|
Bekæmpelsesmiddelforskning fra Miljøstyrelsen nr. 99, 2006 Toksiske effekter af pulseksponering af pesticider- akutte og kroniske effekter hos Daphnia magnaIndholdsfortegnelse2 Udvalgte pesticider og forsøgsorganisme
Bilag 3 Data for ISO-test (akut) Bilag 4 Pulstest - akutte effekter Bilag 5 Gentagne pulseksponeringer Bilag 6 Pulstest - kroniske effekter ForordDenne rapport ”Toksiske effekter af pulseksponering for pesticider - akutte og kroniske effekter hos Daphnia magna” er udarbejdet for Miljøstyrelsen i perioden 1. januar 2004 til 31. marts 2005. Projektet er udført på Institut for Miljø & Ressourcer, Danmarks Tekniske Universitet (M&R DTU) af: Cand.scient. Tina Slothuus (M&R DTU) Laborant Signe Qualmann (M&R DTU) Lektor, civilingeniør, ph.d. Anders Baun (Projektleder, M&R DTU) Civilingeniør Tobias H. Andersen og stud. polyt. Rikke Tjørnhøj har bidraget til med resultater og metodeudvikling især i forbindelse med kroniske test og test med gentagne pulse. Begge takkes for deres store og engagerede arbejdsindsats. Lektor Klaus K. Andersen Institut for Informatik og Matematisk Modellering Danmarks Tekniske Universitet, Kgs. Lyngby har gennemført den statistiske analyse af data fra test for kronisk virkning, og forfatterne takker herfor. Projektet har været fulgt af en følgegruppe bestående af følgende medlemmer: Gitte Larsen, Skov- og Naturstyrelsen Dean Jakobsen, Ferskvandsbiologisk Laboratorium, Københavns Universitet Alf Aagaard og Vibeke Møller, Miljøstyrelsen Peter Wiberg-Larsen, Fyns Amt Jørgen Jakobsen, Danmarks Jordbrugsforskning Niels Martin Frost, Dansk Planteværn Poul Bjerregaard, Biologisk Institut, Syddansk Universitet Ulrik Nørum, Biologisk Institut, Syddansk Universitet Jens C. Streibig, KVL Poul Henning Petersen, Dansk Landbrugsrådgivning Dorthe Lærke Baun, DHI Vand & Miljø René Juhler, GEUS Følgegruppen (i særlig grad Ulrik Nørum, Niels Martin Frost, Alf Aagaard og Vibeke Møller) samt professor Valery Forbes, Roskilde Universitet takkes for konstruktiv kritik af det udførte arbejde og den endelige afrapportering. SammenfatningUdledning af drænvand eller afstrømning fra marker i forbindelse med regnvejr er sammen med udsprøjtning af pesticider eksempler på hændelser, der kan føre til kortvarit høje koncentrationer af pesticider i overfladevand. Flora og fauna i overfladevandet udsættes dermed for en puls af pesticider, og man refererer ofte til dette som en pulseksponering. De standardiserede metoder, der anvendes til undersøgelse af stoffers økotoksicitet, bliver alle udført med en vedvarende (kontinueret) eksponering af test organismerne i et nærmere specificeret tidrum. Derfor tager disse metoder ikke hensyn til evt. effekter, der følger af en pulseksponering. Formålet med det udførte projekt var således at udvikle og afprøve en ny metode til undersøgelse af toksiske effekter af pulseksponering ved anvendelse af freskvandskrebsdyret Daphnia magna. Otte pesticider (dimethoat, chlorpyrifos, pyrazophos, esfenvalerat, pirimicarb, diquat, bromoxynil og azoxystrobin) og to reference stoffer (4-nitrophenol og 3,5-dichlorophenol) blev undersøgt. Ved pulstestene blev nyfødte Daphnia magna (<24 timer gamle) udsat for teststofferne i 0.5-6 timer. Efter disse eksponeringer blev dyrene overført til rent vand. I de følgende 48 timer blev dyrenes bevægelsesevne (mobilitet) observeret. I praksis skete dette umiddelbart efter overførsel (0 timer) og efter 24 og 48 timer. For stofferne dimethoat, pirimicarb, esfenvalerat, chlorpyrifos, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol blev observationsperioden udvidet til 21 dage for at undersøge om pulseksponeringerne førte til kroniske effekter. Ved disse undersøgelser blev følgende observeret: Tiden til første afkom, antal unger samt længde og vægt af moderdyrene efter 21 dage i rent vand. Endelig blev effekten på mobiliteten og dødeligheden af Daphnia magna undersøgt efter at dyrene blev udsat for to pulse af samme varighed og koncentration. Under og umiddelbart efter pulseksponering med dimethoat, pirimicarb, esfenvalerat, bromoxynil, 4-nitrophenol, and 3,5-dichlorophenol var dyrene ikke i stand til at bevæge sig (immobile), men efter 24 timers ophold i rent vand genvandt de mobiliteten. For azoxystrobin blev en sådan genvinding af mobiliteten ikke observeret, og for chlorpyrifos, pyrazophos og diquat steg antallet af immobile og døde dyr i perioden efter pulseksponeringens ophør. Gentagne pulseksponeringer gav mere udtalte effekter på mobiliteten af Daphnia magna en enkelt-pulse. Selv i grupper, hvor den første puls tilsyneladende ikke påvirkede dyrenes mobilitet, var resulatet af anden pulseksponering at en stor del af dyrene blev immobile eller døde. Efter pulseksponering for dimethoat og pirimicarb blev der registreret en reduceret størrelse af moderdyerne. Hos de samme dyr blev der registreret både en forsinkelse og et samlet fald i antal unger produceret i løbet af 21 dage efter at eksponering ophørte. Antallet af unger faldt signifikant jo længere pulsen varede, men selv for pulse af 2 timers varighed kunne signifikante effekter observeres. Der blev fundet en signifikant sammenhæng mellem dyrenes længde efter 21 dage og antallet af unger. Det er muligt, at den nedsatte størrelse skyldes et mindre optag af føde efter pulsenes ophør, og at dette samlet set medfører en mindre produktion af unger, og men dette er ikke blevet undersøgt nærmere i dette projekt. I de udførte test med pesticiderne esfenvalerat, chlorpyrifos, bromoxynil, 4-nitrophenol eller 3,5-dichlorophenol blev der ikke set signifikante effekter på reproduktion efter pulseksponeringer, selvom dyr udsat for 3,5 dichlorophenol var mindre ved testens ophør (21 dage efter eksponering) end ikke-eksponerede dyr. SummaryDrainage from fields, surface runoff after rain events, or spraying of pesticides are all examples of events that may lead to short-term high concentrations of pesticides in surface water. These kinds of events are often referred to as pulses of pesticides. The standardized aquatic toxicity tests used in hazard assessment are performed with a continous exposure of test organims for a specified period of time. Thus, these methods do not take effects of pulse exposure into account. The aim of the present study has been to develop and test a new method to detect toxic effects of pulse exposure to pesticides using the fresh water crustacean Daphnia magna. Eigth pesticides (dimethoate, chlorpyrifos, pyrazophos, esfenvalerate, pirimicarb, diquat, bromoxynil, and azoxystrobin) and two reference compounds (4-nitrophenol and 3,5-dichlorophenol) were tested. In the pulse exposure tests juvenile Daphnia magna (<24 h old) were exposed to the test compounds for 0.5-6 hours. After these exposures, the animals were transferred to clean water and observed for 48 hours. During this post-exposure period changes in the mobility of the animals were registered immediately after the transfer (0 hours) and after 24 and 48 hours. For the compounds dimethoate, pirimicarb, esfenvalerate, chlorpyrifos, bromoxynil, 4-nitrophenol, and 3,5-dichlorophenol the observation period was prolonged to 21 days to study chronic effects of pulse exposure. In these studies, the following endpoints were included: Time to first offspring, number of offspring, length and weigth of surviving mother animals. Furthermore, the effect on the mobility of Daphnia magna of two consequtive pulse exposures to dimethoate, pirimicarb, esfenvalerate, and pyrazophos was studied. During and immidiately after pulse exposures to dimethoate, pirimicarb, esfenvalerate, bromoxynil, 4-nitrophenol, and 3,5-dichlorophenol the animals had lost their mobility, but after 24 hours in clean water they regained mobility. After pulse exposure to azoxystrobin no such recovery was observed during the 48-h post-exposure period, and for chlorpyrifos, pyrazophos, and diquat the number of immobile and/or dead animals increased after the pulse exposure ended. Repeated pulse exposures resulted in more pronounced effect on the mobility of Daphnia magna than single pulse exposures. In groups exposed to short pulses, animals, which apparently were not affected by one pulse, lost their mobility and an increased number of dead animals was observed after a second pulse exposure. Delays in the time to first offspring and reduced number of offsprings were observed in Daphnia magna after pulse exposure to dimethoate and pirimicarb. Significant reduction in the number of offsprings was observed for pulse durations of 2 hours and the number of offspring decreased with increasing pulse duration. Mother animals exposed to pulses of dimethoate or pirimicarb for 2 hours or more were also significantly smaller than unexposed animals, and a correlation between the size of the animals after 21 days and the number of offspring was observed. This effect may be related to a lower uptake of food in the post-exposure period, but this was not studied in the present project. On the other hand, pulse-exposure tests with esfenvalerate, chlorpyrifos, bromoxynil, 4-nitrophenol, and 3,5-dichlorophenol did not reveal any significan decrease in number of offspring after 21 days, eventhough animals exposed to 3,5-dichlorophenol were smaller than non-exposed animals. 1 Indledning1.1 Traditionelle akvatiske toksicitetstests og pulseksponeringer i naturenToksiske effekter afhænger både af koncentrationen af et kemikalie samt af eksponeringstiden (Timbrell, 1990). Typisk vil der således skulle lavere og lavere koncentrationer til af et kemikalie for at opnå en given effekt jo længere eksponeringstiden er (Buhl et al. 1993). Traditionelle akvatiske test til beskrivelse af stoffers toksiske effekter bygger på vedvarende (kontinuerte) eksponeringer af forsøgsorganismerne i et nærmere bestemt tidsinterval, der er som oftest er fastlagt med udgangspunkt i organismens livscyklus frem for eksponeringsformerne i naturen. Der anvendes enkeltarter af ”standardorganismer” (fisk, krebsdyr og alger), og testvilkårene er fastlagte hvad angår fysiske, kemiske og biologiske forhold som f.eks. lys/mørke cyklus, temperatur, medier og fodring under testen. Alle disse forhold fastholdes med henblik på at sikre høj reproducerbarhed i testen (Møhlenberg et al. 2004). Typisk er eksponeringstiden 96 timer for fisk, 24-48 timer for krebsdyr og 72 timer for alger. I naturen optræder forureningshændelser imidlertid ofte med varierende stofkoncentrationer af få timers til få dages varighed - en såkaldt puls (Herricks et al. 1997; Styczen et al. 2003; Slothuus et al. 2004). Eksempler på hændelser, der kan lede til pulseksponeringer i recipienterne kan være: regnskyl, afstrømninger fra marker, drænudledninger, brugen af antibiotika og andre hjælpemidler i dambrug, drift fra udsprøjtninger af pesticider eller udledning fra vaskepladser, hvor sprøjteudstyr fyldes op med pesticider eller rengøres efter brug o.s.v. (McCahon.C.P. et al. 1990; Wiberg-Larsen et al. 1991; Buhl et al. 1992; Herricks et al. 1997; Brent & Herricks 1998; Slothuus et al. 2004). Endvidere er det værd at bemærke, at også pesticider med kort halveringstid i miljøet kan udvise ”pulsadfærd”, hvis de tilføres vandmiljøet (Reinert et al., 2002). Pulsudledninger kan altså give anledning til forbigående, høje koncentrationer af pesticider i vandmiljøet, og det er i flere studier vist, at denne type eksponering kan resultere i større effekter end kontinuerte eksponeringer (jf. beskrivelse hos Tucker & Burton, 1999). Generelt kan det dog konstateres, at de toksiske effekter af pulseksponeringer med pesticider kun i mindre grad er adresseret i den internationale litteratur (Reinert et al., 2002). Dette skyldes sandsynligvis en kombination af, at pulseksponeringer er vanskelige at kvantificere som følge af deres pludselige og kortvarige natur, og at egnede testmetoder til belysning af pulseksponeringers virkning mangler. Selv om pulseksponeringer tildeles mere og mere opmærksomhed, f.eks EU Technical Guidance Document on Risk Assesment of Chemicals (EU, 2003), tager de traditionelle tests ikke hensyn til den varighed eksponeringen af dyrene kan have i naturen. Det er veldokumenteret, at standardiserede laboratoriebaserede metoder, som anvender kontinuerte eksponeringsscenarier for pesticider, som regel ikke resulterer i data, som er egnede til vurdering af effekter af pulseksponeringer (Reinert et al., 2002). Denne problemstilling, og hvad der sker med organsimer efter eksponeringens ophører, er tidligt beskrevet af Wright (1976). Siden 1976 er denne problemstilling adresseret flere gange f.eks. af Buhl et al. (1993) og Brent & Herricks (1999), hvis undersøgelser med indbyggede observationsperioder efter eksponeringens ophør har vist, at der kan ske efterfølgende ændringer i responset. Nærværende projekt er derfor fokuseret på udvikling og afprøvning af en metode til undersøgelse af toksiske effekter af pulseksponering for pesticider. Ved metoden anvendes ferskvandskrebsdyret Daphnia magna som testorganisme, og der inddrages såvel korterevarende (immobilisering) som længerevarende effekter (reproduktion) relateret til pulseksponeringer. 1.2 FormålI dette projekt bliver ændringer i mobiliteten og reproduktionsmønsteret hos Daphnia magna undersøgt efter pulseksponering med pesticider. Formålet er ligeledes at observere hvad der sker med dyrene i løbet af de første 48 timer efter eksponeringens afslutning. I projektet fokuseres i særlig grad på effekten af pulsenes varighed, og i mindre grad på effekten af eksponeringskoncentrationen. De anvendte teknikker bygger på traditionelle ISO standarder og OECD guidelines, og tager ligeledes udgangspunkt i metodikken beskrevet af Brent & Herricks (1999). Formålet med projektet består af tre dele:
2 Udvalgte pesticider og forsøgsorganisme
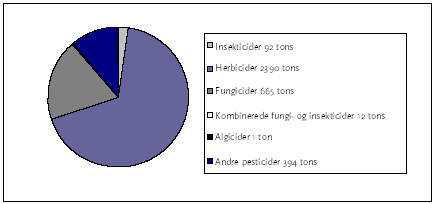
2.1 Pesticidforbrug og -fund i vandmiljøet i DanmarkPesticider er kemiske forbindelser, der bruges til at kontrollere forskellige former for skadevoldere i bl.a. landbruget. Pesticiderne inddeles således i grupper afhængig af den målgruppe, de bl.a. anvendes imod, og i Danmark er herbicider (mod plantevækst), fungicider (mod svampevækst) og insekticider (mod insekter) de hyppigst anvendte. Nogle pesticider er i sig selv tilstrækkelig vandopløselige til at blive opløst og sprøjtet ud i vand, men oftest formuleres de som emulgerbare koncentrater eller som granulater (Walker et al., 1996). Salget af pesticider i Danmark er blevet reduceret fra 6169 tons i 1993 til 3554 tons i 2003, hvilket svarer til et fald på 42%. Faldet skyldes et forsøg på at minimere forekomsten af pesticider i grundvand og vandmiljø. Figur 1 viser fordelingen af salget af pesticider i Danmark i 2003. Det ses at herbicider udgør de mest solgte pesticider herhjemme med 2390 tons virksomt stof, svarende til 67 % af den samlede mængde solgte pesticider (Danmarks Statistik, 2004). Eksempler på hvordan pesticider kan tilføres søer og vandløb kan være: Drift fra udbringning på marker eller punktkilder som vaske eller spulepladser, hvor tømningen og rengøring af tanke og sprøjter foregår. Ligeledes udgør afløb fra dræn samt afstrømningen fra overflader under kraftige regnskyl kildet til pesticider i vandløb. Wiberg-Larsen et al. (1991) skønnede, på baggrund af Fyns Amts vandløbsforureningsrapporter for årene 1988-1990, at ca. 80 km vandløbsstrækning i Fyns Amt årligt forurenes med plantebeskyttelsesmidler. Generelt vil mindre vandløb have en mindre vandføring, og dermed være mere sårbare overfor forureninger end større vandløb, men billedet kan variere betydeligt i forskellige dele af landet.
Figur 1: Det samlede pesticidsalg (ton virksomt stof) i Danmark i 2003 fordelt over pesticidtype (Danmarks Statistik, 2004). I en oversigt over pulskoncentrationer af pesticider fundet i vandløb i Fyns, Århus og Nordjyllands Amt nævnes koncentrationer i vandløb på mellem 0,02 μg/l og 10,0μg/l (Styczen et al., 2003). Blandt de pesticider, der er testet i nærværende projekt, er stofferne dimethoat (0,7 μg/l), pirimicarb (3,0 μg/l), esfenvalerat (0,66 μg/l) og bromoxynil (10 μg/l) påvist og kvantificeret i de angivne maksimumskoncentrationer. Lauridsen & Wiggers (2001) undersøgte ligeledes koncentrationen af pesticider ved normal vandføring og under regnvejrshændelser og fandt koncentrationer mellem detektionsgrænsen på 0,01 μg/l og 5,1 μg/l. Desuden har Lauridsen et al. (2003) opstillet en liste over koncentrationen af en række pesticider i søer og vandhuller, baseret på undersøgelser foretaget af Århus-, Ringkøbing- og Ribe Amt hvoraf det fremgår at koncentrationen i søer ligger i intervallet 0,01-0,09μg/l og i vandhuller i intervallet 0,02-0,92μg/l. 2.2 Undersøgte pesticiderTabel 1 viser de pesticider, der er undersøgt i nærværende projekt, samt fysisk-kemiske parametre for disse. De undersøgte pesticider og referencestoffer er udvalgt i samråd med Miljøstyrelsen, bl.a. ud fra kriterier om anvendelsen i Danmark. I det følgende vil stoffernes virkemekanismer og brug kort blive gennemgået gruppevist, ligesom der også vil blive givet en vurdering af stoffernes skæbne i det anvendte testsystem (se afsnit 2.3). Det skal nævnes, at stoffet diflubenzuron (CAS nr. 35367-38-5) oprindelig var inkluderet i undersøgelserne, men at stoffet efter indledende forsøg udgik, da der ikke kunne observeres effekter på dafniernes mobilitet i koncentrationer op til stoffets vandopløselighed. Tabel 1: Anvendelse, udvalgte fysisk-kemiske egenskaber, EC50, 24h for immobilisering af Daphnia magna og NOEC21d-værdier for reproduktion af Daphnia magna for de undersøgte stoffer. Fysisk-kemiske data for dimethoat, pirimicarb, esfenvalerat, diquat, chlorpyrifos, pyrazophos, azoxystrobin og bromoxynil fra Tomlin 2003; Data for toksicitetstests med disse stoffer er fra Miljøstyrelsen, 2005. Data for 4-nitrophenol fra Verschueren, 1996; data for 3,5-dinitrophenol fra Toxnet, 2005 og fra US-EPA, 2004.
