|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 87, 2004 Adskillelse af effekter af herbicider og kvælstof på vegetation og leddyr i hegn og græslandsvegetationIndholdsfortegnelse2 Vegetation og leddyr i hegn på økologiske og konventionelle bedrifter 3 Eksperimentel adskillelse af herbicid og N-påvirkning 4 Sammenfattende diskussion og konklusioner
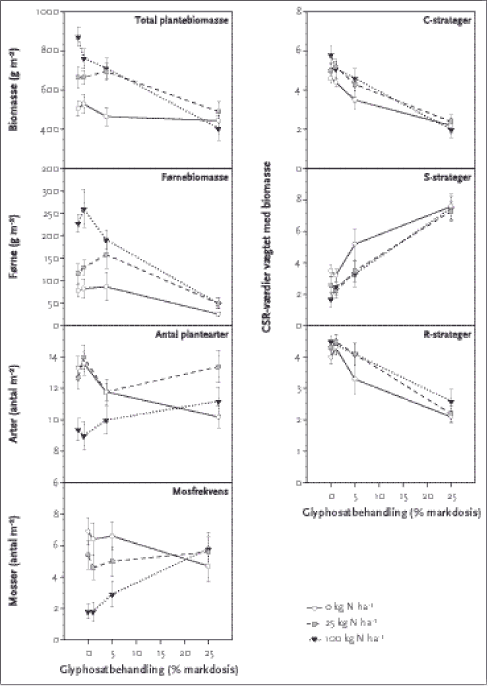
ForordDenne rapport er et resultat af et projekt under Miljøstyrelsens Pesticidforskningsprogram med titlen Adskillelse af effekter af herbicider og ammoniakafdrift på fødekæder i læhegn og engvegetation, som er gennemført i perioden 2001-2004. Projektet blev ansøgt på baggrund af resultater fra andre arbejder under Pesticidforskningprogrammet, pesticidmoniteringsprogram udført af DMU og projekter under Forskningscenter for Økologisk Jordbrug (FØJO). Projektet har været fulgt af en følgegruppe bestående af Kristian Kristensen, DJF, Søren E. Larsen (DMU, indtil 2002), Chr. Kjær, Morten T. Strandberg, Peter Odderskær (fra 2003) DMU, Mogens Erlandsen (Århus Universitet, fra 2003) Niels Lindemark, Dansk Planteværn, Jan Kjær Madsen og Heidi Buur Holbech, Landbrugets Rådgivningscenter, Ditte Olrik, FSL (indtil 2002) , Carsten Suhr Jacobsen, GEUS (2002-03), Allan Prang, GEUS (fra 2003), Peter Esbjerg, KVL (fra 2003), Claus Hansen, MST og Jørn Kirkegaard (Formand), MST. Følgegruppen takkes for engagerede bidrag til rapporten. Projektet har været udført i et meget tæt samarbejde mellem DMU, Afd. for Vildtbiologi og Biodiversitet ved Knud Tybirk (projektleder), Erik Aude, Annie Laursen og studentermedhjælpere og Afd. for Terrestrisk Økologi i Silkeborg ved Marianne Bruus Pedersen og en laborantstab ledet af Inger Møller. Endvidere har der været videnskabeligt samarbejde med Botanisk Institut, KU (Anders Michelsen). Danmarks Jordbrugsforskning, Foulum, har udført kornstørrelsesanalyser. Tak til Landbrugets Rådgivningstjeneste for lån af sprøjteudstyr og til en anonym gårdejer for venlig udlejning af mark til forsøgsareal. Hegnsdelen af projektet har været udført i et samarbejde med Hedeselskabet og Landsforeningen De Danske Plantningforeninger samt en række landmænd, som har stillet deres hegn og viden til rådighed. Disse partnere takkes alle for væsentlige bidrag til rapportens grundlag. SammenfatningDenne projektrapport omhandler 2 års undersøgelser af vegetation og leddyrfauna i løvtræshegn på økologiske og konventionelle landbrug. Endvidere afrapporteres 3 års undersøgelser af effekter af glyphosatafdrift og simuleret ammoniaktilførsel på en eksperimentel vegetation og de tilhørende leddyr. Formålet med projektet var at dokumentere og kvantificere separate og kombinerede effekter af glyphosat og kvælstof på vegetation og leddyrsfauna på en eksperimentel vegetation og relatere dette til de faktiske forhold i marknære biotoper i agerlandet. Undersøgelsen af hegn blev gennemført i 2001 på sandede jorder vest for israndslinien og i 2002 på lerede jorder nord og øst for israndslinien. I alt blev 56 hegn udvalgt ud fra en række fælles kriterier for at få så ensartede hegn som muligt med en ideel alder på 10-15 år og med marker i omdrift på begge sider. Halvdelen af hegnene var på bedrifter, der har været drevet økologisk i mindst 10 år og den anden halvdel på tilsvarende konventionelle. Undersøgelsen på sandjord sammenlignede hegnenes fodpose (lysåben stribe vegetation mellem træer og mark) med felter centralt i hegnet, mens undersøgelsen på lerjord fokuserede på fodposerne. Planternes frekvens blev registreret og leddyr blev samtidigt indsamlet med en sugemaskine i de samme felter. Derudover blev der indsamlet og analyseret jordprøver på såvel fysiske som kemiske karakteristika i alle felter. Den eksperimentelle vegetation blev etableret i 2001 på sandjord efter dybdepløjning og blev sammensat af i alt 30 arter med forskellige funktionelle egenskaber. Arternes forekomst i hegn, spiringsevne og frøtilgængelighed var yderligere kriterier. Der blev etableret 10 gentagelser af 12 unikke behandlinger i felter á 49m², som var alle kombinationer af 0, 25 og 100 kg kvælstof/ha med 0, 1, 5 og 25 % af markdosis af glyphosat. Felterne fik endvidere tilført fosfor, kalium, svovl og kobber. Vegetationen blev registreret og leddyr blev indsamlet årligt i juli/august med samme metoder som i hegnet. Fysiske jordkarakteristika blev indsamlet én gang og kemiske jorddata i år 2001 og 2003. Hegnsundersøgelserne viste, at der var meget klare forskelle i artsdiversiteten af planter på de to driftsformer. Økologiske hegn på sandjord havde 34 % flere arter end tilsvarende konventionelle hegn og på lerjord 50 % flere arter. Forskellene fandtes i den lysåbne fodpose mellem hegn og mark. Der var flere mosser, flere arter der hører hjemme i halvnaturen (eng, mose), flere arter fra marker og ruderater i de økologiske hegn. Der var derimod færre konkurrence-strateger i de økologiske hegn. De økologiske hegn havde en række indikatorarter, hvoraf visse er i generel tilbagegang i agerlandet. Økologiske hegn kan således formentlig være bedre refugier i agerlandet for visse plantearter. De herbivore leddyr fordelte sig generelt efter de samme variable som planterne i hegnene, og der var signifikante forskelle i artssammensætningen i økologiske og konventionelle hegn. Der kunne ikke påvises klare forskelle i tætheden og mangfoldigheden af leddyr mellem de to driftsformer på sandjord. Derimod var der signifikante forskelle mellem prøver taget inde i hegnet og prøver taget i fodposen, idet der var klart flere dyr i fodposen end midt i hegnet. På lerjorden var der dog signifikant flere arter af leddyr i de økologiske hegn (i gennemsnit 240 dyr fordelt på 31 arter pr. prøve af 0,9 m² mod 218 dyr fordelt på 17 arter pr. prøve i konventionelle hegn). Skimmelbillelarver og trips var langt de talrigeste, men også sommerfugle, cikader, visse tægearter, bladbiller, snudebillen Apion flavipes, rovbiller, mariehøns, glimmerbøsser (Meligethes) og voksne skimmelbiller forekom almindeligt. Især for snudebiller var de omgivende markers afgrøde (græsmark – øvrige afgrøder) og dyrkningsform vigtige variable, som ikke er totalt uafhængige, idet de økologiske brug havde flere hegn med græsmark som nabo end på de konventionelle. Den eksperimentelle adskillelse af effekter af kvælstof og glyphosat gav signifikante, interagerende effekter på vegetationens totale biomasse, på førnemængden, på frekvensen af mosser og på artsdiversiteten. Herbicideffekten på biomassen øgedes med øget kvælstofniveau, og effekten af glyphosat viste sig meget forskellig på de udsåede arter. Konkurrence-(C-) og Ruderat- (R-) strateger gik især tilbage med øget N og herbicid, mens Stresstolerante (S-strateger) bedre kunne konkurrere. Fåresvingel (Festuca ovina) viste således tolerans over for glyphosat og dominerede biomassen af nøjsomhedsplanterne. Også mosserne (indvandrede arter) klarede sig tydeligt bedre ved 25 % glyphosat og højt kvælstofniveau end ved lavere doser glyphosat, hvor mosser klarede sig bedst ved lavt kvælstofniveau, fordi der blev plads til at kolonisere, når biomassen var kraftigt reduceret. Desværre var en del af de almindelige hegnsplanter (C- og R-strateger) som kvik, agertidsel, gråbynke og stor nælde ikke særligt veletablerede i den eksperimentelle vegetation. Derfor kan resultaterne ikke direkte overføres til hegnsvegetation. Leddyrenes artsantal steg fra år 2001 til år 2002 og afspejlede vegetationens etablering. Der var dog kun signifikante interaktioner for enkelte grupper. I 2003 var der for frøtæger, blomstertæger, tæger ialt, store løbebiller, små rovbiller, skimmelbiller, antal dyr og antal taxa (arter eller grupper) en signifikant effekt af glyphosatbehandlingen. Der var en signifikant forskel mellem felter tilført forskellig mængde kvælstof for antal dyr i alt, antal arter, antallet af cikader, Scutelleridae-/Pentatomidaetægenymfer, nymfetæger, blomstertæger, små rovbiller, rovbillelarver og skimmelbiller. Antal arter og antal dyr per prøve reduceredes ved den højeste glyphosatdosering, mens felterne, der modtog 100 kg N/ha, havde flere dyr og arter i den usprøjtede situation end de andre kvælstofniveauer. Glyphosatbehandlingen og kvælstoftilførslen havde også en signifikant effekt på artssammensætningen af insekterne og insektarterne fordelte sig mere efter glyphosatgradienten og antal plantearter end efter kvælstofgradienten. Glyphosat kunne alene reducere antallet af plantearter med ca. ¼, og en tilsvarende reduktion ses af kvælstofgødskning. Begge faktorer indvirkede således klart og nogenlunde lige meget rent kvantitativt på diversiteten. Glyphosat har større betydning for plantesamfundets artssammensætning end kvælstof. Glyphosaten alene havde en meget klar effekt på plantebiomassen ved højt gødskningniveau, mens der næsten ingen effekt var ved 0 kg kvælstof. Leddyrene gik tilbage i antal og diversitet ved højt glyphosat-niveau (maks. reduktion hhv. 47 og 26 %). Effekten var specielt tydeligt ved høj kvælstofniveau. Det afspejlede den store ændring af vegetationen under disse forhold og viste leddyrenes afhængighed af vegetationen. Når man sammenfatter de to dele af studiet, når man frem til følgende hovedkonklusioner. Økologiske hegn giver muligheder for, at flere plantearter kan udgøre et alsidigt fødegrundlag for herbivore leddyr og dermed formentlig større diversitet i agerlandets fødekæder. Den interaktive effekt af herbicid og kvælstof fundet i eksperimentet viser, at også gødningstilførsel til hegnene vil påvirke floraen, og at effekten vil være forskellig på økologiske og konven-tionelle bedrifter, selv om gødningsniveauet er det samme. Eksperimentets resultater på vegetations- og leddyrssamfund kan bedst sammenlignes med den forventede udvikling i hegnene de tre første år efter plantning. Hegnsstudiet kan siges at afspejle 'resultatet' efter 10-15 års påvirkninger med (konventionelt landbrug) og uden pesticider (økologisk drift). Studiet viser samlet, at fraværet af pesticider på økologiske bedrifter giver en rigere natur i hegnene. Hvis man yderligere kunne reducere næringsmængden i hegnene kunne vi opnå endnu større mangfoldighed. Sprøjte- og gødningsfrie randzoner eller brak i striber vil kunne forbedre forholdene for naturen i agerlandets småbiotoper. Tilsvarende effekter vil kunne opnåes ved andre tekniske tiltag, som reducerer herbicidafdriften eller gødningstilførslen til hegnene. SummaryThis report contains the results of three years of investigation on the effects of Glyphosate drift and simulated ammonia deposition on experimental vegetation and associated anthropods. Further, the publication reports two years of investigation of vegetation and arthropods in hedges on organic and conventional farms. The aim of the work is to document and quantify separate and combined effects of glyphosate and nitrogen on vegetation and arthropods in small biotopes along fields in the agricultural landscape. The investigation in hedges was carried out in 2001 in the southwestern part of Jutland on sandy Saalian moraines or Weichelian outwash plains and in 2002 on Weichelian moraine loamy soils. A total of 56 hedges were selected, using narrow criteria with an ideal age of 10-15 years and with rotational fields on both sides. Half of the hedges were located on organic farms (minimum 10 years of organic farming period), and the rest on conventional farms. On sandy soils a comparison was made of plots in the centre of the hedges with plots in the open strip of vegetation between the trees and the field (hedge margin). On the loamy soils, the comparison focused on the open strip of vegetation. Frequency of plants was registered and arthropods were collected by vacuum-sucking in the same plots. Soil analysis of physical and chemical characteristics were conducted. The experimental vegetation was established in 2001 on sandy soil after deep ploughing and consisted of 30 species with different ecological attributes. Species abundance in hedges, germination ability and seed availability were additional criteria. Ten replica of 12 unique combinations of 0. 25 and 100 kg N ha-1 yr-1 and 0, 1, 5 and 25 % of field dose of glyphosate in plots of 49m² was established. The plots were fertilised with phosphorus, potassium, sulphur and copper. The vegetation was registered and arthropods were collected yearly with methods resembling the hedge study. Physical soil characteristics were analysed once and chemical characteristics in 2001 and 2003. The hedge studies showed very clear differences in diversity of plants between the two farming practices. Organic hedges had 34 % more plant species than corresponding conventional hedges on sandy soils and 50 % more species on loamy soils. The differences occurred in the hedge margins. More bryophyte species, more species related to semi-natural areas (fens, meadows), more species from fields and ruderal areas were found in the organic hedges. Fewer plant species with a competitive strategy were found in the organic hedges. It was not possible to test for differences between obligatory forest species (hedges are too young) or differences in number of herbicide tolerant species. Many of the indicator species of organic hedges are declining in the agricultural landscape and organic hedges may serve as refuge for certain species. Herbivorous arthropods generally follow plant distribution variables, and species composition differed significantly between organic and conventional hedgerows. Numbers of species per sample, number of animals per sample and Shannon-Wiener diversity index were not affected by farming practice on sandy soils, whereas significant differences were found between internal hedge plots and hedge margin plots, with significantly more arthropods in the hedge margins. On loamy soils, however, more individuals and more species of arthropods were found in organic hedges compared to conventional (average of 240 individuals and 31 species per sample in organic versus 218 individuals and 17 species per sample in conventional hedges). Mould beetle larvae and thrips were the most frequent groups on loamy soils, but also butterflies, cicadae, certain bugs, leaf beetles and adult mould beetles were common. For weevils in particular the crop of associated fields (grass ley versus other crops) had significant impact on abundance. Field crop was an important variable, which was not totally independent of farming practice, as grass ley was more frequent on organic farms. The experiment of separating the effects of glyphosate and nitrogen revealed significant interactions on the biomass of vegetation, litter biomass, frequency of bryophytes and diversity of species. The effects of glyphosate increased by increased nitrogen level, and in general species with a competitive strategy (C) and with ruderal strategy (R) decreased strongly with increased glyphosate level, whereas plants with a Stress-tolerant (S) strategy increased as C and R decreased. Sheep fescue (Festuca ovina) showed remarkable tolerance towards glyphosate and dominated the biomass of the S strategy. Also bryophytes (colonising species) were more frequent at 25 % glyphosate and high nitrogen level compared to lower herbicide doses. With low herbicide doses, the bryophytes had higher frequencies at low N doses. Unfortunately, several common hedge species (C and R strategies) such as couch grass (Agropyron repens), Canada thistle (Circium arvense), mugwort (Artimisia vulgaris) and Common nettle (Urtica dioeca) did not establish very well in the experiment. The plant species react very differently to glyphosate, and therefore the results cannot be compared directly to the conditions in the hedges. The number of arthropods increased from 2001 to 2002, reflecting the establishment of the vegetation. Only in a few groups significant interactions of glyphosate and nitrogen were found. Glyphosate had significant effects on numbers of several groups (several kinds of bugs, large carabid beetles, small rove beetles and mould beetles), numbers of animals and numbers of species in 2003. Significant effects of nitrogen were found for numerous groups (cicadae, several kinds of bugs, small rove beetles, rove beetle larvae and mould beetles) and for numbers of individuals and numbers of species. Numbers of species and individuals per sample were reduced at 25 % glyphosate, whereas plots with 100 kg N had more individuals and species without herbicide than other N levels. Glyphosate and Nitrogen had a significant impact on species composition of arthropods, and the species were distributed more clearly along the herbicide gradient than along the nitrogen gradient. Glyphosate alone could reduce the number of plant species by approximately ¼, and the same reduction could be achieved by nitrogen alone. Both factors influenced significantly and with equal strength on quantitative biodiversity. Glyphosate had a stronger impact on plant community composition as compared to nitrogen. Glyphosate had a very strong impact on plant biomass at high nitrogen level, but hardly any effect on biomass at low nitrogen level. Diversity and quantity of arthropods was significantly reduced at high glyphosate level (47 and 26 %, respectively) – especially at high nitrogen level. This reflected the large change of the vegetation under these circumstances and illustrated the arthropod dependence on the vegetation. When the result of the hedgerow study and the experiment are seen together the following main conclusions are reached. Organic hedges contain more plant species, extending the basis for herbivorous arthropods and adding more diversity to food-web of the agricultural landscape. The interactive effect of herbicide and nitrogen found in the experiment shows that also nitrogen drift into hedgerows affects hedgerow flora, and that the effect will differ between organic and conventional farms, even at similar nutrient levels. The results of the experiment on vegetation and anthropods are comparable to the expected development in planted hedges the first three years after establishment. The hedge studies could reflect the `result' after 10-15 years of influence from conventional agriculture with pesticide drift and organic farming without pesticide drift. The study as a whole showed that absence of pesticides on organic farms gave a richer nature in the hedges. If nutrient levels could additionally be reduced, we expect even higher diversity. Field edge zones free of pesticides and fertilizer may improve the conditions for nature in the small farmland biotopes. Other technological methods reducing herbicide and fertiliser drift into hedgerows will have similar positive effects. 1 Indledning1.1 Baggrund 1.1 BaggrundFor at kunne sikre en afbalanceret forvaltning og understøtte nationale og internationale politiske beslutninger er der et stort behov for viden om naturindholdet i landbrugslandskabet. Gennem de seneste år har folketinget vedtaget fx Pesticidhandlingsplan II og III, Ammoniakhandlingsplan, Wilhjelmudvalgets rapport (Wilhjelmudvalget 2001) og en Biodiversitetshandlingsplan er i skrivende stund i høring. Endvidere vil fremtiden byde på meget store udfordringer for at opfylde Habitatdirektivet og Vandrammedirektivet, hvor landbrugets påvirkninger af internationalt værdsatte naturtyper er meget central. Diskussioner omkring naturkvalitet og biologisk integritet versus traditionelle (arts-) diversitetsmål har bidraget til at nuancere vurderingerne af agerlandets naturindhold (se fx Reddersen m.fl. 1999, Agger m.fl. 1999, Wilhjelmudvalget 2001 og Tybirk 2002). Der er i stigende grad kommet fokus på funktionelle grupper af organismer som bedre udtryk for økosystemernes funktion og kvalitet end blot biodiversitet på artsniveau (Tybirk & Ejrnæs 2001). Højt naturværdi kan findes, hvor der naturligt er få arter, men ofte er få dominerende konkurrencestærke flerårige arter i agerlandets småbiotoper udtryk for en kraftig påvirkning i form af fx herbicid- og ammoniakafdrift. Agerlandets småbiotoper kan potentielt indholde mange arter fra halvnaturarealer (eng, overdrev) og skov. Halvnatur bruges i denne rapport om lysåbne naturtyper der er afhængig af menneskers påvirkning. Flere studier viser, at vi i Danmark har et meget fragmenteret og dynamisk landskab (Agger & Brandt 1988). For at sikre en fremtidig overlevelse af arterne i de isolerede natyrtyper er det nødvendigt, at der kan ske en udveksling af arter via agerlandets småbiotoper. Derfor er det vitigt, at sikre gode betingelser for arterne – også i småbiotoperne. Øget gødnings- og herbicidanvendelse er to væsentlige faktorer, der har påvirket naturindholdet i landskabet gennem de seneste 40 år. Effekterne af både herbicider og kvælstof hver for sig er veldokumenteret i en række tilfælde, mens der i alvorlig grad savnes undersøgelser af kombineret virkning af herbicider og gødning i marknære arealer. 1.1.1 Viden om kvælstofs påvirkning af naturenDer er lavet ganske mange undersøgelser især internationalt af påvirkninger af fx ammoniakafdrift fra landbruget på skoves sundhedstilstand, skovbundsflora, højmoser, heder, overdrev etc. (se sammenfatning i Bak m.fl. 1999, Bobbink m.fl. 1998, Waide m.fl. 1999). Forskningen i kvælstofs påvirkning har i høj grad fokuseret på N-følsomme naturtypers reaktion på øget N-deposition fra luften for at fastsætte tålegrænser for eutrofiering. Der har derimod ikke været lavet mange undersøgelser af kvælstofs betydning for halvnaturen i agerlandet, der som udgangspunkt er stærkt næringsstofpåvirket og ikke omfattet af nationale eller internationale udpegninger. En væsentlig antagelse for denne undersøgelse er, at danske læhegn generelt (både konventionelle og nuværende økologiske) har haft en stor tilførsel af næringsstoffer gennem de seneste årtier i form af ammoniakafdrift og utilsigtet bredspredning af kunstgødskning og/eller husdyrgødning. Derfor er det forventet at næringsstofmængderne ikke på nogen måde er begrænsende, hvilket også viser sig ved at hegn er domineret af almindeligt udbredte konkurrencestærke plantearter (Tybirk m.fl. 2001). Det skal dog i relation til undersøgelsens baggrund bemærkes at konventionelt kvægbrug har større ammoniaktab end tilsvarende økologisk kvægbrug (Kristensen, 2003), men en sådan forskel vil ikke medføre, at hegnsbiotoper kan forventes at være N-begrænsede på økologiske bedrifter. 1.1.2 Viden om pesticiders påvirkning af naturenEn meget bred viden om påvirkninger af herbicider på natur og miljø er systematisk opsamlet i Bicheludvalgets rapport om Miljø og Sundhed fra 1999. Bicheludvalget konkluderer, at afdrift af herbicider kan have betydning for floraen i hegn og småbiotoper, men at der mangler systematiske undersøgelser af, hvorledes pesticider påvirker vilde planter og dertil knyttede dyr i hegn og andre småbiotoper (Bicheludvalget 1999, side 72). Sammenstillinger af resultater fra økologisk jordbrugs påvirkning af naturen i agerlandet har generelt vist positive tendenser sammenlignet med konventionelt landbrug (Stolze m.fl. 2000, Ahnstöm 2002). Langt de fleste studier er fra markfladerne, mens studier af effekter af driftsformer på agerlandets småbiotoper er meget sparsomme. Enkelte undersøgelser (fx Kleijn & Snoijing 1997, de Snoo 1999, Kleijn & Verbeek 2000) har dog påvist, at lysåbne markkantbiotoper generelt er beklædt med vegetation, der klart er domineret af konkurrencestærke planter, der trives med højt næringsindhold i jorden. Der findes kun enkelte studier af driftsformers (mht. pesticider og næringsstoffer) betydning for vegetation i læhegn og skovbryn. I Canada konkluderede Boutin & Jobin (1998), at der var signifikant færre skovarter ved intensiv drift. Den første samlede undersøgelse af vegetationen i 207 danske læhegn baseret på en række enkeltstudier og upublicerede data konkluderede, at meget udbredte konkurrencestærke og forstyrrelsestolerante arter dominerer (Tybirk m.fl. 2001). Kun en enkelt art fra hegnene figurerer på den danske Gulliste (Stoltze & Pihl 1998), mens de 12 mest almindelige arter forekommer i over halvdelen af hegnene. De fleste sjældne arter findes i gamle flerrækkede hegn (>30 år), og hegnsvegetationen er forskellig i Vestjylland og på Sjælland. Et tilbagevendende spørgsmål er, hvorvidt det er generel gødskning eller pesticider, der har medvirket mest til, at hegnsbiotoper er domineret af konkurrencestærke arter. I dag har man bedre muligheder end tidligere for at undersøge dette forhold, da antallet af økologiske brug er steget kraftigt, og det derfor er muligt at finde læhegn og lysåbne småbiotoper af forskellige typer, der gennem længere tid ikke har været påvirket af herbicidafdrift. Hald & Reddersen (1990) fandt, at der var flere ukrudtsplanter og leddyr i kanterne af danske kornmarker end i markens midte. Ligeledes viste en undersøgelse af Hald m.fl. (1994), at sprøjtefrie randzoner i marken resulterede i en rigere flora og som følge deraf en rigere leddyrfauna. Desuden er der generelt en større artsrigdom i økologiske marker end i konventionelt dyrkede (for planter, se fx Hald & Reddersen 1990, Rydberg & Milberg 2000, Jensen & Johnsen 2003, for leddyr, se fx Feber m.fl. 1998, Mäder m.fl. 2002, Navntoft & Esbjerg 2003, Navntoft m.fl. 2003). Derimod var det i samme undersøgelse (Hald & Reddersen 1990) ikke muligt at påvise en effekt af sprøjtefrie randzoner i de tilstødende kantbiotoper, da sådanne virkninger forventes at tage mange år. I den undersøgelse var der ikke taget hensyn til kvælstofafdriften, som kan have stor betydning for eventuelle effekter af reduceret herbicidpåvirkning. Man kan ud fra disse undersøgelser konkludere, at det er nødvendigt at tage udgangspunkt i læhegnene og udvælge disse så ensartet som muligt og udsat for hhv. økologisk og konventionel drift gennem lang tid for at kunne detektere eventuelle forskelle. Forventede positive effekter på flora og leddyrfauna ved fravær af herbicidafdrift på økologiske bedrifter kan potentielt skabe et bedre fødegrundlag for fugle og andre smådyr, der finder deres føde i og omkring marker, og dermed resultere i en større ynglesucces (fx Braae m.fl. 1988, Odderskær m.fl. 1997, Esbjerg & Petersen 2002). Nye studier har vist, at effekten af omlægning til økologisk jordbrug har en meget hurtig effekt på markens plante- og dyreliv (Navntoft m.fl. 2003), selvom jordens frøpuljer ikke kan ændres på en kort årrække. Vegetationen og fluefaunaen i gamle skel og hegn i agerlandet er ændret signifikant blot 3½ år efter omlægning til økologisk drift blandt kvægavlere (Petersen 2003, Petersen m.fl. 2003). Den væsentligste forskel på driften var fravær af pesticider. Resultaterne er overraskende, idet kvægavlere generelt har et lavere pesticidforbrug end planteavlere, og da der forventes en længere omstillingsperiode for naturindholdet i udyrkede småbiotoper end i markfladerne efter omlægning. Dette resultat udfordrer de gængse teorier om konkurrenceforhold og uddøen og indvandring af plantarter i agerlandet og forstærker behovet for eksperimentel adskillelse af de væsentligste påvirkningsfaktorer for at fortolke resultaterne. Generelt er fødekæderne meget væsentlige indikatorer for naturens tilstand i landskabet og for funktionelle interaktioner mellem trofiske niveauer. Der vil dog ikke blive lavet deciderede studier af fødekædeeffekter, men projektet vil diskutere kvaliteten af disse habitater som levested for en række forskellige grupper og funktionelle typer af organismer inden for planter og herbivore leddyr som basis for agerlandets fødekæder. Med henblik på at kvalificere denne diskussion vil nærværende projekt søge at udfylde centrale videnshuller, hvor fokus vil være på om herbicidafdriften eller ammoniakafdrifen til småbiotoper vil betyde mest for dominans af konkurrencestærke artsgrupper på bekostning af arter tilpasset lav næringstilgængelighed eller herbicidfølsomme artsgrupper. 1.2 FormålProjektets formål er at dokumentere og kvantificere herbicid- og ammoniakafdrifts separate og kombinerede effekter på vegetation og overjordisk leddyrfauna på marknære biotoper. Dette overordnede formål belyses gennem to tæt relaterede dele.
