Forurening og formering af mitter i Egelřkke Lung
3 Resultater
3.1 Faunasammensćtning
3.2 Klimaparametre i undersřgelsesperioden
3.3 Larveundersřgelser
3.3.1 Tćtheden af mittelarver i sedimentet
3.3.2 Střrrelsesvariationen af mittelarver
3.4 Registrering med klćkkefćlder
3.5 Registrering med lysfćlde
3.5.1 Aktivitetsfordeling af totale antal arter
3.5.2 Dominantarternes sćsonmćssige fordelingsmřnster
3.6 Resultater af simuleringerne
3.1 Faunasammensćtning
28 mittearter pĺvist – 3 nye for Danniark
I lřbet af undersřgelsesperioden 1996 indsamledes 586 mitter (308 ![]()
![]() , 278
, 278 ![]()
![]() ) i klćkkefćlder og 16393 (1594
) i klćkkefćlder og 16393 (1594 ![]()
![]() , 14799
, 14799 ![]()
![]() ) i lysfćlden (Tabel 3.1). lalt blev 28 arter pĺvist, hvoraf 3 ikke tidligere er fundet i Danmark eller det řvrige Norden (C. submaritimus, C. brunnicans og C. cameroni). Fjorten arter var reprćsenteret i klćkkefćlderne, 24 i lysfćlderne; 4 arter (C. albicans, C. poperinghensis, C. riethi, C. stigma) er kun registreret i klćkkefćlderne og 14 arter kun i lysfćlden (C. submaritimus, C. maritimus, C. chiopterus, C. segnis; C. grisescens, C. scoticus, C. vexans, C. achrayi, C. halophilus, C. brunnicans, C. dewulfi, C, cameroni, C. pulicaris sitosinervis, C. machardyi). Ti arter (C. punctatus, C. duddingstoni, C. obsoletus, C. pictipennis, C. circumscriptus, C. odibilis, C. subfascipennis, C. cubitalis, C. pulicaris, C. nubeculosus) var til stede i fangsterne fra begge fćldetyper.
) i lysfćlden (Tabel 3.1). lalt blev 28 arter pĺvist, hvoraf 3 ikke tidligere er fundet i Danmark eller det řvrige Norden (C. submaritimus, C. brunnicans og C. cameroni). Fjorten arter var reprćsenteret i klćkkefćlderne, 24 i lysfćlderne; 4 arter (C. albicans, C. poperinghensis, C. riethi, C. stigma) er kun registreret i klćkkefćlderne og 14 arter kun i lysfćlden (C. submaritimus, C. maritimus, C. chiopterus, C. segnis; C. grisescens, C. scoticus, C. vexans, C. achrayi, C. halophilus, C. brunnicans, C. dewulfi, C, cameroni, C. pulicaris sitosinervis, C. machardyi). Ti arter (C. punctatus, C. duddingstoni, C. obsoletus, C. pictipennis, C. circumscriptus, C. odibilis, C. subfascipennis, C. cubitalis, C. pulicaris, C. nubeculosus) var til stede i fangsterne fra begge fćldetyper.
Tabel 3.1
Mittearter (Culicoides) registreret i klćkkefćlde og lys/sugefćlde, Egelřkke Lung, Langeland, 1996.
Biting midges (Culicoides) recorded in emergence traps and a light trap, Egelřkke Lung, Langeland, 1996.