Kow: Oktanol-vand fordelingkoefficienten; S: Vandopløselighed; KH: Henry’s lov konstant (fordeling mellem vand- og luftfase). - : ingen data. 2.2.1 Organophosphater
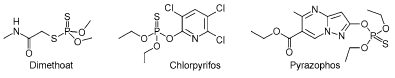
Figur 2: Kemisk struktur af organophosphaterne dimethoat, chlorpyrifos og pyrazophos (Tomlin, 2003). Dimethoat, chlorpyrifos og pyrazophos tilhører gruppen organophosphater, der hovedsageligt anvendes som insekticider (Tomlin, 2003). De fungerer som en acethylcholinesterasehæmmer. Acetylcholin er en neurotransmitter, der overfører impulser mellem neuronen og den motoriske endeplade (musklen). Normalt vil acetylcholin frigives til synapsen mellem to celler og efterfølgende bindes til acetylcholinreceptoren i den postsynaptiske membran. Dette får ionkanaler i cellemembranen, som er selektive for Na+ og K+ til at åbne sig og ionerne passerer dernæst den postsynaptiske membran, hvilket fører til et aktionspotentiale og aktivering af musklen. Virkningen af neurotransmitteren og dermed aktiveringen af musklen, ophører ved at acetylcholinesterase hydrolyserer transmitterstoffet. Er enzymet hæmmet vil denne deaktivering ikke ske (Eckert et al. 1996). Et tegn på forgiftning kan derfor ses som overstimulering af nerverne. Pyrazophos anvendes også som fungicid (Tomlin, 2003). I nogle svampearter virker pyrazophos hæmmende på melaninsyntesen (pigment) men pyrazophos kan også forhindre udviklingen af appressoria (parasitiske svampe) (Tomlin, 2003). Appressoria er specielle hyfer i svampen, der ligger sig langs overfladen af de fotosyntetiske celler i værtsorganismen, og derved muliggør, at svampen kan penetrere plantens celler (Raven et al., 1992, Lawrence, E., 1995). I 2003 blev der solgt 30629 kg dimethoat (93% heraf til landbrugsformål) og 529 kg chlorpyrifos. Dimethoat anvendes især i korn, men også i majs, roer, grøntsager, græs og kløver. Normaldoseringen for dimethoat er ca. 300 g aktivstof/ha (Miljøstyrelsen 2004). Det skal bemærkes, at pyrazophos ikke er godkendt til brug i Danmark, samt at chlorpyrifos ikke er godkendt til landbrugs/udendørs anvendelse. 2.2.2 Pyrethroider
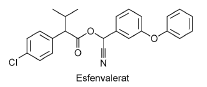
Figur 3: Kemisk struktur for esfenvalerat, et pyrethroidinsekticid (Tomlin, 2003). Esfenvalerat er et pyrethroidinsekticid (Tomlin, 2003), der anvendes i de fleste afgrøder, dog især korn. Pyrethroider er syntetiske udgaver af det naturlige insekticid pyrethrin, som findes i chrysantemum (Walker et al., 1996). Pyrethroiderne er neurotoxiner og virker ved at forlænge den tid Na+ kanalerne er åbne i nervecellerne. Normalt foregår åbningen og lukningen af kanalerne på under et millisekund- den tid en impuls passerer på. Bindingen af et pyrethroid bevirker således at hvilepotentialet ikke indtræffer og der genereres derfor adskillige aktionspotentialer frem for et enkelt. Resultatet er hyperaktivitet, krampe, lammelse og til sidst dør organismen (Connell, 1997; Stenersen, 2004). Esfenvalerat er optaget på Miljøstyrelsens nyeste liste over forbudte plantebeskyttelsesmidler ”Forbudslisten” i juli 2001, og må derfor ikke anvendes til udendørsbrug med undtagelse af anvendelse til dypning af rod og halssprøjtning af nåletræer (Miljøstyrelsen, 2004). Tidligere var doseringen ved udendørs anvendelse 5-20 g/ha. I 2003 blev der solgt 107 kg esfenvalerat (Miljøstyrelsen 2004). 2.2.3 Carbamater
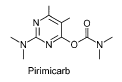
Figur 4: Kemisk struktur for pirimicarb, et carbamatinsekticid (Tomlin, 2003). Pirimicarb er et carbamatinsekticid (Tomlin, 2003). Carbamater er acetylcholinesterasehæmmere ligesom organophosphaterne. De bindes derfor også til det samme aktive site, men er forskellige ved, at carbamatgruppen bundet til acetylcholinesterase hydrolyseres meget hurtigere end organophosphatgruppen (carbon-oxygen-phosphor bindingen) (Stenersen, 2004). Hermed vil virkningen af carbamater aftage hurtigere end virkningen af organophosphater. I 2003 blev der solgt i alt 5183 kg pirimicarb. Hovedparten (4665 kg) blev solgt til landbrugsformål, og pirimicarb var det fjerde mest solgte insekticid næst efter dimethoat, cypermethrin og tau-fluvalinat. Pirimicarb finder især anvendelse på korn men også på afgrøder så som roer og ærter. Normaldosering er 125-250 g aktivstof/ha afhængig af afgrødetype (Miljøstyrelsen 2004). 2.2.4 Bipyridyl
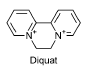
Figur 5: Kemisk struktur for diquat (Tomlin, 2003). Diquat er et herbicid, som virker ved at danne et superoxid under fotosyntesen som skader cellemembranen og cytoplamaet (Tomlin, 2003). I alt blev 12236 kg diquat til landbrugsformål i 2003 i Danmark. Ud af 45 solgte herbicider var diquat dibromid det 16. mest solgte i 2003. Herbicidet bruges hovedsageligt til nedvisning af kartofler, men også til græs og kløver. Normaldoseringen er 1496 g aktivstof/ha (i kartofler) (Miljøstyrelsen 2004). 2.2.5 Strobilurin
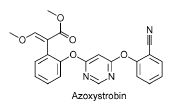
Figur 6: Kemisk struktur for azoxystrobin (Chemfinder, 2004). Azoxystrobin er et fungicid, der tilhører gruppen strubiluriner (Tomlin, 2003). Azoxystrobin virker ved at hæmme respirationsprocessen i mitochondrier. Mitochondrier er cellens ”energifabrik”, og pesticidet virker ved at hæmme elektrontransporten mellem cytochrome b og cytochrome c1. Når elektrontransporten hæmmes, mister cellen således sin evne til at danne energi og dør (Tomlin, 2003, Stryer, 1995). Azoxystrobin anvendes kun i korn og ærter og i 2003 udgjorde salget af azoxystrobin 35941 kg heraf 34941 kg til landbrugsformål. Fungicidet var således det fjerde mest solgte af 16 fungicider til landbruget. Normaldoseringen er 250 g aktivstof/ha (Miljøstyrelsen 2004). 2.2.6 Halogeneret benzonitril
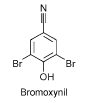
Figur 7: Kemisk struktur for phenoxyherbicidet bromoxynil (Chemfinder, 2004). Bromoxynil er et herbicid (Tomlin, 2003) og tilhører gruppen af halogenerede benzonitriler. Det hæmmer planters vækst ved at hæmme fotosyntesen. Hæmningen sker ved, at herbicidet bindes til fotosystem II og at elektrontransporten blokeres, hvilket stopper CO2- fikseringen samt produktionen af ATP og NADPH2 (Cornell University, 2004). I alt blev der solgt 64101 kg bromoxynil i 2003. Hele salget var til landbruget, hvor bromoxynil var det 6. mest solgte herbicid. Det anvendes til korn, græs og kløver og normaldoseringen er 400 g aktivstof/ha (Miljøstyrelsen, 2004). 2.2.7 Phenoler (referencestoffer)Som reference stoffer er valgt to phenoler (4-nitrophenol og 3,5-dichlorophenol), som begge er særdeles velundersøgte i dafnietest (US-EPA, 2004), og som jf. Tabel 1 har fysisk-kemiske egenskaber, der gør dem relativt nemme at håndtere i forsøgene (dvs. høj vandopløselighed, moderate Kow-værdier og lav flygtighed).
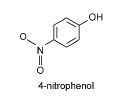
Figur 8: Kemisk struktur af referencestoffet 4-nitrophenol (Chemfinder, 2004). 4-nitrophenol anvendes som syntesekemikalie i forbindelse med fremstilling farmaceutiske stoffer, insekticider og farvestoffer (Rippen, 2004). Stoffet er velundersøgt i økotoksikologiske forsøg (US-EPA, 2004), og optræder bl.a. i miljøet som et nedbrydningsprodukt af organophosphaterne methyl- og ethylparathion.
Figur 9: Kemisk struktur af referencestoffet 3,5-dichlorophenol (Chemfinder, 2004). 3,5-dichlorophenol anvendes ikke til kommercielle formål, men er påvist som nedbrydningsprodukt af højere chlorphenoler, f.eks. pentachlorphenol (Toxnet, 2005). Stoffet anvendes bl.a. som referencestof i de standardiserede testmetoder med bakterier (ISO 1996) og alger (ISO, 1989b). 2.3 Teststoffernes skæbne i testsystemetVed de udførte test blev anvendt et testsystem bestående af 50-100 ml bægerglas fyldt med 25-50 ml vandigt medium. Testorganismerne var udsat for stofferne i 0,5-6 timer, hvorefter de blev overført til rent medium uden teststof. Under alle forsøg er anvendt nominelle stofkoncentrationer, da det af økonomiske årsager ikke har været muligt at foretage kemiske analyser af de aktuelle koncentrationer under og efter eksponering. I det følgende vil de mulige fjernelses- og fordelingsprocesser blive beskrevet (jf. Figur 10) m.h.t. til relevans for de testede stoffer under de anvendte eksponeringsforhold. Vurderingerne er lavet ud fra de fysisk-kemiske data vist i Tabel 1 og stoffernes kemiske strukturer som fremgår af afsnit 2.2.1.-2.2.7.
Figur 10: Mulige fjernelses- og fordelingsprocesser i det anvendte testsystem 2.3.1 FordelingsprocesserReduktion i teststofkoncentrationer som følge af fordampning anses for usandsynlig, når de lave værdier for Henry’s lov konstant (<10-7 atm×m³×mol-1) tages i betragtning. Chlorpyrifos har som eneste stof en Henry’s lov konstant over 10-7 atm×m³×mol-1 og vil derfor kunne have en langsom fordampningshastighed fra vandig opløsning. Fordampning vil dog også kunne udelukkes for chlorpyrifos ved de korte eksponeringstider (£ 6 timer), der anvendes i de udførte forsøg (statiske forsøg, der udføres under låg). Af de testede stoffer er det kun 3,5-dichlorophenol, pyrazophos, chlorpyrifos og esfenvalerat, som er tilstrækkeligt fedtopløselige til potentielt at kunne biokoncentreres (log Kow > 3). Således vil det også kun være for disse stoffer, at sorption til glasvægge vil kunne betyde en nedsættelse af vandfasekoncentrationen. Ligevægtsberegninger (fugacitetsmodel) fortaget på det aktuelle testsystem for samtlige stoffer viser, at der for chlorpyrifos og esfenvalerat kan være betydelig nedsat vandfasekoncentration som følge af sorption. Hvor store afvigelser, der vil være mellem de nominelle og de aktuelle koncentrationer er vanskeligt at afgøre ved teoretiske overvejelser, og kemiske målinger burde inkluderes til bestemmelse af eksponeringskoncentrationerne. Dette er imidlertid faldet uden for rammerne af nærværende projekt. Dog skal det holdes in mente, at eksponeringen af forsøgsdyrene i disse test er af få timers varighed (0,5-6t), og at dyrene ikke får tilført nogen føde som pesticidet evt. ville kunne absorberes af eller adsorberes til under eksponeringen. Begge disse forhold nedsætter risikoen for stoffjernelse ved sorption. 2.3.2 FjernelsesprocesserEksponeringer foregår i syntetiske medier under anvendelse af milliporevand, og hvor forholdene for mikrobiologisk vækst ikke er optimale. Set i lyset af de korte eksponeringstider udelukkes mikrobiel nedbrydning derfor som fjernelsesproces. Eksponeringerne er udført i mørke, og derfor udelukkes også fotokemiske omdannelse som fjernelselsesproces. Flere af de testede pesticider (organophosphaterne, esfenvalerat, pirimicarb og azoxystrobin) vil, ud fra en vurdering af deres kemiske struktur, kunne undergå en hydrolytisk spaltning ved kontakt med vand. Organophophaterne (dimethoat, chlorpyrifos og pyrazophos) vurderes at være de stoffer, for hvilke hydrolysen er af størst betydning, og der vil teoretisk set kunne ske en nedsættelse af toksiciteten over tid. I det anvendte pH interval (pH 7,0-8,5) er målt halveringstider for hydrolyse på 4-118 dage for dimethoat og 22-35 dage for chlorpyrifos (Toxnet, 2005) og halveringstider i samme størrelsesorden må forventes for pyrazophos. Set i forhold til de aktuelle eksponeringstider (£ 6 timer) vurderes det at være rimeligt, at der ses bort fra hydrolyse som fjernelsesproces i de udførte pulsforsøg. 2.4 Daphnia magna som testorganismeSom forsøgsorganisme benyttes Daphnia magna, der er en hyppig brugt testorganisme i økotoksikologiske forsøg. Dermed er den og dens respons overfor kemiske påvirkning vel beskrevet i litteraturen og sammenligninger er derfor mulige. Dafnier forekommer endvidere på mange lokaliteter, og er ligesom mange andre invertebrater mere stedfaste end fisk, der flytter sig meget og forlader et område hvis der opstår en kritisk periode (Miljøstyrelsen, 1998). Arten er økologisk relevant, da dafnierne lever af at græsse planktonalger og detritus og de er derfor vigtige for stofomsætningen der hvor de lever (søer og damme) (Jespersen & Lützen, 1996). Dafnier formerer sig ved parthenogenese (ukønnet formering), og producerer således kloner af sig selv under gunstige forhold (Jespersen & Lützen, 1996). Herved undgås variationer i forsøgsresultater på grund af genetiske forhold, testene bliver mere reproducerbare og der er mulighed for hurtig opformering. Ved pulseksponering forventes et forsinket toksisk respons, og immobilitet, som er det traditionelt anvendte endpoint i dafnietest, vil være mere anvendeligt end f.eks. dødelighed. Ved anvendelse af Daphnia magna kan såvel sub-letale (immobilisering) som kroniske (reproduktion) endpoints undersøges med samme organisme, og da der findes internationalt standardiserede testprocedurer for begge endpoints muliggøres sammenligninger med allerede eksisterende data. Endelig skal som rent praktiske hensyn nævnes, at dafnier ikke er vanskelige at håndtere, og at selv unge dyr har en god størrelse - de kan ses med det blotte øje. 2.5 Pulseksponeringsforsøg beskrevet i litteraturenMed henblik på at afklare eventuelle overlap mellem den foreslåede metode og eksisterende metoder til påvisning af pulseffekter af pesticider på krebsdyr er gennemført et litteraturstudie, som vil blive beskrevet i det følgende. 2.5.1 Akutte effekterDen anvendte metode er udviklet med udgangspunkt i PE-LET50-metoden beskrevet af Brent & Herricks (1999), hvor dyrene (Daphnia magna) observeres i en post-eksponeringsperiode efter en kort pulslignende eksponering. Der er anvendt de samme præliminære tests (OECD/ISO test), eksponeringsscenarier og post-exposure observationsperioder, men hvor Brent & Herricks (1999) udelukkende fokuserer på akutte effekter (immobilsering), er metoden udvidet til også at omfatte kroniske effekter (reproduktion). I artiklen af Brent & Herricks (1999) er fokus rettet mod regnbetingede udledninger fra byområder, og udover en beskrivelse af test med referencestofferne zink og cadmium belyses effekten af enkeltstoffer ikke. I et andet studie udført af Brent & Herricks (1998) er effekterne af en kortvarig eksponering med zink, cadmium og phenol beskrevet for invertebraterne Ceriodaphnia dubia og Hyalella azteca. I dette studie blev også anvendt posteksponerings-observationer, men igen var fokus på immobilisering og ikke på længerevarende effekter. Som det fremgår er phenol det eneste organiske stof for hvilket, der er publiceret resultater fra posteksponeringsstudierne udført af Brent & Herricks (1998, 1999), og effekten af pesticider er således ikke beskrevet tidligere. 2.5.2 Forsinkede effekterForsinkede effekter af pulseksponeringer af andre organismer og ved andre testmetoder er beskrevet i litteraturen. Wright (1976) så således, at graden af toksicitet kan ændres, hvis der inkluderes en observationsperiode efter eksponering, og stoffer med en høj initiel toksicitet virkede således mindre toksiske, når der blev inkluderet en observationsperiode. For phenol fandt Brent & Herricks (1998), at C. dubia genvandt mobilititeten efter den kortvarige eksponering var ophørt. For myggelaver udsat for permethrin, fenitrothion, carbaryl og carbofuran er et ”recovery” efter kortvarig eksponering således beskrevet af Parsons & Surgeoner (1991), mens der for krebsdyr og fisk udsat for kortvarige eksponeringer af lindan (Abel 1980), permethrin (Abel & Gardener, 1986), chlorpyrifos, endrin, fenvalerat (Jarvinen et al., 1988) og esfenvalerat (Holdway et al, 1993) er fundet forøget mortalitet efter posteksponerings-observationer. 2.5.3 Kroniske effekterBlandt de få studier, der beskæftiger sig med kroniske eller sub-kroniske effekter af pesticider ved pulseksponering af invertebrater, skal nævnes undersøgelser udført af Schulz & Liess (2000) med vårfluelarver udsat for fenvalerat i 1 time. Dyrene blev efter eksponering i laboratoriet overført til udendørs mikrokosmer med strømmende vand, og observationsperioden strakte sig over 240 d. Der blev fundet effekter helt ned til 0,001 mg/l i vandfasen, men selvom mulighederne for ekstrapolation af testresultaterne er gode ved denne metode vurderes metoden dog at være for tids- og arbejdskrævende (der kigges på 3 forskellige endpoints og effekten af sedimenttilsætning), hvis effekter af en række pesticider skal undersøges. Med hensyn til langtidseffekter på dafnier efter pulseksponering med pesticider skal undersøgelserne foretaget af Hosmer et al. (1998), Naddy et al. (2000) og Naddy & Klaine (2001) fremhæves. I disse studier anvendes Daphnia magna til undersøgelse af effekten af pulseksponeringer med pesticider (fenoxycarb og chlorpyrifos). Hosmer et al. (1998) undersøgte effekten af fenoxycarb hos Daphnia magna i en simuleret pulseksponering, hvor stoffets halveringstid blev anvendt til dimensionering af pulsen i et flow-through testsystem. Der blev udført reproduktionsstudier med de eksponerede dafnier, men dette studie anvender ikke egentlige posteksponeringsforsøg. Organismer blev således ikke overført til rent medie efter eksponering, men der skete en gradvis reduktion af fenoxycarb koncentrationen over de 21 dage testen varede. Det skal bemærkes, at Hosmer et al. (1998) fremhæver den udtalte datamangel for kroniske effekter efter pulseksponering, samt at deres undersøgelser viste markant lavere toksicitet af fenoxycarb, når pulseksponeringsforsøgene blev sammenlignet med reproduktionstest udført med kontinuert eksponering. Naddy et al. (2000) gennemførte undersøgelser, hvor Daphnia magna blev udsat for små koncentrationer af chlorpyrifos (<1 mg/l) i pulseksponeringer med varigheder fra 1-72 timer. Såvel akutte som kroniske effekter af chlorpyrifos blev inkluderet. I undersøgelsen blev kun akutte effekter fundet, og der blev ikke observeret forsinkede effekter af chlorpyrifos, og altså heller ikke dokumenteret kroniske effekter af pesticidet. Naddy et al. (2000) fandt ingen effekter af chlorpyrifos ved pulseksponeringer på <12 timers varighed. I artiklen af Naddy & Klaine (2001) fokuseres på effekten af gentagne pulseksponeringer af chlorpyrifos. Der anvendes grundlæggende samme metode, som af Naddy et al. (2000), men der anvendes dafnier, som er 7 dage gamle ved eksponeringen, og kun akutte effekter er beskrevet. Det er nemlig væsentligt at bemærke, at Naddy et al. (2000) fandt markante forskelle i testorganismernes respons afhængigt af dyrenes alder på eksponeringstidspunktet, således var 3-, 7- og 14-dage gamle dyr mere følsomme end de nyfødte unger, når overlevelse blev anvendt som endpoint i en 21-dages test. Det skal dog bemærkes, at disse konklusioner udelukkende bygger på, at der var en større effekt på overlevelsen efter en gentagen puls, som desuden var af højere koncentration end den indledende. 3 Materialer og metoder
