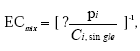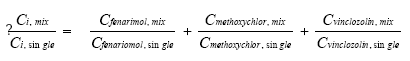|
Hormonforstyrrende effekter af kombinationer af pesticider 2 Mekanismer for kombinationseffekter af hormonforstyrrende stoffer
2.1 Østrogene pesticider testet in vitroDen østrogene virkning af pesticiderne er undersøgt i et MCF-7 celle proliferationsassay. Dette assay er baseret på en human brystcancercellelinie, som differentierer og deler sig når der er østrogen til stede. Proliferation af denne cellelinie er således indikation for, at der er et stof til stede med østrogen aktivitet. Metoden anvendes rutinemæssigt til screening for miljøkemikalier med østrogene egenskaber flere steder, herunder på Afdeling for Miljømedicin på Syddansk Universitet i Odense (Andersen et al. 1999). Kort beskrevet blev MCF-7 celler udsået i mikrotiterplader og efter 24 timer blev en række koncentrationer (0,01– 50 μM) af pesticider tilsat, dels enkeltvis, dels i kombination. Pesticiderne blev testet både med og uden 10 pM 17-østradiol (som inducerer ca 50 % af det maximale proliferationsrespons i cellerne) for at undersøge både agonist og antagonist effekter overfor østradiol. Efter 6 dages inkubation blev cellerne fikseret og farvet. Farveintensiteten, som er et mål for antallet af levende celler, blev aflæst i en ELISA-reader. Hvert forsøg inkluderede en standardrække af 17β-østradiol (1 pM til til 10 nM), hvor maximalt respons sås ved 100 pM. Resultaterne er angivet som den Relative Proliferative Effekt (RPE), hvorved data er normaliseret til både ubehandlede kontrolceller og det maksimale respons induceret af 1 nM 17β-østradiol i det respektive forsøg: RPE = (PEteststof – 1)/(PEmax 17β-østradiol -1), hvor PE er responset normaliseret i forhold til kontrol. Den forventede effekt af blandingerne er beregnet ud fra koncentrations-additions modellen, hvor den følgende ligning gælder for alle effektniveauer:
hvor Ci, mix er koncentrationen af stof i i en blanding, som giver en given effekt og Ci, single er den koncentration, som skal til af det rene stof for at give den samme effekt. Ligningen kan omskrives til:
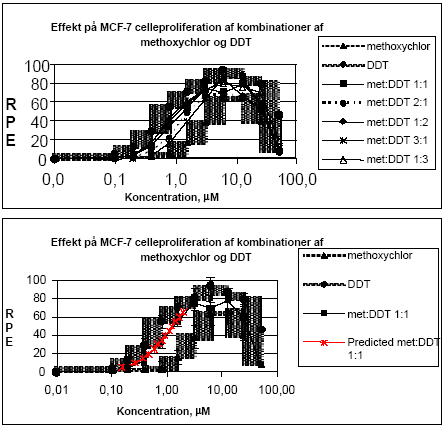
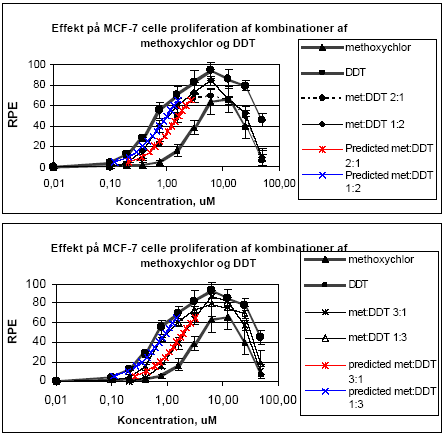
hvor pi er den relative andel af det i 'te stof ud af den totale koncentration af blandingen. Ud fra denne ligning kan den koncentration af blandingen som forventes at give en bestemt effekt beregnes, idet effekten af de enkelte stoffer som funktion af koncentrationen er bestemt ud fra fittede kurver. For pesticidblandinger testet direkte for østrogene effekter var det bedste kurvefit den sigmoide Chapman funktion (3 parametre) mens den sigmoide funktion (4 parametre) er anvendt for agonistiske effekter for blandinger testet sammen med 17β-østradiol. For antagonistiske effekter er funktionen for ekspotentielt henfald (5 parametre) anvendt som bedste kurvefit. Regressionskoefficenterne for de fittede kurver lå mellem 0,89 og 0,99 for de forskellige blandinger. 2.1.1 Effekt af methoxychlor og o,p-DDT på MCF7 celleproliferationMethoxychlor og o,p-DDT er to pesticider med kendte østrogene effekter. En mulig samspilseffekt mellem stofferne med hensyn til deres østrogene virkning er undersøgt i dette assay. Indledningsvist var der nogle metodologiske problemer, idet o,p-DDT afdampede under inkubationen og forurenede de omkringliggende dyrkningshuller på mikrotiterpladen. Hver koncentration af o,p-DDT er derfor blevet testet på separate mikrotiterplader. Pesticiderne blev blandet i molære ratioer på 1:1, 1:2, 2:1, 1:3 og 3:1. Der blev udført tre uafhængige forsøg og data blev analyseret ved isobolmetoden. Både methoxychlor og o,p-DDT stimulerede koncentrationsafhængigt proliferationen af MCF-7 cellerne som tegn på østrogen virkning. For begge pesticider var der et fald i celleproliferationen ved de to højeste koncentrationer (25 og 50 μM) på grund af cytotoksicitet, hvorfor disse koncentrationer ikke blev medtaget i de videre beregninger. Som det ses af Fig 2.1.1 er der god overenstemmelse mellem de målte effekter og de prædikterede værdier for additiv virkning beregnet ud fra isobolmetoden.
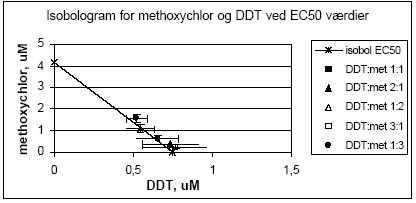
Figur 2.1.1. Effekten af methoxychlor og o,p-DDT samt kombinationer af de to stoffer på proliferationen af MCF-7 celler samt den predikterede effekt under antagelse af additivitet beregnet ved isobolmetoden. De observerede effekter er gennemsnit af mindst tre uafhængige forsøg udført i trippelbestemmelse. Isobolen for de koncentrationer af o,p-DDT og methoxychlor som inducerer 50% stigning i MCF-7 celleproliferationen i forhold til ubehandlede kontrolceller er afbilledet på Figur 2.1.2. Den koncentration af o,p-DDT som øger proliferationen med 50 % er angivet på x-aksen og den tilsvarende koncentration for methoxychlor er angivet på y-aksen. Ved additivitet vil værdierne for blandinger af de to stoffer ligge på den rette linie som forbinder disse to punkter (isobolen). Det ses at de målte værdier for blandingerne ligger tæt på isobolen, hvilket viser god overenstemmelse med additivitet for de to stoffers kombinationseffekt.
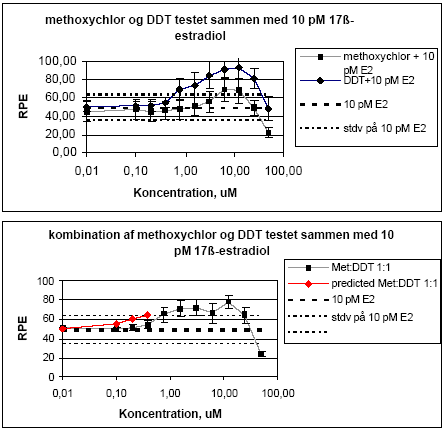
Figur 2.1.2. Isobologram for de koncentrationer af o,p-DDT og methoxychlor som inducerer 50% stigning i MCF-7 celle- proliferationen samt de observerede værdier for blandinger af de to pesticider. Når methoxychlor og o,p-DDT blev testet sammen med 10 pM af 17β-østradiol var der ingen tegn på antagonistisk effekt overfor 17-østradiol i dette assay. Derimod medførte begge stoffer en yderligere stigning i celleproliferationen (Figur 2.1.3). Denne effekt var mest markant for o,p-DDT, som også var mest potent, når stofferne blev testet uden 17β-østradiol. Tilsvarende sås heller ingen antagonistisk effekt overfor 17β-østradiol, når de forskellige blandinger af o,p-DDT og methoxychlor blev testet sammen med 10 pM 17β-østradiol, men et øget respons svarende til det additive respons for blandinger af o,p-DDT og methoxychlor.