De to dele af projektet skulle henholdsvis påvise effekter af pesticidfravær i landbrugsdriften – repræsenteret ved økologisk drift – sammenlignet med konventionelt landbrugspraksis og kvantificere særskilte og interaktive effekter af de to væsentligste påvirkningsfaktorer på hegnsvegetation, nemlig herbicid og kvælstof. 1.3 HypoteserProjektet belyser gennem eksperimentel behandling med herbicid og ammoniak (simuleret ved kontrolleret N-tilførsel) og feltundersøgelser i læhegn flg. hovedspørgsmål: Feltundersøgelsen vil belyse om indførelsen af økologisk jordbrug kan forbedre habitatkvaliteten belyst ved planter og leddyr i læhegn, målt som antal arter, artssammensætningen eller frekvensen af udvalgte arter.
Eksperimentet vil belyse hvilken faktor (herbicidafdrift eller kvælstoftilførsel) der er mest betydende som plante- og leddyrfordelende faktor, målt som antal arter, artssammensætningen og frekvensen af udvalgte arter.
1.3.1 Statistiske overvejelserI hegnsstudiet er indsamlet prøver såvel midt i hegnene som i kanten af hegnene. For at kunne sammenligne kant- og midtprøver skal denne effekt inddrages i den statistiske model. Desuden er kant- og midtprøver indsamlet parvis (ligger ved siden af hinanden) for at kunne fokusere på sammenligningen. For prøver indsamlet i hegnene bliver den statisktiske model således: Yijkl = μ + αi + βj + (αβ)ij + Cik + Dikl + εijkl når prøverne i kant og midte tages parvis. Her er αi effekten af dyrkningsform i, βj er effekten af kant/midt (j), (αβ)ij er interaktionen mellem dyrkningsform og kant/midt, Cik er effekten af hegn nummer k i dyrkningsform i, Dikl er effekten af blok l i hegn k i dyrkningsform i . εijkl er effekten af den enkelte prøve. Cik antages indbyrdes uafhængige og normalfordelte med middelværdien 0 og variansen σB², εijkl antages indbyrdes uafhængige og normalfordelte med middelværdien 0 og variansen σ² mens Dikl antages indbyrdes uafhængige og normalfordelte med middelværdien 0 og variansen σD². Som led i planlægningen af 2. års hegnsundersøgelser blev der lavet en styrkeberegning, som viste, at for at opnå en styrke på 80 % med en Least Significant Difference (forstået som den mindste forskel, der testes som signifikant ved et 5 % signifikansniveau) på 30 % for antal plantearter pr. prøve krævedes 15 hegn af hver driftstype med 5 blokke /hegn. Variationen af gennemsnitlige antal (planter eller leddyr, artsantal såvel som antal individer) i læhegn kan beskrives ved (Yi.) = σB²/h + σ²/h*p, hvor h er antal hegn, p er antal prøver pr. hegn, σ² er variansen mellem gentagne prøver indenfor det samme hegn og σB² er varianskomponenten for hegn, altså den ekstra variation der er mellem prøverne, når de ligger i forskellige hegn. Dvs. antal hegn har større indflydelse på variansestimatet end antal prøver pr. hegn, og et øget antal hegn vil derfor give et bedre estimat af middelværdien. Her blev variabelen inde/ude udeladt, således at undersøgelsen i 2002 udelukkende fokuseres på den lysåbne urtevegetation langs hegnet (fodposen). Modellen blev derfor reduceret til: Yijkl = μ + αi + Cik + εijkl Denne model gælder også for 2001 for analyser af fodposedata alene (sandjord). For den eksperimentelle del af projektet er der udført styrkeberegninger for interaktionen mellem ammoniak og herbicid. Først er udregnet en hjælpestørrelse Φ² = nD²/(2σ²((a-1)(b-1) + 1)), hvor D er forskellen mellem to interaktionsled, σ er spredningen, n er antal replikater, a er antal herbiciddoser, og b er antal ammoniakdoser (Montgomery1984). Ved derefter at aflæse styrken på kurver i Montgomery (1984) har vi fundet frem til, at med α = 0,05 og β = 0.8 vil man kunne detektere en forskel i artsantal på 40 % ved 4 replikater og en forskel på 20 % ved 17 replikater (forudsat a=4 og b=3). Der etableres 10 replikater hvert bestående af 12 permanente prøvefelter med hver sin behandling. Behandlingerne kombineres i et 4x3-faktorielt design, som inkluderer 4 doseringer af glyphosat og 3 kvælstofniveauer. Modellen for test af effekter af glyphosat og kvælstof på planter og dyr er: Yijk = μ + αi + βj + (αβ)ij + Ck + εijk hvor αi er effekten af glyphosatbehandlingen, βj effekten af kvælstoftilførslen, (αβ)ij interaktionen mellem disse, Ck blokeffekten og εijk residualeffekten. Ud over variansanalyserne anvendes gængse multivariate analysemetoder som DCA og CCA til at udtrække og analysere de vigtigste plante- og leddyrfordelende faktorer (forskelle i artssammensætning) i datamaterialet samt opdele og kvantificere variationen i artssammensætningen på forskellige forklarende faktorer. Et økologisk studie hvor der indgår mange variable, nødvendiggør brug af gængse multivariate analysemetoder som DCA (Detrended Correspondence Analysis) (Økland 1990). I en DCA forsøger man at fitte arternes fordeling til en gaussisk responskurve over en kompleksgradient, hvor gradienten er relativ bred. DCA udbygger en CA (Correspondance Analysis) med et par ekstra trin for at afhjælpe nogle problemer ved CA (Jongman et al 1987, Økland 1990, Hill & Gauch 1980). DCA giver derved mulighed for at udtrække og analysere de vigtigste plante- og leddyrfordelende faktorer i datamaterialet. Ved hjælp af CCA (Canonical Correspondence Analysis, der er en indirekte ordination hvor ordinationsakserne er den linearkombination af miljøvariable, der forklarer den floristiske variation bedst) vil det endvidere være muligt at opdele og kvantificere variationen i artssammensætningen på forskellige forklarende faktorer (Borcard m.fl. 1992, Økland & Eilertsen 1994). Det vil således være muligt at vurdere om herbicidafdriften er en afgørende plante- og leddyrfordelende faktor i sammenligning med andre faktorer som fx jordbund og ammoniakkoncentration m.m. Monte Carlo-tests er endvidere et redskab til at kvantificere og teste om herbicidanvendelsen forklarer en signifikant del af variationen i henholdsvis plante- og dyreartssammensætningen. Datainput i gradientanalyserne vil være frekvensdata fra vegetationsundersøgelsen, frekvens- og antalsdata fra leddyrundersøgelsen samt kombinerede data fra vegetationsundersøgelsen og leddyrundersøgelsen. Med henblik på at underbygge og bekræfte om forskellige koncentrationer af herbicid- og ammoniakafdrift har en negativ indvirkning på naturindholdet i vegetationen kan den euclide afstand i et to- eller tredimensionalt ordinationsrum udregnes mellem referencenaturarealer og de observerede herbicid- og ammoniakbehandlede arealer. Referencenaturen til en sådan beregning kan bestå af tilfældigt udtagne prøvefelter fra græslandsnaturtyper med lang kontinuitet uden anvendelse af pesticid og gødning (udtaget fra Danmarks Miljøundersøgelsers vegetationsdatabase). Dette vil give mulighed for at teste (t-test) om der er forskel på økologiske og konventionelle læhegns vegetation i forhold til forskellige plantesamfund i landskabet. For leddyrenes vedkommende anvendes den multivariate programpakke PRIMER version 5.2.9 (Plymout Routines In Multivariate Ecological Research) (fx Clarke 1993, Clarke & Ainsworth 1993). For at reducere indflydelsen fra meget hyppige arter/grupper kvadratrods- eller fjerderodstransformeres data. Data for jordbunds- og plantevariable inddrages som forklaringsvariable. Ved hjælp af Draftsman-plots (Clarke 1993) undersøges den indbyrdes korrelation mellem de tilgængelige data for plantebiomasse og –diversitet samt jordbundsforhold, og for stærkt korrelerede variables vedkommende vælges en enkelt af disse ud til brug i analysen. Ligheden i artssammensætning mellem alle par af prøvefelter udregnes ved hjælp af Bray-Curtis similaritetsindeks (Bray & Curtis 1957), hvilket resulterer i en trekantet similaritetsmatrice. Ikke-metrisk multidimensionel skalering vha. proceduren MDS (Clarke 1993) bruges til at frembringe en ordination af leddyrmatricen, og i ordinationsplottene kan specifikke mijløvariable lægges ind som symboler med en størrelse svarende til deres aktuelle værdi i det pågældende prøvetagningsfelt, så sammenhængen mellem gradienten i artssammensætningen af leddyrene og miljøvariable. På baggrund af similaritetsmatricen undersøges korrelationen mellem artssammensætningen og de udvalgte miljøvariable (relateret til jordbund og planter) vha. BIOENV (Clarke 1993; Clarke & Ainsworth 1993, Somerfield et al. 1994). Denne procedure udregner rank-korrelationer mellem en similaritetesmatrice baseret på miljøvariable og den føromtalte similaritetsmatrice for leddyrsamfundet, hvorved de variable, der bedst forklarer variationen i leddyrsamfundet, kan identificeres. ANOSIM (Clarke & Green 1988) anvendes til at teste, om artssammensætningen af leddyrene er signifikant forskellig mellem foruddefinerede grupper af prøvefelter (fx forskellige doser glyphosat og kvælstof), både overordnet set og i parvise sammenligninger. Ved hjælp af proceduren SIMPER (Clarke 1993) kan de enkelte arters bidrag til forskelligheden i artssammensætningen mellem prøvetagningsfelter med forskellig behandling kvantificeres. 2 Vegetation og leddyr i hegn på økologiske og konventionelle bedrifter2.1 Materiale og metoder Dette kapitel beskriver læhegnsundersøgelserne på sandjord i år 2001 og på lerjord i år 2002. 2.1 Materiale og metoderHerbiciders utilsigtede påvirkning af naturindholdet i småbiotoper belyses i en komparativ undersøgelse af læhegnsvegetation og tilhørende leddyr på økologiske hhv. konventionelle bedrifter. Med henblik på at sikre maksimal sammenligning har en række kriterier været gældende for udvælgelsen af læhegn. Kriterierne for de undersøgte læhegn på sandjord var, at hegnet skulle
De økologiske hegn opfyldte yderligere flg. kriterium
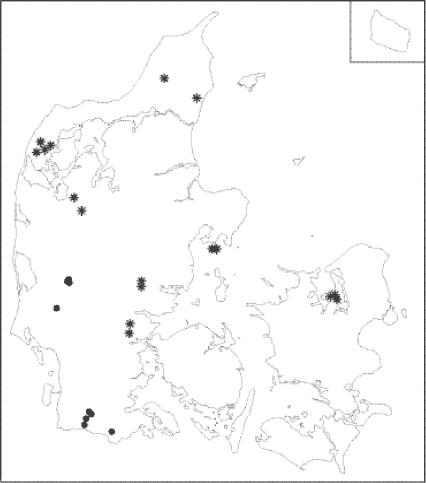
Seksogtyve (2x13) hegn blev undersøgt på konventionelle og økologiske bedrifter i 2001 syd og vest for israndslinien (dvs. JB 1-3(4)). De økologiske hegn blev udvalgt først og derefter blev tilsvarende konventionelle hegn i samme postdistrikt udvalgt. En detaljeret beskrivelse af de undersøgte hegn findes i Bilag A. I år 2002 blev der på baggrund af resultaterne fra 2001 udvalgt hegn efter de samme kriterier, blot med den forskel at hegnene skulle findes nord og øst for israndslinien og jordbundene skulle have et vist lerindhold, baseret på telefoninterview med ejeren (dvs. JB 3-7). Det blev dog i 2002 nødvendigt at udvide alderskriteriet (6-22 år, langt de fleste dog 10-15 år) i enkelte tilfælde for at opnå de 2x15 hegn der var målsat på baggrund af poweranalyser, da det var umuligt at opdrive et tilstrækkeligt antal hegn, der opfyldte samtlige kriterier. Eksempelvis blev der i enkelte tilfælde valgt konventionelle hegn i nabo-postdistrikter. Disse justeringer har været nødvendige for at opretholde minimum 10 års økologisk mark i omdrift på begge sider af hegnet som det væsentligste kriterium i udvælgelsen. Undersøgelsen omfatter dermed stort set samtlige eksisterende danske hegn, der opfylder de opstillede kriterier. Der er i alt i 2001 og 2002 inkluderet 28 hegn på 15 økologiske bedrifter og 28 hegn på 19 konventionelle bedrifter. Det typiske hegn var 14 år og 2,7 m bredt, 3-7 m højt og orienteret nord/syd. Den geografiske placering af hegn kan ses af Figur 2.1.