I det følgende beskrives de materialer og metoder, der er anvendt i projektet. Som en del af projektet er udarbejdet en egentlig testprotokol, som har en større detaljeringsgrad end det nedenfor beskrevne. Testprotokollen findes i Bilag 1. 3.1 Kultivering af testorganismer og algeSom forsøgsorganisme benyttes Daphnia magna, indsamlet i Langedammen ved Birkerød i 1978 og dyrket i laboratoriet siden. Dafnierne holdes i akvarier med kunstigt fremstillet ferskvand (M7-medie, se Bilag 1), der fremstilles i laboratoriet. Dafniestamkulturen dyrkes i klimarum ved en temperatur på 20°C ± 2°C og en 12:12 timers lys/mørke periode (4-6 μE·m-2·s-1). Kulturerne holdes i 4,2 liters akvarieglas eller bægerglas med et volumen på 800 ml. Akvarierne fyldes med 1,6 l M7-medie og pyrexglassene fyldes med 800 ml M7 medie, og der tilsættes maks. 30 hunner pr. akvarie og maks. 20 til pyrexglassene. Akvarieglassene overdækkes med plastiklåg, så der ikke kan komme støv og lignede ned i glassene. Tilsættes der flere hunner, kan konkurrencen mellem dyrene blive for stor, og produktionen af juvenile minimeret. Dyrene får rent medie 2 gange ugentligt med 3-4 dages mellemrum. Det nye medie består af 50% nyfremstillet M7-medie og 50% af det M7-medie, dyrene har opholdt sig i i den forgange periode. Moderdyrene overføres til det nye medie med et nylonfilter. Der laves løbende nye dafniekulturer så produktionen af juvenile holdes på et optimalt niveau (ca. 10-20 unger pr. moderdyr pr. 2-3 dag). Moderdyrene udskiftes når de er ca. 8 uger gamle. Dafnierne får føde i form af grønalgen Pseudokirchneriella subcapitata (tidligere kaldet Selenastrum capricornutum). Algekulturen stammer fra kulturindsamlingen fra Norsk Institut for Vandundersøgelser (NIVA Oslo, Norge). Algekulturen dyrkes til en tæthed på 2-4×106 celler/ml og tilføres dafnierne 3 gange dagligt v.h.a. et pumpesystem. Selve algekulturen befinder sig på en magnetomrører, der sørger for, at algerne forbliver i en homogen suspension. Mængden af føde, der tilføres svarer til 0,1-0,2 mg C /dyr/dag (C = carbon). Hvis det vurderes, at være nødvendigt, f.eks. hvis der ikke produceres tilstrækkeligt med unger, kan der tilsættes en smule tørgær, som ekstra foder. Tørgæret opløses i en smule medie inden det kommes i akvariet. Fremstilling af foder fremgår af Bilag 1. Algekulturen holdes i pyrexflasker ved en temperatur på 4°C, indtil den skal anvendes til foder. Flasken skal rystes før brug, da algen bundfælder under opbevaringen. Dafnier (Daphnia magna), der anvendes i forsøgene, er < 24 timer gamle. Dette sikres ved at fjerne alle unger fra moderdyrene senest 24 timer inden forsøgene sættes i gang (jf. ISO 6341, 1989a). Efter 24 timer vil alle unger i glas med moderdyr således være <24 timer gamle, og disse indfanges og anvendes i de videre forsøg. 3.2 Anvendte kemikalier og testopløsningerDe anvendte referencestoffer og pesticider var alle af analysekvalitet (renhed 98-99,9%) og blev indkøbt fra Sigma-Aldrich A/S, København. Alle øvrige anvendte kemikalier til medier m.v. var ligeledes af analysekvalitet og blev indkøbt fra Merck, København. Gennem hele studiet blev anvendt destilleret vand, som blev ledt gennem et milliporeanlæg inden anvendelse. Stamopløsninger blev fremstillet ved at opløse den ønskede mængde af pesticid direkte i M7-medie. I de tilfælde, hvor et pesticid ikke kunne opløses direkte i M7, blev opløsningsmidlet dimethylsulfoxid (DMSO) anvendt som ”carrier”. En stamopløsning bestående af 10 ml DMSO tilsat pesticid blev fremstillet og ud fra denne blev en endelig stamopløsning fremstillet i M7-medie. Denne stamopløsning indeholdt maksimalt 0,1‰ (v/v) DMSO. 3.3 ValiditetskriterierDer blev målt ilt (Orion elektrode) og pH (PHM 210 standard pH-meter) i testopløsningerne (højeste koncentration) og i kontrollen under alle forsøg. For de kroniske tests blev der desuden foretaget målinger ved hvert medieskift. I følge ISO (1989a) kræves en iltmængde på = 2 mg/l og en pH på 7,8 ± 0,2 under akuttestene. Kravet i OECD guidelinen for kroniske test er en iltkoncentration på 3 mg/l og en pH mellem 6-9, der desuden ikke varierer mere end 1,5 enheder (OECD, 1998). I alle test blev ilt-kravet overholdt (se Bilag 2), idet der blev gennemført test med 7,2-9,8 mg/l ilt. pH-værdien i de udførte test var 7,2-8,9 og varierede ikke mere end 1,5 pH-enhed i hver test. I overensstemmelse med ISO standarden (ISO, 1989a) blev kaliumdikromat (K2Cr2O7) anvendt som referencestof gennem hele projektperioden. I alle tilfælde holdt de funde IC50-værdier sig inden for det af ISO beskrevne interval (IC50, 24 t = 1,47 ± 0,57). 3.4 Akuttest med kontinuert eksponering (ISO test)Formålet med de udførte ISO-test er at bestemme effektkoncentrationer (EC-værdier) for 24 timer og 48 timer ved observation af ekponerede dyrs mobilitet. Disse værdier skal dernæst anvendes til design af pulseksponeringstest. Alle akutte tests blev udført i henhold til ISO-standarden for dafnietest (ISO, 1989a). For at finde det koncentrationsinterval, hvor et respons fra dyrene på de undersøgte pesticider kan forventes, blev der udført ”range finding test” for alle stoffer. Under disse test blev brugt én kontrolgruppe á fem individer og for hver testkoncentration blev der eksponeret én gruppe af fem individer. Ved de definitive test blev anvendt 4 replikater pr. koncentration og 6 replikater i kontrolgruppen. De ekstra replikater i kontrolgrupperne er tilføjet udelukkende med den hensigt at styrke data, da samtlige resultater relateres til kontrolgruppernes respons. Hver replikat bestod af 25 ml testopløsning (M7 tilsat teststof/pesticid) og 5 juvenile Daphnia magna (<24 t). Testene forløb i mørke og dyrene blev eksponeret i 48 timer. Der blev ikke skiftet opløsning i løbet af de 48 timer. Testkoncentrationer fremgår af Tabel 2.Under eksponeringen blev dyrene ikke fodret. I forhold til ISO-testen er antallet af observationstider øget, således at dyrenes mobilitet og dødelighed observeres efter 0, 2, 4, 6, 24 og 48 timers eksponering. Registreringen foregik ved observationer på lysbord, og om nødvendigt v.h.a stereolup. Nedenstående definitioner af immobilitet og dødelighed er anvendt ved alle udførte forsøg, d.v.s. såvel akuttest som pulseksponeringsforsøg: Immobilitet: Dafnier, som ikke kan opretholde deres position i vandsøjlen (svømme) selv efter en forsigtig stimulering (15 sek.), regnes for immobile. Dafnierne kan have krampeagtige bevægelser. Døde dafnier vil indgå i det samlede antal immobile dafnier, men der skelnes mellem døde og immobile dyr ved observationerne. Dødelighed: Dafnier, der ikke har nogen form for bevægelse, som f.eks. bevægelse af antenner, selv 15 sekunder efter stimuli, regnes for døde (Naddy et al., 2000, OECD, 1998). I praksis blev dette overført til, at mobile dyr skal svømme i løbet af de 15 sekunder de observeres men ikke nødvendigvis i alle de 15 sekunder. Dette skyldes, at der ofte blev observeret dafnier, der ikke svømmer i en kort tidsperiode (opholder sig typisk i bunden af glassene) for så pludselig at svømme livligt rundt. Tabel 2: Koncentrationer benyttet under akuttest med Daphnia magna. Ved alle test er inkluderet en kontrolgruppe, der ikke eksponeres for teststof.
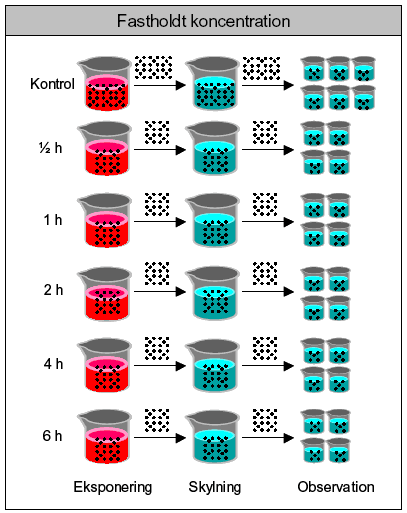
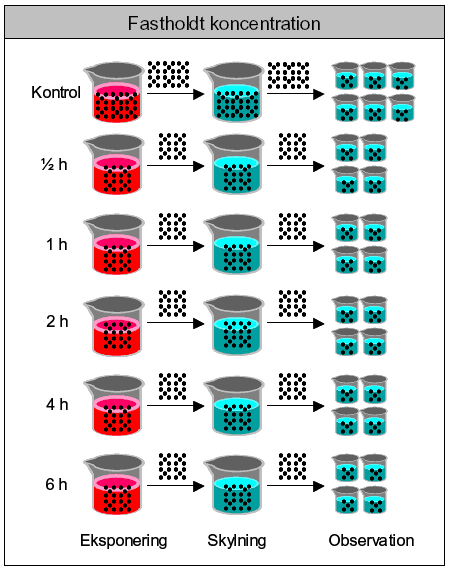
*DMSO anvendt som bærestof (max. 0,1‰ (v/v)). Ved disse test er yderligere inkluderet kontrolgruppe med 0,1 ‰ (v/v) DMSO. 3.5 Pulseksponering3.5.1 Valg af stofkoncentrationerDe anvendte eksponeringskoncentrationer i pulstestene blev udvalgt på baggrund af de indledende akutte tests. Værdierne blev valgt med udgangspunkt i de beregnede IC90, 24t-værdier, men der blev ligeledes taget hensyn til dyrenes tilstand efter 6 timer under akuttestene. I de tilfælde, hvor der kunne observeres en respons, blev der set på om det var i form af dødelighed eller immobilitet. Koncentrationer, hvor immobilisering indtrådte, blev foretrukket, da muligheden for at observere et recovery ellers ikke ville være til stede. Disse kriterier for udvælgelse af koncentrationer medførte i flere tilfælde, at kun et ganske snævert koncentrationsinterval blev aktuelt at undersøge i pulstestene. 3.5.2 Procedure for pulseksponering – korttidseffekterFremgangsmåden for pulseksponeringen er vist i Figur 11. Dyrene blev eksponeret i grupper af 20 individer (30 i kontrolgruppen) og efter endt eksponering blev de indfanget og overført med engangspipette til et skyllekar. Dyrene blev indfanget i skyllekarret v.h.a. et fintmasket net, og dyrene blev delt i 4 grupper pr. eksponeringstid og 6 grupper for kontrolgruppen. De ekstra replikater i kontrolgrupperne er også her tilføjet med henblik på at styrke data, da samtlige resultater relateres til kontrolgruppernes respons. Hver replikat bestod således af 5 juvenile Daphnia magna (<24 t). Kontrolgruppen (30 dyr) blev behandlet på tilsvarende måde som gruppen, der fik en puls af 0,5 time, da det blev antaget, at der vil være størst stress, når dyrene håndteres gentagende gange med korte tidsintervaller (dvs. ”worst case” for kontrollerne). Der blev ikke inkluderet en DMSO kontrol under pulsforsøgene, da der ikke blev registreret en effekt (døde/immobile) i DMSO kontrollerne under de akutte tests. Antallet af immobile og døde dyr blev optalt 0 timer, 24 timer og 48 timer efter pulsens ophør. I posteksponeringsperioden fik dyrene føde i form af algen Pseudokirchneriella subcapitata. Fødemængden svarede til 0,1-0,2 mg C/dag, hvilket er den anbefalede mængde føde i ISO (1989a) og blev tilført til tiden t = 0 timer (pulsophør).
Figur 11: Fremgangsmåde for pulseksponering af dafnier. For hver testet koncentration inddeles dafnierne i grupper afhængig af eksponeringstid. Efter endt eksponering overføres alle dyr til et kar med rent M7 medie (skylning) og dernæst deles de 20 dyr pr. eksponeringstid i 4 replikater á 5 dyr og overføres til glas med rent M7-medie tilsat alge (observation). 3.5.3 Fodring under pulstests.Der blev udført forsøg til bestemmelse af fødens invirkning på mobiliteten hos Daphnia magna, der havde været udsat for en puls af et pesticid (pirimicarb 100 μg/l og dimethoat 30,0 mg/l). Forsøget blev udført som de øvrige pulseksponeringsforsøg, d.v.s samme eksponeringmåde og samme foder og fodermængde (Pseudokirchneriella subcapitata: 0,1-0,2 mg C/dafnie/dag), med undtagelse af at én gruppe i dette forsøg ikke blev fodret efter endt pulseksponering. Den %-vise ændring i mobiliteten (immobile/døde) kunne således blive registreret, sammenlignet for grupperne og relateret til fodringsstatus. Under forsøget med dimethoat blev dyrene eksponeret i en fælles beholder og derefter tilfældigt indsamlet og fordelt i observationsglassene. Under forsøget med pirimicarb blev dyrene delt i to grupper allerede inden eksponeringen for pesticidet. 3.5.4 Gentagne pulse og pulseksponering af 3 dage gamle dafnierPrincippet i dette forsøg svarede til proceduren beskrevet for pulseksponering. Den eneste forskel var, at dyrene i de gentagne pulstest blev eksponeret for to pulse. Efter observationsperiodens ophør, d.v.s. 48 timer efter den første puls, blev dyrene således udsat for endnu en puls af samme koncentration og varighed som den første puls. Forsøgene blev udført med stofferne pyrazophos (2,50 μg/l og 5,00 μg/l), dimethoat (10,0 mg/l og 20,0 mg/l), pirimicarb (40,0μg/l og 70,0μg/l) samt esfenvalerat (10,0 μg/l). I forbindelse med disse forsøg med gentagne pulse, blev der også udført et forsøg med 3 dage gamle dafnier, der fik en enkelt pulseksponering, for at se om der er en aldersafhængig forskel i responset. Forsøgsprincippet er som beskrevet for pulseksponsering af 24 timer gamle dafnier. 3.5.5 Alder og følsomhedISO standarden (ISO 1989a), der benyttes til bestemmelsen af immobiliteten hos Daphnia magna efter kontinuert eksponering for et givent stof, benytter organismer, der er under end et døgn gamle (<24t). Imidlertid er forskellige aldersgruppers varierende følsomhed overfor pulse af pesticider blevet undersøgt og beskrevet i litteraturen (Naddy et al., 2000). På baggrund heraf blev der igangsat en indledende undersøgelse til bestemmelse af følsomheden hos 4 forskellige aldersgrupper hos dafnier: <24t, 3d, 5d og 7d. Dyrene blev pulseksponeret på samme måde som beskrevet i afsnit 3.5.2. 3.5.6 Procedure for pulseksponering – langtidseffekterPulseksponering af <24 t gamle dafnier foregik som beskrevet ovenfor, dog anvendes kun 10 dyr pr. eksponeringstid og i kontrolgruppen. Gruppen af dyr, der skal have en puls af 6 timers varighed, eksponeres som den første og 2 timer senere eksponeres gruppen, der skal have en puls af 4 timers varighed o.s.v. indtil alle grupper er blevet eksponeret. Ved overførsel fra skyllekarret fordeles dyrene i 10 glas pr. eksponeringstid (eller kontrol), d.v.s. at dyrene går enkeltvis i hver deres 50 ml bægerglas i hele posteksponeringsperioden. Dyrene i kontrolgrupperne eksponeres for rent M7-medie på samme måde som dyrene, der eksponeres for et kemikalie. D.v.s. de bliver også overført til et nyt medie og ”skyllet”. På denne måde sikres det, at variationer ikke skyldes forskelle i håndteringen af organismerne (Naddy & Klaine, 2001). Reproduktionen hos Daphnia magna følges efter en pulseksponering (0,5t, 1t; 2t; 4t; og 6t) og 21 dage frem. I posteksponeringsperioden frem til de 21 dage får dyrene føde tilsat i form af algekulturen Selenastrum capricornutum. Mængden af føde svarer til ca 0,1 mg C/dag den første uge og ca. 0,2 mg C/dag de sidste to uger og tilsættes hver anden dag. Følgende parametre blev registreret:
Dagen, hvor de første unger observeres noteres, og juvenile fjernes derefter hver anden dag v.h.a. en pipette. Antallet registreres ved hver oprensning. Moderdyrenes længde på dag 21 er opmålt vha. af et kamera (Leica M26) tilsluttet et mikroskop (Olympus BH-2type). Der blev taget et billede af hver dafnie efter testens afslutning og dafnierne blev opmålt v.h.a. programmet AxioVision LE Rel. 4.1. Vægten af dyrene blev bestemt ved at tage alle de overlevende dyr i en gruppe og veje dem samlet. Filterpapir blev tørret i alubakker ved 105 ºC i en time og overført til nedkøling i en eksikator med silikat inden de blev vejet. Dernæst blev de overlevende dafnier i én gruppe overført til et stykke tørret filterpapir, tørret ved 105 ºC i 1 døgn overført til eksikator og vejet (Dansk Standardiseringsråd, 1980). Dafnierne blev vejet som en samlet gruppe for at være sikker på, at der var dyr nok til, at vejningen kunne gennemføres med tilstrækkelig nøjagtighed. Vægten af dafnierne i en gruppe kunne herved bestemmes som differencen mellem papirets vægt og vægten af dafnier og papir. Ved opstilling af langtidstestene samt ved vurdering af resultaterne herfra er anbefalingerne i OECD’s standard reproduktionstest med Daphnia magna (OECD, 1998) blevet fulgt, d.v.s. de udførte test har overholdt krav om max. 20 % døde moderdyr blandt kontroldyr, krav om min. 60 unger pr. moderdyr i kontrolgruppen, en pH-værdi mellem 6 og 9 (som ikke varierer mere end 1,5 enheder) og et O2-niveau over 3 mg/l. Desuden er der lavet visse modifikationer, da standard reproduktionstesten reelt er designet som en forlænget akuttest, hvor dyrene går i en fast stofkoncentration i modsætning til pulstesten, hvor dyrene eksponeres en given tidsperiode, hvorefter de igen kommer over i rent medie. I OECD guidelines anbefales det, at mediet med kemikalie skiftes tre gange ugentligt for at sikre, at niveauet af kemikalie holdes på et fast niveau. Da dyrene i pulstesten for langtidseffekter kommer over i et rent medie uden kemikalie efter få timer er dette ikke nødvendigt, og mediet skiftes derfor to gange ugentligt på samme måde som stamkulturen med moderdyr. I OECD (1998) er der krav om mindst 10 testdyr pr. koncentration. Dette er overført til 10 testdyr pr. eksponeringstid ved fastholdt koncentration. 3.6 DatabehandlingFor alle udførte forsøg gælder, at det samlede antal forsøgsorganismer i samme kontrolgruppe eller i samme eksponeringsgruppe (koncentration (ISO-test) eller eksponeringstid (pulstest)) blev slået sammen til én gruppe ved databehandlingen (f.eks. betragtes 4 replikater á 5 dyr udsat for samme eksponering ikke som enkelt-replikater, men som én gruppe på 20 dyr). Hvis der var medtaget to forskellige kontrolgrupper, i tilfælde hvor opløsningsmidlet DMSO blev benyttet, blev disse to grupper ikke slået sammen men behandlet hver for sig. Koncentrations-respons kurver for ISO-test estimeres ved hjælp af programmet ToxCalc ver. 5.0. Her udføres Shapiro-Wilk’s test for normalfordeling, test for varianshomogenitet og endelig maximum-likelihood estimation af koncentrations-respons kurven v.h.a. logit-modellen. Ud fra dosis-respons kurven beregner programmet ICx-værdier (immobiliseringskoncentrationer med ”x” fra 1-99% effekt) med tilhørende 95%’s konfidensintervaller. Resultater for ISO-test med kontinuert eksponering og udvidede observationsintervaller er desuden analyseret med programmet DEBtox ver. 1.1 udviklet af Kooijman & Bedaux (1996). Programmet er designet til at analysere data fra standardiserede toksicitetstest med hyppige observationer. Modellen bag programmet bygger på teorien om ”dynamic energy budget” (DEB), og kort fortalt er det med DEBtox muligt at analysere de tredimensionelle effektflader, der fremkommer, når antallet af immobile dyr afbildes mod eksponeringstid og teststofkoncentration (for en fyldestgørende gennemgang af DEB henvises til Kooijman & Bedaux, 1996). I nærværende sammenhæng er DEBtox anvendt til beregning af NEC-værdier (no effect concentration). Fordelen ved at anvende NEC (fremfor f.eks. NOEC eller ICx-værdier) er, at denne parameter er uafhængig af eksponeringstiden (Kooijman & Bedaux, 1996). Under pulstestene registreres det samlede antal immobile (immobile og døde) til tiderne t = 0, 24 og 48 timer efter eksponeringens ophør. Ud fra registreringerne bestemmes den (ikke-lineære) kurve, der beskriver dyrenes respons som funktion af eksponeringstiden. Denne eksponeringstids-respons kurve estimeres ved brug af logit funktionen i programmet ToxCalc ver. 5.0, som beskrevet ovenfor. Herudfra kan immobiliseringstider (ITx-værdier) beregnes, hvor det som oftest er de eksponeringstider, som giver 50%’s effekt (IT50), der anvendes om endpoint for testen. Da der er tale om posteksponeringsforsøg, hvor tiden (t) fra eksponeringens ophør er vigtigt for fortolkningen af resultatet anvendes betegnelsen PEt-IT50 som resultat af testene i overensstemmelse med anbefalinger af Brent & Herricks (1999). PEt-IT50 udtrykker pulslængden, der hæmmer mobiliteten af 50% af forsøgdyrene t timer efter eksponeringens ophør. PEt-IT50 er således udtrykt i timer, og da der i de udførte forsøg er foretaget posteksponeringsobservationer 0, 24 og 48 timer efter eksponeringens ophør vil PE0t-IT50, PE24t-IT50 og PE48t-IT50 blive anvendt i det følgende. Sker der et fald i de beregnede PEt-IT50 med tiden efter eksponering (dvs. PE0t-IT50>PE24t-IT50>PE48t-IT50) beskrives dette i det følgende som en forsinket negativ effekt, idet dyrenes tilstand forværres i posteksponeringsperioden. Observeres derimod en stigning i de beregnede PEt-IT50 med tiden efter eksponering (dvs. PE0t-IT50<PE24t-IT50<PE48t-IT50) betegnes dette som en restitution eller recovery, idet dyrenes mobilitet forbedres i tiden efter eksponeringens ophør. Til vurdering af data opnået ved test for kroniske effekter af pulseksponering blev programmet S-Plus 2000 anvendt til beskrivelse af en mixed-effect model (også kaldet ”repeated measures” analyser). Denne model anvendes ud fra den betragtning, at observationerne i datasættet ikke er uafhængige, da det er sandsynligt, at der er en sammenhæng mellem antal unger produceret efter 14 dage og antal unger produceret efter 21 dage af det samme moderdyr. Ved den anvendte model undersøges om, der er signifikante forskelle i antallet af afkom pr. moderdyr (Yij) som funktion af eksponeringstid og dyrenes kropslængde. Den anvendte model kan kort beskrives som:
- hvor β1 og β2 angiver en evt. effekt af hhv. eksponeringstid og dyrets længde, mens γj angiver dyr nr. j. Endelig angiver εij afvigelsen mellem model og observation. 4 Resultater
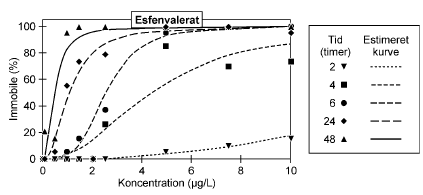
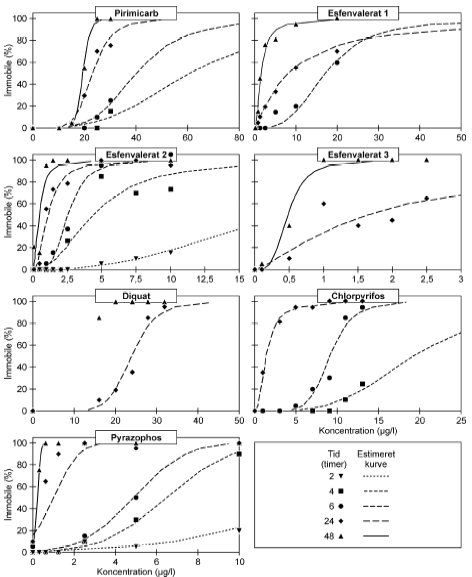
4.1 Akuttest med kontinuert eksponering (ISO test)I Figur 12 er vist et eksempel på de koncentration-responskurver, der er opnået ved akuttest med kontinuert eksponering. Der er tale om en standard ISO-test, men med udvidede observationer, således at dyrenes mobilitet blev registreret efter 2, 4, 6, 24 og 48 timers eksponering. Samtlige koncentrations-responskurver er vist i Bilag 3. Som forventet blev der observeret en klar forøgelse af antallet af immobile dyr jo længere tid dyrene var udsat for stofferne. For esfenvalerat sås effekter allerede efter 2 timer eksponering (Fig. 12), men som det fremgår af Bilag 3 var dette ikke tilfældet for alle de testede pesticider.