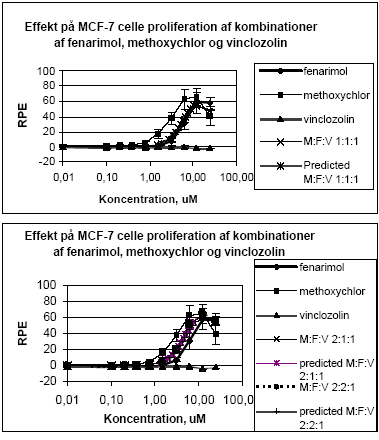
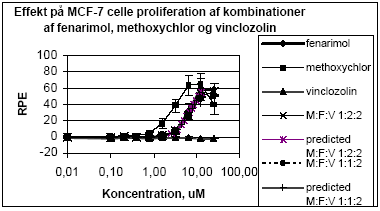
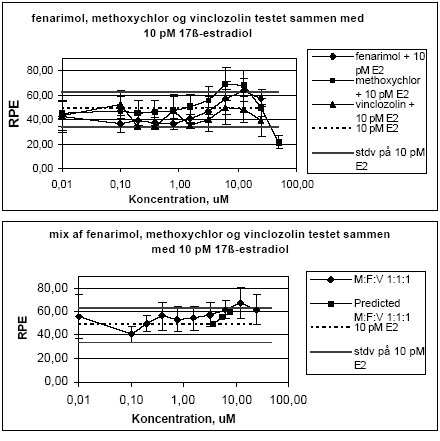
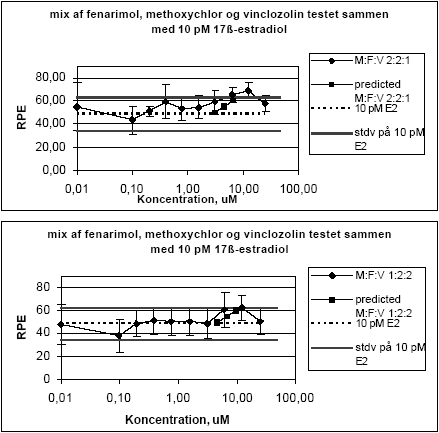
Figur 2.1.3. Effekten af methoxychlor og o,p-DDT samt blandinger heraf testet sammen med 10 pM 17-østradiol på proliferationen af MCF-7 celler samt den prædikterede effekt under antagelse af additivitet beregnet ved isobolmetoden. De observerede effekter er gennemsnit af mindst tre uafhængige forsøg udført i tripelbestemmelse. 2.1.2 Effekt af kombinationer af vinclozolin, methoxychlor og fenarimolSom eksempel på tre pesticider med forskellige hormonforstyrrende virkningsmekanismer blev fenarimol, methoxychlor og vinclozolin valgt. Fenarimol og methoxychlor er begge østrogene. Fenarimol er desuden en aromatasehæmmer og alle tre pesticider har antiandrogene egenskaber. De tre pesticider samt blandinger heraf blev testet i MCF-7 celleproliferationsassayet i 3 uafhængige forsøg. Fenarimol og methoxychlor inducerede begge proliferation af MCF-7 cellerne. Begge pesticider optrådte som partielle agonister idet det maksimale respons kun var ca. 60% af det maksimale respons induceret af 17β-estradiol (Figur 2.1.4). Vinclozolin havde ingen effekt i assayet. Ved testning af 5 forskellige blandingsforhold af de tre pesticider var der god overenstemmelse mellem de registrerede effekter og de prædikterede effekter beregnet ud fra isobolmetoden.
Figur 2.1.4. Effekten af fenarimol, methoxychlor og vinclozolin samt kombinationer af de tre stoffer på proliferationen af MCF-7 celler samt den predikterede effekt under antagelse af additivitet beregnet ved isobolmetoden. De observerede effekter er gennemsnit af tre uafhængige forsøg udført i tripelbestemmelse. Isobolkoeficienter ved de blandingskoncentrationer der giver et respons på henholdsvis 5, 25 og 50 RPE er beregnet for de fem blandinger ud fra formlen:
idet bidraget fra vinclozolin er sat til nul (Tabel 2.1). Tabel 2.1 Isobolkoeficienter ved EC5 , EC25 og EC50 for blandinger af fenarimol, methoxychlor og vinclozolin
Når de tre pesticider enkeltvist blev testet sammen med 10 pM 17β-østradiol sås en tendens til en svag øgning af responset ved de højeste ikke-cytotoksiske koncentrationer (10-25 μM) i forhold til 17β-østradiol alene, undtagen for vinclozolin. Ved de lave koncentrationer af fenarimol og vinclozolin var der en tendens til en reduktion af 17β-østradiol responset, som dog ikke er signifikant da det ligger indenfor standardafvigelsen for 17β-østradiol (Figur 2.1.5). Også for kombinationer af fenarimol, methoxychlor og vinclozolin testet sammen med 10 pM 17β-østradiol sås en tendens til et øget respons som svarede rimeligt godt med det forventede respons under forudsætning af additivitet mellem fenarimol og methoxychlor (Figur 2.1.5).
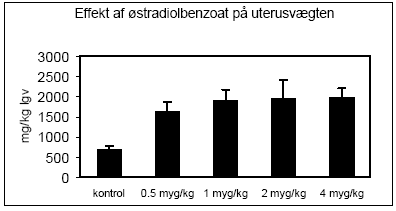
Figur 2.15. Effekten af fenarimol, methoxychlor og vinclozolin samt blandinger heraf testet sammen med 10 pM 17β-østradiol på proliferationen af MCF-7 celler samt den predikterede effekt under antagelse af additivitet beregnet ved isobolmetoden. De observerede effekter er gennemsnit af mindst tre uafhængige forsøg udført i tripelbestemmelse. 2.2 Østradiol-inducerede effekter in vivoDen klassiske uterustest blev anvendt til undersøgelse af samspillet mellem methoxychlor og o,p-DDT in vivo. For at få valideret og endelig indkørt de metoder, der skulle anvendes i forbindelse med uterustest af pesticiderne, blev der udført indledende forsøg med 4 doser østradiolbenzoat. Den anvendte model for uterustesten bygger i dette tilfælde på ovariektomerede voksne Wistar hunrotter der behandles med pesticiderne og et positivt kontrolstof peroralt i 4 dage. Efter aflivning af dyrene blev uterusvægten målt, serum fremstillet til hormonanalyser og biomarkøranalyser og mRNA blev isoleret fra uterus til genekspressionsanalyser. Metoden er blevet etableret ved Danmarks Fødevare- og Veterinærforskning (Hossaini et al. 2000) og har indgået i et interkalibreringsstudie i OECD regi. 2.2.1 Effekt på organvægteMed stigende doser af østradiolbenzoat (s.c.) sås en markant forøgelse af uterusvægten, mens lever- og nyrevægt var uændret. En statistisk signifikant forøgelse af uterusvægten sås allerede ved den laveste dosis på 0.5 g/kg (Fig 2.2.1).
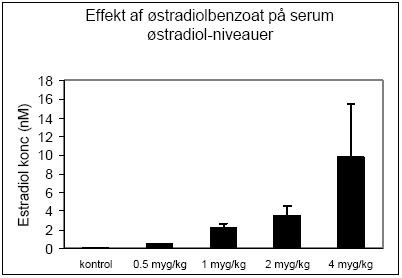
Figur 2.2.1 2.2.2 Effekt på hormonniveauerNiveauer af østradiol i serum fra dyrene blev analyseret ved hjælp af Delfia tidsforsinket fluorescens efter forudgående fast-fase ekstraktion på C18-søjler. Med stigende doser af østradiolbenzoat sås en markant forøgelse af østradiolniveauet, hvilket afspejler de stigende doser af stoffet som dyrene blev doseret med og dermed tjener som en kvalitetskontrol for fremstilling af opløsninger, dosering af dyrene og hormonananalysen (Fig 2.2.2).
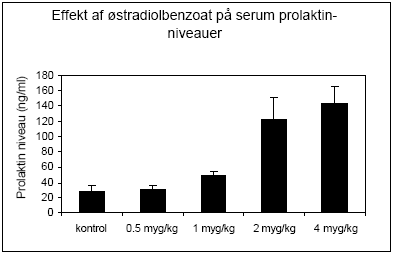
Figur 2.2.2 Hypofysehormonet prolaktin secerneres fra hypofysen efter østrogenbehandling af hunner og kan muligvis bruges som markør for eksponering for østrogene miljøkemikalier. Rotteprolaktin blev i dette tilfælde analyseret ved hjælp af et enzymimmunoassay (BiotrakTM) udviklet af Amersham og der sås stigende niveauer af hormoner ved doser på 1 μg/kg og derover (Fig 2.2.3).
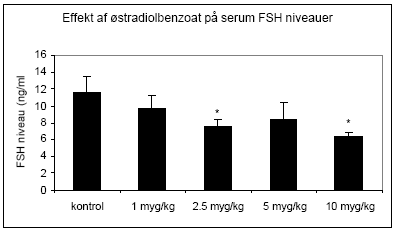
Figur 2.2.3. Follikelstimulerende hormon (FSH) er et andet hypofysehormon, der er under regulation af østradiol, som ved stigende doser bevirker en feedback hæmning af sekretionen af FSH fra hypofysen. Rotte FSH i serum blev analyseret ved hjælp af Delfia tidsforsinket fluorescens og der sås et fald i niveauet, der var statistisk signifikant ved doser på 2.5 μg/kg og derover (Fig 2.2.4).
