Mikrobiologiske plantebeskyttelsesmidlers skæbne i mave-tarm kanalen
3 Resultater
- 3.1 Overlevelse og spiring af Bt og B. cereus celler i tarmen
- 3.2 Ændringer af den normale tarmflora
- 3.3 Enterotoksiner i tarmen
- 3.4 Overførsel af DNA mellem to Bt stammer i kimfri rotter
3.1 Overlevelse og spiring af Bt og B. cereus celler i tarmen
3.1.1 Resultater fra HFA dyr
Kimfri rotter blev først fodret med en human fæces-opslæmning, som fik lov til at etablere sig i rotterne i tre uger, hvorefter dyrene blev fodret med enten B. cereus, Bt subsp. kurstaki eller Bt subsp. israelensis i tre separate forsøg. I hvert forsøg blev dyrene inddelt i fire hold som fik hhv. stråledræbte sporer (Hold 1), ubehandlede sporer (Hold 2), varme-aktiverede sporer (Hold 3) og vegetative celler (Hold 4), se også Figur 2. Dyrene blev fodret med de enkelte bakteriesuspensioner fire dage i træk, halvdelen af dyrene blev aflivet på femtedagen og resten af dyrene to uger efter. Fæces- og tarmprøver blev undersøgt for indholdet af Bt og B. cereus ved at udplade på selektive plader.
3.1.1.1 Genfindelse af Bt og B. cereus i fæces
Som forventet, blev der i ingen af de dyr der blev fodret med stråledræbte sporer (negativ kontrol) detekteret B. cereus lignende bakterier i fæces- eller tarmprøver. I de dyr der fik vegetative celler genfindes ingen eller kun få B. cereus lignende bakterier, dette skyldes formodentlig at de vegetative celler inaktiveres i mavesyren (se afsnit 4.1.2). Nedenstående figurer (Figur 7) viser antallet af de tre undersøgte stammer i fæces i de dyr, der fik levende sporer (Hold 2 og Hold 3). Som det ses af figurerne er der ingen forskel på ubehandlede (Hold 2) og varmebehandlede (Hold 3) sporer mht. genfindelse af stammerne i fæces. Forskellen mellem holdene som ses i de dyr der blev fodret med Bt subsp. israelensis skyldes en forskel i doseringen mellem holdene. Hold 3 blev fodret med ca. 106 varmebehandlede sporer, mens Hold 2 fik ca. 108 ubehandlede sporer. Dyrene i de to andre forsøg (B. cereus og Bt subsp. kurstaki) fik ligeledes ca. 108 bakterier i både Hold 2 og Hold 3. Dyrene blev doseret dag 1-4 (begge dage inkl.) og man ser tydeligt at antallet af B. cereus lignende bakterier er størst i den første uge (ca. 106 cfu/g fæces). Derefter falder antallet, dog med meget stor variation mellem dyrene.
I forsøget med B. cereus F4433/73 er der et dyr hvor bakterien kan genfindes to uger efter sidste dosering, mens det gælder for fem ud af seks dyr i forsøget med Bt subsp. kurstaki, denne forskel kan skyldes forskel i detektionsgrænse mellem de to forsøg (100 hhv. 10 cfu/g fæces) (Figur 7). I et af dyrene fodret med Bt subsp. kurstaki (dyr 15) er koncentrationen af stammen meget højere end de andre dyr i fæces 2 uger efter sidste dosering (Figur 7). I forsøget med Bt subsp. israelensis kunne bakterien ikke detekteres i dyrenes fæces to uger efter sidste dosering.
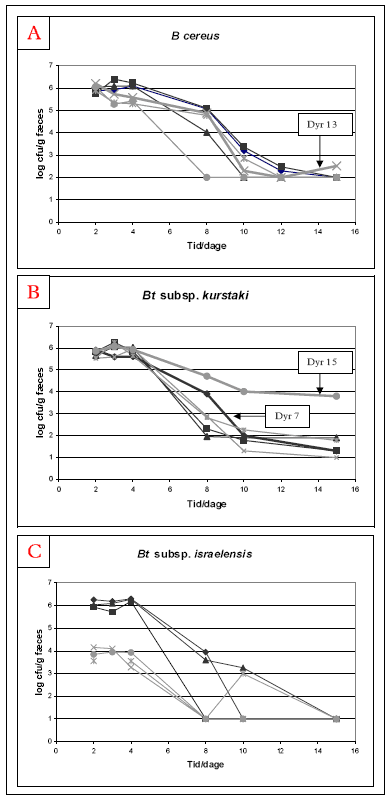
Figur 7. Genfindelse af hhv. B. cereus (A), Bt subsp. kurstaki (B) og Bt subsp. israelensis (C) i fæces fra individuelle dyr doseret dag 1-4 med ubehandlede sporer (sort, Hold 2) og varmebehandlede sporer (grå, Hold 3). Der blev udført tre separate forsøg, et med hver af de tre førnævnte stammer.
3.1.1.2 Genfindelse af stammer i tarmprøver og organer
Halvdelen af dyrene fra hvert hold blev aflivet på femtedagen efter at have fået enten B. cereus, Bt subsp. kurstaki eller Bt subsp. israelensis fire dage i træk. De resterende dyr blev aflivet to uger efter sidste dosering. Prøver blev taget ud fra følgende tarmafsnit: duodenum (øvre tyndtarm), ileum (nedre tyndtarm), caecum (blindtarm) og kolon (tyktarm).