Figur 12: Koncentration-responskurve for immobilisering af Daphnia magna udsat for esfenvalerat i ISO test med kontinuert eksponering. Under akuttesten, blev der observeret forskelle i bevægelsesadfærden mellem de mobile dyr i de eksponerede grupper og de mobile dyr i kontrollen. Dyrene, der blev eksponeret, var langsommere i deres bevægelser eller svømmede mere rykvist end kontrolgruppens dyr. Under testen med esfenvalerat blev det således observeret, at eksponerede dyr bevægede sig langsommere, og jo højere koncentrationen blev, jo mere krampeagtige blev dyrenes bevægelser. Kontrolgruppens dyr var i stand til at svømme unden stimulering, hvorimod nogle af de eksponerede gruppper først svømmede efter en blid stimulering I Tabel 3 er vist de estimerede immobiliseringskoncentrationer (IC-værdier) for alle stoffer efter henholdsvis 24 og 48 timers eksponering. I alle tilfælde ses det, at koncentrationen, der skal til for at immobilisere henholdsvis 10, 50 og 90% af organismerne, falder fra tiden 24 timer og til tiden 48 timer. Disse værdier samt observerede kurveforløb under de første 6 timers eksponering dannede grundlag for udvælgelse af testkoncentrationer i pulstestene. I Tabel 3 er også vist de estimerede værdier for nul-effekt koncentrationen (NEC), og det er værd at bemærke, at der for alle stoffer ses en god overensstemmelse mellem NEC og IC10, 48t. Tabel 3: Estimerede immobiliseringskoncentrationer (IC) med tilhørende 95% konfidensintervaller efter 24 timers og 48 timers kontinuert eksponering. Nul-effekt koncentrationer (NEC) er estimeret ud fra DEBtox ver. 1.1 (Kooijman & Bedaux, 1996)
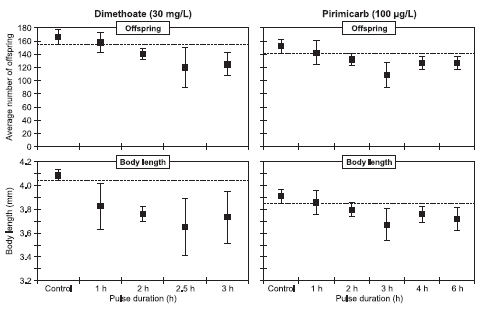
a) 100% hæmning observeret for alle koncentrationer. Angivne værdi er den lavest testede koncentration. b) Langsom kinetik; Enhed: mg·h/l c) Ikke muligt at beregne vha. DEBtox 4.2 Betydningen af fodring i posteksponeringsperiodenTidligere undersøgelser har vist, at fodring af dafnier både kan reducere toksisiteten signifikant (Buhl et al., 1993), men også øge den (Naddy & Klaine, 2001). I ISO-standarden (1989a) udføres akut-testene uden tilsætning af føde til mediet i de 48 timer dyrene eksponeres. Begrundelsen er, at tilsætning af føde (alger) vil kunne reducere mængden af stof dyrene reelt eksponeres for. Da dyrene under pulstesten kommer over i rent vand er der dog ikke risiko for en reduktion i stofkoncentrationen p.g.a. adsorption til føden. I dafniernes naturlige miljø vil der desuden være føde til stede. For at undersøge om dafniernes fodertilstand har en betydning for dyrenes recovery, blev der udført to forsøg. Ét med pirimicarb (100 μg/l) og ét med dimethoat (30 mg/l). Resultaterne for pirimicarb fremgår af Tabel 4. Tabel 4 viser, at der er 8% immobile dyr i den ufodrede kontrolgruppe. Der blev observeret en visuel forskel i størrelsen blandt kontroldyrene. Fodrede dyr var større end ikke fodrede (kontrol) Ligeledes havde ikke fodrede dyr en større tendens til at opholde sig ved bunden frem for at svømme rundt i vandsøjlen. Blandt eksponerde dyr ses der 100% immobilitet uafhængig af eksponeringstid, efter endt pulseksponering. Tabel 4: Andel immobile dafnier (i procent) hhv. fodrede og ikke-fodrede efter pulseksponering med pirimicarb (100 μg/l).
Fodrede grupper genvinder i større grad deres mobilitet sammenlignet med ikke fodrede. Der er således kun 22% og 24% immobile dyr efter 24 timer i grupper, der har fået en eksponering på henholdsvis 6 og 8 timer, og som får foder, sammenlignet med 44% og 61% immobile i ikke fodrede grupper, der har fået en puls af samme varighed. Efter 48 timer er tallet faldet til 8% i alle fodrede grupper, hvor der stadig er henholdsvis 34% og 33% immobile i ikke fodrede grupper. Forsøget blev gentaget med en fastholdt koncentration af dimethoat (30 mg/l) og pulsvarigheder på 3, 4 og 6 timer. Ved disse forsøg blev der ikke set nogen effekt af fodring. På baggrund af forsøgene udført med pirimicarb blev det besluttet at fodre dyrene umiddelbart efter overførslen til rent medie. Hermed opnås også den fordel, at alle gennemførte test har samme udgangspunkt, idet fodring er en nødvendighed i de efterfølgende test med gentagne pulse og med observation over en 21 dages periode. 4.3 PulstestPulseksponeringer af Daphnia magna blev udført for alle teststoffer, og i Figur 13-20 er ændringen i mobiliteten illustreret fra eksponeringen stopper (t = 0) og til observeringsperioden ophører (t = 48). Disse observationer er anvendt til estimation af eksponeringstid-responskurver, og ud fra disse er PE0t -IT50, PE24t-IT50 og PE48t-IT50 værdier beregnet og vist i Tabel 5-8. Alle stoffer er testet i flere koncentrationer, men for overskuelighedens skyld vil der i det følgende kun blive vist resultater for udvalgte koncentrationer for hvert enkelt stof. Data og estimerede tid-responskurver for samtlige test fremgår af Bilag 4. Tabel 5: PEt-IT50- værdier for dimethoat, pirimicarb, chlorpyrifos og esfenvalerat med til hørende 95% konfidensintervaller.
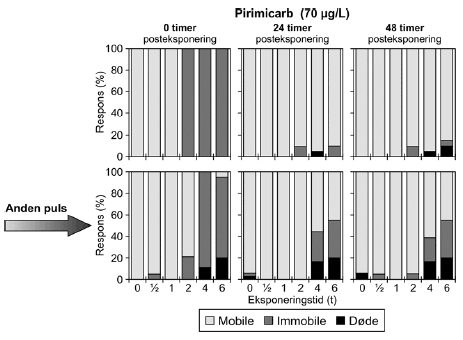
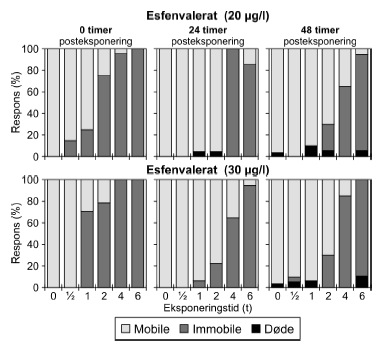
For dimethoat og pirimicarb blev det observeret, at testorganismerne genvinder mobiliteten inden for 24 timer efter overførsel til rent medie. Dette er også afspejlet i de stigende PEt-IT50-værdier (PE24t-IT50 > PE0t-IT50) (Tabel 5). Der er ikke markante forskelle mellem PE24t-IT50 og PE48t-IT50, og dyrenes tilstand forbedres således ikke målbart ved længere ophold i rent medie. Det skal også bemærkes, at de beregnede PEt-IT50-værdier, som forventet, falder med stigende eksponeringskoncentration, d.v.s. at den pulsvarighed, der skal til for at hæmme mobiliteten af 50% af dyrene, forkortes når koncentrationen øges (se Bilag 4). PEt-IT50-værdierne (Tabel 5) afspejler ligeledes, at dafnier eksponeret for pirimicarb genvinder deres mobilitet i højere grad end dafnier, der er blevet eksponeret for dimethoat, selv om der er en reversibel effekt på mobiliteten efter eksponeringen for både pirimicarb og dimethoat. Effekten af pulseksponering af Daphnia magna med dimethoat og pirimicarb er indgående beskrevet i artiklen af Andersen et al. (2005). For en illustration af effekterne af dimethoat og pirimicarb, henvises til Figur 21 og 22, hvor øverste række i begge figurer viser effekterne af en enkelt eksponering med de to pesticider.
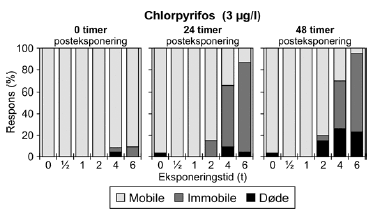
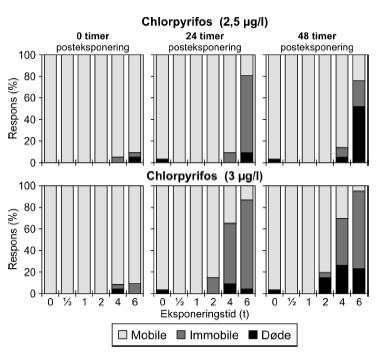
Figur 13: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 3 μg/l chlorpyrifos. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. For dyr udsat for chlorpyrifos i pulstest var der umiddelbart efter overførslen til rent medie (0 timer) kun påvirkning af få dyr ved de længste eksponeringstider (4-6 timer (Figur 13) og ingen påvirkning ved de øvrige eksponeringstider. 24 timer efter eksponeringens ophør steg andelen af immobile og døde dyr markant i grupper udsat for pulsvarigheder af 4-6 timer. Den største effekt blev set efter eksponeringen for 3 μg/l, dog var det øgede respons efter 24 og 48 timer også tydelig ved 2,5 μg/l (Bilag 4.2.1). Som vist i Tabel 5 faldt PEt-IT50-værdierne fra tiden t = 0 timer til tiden t = 24, og dyrenes tilstand forværredes således efter eksponeringens ophør. Der er ikke umiddelbart nogen forskel i de beregnede værdier fra 24 timer til 48 timer, men det er værd at bemærke, at dødeligheden steg i tiden fra 24 timer til 48 timer efter endt eksponering.
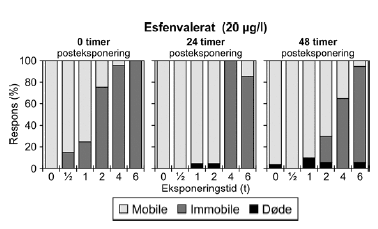
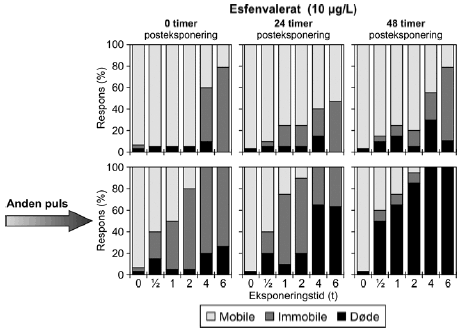
Figur 14: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 20 μg/l esfenvalerat. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. Pulsforsøget med esfenvalerat (Figur 14 og Bilag 4.2.2) viste en markant effekt på mobiliteten efter endt eksponering. Således var henholdsvis 95% og 100% af dafnierne immobile i grupper, der havde modtaget en puls af 20 μg/l og 4-6 timers varighed og 100% af dafnierne var immobile efter en puls af 30,0 μg/l og 4-6 timers varighed. Selv i grupper, der modtog pulse af 1-2 timers varighed 25-75% af dyrene var immobile). Efter 24 timer genvandt nogle af dyrene mobiliteten, som også illustreres af de stigende PEt-IT50-værdier (Tabel 5) fra 0 timer til 24 timer efter eksponering. Fra 24 timer og til 48 timer er der et lille fald i PEt-IT50-værdierne, men som vist i Figur 14 er der grundlag for at udtale sig om en egentlig forværing i dyrenes tilstand i dette tidsrum. Tabel 6: PEt-IT50- værdier for pyrazophos og azoxystrobin med tilhørende 95% konfidensintervaller.
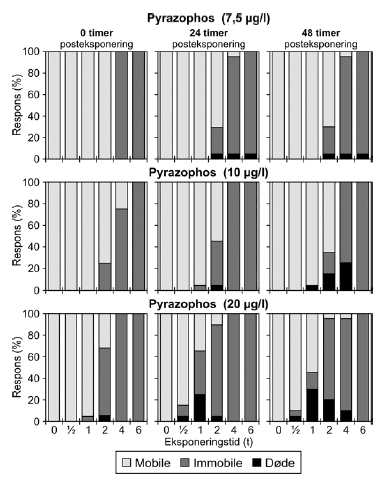
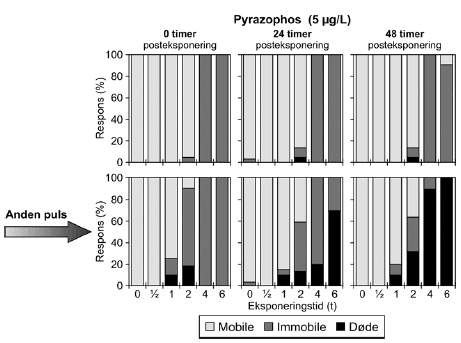
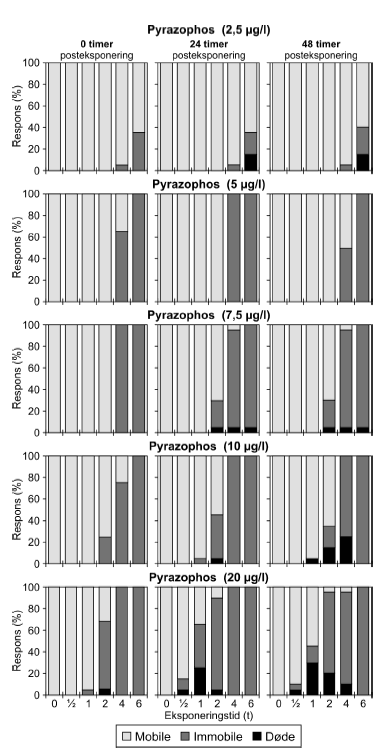
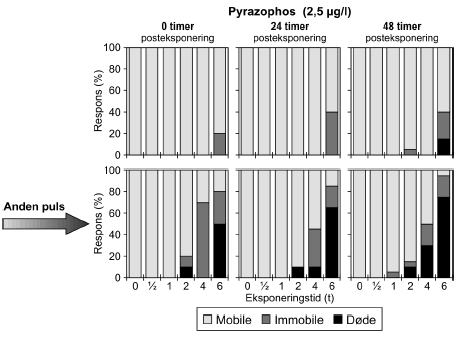
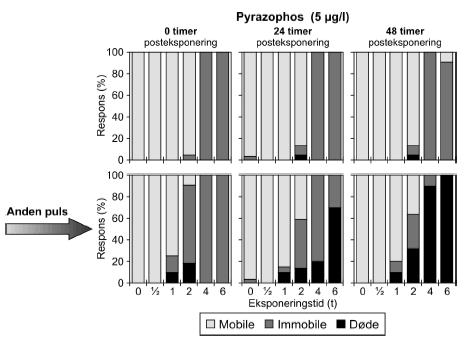
a) Kan ikke estimeres, da færre end to partielle responser. Samtlige dyr udsat for 4- og 6-timers pulse var immobile. Pyrazophos blev testet i fem koncentrationer, og i Tabel 6 er vist resultater for tre af disse, mens der for resultater af de øvrige henvises til Bilag 4.2.3. Effekten af eksponering for pyrazophos blev generelt set i form af immobilisering og ved øgede koncentrationer steg andelen af immobile dyr (Figur 15). Således blev der observeret fuld respons (100% immobilisering) ved koncentrationer ≥ 5 μg/ efter pulseksponeringer af 6 timers varighed. Grupper, der blev eksponeret for koncentrationer mellem 10-20 μg/l, viste ligeledes udprægede forsinkede effekter i form af både immobilisering og dødelighed efter eksponeringen for kortere pulsvarigheder under (2-4 timer), selv om ingen eller kun ringe effekt kunne konstateres efter pulsophør d.v.s til tiden t = 0 timer (jf. Figur 15).
Figur 15: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 7,5-20 μg/l pyrazophos. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. For forsøgene med 10 μg/l og 20 μg/l pyrazophos er ses et fald i PEt-IT50-værdierne fra tiden t = 0 timer og til tiden t = 24 timer (Tabel 6), men yderligere ophold i rent medie overordnet set ikke ændrer på dyrenes mobilitet. Det skal bemærkes, at stigningen i koncentration fra 10 μg/l til 20 μg/l resulterer i lavere PEt-IT50-værdier – altså er det, som forventeligt, kun korterevarende pulse dyrene kan modstå, hvis koncentrationen øges.