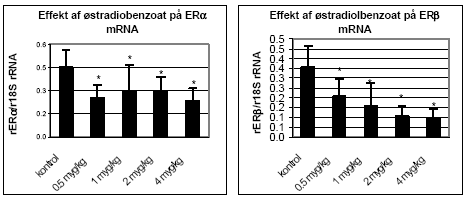
Figur 2.2.4 2.2.3 Effekt på østrogenregulerede generØstrogener, som er kvindelige kønshormoner, påvirker udvikling, differentiering og funktionen af det mandlige og kvindelige reproduktionssystem (Enmark & Gustafsson, 1999). Østrogener spiller også en vigtig rolle i funktionen af knogler, centralnervesystem, immunsystem, hud og det kardiovaskulære system (Ahmed 2000; Enmark & Gustafsson 1999; Gaido et al. 1999; Enmark et al. 1997; Couse et al. 1997). Østrogen receptor (ER) α og β medierer den biologiske effekt af østrogener og er ligand-aktiverede transkriptionsfaktorer, som regulerer ekspression af steroid-responderende gener (Levin 2002; Nielsson et al. 2001). ERα og ERβ har specifikke fordelinger i celler og væv (Enmark et al. 1997) og regulerer forskellige gener (Pettersson & Gustafsson 2001) og det er blevet foreslået, at ERβ modulerer effekter medieret af ERα ved f.eks. at hæmme celleproliferation (Cappelletti et al. 2003; Pettersson et al. 2000). Selv om der er foreslået mange funktioner af ERβ, er denne recptors rolle stadig kontroversiel. Østradiol er vist at nedregulere genekspression af ER i bl.a. MCF-7 celler (Vienonen et al. 2003; Clayton et al. 1997; Donaghue et al. 1999). Hypotesen er, at nedreguleringen af ER mRNA bl.a. involverer en negativ feedback autoregulering af ER på egen transkription, hvor promotor nedregulering tilsyneladende medieres via cellers specifikke transkriptionsfaktorer (Donaghoue et al. 1999). Metoden til at bestemme effekter af miljøkemikalier på genekspression af bl.a. ERα og ERβ og lactoferin er etableret ved Institut for Miljø og Arbejdsmedicin, Aarhus Universitet. Tidligere resultater fra laboratoriet har vist en nedregulering af ER og opregulering af ERα/β mRNA ved eksponering af MCF-7 celler til 17β-østradiol samt effekter på ERα/β mRNA niveau af en række pesticider (Hofmeister & Bonefeld-Jørgensen, 2003; Grünfeld & Bonefeld-Jorgensen, 2003). I dette studie er effekten af østradiol på østrogent-regulerede gener undersøgt i uterus fra eksponerede rotter ved hjælp af semikvantitativ real-time PCR. Lightcycler (Roche) bruges som et thermocycling on-line detektionssystem for at kvantificere RT-PCR produkter og anvendes rutinemæssigt ved Institut for Miljø og Arbejdsmedicin, Århus Universitet (Krüger et al. 2003). Total RNA fra uterus blev isoleret med RNeasy (Qiagen, Merck Eurolab) og omdannet til cDNA med Reverse Transcriptase (Superscript II, LifeTechnologies). Ekspressionen af de østradiol-regulerede gener ER, ER og lactoferrin og det interne kontrolgen ribosomal 18S (r18S) blev målt ved hjælp af genspecifikke primersæt og fluorescerende probersæt (DNA Technologies, Research Park, Aarhus). Da det viste sig, at β2-globulingenet responderede på østradioleksponering og derfor ikke kunne anvendes som intern kontrol anvendtes i stedet r18S genet som intern kontrol. Parallelt i hver PCR-kørsel blev analyseret en standardkurve med fortyndinger af plasmid DNA indeholdende det specifikke genfragment som testes. Desuden blev der parallelt i hver PCR-kørsel analyseret en kendt positiv kontrol. De genspecifikke værdier blev normaliseret til den kendte positive kontrol (for at udligne forskelle imellem kørsler fra dag til dag) og derefter til r18S niveauet (intern kontrol). Hver prøve blev analyseret i mindst 2 uafhængige målinger for hvert gen. Dosis-respons effekt af østradiolbenzoat på uterus ERα/β mRNA genekspression ses på Fig 2.2.5. Ved eksponering af rotter med stigende østradiolbenzoat (0,5 – 4 μg/kg/dag) blev der observeret et signifikant (t-test; p 0,05) fald i både ER og uterus mRNA i forhold til kontroldyr. Resultatet støtter hypotesen om en negativ feedback autoregulering af aktiveret ER.
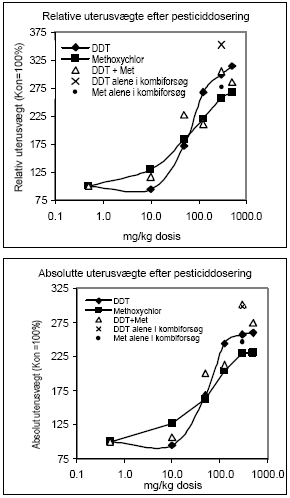
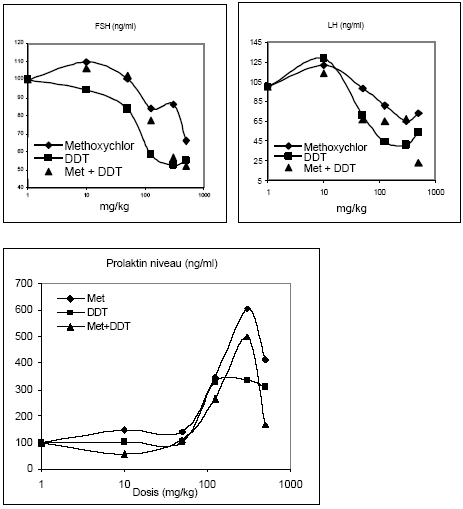
Figur 2.2.5. Dosis-respons af østradiolbenzoat på uterus ERα/β genekspression. Kontrolgruppen har fået jordnøddeoli.Resultatet er gennemsnit af n= 5 dyr, undtagen for det højest doserede hold hvor n=4. *: p 0,05. Det skal bemærkes, at ekspressionsniveauet af mRNA for ER er ca. 1000 gange lavere end for ER. Dette kan ikke ses på diagrammerne pga. normalisering til den kendte positive kontrol. 2.2.4 Effekt på neurotoksiske parametreHjernens signalstoffer (neurotransmittere) er af essentiel betydning for dennes organogenese, kønsdifferensiering og funktion under udvikling og i voksne individer. Vigtige CNS-neurotransmittere er acetylcholin (ACh), noradrenalin (NA), dopamin (DA) og 5-hydroxytryptamin (5HT). Forstyrrelse af funktionen af neurotransmittere kan give anledning til fejludvikling og funktion af hjernen. Indenfor forskningsområdet vedrørende de hormonforstyrrende stoffer ved man endnu meget lidt om stoffernes neurotoksiske effekter og effekter på adfærd, men det er et af de områder, som der højst sandsynligt vil blive fokuseret mere på fremover. Derfor har vi som noget nyt inkluderet disse parametre i dyreforsøgene for at få belyst om nogen af de kendte østrogene stoffer påvirker hjernens funktion. De neurokemiske parametre, der er blevet undersøgt for i udvalgte hjerneregioner, inkluderer neurotransmitterne noradrenalin, dopamin og 5-hydroxytryptamin (Lam et al., 1992), der anvendes som markører på effekt på hjernens noradrenerge, dopaminerge og serotonerge system. Cholinesteraseaktivitet i hjernevæv blev ligeledes målt (Ellman et al., 1961). Acetylcholinesterase (AChE) spalter og inaktiverer signalstoffet acetylcholin (ACh) når dette frigives ved impulsledning i det cholinerge nervesystem. Når dette enzym hæmmes, ophører effekten af ACh meget langsommere, hvilket er den tilgrundliggende mekanisme for flere pesticiders neurotoksiske effekt. Også andre cholinesteraser kan hæmmes af pesticider, f.eks. buturylcholinesterase (BuChE). I de gennemførte forsøg på hunrotter er der foretaget NA, DA, 5HT, AChE og BuChE analyser af alle prøverne af totalhjerner. Resultaterne indikerer, at østradiol ikke påvirker totalhjernens vægt, NA-, DA-, 5HT-koncentration eller AChE- eller BuChE-aktivitet i hunrotter, der har fået fjernet ovarierne. 2.2.5 Diskussion og konklusion på de østradiol-inducerede effekter in vivoOverordnet set gav østradiol-benzoat anledning til de forventede effekter, idet der sås en markant stigning i uterusvægten og hormonanalyserne viste et øget østradiolniveau med stigende dosis, et øget prolaktinniveau og et faldende FSH-niveau. Metoderne var dermed indkørt og kvalitetskontrollen viste, at procedurerne fungerer. Genekspressionsanalyserne viste at ovariektomi af dyrene bevirker en opregulering af både ERα og ERβ (se Fig 2.3.3), mens stigende doser østradiol forårsager en nedregulering af begge gener, hvilket er i overensstemmelse med teorien om, at østradiolniveauet i serum er afgørende for, hvor mange receptorer der dannes. Tidligere cellekulturstudier har vist, at østradiol nedregulerer ERα mRNA og proteinniveau (Vienonen et al. 2003; Clayton et al. 1997; Donaghue et al. 1999; Hofmeister & Bonefeld-Jørgensen, 2003). Hypotesen er, at der foregår en negativ autoregulerende feed-back involverende en hæmning af ER gentranskription, nedsat mRNA stabilitet og en induceret proteolyse af ER protein. Nylige tidsstudier i MCF-7 cellekultur har vist en nedregulering af ERα mRNA og en opregulering efterfulgt af nedregulering af ERβ mRNA ved eksponering for 17β-østradiol. Derudover er set effekter på ERα/β mRNA niveau af en række pesticider inklusive prochloraz (Hofmeister & Bonefeld-Jørgensen, 2003; Grünfeld & Bonefeld-Jørgensen, 2003). I dette studie blev der efter eksponering af ovariektomerede hunrotter for østradiolbenzoat, observeret en signifikant nedregulering af ERα og ERβ mRNA i uterusvæv allerede ved laveste koncentration (0,5 μg/kg/dag). Sammenholdt med, at der ligeledes blev observeret en statistisk signifikant stigning af uterusvægt i de samme østradiolbenzoateksponerede rotter ved samme lave koncentration, tyder disse resultater på at effekter på uterus (ER genekspression og vægt) er relativt følsomme parametre, som sandsynligvis kan respondere på endnu lavere doser østradiol. Uterusvægten og ER mRNA niveauer viste sig klart at være følsomme markører for den østrogene effekt end nogen af hormonanalyserne. Som noget nyt blev testet for neurotoksiske effekter af de østrogene stoffer, hvor det var et åbent spørgsmål om vi ville finde effekt eller ej. Data indikerer, at østradiol ikke påvirker niveauerne af de valgte transmitterstoffer. 2.3 Østrogene pesticider testet in vivo2.3.1 Effekt på organvægteTre dosis-responskurver for østrogene effekter in vivo for hhv. o,p-DDT, methoxychlor og stofferne i en 1:1 kombination blev udført. Den samme dyremodel som beskrevet under 2.2 blev anvendt. Hver gruppe indeholdt 6 dyr og pesticiderne blev doseret oralt i 4 dage i jordnøddeolie med et doseringsvolumen på 5 ml/kg kropsvægt. Pesticider blev doseret med 10, 50, 125, 300 og 500 mg/kg/dag hver for sig og i en 1:1 blanding. Ethinyløstradiol (doseret oralt) 100 μg/kg blev inkluderet i hvert forsøg og gav det forventede positive respons. Nyre-, hjerne- og hypofysevægte var uændrede, mens levervægten steg med både o,p-DDT og methoxychlor hver for sig og sammen. Kropsvægten var faldende for de højeste doser af begge stoffer og der sås toksisk effekt på dyrene doseret med hhv. o,p-DDT 500 mg/kg og o,p-DDT plus methoxychlor 500 mg/kg, mens dyr doseret med methoxychlor 500 mg/kg alene ikke havde iøjnefaldende påvirket almentilstand. Uterusvægten af kontroldyrene lå i de tre forsøg på hhv. 91, 79 og 107 mg. Data er normaliserede til disse værdier og både absolutte og relative uterusvægte er afbildet (Fig 2.3.1). Ved den laveste dosis på 10 mg/kg sås en effekt på uterusvægten af methoxychlor, men ikke af o,p-DDT. Derimod sås en mere markant effekt af o,p-DDT ved de højeste doser. Grunden til, at de relative uterusvægte stiger en anelse mere ved de højere doser end de absolutte vægte skyldes den faldende kropsvægt af dyrene.
