Skade- og nyttedyrfaunaen på økologiske og konventionelle nordmannsgranarealer
5 Resultater
- 5.1 Arealernes sammenlignelighed - vurderet ud fra florasammensætningen
- 5.2 PCA-analyse af floraen
- 5.3 Forekomst af alm. ædelgranlus
- 5.4 Forekomst af andre skadedyr
- 5.5 Faunaen i bankeprøverne
- 5.6 Diversitetsindices
- 5.7 PCA-analyser af invertebratfaunaen
- 5.8 Hegnenes betydning for invertebrat-faunaen i rækkerne
- 5.9 Forekomsten af ædelgranlus som funktion af den øvrige fauna
- 5.10 Juletræernes kvalitet
5.1 Arealernes sammenlignelighed - vurderet ud fra florasammensætningen
Analyser af bunddækkefloraen og dennes affinitet til forskellige økologiske parametre gav følgende resultat:
Fugtighedsforhold er ens for økologiske og konventionelle arealer. Urtesammensætningen (indikatorarter) viser, at der ikke forskel på fugtighedsforholdene på økologiske og konventionelle arealer (fig. 2).
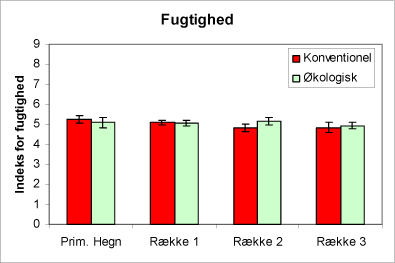
Figur 2. Figuren viser indeks for fugtighed af urtevegetationen i hegnene og i de tre afstande fra hegnet på de økologiske henholdsvis de konventionelt dyrkede arealer. 95% konfidensinterval er angivet.
Urtesammensætningen viser, at de økologiske juletræsarealer generelt har en surere jordbund end de konventionelle arealer, men i hegnene er der ingen forskel mellem driftstyperne (fig. 3). Florasammensætningen indikerer, at de valgte arealer har kvælstofforhold i hegnene, der er næsten ens. På plantagefladen indikerer plantesammensætningen, at de økologiske plantager har lavere kvælstofniveau end de konventionelle arealer (fig. 4). Desuden viser plantesammensætningen, at lystilgangen er ens på de to typer af arealer (fig. 5).
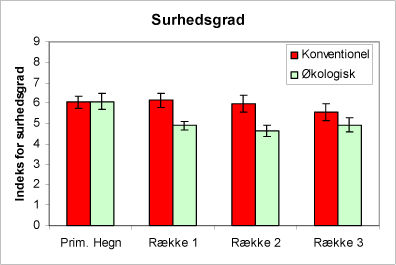
Figur 3. Figuren viser indeks for surhedsgrad af urtevegetationen i hegnene og i de tre afstande fra hegnet på de økologiske henholdsvis de konventionelt dyrkede arealer. SE-værdier angiver 95% konfidensinterval.
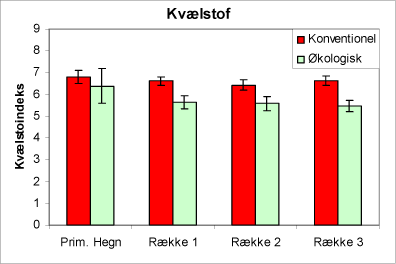
Figur 4. Figuren viser indeks for kvælstof af urtevegetationen i hegnene og i de tre afstande fra hegnet på de økologiske henholdsvis de konventionelt dyrkede arealer. SE-værdier angiver 95% konfidensinterval.
Analyserne af floraen viser, at der ikke er skævhed i materialet. Lokaliteterne synes at være valgt, så de repræsenterer samme gennemsnit og bredde af jordbundstyper.
De forskelle, der er mellem konventionelle og økologiske arealer, er derfor reelle og skyldes ikke skævhed i udvalget af lokaliteter.
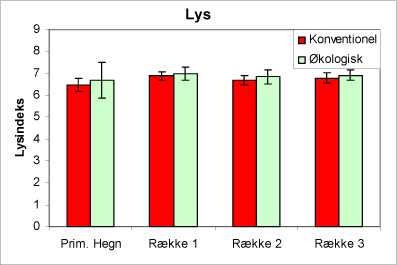
Figur 5. Figuren viser indeks for lystilgang af urtevegetationen i hegnene og i de tre afstande fra hegnet på de økologiske henholdsvis de konventionelt dyrkede arealer. SE-værdier angiver 95% konfidensinterval.
Økologiske bedrifter er karakteriseret ved mindre dækning af NGR, og større urtedækning.
Når man ser på, hvordan dækningen af urter afviger af dækningen af NGR, finder man, at dækningen af bundvegetationen falder med stigende dækning af NGR. Der er den forskel, at plantedækket er ca. dobbelt så stor på økologiske bedrifter ved samme dækning af NGR (fig. 6).
Andelen af énårige planter er signifikant større, jo længere man kommer ud på dyrkningsfladen i de konventionelle plantager (fig. 7). Dette må antages at skyldes, at herbicidanvendelsen og mere intensiv ukrudtsbekæmpelse i almindelighed favoriserer enårige planter.
Artsdiversiteten i bundvegetationen beregnet som ’Shannon-Wiener H’-indekset (se side 21) er større på de økologiske bedrifter end på de konventionelle. For de økologiske bedrifter er indekset omtrent konstant over dyrkningsfladen, mens diversiteten falder fra hegnene på de konventionelle arealer (fig. 8). Det skyldes sandsynligvis en kombination af større behandlingsintensitet med større afstand fra hegnene. Dette er desuden kombineret med større frøspredning og indvækst af planter med udløbere nær hegnene.
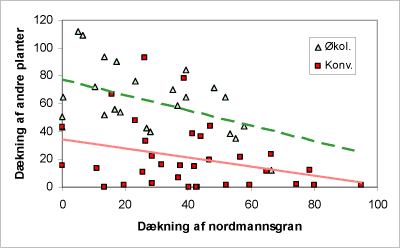
Figur 6. Figuren viser sammenhæng mellem dækningen af nordmannsgran og dækningen af andre planter end nordmannsgran på arealet mellem rækkerne på de to typer af arealer.
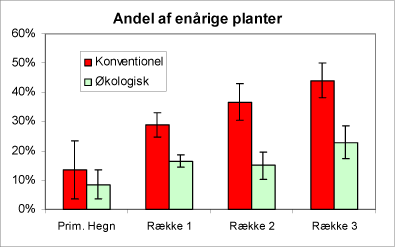
Figur 7. Diagrammet viser andelen (0-1,0) af enårige planter i hegnene og i de tre afstande på de økologiske henholdsvis de konventionelt dyrkede arealer.
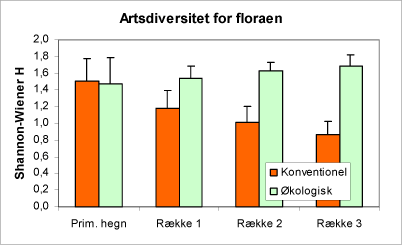
Figur 8. Diagrammet viser artsdiversitet for floraen (Shannon-Wiener H’, se side 21) fordelt på læhegn / rækker af nordmannsgran og driftstype (økologisk / konventionel drift).