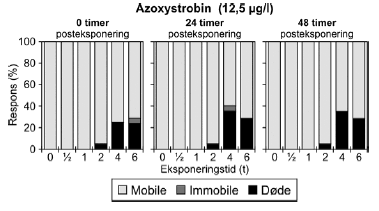
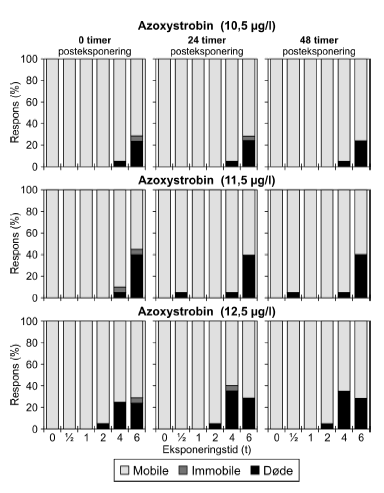
Figur 16: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 12,5 μg/l azoxystrobin. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. Pulseksponering med azoxystrobin blev udført med koncentrationer i intervallet 10,5-12,5 μg/l. Resultatet for 12,5 μg/l fremgår af Tabel 6 og Figur 16, og resultaterne for de øvrige koncentrationer fremgår af Bilag 4.1.6 og 4.2.4. Til trods for at det ikke var muligt at estimere PEt-IT50-værdier (Tabel 6), viser resultaterne i Figur 16, at der er en tydelig effekt af eksponeringen for azoxystrobin hovedsageligt i form af dødelighed. Således medførte pulseksponeringer på 4-6 timers varighed, at ca. 25% af dyrene var døde ved overførsel til rent medie (0 timer). Tabel 7: PEt-IT50- værdier for diquat og bromoxynil med tilhørende 95% konfidensintervaller.
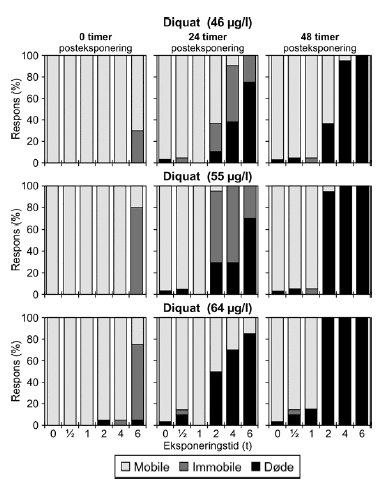
a) Kan ikke estimeres, da færre end to partielle responser. Samtlige dyr var immobile ved efter eksponering for en puls af 6 timeras varighed. b) Konfidensinterval kan ikke estimeres. Diquat er i alt testet i seks koncentrationer fra 32-64 μg/l, men kun resultater af de tre højeste koncentrationer er vist i Tabel 7 og Figur 17. For resultater af samtlige test henvises til Bilag 4.1.7 og Bilag 4.2.5. Ved de indledende forsøg, hvor diquat blev testet i koncentrationerne 32-40 μg/l, kunne der ikke ses nogen klar sammenhæng mellem eksponeringstider og koncentration. Effekten var større ved en eksponering for 32 μg/l end en eksponering for 36 μg/l, og responset efter en eksponering for 40 μg/l minder om responset, der ses efter en eksponering for 32 μg/l. Disse forsøg indikerede dog, at der efter eksponeringen for diquat vil være en forsinket effekt i form af en stigende dødelighed. Med baggrund i disse observationer, blev iværksat yderligere forsøg med højere koncentrationer. Disse forsøg viste, at efter en eksponering for en puls af 6 timers varighed og en koncentration på enten 46 μg/l, 55 μg/l eller 64 μg/l var der henholdsvis 30%, 80% og 75% immobile dyr umiddelbart efter eksponerings ophør (Figur 17). I de resterende grupper var der stort set ingen respons ved t = 0, men tendensen, som blev set ved de lavere koncentrationer, blev nu så udtalt, at en egentlig forsinket negativ effekt nu kunnne observeres i grupper udsat for pulsvarigheder på 2 timer eller mere. PEt-IT50 værdierne kunne dog, som vist i Tabel 7, ikke beregnes for alle koncentrationer, men af Figur 17 fremgår det tydeligt, at responset 24 timer efter pulsens ophør tiltog både i form af immobilitet og dødelighed. Efter 48 timer døde de dyr, der tidligere var immobile (95-100% af de dyr, der har modtaget pulse af 4-6 timers varighed er døde uafhængig af koncentrationen). Efter eksponering for en puls af 2 timers varighed var andelen af døde dyr efter 48 timer, hhv. 37%, 95% og 100% for de tre testede koncentrationer (Figur 17).
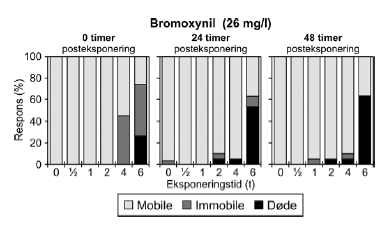
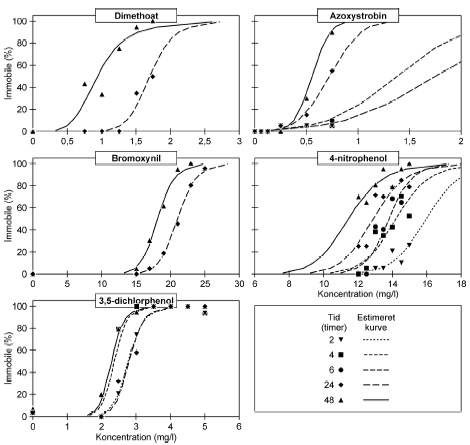
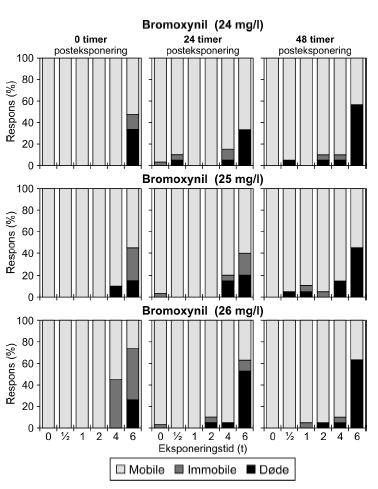
Figur 17: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for diquat (46, 55 og 64 μg/l). Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. Ved pulstest med bromoxynil blev der set en markant effekt på mobiliteten efter eksponeringen for en puls af 6 timers varighed, og dødeligheden steg i tiden efter eksponeringen (Figur 18).
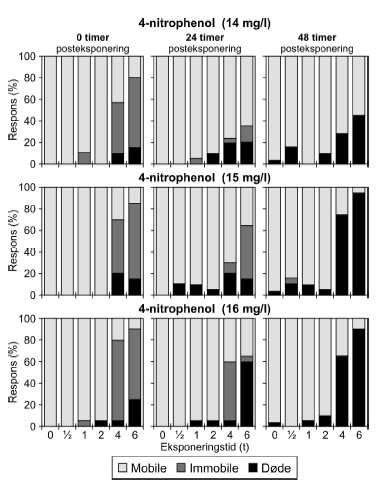
Figur 18: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 26 mg/l bromoxynil. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. Af Figur 18 fremgår ligeledes, at der var forsinkede effekter efter 24 og 48 timer hos de grupper, der har fået en puls af kortere varighed (1-4t). Ved koncentrationen 26 mg/l genvinder størstedelen af dyr eksponeret i 4 timer mobiliteten efter 24 timer, men for de øvrige testede koncentrationer viser de beregnede PEt-IT50- værdier for bromoxynil (Tabel 7 og Bilag 4.1.8) ingen forskelle fra tiden t = 0 timer til tiderne t = 24 timer og t = 48 timer. Tabel 8: PEt-IT50- værdier for referencestofferne 4-nitrophenol og 3,5-dichlorophenol med tilhørende 95% konfidensintervaller.
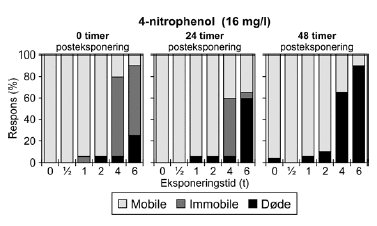
Ved pulstest med 4-nitrophenol, blev der set effekter i form af immobilitet og dødelighed efter pulseksponeringer af 4-6 timers varighed (Figur 19 og Bilag 4.2.7). Med tiden efter eksponering ændredes effekten fra at være domineret af immobilitet (0 timer) til udelukkende at omfatte dødelighed (48 timer). Ligeledes blev der set forsinkede effekter efter eksponeringen for pulse af kortere varighed end 4 timer, når dyrene havde opholdt sig 48 timer i rent medie. PEt-IT50-værdierne viser en forskel fra tiden t = 0 timer til tiden t = 24 timer (Tabel 8 og Bilag 4.1.9), hvor de beregnede PEt-IT50 værdier stiger med tiden, men denne tilsyneladende restitution dækker altså over, at dødeligheden øges markant efter 24 timer og 48 timer i rent medie (Figur 19).
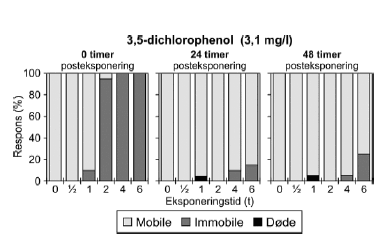
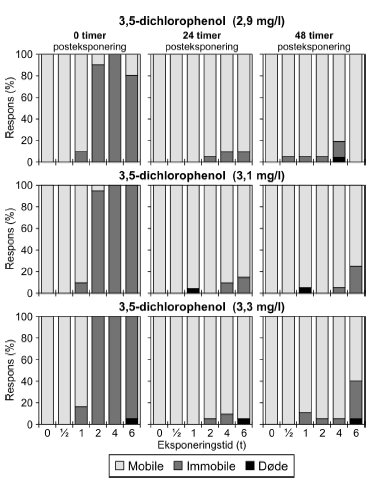
Figur 19: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 16 mg/l 4-nitrophenol. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. For alle de tre testede koncentrationer af 3,5-dichlorophenol (Figur 20 og Bilag 4.2.8) blev set en effekt på mobiliteten hos dafnier eksponeret mere end 1 time. I grupper, der fik en puls af 4-6 timers varighed, var der fra 80-100% immobile til tiden t = 0 timer. Generelt genvandt dyrene deres mobilitet under observationsperioden, men efter 48 timer i rent medie var 25% af dyrene udsat for 6 timers eksponering af 3,1 mg/l stadig immobile. De beregnede PEt-IT50 værdier efter 0 timer (PE0t-IT50) er mindre end værdierne efter 24 timer (PE24t-IT50) for de testede koncentrationer (Tabel 8 og Bilag 4.1.10), som dog kun dækker et meget snævert interval (2,9-3,3 mg/l).
Figur 20: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 3,1 mg/l 3,5-dichlorophenol. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.4 Opsummering af forsinkede effekter af pulseksponeringI Tabel 9 er resultater af pulstest med immobilisering som effekt angivet som ”recovery/restituering”, ”forsinket negativ respons” eller ”intet forsinket respons” vurderet ud fra de opnåede PEt-IT50-værdier. Dafnier eksponeret for enten dimethoat, pirimicarb, esfenvalerat, bromoxynil, 4-nitrophenol eller 3,5-dichlorophenol genvandt mobiliteten under observationsperioden (restituerede). Dette blev indikeret ved stigende PEt-IT50-værdier med stigende observationstider. Ud fra den grafiske fremstilling er dette dog kun at se for dimethoat, pirimicarb (Figur 21-22 samt Figur 9, 10, 11 og 12 Bilag 5), esfenvalerat og 3,5-dichlorophenol. Både eksponering for diquat, chlorpyrifos (3 μg/l) og pyrazophos viste en forsinket negativ effekt under observationsperioden, dette blev både afspejlet i de beregnede PEt-IT-værdier samt i den grafiske fremstilling. For azoxystrobin blev der ikke konstateret forskelle i mobiliteten ved de anvendte koncentrationer efter eksponeringens ophør. Tabel 9: Oversigt over forsinkede effekter på mobiliteten af Daphnia magna for 10 stoffer testet i pulstest med 48 timers posteksponeringsperiode.
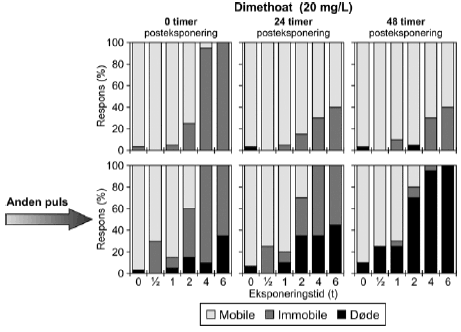
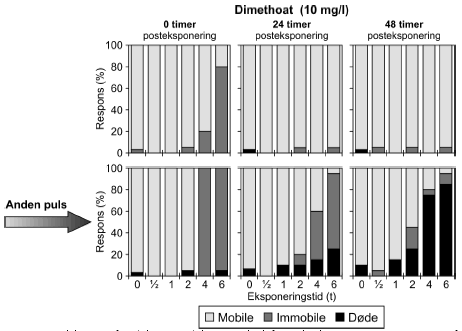
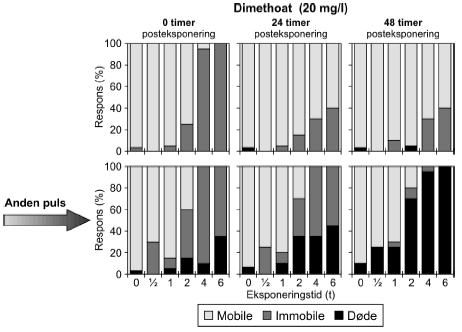
4.5 Gentagne pulseResultaterne af forsøg med gentagne pulseksponeringer med dimethoat (20 mg/l) ses i Figur 21 og øvrige grafer er vist i Bilag 5.1.1. Umiddelbart efter den første puls var henholdsvis 25%, 95% og 100% af dyrene immobile efter en puls af henholdsvis 2, 4 og 6 timers varighed. Efter 24 timer var der 15%, 30% og 40% immobile i de samme grupper og situationen ændredes ikke efter 48 timer på nær i gruppen, der fik en puls på 2 timer, hvor der var 5% døde og ingen immobile. Den anden puls havde en større effekt end den første. Umiddelbart efter pulseksponeringens ophør var der 100% effekt efter en puls på 4 og 6 timer og 30% 15% og 60% effekt i grupperne, der modtog en puls på 0,5, 1 og 2 timer. effekten var hovedsagelig i form af immobilisering, men ved de tre længste eksponeringstider var der 15%, 10% og 35% døde (2, 4 og 6 timer). I løbet af den anden posteksponeringsperiode steg andelen af døde dyrene, således at der efter 48 timer var 25%, 75% og 85% døde ved de tilsvarende tider. Selv ved de laveste eksponeringstider er andelen af døde markant større 48 timers efter anden pulseksponering (25% for både 0,5 og 1 times eksponering).
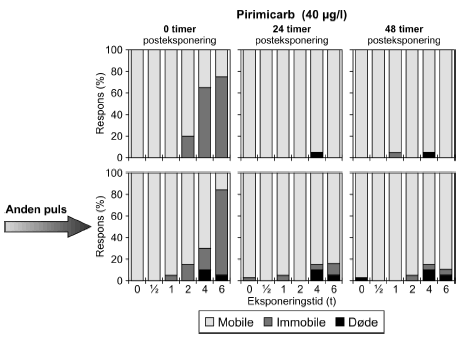
Figur 21: Gentagne pulseksponeringer med dimethoat 20 mg/l med 48 timers mellemrum. De beregnede PEt-IT50-værdier med tilhørende konfidensintervaller (Tabel 11) viser tegn på en koncentrationsafhængig forskel i responset. Således er PEt-IT50-værdierne beregnet for 10 mg/l dimethoat højere end de tilsvarende værdier beregnet for 20 mg/l dimethoat. Dette er gældende for både første og anden puls. Resultaterne for gentagne pulseksponeringer for pirimicarb fremgår af Figur 22 (70 μg/l), mens øvrige resultater er vist i Tabel 11 og Bilag 5.1.2.
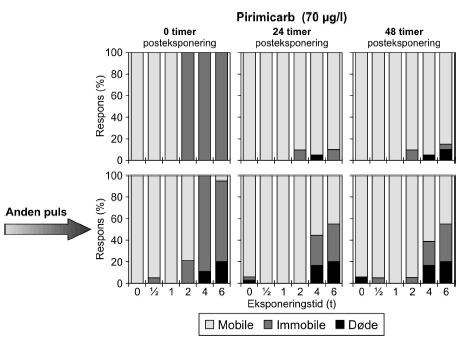
Figur 22: Gentagne pulseksponeringer med pirimicarb (70 μg/l) med 48 timers mellemrum. Tabel 11: Beregnede PEt-IT50 med tilhørende 95% konfidensintervaller for dafnier henholdsvis <24t for to gentagne pulse dimethoat, esfenvalerat, pirimicarb og pyrazophos.
a) Kan ikke estimeres, da færre end to partielle responser. Eksponeringen for 70 μg/l pirimicarb (Figur 22) resulterede i at 100% af dyrene blev immobile i grupperne, der modtog en puls af 2-6 timers varighed. Efter 48 timer faldt responset til 10%, 5% og 15% i de tre grupper. I 6 timers gruppen var der 10% af responset, der skyldes dødelighed og i 4 timers gruppen skyldtes hele responset dødelighed. Endnu en puls resulterede i henholdsvis 21%, 100% og 95% respons i de samme grupper. Henholdsvis 11% og 15% af responset kunne tilskrives dødelighed i 4- og 6 timers grupperne. Alle dyrene i 2 timers gruppen kom sig i løbet af 24 timer, men i 4- og 6 timers gruppen var der 45% og 55% af dyrene, der stadig var påvirket efter 24 timer, heraf var henholdsvis 17% og 20% døde. Situationen er stort set uændret efter 48 timer. De beregnede PEt-IT50-værdier for den anden pulseksponering for pirimicarb er lavere end for den første. For pirimicarb er der, som det var tilfældet for dimethoat, fundet lavere PEt-IT50-værdier ved en koncentrationsstigning.
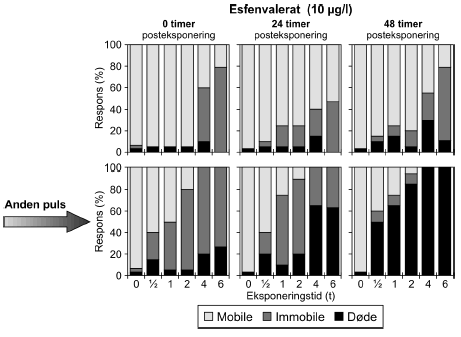
Figur 23: Gentagne pulsekponering med esfenvalerat (10 μg/l) med 48 timers mellemrum. Resultaterne for gentagne pulseksponeringer af Daphnia magna for esfenvalerat ses i Figur 23. Eksponeringen medførte et markant indledende respons ved eksponeringstider >2 timer, hvor der var henholdsvis 50% og 79% immobile. Efter 24 timer var responset faldet ved de længste eksponeringstider (4 og 6 timer), men steget hos de lavere eksponeringstider. Endnu en puls af esfenvalerat efter 48 timer, havde en endnu mere udpræget effekt på dyrene. Således blev et respons på 100% observeret umiddelbart efter eksponeringen (t = 0) for en puls af 4 og 6 timers varighed hvoraf 20% og 26% skyldes dødelighed. Tilsvarende er der 40%, 50% og 80% respons efter 0,5, 1 og 2 timers eksponering, hvoraf 15% er dødelighed i 0,5 timers gruppen og 5% er dødelighed i de to andre grupper. Efter en posteksponeringsperiode på 24 timer er responset stort set uændret i alle grupperne, dog er der tale om en stigning i dødeligheden i grupperne eksponeret for en puls af 4 og 6 timers varighed. Efter 48 timer er dødeligheden endnu mere markant i samtlige grupper. Således ses 50%, 65%, og 85% døde i grupperne eksponeret for en puls af 0,5, 1 og 2 timers varighed og 100% dødelighed ved grupper eksponeret for en puls af 4 og 6 timers varighed. De beregnede PEt-IT50 -værdier for esfenvalerat (Tabel 11) viser, at den anden pulseksponering medfører en endnu lavere mobilitet end ved første puls. Dyrene får det desuden værre med tiden, som også vist i Figur 23, idet PEt-IT50 falder fra 0,88t [0,72; 1,03] til 0,63t [0,53; 0,72]efter en observationstid på 24 timer og til 0,47t [0,35; 0,57] timer efter en observationstid på 48 timer. Et eksempel på resultater opnået ved gentagne pulseksponeringer med pyrazophos er vist i Figur 24, men øvrige resultater er vist i Tabel 11 og Bilag 5.1.4.