Figur 2.3.1 2.3.2 Effekt på hormonniveauerRotte luteiniserende hormon (LH) og follikelstimulerende hormon (FSH) blev analyseret ved hjælp af Delfia tidsforsinket fluorescens (Haavisto et al. 1993) (Fig 2.3.2). Niveauerne af begge hypofysehormoner faldt med stigende doser af o,p-DDT og methoxychlor. FSH niveauerne i kontroldyrene lå mellem 33-39 ng/ml, mens LH niveauerne i kontroldyrene lå mellem 12-20 ng/ml. For begge hormoner sås en tendens til at o,p-DDT havde den største hæmmende effekt. For de dyr, der var doseret med både o,p-DDT og methoxychlor, lå niveauerne af de to hormoner generelt imellem niveauerne for enkeltstofferne. Tages den biologiske variation og dermed spredningen på resultaterne i betragtning, der illustreres af, at CV% for FSH niveauerne i gennemsnit var 21%, mens den tilsvarende gennemsnitlige CV% for LH niveauerne var 47%, kan der ikke påvises nogen signifikant forskel mellem effekt af enkeltstofferne og af blandingen. For at teste om der var statistisk forskel på DDTs og methoxychlors effekt på LH niveauer blev det v.hj.a. en F-test testet om de to regressionslinier for dosis-responskurverne var signifikant forskellige fra hinanden. Der kunne ikke påvises nogen signifikant forskel på DDTs og methoxychlors effekt på serum LH niveau. Både o,p-DDT og methoxychlor gav anledning til stigende prolaktinniveauer i serum, men effekten var først markant ved en dosis på 125 mg/kg (fig 2.3.2). Generelt var der ikke nogen påviselig forskellig effekt af blandingen af pesticiderne sammenlignet med enkeltstofferne.
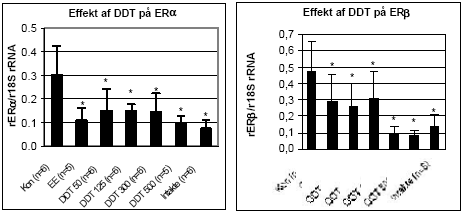
Figur 2.3.2 2.3.3 Effekt på østrogen-regulerede generEffekt af o,p-DDT og methoxychlor på ER mRNA i uterus blev analyseret v.hj.a. den metode der er beskrevet under 2.2.3. mRNA blev oprenset, cDNA syntetiseret og PCR analyser for ERα og ERβ blev udført. Effekten af o,p-DDT på uterus ER/ genekspression ses på Fig 2.3.3. En signifikant (t-test; p 0,05) nedregulering af ERα/β genekspression blev observeret for 100 μg/kg/dag oral eksponering af ethinylestradiol og for alle doser af o,p-DDT (50 mg/kg til 500 mg/kg) i forhold til kontroldyr, som fik indgivet jordnøddeolie. De intakte dyr er ikke-ovariektomerede dyr, som har fået jordnøddeolie. Resultatet viser dermed, at i normale intakte dyr er ERα og ERβ mRNA nedreguleret af østrogener produceret i ovarierne til et niveau svarende til indgivelse af 100μg/kg/dag af ethinylestradiol eller 50 mg/kg/dag o,p-DDT.
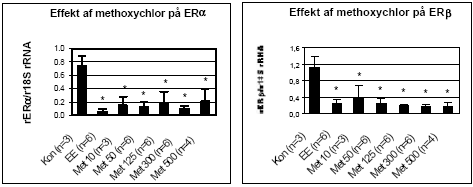
Figur 2.3.3. Effekt af o,p-DDT på uterus ERα og ERβ genekspression. Resultatet er gennemsnittet af det antal dyr, som er angivet i parentesen (n=) under de forskelligegrupper. Kontroldyr: olie; EE: 100 μg/kg/dag ethinyløstradiol; DDT 50: 50 mg/kg etc. for de andre grupper, hvor tallet angiver den totale orale dosis i mg/kg/dag. Alle eksponeringer var signifikant (p 0,05) forskellig fra kontrollen. Da o,p-DDT er vist at udøve en østrogen effekt ved såvel uterus vægtstigning som i MCF7-celleproliferationsanalysen, styrkes hypotesen, at østrogene stoffer der aktiverer ER medfører en negativ feedback autoregulering af ER transkription (mRNA niveau) som er gældende såvel in vitro som in vivo i uterusvæv. Resultatet for ERα og ERβ genekspression i uterus efter eksponering af rotter for methoxychlor ses i Fig 2.3.4. En signifikant nedregulering af begge ER receptorer blev observeret for alle testede koncentrationer.
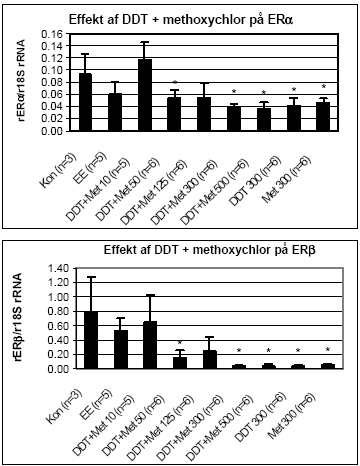
Figur 2.3.4. Effekt af methoxychlor på uterus ERα/β genekspression. Resultatet er gennemsnittet af det antal dyr, som er angivet i parentesen (n=) under de forskellige grupper. Kontrol: olie; EE: 100 μg/kg/dag ethinyløstradiol; Met 10: 10 mg/kg etc. for de andre grupper, hvor tallet angiver den totale orale dosis i mg/kg/dag. Alle eksponeringer var signifikant (p 0,05) forskellig fra kontrollen.
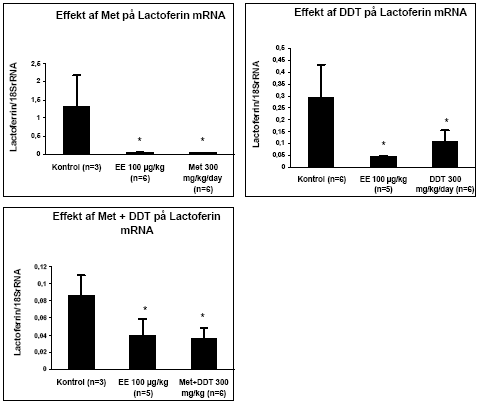
Figur 2.3.5. Effekt af en blanding af o,p-DDT og methoxychlor på uterus ERα/β genekspression. Resultatet er gennemsnittet af det antal dyr, som er angivet i parentesen (n=) under de forskellige grupper. Kon: jordnødde olie; EE: 100 μg/kg/dag ethinyløstradiol;DDT + Met 10: 5 mg/kg DDT plus 5 mg/kg methoxychlor givet oralt. Tilsvarende gælder for de andre grupper, hvor tallet angiver den totale dosis i mg/kg/dag. Alle eksponeringer på 50mg/kg og derover var signifikant (p 0.05) forskellig fra kontrollen. I kombinationsforsøget gav den positive kontrol ethinyløstradiol ikke noget signifikant respons på ERα/β i modsætning til de to tidligere forsøg. Det kan muligvis skyldes, at det kun var muligt at isolere RNA fra 3 kontroldyr i dette forsøg, hvilket resulterer i en høj standardafvigelse. Sammenlignes effekten på både ERα og ERβ af 300 mg/kg af enkeltstofferne (fig. 2.3.3 og fig. 2.3.4) med effekten af blandingen af samme koncentration (fig 2.3.5) ses, at effekten ikke er signifikant forskellig, hvilket indikerer additivitet af stofferne også på gen-niveau. Effekten af o,p-DDT og methoxychlor på lactoferin mRNA blev undersøgt for udvalgte grupper (Fig 2.3.6). Resultaterne viste, at begge pesticider ved en dosis på 300 mg/kg sænkede lactoferin mRNA til et niveau svarende til det, der blev opnået med den positive kontrol ethinyløstradiol. I kombinationsforsøget reducerede både ethinyløstradiol og o,p-DDT og methoxychlor 300 mg/kg hver for sig og sammen signifikant lactoferin mRNA niveau.