Antallet af genfundne bakterier af de tre stammer fra dyr der blev fodret med ca. 108 ubehandlede sporer (Hold 2) ved den første aflivning kan ses i Figur 8 (se også mere udførlige data i Tabel 4). Antallet af de tre stammer i caecum og kolon er ens, hvorimod der er stor forskel på genfindelse af de tre stammer i tyndtarmsprøverne. Dyrene der blev doseret med sporer af Bt subsp. israelensis skiller sig ud ved at have en langt højere koncentration af bakterier i tyndtarmen. Det samme gælder hvis man kigger på dyr fra Hold 3, fodret med varmebehandlede sporer (Tabel 4). På trods af at dyr fodret med Bt subsp. israelensis blev fodret med 100 gange færre sporer end dyr der fik B. cereus og Bt subsp. kurstaki, havde dyrene en højere koncentration i ileum (Tabel 4).
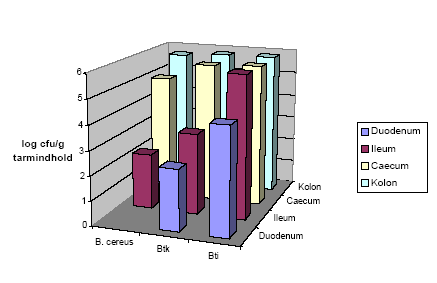
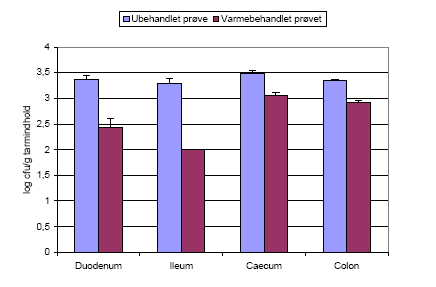
Figur 8. Samlet figur der viser antallet af hhv. B. cereus, Bt subsp. kurstaki (Btk) og Bt subsp. israelensis (Bti) i de tre forsøg. Tallene stammer fra de dyr der blev doseret med ubehandlede sporer (Hold 2). Detektionsgrænsen var 10 cfu/g tarmindhold for B. cereus og Btk, og 10³ cfu/g tarmindhold for Bti.
Ved aflivning af dyr to uger efter sidste dosering genfandtes kun få eller ingen af de tilsatte stammer i tarmprøverne fra dyrene. I dyr fodret med B. cereus fandtes stammen kun i kolon fra et dyr doseret med varmebehandlede sporer og overraskende i de nedre tarmafsnit fra et dyr fodret med vegetative celler (Tabel 5).
I dyr fodret med levende sporer af Bt subsp. kurstaki genfandtes stammen i flere tarmafsnit to uger efter sidste dosering. Og ligesom dyr 15 havde en meget højere koncentration af stammen i fæces (Figur 7B), ligeledes havde dette dyr en meget højere koncentration af Bt subsp. kurstaki end de andre dyr (Figur 9).
Vi fandt ingen Bt subsp. israelensis stammer i de dyr der blev doseret med denne stamme, og dette er i overensstemmelse med resultaterne fra fæcesprøverne.
Tabel 4. Genfindelse af B. cereus og de to anvendte Bt stammer i de forskellige tarmafsnit ved aflivning på Dag 5.
Tabel 5. Antallet af B. cereus og Bt subsp. kurstaki i tarmprøver fra HFA rotter doseret med hhv. ubehandlede sporer, varmebehandlede sporer og vegetative celler ved sidste aflivning (Dag 18).
| Tarmprøve | Koncentration af B. cereus og Bt subsp. kurstaki (log cfu g-1 tarmindhold) | |||||
| B. cereus | Bt subsp. kurstaki | |||||
| Ubehandlede sporer (Hold 2) |
Varme-behandlede sporer (Hold 3) |
Vegetative celler (Hold 4) |
Ubehandlede sporer (Hold 2) |
Varme-behandlede sporer (Hold 3) |
Vegetative celler (Hold 4) |
|
| Duodenum | < | < | < | < | 2,53±1,31 (2/3) | < |
| Ileum | < | < | < | < | 3,39 (1/3) | < |
| Caecum | < | < | 1,36 (1/3) | 1,30 (1/3) | 2,11±1,15 | < |
| Kolon | < | 2,83 (1/3) | 1,85 (1/3) | 1,43±0,38 | 2,64±0,91 (2/3) | < |
*< Indikerer at antallet af celler er under detektionsgrænsen (10 cfu/g tarmindhold)
† Tallet i parentes viser hvor mange dyr der var positive ud af tre.
Figur 9. Antallet af Bt subsp. kurstaki i dyr 15 doseret med varmebehandlede sporer af stammen. Derudover ses effekten af varmebehandling af tarmprøverne på antallet af bakterier. Varmebehandling dræber de vegetative celler, så det der tælles i prøven efter varmebehandling, er antallet af sporer. Hvorimod tælling af prøven uden varmebehandling angiver det totale antal bakterier, både sporer og vegetative celler. Forsøget blev udført to gange.
I forsøgene med de to Bt stammer blev der endvidere udtaget milt og lever fra dyrene og undersøgt for indhold af de tilsatte Bt stammer. Der blev kun fundet en positiv prøve, nemlig dyr 15 fra forsøget med Bt subsp. kurstaki. Dette dyr havde højere koncentration af stammen i både fæces og tarm (se forrige afsnit) og der blev fundet Bt subsp. kurstaki celler i milten.