Figur 2.1. Placeringen af de undersøgte hegn i Danmark . Hegn markeret med cirkel er undersøgt på sandjord i 2001, hegn markeret med stjerne på lerjord i 2002. 2.1.1 Feltundersøgelse2.1.1.1 Prøvefelter 2.1.1.2 Hegnskarakteristika 2.1.1.3 Lysmåling 2.1.1.4 Jordbundskarakteristika Jordprøver indsamledes i oktober 2001 og august 2002. Fem delprøver af jord 0-10 cm under A0 horisonten blev poolet til én prøve. Jorden blev sigtet i laboratoriet gennem 2 mm sigte for at fjerne rødder, sten, mv. pH og ledningsevne blev målt i vandig opløsning. Frisk jord ektraheredes for uorganisk N og P i 1 time i 50 ml 0,5 molær K2SO4. Ekstraktet blev filtreret gennem Whatman GF-D-filter og nedfrosset til analyse for NH4+-N med indophenol metoden, NO3--N med cadmium reduktionsmetoden og PO4-P med molybdæn-blå-metoden (Allen, 1989). PO4-P ekstrationen var meget lav og er derfor udeladt. En delmænge af jorden blev tørret ved 70 grader C og knust. 4 gram heraf brændtes ved 550 grader C. En anden fraktion (200 mg tørvægt) blev opløst i svovlssyre i 1 time og analyseret for total N og P (Kedrowski 1983) med Hitachi U-2000 spektophotometer. C-total måltes med LECO CNS-1000 med metoden efter Sørensen & Bülow-Olsen (1984). Kornstørrelsesfordeling blev foretaget af Forskningscenter Foulum og de kemiske analyser blev foretaget af Økologisk Kemisk Laboratorium, Botanisk Institut, Københavns Universitet. 2.1.1.5 Vegetationsanalyse Til identifikation af planter er anvendt Hansen (1993), for græsser Hubbard (1984), for underarter af Rubus fruticosus (L.) Pedersen & Schou (1988) og for mosser Andersen m.fl. (1976). Der er ikke skelnet mellem underarter af mælkebøtte og rød svingel. Ud fra plantelisterne blev der udregnet en række indirekte hegnskarakteristiska, såsom total diversitet og vægtet gennemsnit af Ellenberg-værdier (Ellenberg m.fl. 1991) for næringsstofniveau (N), temperatur (T), kontinentalitet (K), pH (R), lys (L) og fugtighed (F) for alle prøvefelter. Gennemsnitlige vægtede C-S-R værdier (Grime 1979) blev tilsvarende udregnet for samtlige hegnsprøvefelter med metoden anbefalet af Ejrnæs & Bruun (2000). Total artsliste pr. hegn er fundet ved at registrere samtlige karplanter og mosser på én side i 0,5x100m felt placeret 20 cm fra afgrødekant. Hvert prøvefelt har således i alt 15 direkte og indirekte miljøvariable inklusive jordbundskarakteristika. Habitatpræference for arter er baseret på Hansen (1993) og Smith (1978). Derudover anvendes Grime's (1979) klassifikationssystem, hvor de enkelte arter under britiske forhold tildeles point for hhv C, S og R-strategi. Hansens (1993) klassifikation er enten-eller, mens Grimes er graduereret. Derfor kan en ruderat- eller en ager-art i Hansen godt være delvis C- eller S- strateg i Grimes system. Ukrudtsarter kan forekomme i agerjord men også findes i øvrige naturtyper. Ruderatarter er i øvrige naturtyper beskrevet som også forekommende i ruderate biotoper. Arter klassificeret som halv-natur-arter er ikke i ovennævnte kategorier, men i øvrige lysåbne biotoper. Tilsvarende gælder for skovarter. 2.1.1.6 Arthropod indsamling Leddyrene fra 2001 blev sorteret i henhold til sorteringsskemaet (Bilag C1). På grund af det uventet store antal dyr har vi kun optalt de grupper, som omfatter herbivore arter. De indsamlede prøver fra 2002 er sorteret som prøverne fra 2001, bortset fra, at antallet af bladlus og trips er talt og ikke kun estimeret, mens løbebiller og rovbiller er opgjort i to størrelsesgrupper (Bilag C2). Leddyrene er identificeret vha. Danmarks Fauna (Jensen-Haarup 1912, Hansen 1927; 1951, 1952, 1954, 1965, 1968, Gaun 1974), og nomenklaturen følger denne, bortset fra nogle af tægearterne, som er identificeret vha. Wagner (1961). 2.1.2 DatabehandlingFor vegetation, leddyr og jordbundskarakteristika er effekten af dyrkningsform og placering i hegnene testet i variansanalyse med SAS-proceduren MIXED, idet dyrkningsform og placering i hegn betragtes som systematiske (”fixed”) variable, mens hegnnummer og bloknummer betragtes som tilfældige (”random”) variable (beskrevet i afsnit 1.3.1). Samme type test er brugt til at teste for effekten af andre variable, fx hegnenes ejerforhold. Denne form for analyse forudsætter, at data er normalfordelte, evt. efter passende transformation. Hvis dette ikke kan opfyldes, fx pga. mange nuller i de observerede data, er i stedet anvendt en analyse, som baserer sig på poissonfordelingen (proc GENMOD i SAS), idet antalsdata med mange nuller med rimelighed kan antages at være poissonfordelte. Denne analyse kan ikke på samme måde som proc mixed kvantificere bidraget fra de tilfældige variable til variationen i responsdata. Proc GENMOD er anvendt på alle leddyrdata som supplement til proc MIXED, idet nogle leddyrarter kunne opfylde kravet om normalfordeling for nogle hegn, men ikke for andre. For at tage hensyn til, at der er taget flere prøver i hvert hegn, er anvendt et REPEATED-statement. De sjældneste arter er på grund af de mange nuller ikke egnet til statistisk analyse. I første omgang er data fra sandjord og fra lerjord analyseret hver for sig for at teste effekten af dyrkningsformen i de to geografiske områder. Efterfølgende er de to datasæt slået sammen, og effekten af jordbund er testet som systematisk variabel sammen med dyrkningsform, med hegnnummer hierarkisk til jordbund som tilfældig variabel (se afsnit 1.3.1). I fortolkningen af alle test er anvendt et signifikansniveau på 5 %. Vegetationens diversitet blev beregnet på tre niveauer: α (arter på prøvefelt-niveau), β-diversiteten (artsudskiftningen) og γ-diversiteten (antal arter i hele hegnet). β-diversiteten beregnes ved at udvælge 12 tilfældige sæt af 10 vegetationsprøvefelt fra hver driftstype og beregnes som total artsantal i de 10 vegetationsprøvefelt divideret med gennemsnitligt antal arter i de 12 sæt (Ejrnæs m.fl. 2002). 2.1.1.7 Supplerende analyser af vegetationsdata Plantede træer og buske ligesom dyrkede arter fra landbruget (kulturgræsarter, samt rød- og hvidkløver) blev udeladt af analyserne, men spontane vedagtige arter (kristtjørn, enebær, brombær, hindbær etc.) blev inddraget. Mosser er endvidere udeladt fra denne analyse. 2.1.1.8 Multivariat databehandling Ordinationerne af det totale vegetationsdatasæt (inklusive andre naturtyper) blev foretaget i PCord (McCune & Mefford 1999) uden nedvægtning af sjældne arter. Euclidiske afstande blev beregnet mellem prøvefeltscoren for alle økologiske og konventionelle hegn i dette studie (2x28) i det tre-dimensionelle ordinationsrum i forhold til den gennemsnitlige score for de øvrige naturtyper. Statistiske forskelle mellem Euclidiske afstande blev testet med t-test. Produkt-moment korrelationer beregnedes mellem ordinationsakserne og forklaringsvariable på såvel egne data som det totale datasæt. Signifikante forskelle i forekomsten af enkeltarter blev testet med en indikatorartsanalyse (McCune & Mefford 1999, Dufrene & Legendre 1997). Forskelle i leddyrenes artssammensætning mellem de to dyrkningsformer blev analyseret vha. ANOSIM-analysen i Primer (se 1.3.1.), og arternes bidrag til forskelle mellem dyrkningsformerne blev undersøgt vha. SIMPER. 2.2 Resultater2.2.1 Hegnenes jordbund og strukturAf de i alt 56 hegn der er undersøgt er 25 med græsmark i omdrift som nabo, heraf er 6 konventionelle og 19 økologiske græsmarker. 31 hegn har andre énårige afgrøder (fx. korn, grøntsager) som nabo, heraf 22 konventionelle og 9 økologiske. Hegnene er udvalgt for at ligne hinanden så meget som muligt med pesticidfravær som den væsentligste forskel på de to driftsformer. Forskelle i naboafgrøder er uden tvivl udtryk for de reelle forhold, da det var lettere at finde økologiske hegn der opfylder vore kriterier hos mælkeproducenter med meget kløvergræs. Hegnene var orienteret mod sydøst og bredden af fodposen var i gennemsnit 1,35m. De økologiske hegn var gennemsnitligt 14,7 år gamle og de konventionelle var 13,6. Data for de enkelte hegn forefindes i Bilag A. 2.2.1.1 Hegn på sandjord Tabel 2.1. Middelværdier for jordfysiske og –kemiske målinger i fodposen i hegnene på sandjord samt resultater af variansanalysen af effekter på de fysik-kemiske karakteristika af hegnnummer, blok (tilfældige variabel), dyrkning, position (fodpose eller centralt i hegnet) samt interaktionen mellem disse (systematiske variable). For tilfældige variable er det estimerede procentvise bidrag til den samlede ”tilfældige” variation opgivet. For systematiske variable fremgår signifikansnivauet af variansanalysen. I de tilfælde, hvor der er interaktion mellem de systematiske variable, kan effekten af de enkelte variable ikke bedømmes ud fra p-værdierne.
2.2.1.2 Hegn på lerjord De fleste strukturelle, jordkemiske og jordfysiske karakteristika var ikke signifikant forskellige for de to dyrkningsformer (Tabel 2.2), dog var total-N signifikant højere i de økologiske hegn end i de konventionelle. Variansbidraget fra hegnnummer i forhold til residualbidraget varierede meget mellem de målte responsvariable (Tabel 2.2), men for de fleste bidrog hegnnummeret langt mere til variationen, hvilket viser at der er meget store forskelle mellem hegnene i forhold til variationen inden for hegnene. Tabel 2.2. Resultater af variansanalyse for forskelle i middelværdier for strukturelle, jordfysiske og –kemiske målinger samt effekt af dyrkningsform i hegnene på lerjord. For tilfældige variable er det estimerede procentvise bidrag til den samlede tilfældige variation opgivet. P – Dyrkning = p-værdien for test af forskel mellem økologiske og konventionelle hegn.
2.2.2 Vegetationen2.2.2.1 Artsdiversitet i hegn Af Tabel 2.3 fremgår endvidere, at frekvensen af agerarter og antal arter pr. prøve var en signifikant effekt af dyrkningsformen på sandjorden. Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne, CSR-indikatorerne og antal skovarter. Antallet af arter med dokumenteret viden om herbicidfølsomhed er så lavt (0 i konventionelle hegn, 0,013 pr. prøve i økologiske), at analysen ikke isoleret set giver mening (se dog afsnit 2.2.2.5). Der var klart flere plantearter og højere tæthed i fodposen end midt i hegnene, og forskellen mellem driftsformerne skyldtes forskel i fodposens diversitet. De tilfældige varianskomponenters bidrag fremgår også af tabellen. Variansen mellem hegnene bidrog med mellem 23 og 83 % af den tilfældige variation. Tabel 2.3. Resultater af variansanalyse af udvalgte vegetationsdata for fodposen i hegn på sandjord. Ellenbergværdier er udregnet på basis af tilstede-/ikke tilstedeværelse af arter, CSR-værdier er vægtet med arternes frekvens, opdelt efter forekomst i forskeliige habitattyper, samt antal arter pr. prøvefelt (10m²). For tilfældige variable (hegn og residual) er det estimerede procentvise bidrag til den samlede “tilfældige” variation. For dyrkningsformen fremgår p-værdien af variansanalysen.
På lerjorden fandt vi i gennemsnitligt 36,1 arter i de økologiske hegn mod 24,1 arter i de konventionelle. Artsantallet pr prøvefelt samt antallet af naturarter, ruderatarter og agerarter var signifikant højere i de økologiske hegn på lerjord (Tabel 2.4). Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne, CSR-indikatorerne og antal skovarter. Variansbidraget fra hegnnummer er generelt ikke mere end maksimalt tre gange residualbidraget, hvilket betyder, at variationen mellem hegnene ikke er meget større end variationen inden for hegnene. Tabel 2.4. Resultater af variansanalyse af udvalgte vegetationsdata for fodposen i hegn på lerjord. Ellenbergværdier er udregnet på basis af tilstede-/ikke tilstedeværelse af arter, CSR-værdier er vægtet med arternes frekvens, opdelt efter forekomst i forskeliige habitattyper, samt antal arter pr. prøvefelt (10m²). For tilfældige variable (hegn og residual) er det estimerede procentvise bidrag til den samlede tilfældige variation opgivet. For dyrkningsformen fremgår p-værdien af variansanalysen.
2.2.2.2 Vegetationens sammensætning
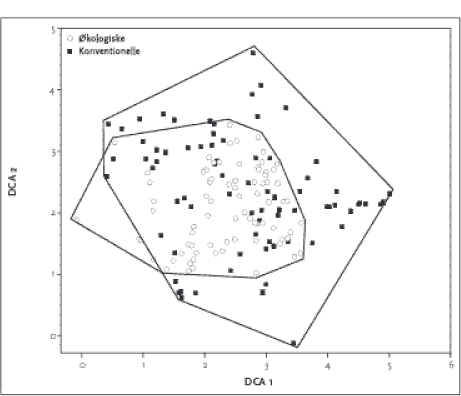
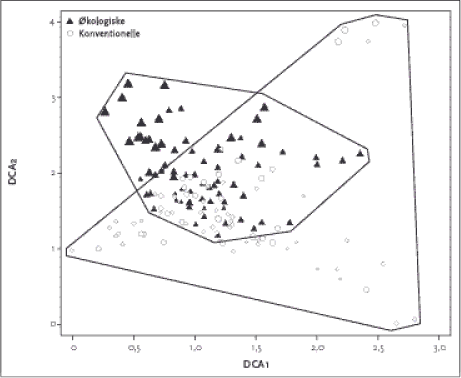
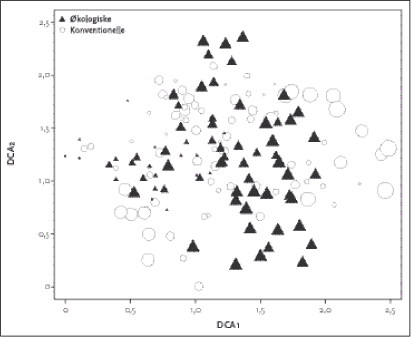
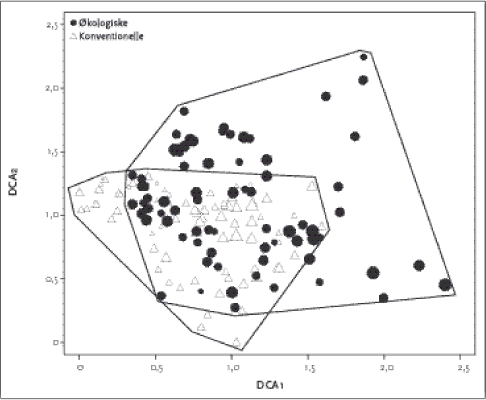
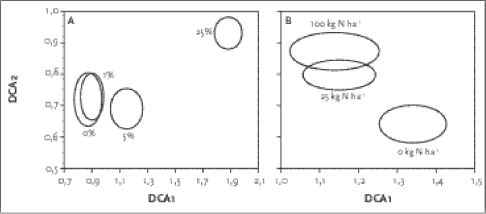
Figur 2.2. Placering af de 155 hegnsprøvefelter på sandjord langs de to vigtigste gradienter i datamaterialet. Polygonerne indikerer det totale areal omfattet af de to dyrkningsformer. Værdierne på DCA1 aksen korrellerede bedst med næringssttofstatus i jorden (Ellenberg N-værdi; P-total), med kornstørrelsesfordeling (mængde grovsand og fint sand) og lys, hvilket indikerer at DCA 1 afspejler en væsentlig gradient i såvel jordbund som lysforhold i artssammensætningen. DCA 2 havde højst korrelation med jordbundsvariable (finsand, P-total) og kanthøjden over markniveau. En DCA analyse af sammensætningen af vegetationen på lerjorden kan ses i Figur 2.3. Førsteaksen er ikke så lang som hos hegn på sandjorden, hvilket delvist skyldes, at der på lerjorden ikke er taget prøver centralt i hegnene. Der var et ganske stort overlap i artssammensætningen hos hhv. økologisk og konventionel drift. Det fremgår også, at der var en længere 1. og 2. akse og dermed større artsforskelle (beta-diversitet) mellem de konventionelle hegn. Produkt-moment korrellationer (Tabel 2.5) viser at DCA 1 på lerjorden primært var korrelleret med Ellenberg-Lys mens andenaksen var bedst korrelleret med Ellenberg-Næringsindhold, som afspejler jordbundsvariation. På sandjord var Ellenberg-Næringsindhold den vigtigste variabel.
Figur 2.3. DCA plot for vegetationssammensætningen for økologisk (trekant) og konventionelle hegn på lerjord. Størrelsen af symbolet angiver artsdiversiteten af de enkelte hegn. Tabel 2.5. Produkt-momentkorrelationskoefficienter mellem kalibrerede Ellenbergværdier og ordinationsscorene (DCA1 & DCA2) fra sand- og lerjorder.
2.2.2.3 Funktionel fortolkning af vegetationen i alle hegn I alt fandt vi 169 arter af urter, heraf var 153 karplanter inklusive Rubus-arter og 16 mosser. I de 28 økologiske hegn blev der fundet 162 arter og tilsvarende blev der fundet 115 arter i de 28 konventionelle hegn. Antallet af arter pr. prøvefelt var signifikant højere (P<0,0001) i økologiske hegn (19,1) sammenlignet med konventionelle (13,2) (Tabel 2,6). Selvom der var flere græsmarker langs de økologiske hegn, gav græsmarkerne ikke i sig selv nogen signifikant højere diversitet sammenlignet med andre afgrøder. Forskellen mellem driftsformerne viste også ved antallet af ruderatarter, naturarter og agerarter og C-strateger er signifikant højere i de økologiske hegn (Tabel 2.6). En uvægtet analyse med CSR-indikatorerne viste signifikant flere S-strateger i de økologiske hegn. Derimod er der ikke signifikante forskelle i Ellenbergindikatorerne og antal skovarter. Variansanalysen viste endvidere at der var signifikant effekt af jordbunden på en række af Ellenberg-indikatorerne, C-strateger samt antallet af ruderat-arter. Variansbidraget fra hegnnummer var op til tre gange residualbidraget, hvilket betyder, at variationen mellem hegnene var større end variationen inden for hegnene. Tabel 2.6. Resultater af variansanalyse i alle økologiske (n=84) og konventionelle (n=84) hegn. For hegnnummer inden for jordtype og residualeffekt fremgår det estimerede procentuelle bidrag til den samlede tilfældige variation. P-værdien for effekt af dyrkningsform (økologisk/konventionel) samt effekt af jordbund (sandjord/lerjord) er opgivet Interaktionsleddet mellem dyrkning og jordbund er ikke medtaget i variansanalysen, da det ikke var signifikant.
2.2.2.4 Vegetationens sammensætning og relationer til andre naturtyper Tabel 2.7 Produkt-moment korrelations koefficienter mellem kalibrerede Ellenberg værdier og ordinations scorer (n =1661) for DCA1-DCA3. Fede tal angiver værdier > 0.5.
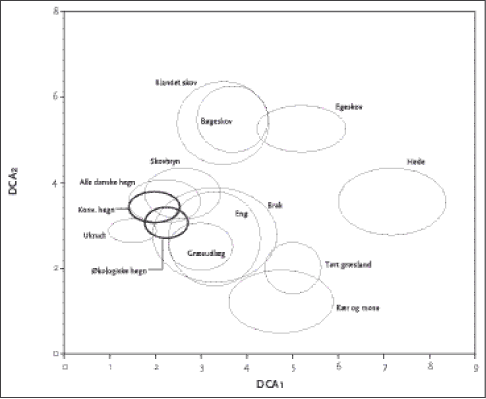
Figur 2.4. DCA figur der viser de undersøgte hegnsdata (de to fede ringe) i relation til en række udvalgte naturtyper. Det ses af Tabel 2.8, at de økologiske hegn havde signifikant større affinitet end konventionelle hegn til lysåbne kultiverede og stærkt påvirkede elementer som ukrudtsflora på marker, græsmark, dyrket eng og mose, samt i mindre grad til tørt græsland og hede. Endvidere havde økologiske hegn større affinitet til skovbryn end konventionelle. Affiniteterne til skovtyperne er relativt perifere, og vi fandt kun meget få obligate skovarter i hegnene, især i de konventionelle. Tabel 2.8. Gennemsnitlig euclidisk afstand mellem ordinationsscoren for hhv. økologiske (n=28) og konventionelle (n=28) og 12 naturtyper. T-test er anvendt til at beregne statistisk forskel mellem middelværdierne. Rækkerne er sorteret efter nærmeste Euclidiske afstand til økologiske hegn N.S.= ikke signifikant, ** = 0.001 <P<0.01, *** = P < 0.001. Fede tale indikerer, hvilken af de to hegnstyper der har signifikant affinitet til naturtyperne.
En indikatorartsanalyse viste, at tyve arter forekom signifikant hyppigere på en af driftstypene (på P<0,1-niveau) (Tabel 2.9). Mosset Brachythecium rutabulum (alm. kortkapsel) og urterne alm. hønsetarm, lav ranunkel, agertidsel, alm. svinemælk og lancetvejbred kan alle siges at være indikatorarter for økologiske hegn. Det er dog kun sidstnævnte der kun forekom i økologiske hegn, mens de øvrige blev også fundet i konventionelle hegn, men signifikant mindre hyppigt. Tabel 2.9. Indikatorarts-analyse af driftsform. Arterne er sorteret efter stigende p-værdi. Indikatorværdi og statistisk signifikans er beregnet med 1000 MonteCarlo permutationer. Arter med P ≥ 0.1 er ikke vist. Plantede hegnsarter og kulturplanter fra græsmarker er udeladt.
2.2.2.5 Herbicidfølsomhed Vi har kunnet konstatere, at der forekommer under 10 'glyphosat-følsomme' arter i vore hegn, og ingen af disse er særligt almindeligt forekommende. Derfor er der ikke foretaget en statistisk analyse af, om der er flere herbicidfølsomme plantearter i økologisk hegn jf. hypotese 2. Vi må konstatere, at den nødvendige viden pt. ikke er tilstede. 2.2.2.6 Sjældne arter 2.2.3 Leddyrfaunaen2.2.3.1 Data fra hegn på sandjord Som det fremgår af Tabel 2.11, var der på sandjord ikke signifikante forskelle mellem dyrkningsformerne på forekomsten af udvalgte arter og gruppe. Antal arter pr. prøve, antal dyr pr. prøve og diversiteten (Shannon-Wiener indeks) var heller ikke signifikant påvirket af dyrkningsformen. Derimod var der for de fleste responsvariable en signifikant forskel mellem prøver taget inde i hegnet og prøver taget i fodposen, og for alle disse gælder, at der gennemsnitligt var flere dyr i i fodposen (Tabel 2.10). Analysen af de tilfældige variable viser, at der ikke er nogen blokeffekt, hvorimod variansen mellem hegnene i de fleste tilfælde udgør en tredjedel af den tilfældige variation eller mere (Tabel 2.11). Numerisk set er variansen mellem hegnene i de fleste tilfælde meget større end variationen mellem dyrkningsformerne. DCA analysen på alle data med undtagelse af Stilbus sp., som lå uden for hovedsværmen, viste, at prøverne fra de konventionelle hegn havde en længere førsteakse end prøverne fra økologiske hegn (Figur 2.5). For andenaksen gør det modsatte sig gældende. Førsteaksen er bedst korreleret til finsand, total-P, jordbundstypen, og lysforholdene (Tabel 2.12), mens andenaksen er bedst korreleret til total-N, ledningsevnen og humusindholdet. At dømme efter størrelsesfordelingen af signaturerne i diagrammet er gradienten i finsand langs førsteaksen ret entydig for økologiske hegn, mens den er temmelig utydelig for de konventionelle hegn. Tabel 2.10. Gennemsnitlige antal af de forskellige leddyrgrupper pr. prøve (0,9 m²) i de to typer hegn på sandjord hhv. i fodposen (ude) og midt i hegnet (inde).
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. Tabel 2.11. Resultater af variansanalyse af udvalgte leddyrdata fra hegn på sandjord. For tilfældige variable er det procentvise bidrag til den samlede tilfældige variation opgivet. For effekten af systematiske variable, dvs. dyrkningsform, position i hegnet (inde/ude) samt interaktionen mellem de to variable, fremgår signifikansnivauet af variansanalysen. I de tilfælde, hvor interaktionen er signifikant, bør man se bort fra p-værdierne for de enkelte variable.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl.
Figur 2.5. DCA analyse på arthropod-data for samtlige 156 prøver på sandjord. Størrelsen af symbolet angiver forekomsten af finsand i feltet (som havde den stærkeste korrelation med 1. aksen, se Tabel 2.12). Hvis man kun ser på prøverne fra fodposerne (Tabel 2.13), er førsteaksen mest korreleret til lysforholdene og total-P og derefter til forekomsten af ruderat-planter. På andenaksen er finsand og grovsand vigtigst, men ellers er det de samme variable som for førsteaksen. Desuden viste ANOSIM-analysen af leddyrdata (se 1.3.1), at der var signifikante forskelle i sammensætningen af leddyrfaunaen mellem hegn på økologiske og hegn på konventionelle bedrifter (p<0,01). Analysen af, hvilke arter der betød mest for forskellene i artssammensætning mellem hegnene i de to dyrkningssystemer på sandjord, viste, at ingen arter eller gruppe bidrog med mere en 10 % (SIMPER, udregnet på baggrund af kvadratrodstransformation, se 1.3.1). Størst forklaringsværdi havde de meget talrige grupper (tæppespindere, stilkhvepse, fluer, myg, bænkebidere, delphacidae-nymfer), der tilsammen bidrog med 31,5 % og hver især med maks. 7,24 %. De enkeltarter, der bidrog mest til forskellen, var snudebillerne Apion flavipes og A. virens, som forklarede hhv. 2,48 og 2,45 %. Som det fremgår af Tabel 2.14, havde ingen af disse de plantearter, der adskilte de to hegnstyper (Tabel 2.9), som værtsplanter. En fjerderodstransformation, som ændrer vægtningen til fordel for sjældnere arter, gav ingen forskel i analysen af enkeltarters bidrag til forskellene i artssammensætning mellem hegnstyperne. Tabel 2.12. DCA-analyse af leddyr overfor udvalgte miljøvariable. ”fr.” = frekvens, ”ts.” = til stede/ikke til stede. Analysen er udført på det fulde datasæt, med undtagelse af Stilbus sp. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
* Udregnet på baggrund af artssammensætningen i feltet Tabel 2.13. DCA-analyse af leddyr overfor udvalgte miljøvariable. ”fr.” = frekvens, ”ts.” = tilstede/ikke tilstede. Analysen er udført udelukkende på data fra fodposen i hegn på sandjord. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
* Udregnet på baggrund af artssammensætningen i prøvefeltet Tabel 2.14. Fødekilder for udvalgte leddyr.