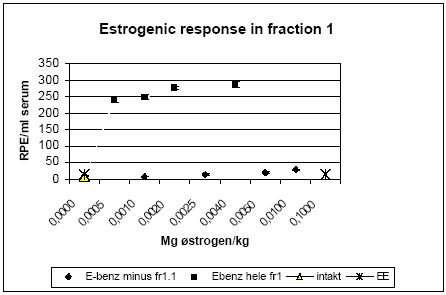
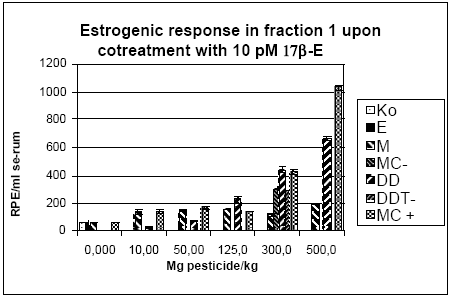
Figur 2.3.6. Effekt af henholdsvis methoxychlor (Met 300 mg/kg), o,p-DDT (300 mg/kg) og stofferne i kombination (150+150 mg/kg) på lactoferin mRNA i rotteuterus. 2.3.4 Effekt på neurotoksiske parametreNeurotransmitteranalyse blev foretaget for forsøgene med o,p-DDT, methoxychlor og kombinationer af stofferne på samme måde som beskrevet i 2.2.4. Data indikerer at hverken o,p-DDT, methoxychlor eller kombinationerne heraf påvirker totalhjernens vægt, proteinkoncentration, NA-, DA-, 5HT-koncentration eller AChE- eller BuChE-aktivitet. 2.3.5 Biomarkør for xenoøstrogent respons i serumPå afdeling for Miljømedicin ved Syddansk Universitet er en biomarkørmetode til måling af xenoøstrogent respons i humant serum indkørt (Rasmussen et al, 2003). Metodens princip er, at endogene østrogener fjernes fra serum ved fastfase ekstraktion og efterfølgende HPLC-fraktionering. Fraktioner uden endogene østrogener testes efterfølgende for østrogen aktivitet i MCF-7 celleproliferationsassayet. Serum fra rotter, der indgik i forsøgene for østrogene kombinationseffekter, er blevet testet ved hjælp af denne metode. Indledningvist blev metoden til rotteserum indkørt ved at teste serum fra rotter doseret med det positive kontrolstof østradiolbenzoat. Der var problemer med et østrogent stof der eluerede i den serumfraktion, hvor en del af pesticiderne eluerede og hvor der ikke i humant serum eluerer østrogener. Det viste sig, at skyldes østradiolbenzoat, som dyrene blev doseret med, og som ikke tilhører de fysiologiske østrogener. Metoden er herefter blevet modificeret, så den fraktion som indeholder østradiolbenzoat blev fjernet i de forsøg hvor rotterne er doseret med østradiolbenzoat som positiv kontrol, inden serumprøven blev testet i MCF-7 celleassayet (Fig 2.3.7). Serum fra hunrotter behandlet med o,p-DDT og methoxychlor alene og i kombination blev analyseret v.hj.a. denne metode. I pesticidforsøgene blev ethinyløstradiol anvendt som positivt kontrolstof, hvorfor den oprindelige fraktioneringsmetode for serumprøverne kunne anvendes. For at få en tilstrækkelig mængde serum til HPLC-fraktioneringen har det været nødvendigt at poole serum fra dyrerne i hver doseringsgruppe. Den poolede serumprøve er herefter blevet fraktioneret. De fraktioner som indeholdt de fysiologiske østrogener samt ethinyløstradiol blev kombinerede og kaldes Fraktion 2. De resterende fraktioner, hvor pesticiderne eluerede, blev kombineret og kaldes Fraktion 1. Begge fraktioner er blevet testet både med og uden 17β-østradiol i en fortyndingsrække, for at sikre at responset ligger indenfor 17β-østradiol standardkurven (som medtages i hvert set-up) og at prøven ikke har toksiske effekter i assayet. Hver fortynding blev testet i tripelbestemmelse. Den fortynding, som gav det højeste respons, er anvendt til de videre beregninger. For fraktion 1 fra negative kontroller (doseret med jordnøddeolie) og postive kontroller sås det maximale respons ved en 5 ganges fortynding. For grupperne doseret med methoxychlor og o,p-DDT (alene og i kombination) sås det maksimale respons ved en 10 ganges fortynding. Fraktion 2 er testet for at sikre at HPLC-fraktioneringen har virket korrekt, dvs at det forventede respons fra ethinyløstradiol ses i denne fraktion og at der ikke er forhøjet respons i de pesticideksponerede dyr. Dette var opfyldt for alle grupperne. Nedenfor vises resultaterne for fraktion 1. Resultaterne er udtrykt som den Relative Proliferative Effekt (RPE) per ml serum efter korrektion for fortynding samt den mængde af serum, der blev fraktioneret. RPE = (PEserumprøve-1)/( PE max 17β-østradiol – 1), hvor PE er responset normaliseret til kontrol.
Figur 2.3.7. Respons i MCF-7 celleproliferationsassayet fra HPLC-fraktioneret rotteserum. Fraktion 1 repræsenterer serumprøven efter fjernelse af de endogent producerede østrogener. Intakt: er 'hele' ikke-ovariektomerede dyr. E-benz-fr1.1 er dyr doseret med østradiolbenzoat (s.c.) og hvor en subfraktion indeholdende østradiolbenzoat er fjernet.
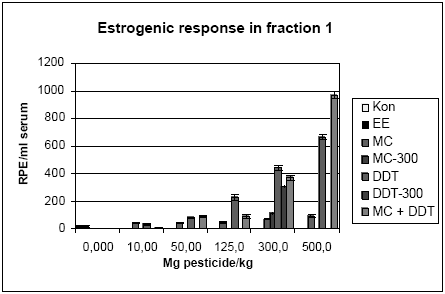
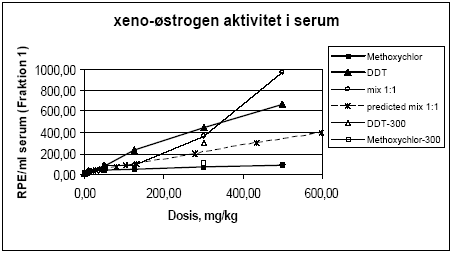
Figur 2.3.8. Respons i MCF-7 celleproliferation assayet fra HPLC-fraktioneret serum.Kon: olie; EE: 100 μg/kg/dag ethinylestradiol; MC: methoxychlor. MC-300 og o,p-DDT-300 er grupper der indgik i kombinationsforsøget men som blev doseret med methoxychlor eller o,p-DDT enkeltvis i en dosis på 300 mg/kg. For hver pesticid doseringsgruppe repræsenterer resultatet responset fra serum poolet fra de 6 dyr i gruppen. For Kon og EE repræsenterer resultatet gennemsnittet for 3 uafhæbgige grupper hidrørende fra de tre forskellige pesticidforsøg. Da der kun er testet en serumprøve per doseringsgruppe er der ikke udført statistiske analyser. Der ses dog et tydeligt dosis-relateret respons for begge pesticider samt for kombinationen af de to pesticider. I lighed med in vitro forsøgene i MCF-7 celle assayet (Fig 2.1.1) ses et kraftigere respons for o,p-DDT end for methoxychlor, når stofferne er doseret enkeltvis til dyrerne. Den kraftigste stigning i respons ses for gruppen doseret med kombinationer af methoxychlor og o,p-DDT. Stigningen i det målte respons er højere end man ville forvente hvis man analyserer resultaterne ved hjælp af isobolmetoden (Fig 2.3.9). Dette kunne tyde på en synergistisk effekt af de to pesticider på det xeno-østrogene respons. Der er imidlertid stor usikkerhed på målingerne, idet der kun er et datapunkt per dosis og fordi der ikke for alle dosisgrupper var 4,0 ml serum til rådighed for analyserne, som er den mængde serum assayet er valideret ved. Generelt var der mindre serum ved de højeste dosisgrupper og for den gruppe rotter som blev doseret med 500 mg/kg af 1:1 blandingen af o,p-DDT og methoxychlor var der kun 0,95 ml serum. Ved beregningen af resultaterne er der taget højde for disse forskelle i prøvemængde men der vil være større usikkerhed på målingerne ved de lave prøvemængder. Den observerede synergistiske effekt skal derfor tolkes med forsigtighed. En mulig biologisk forklaring kunne være at o,p-DDT og methoxychlor måske påvirker hinandens metabolisme så der enten sker en langsommere nedbrydning af o,p-DDT eller en øget dannelse af østrogene metabolitter af methoxychlor, idet methoxychlor vides at blive omdannet in vivo til en række metabolitter hvoraf mindst en er et mere potent østrogen end methoxychlor selv (Gaido et al., 1999).
Figur 2.3.9. Det observerede respons i fraktion 1 fra serum fra rotter doseret med methoxychlor og o,p-DDT enkeltsvist og i 1:1 kombinationer sammenlignet med det prædikterede respons for 1:1 kombinationerne under antagelse af additivitet beregnet ud fra isobolmetoden. For at undersøge eventuelle antiøstrogene eller additive effekter af pesticiderne overfor 17β-østradiol blev fraktion 1 også testet sammen med 10 pM 17β-østradiol på samme måde som i in vitro forsøgene (Fig 2.1.2).