|
Art/species |
Klćkkefćlder/emergence traps |
Lysfćlde/light trap | ||||
|
|
|
|
total |
|
|
total |
|
C. punctatus |
17 |
15 |
32 |
1028 |
9070 |
10098 |
|
C. duddingstoni |
46 |
52 |
98 |
195 |
2437 |
2632 |
|
C. obsoletus |
1 |
0 |
1 |
110 |
966 |
1076 |
|
C. pictipennis |
56 |
42 |
98 |
95 |
646 |
741 |
|
C. circumscriptus |
5 |
0 |
5 |
48 |
461 |
509 |
|
C. submaritimus |
0 |
0 |
0 |
0 |
488 |
488 |
|
C. naritimus |
0 |
0 |
0 |
78 |
178 |
256 |
|
C. chiopterus |
0 |
0 |
0 |
13 |
131 |
144 |
|
C. odibilis |
22 |
24 |
46 |
11 |
124 |
135 |
|
C. segnis |
0 |
0 |
0 |
5 |
111 |
116 |
|
C. subfascipennis |
0 |
5 |
5 |
5 |
105 |
110 |
|
C. cubitalis |
21 |
27 |
48 |
2 |
56 |
58 |
|
C. grisescens |
0 |
0 |
0 |
2 |
0 |
2 |
|
C. scoticus |
0 |
0 |
0 |
0 |
9 |
9 |
|
C. vexans |
0 |
0 |
0 |
0 |
4 |
4 |
|
C. pulicaris |
0 |
1 |
1 |
0 |
2 |
2 |
|
C. nubeculosus |
5 |
3 |
8 |
0 |
2 |
2 |
|
C. achrayi |
0 |
0 |
0 |
0 |
2 |
2 |
|
C. halophilus |
0 |
0 |
0 |
0 |
1 |
1 |
|
C. brunnicans |
0 |
0 |
0 |
0 |
2 |
2 |
|
C. dewulfi |
0 |
0 |
0 |
2 |
1 |
3 |
|
C. cameroni |
0 |
0 |
0 |
0 |
1 |
1 |
|
C. pulicaris sitosinervis |
0 |
0 |
0 |
0 |
1 |
1 |
|
C. machardyi |
0 |
0 |
0 |
0 |
1 |
1 |
|
C. albicans |
0 |
3 |
3 |
0 |
0 |
0 |
|
C. poperinghensis |
0 |
1 |
1 |
0 |
0 |
0 |
|
C. riethi |
134 |
105 |
239 |
0 |
0 |
0 |
|
C. stigma |
1 |
0 |
1 |
0 |
0 |
0 |
|
|
308 |
278 |
586 |
1594 |
14799 |
16393 |
Nogle fĺ arter er dominerende
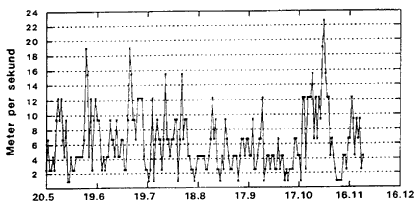
I klćkkefćldematerialet dominerede C. riethi (40.8%), C. duddingstoni (16.7%) og C. pictipennis (16.7%), i lysfćlden C. punctatus (61.7%), C. duddingstoni (16.1%) og C. obsoletus (6.6%); sammen med C. pictipennis (4.5%), C. circumscriptus (3.1%) og C. submaritimus (3.1%) udgjorde de nćvnte arter ca. 95% af det samlede lysfćldemateriale. 10°C er en kritisk grćnsevćrdi Fig. 3.1 og 3.2 samt Appendiks 2-5 viser variationer i klimaet 1996 (Station: Rudkřbing). Appendiks 2 og 3 prćsenterer henholdsvis maksimum- og minimumtemperaturer i de enkelte dřgn gennem sćsonen. Maksimum- og minimumkurverne fřlger i store trćk hinanden gennem hele sćsonen. Larveudviklingen er betinget af gennemsnitstemperaturer i de enkelte dřgn i lřbet af sćsonen; hřje temperaturer vil accelerere udviklingshastigheden i alle stadier inklusiv ćgstadiet. Minimumtemperaturen, der er mĺlt i aften- og natperioderne, hvor de voksne mitter er aktive, er af betydning for flyveaktiviteten. Selv smĺ ćndringer i temperaturen kan have stor betydning for den aktuelle flyveaktivitet, dette gćlder specielt nćr en kritisk lav grćnsevćrdi, der tidligere er mĺlt til ca. 10şC for de fleste danske mittearter (Nielsen, 1982). Flyveaktiviteten afhćnger af vindhastigheden Fig. 3.1 og 3.2 angiver henholdsvis maksimum- og minimum vindhastighed i de enkelte dřgn gennem sćsonen. Her er specielt vindhastigheden i aften- og natperioden, af betydning. Mitternes aktivitetsperiode (primo juni til medio november) er prćget af ret stabile vindforhold