2.2.3.2 Leddyrdata fra hegn på lerjord Variansanalysen viste, at hegnsnummerets bidrag til variationen i leddyrenes forekomst var lige så stor eller op til dobbelt så stort som det residuelle bidrag, dvs. variationen inden for de enkelte hegn (Tabel 2.16). Snudebillen Apion flavipes og det totale antal dyr per prøve var dog mere påvirket af hegnsnummeret. Der var signifikante forskelle mellem de to hegnstyper (økologisk og konventionel) for skumcikader (flest i konventionelle hegn), de tre tægearter Scolopostethus affinis, S. thomsoni og Stenoderma holsatum (flest i økologiske hegn), snudebillerne Apion flavipes, Ceutorrhynchus assimilis (flest i økologiske hegn) og Ceutorrhynchus contractus (flest i konventionelle hegn). Overordnet set var der signifikant flere (15 %) arter/grupper i de økologiske hegn end i de konventionelle. Tabel 2.15. Gennemsnitlige antal leddyr per prøve (0,9 m²) i hhv. økologiske og konventionelle hegn på lerjord.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. Tabel 2.16. Resultat af variansanalyse på udvalgte leddyr fra hegn på lerjord. For de tilfældige variable fremgår det estimerede procentvise bidrag til den samlede tilfældige variation. For den systematiske variabel, dyrkningsformen, er p-værdien for test af forskel mellem økologiske og konventionelle hegn i hhv. proc mixed og proc genmod opgivet.
DCA-analysen på leddyrdata fra lerjorden (Figur 2.6) viste, at både første- og andenaksen var længere for økologiske hegn end for konventionelle. Derved lå en hel del af punkterne for økologiske hegn og kun enkelte af punkterne for konventionelle hegn uden for den fælles punktsværm. Førsteaksen var bedst korreleret med total-P, pH, antal naturarter (planter), antal plantearter ialt og vandprocenten i jorden (Tabel 2.17), mens andenaksen var positivt korreleret med hegnets alder og antal agerarter (planter) og negativt korreleret med fodposebredde og antal skovarter (planter). Med andre ord var begge akser korreleret med både jordbundskarakteristika og planterelaterede variable. ANOSIM-analysen af leddyrdata (se 1.3.1) viste, at der var signifikante forskelle i sammensætningen af leddyrfaunaen mellem hegn på økologiske og hegn på konventionelle bedrifter (p<0.01) Trips, delphacidae-nymfer og små rovbiller, som alle var almindelige i hegnen, var de grupper, der forklarede mest af forskellen i artssammensætning mellem økologiske og konventionelle hegn, nemlig tilsammen 14,3 % og hver for sig maks. 6,45 % (analyseret vha. SIMPER efter kvadratrodstransformation af data, se 1.3.1). Ligesom på sandjorden var snudebillearterne Apion flavipes og A. virens de enkeltarter, der foklarede mest af forskellen, hhv. 3,37 og 2,88 %. Som det fremgår af Tabel 2.14, havde ingen af de plantearter, der adskilte de to hegnstyper (Tabel 2.9), som værtsplanter. En fjerderodstransformation, som ændrer vægtningen til fordel for sjældnere arter, gav ingen forskel i analysen af enkeltarters bidrag til forskellene i artssammensætning mellem hegnstyperne.
Figur 2.6. DCA analyse på arthropod-data for hegn på lerjord. Prøver afbilledet tæt på hinanden har ens sammensætning af leddyrfaunaen. Størrelsen af symbolet angiver artsdiversiteten. Tabel 2.17. DCA-analyse af leddyr fra hegn på lerjordoverfor udvalgte miljøvariable. Kun dyrkningsform samt de 5 variable, der korrelerer mest med hhv. 1. og 2. aksen, er medtaget.
2.2.3.3 Alle leddyr – sand- og lerjord Tabel 2.18. Effekt af dyrkningsform, år og hegnnummer på udvalgte leddyr. For de tilfældige variable (hegnsnummer inden for jordtypen og det residuelle bidrag) er angivet det estimerede procentvise bidrag til den samlede tilfældige variation.For de systematiske variable ses p-værdien fra variansanalysen. ’+’ indikerer flere dyr på lerjord end på sandjord, ’-’ færre dyr på lerjord.
*Samlebetegnelse for mariehøns, glimmerbøsser, skimmelbiller m.fl. 2.2.3.4 Samspil mellem vegetation og leddyr – fødekæder En indikatorartsanalyse afslørede at nogle arter fandtes signifikant (P < 0.05) mere i hegn (kun fodposen) med græsmarker som naboafgrøde, og andre foretrak hegn der grænsede op til andre afgrøder (Tabel 2.20). En tilsvarende analyse mht. dyrkningsform er præsenteret i Tabel 2.21, som viser, at løbebiller samt en enkelt tægeart og kornbladbillen var hyppigere i hegn ved konventionelt dyrkede marker, men bladlopper, svirrefluer og nogle snudebiller foretrak hegn ved økologisk dyrkede marker. Tabel 2.19. Forekomsten af herbivore leddyr i relation til udvalgte strukturelle og planterelatede variable på sandjord.
*Prøver med færre end tre arter og arter som forekommer i færre end tre prøver er ikke medtaget. Tabel 2.20. Indikatorarters fordeling i forhold til om hegnet har græsmark eller andre afgrøder som nabo på sandjord.
Tabel 2.21. Indikatorartsanalyse af betydende arter/dyregruppers fordeling i forhold til dyrkningsform på de marker, der grænser op til hegnet på sandjord. De arter/grupper, der fandtes signifikant (P < 0.05) hyppigere i fodposen i hegn på hhv. økologiske og ikke økologiske bedrifter er vist.
2.3 Diskussion2.3.1 Hegn og jordbundDer var ingen forskelle mellem dyrkningsformen, hvad angår de strukturelle og jordbundsfysiske og kemisk karakteristika i hegn på sandjord. Derimod var der en del forskel mellem fodpose og den centrale del af hegnene, som formentlig skyldes vindaflejret materiale og gødningsspredning i markkanten (Tabel 2.1). På de bedre jorder fandt vi, at total-N var signifikant højest i økologiske hegn (Tabel 2.1 og 2.2), men det er uvist, hvordan denne forskel kan relateres til driftsformen. 2.3.2 Vegetation2.3.2.1 Artsdiversitet Generelt må det forventes, at artspuljen potentielt er næsten den samme for alle de inkluderede hegn, fordi jordbunds- og hegnsvariable ikke viser store forskelle. Der er dog i en række studier fundet flere arter på økologiske marker (Reganolds m.fl. 1993, Hald 1999, Stolze m.fl. 2000, Mäder 2002, Rydberg & Milberg 2000), så puljen af arter, der kan etablere sig fra marker, er lidt større på økologiske bedrifter. Der er ikke tidligere konstateret så klare forskelle mellem konventionelle hegn og økologiske hegns vegetation (Boutin & Jobin 1998, 2001). Andre studier har undersøgt markdriftens indflydelse på hegnsvegetationen over en kort årrække (Hald 1998, Marshall m.fl. 2002), men da hegnsvegetationen har vanskelige vilkår for spredning og etablering i et fragmenteret landskab kan man ikke forvente store effekter på få år (Milchunas & Lauenroth, 1995). Da de undersøgte hegn endvidere var relativt unge, er sandsynligheden, for at der er sket omfattende indvandring af mange arter fra halvnaturtyper og skov, ikke stor og formentligt ikke væsentligt forskellig på de to driftsformer. Udryddelsesraten kan formodes at værre større på konventionelle bedrifter som følge af herbicidanvendelsen, da selv små doser i afdrift kan påvirke den naturlige vegetation (Kleijn & Verbeek 2000, Boutin & Jobin 1998, 2001). Forskellene i artsantal kan således forklares ved forskelle i indvandringsrate af markarter (flest på økologiske bedrifter) og forskelle i udryddelsesrater af herbicidfølsomme arter ved de to driftsformer (Zobel m.fl., 1998). En række studier i England og Frankrig har tidligere vist, at landbrugsdriften har indflydelse på hegnsvegetation (Boatman m.fl., 1994; Cummins & French, 1994; Hegarty m.fl., 1994; McAdam m.fl., 1994; Le Coeur m.fl., 2002), således at mere ekstensive driftsformer giver højere diversitet. De vigtigste påvirkninger er afdrift af pesticider og ammoniak samt fejlplaceret kunstgødning på konventionelle bedrifter (Baudry m.fl., 2000; McCollin m.fl., 2000). Det er generelt accepteret, at en af de væsentlige påvirkninger af markkantvegetation er tilførsel af store næringsmængder (Hegarty m.fl., 1994, Barr m.fl. 1995, Kleijn & Snoeijing 1997, Kleijn & Verbek, 2000; Tybirk m.fl., 2001), som resulterer i dominans af nitrofile C-strateger med eliminering af S-strateger (sensu Grime 1979). Eutrofieringen er også markant i de økologiske hegn, og selvom vi kan konstatere forskelle mellem de to driftsformer er alle hegn eutrofe. I nærværende studie har antagelsen derfor været, at der ikke er mangel på næringsstoffer i de inkluderede hegn, så de alle forventedes at være domineret af mere eller mindre nitrofil vegetation. Driftsformerne var i princippet lige intensive i dette studie mht. næringsstofanvendelse, omend ammoniaktabet er lidt lavere på økologiske bedrifter (Kristensen m.fl. 2003). Driftsformerne var derimod forskellige mht. anvendelsen af pesticider gennem minimum 10 år, så det vi ser kan fortolkes som langtidseffekter af pesticidfravær. Et nyligt afsluttet studie inspireret af tesen om økologisk inerti i agerlandet (Milchunas & Lauenroth 1995, Reddersen m.fl. 1999) har dokumenteret en signifikant ændring i vegetationen og fluefaunaen i gamle skel og hegn på økologiske kvægbedrifter på sandjord allerede 3,5 år efter omlægning til økologisk drift (Petersen 2003, Petersen m.fl. 2003). Metoderne er meget sammenlignelige med vore, og resultaterne var ganske overbevisende på gamle skel efter få års omlægning. For vegetationens vedkommende forstærkes effekterne efter 7 års omlægning. Man kan således konstatere, at herbicidfravær giver effekter meget hurtigere end tidligere forventet. Det kan forklares ved, at hegnsvegetationen løbende rekrutterer arter fra den eksisterende frøbank og via langdistancespredning fra omkringliggende biotoper. I begge tilfælde har de indvandrende planter større overlevelse på økologiske bedrifter pga herbicidfravær, så ekstinktionsraten er lavere. 2.3.2.2 Funktionel fortolkning På trods af en del ligheder i sammenligning med andre naturtyper i landskabet, fandt vi dog også klare forskelle mellem de to hegnstyper. De økologiske hegn havde således flere mosser, flere ukrudts- og ruderatplanter og klarere affiniteter til lysåbne og mere forstyrrelsesprægede landskabselementer som ukrudtssamfund på marker, græsmark og dyrket eng (Tabel 2.3, 2.4 og 2.8, Figur 2.4). Endvidere havde de økologiske hegn flere arter fra de lysåbne halvnaturtyper, og derfor er de økologiske hegn et bedre levested for halvnaturarter. I studiet, som fokuserede på økologiske kvægbedrifter (Petersen 2003), fandtes signifikant flere S-strateger i hegn og skel på de økologiske bedrifter og flere Ruderatstrateger og højere Ellenberg N-værdier i hegn og skel på konventionelle. Billedet i nærværende undersøgelse var ikke så entydigt, både som følge af at jordbunden er meget mere variabel, og fordi vi har inkluderet forskellige bedriftstyper for at opnå tilstrækkelige hegn til sammenligningen. De arter, der fandtes hyppigere i økologiske hegn, var primært natur-, agerlands- og ruderatarter (Tabel 2.6). Dette stemmer fint overens med konklusionerne af Petersen (2003), at arter der har relativt let ved at kolonisere (de fleste ruderater og visse langdistancespredte naturarter), klarer sig bedre uden herbicidafdrift på de økologiske bedrifter. 2.3.2.3 Indikatorarter og refugier Alle hegn i vort studie var eutrofe og temmelig forstyrrede set fra en økologisk vinkel. Langt de fleste arter er meget almindelige og relativt uinteressante i relation til national og international naturbeskyttelse, men diversiteten er naturligvis væsentlig for bredden af de fødekæder der kan etableres. I England har man konstateret en reduktion i hegnsvegetationens diversitet på 14 % mellem 1978 og 1990 (Bunce m.fl. 1999). Vi kender ikke den historiske udvikling i hegnsdiversiteten i Danmark, men man kunne forestille sig et tilsvarende billede med stigende intensitet af landbruget. Der er interessante ligheder mellem de arter der er gået tilbage i England og de arter vi har fundet flere af i de økologiske hegn. 11 af 16 arter, der forekom signifikant mere i danske økologiske hegn, er gået tilbage i England i nævnte periode. Det gælder for alm. røllike, alm. hvene, alm. hønsetarm, agertidsel, rød svingel, fløjlsgræs, lancetvejbred, lav ranunkel, rødknæ, mælkebøtte og mark-ærenpris. Andreasen m.fl. (1996) har desuden fundet signifikant tilbagegang i Danmark for alm. hønsetarm, der var en indikatorart for økologiske hegn. Derimod er burresnerre, som var indikatorart for konventionelle hegn i denne undersøgelse fundet i fremgang i England (Bunce m.fl. 1999). Resultaterne i nærværende studie tyder således på, at økologiske hegn kan spille en væsentlig rolle som et bedre refugium for relativt almindelige arter, som dog er i tilbagegang i agerlandet i dag. Man kan endvidere forestille sig at ældre økologiske hegn kan spille en betydeligt større rolle i denne sammenhæng som refugium for bl.a. skovarter (Stidsen 2004). 2.3.3 LeddyrFor læhegnene på sandjorder (2001) kunne der ikke påvises signifikante effekter af dyrkningsformen på leddyrene via variansanalyserne. Dette skyldes meget små forskelle imellem dyrkningsformerne i de undersøgte variable (Tabel 2.10). På den anden side viste de multivariate analyser, at artssammensætningen var signifikant forskellig i hegnene på de to bedriftstyper, og at variationen i leddyrmaterialet kunne forklares lige så godt eller bedre end variationen i plantedata. Det faktum, at dyrene fordeler sig nogenlunde efter samme miljøvariable som planterne, inklusive dyrkningsformen, er formentlig udtryk for at dyrene fordeler sig efter planterne og ikke efter jordbundskarakteristika i sig selv. Leddyrfaunaen i de to år, dvs. på de to jordtyper, var forbavsende ens, men der var dog større forskelle mellem de to år end mellem de to dyrkningsformer. På lerjorden var der signifikante effekter af dyrkningsformen på artsantallet og på forekomsten af visse arter, hvilket delvis kunne genfindes i den samlede analyse af leddyrene. Forskellen mellem årene (og mellem hegnene) følger forskellen i jordbundsforhold og plantediversitet, og i hegnene på lerjord var der også en signifikant forskel i leddyrenes artssammensætning mellem de to dyrkningsformer. Analyserne af fordelingen af leddyr i forhold til planter og strukturelle variable viste, at de plantefordelende variable (primært jordbund) generelt var de vigtigste for de planteædende leddyr, men især for snudebiller var de omgivende markers afgrøde (græsning – ikke græsning) og dyrkningsform også vigtige variable. Disse to variable er ikke totalt uafhængige, idet frekvensen af hegn med græsmark som nabo var højere på de økologiske brug end på de konventionelle. Visse plantefamilier syntes også at påvirke forekomsten af tæger og snudebiller, uden at der dog nødvendigvis var tale om dyrenes værtsplanter. Snudebillernes præference for græsmarker som nabo (Tabel 2.19 og 2.20) kan evt. hænge sammen med forekomsten af kløver på disse marker, idet kløver er værtsplante for en del snudebillearter (Tabel 2.14). Sammenhængen mellem forekomsten af tæger og græs (poaceae) samt skærmplanter (Apiaceae) i hegnene synes rimelig, idet flere tægearter lever på disse typer af planter. 2.3.3.1 Fødekæder- samspil mellem planter og leddyr 2.3.4 HypoteserneDen første hypotese for hegnene var, at driftsformen ville påvirke artsantallet og artssammensætningen. Hypotesen kan bekræftes for planterne i nærværende undersøgelse. For de herbivore leddyr var det ikke muligt direkte at påvise sådanne effekter på artsantallet på sandjord, hvorimod der på lerjord og i det samlede datasæt var signifikant flere arter i de økologiske hegn end i de konventionelle, ligesom der var en signifikant korrelation mellem artsfordelingen og dyrkningsformen i DCA og signifikante forskelle i leddyrenes artssammensætning mellem de to dyrkningsformer. Den anden hypotese var, at der ville være flere herbicidfølsomme plantearter og flere specialiserede (fx værtsspecifikke herbivore) leddyr i de økologiske hegn end i de konventionelle. Det er vi ikke i stand til at konkludere om, idet vi kun har identificeret få dokumenteret herbicidfølsomme arter blandt de fundne, og vidensgrundlaget generelt har vist sig mangelfuldt til en sådan analyse. Det øgede antal plantearter i økologiske hegn giver i princippet mulighed for flere speciealiserede leddyr, men dette er ikke undersøgt i detaljen. Tredje hypotese forudsagde, at antallet af skovarter ville være større midt i økologiske hegn end midt i konventionelle. På grundlag af denne undersøgelse kan hypotesen imidlertid ikke bekræftes, idet der kun blev fundet 13 skovarter i alt og kun ganske få individer. For plantearter delvist tilknyttet skov ('fakultative' skovarter) var der dog signifikant flere i midten af de økologiske hegn end i de konventionelle. De økologiske hegn viste desuden i den totale DCA analyse bedre korrelationer med skovbryn end konventionelle. Det er klart, at hegnene generelt har været for unge til, at der kan være indvandret ret mange skovarter, der typisk er karakteriseret ved langsom spredning. Det kan derfor ikke udelukkes, at der vil være signifikante forskelle på ældre hegnstyper. 3 Eksperimentel adskillelse af herbicid- og N-påvirkning3.1 Materiale og metoder Den eksperimentelle adskillelse af herbicidbehandling og N-tilførsel afspejler til en vis grad et successionsstudie under forskellige påvirkninger af N og herbicid. I relation til hegnsundersøgelserne kunne forsøgets tre første år afspejle billedet af naturlig indvandring af plantearter i nyplantede hegn på en dybdepløjet jord med forskellige grad af landbrugspåvirkning. I eksperimentet øgede vi dog 'successionshastigheden' ved at udså en lang række arter for hurtigt at få en ensartet vegetation til statistiske sammenligninger. Formålet med undersøgelsen var imidlertid ikke at belyse ændringerne over tid, så der vil for vegetationens vedkommende blive fokuseret på resultaterne efter tredje år. For leddyrenes vedkommende præsenteres resultater for alle tre år (2001-2003), men dog med størst fokus på 2003. 3.1 Materiale og metoder3.1.1 ForsøgsetableringEn eksperimental vegetation blev etableret på nærings- og kalkfattig tør sandjord ved Ebdrup på Djursland. Marken havde tidligere været anvendt til korndyrkning, men havde ligget brak et år. Jorden blev dybdepløjet (60 cm dybt), harvet og tromlet for at begrave eksisterende frøbank fra pløjelaget. 120 permanente prøvefelter á 7x7m med 1,4m mellem prøvefelterne blev udlagt med 9m bufferzone rundt om eksperimentet, der er omgivet af trævækst på alle fire sider. 10 replikater af 12 unikke behandlinger blev etableret i et randomiseret blok design (Mead 1990). Behandlingene var 0, 1, 5 og 25 % af markdosis af glyphosat og 0, 25 og 100 kg N/ha/år. For at undgå begrænsning af andre næringsstoffer fik alle felter hvert år i maj tilført 53 kg Fosfor, 141 kg Kalium, 50 kg Svovl og 0,7 kg Kobber /ha/år. Glyphosat (RoundupBio, glyphosat-isopropylaminsalt, 360 g a.i./l) blev første gang udbragt efter etablering af tæt vegetation 24. august 2001 med vindhastighed på ca. 2 m/s, 50 cm over jordoverflade. Sprøjtebommen var 3 m bred med 50 cm mellem dyserne (Lurmark Lo-drift LD 015 Green) med et tryk på 2.0 bar. Anbefalet markdosis er ifølge producentens vejledning 4-6 l/ha. Vi valgte at bruge 4 l/ha (1440 g a.i./ha) som udgangspunkt for 0, 1, 5 og 25 % markdosis som resulterede i hhv. 0, 4, 20 og 100 ml glyphosat pr. 15 l vand. Anden og tredje glyphosatdosering fandt sted i april 2002 og 2003. 3.1.2 Artssammensætning af eksperimentel vegetationI alt blev 30 arter med forskellige funktionelle egenskaber udsået. Udvælgelsen af arter er foretaget så objektivt som muligt med fokus på, at det skulle blive en græsdomineret vegetation, hvor de vigtigste livsstrategier (funktionelle grupper af planter) var repræsenteret. Denne sammensætning blev suppleret med enkelte kendte herbicid- og/eller nitrogenfølsomme arter, arter i tilbagegang i agerlandet og en sjælden art. Endvidere har aktuel forekomst i læhegn/græsland og tilgængelighed af kommercielle frø spillet en rolle Udfra en bruttoliste på 255 arter blev der indhentet information om arternes livsstrategier. Karplanternes livsstrategier kan forklares med tre funktionelle syndromer, som betegnes CSR-strategier (Konkurrence-, Nøjsomheds- og Pionersyndromet): C for konkurrence (competition) tolerance og S for stresstolerante og R for ruderat/pionertilpassede eller forstyrrelsestolerante arter (Grime 1988, 1997; Ejrnæs 2000). Der findes ”rene” C-strateger som fx alm. kvik, men de fleste arter er en kombination af to eller tre syndromer. CSR-strategierne gives en værdi fra 0-12 for alle tre syndromer, hvor den maksimale sumværdi er12. For at få repræsenteret forskellige strategier i den udsåede vegetation blev arterne udtrukket stratificeret tilfældigt i de forskellige grupper, hvor der først udvalgtes en græsart for hver strategi for at sikre græsdominans. Derefter udvalgtes øvrige urter i de forskellige strategier. 3.1.3 Udsåning og etablering af vegetationSpiringsevnen hos de udvalgte arter blev testet eksperimentelt i petriskåle før udsåning. På trods af at marken stort set var plan og tilsyneladende meget homogen, opstod der en vis vand- og vinderosion efter dybdepløjning og såning. 3.1.4 Dataindsamling3.1.4.1 Jordbundsdata Prøverne for 01 og 03 blev analyseret for pHH2O, ledningsevne, glødetab samt total N og P. Frisk jord extraheredes for uorganisk N og P i 1 time i 50 ml 0,5 M K2SO4. Ekstraktet blev filtreret gennem Whatman GF-D-filter og nedfrosset til analyse for NH4+-N med indophenol metoden, NO3--N med cadmium-reduktionsmetoden og PO4-P med molybdæn-blå-metoden (Allen 1989). PO4-P ekstrationen var meget lav og er derfor udeladt. En delmænge af jorden tørredes ved 70 grader C og blev knust. 4 gram heraf blev brændt ved 550 grader C. En anden fraktion (200 mg tørvægt) blev opløst i svovlsyre i en time for analyse af total N og P (Kedrowski 1983) med Hitachi U-2000 spektophotometer. C-total måltes med LECO CNS-1000 med metoden efter Sørensen & Bülow-Olsen (1984). Kornstørrelsesfordeling blev foretaget af Forskningscenter Foulum og de kemiske analyser af Økologisk Kemisk Laboratorium, Botanisk Institut, Københavns Universitet. 3.1.4.2 Vegetation 3.1.4.3 Leddyr 3.1.5 Databehandling3.1.5.1 Vegetation DCA analysen af den eksperimentelle vegetation blev foretaget med følgende nødvendige forholdsregler, som anbefalet af Økland (1990). I alt 20 sjældne arter blev udeladt (forekom i mindre end 5 prøvefelter), ét prøvefelt blev udeladt (< fem arter til stede). Efter første DCA kørsel skilte fire prøveflader sig klart ud ift andre felter med > 1 s.d. enhed. De fire outliers blev fjernet fra analysen som anbefalet af Økland (1990). Der indgår derfor i alt indgår 115 prøvefelter (af 120 i alt) og i alt 29 arter. Ordinationen blev foretaget i PC-ORD (McCune & Mefford 1999) med standardopsætning. Kendall Tau-korrelationer blev beregnet for diskrete variable og produkt-moment korrelationer for kontinuerte variable. DCA-plottene er afbildet med middelværdien for en behandlingsgruppe som centrum (radius = 1 S.E.) i en cirkel. CCA-analyse blev foretaget på samme datamatrice med CANOCO version 4.02 (ter Braak & Smilauer 1997-99) efterfulgt af fortløbende udvælgelse og Monte Carlo permutationstest for at vise den relative betydning af de enkelte variable og hvorvidt de forklarende variable afviger fra permuterede tilfældige variable (Sokal & Rohlf 1995). Der blev anvendt 999 permutationer uden restriktioner. De forskellige variable blev standardiseret for at sikre ens vægtning af alle variable. For at analysere responsen af enkeltarter blev der lavet en indikator-artsanalyse (INSPAN) (Dufrêne and Legendre 1997) i PC-ORD. INSPAN blev udført på tre undergrupper af data, hvor enten herbicidbehandling eller N-tilførsel blev holdt konstant pga signifikante interaktionseffekter af de to faktorer i variansanalysen. 3.1.5.2 Leddyr Det fulde datasæt, hvor tæger, jordlopper og snudebiller er bestemt til art, blev undersøgt vha. den multivariate programpakke PRIMER version 5.2.9 (se 1.3.1). Data for jordbunds- og plantevariable blev inddraget som forklaringsvariable. Data blev om nødvendigt transformeret inden analyserne (primært for at bringe alle variable til nogenlunde samme størrelsesorden). Den indbyrdes korrelation mellem de tilgængelige data for plantebiomasse og –diversitet samt jordbundsforhold blev undersøgt, og for stærkt korrelerede variables vedkommende blev en enkelt af disse valgt ud. PI en 120120 matrice blev korrelationen mellem artssammensætningen og de udvalgte miljøvariable (relateret til jordbund og planter) undersøgt vha. BIOENV. Ved hjælp af MDS blev fremstillet plots af artssammensætningen i relation til behandlingsvariablene (glyphosat og kvælstof). To-vejs ANOSIM blev anvendt til at teste, om artssammensætningen af leddyrene var signifikant forskellig mellem prøvefelter behandlet med forskellige doser glyphosat og kvælstof. SIMPER blev brugt til at undersøge de enkelte arters bidrag til forskelligheden i artssammensætningen mellem prøvetagningsfelter med forskellig behandling undersøgt. Alle tests er vurderet på et 5 % signifikansniveau. 3.2 Resultater3.2.1 Jordbund og blok-effekterEn variansanalyse på jordbundskarakteristika viste signifikante effekter af kvælstoftilførslen på en række målte jordbundskarakteristika (ledningsevne, pH, NO3 og NH4 og uorganisk N). Det kunne dermed konstateres, at N-tilførslen har givet forskelle i tilgængeligt næringstofmængder. Endvidere viste der sig visse forskelle i kornstørrelsesfordelingen i forsøgsområdet (Figur 3.1).
