Mikrobiologiske bekćmpelsesmidlers effekter pĺ invertebrater
2. Nontarget invertebrater som testorganismer
2.1 Regnorme
2.1.1 Nematoder og Bacillus thuringiensis
2.1.2 Nematoder og aktinomyceter
2.1.3 Regnorme og mikrobielt betingede sygdomme
2.1.4 Mikrobiologiske bekćmpelsesmidlers indirekte effekter pĺ
regnorme
2.1.5 Regnorme og Bacillus thuringiensis
2.1.6 Regnorme og aktinomyceter
2.1.7 Regnorme og Streptomyces griseoviridis
2.2 Nematoder
2.2.1 Nematoder og Bacillus thuringiensis
2.2.2 Nematoder og aktinomyceter
2.1 Regnorme
Allerede klassiske arbejder af Darwin (1881) og danskerne Müller (1878) og Bornebusch (1930) gjorde det klart, at regnorme udgřr et meget vigtigt faunaelement i mange terrestriske řkosystemer. I disse řkosystemer spiller de en betydelig rolle ved (Christensen og Mather, 1994), at
Řkologisk betydning
- pĺvirke jordens fysiske og kemiske egenskaber pĺ adskillige mĺder (beluftning, vandledningsevne, vandkapacitet, aggregatdannelse, mineralisering af nćringsstoffer);
- pĺvirke planterřdder og mikroorganismer, og dermed plantevćkst;
- udgřre en vćsentlig fřdekilde for en lang rćkke dyr, inkluderende alt fra pattedyr til insekter, og dermed overordnet have stor betydning for stof- og energikredslřb.
Med dette som udgangspunkt er det fornuftigt at betragte regnorme som en gruppe af dyr, hvorom det er vćsentligt at have et generelt kendskab og specielt i hvilken grad de kan pĺvirkes af et givent mikrobiologisk plantebeskyttelsesmiddel.
Effekt pĺ regnorme
Mikrobiologiske plantebeskyttelsesmidler kan potentielt set pĺvirke regnorme pĺ flere forskellige mĺder, sĺvel direkte som indirekte, og dermed fĺ betydelige konsekvenser for de řkosystemer, hvori de anvendes; f.eks.
- reducere antallet og biomassen af regnorme (og/eller ćndre deres artssammensćtning) og dermed pĺvirke jordens frugtbarhed i en negativ retning;
- pĺvirke regnormes vćkst og fertilitet og dermed reducere antal og biomasse;
- pĺvirke predatorer pĺ regnorme ved reduktion i regnormebiomasse.
Regnorme som testdyr
Regnorme har en rćkke egenskaber (Christensen og Mather, 1994), der gřr dem ideelle som testorganismer:
- de er almindeligt forekommende i de fleste jorde;
- de er relativt nemme at indsamle og identificere;
- de er relativt nemme at hĺndtere i laboratoriet;
- en rćkke arter kan dyrkes i kulturer;
- der findes en stor baggrundsviden om deres biologi og řkologi;
- de bliver benyttet som testorganismer overfor kemiske pesticider, hvilket betyder, at der eksisterer en erfaring i anvendelse af regnorme som testorganismer.
2.1.1 Regnormene i Danmark
Regnorme er saddelbřrsteorme (Oligochaeta), som tilhřrer ledormene (Annelida). I Danmark forekommer der almindeligt 15 arter fra 9 slćgter (Clausen, 1993); de tilhřrer alle familien Lumbricidae. Se tabel 2.1.
Řkotyper
De danske regnorme kan inddeles i tre forskellige řkologiske typer efter deres vertikale udbredelse i jordbunden (Bouché, 1972; Lee, 1985, Clausen,1993):
- de epigćiske orme, der lever i relativt uomsatte planterester, fřrnelag og jordens allerřverste centimetre;
- de anecike orme, der graver dybe gange i jorden, men primćrt sřger deres fřde pĺ overfladen;
- de endogćiske orme, der laver gangsystemer i jorden, og primćrt finder deres fřde nede i jorden.
Epigćiske regnorme
De epigćiske orme bestĺr af smĺ, hurtigt voksende arter med en kraftig oftest rřdbrun pigmentering. De har en relativt kort levetid, men en stor reproduktionsevne. Til gruppen hřrer D. octaedra, D. rubidus og L. castaneus, arter der primćrt findes i lřvansamlinger, i gřdning, i henrĺdnende trćstubbe og grene og lignende steder. Til gruppen hřrer ogsĺ E. fetida, som specielt er knyttet til kompost og desuden kan findes i mřddinger. Den epigćiske art E. tetraedra findes generelt under meget fugtige omstćndigheder.
Anecike regnorme
De anecike orme bestĺr af store, langsomtvoksende arter, der oftest er brunligt pigmenterede. De har en lang levetid, men en forholdsvis lille reproduktionsevne. Til gruppen hřrer L. terrestris og A. longa. Disse arter findes primćrt pĺ lokaliteter, hvor der tilfřres rigelige mćngder af organisk materiale til jordens overflade, f.eks. lřvskove, grćssede overdrev og enge, samt grćsplćner.
Endogćiske regnorme
De endogćiske orme er mindre ensartede end de to andre grupper. De omfatter sĺvel smĺ som relativt store arter, der dog har det fćllestrćk, at de er upigmenterede eller svagt pigmenterede. De har en temmelig lang levetid og en forholdsvis lille reproduktionsevne. Det fremgĺr af tabel 2.1 hvilke arter, der hřrer til denne gruppe.
Krav til řkosystem
Regnormenes krav til deres omgivelser hćnger naturligvis sammen med hvilken řkotype de tilhřrer. De epigćiske arter er sĺledes ret uafhćngige af fysisk/kemiske forhold i jorden, men er til gengćld afhćngige af tilstedevćrelsen af organisk materiale under nedbrydning. De to andre grupper af regnorme, som har en nćrmere kontakt med jorden, pĺvirkes derimod af jordens fysisk/kemiske forhold med forskelle imellem de enkelte arters prćferencer og tolerancer. Dette medfřrer, at forskellige typer af řkosystemer ikke blot domineres af forskellige řkotyper, men ogsĺ varierer i artssammensćtning (Lee, 1985).
Ernćringsbiologi
Regnorme opdeles ogsĺ i to forskellige typer med udgangspunkt i deres ernćringsbiologi (Lee, 1985):
- de detritivore regnorme, der primćrt konsumerer plantemateriale, som kun i nogen udstrćkning er nedbrudt;
- de geophage regnorme, der primćrt konsumerer jord og plantemateriale, der i střrre udstrćkning er nedbrudt.
Dette betyder, at de detrivore regnorme, i den danske fauna, udgřres af epigćiske og anecike arter, mens de geophage udgřres af de endogćiske arter (Clausen, 1993). Se tabel 2.1.