Figur 24: Gentagne pulseksponering med pyrazophos (5 μg/l) med 48 timers mellemrum. Efter en enkelt puls var der allerede efter endt eksponering 100% immobile dyr i grupperne, som fik en puls af 4-6 timers varighed. Responset var stort set uændret i disse grupper efter 48 timer i rent medie. Endnu en pulseksponering med samme koncentration resulterede ligeledes i et respons i grupper, der modtog pulse af 1-2 timers varighed. Efter 48 timer var det samlede respons stort set uændret i grupperne, men der skete en markant stigning i andelen af dyr, der var døde. Der var således henholdsvis 10%, 32%, 90% og 100% døde dyr ved de fire længste eksponeringstider. De beregnede PEt-IC50-værdier viste ingen forskel fra tiden 0 timer til tiden 24 timer eller 48 timer. Der ses dog et fald i de beregnede værdier med stigende koncentration. Værdierne falder ligeledes fra puls nr.1 til puls nr. 2 (Tabel 11). 4.6 Pulseksponering – aldersvariation i følsomhedDafnier indenfor fire aldersgrupper (<24t, 3d, 5d dog 7d) blev eksponeret for en puls af pirimicarb for at teste en evt. variation i følsomheden blandt disse grupper. Testkoncentrationerne blev valgt ud fra den indledende akuttest med pirimicarb (Tabel 3). Under akuttesten blev IC90,24t bestemt til 29 μg/l, og efter 6 timer var 25% af dafnierne eksponeret for 30,0 μg/l (den højeste testkoncentration under akuttesten) immobile (60% af disse var døde). Til aldersforsøgene blev en koncentration på 40 μg/l derfor anvendt, for at sikre, at der ville være et respons, også hvis dyrene (ældre end 1 dag) skulle være mindre følsomme. Forsøget blev efterfølgende gentaget med en koncentration på 70 μg/l. Tabel 12 viser andelen af immobile og døde blandt dafnier af forskellige aldersgrupper. Den største følsomhed overfor en puls af pirimicarb på 40 μg/l, blev set hos dafnier, der var 3 dage gamle på eksponeringstidspunktet (70% immobile), og den mindste følsomhed hos 5 dage gamle dyr (40% immobile). Såvel 24 timer som 48 timer efter eksponeringens ophør var der stort set ingen immobile eller døde dyr på nær i gruppen med 3 dage gamle dafnier, hvor der var henholdsvis 30% og 25% immobile efter 24 og 48 timer. Ved en koncentration på 70 μg/l var responset størst hos dafnier, der var <24 timer gamle, hvor 65% var immobile sammenlignet med 0% immobile i gruppen med 3 dage gamle dafnier, og 15% immobile i grupperne med henholdsvis 5 og 7 dage gamle dafnier. 48 timer efter eksponeringen var andelen af immobile dafnier faldet i alle grupperne. Der var dog stadig størst respons i gruppen med <24 timer gamle dafnier (15% immobile), men samtidig var der 10% døde i gruppen med 7 dage gamle dyr. Tabel 12: Andelen af immobile/ døde dafnier (i procent) indenfor forskellige aldersklasser efter pulseksponering for pirimicarb (6 timers varighed). n=20 for eksponerede grupper og n=30 for kontrolgrupperne.
a) Antal dyr: n=29. Forsøget blev gentaget med vægt på sammenligning af <24 timer og 3 dage gamle dafnier (alderen dyrene har når de modtager en gentagen puls), der blev eksponeret for en puls af pirimicarb eller dimethoat. Resultaterne fremgår af Tabel 13 og 14, hvor data for eksponeringen af <24 timer gamle dafnier er medtaget - både for enkelt pulstest og gentagne pulstest. Værdierne fundet for 3 dage gamle dafnier, der er blevet eksponeret for 10 mg/l dimethoat (Tabel 13) ligner de værdier, der er fundet for <24 timer gamle dafnier. De beregnede PEt-IT50-værdier for 0 timer er højere for dyr eksponeret for 10 mg/l end dyr eksponeret for 20 mg/l. Med tiden bliver forskellen mindre tydelig. Eksponering for pirimicarb (Tabel 14) viser ikke en forskel i responset mellem dafnier, der er <24 timer gamle, og dafnier, der er 3 dage gamle. PEt-IT50- værdierne beregnet for dyr eksponeret for 40 μg/l er højere end værdierne beregnet for 70 μg/l, til tiden 0 timer, men til tiderne 24 timer og 48 timer er der ingen forskel. Tabel 13: Beregnede PEt-IT50 med tilhørende 95% konfidensintervaller for dafnier henholdsvis <24t og 3 dage gamle eksponeret for dimethoat. Bemærk at data i denne tabel ikke er identiske med data i Tabel 5, da der i denne undersøgelse blev udført nye forsøg.
a) Fundet i pulstest. b) Fundet for første 1. puls i test med gentagen pulseksponering. c) Kan ikke estimeres p.g.a. datas beskaffenhed. Tabel 14: Beregnede PEt-IT50 med tilhørende 95% konfidensintervaller for dafnier henholdsvis <24t og 3 dage gamle eksponeret for pirimicarb. Bemærk at data i denne tabel ikke er identiske med data i Tabel 5, da der i denne undersøgelse blev udført nye forsøg.
a) Fundet i pulstest. b) Fundet for første 1. puls i test med gentagen pulseksponering. c) Kan ikke estimeres pga. datas beskaffenhed. 4.7 Langtidseffekter af pulseksponeringDer er i alt udført 12 forsøg med pulseksponering, hvor dyrene efterfølgende har været observeret i 21 dage i rent medie. Stofferne dimethoat, pirimicarb, esfenvalerat, chlorpyrifos, bromoxynil og 4-nitrophenol er blevet undersøgt. For flere af teststofferne blev der ikke observeret langtidseffekter, og samtlige data vil af pladshensyn ikke blive præsenteret i det følgende. Der vil blive fokuseret på de stoffer, hvor effekter blev fundet, og for resultater af alle de gennemførte test henvises til Bilag 6. Under testene med dimethoat og pirimicarb, blev der tilført alge i overskydende mængder (>0,2 mg C/dag), mens der i de øvrige tests blev fodret i henhold til anbefalingen for reproduktionstest med dafnier (OECD, 1998). 4.7.1 Antal afkom efter pulseksponeringDer blev udført to test med dimethoat ved koncentrationen 30 mg/l, da det ved den første test ikke var muligt at opfylde OECD kravet om minimum 60 unger pr. moderdyr. Den første test viste dog, at antallet af unger var markant lavere hos dafnier, der 14 dage tidligere havde modtaget en puls af 3 timers varighed. 21 dage efter pulseksponeringen var der forskel ved alle eksponeringstider på nær hos dafnier eksponeret 1 og 2,5 time. Anden test med dimethoat (30 mg/l) viste ingen forskelle i antallet af afkom efter 14 dage, men efter 21 dage havde alle grupper eksponeret for en puls ≥2 timer produceret markant færre unger end kontrolgruppen (Figur 25). Pulseksponering af dafnier med pirimicarb (100 μg/l) viste et markant fald i antallet af unger pr. moderdyr efter såvel 14 dage som 21 dage hos grupper, der modtog en puls af 2-6 timers varighed (Figur 25). Pulseksponeringen for 3,5-dichlorophenol (1-6 timer) havde en effekt på antallet af afkom efter 14 dage, mens der efter 21 dage ikke var forskel i antallet af unger produceret som funktion af eksponering for 3,5-dichlorophenol. Hverken efter 14 dage eller 21 dage var antallet af unger fra dyr pulseksponeret med chlorpyrifos, bromoxynil eller 4-nitrophenol forskelligt fra kontrolgrupperne.
Figur 25: Antal unger og længde af moderdyr 21 dage efter pulseksponering med dimethoat (30 mg/l) og pirimicarb (100 μg/l). Gennemsnit og 95% konfidensintervaller er vist (n=10). 4.7.2 Antal dage før første reproduktionEn puls af dimethoat (30 mg/l) af 3 timers varighed forlængede antallet af dage før første afkom fra i gennemsnit 7,70 dage til 9,17 dage (Tabel 15). Pulseksponeringer af kortere varighed medførte ligeledes forsinkelser i reproduktionen, men ikke helt så markant som efter eksponeringen for en puls af 3 timers varighed. Eksponering for 100 μg/l pirimicarb i pulse med varigheder fra 1-6 timer medførte tilsvarende en forsinkelse i antallet af dage før første reproduktion. Kontrolgruppen fik i gennemsnittet deres første afkom efter 8,10 dage sammenlignet med 9,20-9,60 dage for de eksponerede grupper. Af de øvrige testede stoffer (esfenvalerat, chlorpyrifos, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol) var det kun for 4-nitrophenol, at der kunne ses tegn på en effekt på antallet af dage før første reproduktion (2 mg/l (0,5, 4 og 6 timer) samt 8 mg/l (1 og 6 timer)). Der ses dog ingen entydig sammenhæng mellem pulsvarigheden og antallet af dage. Det ses desuden at de grupper, der fik en pulseksponering for enten dimethoat eller pirimicarb, får deres første kuld efter omkring samme tid som kontrolgrupperne for chlorpyrifos, esfenvalerat, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol. At moderdyr i kontrolgrupperne for dimethoat og pirimicarb får unger tidligere end moderdyr i de øvrige forsøg, kan hænge sammen med den øgede fodermængde, der blev tilført under disse forsøg (se afsnit 3.5.6). Der blev dog ikke set nogen forskel på effekten af pulseksponeringer hos fodrede og ikke fodrede dyr, og fodring skulle ud fra disse forsøg ikke have nogen betydning. Tabel 15: Gennemsnitlig antal dage før første reproduktion, med tilhørende 95% konfidensintervaller, efter pulseksponering med udvalgte pesticider og referencestoffer.
4 NP: 4-nitrophenol; 3,5 DCP: 3,5-dichlorophenol a) ikke bestemt 4.7.3 Kropslængde og vægt efter pulseksponeringSom vist i Figur 25, reducerede en dimethoat eksponering på 30 mg/l længden af moderdyrene efter 21 dage for alle pulsvarigheder (1-3 timer). Af samme figur fremgår det, at en pirimicarb eksponering på 100 μg/l tilsvarende reducerede længden af moderdyrene efter 21 dage for pulsvarigheder af 3-6 timer. Pulseksponeringer for esfenvalerat, chlorpyrifos, bromoxynil og 4 nitrophenol viste ingen effekt på længden af moderdyr efter 21 dage. Vægten af overlevende moderdyr blev ikke påvirket af pulseksponeringer for de her undersøgte pesticider (dimethoat, pirimicarb, esfenvalerat, chlorpyrifos, bromoxynil og 4 nitrophenol). 4.7.4 Antal unger, eksponeringstid og kropslængdeSom beskrevet i metodeafsnittet blev det undersøgt om der var en sammenhæng mellem antallet af unger, eksponeringstiden og kropslængden. Parameterestimater for hhv βeksponeringstid og βlængde er angivet i Tabel 16. Det fremgår af tabellen, at der for stofferne bromoxynil, chlorpyrifos, 4-nitrophenol og 3,5-dichlorophenol ikke er en statistisk signifikant sammenhæng mellem eksponeringstiden og antallet af afkom. For dimethoat betød en øget eksponeringstid et signifikant fald i antallet af unger der blev produceret i løbet af de 21 dage testen forløber over (p = 0.002). En tilsvarende effekt sås efter eksponeringen for pirimicarb (p = 0.007). Tilsvarende ses der en signifikant sammenhæng mellem det producerede antal unger og moderdyrets længde efter 21 dage hos dafnier eksponeret for henholdsvis dimethoat, pirimicarb, esfenvalerat, chlorpyrifos og 3,5-dichlorophenol. Dafnier, der har haft en reduceret vækst, og dermed kortere kropslængde efter 21 dage, producerer således signifikant færre unger (p = 0,001 (dimethoat), p < 0,001 (pirimicarb), p = 0,0024 (chlorpyrifos), p =0,024(esfenvalerat) og p = 0,0036 (3,5-dichlorophenol)) end dafnier, der opnår en større kropslængde i løbet af forsøgsperioden. Tabel 16: Parameterestimater for længde og eksponeringstid for de enkelte stoffer i test for kronisk virkning.
*signifikant forskellig fra kontrollen 5 Diskussion
5.1 TestudviklingEn meget væsentlig del af nærværende projekt har været fokuseret på udvikling af en testmetodik til påvisning af toksiske effekter af pulseksponering. Den endelige metode er beskrevet i detaljer i testprotokollen i Bilag 1, og der vil i det følgende blive givet en diskussion af væsentlige forhold og overvejelser, der har indgået i testudviklingen. Generelt vurderes det, at de anvendte tests er lette at udføre indenfor en overskuelig tidsramme (3 dage for akut virkning og 21 dage for kronisk virkning), der anvendes få materialer, og Daphnia magna har vist sig som en velegnet testorganisme. Metodemæssigt er der et vist overlap mellem de gennemførte undersøgelser og den metode Naddy et al. (2000) har anvendt. Deres studie er dog ikke fokuseret mod udvikling af en egentlig testmetodik, men snarere mod beskrivelse af effekten af et enkelt pesticid (chlorpyrifos), og der kan derfor stilles spørgsmålstegn ved generaliseringsværdien af undersøgelserne udført af Naddy et al. (2000). I denne sammenhæng er det af stor betydning, at der for chlorpyrifos ikke blev observeret hverken forsinkede eller kroniske effekter, hvorfor yderligere dokumentation af metoden anvendt af Naddy et al. (2000) har været nødvendig for at vurdere, om metoden er egnet til generelt at beskrive effekter af pulseksponeringer med pesticider. Testmetodikken i Naddy et al. (2000) forekommer umiddelbart at være ganske omstændig og er ikke optimeret i forhold til brug i regulatorisk sammenhæng. Den opkobling til ISO/OECD-metoder, der er anvendt i nærværende rapport, vurderes at give et mere effektivt testdesign uden at væsentlige informationer mistes. Som et eksempel kan nævnes, at de præliminære ISO-test med hyppige observationer indenfor de første 24 timer understøtter valg af relevante testkoncentrationer, således det i vidt omfang blev undgået, at de ressourcekrævende kroniske test blev udført som ”trial-and-error”-forsøg. Dog skal esfenvalerat nævnes som en undtagelse, idet dette stof p.g.a. af ringe vandopløselighed, og sandsynligvis også høj biokoncentrering, voldte problemer ved gennemførslen af kroniske test. 5.1.1 Endpoints for effekter af pulseksponeringI de udførte undersøgelser, blev mobiliteten anvendt som endpoint til vurdering af akutte effekter. I overensstemmelse med ISO-standarden for dafnietest (ISO, 1989a), blev det defineret, at dyrene var immobile, hvis de ikke var i stand til at svømme i løbet af en 15 sekunders periode efter evt. stimulering. Problemet ved at benytte immobilitet er, ud over at det er en mere eller mindre subjektiv vurdering, at nogle dyr måske nok kan svømme, men samtidig udviser en svømmeadfærd, der tydeligvis er forskelligt fra kontrolgruppens adfærd. Vi så således både dyr, der generelt var mere sløve, men også dyr, der svømmede mere ved bunden og i mere roterende bevægelser. Der er ingen tvivl om, at disse dyr har været påvirket af pesticideksponeringen, men da den internationalt accepterede definition på mobilitet blev fastholdt gennem hele studiet, var det ikke muligt at inddrage dette respons i de endelige resultater. Hvis forsøgene var blevet udvidet til at definere en grad af immobilitet som f.eks. mobile (kontrol), mobile (ikke kontrol) og immobile, og rangordnet i henholdt til ”mobilitetsstatus” med værdier (f.eks 0: mobile (kontrol); 1: mobil (ikke kontrol); 2: (immobile)) kunne vi have beregnet en slutværdi for hver gruppe af dafnier (som i flg. eksemplet ville ligge mellem 0 og 2), og som ville fortælle os graden af mobilitet i hver gruppe. Dette ville alt andet lige have resulteret i lavere effekttider og -koncentrationer, end de vi har bestemt ud fra resultaterne i denne undersøgelse. Et andet alternativ ville være at finde et andet endpoint end immobilitet som f.eks. svømmeevne bestemt ved metoden beskrevet af Lauridsen et al. (2003). Det vurderes dog, at denne ændring af pulstestene ville føre til mere arbejds- og omkostningskrævende test, og det meget nære ”slægtskab” med eksisterende internationalt accepterede testmetoder vil også gå tabt ved denne ændring. I forbindelse med en evt. restitution eller en forsinket negativ effekt er det dog nødvendigt at evaluere de anvendte observationstider. I de her udførte test anvendte vi en observationsperiode på 48 timer. De kroniske forsøg med esfenvalerat viste imidlertid, at der efter 48 timer stadig kan ske en ændring i responset. Vi så således en øget dødelighed også selv om der var tale om et recovery i løbet af de første 24 timer efter pulseksponeringer for esfenvaleratkoncentrationer på 1,00-20,0 μg/l. De beregnede PEt-IT50-værdier for de tilsvarende koncentrationer (Bilag 4) viste dog tegn på en yderligere ændring i tidsrummet 24-48 timer. Til sammenligning fandt Naddy & Klaine (2001), at det var nødvendigt med en observationsperiode på mindst 72 timer for at opnå et recovery hos Daphnia magna efter eksponeringer for 0,5 μg/l chlorpyrifos og 96 timer ved en koncentration på 1,0 μg/l. Altså er det for visse stoffer vist, at der kræves en længere observationsperiode efter pulseksponering ved stigende koncentrationer for endeligt at afgøre effekterne af en pulseksponering. Esfenvalerat og chlorpyrifos er klart de mest lipofile af de stoffer, der er testet i nærværende undersøgelse, og længden af den krævede observationsperioden kan meget vel være relateret til teststoffernes fysisk-kemiske egenskaber, hvor fedtopløsligheden (Kow-værdien) kan vise sig at være nøgleparameteren. Undersøgelser af relationer mellem fysisk-kemiske stofegenskaber og effekter af pulseksponering er yderst relevante, men falder dog uden for rammerne af nærværende projekt. De udførte pulstests med reproduktion som endpoint, viste at eksponeringen for stoffer (dimethoat og pirimicarb) som dafnierne umiddelbart var i stand til at komme sig efter, havde en kronisk effekt på reproduktionen. Vi så således et fald i antallet af afkom samt en forsinkelse i reproduktionen. Ligeledes påvirkede eksponeringen dyrenes vækst således at de eksponerede dyr opnåede en kortere længde i løbet af de 21 dage testen forløb over. Statistiske undersøgelser viste ligeledes, en sammenhæng mellem både kropslængde og pulsvarigheden og antallet af unger, der blev produceret, og der er gode perspektiver i at forfølge denne indfaldsvinkel til forudsigelse og fortolkning af reproduktionseffekter af pulseksponering. De kroniske tests kunne desuden udvides til at inkludere reproduktionsevnen hos afkom. Der er i litteraturen beskrevet undersøgelser (kontinuert eksponering 21 dage), hvor anden og tredje generations afkom er blevet inddraget, og hvor effekter i den videre reproduktion er blevet iagttaget (bl.a. Sánchez et al., 1999; Hammer-Wirtz & Ratte, 2000), og det kunne være interessant at se, om dette også er tilfældet efter pulseksponeringer. 5.1.2 Fodring i posteksponeringsperiodenSom et led i testudviklingen blev det undersøgt om fodring i posteksponeringsperioden påvirkede resultaterne opnået ved pulstest. Fodring er en nødvendighed for test af en varighed >48 timer, når der anvendes nyfødte dyr, og er således uundgåelig for test med gentagne pulseksponeringer og test til undersøgelse af kroniske effekter af pulseksponering. For pulstest med immobilisering som endpoint var udgangspunktet, at fodring ikke skulle foretages, da fordelingen af teststof mellem dyr og testmedium kunne forstyrres af dette, og der heller ikke i metodeoplægget har været anvendt fordring (Friis-Nielsen, 2002; Brent & Herricks, 1999). I litteraturen er der eksempler på, at tilstedeværelsen af føde kan øge den toksiske virkning af et stof (Naddy & Klaine, 2001), men tilsvarende er der også eksempler på, at toksiciteten kan blive nedsat (Buhl et al., 1993). Fodringsforsøgene med pirimicarb viste en hurtigere restitution blandt fodrede dyr sammenlignet med dyr, der ikke blev fodret. I modsætning til disse resultater blev der ikke fundet en forskel mellem fodrede og ikke-fodrede dyrs recovery efter eksponeringen for dimethoat. Dyrene, der blev eksponeret for pirimicarb, fik imidlertid en længere eksponering (6-8 timer) end dyrene eksponeret for dimethoat (3-6 timer). Det kunne derfor tyde på at føde kun har en indflydelse på recovery efter pulse af en given varighed (i dette tilfælde >6 timer). Det er dog mere sandsynligt, at variationen skyldes de anvendte koncentrationer og pesticidet. På baggrund af disse undersøgelser blev det besluttet, at fodre dyrene under pulstestene efter de var blevet overflyttet til rent medie. Idet fodring først foretages efter eksponeringen er afsluttet vurderes det, at fodringen kun har ringe indfyldelse på teststoffets fordeling mellem vandfasen og organismerne. 