøjteeffekten.
Der er også i sædskifte 2 og 3 små korttidseffekter af sprøjtescenarierne med svag afdrift med en svagt forbedret blomstring ved brug af glyphosat i majs i stedet for MaisTer, hvilket skyldes den forbedrede konkurrencesituation for mælkebøtten efter sprøjtningen.
I sædskifte 4 er der en forbedret blomstring ved scenariet med glyphosat sammenlignet med konventionel sprøjtning, hver gang roer forekommer, men der kompenseres for denne effekt allerede det følgende år, hvor forholdet er omvendt, dog ikke ved den kraftigste afdrift på 25%, hvor mælkebøtte uddør i det konventionelle sprøjtemønster.
Langtidseffekter
Der ses ingen klare langtidseffekter af ændret sprøjtemønster pga. glyphosatresistente afgrøder i sædskifte 1, 2 og 3, men i sædskifte 4 er der en klar langtidsfordel af glyphosatanvendelsen ved høj afdriftsrate, idet mælkebøtten uddør ved det konventionelle scenarium. Dette skyldes den meget intensive sprøjtning, der foregår i forbindelse med roedyrkningen.
Vild kørvel i konkurrence med kvik
Det var muligt at simulere sameksistens af vild kørvel og kvik ved lave og medium koncentrationer af kvælstof. Ved høje koncentrationer af kvælstof blev kvik udkonkurreret på grund af skyggevirkningen fra den meget højere vild kørvel. Ved vild kørvel var der i simuleringerne tale om en plantepopulation under opbygning, da der fortsat var en stigning i mængden af reproduktive organer (figur 3.45) i de første år ved alle scenarier. Der var ligeledes en stigning i total biomasse med tiden i simuleringerne (ikke vist).
Korttidseffekter
Der er i vinterraps en svag tendens til en større produktion af reproduktiv biomasse hos vild kørvel ved sædskifte 1 med sprøjtescenariet ved anvendelse af glyphosatresistent raps end ved det konventionelle sprøjtescenarium. Effekten er dog ikke så stærk, at den holder sig ved de efterfølgende afgrøder ved alle afdriftskoncentrationer. Ved sædskifte 2 er der ligeledes en svagt positiv effekt på vild kørvel af scenarierne med glyphosat i sammenligning med scenarierne med MaisTer ved afdriftsniveauer på 5% og 25%. Ved 1% afdrift var effekten ikke stærk nok til at slå synligt igennem på figur 3.45.
Langtidseffekter
Der er en positiv langtidseffekt af den tidlige glyphosatsprøjtning (scenarium 3c), idet der er en klart højere maksimal reproduktiv biomasse i de sidste 4 år ved dette scenarium ved 25% afdrift end ved de øvrige scenarier i sædskifte 3. Denne tendens gør sig også gældende ved scenariet med 5% afdrift. En lignende tendens kunne forventes at slå igennem i sædskifte 2, men her bevirker den konventionelle sprøjtning i vårbyg hvert andet år en udjævning af effekten, som derfor ikke slår permanent igennem.
Stor nælde
Det var ikke muligt at simulere sameksistens imellem kvik og stor nælde, idet stor nælde i alle tilfælde konkurrerede kvik ud. Dette støttes af observationer fra naturen, hvor stor nælde er kendt for at være tæt bestanddannende. Ved præsentation af resultaterne fra scenarierne med stor nælde (figur 3.46) er den maksimale skudbiomasse præsenteret. Dette er blevet valgt, fordi skudbiomassen på grund af dens funktion som føde for adskillige almindelige sommerfuglearters larver synes at have større forvaltningsmæssig interesse end frøproduktionen.
Korttidseffekter
Der er en markant negativ korttidseffekt af glyphosat ved roedyrkningen hvert fjerde år i sædskifte 4. Effekten udjævnes allerede det følgende år. Der er også en mindre, negativ korttidseffekt af glyphosat i vinterrapsen i sædskifte 1, men kun ved 5 og 25% afdriftsniveauer.
Langtidseffekter
Ligesom for vild kørvel er der en klar tendens til, at den tidlige glyphosatsprøjtning er mindre skadelig end de øvrige sprøjtescenarier i sædskifte 3. Det er måske ikke så overraskende, at afdriften fra én sprøjtning er mindre skadelig end den halve afdrift fra 2 sprøjtninger, der derved påvirker planterne i længere tid. Men det er overraskende, at der er så stor forskel på den tidlige og den sene glyphosatsprøjtning. Denne forskel skyldes, at giftvirkningen i modellen holder i kortere tid i den tidlige sprøjtning end i den sene sprøjtning set på en graddagsskala. Planternes vækst simuleres på en graddagsskala med 5ºC som nulpunkt (ºD5), og giftens virkning antages i modellen af vare i 21 dage. På 21 dage efter den tidlige sprøjtning forløber der 166ºD5 og på 21 dage efter den sene sprøjtning forløber der 255ºD5. Dette skyldes, at der er en højere gennemsnitstemperatur i denne periode end efter den tidlige sprøjtning. Derved simuleres giftvirkningen til at være mere langvarig ved højere temperatur end ved lav målt på den graddagsskala, som planternes vækst simuleres efter. Hvorvidt det er korrekt at gøre sådan afhænger af, på hvilken tidsskala varigheden af glyphosats giftvirkning skal ses? Hvis nedbrydningen bedst beskrives ved hjælp af timer og dage, er simuleringsresultatet med en forskel imellem scenarium 3c og 3d i overensstemmelse med virkeligheden, men hvis nedbrydningen bedst beskrives på samme graddagsskala som planternes vækst, er forskellen et modelartefakt.