Dyrket jord
Traditionelt dyrket agerjord domineres af endogćiske orme, specielt er A. caliginosa almindelig, men arter som A. rosea og A. chlorotica kan ogsĺ forekomme (Clausen,1993). Pĺ nogle lokaliteter vil de to anecike arter L. terrestris og A. longa ogsĺ kunne findes (Andersen, 1987). Regnormenes antal varierer fra ganske fĺ individer til flere hundrede pr. m2 (Andersen 1983, 1987). Der findes ikke internationalt publicerede undersřgelser over regnormefaunaen i řkologisk dyrket agerjord, men det mĺ forventes, at anecike og epigćiske orme er mere almindelige end i den traditionelt dyrkede jord, og at antallet af individer er hřjere, fordi der anvendes forskellige former for organisk gřdning, og fordi jorden bearbejdes mindre.
Tabel 2.1
Almindeligt forekommende regnormearter i Danmark
Latinsk navn |
Dansk navn |
Řkotype |
Ernćringstype |
Allolobophora chlorotica |
Grřn orm |
En |
Geo |
Aporrectodea caliginosa |
Grĺ orm |
En |
Geo |
Aporrectodea longa |
Lang orm |
An |
De |
Aporrectodea rosea |
Rosa orm |
En |
Geo |
Dendrobaena octaedra |
Mosorm |
Ep |
De |
Dendrodrilus rubidus |
Stuborm |
Ep |
De |
Eisenia fetida |
Brandorm |
Ep |
De |
Eiseniella tetraedra |
Bćkorm |
Ep |
De |
Helodrillus oculatus |
Dyndorm |
En |
Geo |
Lumbricus castaneus |
Lřv regnorm |
Ep |
De |
Lumbricus festivus |
Mřrk regnorm |
Ep, An |
De |
Lumbricus rubellus |
Skov regnorm |
Ep, An |
De |
Lumbricus terrestris |
Stor regnorm |
An |
De |
Octolasion cyaneum |
Blĺ orm |
En |
Geo |
Octolasion tyrtaeum |
Mćlket orm |
En |
Geo |
Řkotyper: En: Endogćisk, An: Anecik, Ep: Epigćisk.
Ernćringstyper: Geo: Geophag; De: Detritivor.
Danske navne efter Bornebusch (1928).
I intensivt drevne gartnerier, hvor der ikke anvendes kompost eller lignende, forventer vi, at regnormefaunaen primćrt bestĺr af endogćiske arter i et relativt begrćnset antal. I dyrkningssystemer, hvor der anvendes kompost og jorddćkke, vil epigćiske og anecike regnorme have gode livbetingelser, og derfor sandsynligvis findes som betydelige populationer. I frugtplantager findes ofte en stor population af den anecike art: L. terrestris (Lee, 1985).
Overdrev og enge
Anecike og endogćiske arter er de dominerende pĺ overdrev og enge, mens artssammensćtning sandsynligvis er afhćngig af jordens fysisk-kemiske forhold. Under dyregřdning (f.eks. kokasser) findes ogsĺ epigćiske arter, samt de to overgangsformer (se tabel 2.1) L. rubellus og L. festivus (Holter 1979, 1983; Hendriksen, 1991). Individantallet og biomassen er ofte hřj.
Lřvskove
I lřvskove pĺ muldjord findes alle tre řkotyper af regnorme ofte i et stort antal med en hřj biomasse og en stor artsdiversitet. Derimod er det hovedsageligt epigćiske orme af arterne D. octaedra, D. rubidus og L. rubellus, i et begrćnset antal, der findes i lřvskove pĺ morbund (Clausen, 1993). I nĺleskove findes der et meget begrćnset antal regnorme, primćrt den epigćiske art D. octaedra (Bornebusch, 1930).
Fysiske krav
Regnormes aktivitet er bestemt af jordens temperatur og fugtighedsforhold. De fleste danske arter er aktive ved temperaturer imellem ca. 0° og 25°C, sĺ lćnge jorden er tilstrćkkelig fugtig. Dette betyder, at regnormene i danske řkosystemer primćrt er aktive i forĺrs og efterĺrsmĺnederne samt i den tidlige og sene sommertid. Hvis jorden er ved at třrre ud eller temperaturerne falder til under 0°C vil ormene enten grave sig ned i dybere liggende jordlag eller forsřge at overleve i en inaktiv tilstand (Edwards og Lofty, 1977; Andersen, 1987).
2.1.2 Regnorme og mikroorganismer
Regnorme lever i et miljř, hvor de er omgivet af et utal af mikroorganismer med en sćrdeles stor artsdiversitet. Litteraturen har hovedsagligt beskćftiget sig med mikroorganismernes interaktioner med regnormenes fordřjelsessystem. Figur 2.1 anskueliggřr de forskellige typer af interaktioner.
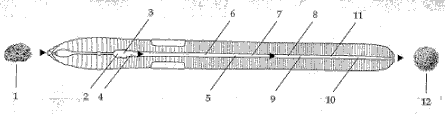
Figur 2.1
Interaktioner imellem regnormes fordřjelsessystem og mikroorganismer.
- Mikroorganismer i fřde
- Mikroorganismer som lysereres i kro/tyggemave
- Mikroorganismer med enzymer der medvirker ved nedbrydning
- Mikroorganismer der passerer ind i tarmen
- Mikroorganismer der gĺr upĺvirkede igennem tarmen
- Mikroorganismer der lyseres i tarmen
- Mikroorganismer der aktiveres
- Mikroorganismer der deler sig
- Midlertidigt residerende mikroorganismer
- Residerende mikroorganismer
- residerende mikroorganismer som koloniserer tarmmateriale
- Mikroorganismer i fćces.
Fordřjelsessystem
Regnorme konsumerer materiale, som indeholder varierende koncentrationer af mikroorganismer (1). Det konsumerede materiale blandes i kro og tyggemave med vand, slim, enzymer og mĺske antibakterielle faktorer. Dette medfřrer, at nogle mikroorganismer lyseres og sandsynligvis optages i tarmen (2); nogle af disse mikroorganismers enzymer kan medvirke ved nedbrydning af specifikke organiske komponenter i tarmen (3). Andre mikroorganismer vil overleve opholdet i den forreste del af fordřjelsessystemet og blive transporteret frem i tarmen (4). Disse kan: (5) gĺ upĺvirkede igennem tarmen og blive udskilt med fćces, (6) blive lyseret og optaget af tarmen, (7) blive aktiveret fra en mindre til en mere aktiv tilstand, (8) dele sig, (9) eller fćstne sig til tarmvćggen i lćngere eller kortere tid, og eventuelt dele sig her (midlertidigt residerende mikroorganismer). Den mikrobielle produktion, der opnĺs ved vćkst (8 & 9), kan blive lyseret og absorberet og/eller blive udskilt med fćces. I tarmen kan der ogsĺ findes residerende mikroorganismer, som danner permanente, relativt stabile populationer. De kan vćre permanent fćstet til tarmvćggen (10) eller kunne kolonisere tarmmaterialet (11); disse bakterier vil i en vis udstrćkning blive udskilt med fćces.