5.2 PCA-analyse af floraen
PCA-analyserne i figur 9A og 9B viser for tilsammen tre akser, at hegnsfloraen placerer sig samlet og adskiller sig fra floraen i rækkerne (fig. 9B).
En sammenligning af de to typer af arealer i et PCA-diagram viser, at de økologiske henholdsvis konventionelle arealer fordeler sig i adskilte grupper (fig. 10). Variationen indenfor de økologiske brug er tydeligvis langt bredere end indenfor de konventionelle.
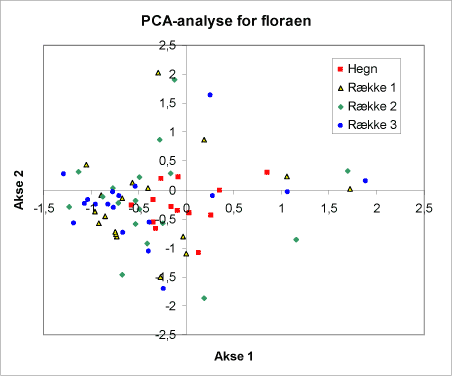
Figur 9A (se teksten figur 9B).
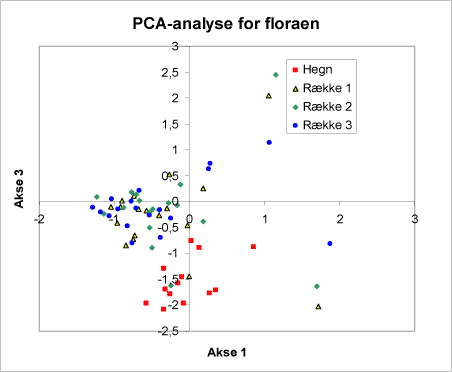
Figur 9B. Diagrammer for PCA-analyse af floraen. De to diagrammer A+B viser for tilsammen tre akser, hvorledes floraen i hegnene placerer sig i forhold til floraen i rækkerne. Bemærk, at punkterne fra hegnene ligger i én gruppe, punkterne fra hegnene i en anden gruppe.
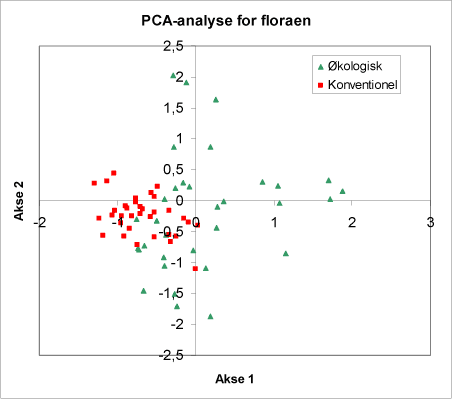
Figur 10. Diagrammet viser en PCA-analyse, der sammenligner floraen på økologiske henholdsvis konventionelle arealer. Bemærk at disse falder i to forskellige grupper, som dog har et overlap. Bemærk desuden, at de punkter fra de økologiske arealer ligger langt mere spredt end punkterne fra de konventionelle.
5.3 Forekomst af alm. ædelgranlus
5.3.1 Sammenligning af bedømmelsesmetoderne for alm. ædelgranlus
I 2001 blev der ved optællingen af de 75 træer pr. areal pr. besøg udelukkende registreret, om der var bladlus tilstede på det enkelte træ eller ej (0-1). I 2002 blev der foretaget en vurdering af tætheden af bladlus på det enkelte træ efter en skala (0-5). På fig. 11 er sammenhængen mellem de to bedømmelsesmetoder illustreret. Det ses, at ved lave tætheder/angreb er der en tilnærmelsesvis lineær sammenhæng mellem de to metoder, men ved højere tætheder, hvor angrebene bliver betydende, bliver afvigelsen mellem de to bedømmelsesmetoder meget stor. Dette betyder, at den mindre tidskrævende bedømmelsesmetode (+/- bladlus) bør suppleres med en subjektiv vurdering af, om angrebet er svagt eller kraftigt i styrke.
At sammenhængen mellem de to metoder ser ud til at være bedst på de økologiske arealer kan forklares ved, at jævnlig bekæmpelse med insekticid vil medføre, at bladlus populationen blive lavere og mere jævnt fordelt på arealet. Forskellen kan også forklares ved at trinnene på 0-5 skalaen ikke er ækvidistante.
I det efterfølgende er resultater, der hidrører fra de to bedømmelsesmetoder, holdt adskilt.
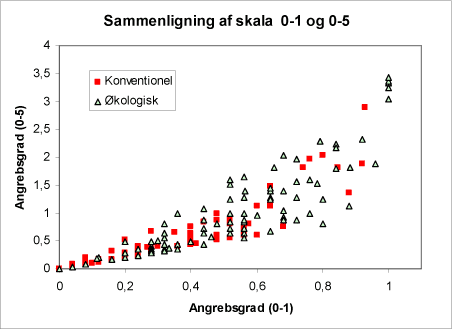
Figur 11. Sammenligning af de anvendte bedømmelsesmetoder for tæthed af alm. ædelgranlus: ’ 0-1’ hvor det blot registreres, om der er bladlus tilstede på de undersøgte træer eller ej. Resultatet kommer derfor ud som” % træer med ædelgranlus”. Ved ”0-5” bedømmes tætheden af ædelgranlus på en skala fra 0-5. Bemærk at ved lave tætheder/angreb er der en tilnærmelsesmæssig lineær sammenhæng mellem de to metoder, men ved højere tætheder, hvor angrebene bliver betydende, bliver afvigelsen mellem de to bedømmelsesmetoder større.
5.3.2 Bladlusforekomst
På figur 12 er vist forekomsten af ædelgranlus på de to typer af arealer gennem sæsonerne 2001og 2002. Det ses, at der i almindelighed er flere træer med bladlus på de økologiske end på de konventionelt drevne arealer. Sæsonudsvingene var i 2001 kun små og den væsentligste udvikling var et fald ved sidste indsamling i begyndelsen af november.
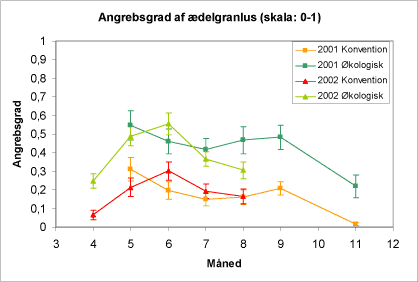
Figur 12. Forekomst af alm. ædelgranlus 2001 og 2002 som procent træer med bladlus gennem sæsonen fordelt på økologiske henholdsvis konventionelle arealer. Diagrammet viser gennemsnit og spredning for de konventionelle (firkanter) og gennemsnit for de økologiske arealer (trekanter).