med vindhastigheder < 4 m/sek. dog afbrudt af perioder af nogle fĺ dages varighed, hvor vindhastigheden stiger til ca. 12 m/sek. Fra medio oktober kan der registreres en lćngerevarende periode med betydelig vindpĺvirkning (> 4 m/sek.) helt op til 16 m/sek. Appendiks 4 og 5 viser udviklingen af relativ luftfugtighed (RH) gennem sćsonen. Her er specielt de maksimale vćrdier af betydning, da de angiver luftfugtighedsforholdene i mitternes aktivitetsperiode i lřbet af dřgnet (specielt aften- og nataktivitet). Der kan registreres et fald i RH fra primo maj til primo august fra gennemsnitlige vćrdier pĺ 95% til 85%. Dette niveau holdes frem til medio oktober, hvorefter den relative luftfugtighed igen stiger til gennemsnitsvćrdier pĺ ca. 95%. Fig. 3.1
Daily maximum wind speed (m/sec), Rudkřbing, Langeland 1996. Fig. 3.2
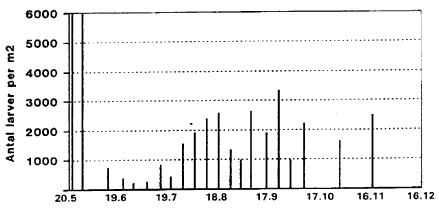
Daily minimum wind speed (m/sec), Rudkřbing, Langeland 1996. Op til 13. 000 larver Fig. 3.3 viser fordelingen af larver gennem sćsonen prćsenteret som et gennemsnit Fig. 3.3
Average number of larvae of biting midges per m2, Egelřkke Lung, 1996. Note: On 24.5 density of larvae is 13000 per m2. Fig. 3.4
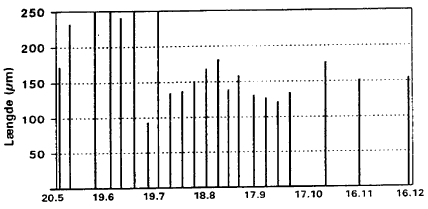
Average number of biting midges Hřje larvetćtheder Fig. 3.4 viser fordelingen af larver frem til starten pĺ nćste aktivitetssćson (13. november 1996 til 20. juni 1997). Fra 5. maj 1997 er det gennemsnitlige antal larver i mosens sediment meget hřjt (ca. 10.000 individer Hovedkapselmĺl afslřrer larvens alder Fig. 3.5 viser střrrelsesfordelingen af larver pĺ forskellige tidspunkter gennem aktivitetsperioden. Střrrelse, der her angives som hovedkapsellćngde, er samtidig et mĺl for larvealder (stadium). De aktuelle gennemsnit pĺ de forskellige indsamlingstidspunkter afspejler sĺledes spredning i alder, men ogsĺ (i mindre grad) forskellig artssammensćtning. Fra indsamlingsperiodens start (24. maj) til 11. juni stiger den gennemsnitlige hovedkapsellćngde (> 200 m) signifikant (P <0.05 KW) og frem til 26. juni er hovedkapselmĺlene af tilsvarende střrrelse. Dette viser, at en meget stor del af de indsamlede larver er i 4. larvestadium. I denne periode er larvetćthederne dog meget lave, jfr. fig. 3.3. Efter medio juli falder den gennemsnitlige hovedkapsellćngde til ca. 140 m, hvilket afspejler, at et střrre antal af larverne befinder sig i tidlige stadier. Fra medio juli til medio august ses en signifikant stigning (P < 0.05 KW) i hovedkapselmĺlene, hvilket indicerer larval vćkst frem mod sidste (4.) larvestadium. Efter dette tidspunkt ses igen et signifikant fald til primo oktober. Dette kan afspejle, at en střrre del af larverne forpupper sig, at voksne mitter klćkkes, samt at en ny reproduktionscyklus resulterer i en tilgang af nyklćkkede larver. Herefter er der igen en stigning frem til ultimo oktober, der kan tolkes som vćkst i sidste larvegeneration. Fig. 3.5
Average length of larval head capsules of biting midges, Egelřkke Lung 1996.