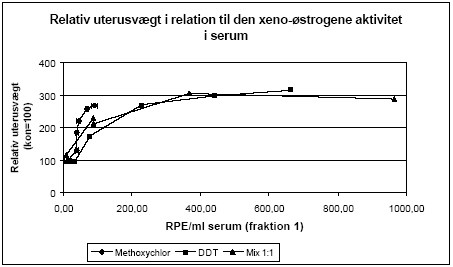
Figur 2.3.10. Respons i MCF-7 celle proliferations assayet fra HPLC-fraktioneret serum testet sammen med 10 pM 17β-østradiol.. Kon: olie; EE: 100 μg/kg/dag ethinylestradiol; MC: methoxychlor. MC-300 og o,p-DDT-300 er grupper, der indgik i kombinationsforsøget, men som blev doseret med methoxychlor eller o,p-DDT enkeltvis i en dosis på 300 mg/kg.For hver pesticiddoseringsgruppe repræsenterer resultatet responset fra serum pooled fra de 6 dyr i gruppen. For Kon og EE repræsenterer resultatet responset gennemsnittet fra 3 uafhængige grupper hidrørende fra de tre forskellige pesticidforsøg. Resultaterne stemmer godt overens med in vitro resultaterne, idet der ses en stigning i MCF-7 celleproliferationen, når serum fra de pesticidbehandlede dyr testes (fig 2.3.10). Dette tyder på at pesticiderne øger det samlede østrogene respons i dyrerne, mens der ikke er tegn på en antiøstrogen virkning. Sammenholdes den relative stigning i uterusvægten hos de doserede dyr med det xeno-østrogene respons i serum ses en stigning i uterusvægten ved RPE-værdier op til ca. 400. Herefter flader kurven ud, hvilket indikerer en mætning af vægtresponset i uterus. I en undersøgelse hvor effekten af lave doser af o,p-DDT og β-HCHs på epithelhøjde i uterus og vaginal epitheltykkelse i mus blev undersøgt som mål for østrogen virkning, sås ligeledes et plateau for epithelhøjden i uterus når blodkoncentrationen nåede et bestemt niveau af β-HCH (Ulrich et al., 2000) mens et tilsvarende plateau ikke sås for o,p-DDT ved de anvendte doseringer. Pesticiderne blev doseret ved subkutan implantation af kapsler hvorfra pesticiderne langsomt blev frigivet og blodkoncentrationerne af pesticiderne var væsentlig lavere end i vores undersøgelse, hvor den maksimale o,p-DDT koncentration kan estimeres til ca. 750 μg/ml serum (vurderet ud fra HPLC-målingerne) mod 622 ng/ml i den refererede undersøgelse.
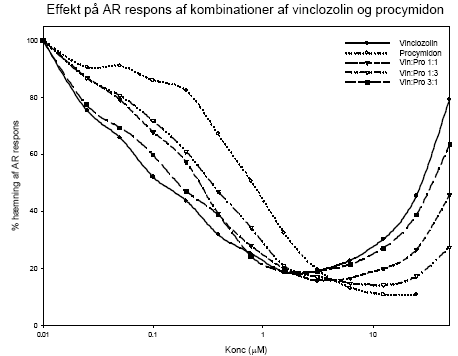
Figur 2.3.11.Stigning i relativ uterusvægt som funktion af respons i MCF-7 celle proliferation assayet fra HPLC-fraktioneret. De opnåede resultater viser, at biomarkørmetoden er anvendelig til at undersøge det samlede østrogene respons fra xeno-østrogener i en serumprøve. Responset ved de højeste pesticiddoser kan i denne undersøgelse være overestimeret, især for gruppen eksponeret for blandingen af de to pesticider, fordi der ikke var den optimale mængde serum til analysen. Under alle omstændigheder ses en klar dosis-relateret stigning i det xeno-østrogene respons i serum. Fordelen ved metoden er, at den tager højde for optagelse i organismen samt dannelse af eventuelle østrogene metabolitter samt nedbrydningen af pesticiderne til inaktive substanser. Den er således egnet til at vurdere den samlede interne eksponering for østrogene stoffer, herunder også af komplekse blandinger. 2.3.6 Diskussion og konklusion på de pesticid-inducerede østrogene effekterI in vitro forsøgene er der god overenstemmelse mellem de målte effekter og de predikterede effekter for additiv virkning beregnet ud fra isobolmetoden. For methoxychlor og o,p-DDT indikerer resultaterne, at der ikke er synergistisk eller antagonistisk interaktion mellem de to pesticider in vitro men en klar additiv virkning. Dette tyder på, at de to pesticiders østrogene virkning skyldes samme virkningsmekanisme, nemlig direkte interaktion af stofferne med ER. Ligeledes indikerer resultaterne for blandinger af methoxychlor, fenarimol og vinclozolin, at de to pesticider med østrogen virkning, fenarimol og methoxychlor, udviser additivitet, mens vinclozolin, som ikke er østrogent, ikke påvirker responset i blandingerne. Også andre in vitro undersøgelser af blandinger af østrogene stoffer har tydeligt vist additivt respons under anvendelse af koncentrations-additionsmodellen. Dette gælder således ved testning af o,p-DDT, p,p-DDT, p,p-DDE og β-HCH i MCF-7 celleassayet (Payne et al., 2001) og for 2-, 3- og 4- komponentblandinger af o,p-DDT, genistein, 4nonylphenol og 4-n-octylphenol i et ER reportergen assay (Payne et al., 2001). I in vivo forsøgene med o,p-DDT og methoxychlor er undersøgt effekt på uterusvægt, effekt på FSH-, LH- og prolaktinniveauer, effekt på ERα og ERβ mRNA i uterus og på udvalgte neurotransmittere og cholinesteraser i hjernevæv. Kort fortalt ses med stofferne en stigning i uterusvægt og prolaktinniveau, mens LH-, FSH-niveau og ERα og β mRNA falder. Effekten af både o,p-DDT og methoxychlor på uterusvægt og LH niveauer sås ved den næstlaveste dosis på 50 mg/kg. Derimod sås effekt allerede ved 10 mg/kg methoxychlor på genekspressionsniveau, hvilket indikerer at dette er den mest følsomme af de undersøgte parametre. Der ses ikke umiddelbart effekt på neurotransmitterniveauer og acetylcholinesteraseaktivitet. Generelt kan der ikke ses bort fra, at den uundgåelige biologiske variation mellem dyrene i disse forsøg bevirker, at en meget præcis og detaljeret undersøgelse af samspilseffekterne er vanskelig og understreger behovet for i in vivo forsøg at undersøge flere dosisniveauer for at få det fulde billede af stoffernes virkning hver for sig og i kombination. De hidtil opnåede data giver dog alligevel et relativt klart billede af samspillet mellem o,p-DDT og methoxychlor. På baggrund af de opnåede resultater for organvægte, hormonniveauer og ER mRNA niveau kan der ikke påvises fravær af additivitet, d.v.s. der er ikke holdepunkter for at antage, at der er nogen synergistiske eller antagonistiske samspilseffekter mellem stofferne. Dette underbygges af, at in vitro forsøgene med de samme stoffer viser samme additivitet. Baseret på in vitro og in vivo studier er en række pesticider herunder o,p-DDT og methoxychlor blevet klassificeret som potentielle miljøøstrogener (Scippo et al. 2003; Diel et al. 2002; Welsh et al. 1969; Gray et al. 1999; Brody and Rudel, 2003). I det aktuelle studie udviste både o,p-DDT og methoxychlor samt en kombination af de to pesticider en signifikant nedregulering af begge østrogenreceptorer ved de lavest testede koncentrationer henholdsvis 50 mg/kg/dag og 10 mg/kg/dag. Resultaterne indikerer, at uterus ER genekspressionsanalyser er meget følsomme og vil være i stand til at respondere til eksponering af disse pesticider ved lavere koncentrationer. Effekter på genekspression af et tredje østrogen-responsivt gen i uterusvæv, lactoferrin, blev ligeledes signifikant nedreguleret og viste den samme høje følsomhed ved eksponering til østradiol, o,p,-DDT og methoxychlor samt en kombination af de to pesticider. De observerede effekter af kombinationer af de to pesticider på ekspressionen af de tre gener indikerer, at der er tale om en additiv effekt. Sammenligning af test for østrogenicitet i tidligere in vitro studier af pesticidet prochloraz (Andersen et al., 2002) med in vitro effekter på ER α/β mRNA (Hofmeister & Bonefeld-Jørgensen, 2003; Grünfeld & Bonefeld-Jørgensen, 2003) og effekter på ERα og ERβ mRNA niveau i uterusvæv (nye upublicerede resultater, Bonefeld-Jørgensen) tyder på, at der er overensstemmelse imellem effekter observeret in vitro og in vivo. 2.4 Antiandrogene pesticider testet in vitroPesticidernes evne til at blokere androgenreceptoren blev undersøgt ved hjælp af et reportergenassay for antiandrogene effekter (Vinggaard et al. 1999). Chinese Hamster Ovary celler (CHO K1) blev udsået i mikrotiterplader og hver brønd blev transfekteret med 75 ng DNA bestående af ekspressionsvektoren pSVAR0 og et MMTV-LUC reporterplasmid i en ratio på 1:100 ved hjælp af 0.15 μl af transfektionsreagenset FuGene. Androgenreceptoragonisten R1881 (0.01 nM) plus/minus pesticider blev tilsat og efter en inkubationsperiode på 24 timer blev cellerne lyseret og aktiviteten af den dannede luciferase blev målt i et BioOrbit Galaxy luminometer. 2.4.1 Effekt af kombinationer af vinclozolin og procymidonDer er udført forsøg til belysning af kombinationseffekter af vinclozolin og procymidon for androgenreceptorinhibering in vitro. De enkelte stoffer og tre kombinationer bestående af 1:1, 1:3 og 3:1 blandinger af pesticiderne blev undersøgt i 12 koncentrationer i området 0.025 – 50 μM (Fig 2.4.1). For procymidon alene ses en antiandrogen effekt i hele koncentrationsområdet, mens der for vinclozolin ses en antiandrogen effekt op til en koncentration på ca. 3 μM, hvorefter hæmningen forsvinder. Da det er metabolitterne af vinclozolin, der er de aktive antiandrogene stoffer, kan dette tænkes at skyldes at vinclozolin-metabolitterne er partielle agonister ved høje koncentrationer, som det også er observeret for det kendte antiandrogene stof hydroxyflutamid. Overordnet set blev de forventede antiandrogene effekter for enkeltstofferne observeret og IC50 for vinclozolin og procymidon fandtes at være henholdsvis 0.1 og 0.6 μM. Data for enkeltstofferne blev normaliseret (da der ses interassay variation) og der blev lavet kurvefitning v.hj.a. non-lineær regression, hvor 'Four parameter logistic curve fit' blev anvendt (F= min + (max-min)/(1+(x/EC50)^Hill-slope)). På baggrund af de fremkomne kurve-fit for enkeltstofferne blev det forventede respons af blandingerne beregnet, under antagelse af at stofferne virker additivt. Herefter blev de forventede og rent faktisk opnåede effekter sammenlignet for 1:1, 1:3 og 3:1 blandingerne (Fig 2.4.2).