5.1.3 Alder og følsomhedI de internationale standarder for dafnietestning (f.eks. ISO (1989a) og OECD (1998)) anvendes dafnier der er <24 timer gamle. Hovedparten af den tilgængelige information om kemiske stoffers toksiske virkning på Daphnia magna omhandler således eksponerering af juvenile dyr. Naddy et al. (2000) fandt imidlertid forskelle i testorganismernes respons overfor pulseksponeringer med chlorpyrifos afhængigt af dyrenes alder på eksponeringstidspunktet. Således var 3, 7 og 14 dage gamle dyr mere følsomme end nyfødte unger og udviste større dødelighed i en 21-dages test. Derfor blev der udført forsøg med 4 aldersklasser (<24t, 3d, 5d og 7d) af Daphnia magna i nærværende undersøgelse. I forsøg med pirimicarb i koncentrationen 40,0 μg/l blev det fundet, at dyr, der var 3 dage gamle, var de mest følsomme overfor pulseksponeringen, men at de <24 timer gamle dyr også havde en markant grad af immobilisering. Disse dyr kom sig dog hurtigere end de 3 dage gamle dyr, hvilket peger på <24 timer gamle dyr som værende mindre følsomme overfor pulseksponering med pirimicarb (6 timers puls ved 40 μg/l). Ved en eksponering for 70 μg/l var det dog tydeligt, at de <24 timer gamle dyr var de mest følsomme, og de 3 dage gamle dyr reagerede ikke på noget tidspunkt på en puls af denne koncentration. Derfor kan det ikke ud af de udførte forsøg udledes om ældre dyr er mere følsomme end unge dyr. Større følsomhed blandt voksne individer sammenlignet med juvenile er dog dokumenteret tidligere for invertebrater (A. aquaticus og G. pulex) eksponeret for ammoniak og hypoxi (Maltby 1995) og Naddy et al. (2000) fandt som tidligere nævnt også en større følsomhed bland 3, 7 og 14 dage gamle individer sammenlignet med <24 timer gamle. En større følsomhed hos nyfødte dafnier sammenlignet med voksne ægbærende dafnier er imidlertid blevet iagttaget efter eksponeringen for cypermethrin (Lauridsen et al. 2003). Det har således ikke på baggrund af nærværende forsøg og litteraturgennemgang været muligt at nå frem til en entydig konklusion vedrørende følsomhed i relation til dyrenes alder på eksponeringstidspunktet. Ved projektets opstart blev det valgt at opbygge testudviklingen omkring <24 timer gamle dyr, og af hensyn til sammenlignelighed med eksisterende data fra internationalt standardiserede test anser vi det for mest hensigtsmæssigt at fastholde anvendelsen af <24 timer gamle dyr ved fremtidige pulstest. Et andet forhold, som måske i højere grad end dyrenes alder, kan påvirke følsomheden, er dyrene skjoldskifte som følge af vækst. Eksponeres dyr (tilfældigvis) umiddelbart efter skjoldskifte vil kemiske stoffer nemmere kunne gennemtrænge overfladen og dermed vil dyrenes eksponering (interne dosis) kunne øges. Dette forhold udjævnes nok til en vis grad i test med kontinuert eksponering, men ved pulstest med så korte pulsvarigheder, som er anvendt i nærværende projekt, kan tidspunktet for skjoldskifte få afgørende betydning. Det er således en mulighed, at de størrelseforskelle inden for grupper, der observeres af Andersen et al. (2005) 3-9 dage efter ophøret af pulseksponering med dimethoat og pirimicarb, og som også er set i dette projekt efter 21 dage, kan henføres til, at en del af dyrene har skiftet skjold umiddelbart før eller under eksponeringen. Testteknisk kunne disse usikkerheder minimeres ved at synkronisere den anvendte kultur m.h.t. skjoldskifte eller ved at opnå en mere ensartet aldersfordeling blandt de <24 timer gamle dafnier, men under de givne forhold vurderes dette ikke at være praktisk gennemførligt. Ovenstående anbefaling af at anvende <24 timer gamle dafnier ved pulstest står derfor stadig ved magt, men det opfordres til at man ved fremtidige test har øget opmærksomhed på størrelseforskelle inden for grupper, der modtager samme puls. 5.1.4 Gentagne pulseI litteraturen er fundet flere undersøgelser, der viser, at pulse i vandmiljøet kan optræde med korte eller længerevarende mellemrum. For eksempel fandt Liess et al. (1999), at der var betydeligt flere pulse af pesticider i vandløbet i perioden april til september, hvor pesticiderne også hyppigt blev anvendt på markerne. Phillips & Bode (2004) har ligeledes dokumenteret forekomsten af gentagne pulse af pesticider med maksimumskoncentration i juni til december i forbindelse med større regnvejrshændelser. Spørgsmålet om effekter af gentagne pulspåvirkninger blev i løbet af projektperioden mere og mere påtrængende også set i lyset af, at der for flere af de undersøgte stoffer blev set et recovery efter eksponering for en enkelt puls. Information om længerevarende effekter, hos dyr der tilsyneladende havde overvundet pulspåvirkningen, blev opnået ved de forsøg, hvor dafniernes reproduktion blev observeret, men hvorvidt en gentagen pulspåvirkning vil have en forstærket effekt bliver ikke belyst ved denne testning. Som en sideaktivitet til det egentlige projekt blev derfor gennemført en række forsøg med gentagne pulseksponeringer. De fire stoffer dimethoat, pirimicarb, esfenvalerat og pyrazophos blev undersøgt i hvad, der må betegnes som indledende forsøg med gentagen eksponering, nemlig en simpel gentagelse af de allerede udførte pulsforsøg. Dyr, der allerede havde modtaget en pulseksponering, blev således udsat for endnu en puls af samme varighed og koncentration 48 timer efter afslutningen af den første eksponering. Som ventet blev der fundet en markant genvinding af mobiliteten efter en enkelt puls af både pirimicarb og dimethoat allerede indenfor de første 24 timer. Der er ingen forskel i responset efter 24 timer og 48 timer i rent medie, hvilket afspejler, at restitutionen sker indenfor de første 24 timer. Efter en gentagen eksponering var der tale om en mere udpræget immobilisering til tiden t = 0 timer. PEt-IT50-værdierne steg ligeledes med stigende posteksponeringstid, hvilket pegede i retning af restituering - dog var der ingen forskelle mellem posteksponeringstiderne. Den tidligere beskrevne forsinkede negative respons i form af et skift fra immobilisering til død fremgår således ikke af PEt-IT50-værdierne fundet for anden puls. Forsøg med gentagne pulse af pirimicarb og dimethoat viser altså, at selv om dyrene genvinder mobiliteten i løbet af 24 timer efter eksponeringens ophør, så er de tilsyneladende så svækkede, at de ikke i stand til at genvinde mobiliteten efter endnu en pulseksponering. For esfenvalerat og pyrazophos var genvindingen af mobiliteten efter første puls mindre udtalt end for dimethoat og pirimicarb, men der blev set samme tendens til at anden puls ændrede responset fra immobilisering til død. Det blev valgt at eksponere dyrene igen lige efter den første pulseksponerings-periode var overstået. Det var dels af hensyn til sammenligning med allerede udførte forsøg og dels for at undgå, at dyrene begyndte at reproducere under testen. Det er muligt, at dyrene ville have udvist en større tolerance over for anden puls, hvis pulsintervallet havde været af længere varighed, således at restitueringsperioden ville have været længere. Ligeledes kan der være tale en aldersafhængig følsomhed, f.eks. beskriver Naddy & Klaine (2001), som nævnt ovenfor, at 3-14 dage gamle dafnier var mere følsomme overfor chlorpyrifos end <24 timer gamle dafnier. De udførte aldersforsøg i nærværende projekt viste dog som tidligere beskrevet ikke en entydig sammenhæng mellem dyrenes alder og deres følsomhed. 5.2 Effekter af enkeltstoffer efter pulseksponering5.2.1 DimethoatDe fundne værdier for toksiciteten af dimethoat i standardtest stemmer godt overens med, hvad der er beskrevet i litteraturen (se Tabel 1 samt Beusen & Neven 1989; Tomlin, 2003). Ved pulstest blev det påvist, at dyrene genvandt mobiliteten efter pulseksponeringer (10-30 mg/l), og denne effekt blev påvist efter at dyrene havde opholdt sig 24 timer i rent vand (PE0t-IT50< PE24t-IT50). En stigende koncentration medførte et fald i de fundne PEt-IT50-værdier, hvilket viser, at der skal kortere tid til at opnå en effekt på 50% af dyrene ved en stigning i koncentrationen fra 10 mg/l til 20 mg/l og 30 mg/l. Det første forsøg, hvor kroniske effekter af dimethoat blev undersøgt, opfyldte ikke validitetskravene fra OECD (1998) på 60 unger pr. moderdyr. Trods for dette kan det dog konkluderes, at der var signifikant færre unger i pulseksponerede grupper end i kontrollen. Den anden test for kroniske effekter af dimethoat bekræftede, at pulseksponeringer af mere end 2 timers varighed havde en effekt på antallet af unger pr. moderdyr efter 21 dage. Resultater fra tidligere undersøgelser med dimethoat, hvor undersøgelserne er udført efter retningslinierne i OECD (1998), har vist 50%’s hæmning i antal afkom ved eksponering for 310 μg/l efter 20 dage (Canton et al., 1980) og ligeledes efter eksponering for 310 μg/l efter 16 dage (Hermens et al., 1984). NOEC for reproduktionen af Daphnia magna efter 21 dages kontinuert eksponering er fundet til 40 μg/l (se Tabel 1). Vore forsøg dokumenterer, at pulseksponeringer i lighed med kontinuerte eksponeringer kan påvirke reproduktionen. Den anvendte koncentration på 30 mg/l er en faktor 100-1000 højere, men der skulle kun en eksponering af 2,5-3 timers varighed til for at reducere antallet af unger med ca. 30% sammenlignet med kontrollen. Det blev ligeledes observeret en forsinkelse i reproduktionen efter en puls på 3 timer (Tabel 15). D.v.s. dyrene, der fik den længste pulseksponering, fik unger på et senere tidspunkt (9,2 dage) end kontrol gruppen (7,6 dage). For dimethoat vil anvendelse af data fra test med kontinuert eksponering således føre til de mest konservative skøn over beskyttelseskoncentrationer. Nærværende projekt bidrager dog med den meget vigtige, og ikke tidligere beskrevne, information, at selv meget kortvarige eksponeringer kan medføre reproduktionsskader hos Daphnia magna. 5.2.2 PirimicarbDer blev fundet IC50-værdier på 21,8 μg/l (24 timer) og 19,4 μg/l (48 timer) ved den standardiserede akuttest med kontinuert eksponering med pirimicarb. Værdierne stemmer godt overens med EC50,48t på 17 μg/l opgivet i Tabel 1 samt værdierne bestemt af Kusk (1996) på 28 μg/l og 21 μg/l for henholdsvis 24 timer og 48 timer. Dafnier restituerede efter pulseksponeringer for pirimicarb, d.v.s. de genvandt deres mobilitet efter overførslen til rent medie. Såvel eksponeringstid som koncentration var betydende for immobilisering og den efterfølgende genvinding af mobiliteten. Ud fra resultaterne konkluderes det, at dyr udsat for pulse af pirimicarb hurtigere genvinder mobiliteten end dyr udsat for dimethoat efter overførslen til rent vand. Som tidligere nævnt binder begge stoffer sig til enzymet acetylcholinesterase, og da hydrolysen af carbamat er hurtigere end af organophosphat-bundne (se afsnit 2.2.3), vil det kunne forventes, at pirimicarb (carbamat) udskilles hurtigere end dimethoat (organophosphat). I overensstemmelse med dette fandt Kusk (1996) en markant frigivelse af pirimicarb efter overførslen af Daphnia magna til rent vand, hvor kun halvdelen af pirimicarb-mængden var tilbage i dyrene efter 24 timer. Der blev produceret et lavere antal unger i løbet af de første 14 dage i grupper, der blev pulseksponeret (2-6 timer) med pirimicarb (100 μg/l). Ligeledes er der tale om en forsinkelse i reproduktionen efter eksponeringer for pirimicarb allerede ved eksponeringer af 1 times varighed. Dyrene i kontrolgruppen fik deres første kuld efter gennemsnitlig 8,1 dage og eksponerede grupper fik unger efter 9,2-9,6 dage. For test med kontinuert eksponering i 21 dage er fundet en NOEC på 0,9 μg/l, og som det var tilfældet for dimethoat vil anvendelse af denne værdi føre til et mere konservativt skøn over beskyttelseskoncentrationer i miljøet end data fra pulstestene. Dette er naturligvis forventeligt, når forskellen i eksponeringstiden (2-6 timer over for 504 timer) tages i betragtning, og derfor er den væsentlige nyhed i de fremkomne data fra pulseksponeringerne, at der kan opstå reproduktionsskabder, selv når dyrene udsættes for pirimicarb i meget kort tid. 5.2.3 ChlorpyrifosUmiddelbart efter endt pulseksponering af Daphnia magna med chlorpyrifos var effekten på mobiliteten mindre markant end efter, at dyrene havde opholdt sig 24 timer i rent medie (PE0t-IT50>PE24t-IT50). Dette fortolkes som et forsinket tab af mobilitet, også selvom der ikke var øget immobilisering efter yderligere 24 timers ophold i rent medie (PE24t-IT50~PE48t-IT50). Kun til tiderne 24 timer og 48 timer er der tale om en koncentrationsafhængig forskel i responset, hvilket afspejles i en lavere PEt-IT50 værdi for 3 μg/l sammenlignet med 2,5 μg/l. Van der Hoeven & Gerritsen (1997) undersøgte evnen til recovery hos Daphnia pulex efter eksponeringen (1-3 dage) for chlorpyrifos. Resultatet af deres forsøg var, at et recovery næsten aldrig indtraf, og at dyrene ofte blev immobile efter eksponeringens ophør også selv om de var mobile til tiden t = 0 timer. Altså et mønster, der svarer til det, der i nærværende undersøgelse blev set efter pulseksponeringer. Van der Hoeven & Gerritsen (1997) fandt ingen effekter (hverken positive eller negative) efter kontinuerte eksponeringer for en koncentration på 0,5 μg/l. I modsætning til dette fandt Naddy & Klaine (2001) et recovery på >80%, op til 7 dage efter eksponeringen af Daphnia magna for en puls af chlorpyrifos (0,5 μg/l). Proceduren anvendt i nærværende undersøgelse ligner den anvendt af Naddy & Klaine (2001), dog blev chlorpyrifos testet i koncentrationer, der er 5-6 gange lavere. For chlorpyrifos viser de beskrevne studier sammenholdt med vore undersøgelser således, at det kun er muligt for dafnier at genvinde mobiliteten efter pulseksponeringer for chlorpyrifos under en vis koncentration, og at højere koncentrationer kan give anledning til forsinkede negative effekter. I lighed med undersøgelser udført af Naddy & Klaine (2001) blev der heller ikke i nærværende undersøgelse påvist effekter på de undersøgte parametre for reproduktionseffekter som følge af pulseksponering med chlorpyrifos, men det er værd at bemærke, at der ved kontinuert eksponering i 21 dage er fundet en NOEC på 0,052 μg/l (se Tabel 1). 5.2.4 EsfenvaleratEsfenvalerat er et neurotoksin og virker som beskrevet i afsnit 2.2.2 ved at forlænge åbningen af Na+ kanaler i nervecellen så der opstår adskillige aktionspotentialer. Vi så også, at immobiliseringen blandt dafnierne viste sig i form kramper. IC50,24t og IC50,48t for esfenvalerat blev beregnet til 1,58 μg/l og 0,70 μg/l. Værdien på 0,24 μg/l opgivet i Pesticide Manual for 48 timer (Tomlin, 2003) ligger indenfor det beregnede konfidensinterval [0,07; 1,53] for vores test, hvilket også er tilfældet for de 0,9 μg/l, der fremgår af Tabel 1. Pulstestene med esfenvalerat (20 μg/l og 30 μg/l) viser en lille restituering i løbet af de første 24 timer af observationsperioden. Herefter forringes dyrenes tilstand noget (PEt-IT50-værdien falder), men der er stadig flere mobile dyr 48 timer efter pulseksponeringens ophør, end der var til t = 0 timer. Under de indledende reproduktionstest med esfenvalerat (1 μg/l, 5 μg/l og 20 μg/l) blev der imidlertid observeret en så markant grad af immobilisering og dødelighed i løbet af de første 4-6 dage, at forsøgene måtte afsluttes, da de valgte koncentrationer tilsyneladende var for høje til, at en test af 21 dages varighed kunne gennemføres. Beregning af PEt -IT50-værdier for eksponeringen for 1 μg/l og 5 μg/l viser ligesom de indledende tests med 20 μg/l og 30 μg/l en lille forbedring i mobiliteten i løbet af de første 24 timer efterfulgt af et fald fra tiden t = 24 til t = 48 timer (Bilag 6.1.4), hvilket afspejles ved en stigende PEt-IT50-værdi fra tiden t =0 til tiden t = 24 timer, der så igen falder til tiden 48 timer. Kun værdierne for 5 μg/l adskiller sig fra værdierne for 20 μg/l og 30 μg/l (på nær til tiden t = 24t for 30 μg/l). Der kunne ikke konstateres sammenhæng mellem koncentration og PEt-IT50-værdien. Nørum & Bjerregaard (2003) testede effekten af pyrethroiderne esfenvalerat og cypermethrin på mobiliteten hos Gammarus pulex og Asellus aquaticus. De så at immobilisering indtraf tidligere og tidligere ved eksponeringen af Gammarus pulex for koncentrationer der var ≥1 μg/l og Asellus aquaticus for koncentrationer ≥0,3 μg/l. Tilsvarende fandt Cold & Forbes (2004) en markant stigning i dødeligheden hos Gammarus pulex efter overførslen til rent vand, selv om der var 100% overlevelse umiddelbart efter pulseksponeringer for esfenvalerat på op til 2 μg/l. I vore forsøg blev set en lille stigning i mobiliteten (Tabel 5) efter overførslen til rent vand fra en eksponeringskoncentration, der var over en faktor 10 højere end koncentrationen Cold & Forbes anvendte. Vi fandt dog ingen markant dødelighed efter 48 timer i rent vand (pulstest med 20 μg/l og 30 μg/l samt kontinuerte tests med 1 μg/l, 5 μg/l og 20 μg/l). Dette rejser spørgsmålet, om stoffet har været til stede i mediet i de valgte koncentrationer under pulstestene eller om f.eks. adsorption til testbeholderne har reduceret den reelle eksponeringskoncentration. Af økonomiske årsager har det ikke har været muligt at foretage kemiske analyser i de udførte test, men tages esfenvalerats fysisk-kemiske data i betragtning (især den høje Kow-værdi) er sorption til glasvægge en realistisk fjernelsesproces (jf. afsnit 2.3). I den anvendte testprocedure defineres graden af immobilitet ikke, og der skelnes derfor ikke mellem dyr, der har en nedsat bevægelse sammenlignet med kontrollen og dyr der tydeligvis er ved at dø. Som indikeret af reproduktionstestene er det derfor muligt, at dyrene eksponeret for 20 μg/l og 30 μg/l ville have udvist en markant større dødelighed, hvis posteksponeringsperioden var blevet udvidet i forhold til de 48 timer. Endelig skal det nævnes, at forskellen mellem de her beskrevene tests med Daphnia magna og forsøgene udført af Nørum & Bjerregaard (2003) & Cold & Forbes (2004) med Gammarus pulex og Asellus aquaticus kan skyldes, at arterne har forskellig følsomhed overfor stoffet (jf. f.eks. Stephenson, 1982; Nørum & Bjerregaard 2003). Gentagne pulseksponeringer med esfenvalerat viste større effekt på mobiliteten hos dafnier end en enkelt puls (t = 0 timer). Ligeledes kunne markante effekter ses ved lavere eksponeringstider end efter en enkelt puls. Efter 24 timers observation er den samlede effekt stort set uændret. Èn markant forskel er dog ændringen i andelen af dyr, der går fra immobilisering til død. Ud fra resultaterne nævnt i overstående, kan vi imidlertid ikke udelukke, at vi ville have set en tilsvarende effekt med tiden hvis observationsperioden, efter den første puls (pulstest med 20 μg/l og 30μg/l), var blevet udvidet. Som tidligere nævnt var der generelt stor dødelighed under tests for kroniske effekter af esfenvalerat, og flere test blev stoppet inden udløbet af de 21 dage p.g.a. for stor dødelighed (Bilag 6.1.4). Testen med en pulseksponering på 0,1 μg/l, var den eneste test der forløb over alle 21 dage, og selv i denne test blev OECD-standardens krav om maksimal 20% døde i kontrolgruppen ikke opfyldt (Bilag 6). På baggrund af resultater opnået med de overlevende moderdyr, kan det dog konkluderes, at der ikke blev fundet effekter på antallet unger, størrelse eller vægt efter 21 dage, men det skal understreges, at der kræves yderligere undersøgelser inden endegyldige konklusioner vil kunne drages. I test med kontinuert eksponering er NOEC fundet til 0,052 μg/l for 21-d testen med Daphnia magna (Tabel 1). Kroniske effekter af pulseksponeringer for esfenvalerat er tidligere blevet fundet ved test med krebsdyr. Cold & Forbes (2004) eksponerede Gammarus pulex indenfor koncentrationsintervallet 0,1-0,6 μg/l (1 time), og så effekter på overlevelse, pardannelse og reproduktion. Disse effekter kunne ses mindst to uger efter pulsens ophør. Barry et al. (1995) undersøgte effekten af pulseksponeringer for esfenvalerat på regnbuefisken Melanotaenia fluviatilis, og fandt en negativ korrelation mellem koncentration og fekunditet og det totale antal yngel. 