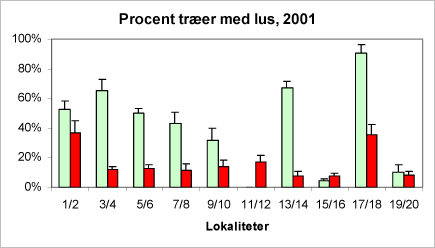
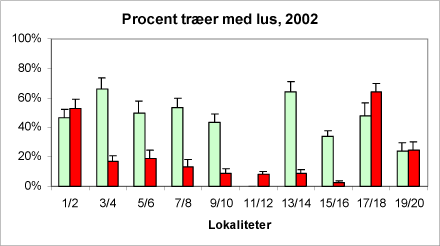
Figur 13A + B. Parvis sammenligning af ædelgranlus forekomsten på økologiske nordmannsgran-arealer (lyse/grønne søjler) og konventionelle (mørke/røde søjler) i hhv. 2001 (A) og 2002 (B). Tallene på abcissen angiver forsøgsværterne (jævnfør kortet bilag A). De viste usikkerhedsangivelser dækker over spredningen på de eksempelvis seks optællinger på den pågældende lokalitet i løbet af sæsonen 2001.
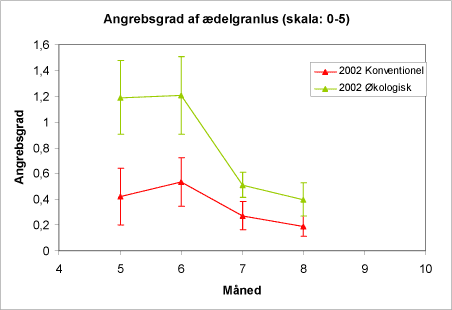
Figur 14. Forløbet i forekomsten af alm. ædelgranlus 2002 på de to typer af lokaliteter. Figuren viser gennemsnit og spredning. SE-værdier angiver 95 % konfidensinterval.
Oversigtsfiguren over forløb over forekomsten af alm. ædelgranlus i 2002 viser – sammenlignet med 2001 - et mere dramatisk fald i juni-juli såvel på konventionelle som på økologiske arealer (fig. 12 og 14). Dette "crash" i bladlusbestanden var generelt for mange typer af kulturer og arter af bladlus det pågældende år, og tilskrives at de aktuelle vejrforhold havde begunstiget først en udvikling i mange bladluspopulationer og senere en positiv udvikling af nyttedyr-populationerne.
En variansanalyse af forekomsten af ædelgranlus på de ti par af økologiske/konventionelle producenter viste, at antallet af træer angrebet af ædelgranlus var betydeligt større ved økologisk dyrkning. Forskellen mellem driftsystemerne er stærk signifikant (***).
Desuden fremgår det af figur 13A+B, at der er en stor geografisk forskel, som kan skyldes graden af isolation for den enkelte plantage eller andre lokale forhold.
Hvis man sammenligner forekomsten af ædelgranlus i rækkerne i de tre afstande fra hegnet, viser det sig, at bladlustætheden tiltager, jo længere væk fra hegnet man kommer (fig. 15 og 16). Når man tager for højde for driftsform og den geografiske placering, bliver denne forskel signifikant (*) for materialet som helhed.
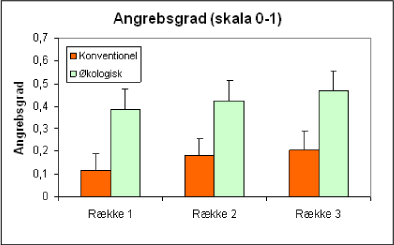
Figur 15. Angreb af alm. ædelgranlus i de tre afstande fra hegnet. Gennemsnit for samtlige optællinger 2001, hvor der blev registreret +/- forekomst af ædelgranlus (skala 0-1) (orange/lmørke søjler = konventionelle; grønne/lyse søjler = økologiske).
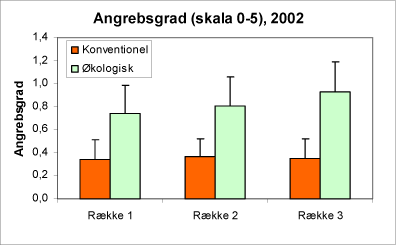
Figur 16. Angreb af alm. ædelgranlus i de tre afstande fra hegnet. Gennemsnit for samtlige prøver 2002, hvor skalaen 0-5 blev anvendt (orange/lmørke søjler = konventionelle; grønne/lyse søjler = økologiske).
5.4 Forekomst af andre skadedyr
Der blev ved den visuelle bedømmelse kun fundet få og spredte symptomer på forekomst af andre skadedyr (galmider, snudebillegnav og minering af ædelgrannålevikere). Det ringe omfang tillader ikke en statistisk behandling.
5.5 Faunaen i bankeprøverne
Der blev indsamlet, udsorteret, identificeret og optalt i alt ca. 150.000 individer af leddyr. Disse fordeler sig på 312 arter eller artsgrupper. Billerne alene omfatter 168 arter heraf. Arterne er blevet samlet i taksonomiske grupper, funktionelle grupper samt efter deres fordeling i hegnet og afstanden fra dette (se bilag B).
Den største interesse samler sig naturligt om de dyr, der kan optræde som henholdsvis skadedyr og nyttedyr i forhold til nordmannsgran.
I det efterfølgende præsenteres de faunagrupper, der er mest relevante i denne sammenhæng, og hvor datamaterialet er tilstrækkelig stort til at kunne bære en præsentation.
5.5.1 Nyttedyr
En af de væsentligste grupper udgør mariehønsene. Vi ved fra tidligere undersøgelser, at såvel voksne som larver er villige predatorer på alm. ædelgranlus (Ravn et al. 2004). Fra opgørelsen over bankeprøverne (Bilag B) fremgår det, at mariehønsene optræder mest talrigt på de økologiske lokaliteter (figur 17). Der er ikke en generel tendens til at mariehønsenes antal på nordmannsgran afhænger af afstanden fra hegnet. En nøjere analyse viste dog, at der var et stigende antal væk fra hegnene for de økologiske lokaliteter i 2002.
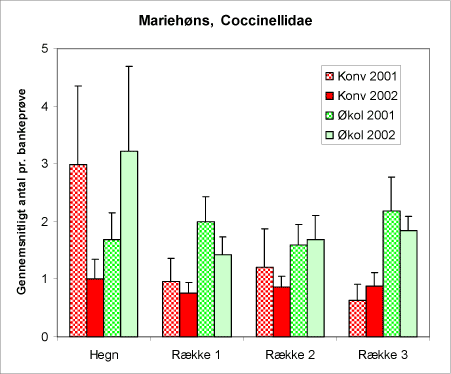
Figur 17. Forekomst af mariehøns i bankeprøverne på de to typer af lokaliteter begge år. Ordinat-aksen angiver det gennemsnitllige antal pr. bankeprøve. På abcissen er angivet hvor prøven er taget: i hegnet eller i rækkerne i de tre afstande fra hegnet.
Mariehøns udviser store forskelle i populationstæthed. Det fremgår også af denne undersøgelse (figur 18). Store tætheder af voksne mariehøns i juli-august er især betinget af gode fødeforhold for larverne i maj-juni måned. Dette giver en god baggrund for at forklare den voldsommere nedgang i forekomsten af alm. ædelgranlus i 2002 sammenlignet med 2001 og forskellen på pesticidfri arealer sammenlignet med konventionelle (figur 14).