Figur 3.1. Fordeling af fint sand (venstre) og ler (højre) i forsøgsarealet. Langs 'x- aksen' er de 12 behandlinger fordelt på de 10 tilfældigt placerede replica på 'y-aksen' 3.2.2 Etablering af plantearterI Bilag B findes fotos af den eksperimentelle vegetation. Tre udsåede arter (langbladet vortemælk Euphorbia esula, almindelig agermåne Agrimonia eupatoria og almindelig torskemund, Linaria vulgaris) spirede aldrig i petri-skålene, men blev etableret i felten. Fire måneder efter såning i 2001 var 23 ud af de 30 (77 %) udsåede arter etableret. Efter 3 vækstsæsoner var over 90 % af de udsåede arter etableret og alle fire grupper (C, S, R og blandet) af plantestrategier var vel repræsenteret (Tabel 3.1). Tabel 3.1. De udsåede arters højde, etablering og total biomasse efter 3 vækstsæsoner. (C = Competitiv strategi, S = Stress tolerant strategi, R = Ruderat strategi) ifølge Grime (1988) og Ejrnæs and Bruun (2000). Højde iflg. Lid (1987). Frøspiring er procent spiring af 20 frø i petriskål efter 59 dage (n=2). Etablering er udtrykt som procent af prøvefelter med succes fire mdr. og 2,5 år efter udsåning. De udsåede arters relative dominans er udtrykt som % af total høstet biomasse. Mindst 3 arter fra hver gruppe blev etableret i min. 40 % af felterne efter 4 mdr. En græsart fra alle fire funktionelle grupper etablerede sig, men kvik (Elytriga repens) blev dog dårligt etableret. Dette blev delvist kompenseret ved en kraftig etablering af stortoppet hvene (Agrostis gigantea med en CR strategi), der var iblandet frø af alm. hvene (A. capillaris). De tre dominerende græsser udgjorde 86 % af den totale høstede biomasse. Af de enårige blev almindelig dværgløvefod (Aphanes arvensis) aldrig etableret, mens enårig rapgræs (Poa annua) og kornblomst (Centaurea cyanus) blev etableret, men forsvandt stort set igen efter 2,5 år. Derimod har to andre enårige, salomons lysestage (Lepidium campestre) og mark-forglemmigej (Myosotis arvensis), formået at forblive omend kun som en sporadisk del af biomassen. 31 arter indvandrede til prøvefelterne, heraf 11 mosser, men ingen af de indvandrede arter formåede at dominere vegetationen (99,5 % af biomassen stammer fra de udsåede arter). De fem mest almindelige mosser var Ceratodon purpureus (104 prøvefelter), Bryum sp. (24 prøvefelter), Brachythecium rutabulum (17 prøvefelter), Ditrichum heteromallum (9 prøvefelter) og Polytrichyum piliferum (8 prøvefelter). 3.2.3 Klassisk analyse af vegetationsdataResultater af variansanalysen på vegetationsdata viste en signifikant interaktion af herbicidbehandling og N-tilførsel (Tabel 3.2). Interaktionen bestod i, at forskellene mellem N-niverauerne mindskedes med øget herbiciddosering, eller sagt på en anden måde, at herbicideffekten (skønt overordnet set ikke signifikant) steg med N-tilførslen (Figur 3.2). For førnemængden var der ligeledes signifikant interaktion mellem herbicid- og kvælstofeffekterne (Tabel 3.2). Der var en markant stigning i førneproduktionen med øget N- tilførsel ved lave herbicidtilførsler, og ligesom for den totale biomasse øgedes herbicideffekten ved stigende N-tilførsel (Figur 3.2). Tabel 3.2. Resultater af variansanalyse (Generaliseret lineær model LME -Linear Mixed Effect model) for vegetation på de eksperimentelle forsøgsfelter som funktion af herbicid behandling x produktivitet (N-tilførsel). Fed angiver statistisk signifikans. Ved signifikant interaktion mellem herbcid- og kvælstofeffekt giver det ikke mening at se på de separate effekter.
For karplantediversiteten bestod interaktionen mellem de to variable i, at artsantallet faldt med øget N-tilførsel, når der ingen herbicidtilførsel var, mens N-effekten faldt ved øget herbiciddosering (Figur 3.2). Sagt på en anden måde var der en negativ effekt af øget herbiciddosering i de ugødede felter, mens der ved 25 kg N/ha stort set ingen herbicideffekt var, og ved 100 kg N/ha var der en positiv effekt af øget herbiciddosering på antallet af karplantearter. Mosfrekvensen var signifikant påvirket af inteaktionen mellem N-tilførsel og herbiciddosering (Tabel 3.2, Figur 3.2), idet der i de ugødede felter var en negativ effekt af øget herbicidpåvirkning (især 25 % markdosis), mens der ved 25 kg N/ha ingen herbicideffekt var, og ved 100 kg N/ha var der en positiv effekt af herbiciddoseringen på mosfrekvensen.
Figur 3.2. Effekter af glyphosatbehandling (markdosis 1440 g a.i./ha) på totalbiomasse, førneproduktion, antal karplanter og mosfrekvens samt effekter på konkurrenceplanter (C-strrateger), nøjsomhedsplanter (S-strateger) og ruderatplanter (R-strateger) beregnet som vægtet biomassse ved tre niveauer af N-tilførsel (0, 25 og 100 kg N/ha/år). For de tre hovedgrupper af plantestrategier blev der fundet en signifikant effekt af både herbicidbehandling og N-tilførsel på den fremherskende plantestrategi (Tabel 3.2). Herbicideffekten var for alle grupper uafhængig af N-tilførslen. Den biomassevægtede C- og R-værdi faldt med øget herbiciddosering, mens S-værdien steg (Figur 3.2). Det betyder, at øget pesticidanvendelse giver lavere C- og R-værdier; men højere S-værdi i plantesamfundet. Glyphosat kunne alene reducerede artsantallet med maks. 23 %, og kvælstofgødskning reducerede artsantallet med maks. 29 % (Figur 3.2). De to faktorer indvirker således klart og nogenlunde lige meget rent kvantitativt på diversiteten. N alene virkede stærkt på plantebiomassen (fra ca. 500 til 830 g/m²), mens glyphosat ved højt N kunne reducere biomassen tilsvarende. Af Figur 3.2 kan det endvidere ses, at ved 0, 1 og 25 % herbicid var der ingen klar effekt af kvælstofgødskning på de tre strategier, mens der ved 5 % glyphosat var en tendens til at ved øget N-tilførsel steg andelene af C og R, mens S faldt.Ved 0 Herbicid: R var næsten konstant ved øget N-tilførsel, mens C udkonkurrerede S ved højere N.Ved 25 % herbicid: R næsten konstant ved øget N-tilførsel, S udkonkurrerede C ved højere N. S-strateger udkonkurrerede altså C- og til dels R-strateger ved høj herbicidbelastning ved såvel oligo- som eutrofe forhold. Her skal man dog være opmærksom på, at de enkelte strategiers biomasseproduktion primært udgjordes af fåresvingel, stortoppet hvene og rejnfan. En tilsvarede situation med få arter, der dominerer, er dog ikke ualmindeligt i agerlandet. En analyse af gennemsnitsværdier af total biomasse og diversitet for de 12 forskellige kombinationsbehandlinger viste, at uden N-tilførsel var der maksimal biomasse og diversitet ved ingen eller 1 % herbicid (Figur 3.3). Endvidere var der ved 25 kg N-tillførsel maximal biomasse ved 5 % herbicid, mens artsantallet var relativt ens for de fire herbicid-niveauer. Ved 100 kg N-tilførsel var der højst artsantal ved 25 % herbicid, men biomasse maximum ved 0 herbicid.
Figur 3.3: Gennemsnitsværdier af total biomasse for de 12 forskellige kombinationsbehandlinger (markdosis 1440 g a.i./ha) med indikation af artsrigdom. Symbol størrelsen er proportional med artsantallet, cirkler = 0 N, trekanter = 25 Kg N ha-1,firkant 100 kg N ha-1 Analysen af plantebiomasse, artsrigdom og mosfrekvens mod de forklarende variable viste, at biomassen korrelerede kraftigt og positivt med førne, C- og R-strategier samt ler, men korrelerer negativt med S- strategien (Tabel 3.3). Artsrigdommen og mosfrekvensen korrelerede ikke stærkt med nogle af de viste variable, men mosser havde dog en vis positiv korrelation med fint sand og negativ korrelation med førne. Tabel 3.3. Korrelationskoefficienter mellem biomasse, artsdiversitet, mosfrekvens og forklarende variable. Grime indikator værdier er baseret på vægtet gennemsnit af prøvefelter udregnet på biomassen.
3.2.4 Multivariat behandling af vegetationsdata3.2.4.1 DCA-analyse Tabel 3.4. Korrellationskoefficienter mellem de to første ordinationsakser og forklarende variable. Alle de viste variable er signifikant korrelleret til en af de to akser med p-værdier < 0.01. De forklarende variable er sorterede ift værdien i DCA 1 søjlen. Fed indikerer de variable med størst korrellation med DCA 1 og 2.
Dette afspejledes tydeligt i DCA plottene, når de fire glyphosatbehandlinger afbildedes (Figur 3.4a). Her fremgår det, at DCA 1 primært er en glyphosatrelateret gradient, men også at glyphosat i et vist omfang påvirker anden-aksen, hvor 25 % herbicid placerer sig relativt isoleret fra de øvrige herbicidbehandlinger. Andenaksen korrelerede svagt positivt med nitrogentilførsel og nitrat i jorden og svagt negativt med pH i 2003 og andelen af fint sand. Dette afspejledes mere tydeligt i DCA-plottene (Figur 3.4b) hvor de tre N-tilførselsniveauer blev afbildet. Der var en tydelig N-gradient i materialet på begge akser, hvor især 0 kg N isolerede sig fra 25 og 100 kg N (interaktion mellem herbiciddosering og kvælstoftilførsel).
Figur 3.4. DCA analyser af vegetationsdata for eksperimentet. Hver ring har middelværdien af de 10 replika som centrum og 1 standardfejl som radius. A viser herbicidbehandlingen (markdosis 1440 g a.i./ha), B de forskellige N-tilførselsniveauer. 3.2.4.2 CCA-analyse Tabel 3.5. Forklarende variable og ranket varians forklaret af de enkelte variable beregnet med fortløbende selektion af variable enkeltvis efterfulgt at Monte Carlo Permutations test. Variable der forklarer < 1 % er opført alfabetisk. N.S.= Ikke signifikant, * betyder p<0,05, *** betyder p<0,001. 3.2.4.3 Indikatorartsanalyse phosatfølsom, mens fåresvingel var meget robust over for glyphosat. Tabel 3.6. Indikatorartsanalyse i de forskellige herbicid-doseringsgrupper (markdosis 1440 g a.i./ha) under oligotrofe (0 kg N) forhold. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans (p < 0,05).
Tabel 3.7. Indikatorartsanalyse i de forskellige herbicid-doseringsgrupper (markdosis 1440 g a.i./ha) under eutrofe (100 kg N) forhold. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans (p < 0,05).
Andre arter reagerede forskelligt under de to N-niveauer – omend billedet ikke var så statistisk klart. Alm. kongepen (Hypochoeris radiata), hvidmelet gåsefod (Chenopodium album) og mosset Ceratodon purpureus klarede sig signifikant bedre i konkurrencen med højt glyphosatniveau og høj N-tilførsel, mens dette ikke var tilfældet ved lavt N-niveau, hvor der var en ikke-signifikant tendens til det modsatte. Uden herbicidbehandling kunne vi finde indikatorarter med klar respons på forskellig N- tilførsel (Tabel 3.8). Alm. kongepen (Hypochoeris radiata), alm. kællingetand (Lotus coniculatus) og mosset Ceratodon purpureus klarede sig signifikant bedre i konkurrencen uden N. Kvik (Elytrigia repens) og rejnfan (Tanacetum vulgare) reagerede derimod positivt på N- tilførsel. Prikbladet perikon (Hypericon perforatum) reagerede intermediært ved at have optimum ved 25 kg N. Tabel 3.8. Indikatorartsanalyse i de forskellige N-tilførselsniveauer uden herbicidbehandling. Kun arter med Indikatorværdi (IV) >20 er medtaget. i indikerer arter der ikke er udsået. Fed angiver signifikans.
3.2.5 LeddyrBlandt de grupper af leddyr, der er sorteret (dvs. herbivore gruppe, løbebiller og rovbiller), var tæger de talrigeste med i gennemsnit 67 dyr per prøve i 2003, hvoraf de fleste var blomstertæger (48 per prøve) efterfulgt af frøtæger (12 per prøve). Cikaderne var også talrige (gennemsnitligt 56 per prøve), rovbiller forekom med et gennemsnit på 7 per prøve, Clavicornia-gruppen (skimmelbiller, mariehøns og glimmerbøsser) havde et gennemsnit på 4 per prøve, hvoraf 3 var skimmelbiller, og de øvrige grupper (bier/hvepse, svirrefluer, sommerfugle, bladlopper, løbebiller, smeldere, bladbiller, jordlopper og snudebiller) forekom alle i et gennemsnitligt antal under 1 per prøve. 3.2.5.1 Udvikling over år Set over alle tre år var der signifikante pesticideffekter på bier og hvepse, svirrefluer, cikader, de fleste tægegrupper, jordlopper, små rovbiller, rovbillelarver, alle Clavicorniagrupper (skimmelbiller, glimmerbøsser og mariehøns), det samlede antal leddyr og antallet af arter (Tabel 3.9). Der var i alle tilfælde tale om et fald i antallet ved højeste pesticiddosering sammenlignet med de usprøjtede felter (se Figur 3.5). For nogle grupper var der tale om et begyndende fald i antalllerede ved 1 % af markdosis, mens der for andre først var en negativ effekt ved 25 % af markdosis. Effekten af kvælstof var signifikant for næsten de samme grupper som pesticideffekten, med undtagelse af bier/hvepse og svirrefluer, som ikke var påvirket af kvælstofniveauet, og sommerfugle og løbebillelarver, som var signifikant påvirket af kvælstof, men ikke af herbicidet (Tabel 3.9). For de fleste grupper steg det gennemsnitlige antal med stigende kvælstoftilførsel, men for Scutelleridae-/Pentatomidae-tægenymfer var effekten omvendt, og løbebillelarvernes antal steg ved 25 kg N/ha sammenlignet med 0, men faldt så ved 100 kg N/ha. Tabel 3.9. P-værdier for effekt af pesticid- og N-behandling på leddyrfaunaen. Testen er udført på data fra årene 2001-2003. I de tilfælde, hvor interaktionleddet har en signifikant effekt, kan effekten af de enkelte variable ikke vurderes ud fra tabellen.
* Testet med interaktionsleddet Af Tabel 3.10 fremgår, at blokeffekten maksimalt udgjorde 10 % af den samlede tilfældige varians, mens forskelle i forekomsten af leddyr mellem årene gennemgående udgjorde en større del (op til 56 %). Blokeffekten varierede mellem årene (interaktion), idet der i 2001 og 2003 var en kraftigere blokeffekt end i 2002. Variationen mellem felter inden for blokken (felt(blok) i Tabel 3.10) var af cirka samme størrelse som variationen mellem blokke. Tabel 3.10. Procentvis bidrag til den samlede ”tilfældige” variation for det samlede leddyrdatasæt (2001-2003). Data er testet uden interaktionsleddet glyphosat*NH4 som systematisk variabel.
* Data for alle år 3.2.5.2 Leddyr år 2003 Tabel 3.11. P-værdier (Proc MIXED) for effekt af glyphosatbehandling og kvælstoftilførsel på leddyr indsamlet i 2003. Effekten af hver af de to variable kan kun vurderes i de tilfælde, hvor interaktionen ikke var signifikant. Tal i parentes indikerer problemer med testen (for få dyr, manglende konvergens el. lign.).
* Testet med interaktionsleddet glyphosat*kvælstof som systematisk variabel. Af Figur 3.5 ses, at antal arter og antal dyr per prøve er reduceret ved den højeste glyphosatdosering. Felterne, der modtog 100 kg N/ha, havde flere dyr og arter i den usprøjtede situation end de andre kvælstofniveauer, men faldet i antal dyr og arter som funktion af stigende pesticiddosering var også størst i 100 kg N-felterne (Figur 3.5). Den maksimale herbicideffekt var 47 % reduktion i antallet af dyr og 26 % reduktion af artsantallet. Kvælstoftilførsel resulterede i en øgning af antal leddyr på maks. 55 % og maks. 21 % flere arter (cf. Figur 3.5). Den pesticideffekt, der ses for antal dyr ialt, går igen for de fleste øvrige grupper med signifikant pesticideffekt. For bier/hvepse var der dog ingen klar tendens, mens store løbebiller (for få dyr), og skimmelbiller forekom i mindre antal ved 1 og 5 % markdosis end i de usprøjtede plots, men i højest antal i plots behandlet med 25 % af markdosis (middelværdier hhv. 3,1; 2,3; 1,8 og 4,1). Antallet af de forskellige grupper steg generelt med stigende kvælstoftilførsel, med undtagelse af Scutelleridae-/Pentatomidaetægenymfer, som forekom i faldende antal med stigende kvælstoftilførsel. Den tilfældige variation mellem prøverne udgjordes for op til 30 %s vedkommende af variation mellem blokke (Tabel 3.12), mens variationen mellem felter inden for blokke udgjorde op til 64 %. Begge variationsparametre varierede meget mellem de forskellige grupper af leddyr.