Mikrofloraen i fćces (12) vil sĺledes kunne bestĺ af: Mikroorganismer der gĺr upĺvirkede igennem fordřjelsessystemet (5); mikroorganismer der er blevet aktiveret (6); mikroorganismer der har delt sig og sandsynligvis dermed er blevet forřget i antal (8); samt residerende mikroorganismer, der har koloniseret tarmindholdet (9, 11). Disse forskellige grupper af mikroorganismer vil findes i en blanding, der vil afhćnge af hvilke mikroorganismer, der er blevet konsumeret, og deres skćbne i tarmen.
I det fřlgende vil det blive beskrevet hvilken evidens, der findes for de forskellige typer af relationer imellem regnorme og mikroorganismer, og eksempler pĺ hvilke mikroorganismer, som er kendt for at udvise det beskrevne respons, vil blive givet.
Mikroorganismers skćbne
En rćkke mikroorganismer reduceres betydeligt i antal (mere end en faktor 100) under ophold i regnormes fordřjelsessystem (2), f.eks. mange svampehyfer (Domsch og Banse, 1972; Dash et al., 1979), de gram-negative-bakterier Eschericia coli (Brusewitz, 1959; Pedersen og Hendriksen, 1993), Serratia marcessens (Day, 1950) og Pseudomonas putida MM11 (Pedersen og Hendriksen, 1993) samt den gram-positive bakterie Bacillus mycoides (Day, 1950). Denne antalsmćssige reduktion, der sandsynligvis primćrt forĺrsages af en lysering af cellerne, synes primćrt at foregĺ i den foreste del af fordřjelsessystemet (Pedersen og Hendriksen, 1993). Det er ikke specifikt dokumenteret for regnorme, at nogle af de lyserede organismers proteolytiske enzymer medvirker ved nedbrydning af specifikke komponenter i tarmen (3); men det er kendt fra en rćkke andre invertebrater (Martin, 1983), og resultater angĺende specifikke enzymers tilstedevćrelse i tarmen hos den tropiske art Pontoscolex corethrurus (Zhang et al., 1993) giver en betydelig indirekte evidens herfor.
Effekt pĺ sporer
Det er primćrt sporer af sĺvel bakterier som svampe, der gĺr upĺvirkede igennem fordřjelsessystemet (5) (Brown, 1995), men ogsĺ bakterien Aeromonas hydrophila (Pedersen og Hendriksen, 1993), som ikke danner sporer, blev ikke antalsmćssigt pĺvirket af passagen; dette kan dog vćre et nettoresultat af en kombination af bakteriel vćkst og dřd i tarmen. Tilsvarende synes nogle bakterier i mindre aktive tilstandeat gĺ upĺvirkede igennem tarmen (Fischer et al., 1995). Det er derimod kun dokumenteret med sikkerhed for P. fluorescens KTG (Clegg et al., 1995), at der sker et betydeligt fald i antal i selve tarmen (7).
Aktivering ved tarmpassage
Det er ikke dokumenteret for en enkelt stamme af mikroorganisme, at den bliver aktiveret fra en mindre til en mere aktiv tilstand under tarmpassagen; men en rćkke forhold taler for, at det er tilfćldet for en rćkke bakterier: (a) bakteriers dyrkbarhed forřges uden, at der samtidig sker en forřgelse i biomasse eller antal opgjort ved direkte tćllinger (Daniel og Anderson, 1991; Pedersen og Hendriksen, 1993); (b) den mikrobielle respiration forřges i fćces uden en korresponderende forřgelse i biomasse (Scheu, 1987) og (c) antallet af detekterbare bakterier, opgjort ved hjćlp af rRNAprober, forřges uden en tilsvarende forřgelse ved direkte tćlling (Fischer et al., 1995).
Flere undersřgelser beskriver, at der sker en betydelig vćkst af mikroorganismer i regnormes fordřjelsessystem (8), primćrt af bakterier og aktinomyceter (Brown, 1995). Men da de fleste undersřgelser ikke pĺ en overbevisende mĺde opgřr antallet af mikroorganismer i det konsumerede materiale, og ikke tager hřjde for en eventuel lysering i tarmsystemets forreste del og ikke tager hensyn til en reaktivering i tarmen, er det vanskeligt at benytte denne evidens som udtryk for en reel vćkst i regnormes tarm. Pedersen og Hendriksen (1993) sandsynliggřr dog, at der sker en vćkst af bakterien Enterobacter cloacae imellem fortarm og bagtarm.
Mikroorganismer i tarm
Det er sandsynligt, at der eksisterer residerende mikroorganismer i regnorme (10, 11), da Máriaglieti (1979) og Contreras (1980) overvejende fandt bakterier fra slćgten Vibrio og aktinomyceten Streptomyces lipmanii i tarmmateriale fra E. lucens. Desuden fandt Jolly et al. (1993) og VinceslasAkpa og Loquet (1995) segmenterede filamentřse mikroorganismer fasthćftet til bagtarms epitelet af O. cyaneum, L. terrestris og E. foetida; disse organismer er ikke blevet bestemt, men ligner morfologisk, hvad der findes hos rotter og mus og dér kaldes segmenterede filamentřse bakterier. Henschke et al. (1989) giver evidens for, at bakterier ogsĺ kan danne midlertidigt residerende populationer i tarmen, idet de kunne detektere P. fluorescens DSM 50148 i fćces fra L. terrestris 50 dřgn efter, at den var blevet konsumeret.
Mikroorganismer i fćces
Mikrofloraen i fćces er primćrt blevet betragtet, som vćrende en afspejling af de mikroorganismer, der findes i det konsumerede materiale (Brown, 1995; Satchell, 1983; Edwards og Fletcher, 1988). De mange forskellige relationer der eksisterer imellem regnormes fordřjelsessystem og mikroorganismer, sandsynliggřr, at dette er en alt for simpel betragtning af denne mikroflora. Efter defćkation synes der fřrst at ske en betydelig bakteriel vćkst efterfulgt af en vćkst af svampe (Brown, 1995; Satchell, 1983; Fletcher og Edwards, 1987).