3.1.1.3 Spirer sporerne i tarmen på HFA rotter?
I enkelte dyr viste udpladninger fra fæces (Figur 7) at sporerne må have spiret i tarmen. F.eks. dyr 13 doseret med varmebehandlede B. cereus sporer viste i slutningen af forsøgsperioden en stigning i antallet af bakterier i dyret, tydende på at der er sket spiring og vækst i mave-tarm kanalen på dette dyr. Et andet meget tydeligt eksempel er dyr 15 (fodret med varmebehandlede Bt subsp. kurstaki sporer), hvor der var en meget højere koncentration af bakterierne i både fæces og tarm end de andre dyr. Desuden viste varmebehandling (som dræber de vegetative celler) af tarmprøver fra dette dyr, at i tyndtarm var ca. 90% af bakterierne i vegetativ fase (Figur 9), mens de længere nede i tarmen igen havde sporuleret og dannet sporer. Dvs. at i dette dyr har Bt subsp. kurstaki haft en hel vækstcyklus fra spiring af sporen til vegetative celler, vækst af vegetative celler og igen sporulering af de vegetative celler til sporer.
Varmebehandling af tarmprøver fra første aflivning af dyr fodret med ubehandlede sporer af Bt subsp. israelensis viste også at sporerne var spiret i tarmen (Figur 10), da antallet af bakterier bliver reduceret efter varmebehandling, dvs. der er vegetative celler til stede i prøverne, der bliver dræbt under varmebehandlingen.
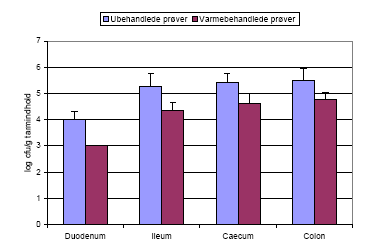
Figur 10. Tarmprøver fra dyr doseret med Bt subsp. israelensis sporer blev talt både før og efter varmebehandling. Tallet (log cfu/g tarmindhold) før varmebehandling giver det totale antal bakterier, sporer og vegetative celler, mens tallet efter varmebehandling angiver antallet af sporer i prøven, da de vegetative celler bliver dræbt af varmebehandlingen.
3.1.2 Overlevelse af sporer og vegetative celler i simuleret mavevæske
Da forsøgene viste at i dyr doseret med vegetative celler kunne de doserede stammer ikke genfindes, blev der lavet forsøg med overlevelse af bakterierne i simuleret mavevæske (SGF, simulated gastric fluid). Figur 11 viser at de vegetative celler hurtigt bliver inaktiveret i SGF, hvorimod både ubehandlede og varmebehandlede sporer har en god overlevelse. Dette understøtter resultaterne fra dyreforsøget med HFA rotter hvor ingen eller kun få bakterier kunne genfindes efter dosering med vegetative celler.
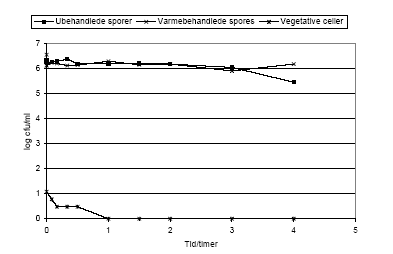
Figur 11. Stabilitet af sporer og vegetative celler af B. cereus F4433/73 i simuleret mavevæske ved pH 1.5.
3.1.3 Bt subsp. israelensis i kimfri dyr
Kimfri dyr blev fodret med en Bt subsp. israelensis stamme, som havde fået indsat et plasmid med et gfp gen, som får bakterien til at lyse grønt i vegetativ fase. Dyrene blev kun doseret med ubehandlede sporer, da de forrige forsøg viste at vegetative celler ikke overlever i mavesyren og der ikke var forskel på ubehandlede og varmebehandlede sporer.
Dyrene blev doseret fire dage i træk med ca. 107 sporer af Bt subsp. israelensis, dagen efter blev halvdelen af dyrene aflivet og de resterende dyr efter to uger.
Bt subsp. israelensis blev opretholdt stabilt i dyrene i hele forsøgsperioden på ca. 106 cfu/g fæces, og der sås ingen nedgang i antallet af bakterier som i forsøgene med HFA dyrene, så Bt klarer sig bedre i kimfri rotter uden en naturlig tarmflora (data ikke vist). Også tarmprøverne (Figur 12) viste at Bt subsp. israelensis har etableret sig i dyrene.
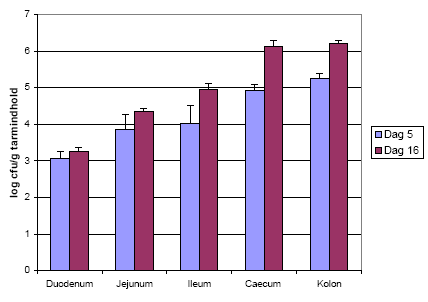
Figur 12. Antallet af Bt subsp. israelensis i tarmen fra kimfri dyr efter aflivning på hhv. dag 5 og dag 16.