Ingen larvevćkst i vintermĺnederne Resten af sćsonen 1996 frem til 20. december kan der ikke mĺles signifikant ćndring i hovedkapselmĺlene, hvilket kan indicere en temperaturbetinget stagnation af larvevćkst, samt at der i denne periode ikke sker tilgang af nyklćkkede larver. En artsbestemmelse af larvematerialet vil give et endnu klarere billede af larvevćksten gennem sćsonen. Tćtheden af klćkkede mitter Tabel 3.2 angiver artssammensćtning og individantal af voksne mitter registreret med klćkkefćlder pĺ de forskellige stationer, samt det totale antal larver indsamlet pĺ disse. Pĺ de fleste stationer er larveantallet nogenlunde ensartet, men station 6 og 8 adskiller sig fra de řvrige ved henholdsvis et meget stort og et meget lille antal larver. Fra station 7, hvor klćkkefćlderne mĺtte fjernes, foreligger kun larvedata (936 larver indsamlet). Da antallet af klćkkefćlder pĺ stationerne er det samme, kan tćtheden af klćkkede individer Individtćthed afflćnger af sedimenttype Stationerne med relativt hřje vćrdier ligger spredt i řstlige, midterste og sydlige bassin; et fćlles trćk synes at vćre forekomst af finkornet sediment (jćvnfřr 2.2.2). Pĺ de resterende 3 stationer er individtćthed og artsantal meget lavere end pĺ ovennćvnte stationer: Station 2 (33.4 Artssammensćtningen forskellig pĺ stationerne Sammensćtningen af mittefaunaen klćkket fra de individ- og artsrige stationer (1, 3, 4 og 6) og fra de individ- og artsfattige stationer (2, 5 og 8) er ikke fundamentalt forskellig, men blot noget forarmet pĺ sidstnćvnte (Tabel 3.2), idet der ialt er fundet henholdsvis 12 og 8 arter pĺ de to typer af stationer. Pĺ de individ- og artsrige stationer varierer arternes dominansforhold betydeligt: Pĺ station 1 og 3 er C. riethi klart dominerende efterfulgt af henholdsvis C. odibilis og C. duddingstoni, pĺ station 4 forekommer ikke mindre end 4 arter i antal (C. cubitalis, C. pictipennis, C. duddingstoni og C. odibilis) og pĺ station 6 er C. pictipennis og C. duddingstoni dominantarter. Pĺ de individ- og artsfattige stationer (station 2, 5 og 8) er C. punctatus og C. duddingstoni gennemgĺende dominerende. De klćkkede mittearters rumlige fordeling i mosen er meget forskellig (Tabel 3.2). C. duddingstoni og C. punctatus er řjensynligt vidt udbredt i bassinerne, idet de er registreret pĺ henholdsvis 7 og 6 af de undersřgte stationer, men ogsĺ C. cubitalis, C odibilis og C. pictipennis, der er fundet pĺ 5 stationer, synes meget udbredte. De nćvnte arters kvantitative forekomst pĺ stationerne varierer betydeligt, f.eks. er C. cubitalis kun talrig pĺ station 4, mens de řvrige arter forekommer i antal pĺ flere af stationerne. De resterende arter er registreret pĺ 1-3 stationer; blandt disse mĺ C. riethi, der er fundet pĺ 3 stationer – og i stort antal pĺ 2 – fremhćves, idet den udgřr > 40% af det samlede antal klćkkede mitter. Arterne Calbicans, C. circumscriptus, C. nubeculosus, C. obsoletus, C. poperinghensis, C. pulicaris, C. stigma og C. subfascipennis registreres kun spredt og i ringe antal i klćkkefćlderne. Tabel 3.2
Number of adult biting midges collected in emergence traps in 7 sites, Egelřkke Lung, 1996. Number of larvae collected throughout the season presented.