Mikroorganismer pĺ orme
Det er dog ikke kun regnormenes fordřjelsessystem, der kan pĺvirke mikroorganismer. Fra ormenes overflade udskilles slim, som indeholder labile organiske forbindelser, der kan fremme mikrobiel vćkst, og deres ekskretionsprodukter indeholder mobiliserede nćringsstoffer, der ligeledes vil pĺvirke mikroorganismerne (Scheu, 1991; Lee, 1985). Det er endvidere blevet sandsynliggjort, at Flavobacterium sp. og P. cepacia kan blive transporteret rundt i jord pĺ regnormes overflade (Heijnen og Marinissen, 1995). Desuden vil regnormenes graveaktivitet og specielt de anecike regnormes transport af organisk materiale fra overfladen ned i jorden have en stor betydning for mikroorganismernes aktivitet (Hendriksen, 1996; Devliegher og Verstrate, 1995; Winding et al., submitted).
Mikroorganismers spredning
Relationerne imellem regnorme og mikroorganismer antyder kraftigt, at regnorme kan have stor betydning for specifikke mikroorganismers overlevelse og spredning i jorden. Det er da ogsĺ blevet vist, (1) at regnorme kan pĺvirke spredningen af en rćkke plantepatogene mikroorganismer (Brown, 1995), (2) spredning og dermed sygdomsbekćmpelseseffektivitet af P. corrugata 2140R (en bakterie der kan bruges til bekćmpelse af svampesygdommen »Take-All«) (Doube et al., 1994), (3) spredning og noduleringseffektivitet af rhizobier (Doube et al., 1994), samt (4) spredning og overlevelse af A. hydrophila (Hendriksen, 1995).
De mange forskellige interaktioner imellem regnorme og mikroorganismer, ikke mindst i deres fordřjelsessystem, mĺ have en afgřrende betydning for den enkelte mikroorganismes muligheder for at pĺvirke regnorme i en negativ retning. Det har dog ikke vćret muligt at finde undersřgelser som belyser samspillet imellem disse interaktioner og mikroorganismers patogene egenskaber.
2.1.3 Regnorme og mikrobielt betingede sygdomme
Patogen typer
Mikroorganismer kan vćre patogene overfor regnorme, dvs. de kan invadere regnorme og forĺrsage sygdom. Mikroorganismer er meget forskellige med hensyn til patogenecitet (evne til at forĺrsage sygdom). Dette kan dels skyldes forskelle i evnen til at invadere regnormene (trćnge ind i regnormenes kropshule og/eller celler) og dels forskelle i deres evne til at vćre aktive i regnormes kropshule. Mikroorganismer kan opdeles i potentielle, fakultative og obligate patogener. Potentielle patogener er mikroorganismer, som kan invadere vćrten, uden at vćrten af andre ĺrsager er svćkket; mens fakultative patogener kan invadere vćrten, hvis den af andre grunde allerede er svćkket. Sĺvel potentielle som fakultative patogener kan overleve og dele sig udenfor vćrterne, og de kan derfor ogsĺ dyrkes pĺ kunstige medier. Obligate patogener er derimod helt afhćngige af levende celler for at overleve, dele sig og vćre patogene. (Afsnittet er baseret pĺ Tanada og Kaya, 1993). Meget lidt er reelt kendt angĺende mikroorganismers patogenecitet overfor regnorme.
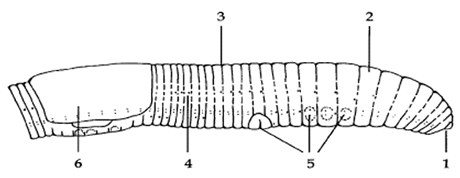
Figur 2.2
Mulige invasionsveje i regnorme for mikroorganismer
- Vćgge i fordřjelsessystemet
- Epidermis
- Dorsalporer
- Nephridieporer
- Křnsporer
- Kokoners dannelse
Invasionsveje
Dales og Kalaç (1992) definerer mikroorganismer, der er patogene for regnorme, som mikroorganismer, der kan trćnge ind i coelomet, reproducere sig og danne en blivende population i coelomvćsken. Fig. 2.2 anskueliggřr de forskellige invasionssveje, der mĺ findes hos regnorme. Mikroorganismer kan blive konsumeret og invadere selve regnormen over tarmvćggen (1). Andre mikroorganismer kan ved hjćlp af enzymer penetrere epidermis og pĺ den mĺde trćnge ind i ormen (2). Pĺ regnormes overflade findes ĺbninger ind til coelomhulen (dorsalporer (3) og nephridieporer (4)), hvorigennem mikroorganismer kan trćnge ind. Desuden kan křnsorganerne invaderes igennem křnsporerne (5). En invasion af regnormenes kokoner kan tćnkes primćrt at foregĺ under kokonernes dannelse (6). Af disse invasionsveje er kun tarmvćggen blevet dokumenteret som en sandsynlig invasionsvej til coelomet (Dales og Kalaç, 1992, Smirnoff og Heimpel, 1961). I coelomvćsken, primćrt fra E. fetida, er en rćkke forskellige hemolytiske, hemoagglutinerende og antibakterielle molekyler, samt celler med fagocytotiske egenskaber blevet identificeret (Valembois et al. 1986; Dales og Kalaç, 1992). Disse faktorer udgřr et forsvarssystem imod bakterielle infektioner. Pĺ samme mĺde er der blevet isoleret bakterier fra kokoners lumen; i dette tilfćlde ogsĺ bakterier der er blevet udsat i den omgivende jord (Zachmann og Molina, 1993); disse bakterier vil sandsynligvis kunne invadere kokonen under dens dannelse, som det er blevet postuleret for parasitiske mider (Gjelstrup og Hendriksen, 1991). Vćkst i coelomvćsken er dels betinget af mikroorganismens mulighed for at dele sig, dels af regnormes forsvarssysstems mulighed for at bekćmpe organismen, hvilket synes betinget af ikkespecifik phagocytose, som bliver hjulpet af forskellige antimikrobielle faktorer, samt muligvis af stoffer med opsoninlignende egenskaber (Dales og Kalaç, 1992). Mikroorganismer med patogene egenskaber synes sĺledes, at blive phagocyteret med en lavere hastighed end ikkepatogene mikroorganismer (Dales og Kalaç, 1992).
Mikrobielle toksiner
Mikroorganismer kan ogsĺ forĺrsage sygdom ved at danne stoffer, som er toksiske overfor organismen. I sĺdanne tilfćlde er det ikke en nřdvendig forudsćtning for virkningen, at mikroorganismen invaderer organismen. Toxinet kan enten blive produceret og/eller tilfřrt regnormens omgivelser eller blive produceret i regnormenes fordřjelsessystem og derfra pĺvirke organismen. Definitionsmćssigt findes der to forskellige typer af toxiner: (1) de anaboliske toksiner, som er stoffer, der produceres af selve mikroorganismen og (2) kataboliske toksiner, der produceres som en funktion af mikroorganismens nedbrydning af substrat, f.eks kan nedbrydning af kulhydrat medfřre dannelse af toksiske alkoholer.