Agertidsel
Det var ikke muligt at simulere sameksistens imellem agertidsel og kvik i FieldEco med mindre kvælstofniveauet blev sat ekstremt lavt. Ved realistiske kvælstofniveauer udkonkurrerede agertidslen kvik i løbet af to eller tre år. Dette skyldes måske, at den tidselgruppe (måske en klon), der blev benyttet til parameterisering var meget kraftig og tæt bestandsdannende. Da der i naturen kan findes sameksisterende bestande af tidsler og kvikgræs, er der muligvis forskel på forskellige tidselkloners konkurrenceevne.
I scenarierne blev det besluttet at holde fast ved den parameterisering af tidsel, som var blevet gennemført, da der ikke fandtes oplysninger i litteraturen om, hvordan parameteriseringen skulle være for at kunne simulere sameksistens. Resultaterne i scenarierne er derfor baseret på effekterne af herbiciderne på agertidsel voksende uden interspecifik konkurrence (fig. 3.47).
Korttidseffekter
Der er korttidseffekter på blomstringen af agertidsel i sædskifte 2, hvor effekterne af sprøjtescenarierne med glyphosat er mindre skadelig end sprøjtescenariet med MaisTer, idet der er et forøget maksimum af reproduktive organer ved scenarierne med glyphosat i 5% og især i 25% afdriftsscenariet. En markant korttidseffekt findes i sædskifte 4, hvor sprøjtningen med glyphosat i forbindelse med roedyrkningen har en negativ effekt på tidslernes reproduktion; effekter er dog overvundet allerede det følgende år i forbindelse med vårbyggen.
Langtidseffekter
Det eneste sted, hvor der forekommer langtidseffekter på tidslernes produktion af reproduktive organer, er i sædskifte 3 med permanent majs. I dette sædskifte slår forskellen mellem scenarierne med MaisTer og glyphosat fra sædskifte 2 permanent igennem. De tilsvarende effekter i sædskifte 2, hvor majsdyrkningen kun forekommer hvert andet år, forsvinder i år uden majs.
Rejnfan
Det var ligesom for agertidsel og stor nælde ikke muligt at simulere sameksistens mellem rejnfan og kvik. Også i dette tilfælde blev kvik udkonkurreret af en tæt bestanddannende art, hvilket er i overensstemmelse med observationer fra naturen, hvor rejnfan som regel forekommer i tætte bestande.
Korttidseffekter
Der var ingen klare korttidseffekter af sprøjtescenarier med glyphosat på rejnfan.
Langtidseffekter
Der var en langtidseffekt i form af et generelt lavere frugtsætningsniveau ved 25% afdrift i alle sædskifter (fig. 3.48). Det vil sige, at pesticidet påvirker frugtsætningen, men at der generelt ikke er de store forskelle imellem sprøjtescenarierne inden for sædskifterne.
Figur 3.44.
Sammenligning imellem den årlige maksimale frugtsætning hos almindelig mælkebøtte i konkurrence med kvik ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.45.
Sammenligning imellem den årlige maksimale frugtsætning hos vild kørvel i konkurrence med kvikgræs ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.46.
Sammenligning imellem den årlige maksimale frugtsætning hos stor nælde ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.47.
Sammenligning imellem den årlige maksimale frugtsætning hos agertidsel ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
Figur 3.48. Sammenligning imellem den årlige maksimale frugtsætning hos rejnfan ved anvendelse af glyphosatresistente og konventionelle afgrøder i 4 sædskifter med 1%, 5% og 25% afdrift af markdosis.
3.2.5.4 Konklusion på simuleringer
Der blev i simuleringerne fundet små korttidsforskelle på de flerårige tokimbladede hegnsplanter i sædskifter med majs ved anvendelse af glyphosat i stedet for MaisTer, hvilket overvejende skyldes relativt svagere sensitivitet overfor glyphosat hos disse arter. I sædskifter med vinterraps og roer er billedet ikke så klart, da der blev fundet mindre skadelige effekter af glyphosat end af konventionel sprøjtning ved nogle kombinationer af arter og sædskifter og mere skadelige effekter ved andre kombinationer.
Hovedparten af de fundne forskelle imellem effekter af glyphosat i forhold til konventionel behandling var dog af kortvarig karakter og forsvandt igen i det følgende år, hvor der i tre af sædskifterne forekommer en kornafgrøde med konventionelt sprøjtemønster i alle scenarier. Simuleringerne peger kun på langtidsforskelle imellem effekterne af sprøjtningerne ved dyrkning af konventionelle afgrøder og glyphosatresistente afgrøder i forbindelse med sædskiftet med permanent majs, hvor der ikke er en kornafgrøde i sædskiftet til at rette forskellen op.
Der blev ikke fundet særlig markante forskelle imellem konventionel sprøjtning og glyphosatsprøjtninger ved 1% og 5% afdrift, men ved 25% begyndte forskellene at slå klart igennem. Det blev ikke undersøgt ved simulering, hvorledes planternes vækst ville have været helt uden herbicidpåvirkning.
Version 1.0 November 2008, © Miljøstyrelsen.