3.2 Klimaparametre i undersřgelsesperioden
Daglig maksimum vindhastighed (m/sek), Rudkřbing, Langeland 1996.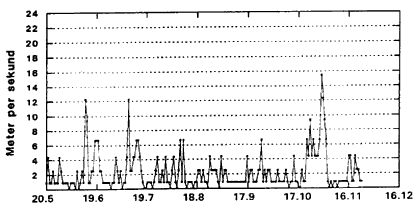
Daglig minimum vindhastighed (m/sek), Rudkřbing, Langeland 1996.3.3 Larveundersřgelser
3.3.1 Tćtheden af mittelarver i sedimentet
Gennemsnitlige antal mittelarver 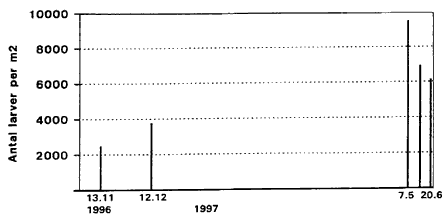
Gennemsnitlige antal mittelarver 3.3.2 Střrrelsesvariationen af mittelarver
Gennemsnitslćngde af mittelarvernes hovedkapsel, Egelřkke Lung 1996.3.4 Registrering med klćkkefćlder
Antal voksne mitter indsamlet i klćkkefćlder pĺ 7 stationer, Egelřkke Lung, 1996. Antallet af larver indsamlet gennem sćsonen angivet.
|
Art/species |
St. 1 |
St. 2 |
St. 3 |
St. 4 |
St. 5 |
St. 6 |
St. 8 |
Total |
|
C. albicans |
||||||||
|
|
1 |
0 |
0 |
0 |
0 |
2 |
0 |
3 |
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
C. circumscriptus |
||||||||
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
1 |
0 |
2 |
0 |
2 |
0 |
0 |
5 |
|
C. cubitalis |
||||||||
|
|
7 |
1 |
1 |
16 |
0 |
2 |
0 |
27 |
|
|
2 |
0 |
1 |
18 |
0 |
0 |
0 |
21 |
|
C. duddingstoni |
||||||||
|
|
0 |
3 |
9 |
7 |
2 |
29 |
2 |
52 |
|
|
1 |
0 |
12 |
12 |
3 |
18 |
0 |
46 |
|
C. nubeculosus |
||||||||
|
|
0 |
0 |
3 |
0 |
0 |
0 |
0 |
3 |
|
|
2 |
0 |
2 |
1 |
0 |
0 |
0 |
5 |
|
C. obsoletus |
||||||||
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
|
C.odibilis |
||||||||
|
|
9 |
1 |
3 |
8 |
0 |
3 |
0 |
24 |
|
|
7 |
1 |
6 |
5 |
0 |
3 |
0 |
22 |
|
C. pictipennis |
||||||||
|
|
1 |
0 |
6 |
7 |
0 |
28 |
0 |
42 |
|
|
0 |
0 |
5 |
14 |
0 |
35 |
2 |
56 |
|
C. poperinghensis |
||||||||
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
0 |
1 |
0 |
0 |
0 |
0 |
0 |
1 |
|
C. pulicaris |
||||||||
|
|
0 |
0 |
0 |
0 |
1 |
0 |
0 |
1 |
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
C. punctatus |
||||||||
|
|
0 |
0 |
3 |
4 |
3 |
3 |
2 |
15 |
|
|
0 |
6 |
0 |
6 |
2 |
3 |
0 |
17 |
|
C. riethi |
||||||||
|
|
53 |
0 |
50 |
2 |
0 |
0 |
0 |
105 |
|
|
64 |
0 |
69 |
1 |
0 |
0 |
0 |
134 |
|
C. stigma |
||||||||
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
0 |
0 |
1 |
0 |
0 |
0 |
0 |
1 |
|
C. subfascipennis |
||||||||
|
|
0 |
0 |
0 |
0 |
0 |
2 |
3 |
5 |
|
|
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Total |
148 |
12 |
173 |
102 |
13 |
128 |
9 |
586 |
|
Larver/larver |
795 |
722 |
623 |
851 |
738 |
1148 |
223 |
Tidlige og sene arter
Dominantarterne har klćkningsmaksima pĺ forskellige tidspunkter i lřbet af sćsonen (Appendiks 6 til 11): C. riethi og C. pictipennis har en aktivitetstop tidligt pĺ sćsonen med toppunkt medio juni, C. odibilis har en mindre klćkningstop medio juni og en mere markant omkring medio juli, mens C. duddingstoni, C. punctatus og C. cubitalis har relativt sene klćkningstoppe henholdsvis 25. juli, 1. august og 8. august.