Infektiv dosis
En enkelt celle fra en mikroorganisme kan principielt skabe en infektion, men generelt set er dette meget sjćldent tilfćldet. I stort set alle tilfćlde vil det krćve et minimum af infektive mikroorganismer, som invaderer organismen, fřr de forĺrsager en infektion. Denne infektive dosis udtrykkes ofte kvantitativt som letal dosis (LD), effektiv dosis (ED), eller infektiv dosis (ID) (Tanada og Kaya, 1993). Disse begreber er reelt endnu ikke taget i anvendelse i relation til regnorme.
Symptomer
Efter en mikroorganismes invasion af en regnorm, og efterfřlgende etablering, kan infektionen deles i fřlgende faser: inkubationsperioden, sygdommens udbrud, sygdommens hřjdepunkt og sygdommens ophřr. Inkubationsperioden strćkker sig fra regnormen er blevet invaderet til de fřrste tegn pĺ sygdom er synlige. Sygdommens udbrud er tidspunktet, hvor de fřrste tegn pĺ sygdommen bliver synlige. Sygdommens hřjdepunkt er den periode, hvor regnormen udviser de maksimale tegn pĺ sygdommen. Sygdommen kan enten blive kronisk eller afsluttes ved at ormen dřr eller den bliver rask. Tegnene pĺ sygdom kan enten vćre karakterer der er (1) morfologisk erkendelige f.eks. pletter, blodudtrćdninger, indsnřringer og lignende, (2) vćre erkendelige ved dyrenes aktivitet, (3) nedsćttelse af vćkstrate og/eller (4) fertilitet (kokonproduktionsrate) eller (5) dřd.
Tabel 2.2
Mikroorganismer med patogene egenskaber overfor regnorme
Mikroorganisme |
Pĺvirket regnorm |
Reference |
Corynebacterium diphteriae |
L. terrestris |
Cameron, 1932 |
Brevibacterium sp. |
E. fetida |
Roch, 1980 |
Serratia marcescens |
E. fetida |
Çotyk og Dales, 1984 |
Bacillus megaterium |
E. fetida |
Valembois et al., 1986 |
Arthrobacter sp. |
E. fetida |
Valembois et al., 1986 |
Serratia sp. |
E. fetida |
Valembois et al., 1986 |
Aeromonas sp. |
E. fetida |
Valembois et al., 1986 |
Aeromonas hydrophilia |
E. fetida |
Valembois et al., 1986 |
Yersenia ruckeri |
D. venata |
Çotyk og Kalaç , 1990 |
Yersenia ruckeri |
E. fetida |
Dales og Kalaç , 1992 |
Bacillus thuringiensis |
L. terrestris |
Smirnoff og Heimpel, 1963 |
Enterobacter aerogenes |
H. suctoria |
Rao et al. , 1983 |
Patogene mikroorganismer
Kun ganske fĺ mikroorganismer er dokumenteret patogene overfor regnorme. Tabel 2.2 er en oversigt over de af os kendte eksempler. Det fremgĺr af tabellen, at den kun omfatter bakterier, men sĺvel grampositive som gramnegative. De fřrste ti eksempler er bakterier, der kan forĺrsage dřd hos regnorme (primćrt E. fetida) ved injektion af bakterierne i coelomvćsken; det er ikke blevet undersřgt hvorvidt disse bakterier kan inficere regnorme pĺ en mere naturlig mĺde. Hendriksen (1995) og Pedersen og Hendriksen (1993) har foretaget fodringsforsřg med A. hydrophila, de fandt ingen effekter pĺ regnormene, sĺ dette kan betyde, at der enten er forskelle imellem forskellige stammer af bakterierne eller at A. hydrophila ikke er invasiv. B. thuringiensis synes at kunne invadere coelomet efter konsumption og derved forĺrsage dřd. E. aerogenes er den eneste mikroorganisme, hvorom det med stor sikkerhed kan siges, at den er patogen overfor regnorme (den tropiske art Hoplochaetella suctoria), idet en grundig undersřgelse har kundgjort, at E. aerogenes opfylder alle fire betingelser (Koch's postulater) for at kunne etablere en kausal sammenhćng imellem sygdom og mikroorganisme: (1) Den specifikke mikroorganisme skal kunne isoleres fra samtlige tilfćlde af sygdommen; (2) mikroorganismen skal kunne dyrkes i kultur; (3) mikroorganismen skal kunne give ikke syge organismer sygdommen eksperimentelt (smitteoverfřrsel); og (4) mikroorganismen skal ogsĺ kunne isoleres fra de eksperimentelt smittede organismer (Tanada og Kaya, 1993). En patogen hyphomycet Exophiala jeanselmei er blevet isoleret fra naturligt inficerede O. tyrtaeum og E. foetida kokoner, og det er blevet vist eksperimentelt, at den kan inficere kokoner fra seks forskellige arter af regnorme (Vakili, 1993).
2.1.4 Mikrobiologiske bekćmpelsesmidlers indirekte effekter pĺ regnorme
Mikroorganismer kan pĺvirke regnorme indirekte ved at influere pĺ faktorer, der har afgřrende betydning for regnormes livsprocesser. Det kan f.eks. vćre: (1) produktion af kataboliske toksiner, (2) produktion af antibiotika, der pĺvirker den mikrobielle samfundsstruktur og dermed regnormes levevilkĺr, (3) pĺvirkning af nedbrydningsprocesser og stofkredslřb. Der findes ikke undersřgelser, der belyser en eller flere af disse problemstillinger.
2.1.5 Regnorme og Bacillus thuringiensis
Et mindre antal undersřgelser har beskrevet B. thuringiensis effekt pĺ regnorme, men det er ikke muligt at drage endelige konklusioner angĺende denne bakteries patogenecitet overfor regnorme. Der findes dog en rćkke indicier for, at B. thuringiensis kan pĺvirke regnorme, muligvis ved at vćre patogen.
»Blisterdisease«
Heimpel (1966) isolerede to stammer af B. thuringiensis fra E. foetida fra regnormekulturer, som udviste en »blister-disease«. En sygdom der řjensynligt fřrer til massedřd i kulturerne. Stammerne blev begge henfřrt til B. thuringiensis subsp. thuringiensis. Isoleringen af disse to stammer blev dog ikke fulgt op af eksperimentelle undersřgelser, som kunne have eftervist, hvorvidt disse isolater forĺrsagede sygdommen. Der mangler sĺledes en eksperimentel eftervisning af Koch's postulater. Çotyk og Dales (1984) injicerede dog en af stammerne i E. fetida i relativt hřje koncentrationer (9 x 106/ml) uden at det forĺrsagede dřd; dette kan skyldes en rćkke forskellige ĺrsager, som ikke nřdvendigvis er knyttet til bakteriens patogene egenskaber: F.eks. forskelle imellem forskellige populationer af E. fetida's respons, at stammen af B. thuringiensis har ćndret sig ved gentagne opformeringer i kultur, eller at sporer og vegetative celler af B. thuringiensis giver forskelligt respons hos ormen.