3.1.3.1 Spirer sporerne i tarmen på kimfri rotter?
Flowcytometri (KU, phd studerende Martin Bahl) viste at grønne bakterier, dvs. vegetative celler kunne detekteres i alle tarmafsnit ved aflivning på dag 5 (Tabel 6) dvs. at sporerne var spiret og havde dannet aktive vegetative celler. Endvidere viste varmebehandling af prøverne fra tarm og fæces at der kom sporer ud med fæces som havde tabt gfp-plasmidet, dvs. at bakterien havde undergået en hel livscyklus i tarmen: sporerne var spiret til vegetative celler, som så igen havde sporuleret til sporer.
Varmebehandling (80 °C, 15 min) viste endvidere at de indgivne sporer havde spiret i tyndtarmen og selv i fæces fandtes størstedelen af bakterierne som vegetative celler.
Tabel 6. Resultater af flowcytometri og varmebehandling på prøver fra første aflivning af dyr fodret med sporer af Bt subsp. israelensis.
| Dyr | Prøve | Antal grønne bakterier = vegetative celler (log cfu/g tarmindhold) bestemt vha. flowcytometri | Antallet af vegetative celler (log cfu/g tarmindhold) bestemt vha. varmebehandling af tarmprøver |
| 3 | Duodenum | < | i.b. |
| Jejunum | < | i.b. | |
| Ileum | < | 3,07 | |
| Caecum | 3,60 | 4,62 | |
| Kolon | 4,47 | 5,03 | |
| 4 | Duodenum | < | i.b. |
| Jejunum | 2,45 | i.b. | |
| Ileum | 2,45 | i.b. | |
| Caecum | 4,07 | 4,50 | |
| Kolon | 4,46 | 4,77 | |
| 5 | Duodenum | < | i.b. |
| Jejunum | 3,23 | 3,61 | |
| Ileum | 3,23 | 3,48 | |
| Caecum | 4,73 | 4,37 | |
| Kolon | 4,62 | 4,39 |
Endvidere ses ved varmebehandling og udpladning på selektive plader, at der i løbet af forsøget udskilles sporer uden plasmid, og da celler kun taber plasmider ved deling betyder det at sporerne har spiret til vegetative celler, som har groet og så igen sporuleret (Figur 13).
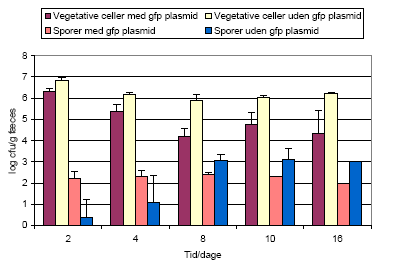
Figur 13. Bt subsp. israelensis i kimfri rotter. Antallet af sporer og vegetative celler udtrykt i cfu/g i fæces med og uden gfp plasmidet pAD44-12.
3.1.4 Opsummering
De tre dyreforsøg med HFA rotter doseret med a) B. cereus F4433/73R, b) Bt subsp. kurstaki og c) Bt subsp. israelensis, viste alle tre at i dyr fodret med vegetative bakterier genfandtes ingen eller kun få bakterier i fæces- og tarmprøver. Forsøg med sporer og vegetative celler af B. cereus og Bt subsp. kurstaki i simuleret mavevæske viste at de vegetative celler hurtig blev dræbt. Dette er i overensstemmelse med et andet studie [Clavel et al., 2004], som viste at under pH 4,5 har vegetative celler af B. cereus og Bt svært ved at overleve. Derfor kan den manglende genfindelse af B. cereus og Bt forklares ved at de vegetative celler ikke overlever passagen gennem mavesækken.
Varmeaktivering af sporerne før dosering havde ingen indflydelse på deres overlevelse og spiring i tarmmiljøet, dvs. at varmebehandling af fødevarer f.eks. ved tilberedning ikke øger risikoen for spiring af Bt i tarmen. Både ubehandlede og varmebehandlede sporer kunne detekteres med den højeste koncentration den første uge med dosering, hvorefter antallet hurtig faldt. Dog kunne vi selv to uger efter sidste dosering genfinde B. cereus og Bt i enkelte dyr.
I forsøget med Bt subsp. israelensis i HFA rotter kunne bakterien ikke detekteres i dyrenes fæces to uger efter sidste dosering, dette skyldes dog formodentlig at bakterien i modsætning til de to andre undersøgte stammer ikke var rifampicin resistent men streptomycin resistent. Streptomycinresistente bakterier er almindelig blandt en normal menneske tarmflora, og disse har formodentlig hindret den resistente subsp. israelensis stamme i at vokse op på de selektive plader. Det kan derfor ikke udelukkes at der ligesom i de to andre forsøg stadig er Bt subsp. israelensis stammer tilbage i dyrene to uger efter dosering.
Der var et enkelt dyr doseret med varme-aktiverede sporer af Bt subsp. kurstaki, som havde en meget højere koncentration af denne stamme i fæces end de andre dyr. Ved aflivning to uger efter dosering blev den samme høje koncentration fundet i alle tarmafsnit og varmeaktivering af tarmprøver fra tyndtarmen, viste at 90% af cellerne her var i den vegetative fase. Dette viser at i dette dyr er der sket spiring og re-sporulering. Desuden blev der i dette dyr fundet Bt subsp. kurstaki celler i milten, dvs. at Bt subsp. kurstaki celler kan passere tarm-barrieren og nå andre organer. Passage af tarm-barrieren blev ikke observeret for Bt subsp. israelensis i hverken HFA eller kimfri dyr.