3.5 Registrering med lysfćlde
3.5.1 Aktivitetsfordeling af totale antal arter
De fřrste mitter registreres primo juni
Det totale antal mitteregistreringer (alle arter) gennem sćsonen viser betydelige udsving i sommerens lřb (Fig. 3.6). De fřrste registreringer opnĺs primo juni, men frem til primo juli registreres mitterne kun i beskedent antal (<100 individer per dřgn). Herefter ses en markant registreringstop pĺ op til 600 individer. I en periode frem til 22. juli er fangsttallene atter lave, hvorefter en ny markant aktivitetsperiode med fangsttal pĺ op til 600 individer strćkker sig frem til den 8. august. Efter en ca. 7 dages periode med forholdsvis lave registreringstal indtrćder en markant aktivitetsperiode med fangster pĺ > 800 individer per dřgn, der strćkker sig frem til ca. 1. september. Efter en kortvarig periode med fangster pĺ <200 individer per dřgn frem til ca. 10. september stiger aktiviteten igen omkring midten af september med toppe pĺ op til 600 individer per dřgn. Aktiviteten er derefter meget lav i slutningen af september og i fřrste uge af oktober, men omkring midten af oktober indtrćder ĺrets sidste aktivitetsperiode med fangster pĺ op til 200 individer per dřgn.
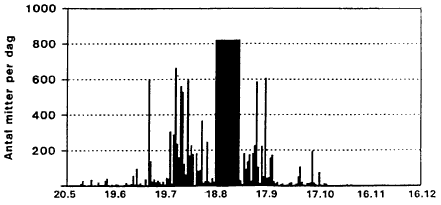
Fig. 3.6
Daglig fangst af mitter i lysfćlde, Egelřkke Lung, 1996.
Daily catch of biting midges in a light trap, Egelřkke Lung, 1996.
Mitteaktivitet i 5 mĺneder
Mitteaktiviteten i omrĺdet strćkker sig sĺledes over nćsten 5 mĺneder formodentlig med 5 aktivitetsmaksima. Relateres mitternes flyveaktivitet til fremherskende vindpĺvirkning ses det tydeligt, at vindstyrken er af afgřrende betydning for flyveaktiviteten. Dette bekrćfter, at mitterne er sćrdeles fřlsomme overfor selv smĺ vindpĺvirkninger. Det kan sĺledes demonstreres, at i perioder, hvor der er tale om kraftig vindpĺvirkning, er mitternes flyveaktivitet sćrdeles ringe ofte lig nul (sml. Fig. 3.1 og 3.2). Derimod spiller lufttemperatur og relativ luftfugtighed tilsyneladende ingen střrre rolle for mitternes flyveaktivitet (Appendiks 2-5).
3.5.2 Dominantarternes sćsonmćssige fordelingsmřnster
Arterne har 1-4 ĺrlige aktivitetstoppe
For at forstĺ mittefaunaens sćsondynamik prćsenteret i det foregĺende, er en nćrmere analyse af dominantarternes (jćvnfřr 3.1 og Tabel 3.1) sćsonmćssige fordeling nřdvendig. Endvidere mĺ lysfćldematerialet af disse arter opdeles efter křn og hunnerne yderligere efter fysiologiske stadier (blodsugningsaktive hunner, det vil sige hunner, der sřger efter et blodmĺltid fřr ćglćgning – nullipare/pare individer samt gravide hunner).