B. t. subsp. Thuringiensis
Smirnoff og Heimpel (1961) undersřgte eksperimentelt effekten af det mikrobiologiske bekćmpelsesmiddel Thuricide, som indeholder B. thuringiensis subsp. thuringiensis, pĺ L. terrestris. De fandt, at hřje ikke realistiske koncentrationer (120 vćgt%) Thuricide iblandet jorden, svarende til ca. 4x108 8x109 B. thuringiensis/g jord, alle medfřrte 100% mortalitet i lřbet af ca. 2 mĺneder, selv ved den laveste pĺvirkning. Histologiske undersřgelser af de dřde orme viste, at B. thuringiensis invaderede coelomet via tarmvćggen og forĺrsagede dřd pĺ grund af blodforgiftning. Krieg (1983; citeret i Addison (1993)) foreslĺr dog, at invasionen ikke var forĺrsaget af selve bakterien, men af diatoméjorden som blev benyttet som bćrestof i formuleringen af Thuricide.
B. t. subsp. Galleriae
Atlavinyte et al. (1982) undersřgte eksperimentelt effekten af det mikrobiologiske bekćmpelsesmiddel Entobacterin, som indeholder B. thuringiensis subsp. galleriae, pĺ A. caliginosa. Deres resultater er desvćrre tvetydige. De fandt sĺledes i ét eksperiment at Entobacterin ingen eller ringe effekt havde pĺ ormene, mens det i at andet eksperiment havde en effekt pĺ sĺvel overlevelse som vćkst. Effekten var isćr udtalt med koncentrationer svarende til ˝ og 1 gange feltdosis, mens 2 gange feltdosis havde en ringe effekt. Det blev ikke undersřgt, hvorvidt bakterien inficerede ormene. Det blev derimod vist, at mellemkoncentrationerne af Entobacterin negativt pĺvirkede parametre som nedbrydningsraten af halm, tćthederne af mider, collemboler, insektlarver og mikroorganismer (opgjort ved pladespredning) og visse jordbundskemiske parametre. Dette kunne antyde, at den viste effekt pĺ A. caliginosa af Entobacterin snarere er en indirekte effekt end en mere direkte.
Der er endvidere blevet gennemfřrt et par feltundersřgelser vedrřrende B. thuringiensis effekt pĺ regnorme (White, 1961 (citeret i Benz og Altweg, 1975); Benz og Altweg, 1975). Ingen af disse undersřgelser er gennemfřrt pĺ en sĺdan mĺde, at det er muligt at give en biologisk meningsfuld fortolkning af resultaterne, da de ikke angiver hvilke regnorme (řkotyper), der er blevet fundet pĺ lokaliteterne, og det dermed ikke kan vurderes, hvorvidt ormene reelt har vćret eksponeret for bakterierne.
B. thuringiensis i fćces
Hendriksen (upubl.) fandt B. thuringiensis (identificeret v.h.a. kolonimorfologi og tilstedevćrelse af krystaller) i fćces fra fřlgende arter af aktive regnorme indsamlet pĺ forskellige lokaliteter: L. rubellus, L. festivus, L. terrestris, A. caliginosa, A. chlorotica, A. longa og O. cyaneum. Forekomsten var mest talrig i de endogćiske arter. Dette antyder, at der forekommer stammer af B. thuringiensis i det materiale, som regnorme konsumerer, og at denne forekomst ikke umiddelbart synes at pĺvirke regnormene.
B. thuringiensis varianter
Da der findes mange forskellige underarter af B. thuringiensis, som producerer d-endo-toxiner med forskellige egenskaber (Ellar, 1990), og da det enkelte isolat desuden kan producere en rćkke andre stoffer med toksiske egenskaber, primćrt i den vegetative vćkstfase (Hansen et al., 1996), er det meget sandsynligt, at der vil vćre stor variation imellem forskellige underarters og isolaters effekt pĺ regnorme. Dette forhold kompliceres yderligere af, at det ikke med udgangspunkt i den tilgćngelige viden er muligt at afgřre hvilke forhold (d-endo-toksiner, andre toksiner, indirekte effekter) som er involveret i B. thuringiensis mulige effekt pĺ regnorme.
2.1.6 Regnorme og aktinomyceter
Aktinomyceter er ofte blevet isoleret fra tarmmateriale fra regnorme (Brown, 1995); og det er sandsynligt, at nogle aktinomyceter kan vokse i tarmmiljřet (Parle, 1963). Isćr er forekomsten af aktinomyceter knyttet til regnorme blevet studeret i Centraleuropa (Contreras, 1980; Kristufek et al. 1990; Ravasz og Toth, 1990; Kristufek et al., 1993). Disse undersřgelser viser, (1) at det blandt aktinomyceterne isćr er arter fra slćgten Streptomyces, der er almindeligt forekommende i tarmmaterialet, (2) at Streptomyces arterne der findes i regnormene oftest ogsĺ er almindelige i jorden, men at forholdet imellem arterne varierer imellem regnorme og jord, (3) at der er forskel i Streptomycet-floraen imellem forskellige arter af regnorme, hvilket isćr synes knyttet til forskelle i řkotyper, (4) at nogle fĺ Streptomycet arter mĺske kan danne residerende populationer i tarmen hos regnorme.
2.1.7 Regnorme og Streptomyces griseoviridis
Streptomyces griseoviridis er ikke blandt de isolater som er blevet isoleret og identificeret fra regnorme (adskillige hundrede isolater) (se ovenfor).
Det er, sĺ vidt vides, aldrig blevet undersřgt, hvorvidt aktinomyceter kan pĺvirke regnorme i en negativ retning (vćre patogene eller pĺ anden mĺde pĺvirke regnorme). Det er sĺledes ikke muligt med baggrund i litteraturen at udtale sig om, hvorvidt S. griseoviridis har potentiale for at have negative effekter pĺ regnorme. Streptomyceter synes dog ikke at vćre stćrkt virulente overfor invertebrater, da der ikke er beskrevet sygdomstilfćlde hos invertebrater, der er sat i forbindelse med infektion med en art af Streptomyces (KornWendisch og Kutzner, 1992). Med udgangspunkt i dette, og S. griseoviridis antibiotiske egenskaber (Lübeck, 1994), sĺ synes indirekte effekter mere sandsynlige end direkte patogene effekter.