Forsøgene med Bt subsp. israelensis mærket med et grønt fluorescerende protein og undersøgt i kimfri rotter viste tydeligt at indgivne sporer både spirer og re-sporulerer, dvs. at de har en hel livscyklus i tarmmiljøet. Selv i fæces fandtes bakterierne at være i det vegetative stade. Denne forskel fra resultaterne med HFA dyr skyldtes sandsynligvis fraværet af en omkringliggende tarmflora i de kimfri dyr, hvilket giver Bt meget bedre vækstbetingelser i hele mave-tarm kanalen.
3.2 Ændringer af den normale tarmflora
En eventuel påvirkning af den normale tarmflora efter dosering med B. cereus F4433/73, Bt subsp. kurstaki eller Bt subsp. israelensis blev studeret både ved simple kimtællinger af bestemte bakteriegrupper og ved en molekylærbiologisk metode, som i princippet skulle fange alle bakterier i prøven. Der blev taget fæcesprøver før, under og efter dosering, derudover blev tarmprøver også undersøgt.
3.2.1 Kimtællinger af bakteriegrupper
Som beskrevet i Materialer og Metoder blev følgende bakteriegrupper undersøgt for eventuelle ændringer efter dosering med B. cereus og Bt: Total aerobe, total anaerobe, laktobaciller, koliforme og enterokokker. Disse blev udvalgt da de er dyrkbare og hører til de store grupper af bakterier i tarm.
Ingen store ændringer blev observeret i fæces- og tarmfloraen ved dyrkning af de ovenstående nævnte bakteriegrupper før, under og efter dosering. Der blev fundet enkelte små ændringer som er opsummeret i Tabel 7. Ved aflivning to uger efter sidste dosering blev der ikke observeret forskelle mellem holdene.
Tabel 7. Observerede ændringer af bestemte bakteriegrupper i fæces- og tarmprøver. Ændringer i bakteriesammensætningen i tarmprøverne ses kun ved første aflivning på Dag 5.
| Doseret bakteriestamme | Tarmafsnit | Ændring |
| B. cereus F4433/73 | Ileum | Nedgang i antalllet af laktobaciller i dyr doseret med varmebehandlede sporer |
| Caecum | Nedgang i antallet af koliforme i dyr doseret med levende sporer | |
| Kolon | Nedgang i antallet af totale aerobe og anaerobe bakterier i dyr doseret med varmebehandlede sporer | |
| Bt subsp. kurstaki DMU67 | Caecum og kolon | Nedgang i antallet af aerobe, anaerobe, laktobaciller og koliforme i dyr doseret med ubehandlede sporer |
| Bt subsp. israelensis HD567 | Fæces i doseringsperioden | Øget antal enterokokker i dyr doseret med sporer (både inaktiverede og levende) |
3.2.2 Ændringer af tarmflora studeret vha. DGGE
Da en stor del af bakterierne i tarmen ikke kan dyrkes på næringsmedier i laboratoriet, har vi anvendt en molekylærbiologisk metode, DGGE (denaturerende gradient gel elektroforese) som i princippet fanger alle bakterier, der udgør mere end 1% af populationen.
I projektet har vi undersøgt fæcesprøver fra alle tre forsøg med HFA dyr. Der er blevet taget prøver til DGGE før, under og efter dosering. Prøverne blev i forsøget med B. cereus F4433/73 undersøgt både med generelle primere der detekterer hele bakteriepopulationen og med specifikke primere til at finde ændringer i Bacteroides, laktobaciller og bifidobakterier. Disse tre grupper af bakterier menes at have betydning for menneskers tarmfunktion og immunstatus. I de to forsøg med Bt er prøverne kun analyseret vha. generelle primere, da forsøget med B. cereus ikke viste ændringer i de førnævnte specifikke populationer.
3.2.2.1 B. cereus
Som sagt var der ikke ændringer i de specifikke grupper (Bacteroides, laktobaciller og bifidobakterier) efter dosering med B. cereus F4433/73. Det samme gjaldt for hele bakteriepopulationen i de dyr der blev doseret med inaktiverede (strålesteriliserede) sporer og vegetative celler. Derimod viste PCR-DGGE af prøver fra dyr doseret med levende sporer (enten ubehandlede eller varmebehandlede) en klar gruppering af bakteriepopulationerne i grupper før, under og efter dosering (Figur 14, Bilag A).
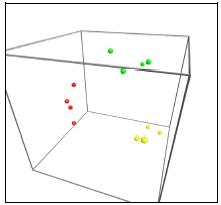
Figur 14. Computeranalyse af tarmfloraen hos rotter med menneske-flora før, under og efter fodring med sporer af B. cereus. Prøver fra før, under og efter fordring er markeret med henholdsvis rødt, grønt og gult. Jo tættere prikkerne ligger på hinanden, des mere ens var floraen i fæcesprøverne.
Disse grupperinger kunne tilskrives ændringer i tilstedeværelsen af bånd eller intensiteten af bånd på DGGE gelen (Figur 15). Kloning, sekventering og sammenligning med kendte sekvenser i databaser viste at bakterierne i de udskårne bånd var identiske med forskellige anaerobe ikke-dyrkbare tarmbakterier.