Figur 3.5. Antal leddyr (minus bladlus og trips) og antal leddyrarter per prøve i årene 2001-2003 som funktion af glyphosatdosis (markdosis 1440 g a.i./ha) ved de tre forskellige gødskningsniveauer. Punkterne repræsenterer rmiddelværdier af 10 prøver, de vertikale linjer er standardfejlen på middeltallet. MDS-plottene (Figur 3.6) viser, at der er en sammenfaldende gradient i artssammensætning på den ene side og glyphosatbehandling eller plantebiomasse på den anden, mens det ikke er tilfældet for kvælstoftilførslen. Med andre ord er forskelle i artssammensætning bedre korreleret med pesticidbehandlingen og plantebiomasse end med kvælstoftilførslen. Den mest forklarende enkeltvariabel blandt de undersøgte jordbunds- og plantevariable (miljøparametre) var total plantebiomasse (korrelation 0,247 i BIOENV), efterfulgt af pesticidbehandlingen (korrelation 0,202). Ingen andre miljøvariable havde en korrelation på over 0,2 med artssammensætningen af leddyr. Når det maksimale antal forklarende variable blev sat til tre, var artssammensætningen af leddyr bedst korreleret med pesticidbehandlingen, den totale plantebiomasse og førnebiomassen (korrelation 0,351). Da der var en vis sammenhæng mellem de to biomassemål, blev førnebiomassen efterfølgende udelukket fra analysen, hvorefter pesticidbehandlingen, den totale plantebiomasse og det i 2003 målte NO3-N forklarede artsfordelingen bedst (korrelation 0,336). Når maksimalt fem forklarende variable indgik i analysen, var korrelationen bedst med en kombination af pesticidbehandling, kvælstoftilførsel, antal plantearter, førnebiomasse og bloknummer (korrelation 0,396). En opdeling af plantebiomassen i græsser, øvrige urter (ikke-græsser), mosser og træer og efterfølgende analyse viste, at artssammensætningen af leddyr var bedst forklaret af græsbiomassem (korrelation 0,14 i BIOENV), efterfulgt af den samlede plantebiomasse (0,127) og biomassen af øvrige urter (0,077).
Figur 3.6. Multidimensionel scalings-plots med hhv. pesticidbehandling (A), gødskning (B) og total plantebiomasse (C) lagt over, dvs. afstanden mellem boblerne er proportional med forskellen i artssammensætning, og størrelsen af boblerne svarer til behandlingsintensiteten/plantebiomassen. To-vejs ANOSIM (Tabel 3.13) gav en overordnet R på 0,188 for effekten af glyphosatbehandlingen (maks. 1,0) på artssammensætningen og et signifikansniveau på 0,001 (testet på tværs af kvælstoftilførselsniveauerne). Artssammensætningen i de usprøjtede felter samt felter behandlet med 1 og 5 % af markdosis var signifikant forskellig fra felterne behandlet med 25 % af markdosis. For kvælstoftilførslen var der en R på 0,177 og et overordnet signifikansniveau på 0,001 (testet på tværs af pesticidniveauerne). Der var signifikante forskelle i artssammensætningen mellem alle kvælstoftilførselsniveauer. Ingen enkeltarter bidrog iøjnefaldende til forskellene i artsammensætning mellem felter behandlet med forskellige glyphosat- og kvælstofdoser (SIMPER). Det maksimale bidrag var 11 % af forskellen mellem felter sprøjtet med 25 og 5 % af markdosis forklaret af Stenodema-nymfer (blomstertæge, lever på græsser), baseret på en gennemsnitlig frekvens på hhv. 4,21 og 6,12 i de to pesticidniveauer. Denne type tæge var i alle, undtagen et, tilfælde den mest forklarende for artsammensætningsforskelle mellem såvel pesticid- som kvælstoftilførselsniveauer, efterfulgt af andre tæger, cikader og små rovbiller i varierende rækkefølge. Tabel 3.12. Procentvis bidrag til den samlede ”tilfældige” variation for leddyr indsamlet i 2003.
Tabel 3.13. ANOSIM-test af effekten af glyphosat (markdosis 1440 g a.i./ha) og kvælstof på artssammensætningen af leddyr.
3.3 Diskussion af felteksperimentets resultater3.3.1 Jordbund og forsøgsetableringEksperimentet har generelt været meget vellykket i forhold til etablering og udvikling af vegetationen. Enkelte arter havde svært ved at etablere sig fra frø (fx kvik, stor nælde, gråbynke og agertidsel som er udbredte og konkurrencestærke hegnsplanter). Såbedet var stærkt soleksponeret, og få dages udtørring af frø kan have haft afgørende betydning for nogle arters etablering. Flere af C-strategerne formerer sig vegetativt, så de er vanskelige at etablere fra frø. De udsåede arter dominerer vegetationen og græsser opnåede den ønskede dominans – mens indvandringen af arter, specielt mosser, gav yderligere dimensioner til eksperimentet mht. koloniseringsmuligheder for planter. Selvom arealet var udvalgt som værende homogen mht. jordbund, viste der sig dog visse forskelle i jordbunden. Den tilførte kvælstof har derudover haft tilsigtede signifikante effekter efter 3 år på kvælstofniveauet og dertil knyttede sideeffekter på pH og ledningsevne. 3.3.2 Effekter af herbicidbehandling og N-tilførsel på vegetation og leddyrHovedformålet med eksperimentet var at belyse effekten af herbicidafdrift og kvælstoftilførsel. Eksperimentet har påvist klare interaktioner mellem N- og glyphosatpåvirkningen for plantebiomasse, førne, karplantediversitet, mosfrekvenser og dominansen af konkurrencestærke plantearter. For plantebiomasse og førne var der en meget markant reduktion ved 25 og 100 kg N i kombination med 25 % glyphosat, men denne effekt udeblev ved 0 kg N. For mosserne var frekvensen ved høje kvælstofniveauer klart højere ved 25 % glyphosat end ved lavere doser, hvorimod det omvendte var tilfældet ved lavere kvælstofdoser (Figur 3.2). Glyphosat ændrer totalt konkurrenceforholdene og dermed artssammensætningen af planter. Det skyldes sandsynligvis, at fåresvingel er både nøjsom og tilsyneladende relativt glyphosat-tolerant, således at den på trods af 25 % markdosis opnår en rimelig biomasse og dominerer felterne (Tabel 3.6), hvorimod de herbicidfølsomme arter forsvinder. Glyphosat forhindrer en række arter i at etablere sig og mindsker artsdiversiteten ved 0 kg N-tilførsel. Ved 25 kg N er der stort set ingen effekt af glyphosatbehandlingen, mens der ved 100 kg N er en positiv effekt af stigende glyphosatdosering, således at artsantallet ved 25 % af markdosis er sammenligneligt med antallet ved 0 og 25 kg N (Figur 3.2). Dette skyldes formentlig, at glyphosaten virker så negativt på nogle arter (konkurrencestrategerne), at deres tilbagegang giver nicher til relativt mere glyphosattolerante arter (bl.a. R- og S-strateger). Vi skal dog bemærke, at det især er meget få arter fra hver strategi, der dominerer vegetationen, så billedet kunne være anderledes med andre arter, selvom få arter, der dominerer i hver strategi, ikke vil være usædvanligt i hegnsvegetation. Glyphosat kan alene reducere artsantallet med ca ¼, og en tilsvarende reduktion i artsantal ses af kvælstofgødskning (Figur 3.2). Begge faktorer indvirker således klart og nogenlunde lige meget rent kvantitativt på diversiteten. Glyphosatens isolerede betydning på plantebiomassen er meget klar ved højt N-nivau, men næsten fraværende ved 0 kg N. En mulig forklaring på, at C-strategerne tilsyneladende påvirkes mest af glyphosatbehandlingen kunne være, at C-strategerne oftest er højere og derfor måske kan 'skygge' for glyphosatafdrift på R- og S-strategerne. Vi har ingen målinger af afsat mængde glyphosat på de forskellige planter, men det er ikke vort indtryk, at der har været væsentlig forskel i afsætningen af glyphosat på de enkelte arter. Udbringningen er sket om foråret (maj), hvor de fleste planter endnu er små. Den potentielle forskel i eksponering bliver først mere udpræget senere i vækstsæsonen. Måske er konkurrenceplanterne generelt mere følsomme end de øvrige, fx pga. deres højere vækstrate. En sådan forklaring stemmer overens med, at ved stigende glyphosatdosering forsvinder den positive effekt af N-tilførsel på den totale plantebiomasse (Figur 3.2). En anden mulig forklaring på den interaktive effekt af N-tilførsel og glyphosatdosering er, at selv beskedne glyphosatdoser hindrer en konkurrencerelateret uddøen (competitive exclusion), som ellers typisk er resultatet af tre år med øget N-tilførsel, fordi de dominerende nitrofile arter bortskygger små og lave arter. Glyphosatbehandlingen vil på den måde kunne åbne vegetationen for mindre konkurrencestærke arter selv ved 100 kg N/ha/år. Med de multivariate analyseteknikker viste det sig, at glyphosat er den væsentligste variabel til ændringer i vegetationssammensætningen (Tabel 3.6). Jordbundskarakteristika og N-tilførsel viste sig således at være af sekundær betydning i forhold til de anvendte glyphosatkoncentrationer. Det skyldes, at glyphosat ved 0 kg kvælstof øger uddøen af herbicidfølsomme arter og ved 100 kg N virker glyphosat især ved at reducere biomasse og førne. Dette åbner mulighed for øget indvandring/etablering af nye (nitrofile) arter, som ikke findes ved 0 kg N. Glyphosat kan således virke både reducerende og fremmende afhængigt af N-niveauet. Indikatorartsanalysen viste, at nogle arter reagerer meget forskelligt på herbicidbehandling afhængig af N-tilførslen. Dette kan have været medvirkende årsag til, at de forskellige funktionelle plantegrupper har reageret så markant, idet deres biomasse ofte er domineret af en eller meget få arter. Hvis vi eksempelvis havde udsået en anden, mere glyphosatfølsom nøjsomhedgræs end fåresvingel, havde billedet måske set anderledes ud. Hvis stor nælde, agertidsel, gråbynke eller kvik var slået an, kunne billedet også have været anderledes. Derfor skal dette resultat tolkes med varsomhed. Det ændrer dog ikke ved, at arter har forskellige reaktionsmønstre på glyphosat og næringsberigelse, og at en vegetation, der udsættes for herbicidafdrift, meget vel kan være domineret af en eller få arter. For leddyrene var der signifikante effekter af glyphosatbehandlingen på en lang række grupper, på antal dyr og på diversiteten, uafhængigt af N-tilførslen (Tabel 3.9, Figur 3.5). Specielt 25 % glyphosat slog kraftigt igennem på leddyrene. Eftersom glyphosat formentlig ikke er giftigt for leddyrene, må der være tale om en indirekte effekt via effekten på planterne, førnen og (indirekte) mikroklimaet. Dette underbygges af, at artsammensætningen af leddyrene i høj grad er korreleret med plantebiomasse og førnebiomasse. Den overvejende negative effekt af herbicidbehandlingen på leddyrene stemmer overens med fx resultaterne i Reddersen m.fl. (1998). Der findes kun ganske få studier, der tidligere har beskæftiget sig med effekter af lav- herbiciddosis på vegetation sammensat af arter, der findes i (halv-) naturen og småbiotoper i agerlandet (Bicheludvalget 1999). Langt den overvejende del af pesticidforskningen har været rettet mod arter på markfladen, såvel ukrudtsarter som skadedyr og svampe. Der er lavet talrige dosis-respons studier af de forskellige midlers effekter på veldefinerede målorganismer som en del af godkendelsesprocedurer for pesticider. Derimod er denne markante effekt på dyre- og plantesamfund ikke påvist før. Endvidere har der været lavet en række studier af indirekte effekter af herbicider på leddyr og fugle i agerlandet (se review i Bicheludvalget 1999), men kun et enkelt studie (Kleijn & Snoeijing 1997) har tidligere forsøgt at kvantificere effekten af et herbicid (Starane 200; fluoroxypyr) på agerlandets kantvegetation. Kleijn & Snoeijing (1997) fandt et fald i artsrigdom med øget fluoroxypyrdosering, mens biomasse produktionen steg for græssers vedkommende og faldt for andre urter under højproduktive forhold. De fandt endvidere, at 5 og 10 % fluoroxypyr (af markdosis) reducerede biomasseproduktionen af indvandrende urter og øgede ekstinktionsraten af de tilstedeværende arter. Forsøget anvendte årlig slåning og fjernelse af biomassen, hvilket også påvirkede konkurrencen signifikant, så derfor kan resultaterne ikke direkte sammenlignes med nærværende forsøg. Eksperimentet i dette projekt har ved at tildele varierende doser af N og ensartede doser af P og K kunnet påvise en række forventede effekter af kvælstof på levende biomasse og på førne. Nitrogentilførsel alene (dvs. uden glyphosatbehandling) øgede produktiviteten af en sådan vegetation med ca. 65 %, når vegetationen som i dette tilfælde var N-begrænset. Konkurrencestrategerne (især repræsenteret ved stortoppet hvene) viste en stigende dominans med øget N-tilførsel, mens Nøjsomhedsplanterne (S-strateger især repræsenteret ved fåresvingel) viste en vigende tilstedeværelse. Ruderaterne viste ingen signifikante ændringer. Artsrigdommen for karplanter og frekvensen for mosser er højst ved 0 kg N/ha og stærkt faldende ved stigende N-tilførsel. Dette bekræfter således teorierne om tålegrænser for luftbårent kvælstof til N-begrænsede økosystemer (Bobbink m.fl. 1998). Der er næppe tvivl om at effekterne af øget N-tilførsel på de undersøgte leddyrgrupper, som jo primært består af herbivore arter, er afledt af den øgede plante- og førnebiomasse og faldende plantediversitet ved øget N (Figur 3.2 og 3.5). De fleste arter har gavn af den øgede mængde tilgængelig føde, bedre mikroklima og mere stratificerede vegetationsstruktur, der opstår ved øget vækst, mens nogle få arter kan tænkes at være negativt påvirkede i kraft af, at plantediversiteten falder. I nærværende studie sås kun et konsekvent fald for en type tæger (Scutelleridae/Pentatomidae), som lever på græs, hvilket ikke umiddelbart kan forklares af ovenstående argumentation. Den overvejende positive effekt i dette eksperiment kunne meget vel hænge sammen med, at vegetationen endnu ikke var fuldt udviklet, og biomassetilvæksten derfor har været den mest betydende faktor for leddyrene. En sådan effekt stemmer overens med resultaterne i Esbjerg & Petersen (2002), som fandt en positiv korrelation mellem plantebiomasse og leddyrbiomasse for visse herbivore grupper. Man kan sagtens forestille sig, at den forarmning af floraen, der sker ved længere tids N-påvirkning, på lang sigt vil resultere i en tilsvarende forarmning af leddyrfaunaen. Andre studier (Koricheva m.fl. 2000, Siemann 1998 og Haddad m.fl. 2000) tyder imidlertid på, at kun meget stationære leddyr vil påvirkes af en lokal forarmning af floraen som følge af fx N-tilførsel. I et studie af kvik-domineret flora sammenlignet med en artsrig ”naturlig” flora fandt Lagerlöf & Wallin (1993), at der godt nok var flere leddyrarter i plots med ”naturlig” flora, men kvik viste sig også at have en stor betydning som skjul for mange leddyrgrupper, primært rovlevende insekter. Størrelsen af afdriften af ammoniak fra markflader og fejlplaceret gødskning er meget dårligt dokumenteret, hvorimod effekten af gødskning er undersøgt meget grundigt i en lang række undersøgelser (se reviews af Waide m.fl. 1999, Bobbink m.fl. 1998). De fleste er enige, om at produktivitet påvirker diversitet, men de underliggende mekanismer og mønstre er man ikke enige om (Waide m.fl. 1999). En stor del af undersøgelserne opererer med NPK-gødskning, mens forskning i N-tilførsel alene hovedsageligt er lavet på N-følsom vegetation i kvælstofbegrænsede økosystemer (højmoser, heder, skove etc.) i relation til at fastsætte tålegrænser for N-tilførsel fra luften, (Bak m.fl. 1999). Kleijn & Snoeijing (1997) har i lighed med dette projekt undersøgt kombinationer af herbicid (fluoroxypyr, 0-50 % markdosis) med produktivitet (0, 25 og 50 % af markdosis NPK-tilførsel), hvor resultaterne som forventet gav meget signifikante udslag med øget biomasseproduktion og faldende artsdiversitet (specielt lave arter blev udkonkurreret) med øget produktivitet. Effekten her kan dog ikke umiddelbart sammenlignes med nærværende forsøg, da der er tale om et NPK-forsøg. 3.3.2.1 Funktionel fortolkning af resultaterne Resultaterne af glyphosatbehandlingen kan derudover tolkes som en forstyrrelse af økosystemet, der også har forskellig virkning på de forskellige plantestrategier. Forstyrrelser (fx. græsning, brand, slåning, herbicider) har oftest været undersøgt som enten-eller begivenheder, og en del undersøgelser påpeger, at mange halv-naturlige økosystemer har behov for forstyrrelser for at kunne opretholde artsdiversiteten (Mackey & Currie 2000). En eng har behov for græsning og/eller høslet for at opretholde sin struktur og funktion og de tilknyttede og tilpassede arter. Mangel på forstyrrelser går især ud over rosetplanter (typisk nøjsomhedsplanter) og ruderater samt mosser i græsland (Jacquemyn m.fl. 2003). Nogle typer af forstyrrelser forbedrer med andre ord vilkårene for visse plantearter og andre forringer mulighederne (Jutila & Grace 2002). Hvis man opfatter glyphosatbehandlingen som en forstyrrelse, kan resultatet af 25 % markdosis forklares med forbedrede konkurrencevilkår for en relativ glyphosattolerant nøjsomhedsplante (fåresvingel,) idet konkurrenceplanterne ikke kan klare forstyrrelsen. Resultaterne af den eksperimentelle undersøgelse viser, at indvandring (med mosser som eksempel) er størst ved lave N-niveauer pga. lav biomasse (og dækning) af vegetationen. Derimod har mange planter som udgangspunkt svært ved at etablere sig ved høj N-tilførsel pga. konkurrencen fra C-strategerne, som imidlertid viser sig mere følsomme over for glyphosat i eksperimentet. Derved får mosserne muligheden for at etablere sig ved moderate herbiciddoser. Ved 5 % glyphosat-dosis var effekten af kvælstofgødskning markant, således at andelen af C- og R-strateger stiger ved øget N-tilførsel , mens S-strategernes andel falder (Figur 3.2). Med andre ord: S-strateger udkonkurrerede altså C- og til dels R-strateger ved høj herbicidbelastning ved såvel lav som høj N-tilførsel. Vi har i denne rapport valgt at fokusere på år 3 (det sidste projektår), hvilket under alle omstændigheder kun er et billede i en succession, som kræver yderligere opfølgning, fx efter 5 og 10 år, specielt hvis resultaterne af eksperimentet skal sammenholdes med hegnsundersøgelsen (se kap. 4). Endvidere kunne det være særdeles interessant at indlægge en yderligere variabel, fx slåning i eksperimentet idet én af mulighederne for at forvalte eutrofieret vegetation er slåning og fjernelse af biomasse. Dette kan gælde for vegetationen i enge såvel som for vejkants- og hegnsfodposevegetation. 3.3.3 HypotesenePå baggrund af resultaterne af eksperimentet kan hypotesen om, at herbicidafdriften vil påvirke diversitet og dominans af en græslandsvegetation og de tilknyttede leddyr, bekræftes med den modifikation, at især 25 % af markdosis glyphosat gav de forventede effekter. I de fleste tilfælde var der ingen signifikant effekt af 1 % dosis, hvorimod 5 % fx gav signifikante ændringer i artsdominansen. Hypotese 2 vedrørende reduktion i artsantallet som følge af ammoniakafdrift simuleret ved N-tilførsel blev delvist bekræftet af experimentet. N-tilførslen reducerede artsantallet af en græslandsvegetation ved at øge dominansen af flerårige, konkurrencestærke plantearter på bekostning af mangfoldighed af planter og leddyr. Som forventet var denne reaktion stærkest hos planterne, da leddyrene formentlig reagerede på planternes vækst og tilstedeværelse. Eksperimentets hypotese H3, der forudsiger interaktioner mellem herbicid- og kvælstofpåvirkning, blev fuldt ud bekræftet af eksperimentet. 4 Sammenfattende diskussion og konklusioner4.1 Sammenfatning af resultater fra hegnsundersøgelser og eksperiment 4.1 Sammenfatning af resultater fra hegnsundersøgelser og eksperimentI hegnene fandt vi, at driftsformen påvirkede artsantallet og artssammensætningen af planterne i hegns fodpose (Tabel 4.1) med en lille overvægt af konkurrencestærke arter i de konventionelle hegn, men flere natur- og agerlandsarter i de økologiske hegn. Vi kunne derimod ikke med sikkerhed sige, om der var flere herbicidfølsomme plantearter eller flere skovarter i de økologiske hegn, idet der var for få arter med sikker viden om herbicidfølsomheden og for få skovarter i de unge hegn. Dog havde de økologiske hegn en større lighed med skovbryn end de konventionelle hegn. Tabel 4.1. Maksimale forskelle i antal plantearter, antal leddyrarter og tætheden af leddyr i hhv. hegnsstudiet (forskellen mellem økologisk og konventionel i forhold til konventionel) og eksperimentet (effekter af stigende kvælstof- og glyphosattilførsel).