Figur 2.4.1. Androgenreceptor blokering induceret af stigende koncentrationer af vinclozolin eller procymidon eller de to stoffer i kombination bestemt i tilstedeværelse af 0.01 nM R1881. Data repræsenterer gennemsnittet af n=4-6 forsøg hver udført i firdobbelt bestemmelse. Figur 2.4.2 Sammenligning af observeret respons og prædikteret respons for androgen receptor blokering induceret af blandinger af vinclozolin og procymidon. Data repræsenterer mean ±SD for n= 4-6 forsøg.
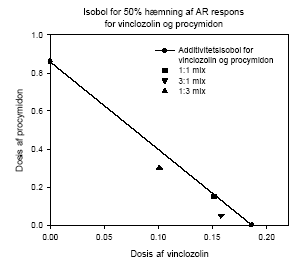
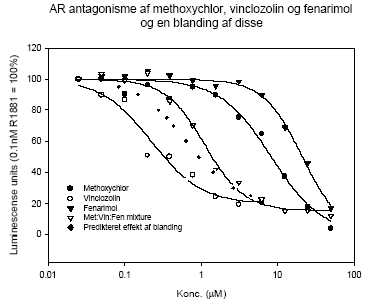
Figur 2.4.3 Additivitetsisobol for 50% hæmning af androgen receptor aktivering for blandinger af vinclozolin og procymidon til sammenligning med de faktisk observerede effekter af blandingerne. Isobolen som afbilder de effekter der angiver 50% hæmning af det R1881-inducerede respons ses i fig 2.4.3. Den dosis af vinclozolin der giver 50% androgen receptor blokering er afbildet på x-aksen, mens den tilsvarende dosis af procymidon er angivet på y-aksen. Punkterne for blandingerne ses at ligge tæt op ad additivitets-isobolen, som er den rette linie mellem punkterne på x- og y-aksen. Resultaterne i fig 2.4.2 og 2.4.3 indikerer, at vinclozolin og procymidon virker additivt og at der ikke er nogen interaktion mellem stofferne. I dette tilfælde opnås således i in vitro forsøg det predikterede additive respons, hvilket indikerer at der ikke umiddelbart er nogen grund til at tro, at synergistiske eller antagonistiske samspilseffekter for disse antiandrogene stoffer finder sted ved blokering af androgenreceptoren in vitro. 2.4.2 Effekt af kombinationer af vinclozolin, methoxychlor og fenarimolFormålet med dette studie var at undersøge kombinationeffekt in vitro af 3 pesticider med forskellig virkningsmekanisme. Udover at de 3 pesticider alle virker antiandrogent in vitro og hæmmer aktivering af androgen receptoren, virker både methoxychlor og fenarimol også østrogent in vitro og fenarimol har deuden den egenskab at den hæmmer aromataseaktivitet. Resultaterne i fig 2.4.4 viser, at når den forventede effekt af blandingen sammenlignes med den rent faktisk observerede effekt af blandingen under forudsætning af additivitet, så ses en effekt der er mindre end en additiv effekt. Isobolkoefficienterne som er opgivet i tabellen er generelt større end 1 og samlet indikerer disse data at de 3 pesticider virker antagonistisk i dette assay. En forklaring på dette kan være at den metaboliske kapacitet i cellerne har nået sin grænse, idet det gælder for både vinclozolin og methoxychlor at det er metabolitterne, der er de mest aktive antiandrogene stoffer.
Figur 2.4.4 Koncentrations-responskurver for hhv. vinclozolin, methoxychlor og fenarimol og en blanding af stofferne i æquimolært forhold (1:1:1). Androgenreceptor inhibering er analyseret i reportergen assay i tilstedeværelse af 0.1 nM R1881. Data repræsenterer 3 uafhængige forsøg udført i 4-dobbelt bestemmelse.
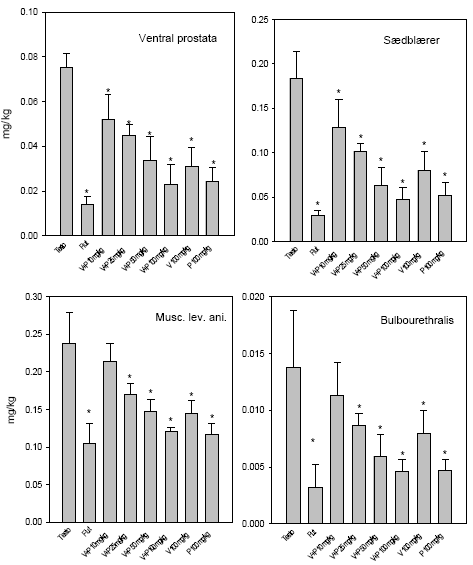
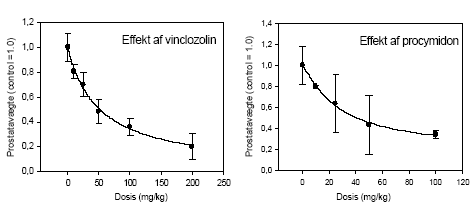
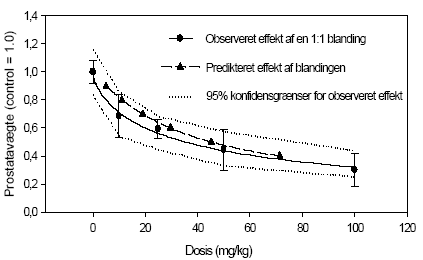
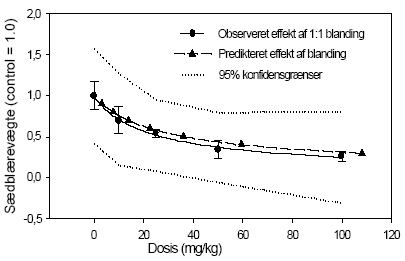
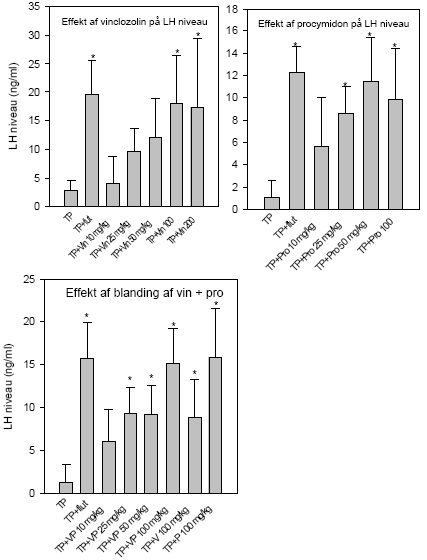
2.5 Antiandrogene pesticider testet in vivoAntiandrogene effekter in vivo er undersøgt ved hjælp af den klassiske Hershberger test. Metoden er baseret på ikke-kønsmodne kastrerede Wistar hanrotter (28 dage gamle ved kastrationen) som doseres dagligt i 7 dage (fra dag 42-48) med testosteron med og uden teststof. Det kendte antiandrogen flutamid bruges som den positive kontrol. Blod opsamles til hormonanalyser og reproduktionsorganerne: ventral prostata, sædblærer, musc. levator ani/bulbocavernosus og bulbourethralkirtlen udtages og vejes. Ventral prostata gemmes til efterfølgende genekspressionsanalyse. I kombinationsforsøget blev hjerner udtaget til analyser for neurotransmittere. Denne dyreforsøgsmodel er etableret på DFVF (Hossaini et al., Abstract 2000; Vinggaard et al. 2002) og har indgået som en del af et interkalibreringsstudie i OECD regi. Der er i alt udført 3 dosis-respons forsøg, 1 med vinclozolin og 1 med procymidon alene (0, 10, 25 ,50, 100 og 300 mg/kg) og 1 kombinationsforsøg med en 1:1 blanding i samme doser som beskrevet for enkeltstofferne. I det sidste forsøg blev også inkluderet to grupper, der fik administreret pesticiderne alene i en dosis på 100 mg/kg. 2.5.1 Effekt på organvægteEffekterne af vinclozolin på vægten af ventral prostata, sædblærer, musc.lev.ani og bulbourethralis var signifikant forskellige ved doser på hhv. 10, 10 , 25 og 25 mg/kg (Fig 2.5.1). Effekterne af procymidon på de tilsvarende organer var signifikant allerede ved 10 mg/kg i alle tilfælde. Kombinationen (1:1 blanding) af de to stoffer havde effekt på ventral prostata og sædblærer ved 10 mg/kg, mens effekten på musc.lev. ani/bulbocavernosus og bulbourethralkirtlen først opstod ved en 25 mg/kg dosis. Analyse af kombinationseffekterne på prostatavægte og sædblærevægte v.hj.a. isobolmetoden og efterfølgende sammenligning af prædikteret og faktisk opnået respons (fig 2.5.3 og 2.5.4) viser, at der er grundlag for at antage at de antiandrogene stoffer også in vivo virker additivt. Der er en tendens til - ved de lavere doser - at effekten er en smule kraftigere end den predikterede, men SD'erne for det opnåede respons overlapper det predikterede, og endelig ligger det predikterede respons (under antagelse af additivitet) indenfor 95% konfidensgrænserne for det faktisk opnåede respons.
Figur 2.5.1. Effekter af stigende doser af en 1:1 blanding af vinclozolin og procymidon på vægte af hhv. ventral prostata, sædblærer, m. levator ani/bulbocavernosus og bulbourethralkirtlen. De sidste to søjler i hver figur viser effekten af hhv. vinclozolin 100 mg/kg og procymidon 100 mg/kg doseret enkeltvis til dyrene og skal sammenlignes med den 3. sidste søjle hvor de er givet i kombination (50+50 mg/kg). Hver søjle repræsenterer middel ±SD for n=6 dyr.