2.2 Nematoder
Diversitet
Nematoder kan ikke pĺ samme mĺde som regnorme betragtes som en gruppe af dyr med en relativt ensartet effekt pĺ řkosystemer. Nematoder er en dyregruppe, der indeholder arter, som regnes for at vćre skadedyr (en rćkke parasitćre arter pĺ sĺvel hřjere dyr som planter). Nematodarter, der parasiterer artropoder og mollusker, kan anvendes til biologisk bekćmpelse af en rćkke forskellige arter af disse invertebrater og udnyttes i denne forbindelse kommercielt. Gruppen indeholder ogsĺ en lang rćkke fritlevende arter, der kan have andre mere gavnlige effekter pĺ řkosystemers funktion.
Funktion
De fritlevende nematoder udgřr imellem en og ti millioner individer/m2 i de allerfleste terrestriske řkosystemer (Nicholas, 1975), hvor deres biologiske rolle er at indgĺ i den terrestriske detritusfřdekćde pĺ flere forskellige niveauer og ved at pĺvirke mineralisering af nćringsstoffer (Griffiths, 1994). De fritlevende nematoder er en yderst divers gruppe af dyr, der dels er vidt forskellige i en taksonomisk sammenhćng, dels har en meget forskelligartet ernćringsbiologi. Der kan f.eks. skelnes imellem bakteriećdere, svampećdere, algećdere, rodsugere, predatorer og nematoder med et blandet fřdeindtag (Nicholas, 1975). Nematoders forekomst og řkologi i Danmark er řjensynligt ikke blevet undersřgt siden Nielsen (1949). Det mĺ forventes, at den danske nematodfauna omfatter adskillige tusinde arter.
Derimod pĺvirker nematoder, i modsćtning til regnorme, ikke direkte jordens kemiske og fysiske egenskaber, da de pĺ grund af deres ringe střrrelse ikke har jordbundsdannende egenskaber, som det er tilfćldet med regnorme.
Effekt pĺ nematoder
Mikrobiologiske plantebeskyttelsesmidler kan potentielt set pĺvirke nematoder pĺ mange forskellige mĺder, f.eks. ved at reducere antallet (enten direkte eller ved nedsat vćkst og fertilitet) og/eller ved at ćndre artssammensćtningen og dermed pĺvirke detritusfřdekćden.
Nematoder som testdyr
Nematoder har nogle egenskaber, der gřr dem fornuftige at benytte som testorganismer i denne sammenhćng:
- de er almindeligt forekommende i de fleste jorder;
- de er relativt simple at indsamle;
- en rćkke arter kan holdes i kultur, sĺvel xenisk som axenisk;
- mange arter har en relativ kortvarig livscyklus, hvilket gřr det nemmere at studere vćkst og frugtbarhed.
Men desvćrre er der ogsĺ en rćkke egenskaber, der gřr dem mindre egnede som testorganismer:
- det er en meget divers gruppe af dyr, bestĺende af mange arter, med mange forskellige typer af biologi;
- det er yderst vanskeligt at identificere nematoder;
- kendskabet til de fritlevende nematoders taxonomi, udbredelse, biologi og řkologi er begrćnset.
2.2.1 Nematoder og Bacillus thuringiensis
En rćkke undersřgelser har klargjort, at der kan forventes en vis grad af pĺvirkning af nematoder ved anvendelse af B. thuringiensis. Reprćsentanter for bĺde fritlevende-, planteparasitiske- og zooparasitiske nematoder har vist sig, et eller flere steder i deres livscyklus at vćre fřlsomme, i střrre eller mindre grad, over for visse B. thuringiensis stammer.
Effekt pĺ nematoders ćg
En rćkke rapporter peger pĺ, at B. thuringiensis d-endotoksiner kan vćre toksiske isćr over for nematoders ćg. Bottjer et al. (1985) undersřgte in vitro effekten af 30 sporulerede B. thuringiensis varieteter pĺ ćg fra den zooparasitiske Trichostrongylus colubriformis. Alle 30 varianter af bakterien viste sig at vćre toksiske overfor ćggene. Den toksiske aktivitet af d-endotoksin berigede fraktioner varierede fra en LD50 pĺ 0.025 til 2100 ng total protein per ml, dvs en variationsfaktor pĺ 84000. To af de kommercielt anvendte B. thuringiensis varieteter, israelensis og kurstaki havde LD50 vćrdier pĺ henholdsvis 1.27 og 62 ng totalprotein per ml. Yderligere undersřgelser med B. thuringiensis var. israelensis toksinpreparationer og ćg fra seks zooparasitiske og en fritlevende nematod(er) viste toksisitet over for alle syv arter af nematoder med LD50 vćrdier i omrĺdet 0.38-10.6 ng totalprotein per ml. B. thuringiensis letale effekt pĺ nematoders ćg er for T. colubriformis forĺrsaget af en ćndring af strukturen af ćggeskallen, hvilket medfřrer en ćndret permeabilitet (Bone et al., 1987, Wharton og Bone, 1989). Bone et al. (1986) fandt ved bioassays med T. colubriformis og forskellige fraktioner af B. thuringiensis var. israelensis kulturer, at effekten pĺ nematoders ćg kan henfřres til proteinkrystallen, som ikke kun indeholder insektspecifikke endotoksiner. Det blev ogsĺ vist at toksisiteten kan henfřres til et protein, da aktiviteten blev reduceret af proteolytiske enzymer. Yderligere fandt man, at den relevante fraktion ikke udviste effekt over for myggen Aedes aegypti, hvilket er et indicie for, at det ikke er de Diptera aktive toksiner, som udviser effekt pĺ nematoders ćg. Pĺ samme mĺde havde oprensede B. thuringiensis var. israelensis Diptera aktive proteiner ingen effekt over for nematoderne.
b-exotoksin
Visse B. thuringiensis typer, som f.eks. varieteten thuringiensis producerer b-exotoxin, som kan udskilles i dyrkningsmediet. Dette toxin er en RNA syntese inhibitor (Bone, 1989). b-exotoksin produceres ikke af hovedparten af de kommercielt anvendte stammer, som f. eks B. thuringiensis var. israelensis (Lisansky et al., 1993). Toksinet inhiberer ćg-klćkning og drab af andet trin larver af den plantepatogene nematod Meloidogyne spp. (Prasad et al., 1972). Ligeledes var b-exotoksin ĺrsag til, at vćksten i kulturer af den fritlevende nematode Panagrellus redivivus blev signifikant hćmmet, at den tomat patogene Meloidogyne incognita ikke dannede galler pĺ tomatrřdder, og at den mycophage Aphelenchus avenae blev hćmmet markant i vćkst (Ignoffo og Dropkin, 1977). Det kommercielle produkt Thuringiensin, som indeholder b-exotoksin, var i stand til at reducere antallet af N. brasiliensis i tarmsystemet hos mus (Bone, 1989).