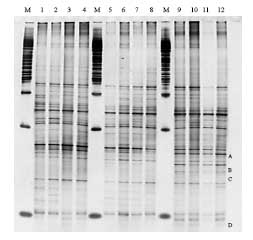
Figur 15. PCR-DGGE profiler genereret fra fæcesprøver af dyr doseret med varmebehandlede sporer af B. cereus F4433/73. Bane 1-4: prøver fra individuelle dyr før dosering; bane 5-8: poolede prøver fra dyr i doseringsugen; bane 9-12: poolede prøver fra dyr efter dosering. M: markør. Båndene blev bestemt til at tilhøre følgende genera af bakterier: Bånd A: Ruminococcus obeum; bånd B: Fusobacterium sp.; bånd C: Clostridiacease familien; bånd D: genus annerella.
3.2.2.2 Bt subsp. kurstaki
I modsætning til forsøget med B. cereus har vi med prøverne fra Bt forsøgene ikke anvendt samlede prøver fra flere dyr, men set på fæcesprøver på enkeltdyr fra før, under og efter dosering med Bt.
PCA-analyse (principal komponent analyse) af DGGE gelerne viser at for de dyr der fik vegetative celler og inaktiverede sporer sker der en ændring i floraen i undersøgelsesperioden, hvor floraen efter dosering adskiller sig fra floraen før og under.
I de rotter der fik levende sporer (både ubehandlede og varmebehandlede) ses i nogle af dyrene en ændring af bakteriesammensætningen under dosering (Bilag A). Dette gælder for dyr 7 fra Hold 2 og dyr 15 fra Hold 3, som begge i perioder har en højere koncentration af Bt subsp. kurstaki i fæces end de andre dyr i holdene (Figur 7B). Som det ses af Bilag A (Figur 1.8.3 og 1.8.6) er der bånd der bliver kraftigere under dosering. Kloning og sekventering af disse bånd viste op til 99% identitet med Faecalibacterium prausnitzii, en dominerende gruppe i en normal human fæcesflora.
3.2.2.3 Bt subsp. israelensis
Også i forsøget med underarten israelensis sås en ændring i bakterie-sammensætning under dosering med sporer af bakterien (Bilag A), men da der også sås en ændring i kontrolgrupperne fodret med hhv. vegetative celler og inaktiverede sporer er det uvist om ændringen skyldes doseringen med stammen.
3.2.3 Opsummering
Der blev kun observeret få og midlertidige ændringer af den omkringliggende bakterieflora, og vi kan konkludere at påvirkningen af den normale tarmflora er minimal. Nogle stammer af B. cereus bliver som sagt brugt som probiotika til mennesker og dyr og et studie med en sådan probiotisk stamme har vist en effekt på E. coli og enterokok populationen [Gedek et al., 1993]. Endvidere har et studie vist en signifikant reduktion i antallet af koliforme bakterier i tarmprøver fra smågrise fodret med en probiotisk B. cereus stamme (Jadamus et al, 2002). Den samme nedgang i antallet af koliforme bakterier har vi observeret i vores forsøg med B. cereus, dog var reduktionen kun midlertidig. Det er usikkert hvorvidt denne påvirkning af den omkringliggende bakterieflora skyldes vegetative celler eller sporer af B. cereus og Bt. Som forsøgene viser er bakterierne i de fleste dyr primært tilstede som sporer, og det er derfor sandsynligt at det er disse der er ansvarlige for ændringerne, evt. ved at binde sig til tarmepitelceller og derved optage plads for andre bakterier.
I to rotter fodret med sporer af Bt subsp. kurstaki sås en ændring af bakteriesammensætningen i fæces under dosering ved DGGE. Dette gælder for dyr 7 fra Hold 2 og dyr 15 fra Hold 3, som begge i perioder har en højere koncentration af Bt subsp. kurstaki i fæces end de andre dyr i holdene. Under dosering er der bånd der bliver kraftigere på DGGE gelen. Kloning og sekventering af disse bånd viste op til 99% identitet med Faecalibacterium prausnitzii, en dominerende gruppe i en normal human fæcesflora. F. prausnitzii er en butyrat-producerende bakterie, som menes at danne energi til tarmepitelceller og har en god indflydelse på sundhedstilstanden i tyktarmen. Da der ses en stigning af denne bakterie i doseringsperioden kunne det tyde på at Bt subsp. kurstaki har en positiv effekt på netop denne bakterie.
PCA-analyse (principal komponent analyse) af DGGE gelerne viser at for de dyr der fik vegetative celler og inaktiverede sporer sker der en ændring i floraen i undersøgelsesperioden, hvor floraen efter dosering adskiller sig fra floraen før og under. Dette er formodentlig en naturlig ændring der sker i tarmfloraen på HFA rotter, en bakteriesammensætning, som i forhold til konventionelle rotter ikke er helt stabil, da den ikke er naturlig for rotter.
3.3 Enterotoksiner i tarmen
Inden der blev kigget efter enterotoksiner i tarmprøver vha. Vero-celle assay og et kommercielt kit fra Oxoid (BCET-RPLA) var det nødvendigt at lave en del indledende forsøg for at optimere forsøgsbetingelserne, bestemme detektionsgrænser m.m. Disse forsøg er ikke beskrevet her, men i afsnittet om Materialer og Metoder, paragraf 3.5.