C. punctatus (Fig. 3.7-3.9). De blodsugningsaktive hunner (Fig. 3.7) har 5 aktivitetstoppe: Omkring 1. juli, 25. juli, 22. august, 20. september og – mindre udtalt – omkring 20. oktober. Gravide hunner (Fig. 3.8) har et markant toppunkt omkring 12. september. Hanner (Fig. 3.9) har aktivitetstoppunkter omkring 1. august, 20. august, 20. september samt 20. oktober.
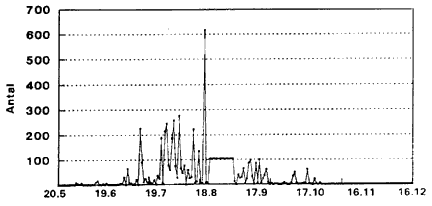
Fig. 3.7
Daglig fangst af C. punctatus ![]()
![]() (blodsugningsaktive) indsamlet i lysfćlde, Egelřkke Lung, 1996.
(blodsugningsaktive) indsamlet i lysfćlde, Egelřkke Lung, 1996.
Daily catch of C. punctatus (nulliparous/parous empty) in a light trap, Egelřkke Lung, 1996.
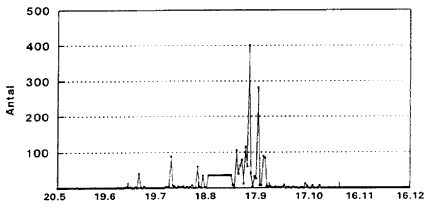
Fig. 3.8
Daglig fangst af C. punctatus ![]()
![]() (gravide) indsamlet i lysfćlde, Egelřkke Lung, 1996.
(gravide) indsamlet i lysfćlde, Egelřkke Lung, 1996.
Daily catch of C. punctatus ![]()
![]() (gravid) in a light trap, Egelřkke Lung, 1996.
(gravid) in a light trap, Egelřkke Lung, 1996.
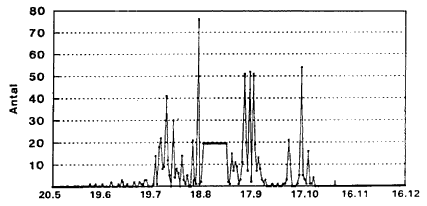
Fig. 3.9
Daglig fangst af C. punctatus ![]()
![]() indsamlet i lysfćlde, Egelřkke Lung, 1996.
indsamlet i lysfćlde, Egelřkke Lung, 1996.
Daily catch of C. punctatus ![]()
![]() collected in a light trap, Egelřkke Lung, 1996.
collected in a light trap, Egelřkke Lung, 1996.
C. duddingstoni (Appendiks 12-14). De blodsugningsaktive hunner (App. 12) har 2 aktivitetstoppe omkring 25. juli og 22. august. Gravide hunner (App. 13) har toppunkter pĺ de samme tidspunkter som de blodsugningsaktive hunner, dog med den mest markante top omkring d. 22. august. Hannernes fordelingsmřnster (App. 14) svarer nřje til fordelingen af gravide hunner.
C. submaritimus (Appendiks 15-17). De blodsugningsaktive hunner (App. 15), gravide hunner (App. 16) samt hanner (App. 17) har en enkelt aktivitetstop omkring 22. juli, men fangsttallene er smĺ.
C. obsoletus (Appendiks 18-20). De blodsugningsaktive hunner (App. 18): Der registreres 2 smĺ aktivitetstoppe i perioden 1. juni til 1. juli samt maksima omkring 1. august, 22. august, 15. september og 15. oktober. Fangsttallene af gravide hunner (App. 19) er smĺ, řjensynlig er der et toppunkt omkring 11. august samt et mindre omkring 10. september. Hanner (App. 20) har et markant aktivitetstoppunkt omkring 15. august og to mindre omkring 12. september og 7. oktober.
C. pictipennis (Appendiks 21-23). De blodsugningsaktive hunner (App. 21) har 3 aktivitetstoppe omkring 7. juni, 2. juli samt 2. august. Gravide hunner (App. 22) har en markant aktivitetsperiode 1.-15. august. Det samme gćlder hanner (App. 23).