Hos blødvinger, Cantharidae (figur 19), kan såvel larver som voksne optræde som bladluspredatorer. De forekommer på nordmannsgran – lejlighedsvis i større tætheder, hvor de grundigt efterstræber bl.a. brun ædelgranlus, Cinara confinis (Ravn 2002). Det er dog ikke undersøgt i hvilket omfang alm. ædelgranlus indgår i blødvingernes foretrukne bytte. Gruppen rummer mange arter, og antageligt er der artsspecifikke præferencer for visse bladlusgrupper. Denne insektgruppe forekommer hyppigst i hegnene og er lige hyppig på de to typer af arealer.
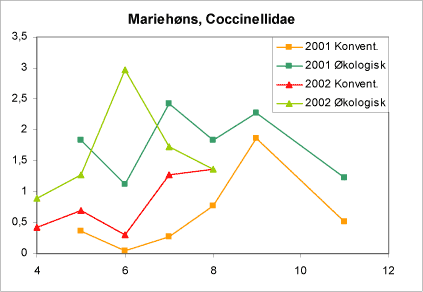
Figur 18. Månedsfordeling af mariehøns (Coccinellidae) i bankeprøverne fra økologiske og konventionelle brug 2001 og 2002. Abcissen angiver måned (4=april) og ordinaten det gennemsnitlige antal mariehøns pr. bankeprøve. Kun antallet af mariehøns i Nordmannsgranrækkerne er indgået i beregningerne.
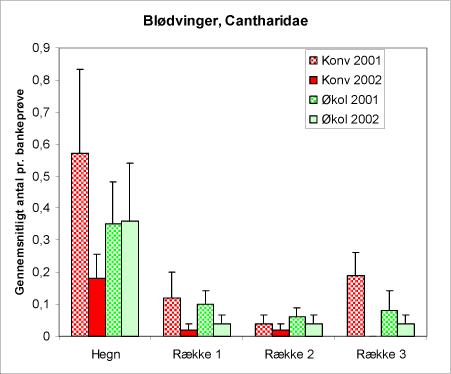
Figur 19. Forekomst af blødvinger i bankeprøverne fra hegnene og i tre afstande fra disse. Diagrammet viser antallet af blødvinger i bankeprøver fra de to typer af driftsformer (konventionel hhv. økologisk) og fra begge indsamlingsår (2001 og 2002).
Netvingede, Neuroptera (figur 20) rummer en række familier – guldøjer Chrysopidae og florvinger, Hemerobiidae, hvoraf de fleste har larver, der lever af bladlus. Der er tilsyneladende ikke den store forskel på denne gruppes forekomst på de to typer af arealer.
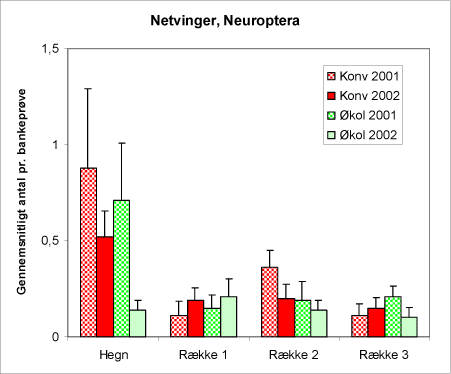
Figur 20. Netvingede (Neuroptera) i bankeprøverne. Hovedparten er Guldøjer (larver og voksne).
Gruppen snyltehvepse (figur 21), (udgør størstedelen af Apocrita) rummer især individer fra familierne Chalcidoidea, Proctrupoidea og Ichneumonoidea. Gruppen udviser ikke signifikant forskel mellem de to arealtyper, men aftager signifikant med afstanden fra hegnene.
Ørentviste (Dermaptera) (figur 22) er overvejende nataktive dyr og relativt altædende, men deres rolle som predatorer overfor alm. ædelgranlus er velkendt (Smetana & Jensen, 1998). Alm. ørentvist, Forficula auricularia udgør langt hovedparten af individerne i denne gruppe. Det ses af bankeprøverne, at ørentvistene er hyppigst i hegnene og på de økologiske arealer.
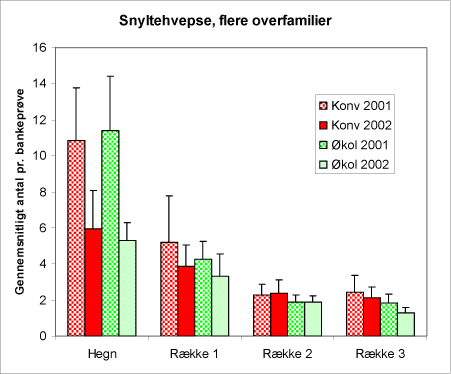
Figur 21. Fordelingen af snyltehvepse i bankeprøverne begge år og på de typer af arealer. Bemærk, at hyppigheden aftager signifikant med afstanden fra hegnene.
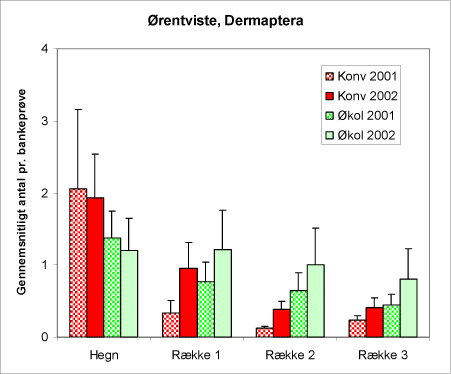
Figur 22. Antallet af ørentviste (Dermaptera) i bankeprøverne fra begge år og begge arealtyper.
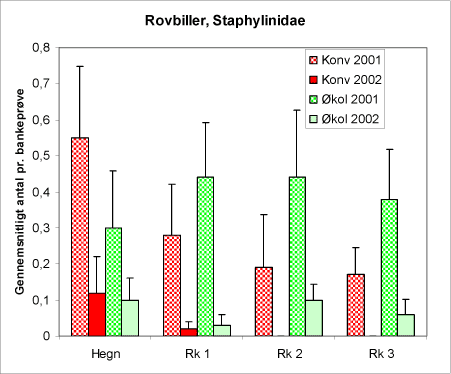
Figur 23. Antallet af rovbiller (Staphylinidae) i bankeprøverne fra begge år og begge arealtyper.
Rovbillerne, Staphylinidae, (figur 23) er en meget artsrig gruppe og trods navnet er de ikke alle predatorer. Oppe i vegetationen er arter af slægten Tachyporus hyppigt forekommende. De lever af bladlus – måske også alm. ædelgranlus. De fleste af rovbillerne i bankeprøverne bestod af individer af denne slægt. Som det ses af figur 23, findes der begge år flere rovbiller på det økologiske areal i forhold til det konventionelle. I hegnene er det omvendt.
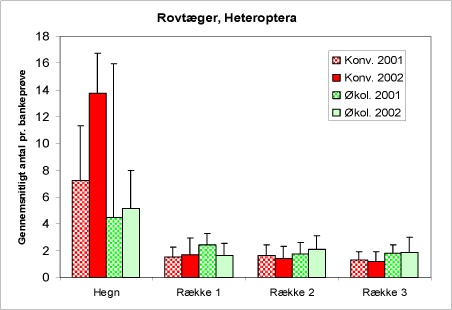
Figur 24. Forekomst af rovlevende tæger (Heteroptera) i bankeprøverne i begge år og på begge typer af arealer.