Figur 2.5.2. Dosis-responskurver for effekt på prostatavægte af hhv. vinclozolin og procymidon. Se i øvrigt figurteksten 2.5.1.
Figur 2.5.3. Sammenligning af observeret effekt og forventet effekt under antagelse af additivitet på prostatavægte for en 1:1 blanding af vinclozolin og procymidon.
Figur 2.5.4. Sammenligning af observeret effekt og forventet effekt under antagelse af additivitet på sædblærevægte for en 1:1 blanding af vinclozolin og procymidon. 2.5.2 Effekt på hormonniveauerLH og FSH i serum blev analyseret v.hj.a. Delfia metoden (af Pirji Pakarinen, Turku Universitet, Finland). Begge pesticider gav som ventet en signifikant stigning i LH niveau, der opstod ved hhv. 100 mg/kg og 25 mg/kg for vinclozolins og procymidons vedkommende (fig 2.5.5). Ved dosering med en blanding af pesticiderne sås signifikant effekt ved doser på 25 mg/kg og opefter. Mht. FSH analyserne sås generelt mindre markante effekter end de der blev opnået med LH, men der sås en signifikant stigning af FSH for både vinclozolins og procymidons vedkommende ved 25 mg/kg (data ikke vist). Der var dog ikke nogen klar dosis-respons sammenhæng. I kombinationsforsøget sås en statistisk signifikant effekt allerede ved en total dosis på 10 mg/kg. Generelt for disse FSH data gælder, at der ikke fås nogen pæn dosis-responskurve for nogen af stofferne, f.eks. ses effekt af 50 og 200 mg/kg vinclozolin, men ikke af 100 mg/kg. For procymidons vedkommende ses effekt ved 25 mg/kg, men ikke ved andre doser. I kombinationsforsøget er alle doser derimod signifikant forhøjede, og der er åbenbart opnået maksimal effekt allerede ved laveste dosis for der er ingen hældning på kurven. Pga. den manglende dosis-respons effekt på FSH niveau og pga. de mindre konsistente data, kan der ikke drages nogen endelig konklusion udfra disse data.
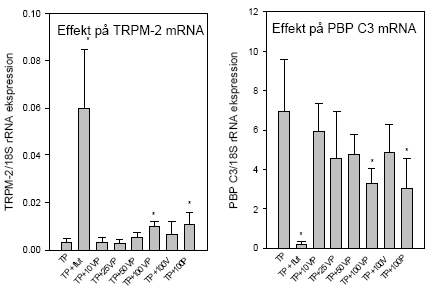
Figur 2.5.5 Effekt af pesticider på serum niveau af luteiniserende hormon (LH). Øverst til venstre:Dosis-respons af vinclozolin alene. Øverst til højre: Dosis-respons af procymidon alene. Nederst: Dosis-respons af 1:1 blandinger af vinclozolin og procymidon plus en enkelt gruppe der fik pesticiderne hver for sig (100 mg/kg).'TP': testosteronpropionat 'Flut': flutamid. 2.5.3 Effekt på genekspressionLightcycler-teknologien, der er beskrevet i afsnit 2.2.3 blev anvendt til at kvantificere den relative ekspression i prostata af de androgent-regulerede gener, testosterone-repressed prostatic message 2 (TRPM-2) og prostata binding protein polypeptide C3 (PBP C3) sammenlignet med ekspressionen af 18S-mRNA (Nellemann et al. 2000). TRPM-2 udtrykkes ikke normalt i rottens ventrale prostata men induceres ved kastration og er sandsynligvis involveret i apoptosis. PBP C3 udgør ca. 50% af secerneret protein fra den normale rotteprostata og er derfor det mest forekommende androgent-regulerede gen i dette væv. Data viser, at flutamid som ventet giver et øget TRPM-2 mRNA niveau, mens PBP C3 mRNA blev signifikant nedsat (fig 2.5.6). Vinclozolin alene gav ikke noget signifikant respons ved 100 mg/kg, mens procymidon (100 mg/kg) gav signifikant forøget TRPM-2 og nedsat PBP C3 mRNA i forhold til testosteron-kontrollen. Disse resultater indikerer, at effekten på genekspressionsniveau i dette tilfælde indtræder ved højere doser end de der skal til for give effekt på organniveau og på LH niveauer.
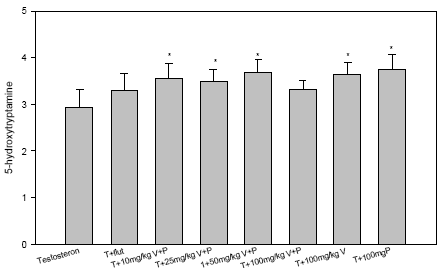
Figur 2.5.6 Dosis-respons effekt af blandingen af vinclozolin og procymidon på TRPM-2 mRNA og PBP C3 mRNA i prostata. 2.5.4 Effekt på neurotoksiske parametreHverken vinclozolin eller procymidon hver for sig eller sammen påvirkede totalhjernens vægt, proteinkoncentration, NA-, DA-koncentration eller AChE- eller BuChE-aktivitet i disse forsøg. Dette indikerer, at de noradrenerge, dopaminerge, og cholinerge dele af CNS ikke påvirkes markant af pesticiderne. Multipel statistisk analyse (Dunnett's Metode) viste, at holdene doseret med 10, 25 og 50 mg/kg vinclozolin og procymidon i kombination og med 100 mg/kg vinclozolin eller 100 mg/kg procymidon alene statistisk signifikant afviger fra kontrolholdet doseret med testosteron med hensyn til totalhjernens 5HT- koncentration (fig 2.5.7). Der er efterfølgende udtaget totalhjerner fra ikke-behandlede og testosteron-behandlede kastrerede dyr for at få en afklaring af, om testosteronbehandlingen i sig selv påvirker hjernens 5HT-koncentration i forhold til kontrolbehandling. Dette viste sig ikke at være tilfældet. 5-HT vides at regulere LHRH sekretion fra hypothalamus i hunrotter (Moguilevsky & Wuttke, 2001) og også sekretion af hypofysehormoner og en effekt på 5-HT niveauet, kan ikke udelukkes at være involveret i stoffernes effekter. Flutamid gav dog ikke nogen effekt på 5-HT niveauet, hvilket tyder på at der ikke er tale om en generel effekt af antiandrogener.
Figur 2.5.7. Effekt af blandingen af vinclozolin og procymidon på 5-hydroxytryptamin niveau (serotonin) i hjernehomogenater. 2.5.5 Diskussion og konklusion på de pesticid-inducerede antiandrogene effekterI in vitro forsøgene for kombinationer af vinclozolin og procymidon er der god overenstemmelse mellem de målte effekter og de prædikterede værdier for additiv virkning beregnet ud fra isobolmetoden. Dette indikerer, at der ikke er synergistisk eller antagonistisk interaktion mellem de to pesticider men en klar additiv virkning. Dette tyder også på, at de to pesticiders antiandrogene virkning skyldes samme virkningsmekanisme. I in vivo forsøgene med vinclozolin og procymidon er undersøgt effekt på vægt af reproduktionsorganer, effekt på FSH- og LH-niveauer, effekt på TRPM-2 og PBP C3 mRNA og på udvalgte neurotransmitterniveauer og acetylcholinesteraseaktivitet. Kort fortalt ses med stofferne et fald i vægte af alle reproduktionsorganer, en stigning i LH- og FSH-niveau og hhv. et fald og en stigning i mRNA niveauer af PBP C3 og TRPM-2. Følsomheden af de tre nævnte parametre for antiandrogener var i disse forsøg: reproduktionsorganvægte > serum LH niveau > genekspression i prostata. Hverken noradrenalin-, dopaminkoncentration, acetyl- eller butylcholinesterase-aktivitet var påvirkede efter vinclozolin og procymidon behandling, hvilket indikerer at de noradrenerge, dopaminerge, og cholinerge dele af CNS ikke påvirkes markant af pesticiderne. Dog gav vinclozolin og procymidon enkeltvis og i kombination en signifikant forøgelse af totalhjernens serotoninkoncentration. Da serotonin regulerer sekretionen af hypothalamus- og hypofysehormoner på en kompleks måde (Moguilevsky & Wuttke, 2001), kan det ikke udelukkes, at effekt på denne neurotransmitter indirekte kan bidrage til nogle af de hormonforstyrrende effekter af pesticiderne. Generelt vurderes, at der med det anvendte eksperimentelle design ses en biologisk variation mellem dyrene på organvægtniveau, der er af en størrelsesorden, som tillader en relativ præcis og detaljeret undersøgelse af samspilseffekterne. De opnåede data giver således et klart billede af samspillet mellem vinclozolin og procymidon. På baggrund af de opnåede resultater for organvægte, hormonniveauer og mRNA niveauer kan der ikke påvises fravær af additivitet, d.v.s. der er ikke holdepunkter for at antage at der er nogen synergistiske eller antagonistiske samspilseffekter mellem stofferne. Dette underbygges af, at in vitro forsøgene med de samme stoffer viser samme additivitet. Ovennævnte resultater er publiceret i Nellemann et al. 2003. In vitro forsøget, hvor blandinger af pesticider med forskellig virkningsmekanisme blev undersøgt (vinclozolin, menthoxychlor og fenarimol) viste dog ikke en klar additiv effekt, men der var tendens til antagonistiske effekter. Dette indikerer at pesticider i visse tilfælde kan spille sammen på en sådan måde at den totale effekt bliver mindre end man skulle forvente ud fra additivitetsprincippet. Dette understreger behovet for at undersøge kombinationer af pesticider med både samme og forskellig virkningsmekanisme.
|