3.6 Resultater af simuleringerne
Det daglige antal voksne mitter simuleres
Simuleringerne blev startet 19. marts (hvor temperaturen overstiger de simulerede mitters temperaturnulpunkt) med 20.000 nyklćkkede larver
Sammenfald mellem simulering og observation
Modellen foretager daglige beregninger af antallet af individer i alle stadier. Pĺ Fig. 3.10 ses en sammenligning af det simulerede antal voksne (sum af alle voksenstadier) og fangster af C. punctatus i lysfćlder. Der er et godt sammenfald mellem de simulerede toppe og de observerede flyvetoppe, hvorfor det ser ud til, at de anvendte udviklingstider ikke er langt fra virkeligheden. Der kan dog ikke forventes direkte sammenfald af simulerede forekomster og de observerede data. Dette skyldes, at de observerede data ikke er observationer af populationstćtheder, men af flyveaktivitet. Flyveaktiviteten er ikke blot afhćngig af populationstćtheder, men ogsĺ af temperatur og vind. Derfor kan der under ugunstige vejrforhold godt findes mange mitter, der blot ikke flyver og derfor ikke fanges i lysfćlden.
Hřjderne af de simulerede toppe viser populationstćtheder, hvilket kan anses for et mĺl for det potentiale for flyvende individer, der findes i tilfćlde af ideelt flyvevejr. Vćrdierne i den simulerede kurve er divideret med 10 for at gřre kurven sammenlignelig med de observerede vćrdier. Der skal i fortolkningen af disse simuleringer ikke lćgges vćgt pĺ de prćcise vćrdier af kurven, men blot lćgges mćrke til toppenes placering, indbyrdes afstand og relative hřjde i sammenligning med lysfćldefangsterne.
Dřdelighed og diapause bestemmer toppenes hřjde
Toppenes hřjde er stćrkt afhćngig af dřdeligheden i systemet, og deres relative hřjde er stćrkt afhćngig af diapauseprocenterne i lřbet af sommeren. Dette betyder, at hvis diapauseprocenterne bliver lavere end de her benyttede, bliver forekomsten af mitter betydeligt střrre i sidste halvdel af sommeren (Fig. 3.11).
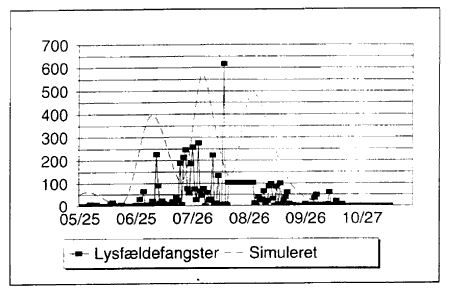
Fig. 3.10
Simuleret kurve over det daglige antal af voksne mitter sammenlignet med fangst af C. punctatus ![]()
![]() (sum af alle fysiologiske stadier) i lysfćlde, Egelřkke Lung, 1996.
(sum af alle fysiologiske stadier) i lysfćlde, Egelřkke Lung, 1996.
Simulation of the daily number of adult biting midges and the catch of C. punctatus ![]()
![]() (all gonotrophic stages) in a light trap, Egelřkke Lung, 1996.
(all gonotrophic stages) in a light trap, Egelřkke Lung, 1996.
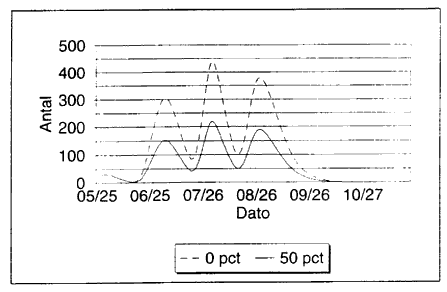
Fig. 3.11
Simulerede kurver over populationsudviklingen hos voksne mitter. (C. punctus). Ved de to simuleringer er benyttet 0% og 50% diapause hos de ćg, der lćgges frem til 23.6. Derefter benyttes samme diapauseprocent.
Simulated population curves of adult biting midges (C. punctatus). In the two simulations 0% and 50% egg diapause, respectively, is assumed for eggs produced until 23.6. Later, the same % egg diapause is used.