Tægerne, Heteroptera, tæller såvel plantesugere som rovlevende insekter i gruppen, nogle arter – f.eks inden for familien Miridae – kan optræde både som nytte- og skadedyr. De udprægede rovlevende familier (f.eks. næbtægerne Anthocoridae og rovtægefam. Reduviidae og Nabidae) er præsenteret på figur 24. Det ses, at individer af denne gruppe i høj grad holder sig til hegnene – de er typiske løvtræsinsekter. På dyrkningsfladen er der ikke forskel på forekomsten på de typer af arealer.
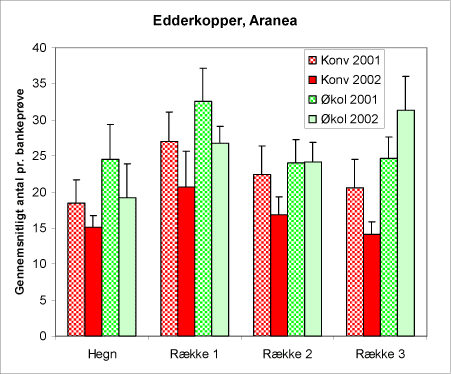
Figur 25. Forekomst af edderkopper i bankeprøverne i begge år og på begge arealtyper.
Edderkopperne (figur 25) regnes normalt blandt nyttedyrene, i hvert fald er de rovlevende. Imidlertid er bladlus ikke deres foretrukne bytte. De er polyfage rovdyr, og fortærer et bredt udsnit af byttedyr. Bl.a. fanger og fortærer de fluer og andre insekter, der bevæger sig hurtigt. I juletræsplantagerne er den store tæthed af edderkopper iøjnefaldende. Der er registreret næsten 20.000 edderkopper i bankeprøverne. Antagelig giver juletræsplantagens åbne struktur rigtigt gode muligheder for de netspindende arter. Edderkopper er relativt påvirkelige af pesticidanvendelse, det er måske det, der afspejler sig i den hyppigere forekomst på de økologiske juletræsarealer.
5.5.2 Skadedyr
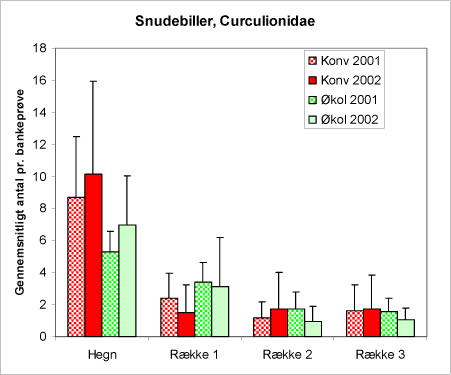
Figur 26. Antallet af snudebiller pr. bankeprøve på de to typer af arealer for de to år.
Gruppen snudebiller – Curculionidae – rummer et antal arter, der kan optræde som skadevoldere i juletræskulturer. Flere arter af løvsnudebiller er polyfage og begnaver i det tidlige forår mange forskellige arter af løv- og nåletræer – også nordmannsgran. En række af de fundene snudebillearter kan optræde mere eller mindre skadeligt på juletræerne, det gælder f.eks. repræsentanter fra slægterne: Barypeithes, Cryphalus, Phyllobius, Polydrosus og ikke mindst Strophosoma. Som det fremgår af diagrammet, figur 26, er Snudebillerne er talrigst i hegnene og aftager ind i plantagen. Der er ikke forskel i forekomsten på de to typer af arealer.
5.5.3 Andre faunagrupper
En række store insektgrupper er ikke udprægede nytte- eller skadedyr i relation til juletræerne. Det gælder f.eks. de fleste af tovingerne (figur 27). Ordenen tovingede forekommer med relativt stort individantal i prøverne, selv om bankeskærm ikke er den bedste metoder til at indsamle denne gruppe. Af diagrammet ses, at der ikke er forskel i tovingernes forekomst på de to typer af arealer, men derimod en tydelig hegns effekt.
Den næste gruppe – springhalerne (figur 28) – udgør ubetinget den hyppigste enkeltgruppe. Springhalerne er utroligt talrige i plantagerne. Der er blevet registreret 67.000 springhaler i bankeprøverne. Når vi finder dem oppe i træerne, er det fordi, de lever af alge- og svampebelægninger. De er således indirekte nyttedyr. De udgør desuden alternativ føde til en række polyfage, rovlevende dyr. Springhalerne er en af de få faunagrupper, hvor vi finder den største hyppighed inde i plantagen i forhold til hegnet. Der er derimod ikke signifikante forskelle mellem de to arealtyper.
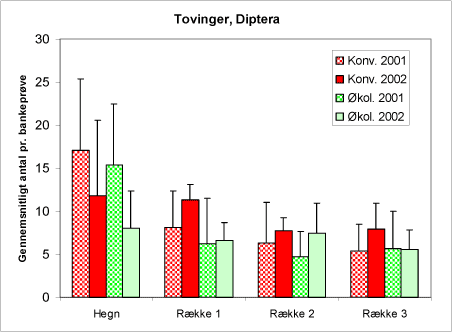
Figur 27. Fund af tovinger (fluer, myg og stankelben), Diptera i bankeprøverne på begge typer af arealer og begge år.
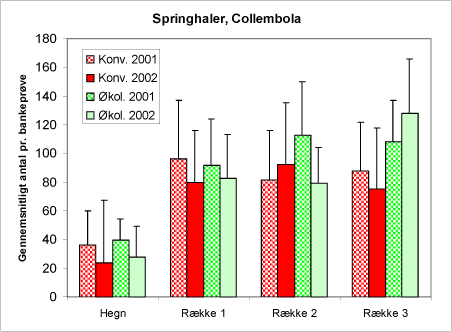
Figur 28. Antallet af springhaler, Collembola, der er registreret i bankeprøverne de to år på de to typer af arealer.
5.5.4 Generelle faunaovervejelser
Nogle få insektgrupper har et atypisk billede i forekomsten.
Barklus, Psocoptera, lever af pollen, alger og laver på bark og blade på træerne. I gamle fuglereder findes der store mængder. Nogle arter er mellemvært for fårebændelorm.
Barklus optræder relativt hyppigt i bankeprøverne. Det er derfor relativt sikkert, når diagrammet (figur 29) viser, at der er flere barklus i plantagen end i hegnene, og der er flere barklus på de økologiske arealer i forhold til de konventionelle samt at forekomsten i bankeprøverne tiltager ind på arealet på de økologiske arealer og er konstant eller aftager på de konventionelle. Det kunne tyde på, at barklus er følsomme for pesticidanvendelse. Denne hypotese, har dog ikke kunnet bekræftes, måske er det andre forhold, der spiller ind.
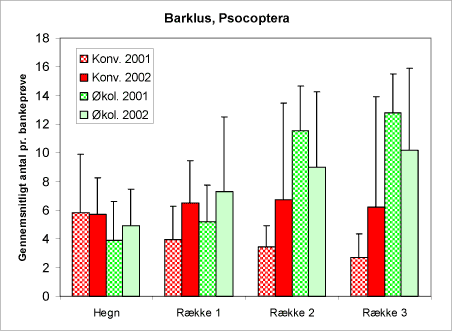
Figur 29. Diagram over forekomsten af barklus i bankeprøverne fra hegnene og nordmannsgranerne i de tre afstande på de to typer af arealer og de to år.