Effekt pĺ larver og voksne
Det er imidlertid ikke kun B-exotoksin, som kan hćmme larver og voksne nematoder. Meadows et al. (1990) fandt sĺledes, at oprensede d-endotoksiner fra B. thuringiensis varieteterne israelensis, kurstaki, og morrisoni var i stand til effektivt at reducere den fritlevende nematod Turbatrix aceti i antal. Varieteten israelensis effekt pĺ sĺvel larver som voksne nematoder var mere udtalt end kurstaki's; dette svarer til effekterne overfor ćg, som er beskrevet tidligere.
Spore spiring
I ovenstĺende undersřgelser er der arbejdet med mere eller mindre oprensede toksiner. I nogle tilfćlde er der anvendt sĺkaldt toksinberigede fraktioner, hvor sporerne er sřgt fjernet ved gradient centrifugeringer. I disse tilfćlde kan det ikke udelukkes, at effekterne pĺ nematoderne er forĺrsaget af en kombination af toksiner og spirede B. thuringiensis celler. I forbindelse med en screening af B. thuringiensis isolater for aktivitet overfor nematoder fandt Borgonie et al. (1995) sĺledes spirende sporer i forbindelse med alle 12 arter af nematoder, uanset om spore-krystal blandingen var toksisk for nematoderne eller ej. Hyppigheden af sporespiring var nematodarts afhćngig. I en anden rapport (Leyns et al., 1995), testedes aktiviteten af 128 forskellige B. thuringiensis spore-krystal preparationer over for juvenile og voksne Caenorhabditis elegans (en fritlevende nematod). Af de 128 B. thuringiensis isolater viste 109 stort set ingen effekt overfor nematoder, 16 isolater udviste en mortalitet pĺ mellem 10 og 20%, mens tre isolater resulterede i mortaliteter pĺ henholdsvis 21, 47 og 59%. I denne forsřgsrćkke blev der ikke observeret letale effekter over for nematoders ćg. Dette skyldes sandsynligvis, at sporespiring og vćkst var inhiberet i forsřgene, idet der var tilsat et antibiotikum. Det er derfor meget sandsynligt, at visse varianter af B. thuringiensis effekt overfor nematoders ćg er knyttet til bakteriens spiring og vćkst. I overensstemmelse hermed har vi observeret (se bilag 2), at alle afprřvede B. thuringiensis isolater var letale over for voksne Panagrellus redivivus nĺr der ikke var tilsat et antibiotikum, mens kun fĺ bakterier var letale nĺr der var tilsat et antibiotikum.
Specifik aktivitet
Flere af de bakterier, som vi har anvendt i testen over for P. redivivus, er tidligere blevet patenteret som B. thuringiensis stammer med en specifik aktivitet overfor nematoder (Edwards et al., 1990). Yderligere en rćkke bakterier og gener med nematicidal aktivitet er sřgt patenteret (Narva et al., 1991). Analyserne, som ligger til basis for patentet (Edwards et al., 1990), er blevet foretaget med sporekrystal blandinger, hvor sporene er blevet inaktivteret ved 60Co bestrĺling. Det fremgĺr ikke tydeligt af patentet, men vi skřnner, at B. thuringiensis dřdesporerkrystal blandingen mĺ have en tćthed pĺ 1081010/krystaller per ml for at kunne inaktivere alle nematoderne ved in vitro bioassays. Det er sĺledes vist, at der findes specielle varieteter af B. thuringiensis, som udviser en aktivitet overfor nematoder, der er knyttet til specifikke toksiner, hvor man har kendskab til generne som koder for toksinerne. Det er blevet sandsynliggjort, at B. thuringiensis varieteter med gener kodende for disse toksiner er almindeligt forekommende i jord (Hansen et al., 1996).
Krystal proteiner
Ved bioassays, med krystalberigede fraktioner, hvor eventuelle sporer er strĺledrćbte, anvendes omkring 150 nematoder og 100 mg protein, hvilket resulterer i en mortalitet pĺ 99-100%. Selv om voluminer ikke er angivet, skřnnes det pĺ baggrund af metodebeskrivelserne i andre rapporter vedrřrende B. thuringiensis-nematode bioassays, at proteinkoncentrationen er ca. 1 mg per ml. Disse proteinmćnger er i střrrelsesordenen 106 gange střrre end de LD50 vćrdier som Bottjer et al. (1985) fandt. Dette er endnu et indicium for, at af sporespiring og vćkst har betydning for B. thuringiensis effekter pĺ nematoder. Leyns et al. (1995) fandt ogsĺ, at relativt hřje doser er nřdvendige for at se effekt pĺ nematoder. Der var ingen effekt med en sporedensitet pĺ 107 per ml, og fřrst ved densiteter pĺ over 108 var der signifikante effekter pĺ nematoderne. Ved disse forsřg blev der, som tidligere nćvnt, anvendt antibiotika. I vore egne bioassays blev der ogsĺ anvendt antibiotika for at forhindre bakterievćkst (bilag 2).
In vitro – in vivo
Det skal understreges, at hovedparten af data, der er refereret hidtil i dette afsnit, er baseret pĺ in vitro undersřgelser. Bone (1989) fandt, at et kommercielt produkt baseret pĺ B. thuringiensis var. israelensis havde lavere nematicidal aktivitet in vivo i mus inficeret med N. brasiliensis end in vitro. Det omvendte forhold gjorde sig gćldende for et produkt, hvor toxiciteten var baseret pĺ d-exotoksin.
Sprřjte dosis
I forbindelse med dosisaktivitets forhold er det relevant at nćvne, at umiddelbart efter kommerciel flysprřjtning af et skovomrĺde i Québec med B. thuringiensis var. kurstaki fandt man 7×105 CFU per cm2 skovbund (Cardinal and Marotte, 1987). Ved vore feltforsřg med samme bakterie-type indeholdt sprřjtevćsken 6.5×107 CFU per ml, og de hřjeste antal genfundne bakterier pĺ blade og i jord lige efter sprřjtning var henholdsvis 107 og 104 per gram (Pedersen et al., 1995).
Reel dosis
Det skal her understreges, at selv om en sporedensitet pĺ 107 per ml ikke har nogen effekt in vitro er det ikke ensbetydende med, at en langt lavere sporedensitet pr. gram jord nřdvendigvis ikke har aktivitet. Dette skyldes, at vi ikke har et kendskab til sporer og krystallers fordeling i jorden. Hvis f.eks B. thuringiensis primćrt findes i jordens vandhinder, sĺ vil der i en jord med lavt vandindhold sandsynligvis vćre hřje densiteter af B. thuringiensis i de vandhinder, hvor bakterien findes. Som en konsekvens heraf vil effekten pĺ nematoder in vivo kun kunne bestemmes eksperimentelt. Sĺdanne undersřgelser er ikke řjensynligt ikke blevet gennemfřrt.
2.2.2 Nematoder og aktinomyceter
Vi har ikke fundet litteratur, der belyser interaktioner imellem aktinomyceter og nematoder, og dermed naturligvis heller ikke angĺende S. griseoviridis.