3.3.1 Vero-toksiciteten af forskellige bakterier
Vero-toksiciteten af forskellige stammer dyrket under optimale betingelser er vist i Tabel 8. Bt subsp. israelensis HD-567 havde den højeste aktivitet, dette kan sandsynligvis tilskrives en celledræbende faktor som produceres samtidig med krystal-toksinet. Ingen af de andre stammer har denne faktor. De to andre kommercielle stammer, subsp. kurstaki og subsp. tenebrionis har lavere aktivitet end det sygdomsfremkaldende isolat B. cereus F4433/73. Endelig har også den probiotiske stamme B. cereus BCIP 5832 lav Vero-toksisk aktivitet.
Tabel 8. Vero-toksicitet af referencestammer og stammer brugt til rotteforsøg. Fortyndingsfaktoren er det antal gange bakteriesuspensionen er blevet fortyndet for at opnå mindst 50% overlevelse af Vero-cellerne. Dvs. jo højere fortyndingsfaktor jo mere toksisk er stammen.
| Stamme | Fortyndingsfaktor |
| B. cereus F4433/73 | 160 |
| B. cereus BCIP 5832 | 20 |
| B. thuringiensis subsp. Kurstaki HD-1 | 80 |
| B. thuringiensis subsp. Kurstaki DMU67R | 80 |
| B. thuringiensis subsp. israelensis HD-567 | 640 |
| B. thuringiensis subsp. tenebrionis NB-125 | 80 |
3.3.2 Detektion af Vero-toksisk aktivitet i tarmprøver fra rotter
I ingen af forsøgene med HFA rotter kunne der detekteres Vero-toksisk aktivitet i prøverne fra tarm og fæces. Hverken i de dyr der var doseret med den sygdomsfremkaldende B. cereus F4433/73 eller i de dyr doseret med de to underarter af Bt (subsp. kurstaki og subsp. israelensis).
I de kimfri rotter doseret med sporer af Bt subsp. israelensis blev komponenten L2 fra HBL detekteret i et enkelt dyr fra tyndtarmsprøve vha. det kommercielle kit BCET-RPLA (Tabel 9).
Tabel 9. Resultat af Vero-assay og Oxoid test af tarmprøver fra mono-associerede rotter. Fortynding: Tarm-prøve ekstrakterne fortyndes 10, 20, 40, 80, 160, 320, 640 og 1280 gange. Den angivne fortynding nedenfor angiver ved hvilken fortynding der er mere end 50% af cellernes aktivitet tilbage. Kun de ekstrakter hvor der enten står et ”-” eller et ”+” er blevet testet i OXOID analysen.
| Dyr | prøve | Fortynding | Oxoid test |
| 1 | J | 40 | - |
| 1 | I | Mindre end 10 | - |
| 1 | Ca | Mindre end 10 | - |
| 3 | J | 40 | - |
| 3 | I | 20 | + |
| 3 | Ca | Mindre end 10 | - |
| 4 | J | 40 | - |
| 4 | I | Mindre end 10 | |
| 4 | Ca | Mindre end 10 | |
| 5 | J | 20 | - |
| 5 | I | Mindre end 10 | |
| 5 | Ca | Mindre end 10 | |
| 6 | J | 80 | - |
| 6 | I | Mindre end 10 | |
| 6 | Ca | Mindre end 10 | |
| 7 | J | 40 | - |
| 7 | I | Mindre end 10 | |
| 7 | Ca | Mindre end 10 | |
| 8 | J | 40 | - |
| 8 | I | Mindre end 10 | - |
| 8 | Ca | Mindre end 10 | - |
| 6 | Kolon | Mindre end 10 | |
| 7 | Kolon | Mindre end 10 | |
| 8 | Kolon | Mindre end 10 |
Desuden sås udslag i Vero-celle assayet i jejunum (midterste del af tyndtarm) i alle dyr, også i kontroldyr 1, som ikke blev doseret med Bt subsp. israelensis (Tabel 9).
3.3.3 Opsummering
Inden prøver fra dyr kunne testes for tilstedeværelsen af enterotoksiner var det nødvendigt at udføre en del indledende forsøg, som beskrevet i Materialer og Metoder. I disse indledende forsøg blev forsøgsbetingelserne optimeret, det blev vist at fæces- og tarmekstrakt ikke influerer på genfindelse af enterotoksiner, at fæcesprøver ikke i sig selv er giftige for cellerne, og at der skal være mindst 106 bakterier per ml til stede for at der kan måles Vero-toksisk aktivitet.
Vero-celle assay på de anvendte stammer viste at den højeste toksin-aktivitet fandtes i Bt subsp. israelensis, formodentlig pga. et nonspecifikt cytolytisk toksin der dannes samtidig med krystaltoksinet. Denne stamme bruges i Danmark kun på prydplanter. De andre kommercielle stammer, subsp. kurstaki og subsp. tenebrionis, som benyttes til spiselige afgrøder, og hvoraf kun subsp. kurstaki er tilladt på det danske marked, havde en lavere aktivitet end det B. cereus isolat der har givet anledning til diarré.
At der ikke kunne detekteres Vero-toksisk aktivitet i HFA dyrene kan skyldes at vegetative former af de tilsatte bakterier ikke fandtes i tilstrækkelig høj densitet i tyndtarmen, hvor produktionen af enterotoksiner menes at ske [Granum and Lund, 1997]. De indledende forsøg viste at der skal være mindst 106 bakterier per ml til stede for at der kan detekteres cytotoksisk effekt på Vero-celler, og i tyndtarmsprøverne detekterede vi højst 104 bakterier per g tarmindhold.