5.6 Diversitetsindices
En sammenligning af resultatet fra optællingen af bankeprøverne viser, at hvis man betragter fauna-familierne, træffer vi ikke et signifikant større antal forskellige familier på de økologiske arealer i forhold til de konventionelle (figur 30). En sammenligning af arealtyperne ved hjælp af de beregnede diversitetsindices (figur 31 og 32) viser ligeledes, at der – bedømt ud fra faunaen i bankprøverne – kun er en lille smule hvis nogen overhovedet overvægt i mangfoldigheden på de økologiske arealer.
Sammensætningen af træ- og buskarterne i hegnene varierer kraftigt mellem lokaliteterne. På figur 33 er Shannon-Wiener H’ diversitetindekset for hegnene på lokaliteterne 7-20.
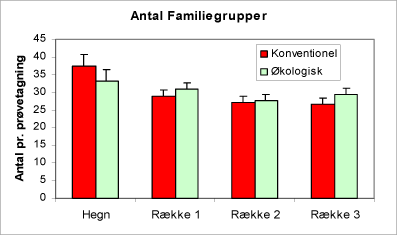
Figur 30. Diagrammet viser antal familiegrupper pr. prøvetagning. Bemærk, at der er flere invertebratfamilier i hegnene end i rækkerne. Der er ingen gradient fra række 1 til 3.
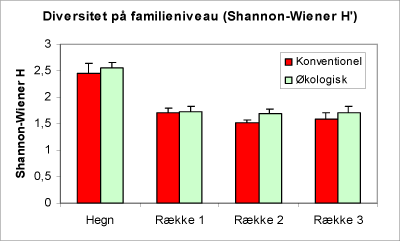
Figur 31. Diagram over diversitet på familieniveau (Shannon-Wiener H’, se side 21). Bemærk, at der ingen forskel er på driftstyperne hverken i hegn eller i rækkerne og ingen gradient fra række 1 til 3. Hegnene har den højeste diversitet, når der vurderes ud fra indholdet af bankeprøverne.
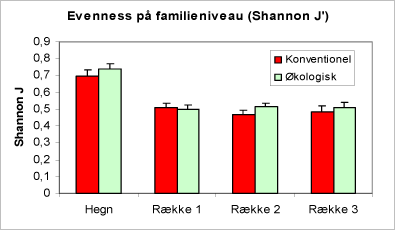
Figur 32. Evenness på familieniveau (Shannon J’, se side 21). Der er ingen forskel på driftstyperne hverken i hegn eller i rækkerne og ingen gradient fra række 1 til 3. Hegnene har den højeste Evenness.
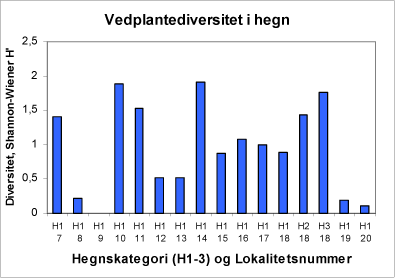
Figur 33. Diversitet af træ- og buskarter i hegnene.
5.7 PCA-analyser af invertebratfaunaen
I et PCA-diagrammerne har punkter, der ligger sammen, fælles affinitet til de omverdens faktorer, der er inddraget i analysen. PCA-analyserne af invertebratfaunaen i familiegrupper viste, at en af de mest betydende faktorer var en stor variation mellem de to år (figur 34). Desuden faldt hegnene henholdsvis rækkerne med nordmannsgran i hver sin vel adskilte gruppe (figur 35). Man kunne ikke erkende forskelle mellem driftstyperne økologisk eller konventionel drift (figur 36). Punkterne fra de økologiske henholdsvis konventionelle arealer danner én sammenfaldende punktsværm.
På grund af den store årstidsvariation blev ”år” taget ind som en covariabel, så eventuelle forskelle mellem rækkerne eller drifttyperne blev fremhævet. Resultatet var stadig det samme, nemlig, at man ikke kunne identificere forskelle mellem rækkerne indbyrdes. Der var heller ingen forskel mellem drifttyperne.
I en særlig analyse blev datamaterialet splittet op i hegn og rækker, som blev analyseret hver for sig. Herefter blev det undersøgt, om de vigtigste akser i analysen af faunaen i hegnene kunne forklare fordelingen af faunaen i rækkerne. Der var en ikke signifikant tendens til sammenhæng. Ud fra faunasammensætningen blev det vurderet, at denne tendens til sammenhæng var en følge af nærhed til vand i omgivelserne. Sammenhængen skyldes således snarere, at faunaen i hegn og rækker reagerer på de samme omgivelsesfaktorer. Det kunne således ikke påvises, at faunaen i rækkerne som helhed afhang af faunasammenhængen i hegnene. Som det er vist andetsteds, er der dog for enkelte organismegrupper en sammenhæng.
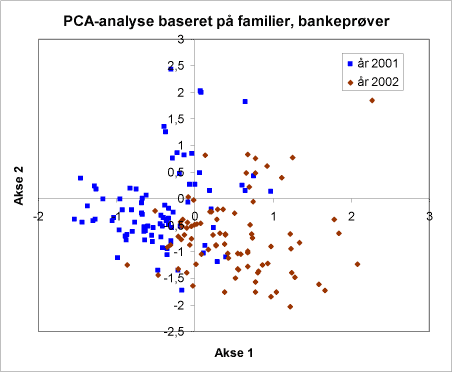
Figur 34. Fordeling af bankeprøver efter PCA-analysens to vigtigste ordinationsakser. Signaturerne viser hvilket år prøven er taget. Der er en tydelig forskel mellem årene. Hvert punkt repræsenterer gennemsnittet af 4 måneders bankeprøver i perioden maj-august for hvert år i en række eller et hegn på en lokalitet.
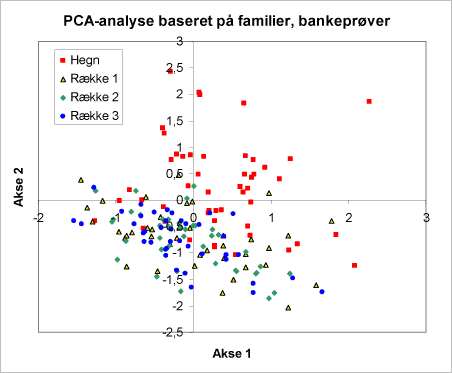
Figur 35. Som figur 34. Signaturerne viser her om bankeprøven er taget i hegn eller i en bestemt række. Umiddelbart er der en klar forskel mellem rækkerne på den ene side og hegnene på den anden. Man kan ikke umiddelbart se forskel mellem de tre rækker.