Eksperimentet har klart påvist interaktioner mellem kvælstof- og glyphosatpåvirkningen for plantebiomasse, førne, karplantediversitet, mosfrekvenser og dominansen af konkurrencestærke plantearter. For plantebiomasse og førne var der en meget markant reduktion ved 25 og 100 kg N i kombination med 25 % glyphosat, men denne effekt udeblev ved 0 kg N. Det skyldes sandsynligvis, at fåresvingel var relativt glyphosat-tolerant, og at den på trods af 25 % markdosis opnåede en rimelig biomasse og dominerede felterne. For mosserne var der ligeledes en markant interaktion, idet frekvensen ved høje kvælstofniveauer var klart højere ved 25 % glyphosat end ved lavere doser, hvilket var omvendt ved lavere kvælstofdoser (Figur 3.2). Mht. artsrigdommen af planter sås en negativ effekt af glyphosat ved 0 kg N, men en positiv effekt ved 100 kg N. Afdriften har i forsøg vist sig at ligge på omkring 1-16 % af markdosis i en afstand af 1 m fra den yderste dyse (Marrs m.fl. 1989, Hald m.fl. 1988), så mængderne anvendt i dette forsøg synes umiddelbart realistiske i forhold til forventelig afdrift til hegnene. Miljøstyrelsen (1999) angiver, at man i en tysk undersøgelse (Ganzelmeier m.fl. 1995) har påvist afdrift på 1,8 % af markdosis ved 2 m afstand til afgrøden, faldende til 0,1 % i 20 m afstand. Disse resultater baserer sig imidlertid på forsøg med kun et sprøjtespor. Nordby & Skuterud (1975) fandt, at ved sprøjtning i flere spor, som det foregår ved normal anvendelse af herbicider, er afdriften til hegnene op til 3 gange så stor. Den dosis, der er anvendt som udgangspunkt for 'afdriftsdoserne', var den anbefalede ifølge producentens vejledning (1440 g a.i./ha), mens man i PC planteværn (www.lr.dk) angiver den højest anbefalede dosering i stub til 900 g a.i./ha. Dette tyder på, at 'afdriftsprocenterne' anvendt i eksperimentet snarere var 1,6 %; 8 % og 40 % i forhold til nuværende praksis, og dermed var den højeste anvendte glyphosatdosis måske høj i forhold til forventelig afdrift. Den høje afdriftsprocent blev som udgangspunkt valgt for at 'sikre' en klar glyphosateffekt på vegetationen, hvilket også var tilfældet. På trods af de forskelle mellem forholdene i hegnene og i eksperimentet, som allerede er nævnt (afsnit 3.3), kan den udvikling i flora og fauna, som er belyst via eksperimentet, i nogen grad simulere, hvad der sker i nyetablerede hegn, når der sprøjtes med glyphosat. Formentlig gælder dette også for ukrudtssprøjtning mere generelt, selv om der er store forskelle i sprøjtemidlers effekter og plantearters følsomhed over for ukrudtsmidler. Desværre blev nogle vidt udbredte hegnsarter såsom kvik, stor nælde, agertidsel og gråbynke kun sporadisk etableret i eksperimentet. Disse arter er konkurrencestrateger med stor vægt på vegetativ spredning og derfor vanskeligere at etablere fra frø. Derfor bliver sammenligningen med hegn mindre ligetil, idet de enkelte arters specifikke reaktion på glyphosat kan være ret afgørende for eksperimentets resultater. I princippet vil de samme plantekonkurrencestrategier kunne være repræsenteret i hegnene, men der er ingen tvivl om at den eksperimentelle vegetation som udgangspunkt har en klar overvægt af nøjsomhedsplanter og mere eller mindre sjældne planter ift. de undersøgte hegn. Disse arter kan potentielt findes i hegnene, men vil ofte være blevet udkonkurreret pga. de eutrofe forhold. Det var ikke muligt at bekræfte artsforskellene i hegnene eksperimentelt. Det skyldes formentlig en række faktorer. Ved 100 kg N, som formodes at være sammenligneligt med eutrofieringsniveauet i de undersøgte hegn, fandt vi i eksperimentet en positiv effekt af glyphosatanvendelsen på artsantallet, således at artsantallet nåede op på samme niveau som ved 0 kg N/ha. Her skal man dog tage i betragtning, at udviklingen i artsantal og artssammensætning i eksperimentet formentlig kun afspejler den tilsvarende udvikling i de første 3 år af et hegns levetid, hvilket ikke er repræsentativt for situationen 10-15 år senere (dvs. på det tidspunkt, vi undersøgte hegnene). En anden mulighed er, at det ikke udelukkende er fraværet af pesticidafdrift på økologiske bedrifter, som forårsager den højere artsdiversitet. Det kunne være forskelle i artspuljer og måske landskabelige forhold (afstande til halvnatur), der influerer. Der er igangsat forskning for at afdække, om økologiske bedrifter er lokaliseret anderledes i landskabet og/eller indeholder flere småbiotoper. Det skal desuden understreges, at de konventionelle hegn formentlig gennem de 2-3 første leveår har fået tilført herbicid svarende til 100 % markdosis af herbicid (evt. glyphosat) for at holde ukrudtet væk, mens hegnet voksede til. På den anden side ville en økologisk landmand tilsvarende have udført mekanisk rensning flere gange i løbet hegnets første år for at holde ukrudtet i ave. Mekanisk renholdelse anvendes i såvel økologiske som konventionelle hegn, hvilket også påvirker bundvegetationens etablering og udvikling, omend ikke nødvendigvis i samme grad og retning som herbicidbehandlingen. Det er endvidere ikke ualmindeligt, at konventionelle mælkeproducenter anvender 100 % herbiciddosis under el-hegnet (fx langs hegn) for at undgå uønsket el-afledning af høj vegetation. Ud over effekter på den vegetative del af planten er det velkendt, at subletale doser kan have afgørende effekt på planters frøproduktion (referencer i Bicheludvalget 1999) og dermed påvirke arters indbyrdes konkurrence. Vi kan ikke afvise, at en del af de ændrede konkurrenceforhold mellem arterne under de forskellige driftsformer skyldes sådanne effekter, som på langt sigt kan føre til uddøen af visse arter på lokaliteten. Forskellige herbicider har forskellig virkning på (non-target) organismer, og glyphosat som modelherbicid i eksperimentet kan kun give et groft fingerpeg. Hvis man fortolker herbicidtilførslen som en forstyrrelse, kunne man have forventet at R-strategerne generelt ville klare sig bedre med højere herbicidtilførsel i eksperimentet, men det kunne ikke bekræftes. Herbicidtilførslen bør måske snarere fortolkes som et (økotoksikologisk) stress, hvor netop de stress-tolerante bliver favoriseret af højt glyphosatniveau. Et billede der svarer til resultaterne på kobberforurenet jord, hvor stresstolerante arter kommer til at dominere (Strandsby & Axelsen 1998). For leddyrenes vedkommende var tætheden og antallet af arter nogenlunde sammenlignelige i hegnene og eksperimentet. Artssammensætning var imidlertid noget forskellig i hegn og eksperiment. Tæger var således langt mere talrige i eksperimentet end i hegnene, og antallet af cikader var sammenligneligt, mens der for rovbiller, løbebiller, Clavicornia, jordlopper og snudebiller var langt flere dyr per prøve i hegnene end i eksperimentet. Resultaterne fra eksperimentet kan på den baggrund ikke direkte overføres til forholdene i hegnene, men de mekanismer, der kobler flora og fauna, må formodes at være de samme i de to typer lokaliteter, idet leddyrfaunaen i eksperimentet formentlig ikke er væsentlig forskellig fra faunaen i et nytilplantet hegn. I eksperimentet var der en reduktion i antal dyr og antal taxa som følge af glyphosatbehandlingen (Tabel 4.1), mens både antal dyr og taxa steg som følge af N-tilførsel. Tætheden og diversiteten af leddyr fulgte i store træk mønstret for plantebiomasse. Der var således flere dyr og flere arter ved stigende plante- og førnebiomasse, formentlig som følge af større fødemængde eller forbedrede mikroklimatiske forhold, mens plantediversiteten, som faldt ved stigende kvæstofniveau, ikke syntes at have nogen stor betydning for de fleste dyregrupper. For de herbivore leddyr var der i hegnenes fodposer en signifikant forskel i artssammensætningen på sandjord, mens der på lerjord og i det samlede datasæt derudover, lige som for planternes vedkommende, var signifikant flere arter i de økologiske hegn end i de konventionelle (Tabel 4.1). Der var altså tilsyneladende en forskellig sammenhæng mellem antal arter af planter og leddyr i eksperimentet og i hegnene. Dette hænger formentlig sammen med, at vegetationen i eksperimentet stadig var under etablering (plantedækket var ikke 100 %). Man kan således forvente, at der med tiden vil være en positiv sammenhæng mellem antallet af plantearter og antallet af leddyrarter, hvilket stemmer overens med, at glyphosatdoseringen og antallet af plantearter forklarede leddyrenes artssammensætning bedre en kvælstofniveauet. Den relativt lille forskel mellem økologiske og konventionelle hegn i artsantallet af leddyr hænger formentlig sammen med, at fodposens flora i begge dyrkningssystemer domineres af græsser og ikke indeholder mange urter, som vil kunne tiltrække mere specialiserede herbivore leddyr. Indikatorplanterne i hhv. økologiske og konventionelle hegn vil dog, som beskrevet i 2.3.3.1., potentielt kunne være værter for specialiserede herbivore insekter. Man kan man således ikke udelukke, at enkelte specialister vil kunne etablere sig, hvis netop deres værtsplante får plads i fodposen. I og med at der er en stor udveksling af dyr mellem hegn og mark, og mange marklevende leddyr overvintrer i hegnene (Hald m.fl. 1998), vil en ændring af levevilkårene for leddyrene i hegnene potentielt kunne have en stor betydning for agerlandets leddyrfauna. Desuden tyder resultaterne fra eksperimentet på, at regulering af gødskningen af hegn vil kunne have lige så stor gavnlig effekt på leddyrenes diversitet som fraværet af herbicider. 4.2 KonklusionerI forhold til projektets indledende hypoteser er der opnået en række markante resultater som har praktiske implikationer. Et væsentligt resultat af dette studie er, at vi ud over de forventede effekter af N og Glyphosat hver for sig kan konstatere signifikant interaktion af N-tilførsel og glyphosat, som ikke tidligere har været påvist nationalt eller internationalt (Bicheludvalget 1999, Kleijn & Snoeijing 1997). Nærværende studie har klart bekræftet en formodet forskel i floraen mellem hegn på økologiske og konventionelle bedrifter, som dog aldrig tidligere har været klart dokumenteret. Data indsamlet i nærværende hegnsstudier har godtgjort, at en række af de arter der fandtes signifikant mere i økologiske hegn er arter der er konstateret i tilbagegang i britiske overvågningsdata for hegn (Aude m.fl. 2003b, Bunce m.fl. 1999). Det tyder altså på, at økologiske hegn kan fungere som refugier og som levesteder for truede plantearter i agerlandet (Aude m.fl. 2003, Tybirk m.fl. 2003), og at agerlandets småbiotoper får en højere kvalitet ved at omlægge til økologisk jordbrug. Den samme effekt kan formentlig opnås ved at reducere herbicidafdriften væsentlig ved at etablere sprøjtefrie randzoner. Da den største forskel mellem de to hegnstyper er fraværet af kemisk ukrudtsbekæmpelse i de økologiske hegn, skyldes forskellene i floraens artssammensætning primært denne variabel. Den interaktive effekt af herbicid og kvælstof fundet i eksperimentet viser imidlertid, at gødningstilførsel til hegnene påvirker floraen i lige så høj grad som herbicidafdriften ved de anvendte N- og glyphosatbehandlinger, som ikke er urealistiske for forholdene i hegnene, og at glyphosateffekten er større ved øget N-tilførsel. Effekten af næringsstoftilførsel vil således være forskellig på økologiske og konventionelle bedrifter, selv om gødningsniveauet er det samme, og en kombination af sprøjtefrie randzoner med slåning eller gødningsfrie zoner vil optimere den positive effekt på hegnsflora og -leddyr. Tidligere danske studier af sprøjtefrie randzoner har antydet, at disse kunne have en betydning for vegetationen og leddyrfaunaen i hegn og skel (Hald m.fl. 1994). Hald og kolleger kunne dog kun svagt påvise en forskel mellem hegn langs sprøjtede og usprøjtede randzoner. En af forklaringerne var økologisk inerti i eksisterende vegetation og at studiet kun forløb gennem et kortere tidsrum (4 år) samt at hegnene, der blev undersøgt, ikke var udvalgt pga. ensartethed. Udgangspunktet for hegnsstudierne i dette projekt har været en generel opfattelse af, at der er stor økologisk inerti (Reddersen m.fl. 1999, Bicheludvalget 1999, Tybirk 2002) mht. ændringer i plantesamfund i agerlandet. Inertien forventes fordi biotoper, der er tilført store næringsmængder, ikke på kort sigt vil kunne reducere fertiliteten. Derved vil konkurrenceplanter have en selektiv fordel. Samtidig har mange (plante)arter (specielt nøjsomhedsarterne) ikke noget stort spredningspotentiale, og derfor vil denne gruppe have meget svært ved at etablere sig i fx hegn efter reduktion af næringsstof- og pesticidtilførslen. Dette studie dokumenterer, at på 10-15 år kan man opnå en signifikant øget diversitet i hegn ved at hegn drives økologisk, og det tidligere omtalte studie (Petersen 2003) tyder på, at en omlægning til økologisk driftsform ganske hurtigt vil kunne give positive resultater for vegetationen og fluefaunaen. Vi er på den baggrund måske nødt til at revurdere tesen om, at der generelt er stor økologisk inerti i agerlandets hegn og småbiotoper. Vi kan i hvert fald konstatere, at pesticidfravær giver nogle markante og hurtige ændringer af plantesamfund og til dels leddyrssamfund. Det er dog klart, at næringsstofferne ikke kan fjernes umiddelbart, så den yderligere positive effekt, som kan forventes ved et sænket N-niveau i hegnene, vil tage tid at opnå. Sammenligner man dette projekts resultater med internationale studier på området kan det således slås fast at
I modsætning til den almindelige antagelse om økologisk inerti viser studiet, at herbicidfravær hurtigt giver signifikante, positive ændringer af diversiteten i hegn.
4.3 PerspektiveringPå baggrund af studiets resultater vil vi anbefale, at man overvejer at inddrage effekter på vegetation og fauna i marknære habitater i godkendelsesprocedurer for sprøjtemidler. Derudover giver resultaterne anledning til en række anbefalinger for praktisk landbrugsdrift og yderligere videnopbygning. Driftsform Randzoner og tekniske foranstaltninger Eksperimentet i nærværende rapport viser, at en reduktion af N-indholdet og/eller N-tilførslen kan give yderligere fordele for biodiversiteten i hegnene. Derfor kan det anbefales, at der i de udlagte SFL-områder ansøges om både sprøjte- og gødningsfrie randzoner til gavn for naturkvaliteten og mangfoldigheden af potentielle fødekæder i agerlandet. Andre dyrkningsmæssige tiltag, der reducerer tilførslen vil selvsagt have samme effekt. Derudover vil tiltag, der fjerner den tilførte næring, fx slåning og efterfølgende fjernelse af vegetationen, på længere sigt reducere næringsstofniveauet og dermed fremme den biologiske mangfoldighed. Af andre realistiske muligheder kan nævnes udlægning af brak i striber på 10 m langs hegn og andre småbiotoper, som der netop er åbnet mulighed for fra 2004 (www.lr.dk). Dette vil medføre reduktion af både herbicid- og gødningspåvirkning. Videnbehov For at kunne undersøge om hypotesen om økologisk inerti skal revurderes, vil længere tidsserier af studier såvel eksperimentelt som i agerlandets småbiotoper (fx værdien af gamle hegn (økologiske og konventionelle) for biodiversitetsbevarelse) være nødvendige. Flere og mere detaljerede studier af naturindholdet på økologiske og konventionelle bedrifter i relation til naturbeskyttelsesinteresser omfattet af Habitatdirektivet vil være nødvendige for at sikre gunstig bevaringsstatus for de udpegede naturtyper og arter i NATURA 2000-områder. 5 LitteraturAgger, P. and Brandt, J. 1988. Dynamics of small Biotopes in Danish agricultural landscapes. Landscape Ecology 1: 227-240. Agger, P., Andersen, K.B., Primdahl, J. & Petersen, E.H. 1999. Natur og Landbrug. Temarapport nr. 1/1999, Naturrådet. Agger, P., Christensen, P., Reenberg, A. Aaby, B. 2002. Det fede landskab – landbrugets næringsstoffer og naturens tålegrænser. Vismandsrapport 2002. Naturrådet. Ahnström, J. 2002. Ekologisk lantbruk och biologisk mångfald - en litteraturgenomgång. SLU, Sveriges Lantbruksuniversitet, Uppsala. Allen, S.E. 1989. Chemical analysis of ecological material, 2nd ed. Blackwell Scientific Publications, Oxford. Andersen, A. G., Boesen, D. F., Holmen, K., Jacobsen, N., Lewinsky, J., Mogensen, G., Rasmussen, K., & Rasmussen, L. 1976. Den Danske Mosflora 1. Bladmosser Gyldendal, København. Andreasen, C., Stryhn, H., & Streibig, J. C. 1996. Decline of the flora in Danish arable fields. Journal of Applied Ecology 33, 619-626. Aude, E., Tybirk, K., & Pedersen, M. B. 2003. Vegetation and diversity of conventional and organic hedgerows. Agriculture, Ecosystems & Environment 99(1-3): 135-147. Aude, E., Tybirk, K., Michelsen, A., Ejrnæs, R., Hald, A.B., Mark, S. In press. Conservation value of spontaneous vegetation in hedgerows – does organic farming make a difference? Biological Conservation. Bak, J., Tybirk, K., Gundersen, P. Asman, W.A.H., Jensen, J.P. & Conley, D. 1999. Natur- og miljøeffekter af ammoniak. Ammoniakfordampning – redegørelse nr. 3. DJF/DMU rapport, 66 pp. Barr, C. J., Britt, C. P., Sparks, T. H. 1995. Hedgerow management and wildlife. A review of research on the effects of hedgerow management and adjacent land on biodiversity. Institute of Terrestrial Ecology and ADAS. 125 pp. Baundry, J., Bunce, R. G. H., & Burel, F. 2000. Hedgerows: An international perspective on their origin, function and management. Journal of Environmental Management 60, 7-22. Bicheludvalget 1999. Rapport om Miljø og sundhed. Miljø- og Energimisteriet. Boatman, N. D., Blake, K. A., Aebisher, N. J., & Sotherton, N. W.1994. Factors affecting the herbaceous flora of hedgerows on arable farms and its value as wildlife habitat. In Hedgerow management and nature conservation, eds. T. A. Watt & G. P. Buckley, pp. 33-46. Wye College Press, London. Bobbink, R., Hornung,M. & Roelofs, J.G.M. 1998. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. Journal of Ecology 86: 717-738. Borchard, C.H., Legendre, P. & Drapeau, P., 1992. Partialling out the spatial component of ecological variation. Ecology 73(3): 1045-1055. Boutin, C. & Jobin, B. 1998. Intensity of agricultural practices and effects on adjacent habitats. Ecological Applications 8: 544-557. Boutin, C. & Rogers, C. A. 2000. Pattern of Sensitivity of Plant Species to Various Herbicides - an Analysis With Two Databases. Ecotoxicology 9: 255-271. Boutin, C. Elmegaard, N. & Kjær, C. in prep. Patterns of terrestrial plant sensitivities to six herbicides with different modes of action. Boutin, C., Jobin, B., Belanger, L., Baril, A. and Freeemark, K. 2001. Hedgerows in the farming landscape of Canada. In: Barr, C. J. and Petit, S. (eds.), Hedgerows of the world. IALE(UK), pp. 33-42. Braae L., Nøhr H., Petersen B.S. 1988. Fuglefaunen på konventionelle og økologiske landbrug. Sammenlignende undersøgelser af fuglefaunaen, herunder virkninger af bekæmpelsesmidler. Miljøprojekt nr. 102, Miljøstyrelsen. Bray R.J. and Curtis J.T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs 27: 325-349. Bunce, R. G. H., Smart, S. M., van de Poll, H. M., Watkins, J. W., & Scott, W. A. 1999, Measuring change in British vegetation. Centre for Ecology and Hydrology & Natural Environment Research Council, Cumbria, 2. Chiverton, P. A. & N. W. Sotherton 1991. The Effects on Beneficial Arthropods of the Exclusion of Herbicides from Cereal Crop Edges. J. Appl. Ecol. 28: 1027-1039. Chiverton, P.A. 1999. The benefits of unsprayed cereal crop margins to grey partridges Perdix perdix and pheasants Phasianus colchius in Sweden. Wildlife Biology 5:2, 83-92. Clarke KR and Ainsworth M. 1993. A method of linking multivariate community structure to environmental variables. Marine Ecology Progress Series 92: 205-219. Clarke KR and Green RH. 1988. Statistical design and analysis for a `biological effects' study. Marine Ecology Progress Series 46: 213-226. Clarke KR. 1993. Non-parametric multivariate analyses of changes on community structure. Australian Journal of Ecology 18:117-143. Cummins, R. P. & French, D. D.1994. Floristic diversity, management and associated land use in British hedgerows. In: Hedgerow management and nature conservation, eds. T. A. Watt & G. P. Buckley, pp. 95-106. Wye College Press, London. Cunha, A., Power, S.A., Ashmore, M.R., Green, P.R.S., Haworth, B.J. & Bobbink, R. 2002. Whole Ecosystem Nitrogen manupulation: asn update review. JNCC Report 331 (www.jncc.gov.uk). de Snoo, G. R. 1999. Unsprayed field margins: effects on environment, biodiversity and agricultural practice. Landscape and Urban Planning 46: 151-160. Dietrick, E.J. 1961. Animproved backpack motorised fan for suction sampling af insect populations. Journal of Economic Entomology 54(2): 394-395. DFFE 2003. Hektarstøtte og anmendelse af foderarealer. Direktoratet for Fødevareerhverv, København. Dufrene, M. & Legendre, P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67: 345-366. Ejrnæs, R. 2000. Agerlandets Natur. I: Holten-Andersen, J., Christensen, H.S., Pedersen, T.N. & Manninen, S. (red.), Dansk Naturpolitik. Viden og vurderinger, s. 218-231. Naturrådet. Ejrnæs, R., Aude, E., Nygaard, B., & Münier, B. 2002. Prediction of conservation value using ordination and neural networks. Ecological Application 12: 1180-1187. Ejrnæs, R., Bruun, H. H. 2000. Gradient analysis of dry grassland vegetation in Denmark. Journal of Vegetation Science 11, 573-584. Ellenberg, H., Weber, H. E., Düll, R., Wirth, V., Werner, W., & Paulissen, D. 1991. Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica XVIII. Esbjerg, P. & Petersen, B.S. 2002. Effects of reduced pesticide use on flora and fauna in the agricultural fields. Bekæmpelsesmiddelforskning fra Miljøstyrelsen Nr. 58 2002. Feber, R. E., Bell, J., Johnson, P. J., Firbank, L. G., & Macdonald, D. W. 1998. The Effects of Organic Farming on Surface-Active Spider (Araneae) Assemblages in Wheat in Southern England, UK. Journal of Arachnology 26: 190-202. Fletcher, J. S., Pfleeger, T. G., Ratsch, H. C. and Hayes, R. 1996. Potential impact of low levels of chlorsulfuron and other herbicides on growth and yield of nontarget plants. - Environmental Toxicology and Chemistry 15: 1189-1196. Ganzelmeier, H., Rautmann, D. Spangenberg, R., Streloke, M, Herrmann, M. Wenzelburger, H.-J. & Walter, H.-F. 1995. Studies on the spray drift of plant protection products. Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft, Berlin-Dahlem. Blackwell Wissenschafts-Verlag, Berlin. Gaun, S. 1974. Blomstertæger. Danmarks Fauna. GEC Gad, København. Grime, J. P. 1979. Plant strategies & vegetation processes John Wiley & Sons. Grime, J. P. 1988. Comparative Plant Ecology. A functional approach to common British species Unwin Hyman, London. Haddad, N.M., Haarstad, J. & Tilman, D. 2000. The effects of long-term nitrogen loading on grassland insect communities. Oecologia 124: 73-84. Hald A.B., Reddersen J. 1990. Fugleføde i kornmarker – insekter og vilde planter. Undersøgelser på konventionelle og økologiske landbrug 1987-88. Miljøprojekt nr. 125, Miljøstyrelsen. Hald, A. B. , Nielsen, B.O., Samsøe-Petersen, L., Hansen, K., Elmegaard, N & Kjølholt, J. 1988, Sprøjtefri randzoner i kornmarker. Miljøprojekt nr. 103, Miljøstyrelsen. Hald, A. B. 1998, Sustainable agriculture and nature values - using Vejle county as study area. Faglig Rapport fra DMU nr. 222. Hald, A. B. 1999. Weed vegetation (wild flora) of long established organic versus conventional cereal fields in Denmark. Annals of Applied Biology 134: 307-314. Hald, A.B., Pontoppidan, H., Reddersen, J. & Elbek-Pedersen, H. 1994. Sprøjtefrie randzoner i sædskiftemarker. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 6. Hansen, K. 1993. Dansk feltflora. Gyldendalske boghandel, Nordisk Forlag, Copenhagen. Hansen, V. 1927. Biller VII. Bladbiller og Bønnebiller. Danmarks Fauna. GEC Gad, København. Hansen, V. 1951. Biller XV. Rovbiller 1. del. Danmarks Fauna. GEC Gad, København. Hansen, V. 1952. Biller XVI. Rovbiller 2. del. Danmarks Fauna. GEC Gad, København. Hansen, V. 1954. Biller XVII. Rovbiller 3. del. Danmarks Fauna. GEC Gad, København. Hansen, V. 1965. Biller XXI. Snudebiller. Danmarks Fauna. GEC Gad, København. Hansen, V. 1968. Biller XXIV. Sandspringere og Løbebiller. Danmarks Fauna. GEC Gad, København. Hegarty, C. A., McAdam, J. H., & Cooper, A. 1994. Factors influencing the plant species composition of hedges - implications for management in environmentally sensitive areas. Boatman, N. (Ed.) Field margins: integrating agriculture and conservation. Monograph No 58, 227-234. Surrey, BCPC Publications. Hill, M. O. 1979. DECORANA - A FORTRAN program for detrended correspondence analysis and reciprocal averaging. Ithaca, New York, Cornell University. Hill, M.O. & Gauch, H.G. 1980. Detrended correspondance analysis: an improved ordination technique. Vegetatio 42: 47-58. Hubbard, C. E. 1984. Grasses. A guide to their structure, identification, uses and distribution in the British Isles Penguin Books. Jacquemyn, H., Brys, R. & Hermy, M. 2003. Short-term effects of different management regimes on the response of calcareous grassland vegetation to increased nitrogen. Biological Conservation, 111: 137-147. Jensen, A.-M. M. & Johnsen, I. 2003. Wild Flora. I: Navntoft, S., Esbjerg, P., Jensen, A.M. M., Johnsen, I. & Petersen, B.S. 2003. Flora and fauna changes during conversion from conventional to organic farming. Pesticides Research 74, pp 17-31. Jensen-Haarup, AC. 1912. Tæger. Danmarks Fauna. GEC Gad, København. Jongman, R.H.G., ter Braak, C.J.F. & van Tonbgeren, O.F.R. 1987. Data analysis in community and landscpae ecology. Pudoc. Wageningen. Kedrowski, R.A. 1983. Extraction and analysis of nitrogen, phosphorous and carbon fractions in plant material. Journal of Plant Nutrition 6: 989-1011. Kleijn, D. & Snoeijing, I. J. 1997. Field boundary vegetation and the effect of agrochemical drift: botanical change caused by low levels of herbicide and fertilizer. Journal of Applied Ecology 34: 1413-1425. Kleijn, D. & Verbeek, M. 2000. Factors affecting the species composition of arable field boundary vegetation. Journal of Applied Ecology 37: 256-266. Koricheva, J., Mulder, C.P.H., Schmid, B., Joshi, J. & Huss-Danell, K. 2000. Numerical responses of different trophic groups of invertebrates to manipulation of plant diversity in grasslands. Oecologia 125, 271-282. Kristensen, I.S. Halberg, N Nielsen,A.H. Dalgaard, R. og Hutchings, N. 2003 Agricultural Nutrient management in Denmark with special reference to intensive dairy farming http://www.agrsci.dk/jbs/isk/DK_country_report_partII.pdf. Kudsk, P. pers. komm. Seniorforsker, DJF. Lagerlöf, J. & Wallin, H. 1993. The abundance of arthropods along two field margins with different types of vegetation composition: an experimental study. Agriculture, Ecosystems and Environment 43: 141-154. Le Coeur, D., Baundry, J., Burel, F., & Thenail, C. 2002. Why and how we should study field boudary biodiversity in an agrarian landscape context. Agriculture, Ecosystems and Environment. 89: 23-40. LI-COR 1991. LI-COR radiation Sensor instruction manual. LI-COR, Lincoln, Nebraska. Lid, J. 1987. Norsk, svensk, finsk flora. Det Norske Samlaget, Oslo. Mackey, R.L. & Currie, D.J. 2000. A re-examination of the expected effects of disturbance on diversity. Oikos 88: 483-493. Mäder, P., Fliessbach, A., Dubois, D., Gunst, L., Fried, P., & Niggli, U. 2002. Soil Fertility and Biodiversity in Organic Farming. Science 296, 1694-1697. Marrs, R. H., Frost, A. J., Plant, R. A., & Lunnis, P. 1993. Determination of buffer zones to protect seedlings of nontarget plants from the effects of glyphosate spray drift. Agriculture Ecosystems & Environment 45: 283-293. Marrs, R.H., Williams, C. T., Frost, A. J., & Plant, R. A. 1989. Assessment of the effects of herbicide spray drift on a range of plant species of conservation interest. Environmental Pollution 59: 71-86. Marshall, J., Baudry, J., Burel, F., Joenje, W., Gerowitt, B., Paoletti, M., Thomas, G., Kleijn, D., Le Coeur, D., & Moonen, C. 2002. Field boundary habitats for wildlife, crop and environmental protection. ed. Landscape ecology in agroecosystems management, pp. 219-247. CSR Press. McAdam, J. H., Bell, A. C., & Henry, T. 1994. Field margin flora and fauna changes in response to grassland management practices. Boatman, N. (ed.) Field margins: integrating agriculture and conservation. Monograph No 58, 153-158. 1994. Surrey, BCPC Publications. McCollin, D. 2000. Editorial: Hedgerow policy and protection - changing paradigms and the conservation ethic. Journal Of Environmental Management 60: 3-6. McCune, B. and Mefford, M. J. 1999. PC-ORD for windows. Gleneden Beach, Oregon, USA, MjM Software. Mead, R. 1990. The design of experiments. Statistical principles for practical application. University Press, Cambridge. Milchunas, D. G. & Lauenroth, W. K. 1995. Inertia in plant community structure: State changes after cessation of nutrient-enrichment stress. Ecological Applications 5, 452-458. Miljøstyrelsen 1999. Rammer for vurdering af plantebeskyttelsesmidler. WWW.mst.dk. Mineau, P. & A. McLaughlin 1996. Conservation of Biodiversity Within Canadian Agricultural Landscapes - Integrating Habitat For Wildlife. Journal of Agricultural & Environmental Ethics 9(2): 93-113. Montgomery, D. 1984. Design and Analysis of Experiments. Wiley, 2nd ed., p. 209. Navntoft, S. & Esbjerg, P. 2003. Arthropods. I: Navntoft, S., Esbjerg, P., Jensen, A.M. M., Johnsen, I. & Petersen, B.S. 2003. Flora and fauna changes during conversion from conventional to organic farming. Pesticides Research 74, s. 33-50. Navntoft, S., Esbjerg, P., Jensen, A.M. M., Johnsen, I. & Petersen, B.S. 2003. Flora and fauna changes during conversion from conventional to organic farming. Pesticides Research 74. Nordby, A. % Skuterud, R. 1975. The effects of broom height, working pressure and wind speed on spray drift. Weed Research 14, 385-95. Odderskær P., Prang. A., Elmegaard N., Nørmark Andersen P. 1997. Skylark reproduction in pesticide treated and untreated fields. Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 32. Økland, R. & Eilertsen, O. 1994. Canonical Correspondence Analysis with variation partitioning: some comments and an Aplication. Journal of Vegetation Science 5: 117-126. Økland, R. 1990. Vegetation Ecology: Theory, Methods and Applications with reference to Fennoscandia. Sommerfeltia Supplement 1: 1-233. Pedersen, A. & Schou, J. C. 1988. Nordiske brombær (Rubus sect. Rubus. sect Corylifolii og sect Caesii) Aarhus University Press. AaU reports. Botanical Institute, Aarhus University. Aarhus. Petersen, S. 2003. Effekten af økologisk jordbrug på vegetation og fluefauna i læhegn og skel. Specialerapport, Københavns Universitet. Petersen, S., Axelsen, J.A., Tybirk, K., Aude, E., Vestergaard, P. In prep. Effects of duration of organic farming practice on vegetation in Danish field boundaries. To be subm. AGEE. Pinheiro, J.C. & Bates, D.M. (2000) Mixed-Effects Models in S and S-Plus. Springer, New York. Raunkjær, C. 1910. Formationsundersøgelse og formationsstatistik. Botanisk Tidsskrift 30: 20-132. Ravn, H. W. & Løkke, H. in prep. Biomarker patterns in plants exposed to glyphosate. Manus for Chemosphere. Reddersen, J, Elmholt, S, Holm, S. 1998. Indirect Effects of Fungicides and Herbicides on Arthropods. Response to treatment-induced variations in leaf fungi weeds in winter wheat 1994-1995. Bekæmpelsesmiddelforskning fra Miljøstyrelsen, no. 44. Reddersen, J., Tybirk, K., Halberg, N., & Jensen, J., 1999. Mere og bedre natur i landbrugslandet. Dokumentation for en bedre indsats. DMU faglig rapport no. 288, 109 pp. Reganold, J. P., Palmer, A. S., Lockhart, J. C., & Macgregor, A. N. 1993. Soil Quality and Financial Performance of Biodynamic and Conventional Farms in New-Zealand. Science 260: 344-349. Rydberg, N. T. & Milberg, P. 2000. A survey of weeds in organic farming in Sweden. Biological Agriculture & Horticulture 18: 175-185. SAS Institute. 1989. SAS/STAT User's Guide, Version 6 fourth edition. Cary NC, SAS Institute Inc. Siemann, E. 1998. Experimental tests of effects of plant productivity and diversity on grassland arthropod diversity. Ecolgy 79(6): 2057-2070. Smith, A. J. E. 1978. The moss flora of Britain & Ireland Cambridge University Press, Cambridge. Sokal, R.R. & Rohlf, F.J. 1995. Biometry. W. H. Freeman and Company, New York. Somerfield, P.J., Gee, J.M., Warwick, R.M. 1994. Soft sediment meiofaunal community structure in relation to a long-term heavy metal gradient in the Fal estuary system. - Marine Ecology Progress Series 105: 79-88. Sørensen, N. K. & Bülow-Olsen, A. 1994. Fælles arbejdsmetoder for jordbundsanalyser. Plantedirektoratet. Landbrugsministeriet, København. Stidsen, A.N. in prep. Vegetationen i gamle økologiske hegn. Specialestudie, Århus Universitet/DMU. Stoltze, M. and Pihl, S. Rødliste 1997 over planter og dyr i Danmark. MEM. Stolze, M., Piorr, A., Häring, A., & Dabbert, S. 2000. The environmental impact of organic farming in Europe. Dabbert, S., Lampkin, N, Michelsen, J, Nieberg, H., and Zanoli, R. (eds.) Organic farming in Europe: Economics and policy. 6, 1-125. Stuttgart-Hohenheim, Universtity of Hohenheim. Strandberg, B., Strandsby , E.B., & Axelsen, J.A. 1998. Biomonitering af forurenede grunde ved brug af planter og jordboende dyr. ATV møde, Vintermøde om grundvandsforurening, Vingstedcentret 10.-11. marts 1998. Pp. 265-274. Strandsby, E.B., & Strandberg, B. 1998. Biomonitering på tungmetal- og tjæreforurenede grunde. ATV møde, Vintermøde om grundvandsforurening, Vingstedcentret 10.-11. marts 1998. Pp. 275-287. ter Braak, C.J.F. & Smilauer, P. 1997-99. CANOCO for Windows Version. Centre for Biometry, Wageningen, Holland. Tybirk, K. & Ejrnæs, R. 2001. Hvordan bidrager økologisk jordbrug til naturbevarelse?. I: Tybirk, K. & Alrøe, H.F. (red.) Naturkvalitet i økologisk jordbrug. FØJO/DARCOF report no 9, s. 37-54. Tybirk, K. 2002. Naturplaner: integration af naturhensyn i landbruget. Flora & Fauna 108(3): 65-76. Tybirk, K., E. Aude, R. Ejrnæs, J. Reddersen, A.B. Hald, L. Riberholdt, C. Jönsson, J.E. Jørgensen, M. Gramstrup, P.R. Andreasen, L.R. Vind , A. Dalsgaard , N.V. Jensen, P. Odderskær & H. Sell. 2001. Botanical conservation values in Danish hedgerows. IBarr, C. & Petit, S. (eds.). Hedgerows of the World: their ecological functions in different landscapes. Proc. 2001 UK-IALE Conference, Univ. Birmingham, Pp 299-308. UK-IALE/ CEH. Wagner E. 1961. Heteroptera und Hemiptera. Die Tierwelt Mitteleuropas. Vol 4. P. Brohner, et al. (eds.) Quelle & Meyer, Leipzig. Waide, R.B., Willig, M.R., Steiner, C.F., Mittelbach, G., Gough, L., Dodson, S.I., Juday, G.P. & Parmenter, R. 1999. The Relationship Between Productivity and Species Richness. Annual Review of Ecology and Systematics 30: 257-300. Wilhjelmudvalget 2001.En rig natur i et rigt samfund. 1-68. MEM. Zobel, M., Van der Maarel, E., & Dupré, C. 1998. Species pool: the concept, its determination and significance for community restoration. Applied Vegetation Science 1: 55-66. Bilag A Beskrivelse af de undersøgte læhegnUdvælgelsen af de økologiske læhegn der opfyldte de opstillede kriterier var ikke let. Dels findes der ikke mange af slagsen, dels var de vanskelige at opspore. Derimod var det ikke vanskeligt at finde tilsvarende konventionelle hegn. De udvalgte læhegn er beskrevet i tabel 1. Generelt om hegnene kan det siges, at der ikke er nogen forskel mellem økologiske og konventionelle hegn i anlæggelsesfasen fra Hedeselskabets side med hensyn til planteafstand, planteteknik, artsvalg mv. Se nyskrevet bog om danske hegns historie, hvor det moderne koncept er beskrevet (Fritzbøger 2002). 1.1 Anlæg af læhegnKort beskrivelse og kvalitetetsspecifikationer af anlæggelse af kollektive læplantninger, etableret efter reglerne i "Lov om læhegn og tilskud til læplantning" med tilhørende bestemmelser. 1.1.1 JordarbejdeJordarbejdet udføres som en kombination af fræsning og harvning, reolpløjning til en dybde af normalt min. 50 cm og efterharvning. Hvor jordtypen gør en dybdepløjning uhensigtsmæssig (f. eks. svær lerjord) kan reolpløjning erstattes af anden dybdebearbejdelse og pløjning i mindre dybde. Plantebedet skal være gennembearbejdet og jævnt. 1.1.2 PlanteplanPlanteplanen skal være i overensstemmelse med den neden for angivne læhegnsmodel/principskitse. Plantesammensætningen skal tage hensyn til lokaliteten og det omgivne landskab. 1.1.3 PlanterI Landsforeningen De Danske Plantningsforeningers "Kvalitetskrav til læplanter" er kravet til de enkelte plantearter til læplantning beskrevet. Planterne leveres i de til læplantninger i det åbne land egnede arter og provenienser. Planterne skal leveres i henhold til den af LDDP godkendte artssammensætning, der gælder for lauget. 1.1.4 PlantningPlantningen udføres manuelt eller maskinelt med de plante- og rækkeafstande, der er aftalt for lauget, normalt en planteafstand på ca. 1,0 m og en rækkeaftsand på ca. 1,2 m. 1.1.5 EfterbedringDer efterbedres i den 3-årige vedligeholdelsesperiode, hvor manglende eller udgåede planter erstattes efter behov og med hensyn til mængde og type. Ved efterbedring bør der ikke plantes nærmere naboplanter end disses højde. Hvis der mangler 1 plante, efterbedres ikke, med mindre der er stort udfald af en hovedtræart. Hvis der mangler 2 planter, efterbedres med 1 plante. Hvis der mangler 3 planter, efterbedres med 2 planter. 1.1.6 RenholdelseLæplantningerne renholdes i 3 vækstsæsoner, indtil afleveringsforretningen. Første år renholdes mekanisk/manuelt. 2. og 3. år kan der renholdes mekanisk, manuelt og/eller kemisk, ligesom biologisk nedbrydelige dækmaterialer kan anvendes. Ved økologiske avlere anvendes ikke kemisk ukrudtsbekæmpelse. Ukrudtsvegetationen må derfor aldrig nå en størrelse og/eller tæthed, som vil vanskeliggøre mekanisk rensning. 1.1.7 Læhegnsmodel/principskitseI de tre- og flerrækkede hegn kan de enkelte rækker have forskellige funktioner, idet vindpresset og lysforholdene ikke er ens i de 3 rækker. Selv om hegnet i det følgende er lidt skematisk opdelt, må det dog aldrig glemmes, at hegnet er et organisk hele, idet et 3- og flerrækket hegn skal opfattes som et enkeltrækket med 3 eller flere rødder. Af hensyn til beskrivelsen kan det være hensigtsmæssigt at opdele hegnet således: 1.1.7.1 Vindrække(r) (vestvendt(e) række(r) Hovedformålet er at give et stabilt læ for vestenvinden/den dominerende vindretning. Arter til denne/disse række(r) skal være hårdføre, vindfaste og længelevende med mulighed for rimelig højdeudvikling. 1.1.7.2 Højderække(r) Hovedformålet er at give læhegnet en betydelig højde. Arter til denne række skal være højdegivende, længelevende og med rimelig hårdførhed og vindfasthed. 1.1.7.3 Karakterrække(r) Hovedformålet er at afrunde og stabilisere hegnet i læsiden. I denne/disse række(r) er der mulighed for indplantning af blomstrende buske og træer, ligesom det er muligt at tage egnsindividuelle hensyn, dog sådan at hegnets primære opgave som lægiver ikke sættes til. Afhængig af klimatiske forhold kan hegnet bestå af 1-3 vindrækker, 1-3 højderækker og 1-2 karakterrækker, iflg. læplantningsloven dog sammenlagt normalt ikke over 7 rækker. Principskitse for 3 rk. læplantning • = buske, ⊕ = varige bestandstræer, + = ammetræer
1.2 De undersøgte hegn1.2.1 TræartssammensætningenDer er ikke foretaget analyse af den reelle træartssammensætning i de enkelte hegn, da det er vurderet, at de strukturelle variable er langt mere betydende for urtevegetationen og den tilknyttede fauna. Der har ikke efterfølgende været muligt at få oplyst præcise planteplaner for de enkelte hegn, men ifølge Hedeselskabets lokale konsulenter er der sjældent den store variation inden for et område. Tabel A.1. Typisk sammensætning for 3-rækkede læhegn i undersøgelsesområderne omkring 1990. Der kan dog være tale om visse lokale variationer, som dog formentlig ikke betyder meget for urtevegetationens sammensætning.
1.2.2 HegnskarakteristikaTabel A.2 viser grundoplysninger for de undersøgte hegn. Udvælgelse af økologiske-konventionelle er bestræbt inden for samme postdistrikt, men i enkelte tilfælde har der været tale om nabodistrikter. Tabel A2 Grundoplysninger om de enkelte hegn. Forklaring til tabel A.2.
Bilag B Fotos af eksperiment juli 2002Tabel B1. Oversigt over behandlinger. Der er 10 gentagelser af hver behandling.
Nedenstående fotos viser eksempler på vegetationens fremtræden sommer 2002. Foto B1. Felt B1. 0% Glyphosat, 0 kg N
Foto B2. Felt B5. 0% Glyphosat, 25 kg N
Foto B3. Felt B7. 5% Glyphosat, 25 kg N
Foto B4. Felt B 11. 5% Glyphosat, 100 kg N
Foto B6. Felt B 12. 25% Glyphosat, 100 kg N
Foto B7. E7. 5% Glyphosat, 25 kg N
Foto B8. E8 25% Glyphosat, 25 kg N
Bilag C1 Leddyr sortering af D-vac prøver 2001
Bilag C2 Insektsortering, hegnsprojektet 2002Bladlus: Prøven deles i fjerdedele, hvoraf en tælles Thrips: Ditto Dipterer: Sum svirrefluer, sum bier hvepse Edderkopper: Sum jagtedderkopper, sum tæppespindere Sommerfugle: Sum voksne, sum larver Bladhvepse: Sum voksne, sum larver Cikader: Deles op i undergrupper – ved mange individer kan prøven opdeles i fjerdedele, hvoraf en tælles Bladlopper: Sum Tæger: Så vidt muligt til art, især de talrige Løbebiller: Sum små, sum store, artsnavn for de 1-3 mest almindelige Rovbiller: Sum små, sum store, artsnavn for de 1-3 mest almindelige Smeldere: Sum Blødvinger: Sum Jordlopper: Så vidt muligt til art, især de talrige Bladbiller: Så vidt muligt til art, især de talrige Snudebiller: Så vidt muligt til art, især de talrige Clavicornia: Sum mariehøns, sum glimmerbøsser, sum resten (dvs. skimmelbiller)
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||