I forsøgene med kimfri rotter associeret med Bt subsp. israelensis sås udslag i Vero-celle assayet i jejunum (midterste del af tyndtarm) i alle dyr, også i kontroldyr 1, som ikke blev doseret med Bt subsp. israelensis (Tabel 9). Derfor er det sandsynligvis ikke bakterien der har en negativ effekt på Vero-celler, men kan måske skyldes galde-salte eller pankreas-enzymer der er blevet udskilt til tyndtarmen. Kun i et dyr ud af seks fra forsøgene med kimfri rotter fandtes cytotoksisk aktivitet i Vero-celle assay, samt positivt respons fra Oxoid kittet som detekterer L2 komponenten fra enterotoksin HBL.
3.4 Overførsel af DNA mellem to Bt stammer i kimfri rotter
3.4.1 Dyreforsøg med kimfri dyr
Bakterierne i B. cereus gruppen er kendt for at indeholde plasmider (cirkulært DNA) hvoraf mange kan overføres fra den ene bakterie til den anden ved en proces der kaldes konjugation. Der findes mange artikler som beskriver denne overførsel i laboratoriet, og det er også blevet vist at der kan ske overførsel vha. konjugation i tarmen på insekter. Det er dog aldrig blevet undersøgt i tarmen hos pattedyr, og det er interessant fra et evolutionært og økologisk synspunkt hvorvidt Bt er i stand til at udveksle DNA i et sådant miljø, og er en vigtig faktor i risikovurderingen af Bt midler. Derudover afspejler det bakteriel aktivitet. En eventuel overførsel viser nemlig at de indgivne sporer må have spiret og dannet vegetative celler, idet kun aktive celler kan overføre DNA.
For at øge muligheden for at detektere en eventuel overførsel mellem to Bt stammer valgte vi at bruge kimfri rotter, som blev doseret med to isogene stammer, dvs. bakterier som er ens bortset fra indholdet af plasmider. Seks dyr blev først doseret med sporer af recipientstammen fire dage i træk, ugen efter fik dyrene så donorstammen som sporer fire dage i træk. Donorstammen indeholdt det konjugative plasmid pXO16, som i laboratoriet overføres med meget høj frekvens til recipientstammen. Halvdelen af dyrene blev aflivet dagen efter sidste dosering med donorstammen og de resterende tre en uge efter.
Der blev detekteret transkonjuganter (dvs. recipienter som har modtaget plasmidet pXO16 fra donoren) i alle dyr ved den første aflivning (Figur 16). Som det ses af figuren (store standardafvigelser) dækker tallene over store individuelle forskelle i specielt antallet af donorer og transkonjuganter i de enkelte dyr. Der blev dog fundet transkonjuganter i alle dyr i alle tarmafsnit, på nær et dyr hvor der ikke kunne detekteres transkonjuganter i den øverste del af tyndtarmen (duodenum). Ved aflivning en uge efter sidste dosering med donorstammen blev der kun fundet få donorceller og ingen transkonjuganter.
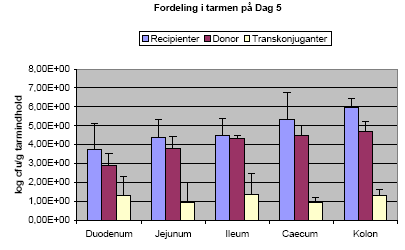
Figur 16. Antallet af recipienter, donorer og transkonjuganter i de forskellige tarmafsnit ved aflivning dagen efter sidste dosering med donorstammen. Gennemsnit af tre dyr.
Transkonjuganterne blev isoleret ved at plade ud på selektive plader, som kun transkonjuganterne kunne vokse på. For at sikre at det var ægte transkonjuganter og ikke spontane mutanter, blev der lavet plasmid-oprensninger som blev undersøgt ved gel-elektroforese. Dette viste at alle transkonjuganter var ægte transkonjuganter indeholdende plasmidet pXO16 (Figur 17).
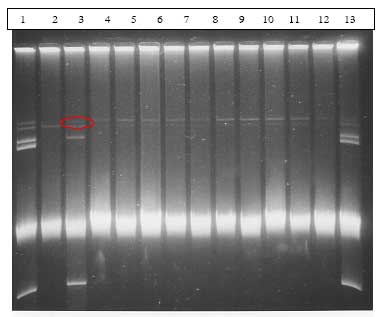
Figur 17. Gelbillede af transkonjuganter opnået i dyreforsøg med en Bt subsp. israelensis donor med det konjugative plasmid pXO16 og en Bt subsp. israelensis recipient. Bane 1 og 14: vildtypestamme af Bt subsp. israelensis (AND508), Bane 2: recipient, Bane 3: donor, Bane 4-13: transkonjuganter. Det konjugative plasmid pXO16 kan svagt detekteres i donorstammen (rød ring), i den oprindelige vildtypestamme, samt i alle transkonjuganter.
3.4.2 Opsummering
Forsøget med kimfri rotter viste at Bt stammer er i stand til at udveksle plasmider in vivo i en tarm fra pattedyr. Forsøget viste dog også at bakterier indeholdende det konjugative plasmid, pXO16, ikke havde en vækstfordel i tarmmiljøet, da der i slutningen af forsøget primært kunne detekteres recipientstammer i dyrene, kun få donorstammer og ingen transkonjuganter.
Version 1.0 Januar 2007, © Miljøstyrelsen.