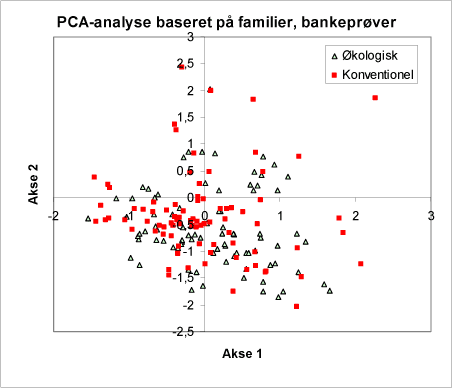
Figur 36. Som figur 34. Signaturerne viser her om bankeprøven er taget på en lokalitet med økologisk drift eller med konventionel drift. Bemærk, at punkterne udgør én samlet gruppe – dvs. umiddelbart er der ingen forskel mellem driftstyperne.
5.8 Hegnenes betydning for invertebrat-faunaen i rækkerne
Da det ikke kunne påvises, at faunaen i rækkerne som helhed afhang af faunasammenhængen i hegnene, blev det undersøgt, om der var en signifikant forskel på forekomsten af antal af individer i en række faunagrupper i række 1 og række 3 (tabel 1). Der var adskillige grupper med signifikant forskel. Kun en af de signifikante faunagrupper (Barklus, Psylloidea) er der en negativ t-værdi, så der er færrest i række 1. For alle de øvrige signifikante forskelle gælder det at der er flere individer i række 1 end i række 3.
Dernæst blev det undersøgt om kontrasten mellem faunaen i række 1 og 3 kunne forklares ved invertebratfaunaen i hegnene. Hvis hegnene har en indflydelse på rækkernes fauna må det nemlig antages, at denne indflydelse falder med afstanden fra hegnet.
Kontrasten mellem række 1 og række 3 blev for hver familiegruppe (eller nærmest højere systematiske niveau) udtrykt ved følgende faunaratio:
log((Nmiddel,,række 1 + 0,1)/((Nmiddel,,række 3 + 0,1))
hvor Nmiddel,,række 1 eksempelvis er middelværdien for en familiegruppe over begge år og månederne 5-9 i række 1.
Tabel 1. Faunaforskelle mellem række 1 og 3. Parvis t-test over forskel i ratioen log(Nrk+0,1)/(Nhegn+0,1) mellem række 1 og række 3. Data er gennemsnittet for maj – september begge år. Der indgår således en observation for hver lokalitet (N=20). Mht. respons er der meget lidt forskel mellem årene. En beregning hvor begge de to år indgik gav næsten identisk resultat (N=40). Faunagrupper med få data er udeladt.
| Gruppe | T(N=20) | P(N=20) | Rk1>Rk3 | Rk1=Rk3 | Rk1<Rk3 |
| _HETEROP | 1,0772 | 0,2949 | 10 | 2 | 8 |
| _HOMOPTE | 0,9613 | 0,3485 | 5 | 12 | 3 |
| ANTHOCOR | -1,6451 | 0,1164 | 3 | 10 | 7 |
| APHIDOID | -0,2345 | 0,8171 | 11 | 0 | 9 |
| APOCRITA | 2,9954 | 0,0074 | 14 | 0 | 6 |
| ARANEA | 2,1702 | 0,0429 | 14 | 0 | 6 |
| CERCOPID | 0,8302 | 0,4168 | 10 | 1 | 9 |
| CICADELL | 5,4173 | 0,0000 | 18 | 0 | 2 |
| COCCINEL | -0,6216 | 0,5416 | 9 | 0 | 11 |
| COLEOPTE | 2,6462 | 0,0159 | 16 | 1 | 3 |
| COLLEMBO | -0,0051 | 0,9960 | 11 | 0 | 9 |
| CORTICAR | 4,0726 | 0,0006 | 17 | 0 | 3 |
| CURCULIO | 2,1076 | 0,0486 | 15 | 0 | 5 |
| DIPTERA | 1,0211 | 0,3200 | 13 | 0 | 7 |
| FORFICUL | 0,2341 | 0,8174 | 5 | 9 | 6 |
| FORMICID | -0,7586 | 0,4574 | 6 | 4 | 10 |
| HYMENOPT | 0,9843 | 0,3373 | 6 | 10 | 4 |
| LEPIDOPT | 2,8667 | 0,0099 | 14 | 1 | 5 |
| MIRIDAE | 1,7603 | 0,0944 | 13 | 1 | 6 |
| NITIDULI | 0,1634 | 0,8719 | 7 | 6 | 7 |
| OPILONES | 3,5071 | 0,0024 | 14 | 0 | 6 |
| PSOCOPTE | -0,6560 | 0,5197 | 10 | 0 | 10 |
| PSYLLOID | -1,7886 | 0,0896 | 8 | 0 | 12 |
| TENTHRED | 0,8360 | 0,4136 | 5 | 13 | 2 |
| _NON_CRY | -1,2364 | 0,2314 | 8 | 3 | 9 |
| ACARI | -0,6605 | 0,5168 | 9 | 1 | 10 |
| BRACHYCE | 0,2286 | 0,8217 | 10 | 1 | 9 |
| BRENTIDA | 2,1632 | 0,0435 | 11 | 4 | 5 |
| CRYPTOST | 2,1518 | 0,0445 | 12 | 1 | 7 |
| DERMAPTE | 2,1386 | 0,0457 | 13 | 3 | 4 |
| GASTROPO | 1,7106 | 0,1034 | 12 | 0 | 8 |
| CHRYSOME | 0,7559 | 0,4590 | 12 | 3 | 5 |
| NEMATOCE | 1,2844 | 0,2144 | 10 | 2 | 8 |
| CHALCIDO | 3,4677 | 0,0026 | 16 | 0 | 4 |
| HEMIPTER | 1,2284 | 0,2343 | 7 | 8 | 5 |
| LYGAEIDA | 1,7648 | 0,0937 | 6 | 12 | 2 |
| ISOPODA | 2,2668 | 0,0353 | 7 | 12 | 1 |
| DIPLOPOD | 0,4497 | 0,6580 | 3 | 16 | 1 |
En PCA-analyse blev udført for disse faunaratioer. Herved blev faunakontrasten mellem række 1 og 3 udtrykt ved de fire vigtigste ordinationsakser. Disse fire akser forklarer tilsammen 46,7% af den samlede variation.
Ved en ordination af hegnenes faunasammensætning midlet over de samme måneder blev den vigtigste variation i hegnenes fauna reduceret til 4 akser (tabel 2). De fire akser forklarer tilsammen 90,5% af faunavariationen i hegnene. Samtidig forklarer de fire akser for invertebratfaunaen 64,2% af faunakontrasten mellem række 1 og 3.
Ved en ordination af træartssammensætningen i hegnene forklarede de fire vigtigste akser 51,7% af variationen. Træartssammensætningen udtrykt ved disse fire variable kunne tilsammen forklare 37,7% af variationen i faunakontrasten mellem række 1 og række 3.
Tabel 2. Forklaringsprocenter for de fire vigtigste akser i ordinationsanalysen
| Akse 1 | Akse 2 | Akse 3 | Akse 4 | |
| Faunakontrast mellem række 1 og 3 | 16,8 | 27,9 | 38,4 | 46,7 |
| Faunaen i hegn forklarer af denne: | 23,0 | 42,0 | 46,2 | 64,2 |
| Træartssammensætningen forklarer af denne: | 8,5 | 15,8 | 29,6 | 37,7 |
| Faunaen i hegn | 53,3 | 77,0 | 86,1 | 90,5 |
| Træartssammensætningen i hegn | 17,6 | 30,2 | 41,6 | 51,7 |
Som man kan se, er en stor del af variationen i faunakontrasten mellem række 1 og 3 forklaret ved faunaen i hegnene. Derimod kan træartssammensætningen i hegnene kun forklare en mindre del af variationen i faunakontrasten mellem række 1 og 3.
5.9 Forekomsten af ædelgranlus som funktion af den øvrige fauna
Der er ingen overordnet sammenhæng mellem mængden af bladlus og ratioen predatorer/herbivorer eller ratioen for nordmannsgran specifikke nyttedyr/skadedyr (jævnfør bilag B) (r = -0,04 ns).
For at teste om ratioen af ædelgranlus i række 1 og række 3 (Nrække 1 +0,01)/(Nrække 3+0,01) afhænger af faunaratioen mellem række 1 og 3 er der udført en lineær regression, hvor de fire vigtigste PCA-ordinationsakser over faunakontrasten mellem række 1 og 3 indgår som uafhængige variable.
Regressionen viste, at forskellen på angreb af bladlus i række 1 og 3 havde en signifikant sammenhæng med forskellen i insekter i række 1 og 3 (F4,19= 3.51, P=0,0325). Det er især akse 1 og akse 4, der er betydende, og begge har positiv sammenhæng med ædelgranlus-ratioen. Jo flere bladlus der er i række 1 i forhold til række 3 jo større er værdierne på ordinationsakse 1 og 4.
Der var signifikant korrelation (Spearman Rank Correlation) mellem forskellen i angreb af alm. ædelgranlus i række 1 og 3 og faunakontrasten mellem række 1 og 3 af adskillige insektgrupper i rækkerne: Den inhomogene gruppe ”Røde mider”(rs= -0,56), Non-cryptostigmata (rs= +0,56), Cydnidae (rs= -0,49), Plecoptera (rs= +0,46), Lathridiidae (r= +0,46) Brentidae (rs= +0,46), Acanthosomatidae (rs= -0,45), Staphylinidae (rs= -0,45) Chalcidoidea (rs= -0,45). Det skal bemærkes, at der var også en tendens til sammenhæng med Coccinellidae (rs= -0,44). Med positivt fortegn betyder det, at jo flere individer af gruppen, der er i række 1 i forhold til række 3, jo flere ædelgranlus vil der være i række 1 i forhold til række 3. Et negativt fortegn antyder derfor at invertebratgruppen dæmper antallet af bladlus. Rovbillerne er et eksempel på en gruppe, der er kandidat til at holde bladlus nede og blive begunstiget i række 1.
Der var derimod ingen samlet signifikant sammenhæng mellem forskellen i angreb af ædelgranlus i række 1 og række 3 og insekter i hegnene udtrykt ved de fire vigtigste ordinationsakser. Der var dog en tendens til sammenhæng (Lineær regression, F4,19=2.55, P=0,0825). Isoleret set var sammenhængen med akse 2 signifikant.
Der var kun signifikant korrelation mellem forskellen i angreb af bladlus i række 1 og 3 med mængden af en enkelt invertebratgruppe i hegnene: Mejere, Opiliones (rs= -0,46). Jo færre bladlus i række 1 i forhold til række 3, jo færre mejere skulle der være i hegnene. Denne gruppe kan dog ikke antages, at have nogen form for direkte årsagssammenhæng med forekomstens af ædelgranlus.
Der var ingen samlet signifikant sammenhæng mellem forskellen i angreb af ædelgranlus i række 1 og række 3 og træarter i hegnene udtrykt ved de fire vigtigste ordinationsakser (Lineær regression, F4,19=1.02, P=0,43).
Der fandtes heller ingen signifikante korrelationer mellem forskellen i angreb af ædelgranlus i række 1 og 3 med mængden af enkelttræarter i hegnene (der blev anvendt nonparametirsk korrelation (Spearman Rank Correlation) idet datasættet indeholdt mange 0-værdier.
5.10 Juletræernes kvalitet
I 2001 træerne blev klassificeret I-III, hvor I er primatræer, II er standard, og III er uklassificeret – og 2002 det blev vurderet hvor kraftig en gulfarvning (0-2) træerne havde. Af figur 37 fremgår, at variationen er kolossal. Der er dog ikke markant færre primatræer på de økologiske arealer.
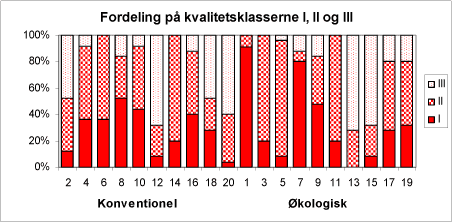
Figur 37. Fordeling af juletræerne på kvalitetsklasserne I, II, og III i 2001. De lige numre er de konventionelle arealer og de ulige er de økologiske arealer.
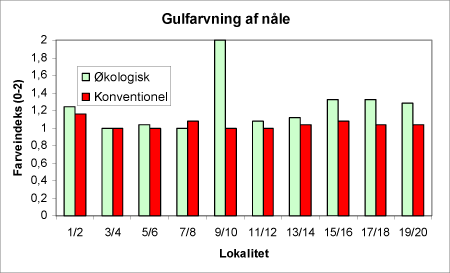
Figur 38. Gulfarvning af træerne på de enkelte lokaliteter i 2002 vurderet efter en 0-2 skala, hvor 2 er de mest gule træer.
Til gengæld er der en klar tendens til at problemer med gulfarvning af nålene er mere udtalt på de økologiske arealer end på de konventionelle (figur 38).
Åbenhed i toppen er forholdet mellem årstilvæksten i fht. træets højde (figur 39 og 40). Der ses i begge tilfælde en tendens til at de konventionelle træer placerer sig som træerne med længste årsskud i fht. de økologiske.
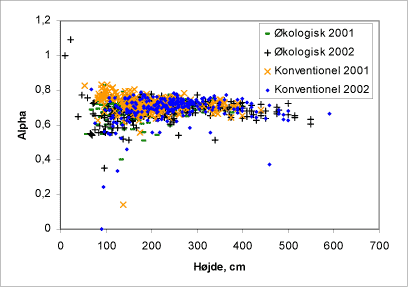
Figur 39. Træernes åbenhed (a) i forhold til træhøjden.
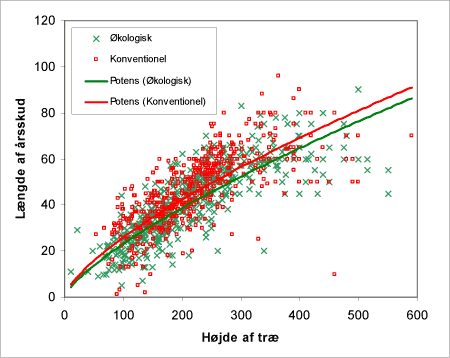
Figur 40. Åbenhed i toppen bestemt på baggrund af længden af sidste årsskud i forhold til træets højde.
Version 1.0 November 2006, © Miljøstyrelsen.