5.2.5 PyrazophosBaseret på en IC50,24t-værdi på 0,62 μg/l opnået i ISO-test med kontinuert eksponering blev pyrazophos fundet at være det mest toksiske af alle testede stoffer. I Tabel 1 er opgive en værdi på 0,36 μg/l, som ligger inden for 95%’s konfidensintervallet for de opnåede resultater i dette projekt I pulstest med immobilisering som endpoint udviste pyrazophos en forsinket negativ respons fra tiden t = 0 timer til tiden t = 24 timer efter eksponeringer for 10 μg/l eller 20 μg/l. Den grafiske fremstilling viser også tendenser til forsinkede negative effekter ved koncentrationerne 2,5 μg/l, 5 μg/l og 7,5 μg/l. Generelt blev der kun set effekter efter eksponeringer for pulse af 2-6 timers varighed, men i løbet af posteksponeringperioden ses det, at mobiliteten forringes hos dafnier, der har modtaget korterevarende pulse. Der er derfor også tale om en effekt på længere sigt hos grupper, der ikke udviste respons umiddelbart efter eksponeringens ophør. Udsættes dyrene for endnu en puls af samme koncentration og varighed forøges responset betydeligt også i grupper, der ikke blev påvirket af den indledende puls (1-2t). Ligeledes stiger responset med tiden, og efter 48 timer skyldes størsteparten af responset dødelighed. Key & Fulton (1993) viste, at gentagne pulse af chlorpyrifos, der ligesom pyrazophos er en acetylcholinesterasehæmmer, øger toksiteten af pesticidet. Tilsvarende undersøgte Naddy & Klaine (2001) effekten af gentagne chlorpyrifos-eksponeringer, og fandt, at gentagne pulse er mere toksiske end en enkelt puls. Naddy & Klaine (2001) fandt for chlorpyrifos, at dyrene genvandt mobiliteten mellem to pulse, hvilket er et andet mønster end hvad vi observerede for pyrazophos, til trods for at de pesticider formodes at have samme virkningsmåde på Daphnia magna. 5.2.6 AzoxystrobinDe fundne IC50-værdier for 24 og 48 timer for azoxystrobin stemmer godt overens med værdierne fundet af Lauridsen et al. (2003) ligesom der også er en rimelig overensstemmelse med værdierne angivet i Tabel 1. Pulseksponeringer for azoxystrobin (10,5-12,5 μg/l) viste kun en effekt ved pulse af 4-6 timers varighed. De beregnede PEt-IT50-værdier viste hverken tegn på restituering eller en forsinket negativ effekt efter pulseksponeringer for azoxystrobin i løbet af den undersøgte observationsperiode. Dog er effekten af eksponeringen hovedsagelig i form af dødelighed. Vi ser derfor at de dyr, der har kunnet klare eksponeringen for azoxystrobin, ikke får en forsinket effekt, hvorimod dyrene, der har været mere sensitive dør straks. Som omtalt i afsnit 2.2.5 virker azoxystrobin ved at hæmme den mitrochondrielle respiration, hvorved energidannelsen i cellen hæmmes og organismen dør. Dette er ligeledes i overenstemmelse med de her opnåede resultater. Nørum & Bjerregaard (2003) undersøgte effekten af pulseksponeringer med azoxystrobin på bevægelsesadfæren hos Gammarus pulex. Under eksponeringen af dyrene for en azoxystrobin-koncentration på 100 μg/l blev der ikke observeret nogen adfærdsændringer. Adfærdsforsøg udført af Lauridsen et al. (2003) viste, at aktiviteten af hjerte, brystlemmer og mandibler hos ægbærende Daphnia magna (8 ± 1 dag) blev reduceret ved azoxystrobinkoncentrationer på 0,5 mg/l (24 timers eksponering). Tilsvarende blev svømmeaktiviteten reduceret med ca. 15% efter eksponering for 1,0 mg/l. Hverken Nørum & Bjerregaard (2003) eller Lauridsen et al. (2003) har undersøgt restituering efter pesticideksponeringerne. 5.2.7 DiquatIC50,24t-værdien for diquat blev fundet til 23,9 μg/l, hvilket er klart lavere end værdien opgivet i Tabel 1. Det var ikke muligt at beregne en tilsvarende værdi for 48 timer, da der var 100% respons i alle grupper på nær den laveste testkoncentration. Ud fra de beregnede PEt-IT50-værdier alene er det svært at drage entydige konklusioner. Værdierne peger dog i retning af en forsinket negativ effekt. Dette underbygges af den grafiske fremstilling (46-64 μg/l), der viser en markant forsinket effekt i grupper, der har modtaget en puls af 6 timers varighed, hvor vi efter endt eksponering allerede har en effekt på henholdsvis 30%, 80% og 75% (immobilitet) og hvor responset når op på 100% (immobilitet og dødelighed) efter 48 timer. Ligeledes ses der en effekt efter 48 timer i grupper, der har modtaget en puls af 2-4 timers varighed. Der er ikke fundet tilsvarende undersøgelser med diquat i litteraturen, men det skal understreges, at de ovenfor beskrevne effekter optræder ved koncentrationer, der ligger en faktor 5-10 under de effektkoncentrationer (både akutte og kroniske), der opgivet i litteraturen for test med kontinuert eksponering (se Tabel 1). 5.2.8 BromoxynilVed vore test med kontinuert eksponering er bromoxynil fundet at være lidt mindre akut toksisk end opgivet i litteraturen (IC50,24t =18,9 mg/l mod IC50,24t=12,5 mg/l opgivet i Tabel 1), men værdier er i samme størrelsesorden. Pulsforsøg udført med bromoxynil viser et recovery efter eksponeringen for 26 mg/l, hvis vurderingen udelukkende baseres på de fundne PEt-IT50-værdier (Tabel 8). Figur 18 viser imidlertid også, at der i løbet af de 48 timer efter eksponeringens ophør, sker et skift således, at dyr, der initielt er immobile, dør, i gruppen eksponeret for en 6-timers puls. Om dette skift skal kaldes en forsinket negativ respons kan diskuteres, da immobilisering sandsynligvis også ville gøre, at dyrene ikke er funktionsdygtige i vandmiljøet. På den anden side set bør letale effekter beskrives som forsinkede negative effekter, da restitution – i sagens natur – ikke længere er mulig. Resultaterne tyder derfor på, at der ved pulse af bromoxynil af kortere varighed end 6 timer er tale restitution, mens længerevarende pulse fører til forsinkede negative effekter. Buhl et al.(1993) viste, at Daphnia magna, der var blevet immobiliserede af eksponering for op til 13 mg/l bromoxynil octanoate, kunne genvinde deres mobilitet i løbet af 24 timer, hvis eksponeringen var af 1,5-6 timers varighed. Enkelte af dyrene, der fik en eksponering af 6 timers varighed og 7,8-13 mg/l i disse forsøg var immobile efter 48 timer, og disse resultater er altså samstemmede med, hvad der blev fundet i vore undersøgelser. Dyr udsat for pulse af bromoxynil på 20 mg/l af 0,5-6 timer varighed viste ikke tegn på skader på reproduktionen, mens der for test med kontinuert eksponering er rapporteret en NOEC efter 21 dage på 3,1 mg/l (se Tabel 1). 5.2.9 4-nitrophenol og 3,5-dichlorophenolBaseret på PEt-IT50-værdierne fundet ved pulstest ses det, at dafnier udsat for 4-nitrophenol genvandt mobiliteten efter pulseksponering. Restitutionen er kun tydelig mellem tiderne t = 0 timer og t = 24 timer og bliver mindre udpræget ved stigende koncentrationer. Responset for 4-nitrophenol ændres dog fra immobilitet (t = 0) til dødelighed med tiden (t = 48 t), hvilket også blev fundet efter eksponeringerne for bromoxynil. For 3,5-dichlorophenol ses tydeligt, at dyrene genvandt mobiliteten efter overførsel til rent vand. Dette gælder for alle testede koncentrationer på nær for den højeste koncentration (3,3 mg/l), hvor det ikke var muligt, at beregne en PEt-IT50-værdi til tiden t = 0 timer. Igen sker restitutionen mellem tiderne t = 0 timer og t = 24 timer efter pulsens ophør. 5.3 Perspektiver og anbefalinger i relation til risikovurdering og godkendelse af pesticider5.3.1 Effekter af pulseksponeringer – et nyt internationalt forskningsområdeStadigt flere artikler i den videnskabelige litteratur diskuterer problemstillingen omkring vurderingen af episodiske pulseksponeringer i det akvatiske miljø, hvor koncentrationen af et eller flere stoffer stiger til et højt niveau i en kortere tidsperiode (Reinert et al., 2002). Spørgsmålet er, hvor stor betydning disse eksponeringer skal tildeles, og om viden fra eksisterende toksicitetstest kan anvendes, når effekten af episodiske eksponeringer skal vurderes. De undersøgelser, der er publiceret i de seneste år har ofte udgangspunkt i, at eksisterende toksicitetstest ikke er tilstrækkelige til vurdering af effekter af pulseksponering, da disse test udelukkende udføres med kontinuerte eksponeringer og ikke tager højde for at eksponeringen i miljøet ikke nødvendig vis er kontinuert (Brent & Herricks, 1999; Naddy & Klaine, 2001). I nogle få af undersøgelserne på akvatiske organismer er der blevet set på langtidseffekter af kortvarigt høje eksponeringer, d.v.s. der er blevet indbygget en observationsperiode, hvor det er blevet testet om dyrene får det bedre eller værre med tiden. Varigheden af disse post-eksponeringsperioder med observation afhænger typisk af den anvendte testorganisme. Inddragelsen af sådanne observationsperiode har, som også vist i nærværende undersøgelse, medført, at effekter, der ikke umiddelbart er synlige efter eksponeringens ophør, er blevet detekteret (Naddy et al. 2000). Ligeledes er der blevet set på kroniske effekter og reproduktionsparametre efter eksponeringen for en puls af et kemikalie (Brent & Herricks, 1999; Schultz & Liess, 2000; Cold & Forbes, 2004; Andersen et al., 2005). 5.3.2 Pulsvarighed og - koncentrationSom nævnt i indledningen har nærværende projekt i højere grad fokuseret på effekterne af pulsvarighed end på effekter af koncentrationer. Dermed har det været ønskeligt, at de anvendte eksponeringstider også har relevans i forhold til eksponeringssituationen i danske vandløb og søer. At bestemme realistiske pulsvarigheder er imidlertid vanskeliggjort af, at kun ganske få undersøgelser af pulshændelser er beskrevet i litteraturen, men også af, at pesticidernes afstrømningsmønstre afhænger af såvel stoffernes fysisk-kemsike egenskaber som deres anvendelse. Ud fra de pulsvarigheder, der er trods alt er beskrevet i litteraturen, er pulsvarighederne anvendt i nærværende projekt i en størrelseorden, der kan være realistisk for danske forhold. Styczen et al. (2003) fandt således pulse af pesticider fra 4-6 timers og op til få dages varighed i vandløb efter regnvejrshændelser, og Slothuus et al. (2004) så pulse af ca. 3 timers varighed efter brugen af blåsten i dambrug. Situationen i søer og vandhuller ser imidlertid anderledes ud, da det typisk vil tage nogle dage fra pesticidet tilføres og til det er jævnt fordelt. I undersøgelsen af Styczen et al. (2003) blev det fundet, at der herefter gik omkring 2 uger inden stoffet igen var ”forsvundet”. Som tidligere beskrevet er der i vandløb i Fyns, Århus og Nordjyllands Amt fundet pulskoncentrationer af pesticider mellem 0,01 μg/l og 10,0 μg/l (Styczen et al.,2003; Lauridsen & Wiggers 2001). Ikke alle de pesticider, der er anvendt i nærværende projekt, er fundet i ovennævnte undersøgelser. Blandt de fundne stoffer er dog dimethoat, pirimicarb, esfenvalerat og bromxynil, og for disse stoffer gælder det, at de fundet i lavere koncentrationer end anvendt i denne rapport (se afsnit 2.1). For nogle pesticider er det muligt, at pulse af kortere varighed (0,5-1 time) vil kunne udløse tilsvarende effekter som længerevarende eksponeringer for lavere koncentrationer. Ud fra de opnåede erfaringer i nærværende projekt var det dog kun i få tilfælde muligt at påvise effekter af pulsvarigheder <1 time. Ved fremtidig anvendelse af pulstest kan eksponeringen ved 0,5 time med rimelighed undlades. Især effekter af længerevarende pulse (flere timers varighed) påkalde sig opmærksomhed i forbindelse med godkendelse og risikovurderinger pesticider. Afhængigt af pesticidernes fysisk-kemiske egenskaber og anvendelsesmønstre vil der kunne forekomme ganske høje koncentrationer i recipienterne. Dette understøttes af modelberegninger foretaget med PestSurf, som simulerer eksponering i vandløb i hhv. et sandet og et leret opland, som belastes med pesticider (Styczen & Poulsen, 2005). Med undtagelse af bromoxynil er disse modelkørsler er ikke foretaget for de stoffer, som indgår i nærværede undersøgelse. I det følgende vil resultater af disse modelkørsler for bromoxynil, alpha-cypermethrin og tribenuron-methyl kort blive omtalt. For data vedr. bromoxynil henvises til Tabel 1. Alpha-cypermethrin er et lipofilt stof (logKow = 5.5), har en høj sorption (Koc = 57889) og en relativ lang halveringstid (DT50=103 d) (Styczen & Poulsen, 2005). I modsætning hertil er tribenuron-methyl hydrofilt (logKow=0.78), er mobilt (Koc = 22) og har en kort halveringstid (DT50= 9 d). (Styczen & Poulsen, 2005). Udover at disse to stoffer dækker et stort spektrum af fysisk-kemiske egenskaber, kan disse to stoffers opførsel i miljøet med rimelighed sammenlignes med hhv. esfenvalerat’s og dimethoat’s skæbne i miljøet, når stoffernes fysisk-kemiske data tages i betragtning. Tabel 17. Maksimumskoncentration i vandløb med hhv. leret og sandet opland beregnet i PestSurf samt anvendt dosering ved modelberegninger (Styczen & Poulsen, 2005). Sammenligneligt stof med tilhørende normal dosering er vist i sidste kolonne.
1Maksimumskoncentration beregnet i et tidsrum på 24 timer. I Tabel 17 er vist resultater fra PestSurf-modellen i form af koncentrationer i vandløb med hhv. leret og sandet opland med den angivne dosering. For bromoxynil er resultaterne naturligvis direkte anvendelige, for alpha-cypermethrin er den anvendte dosering i overensstemmelse med normal doseringen af esfenvalerat, mens der for tribenuron-methyl er regnet på en dosering, der er 20 gange mindre end normaldoseringen af dimethoat. Herbicidet bromoxynil findes således i koncentrationer, der er mere end 106 gange lavere end de koncentrationer, der er undersøgt i pulstestene, og pulseksponering af krebsdyr med bromoxynil forventes derfor ikke at give anledning til effekter på mobiliteten af krebsdyr i vandmiljøet. For insekticiderne dimethoat og esfenvalerat var det overordnede billede efter pulstest, at dyrene genvandt mobiliteten, men for eksponeringer af 6 timers varighed med 20 mg/l dimethoat var 40% af dyrene immobile (Figur 21) og 90% immobile efter eksponering for 20 μg/l esfenvalerat (Figur 14). De anvendte koncentrationer ved pulstestene er i størrelsesordenen 104 højere end de i Tabel 17 angivne (efter korrektion for dosering af dimethoat). Dette vil normalt blive betragtet som en tilfredsstillende sikkerhedsmargin (margin of saftety, MOS). I denne forbindelse skal observationstiden efter pulseksponeringen i dafnietestene dog inddrages (se afsnit 5.3.3) ligesom også resultater opnået ved forsøg med gentagne pulse bør tillægges stor vægt. Ved disse forsøg blev genvinding af mobiliteten ikke set ved overførsel til rent vand efter anden puls, og der var en udpræget tendens til øget dødelighed i grupper udsat for pulseeksponeringer af mere end 2 timers varighed. Dette mønster blev set for såvel dimethoat (Figur 21) og esfenvalerat (Figur 23). At gentagne pulse forekommer, er også illustreret modelberegninger gennemført i PestSurf af Styczen & Poulsen (2005), men ”pulsadfærden” af et stof afhænger mange parametre såsom oplandets karakteristika, regnhændelsernes varighed og intensitet og stoffernes fysisk-kemiske egenskaber. Det ligger udenfor dette projekts rammer at beskrive disse sammenhænge, men de opnåede resultater understreger, at gentagne pulseksponeringer vil have en markant større effekt end de enkelt-stående pulse. Den ovenfor angivne sikkerhedsmargin for dimethoat og esfenvalerat vil derfor blive mindre, hvis gentagne pulse tages i betragtning. Der mangler dog yderligere undersøgelser af effekten af gentagne pulse for kunne kvantificere, hvor meget sikkerhedsmarginen reduceres. 5.3.3 Observationstid efter pulseksponeringVed vurdering af forsinkede effekter efter pulseksponeringens ophør er det dog nødvendigt at evaluere de anvendte observationstider. I de her udførte test er anvendt en observationsperiode på 48 timer i forbindelse med vurdering af kortidseffekter. For mest lipofile stoffer (esfenvalerat og chlorpyrifos) kan længden af observationsperioden have været for kort til at se evt. forsinkede effekter af pulseksponeringen. Yderligere undersøgelser med fokus på relationer mellem fysisk-kemiske stofegenskaber og effekter af pulseksponering er derfor påkrævede (se også afsnit 5.3.2). I den forbindelse vil det være et krav, at de biologiske observationer følges med koncentrationsmålinger. De stoffer, for hvilke observationsperiodens længde er af afgørende betydning, er nemlig også er de stoffer, som er de vanskeligste at håndtere i laboratoriet (pga. høj fedtopløselighed og stor sorption til alle overflader). I den forbindelse kan anvendelsen af radioaktive isotoper være særdeles fordelagtig, da en komplet redegørelses for stoffernes opførsel før, under og efter eksponering er ønskelig. 5.3.4 Risiko for effekter som følge af pulseksponeringI forbindelse med vurdering af de anvendte koncentrationers ”miljørealisme”, skal det understreges, at testudviklingen og den gennemførte testning i nærværende projekt har været rettet mod regulatorisk anvendelse af resultater fra toksicitetstest snarere end simulering af miljørealistiske koncentrationer. Som ved den øvrige regulatoriske testning anvendt ved risikovurdering af kemiske stoffer (standardtest) er formålet med nærværende undersøgelser ikke i sig selv at beskytte dafnier under naturlige forhold. Formålet med standardtest er at levere data, der kan bruges ved vurdering af, hvordan økosystemets arter og dets struktur kan beskyttes. I denne sammenhæng anvendes dafnier som testorganismer ud fra de (overvejende praktiske) hensyn, der er beskrevet i afsnit 2.4. Ved den videre anvendelse af resultater opnået ved pulstest med såvel akutte som kroniske endpoints skal data derfor kunne passes ind i den nugældende risikovurderingspraksis. Dette vil typisk indebære, at der anvendes sikkerhedsfaktorer (også kaldet usikkerhedsfaktorer eller applikationsfaktorer) for at ekstrapolere fra de anvendte laboratorietest og testorganismer til vandmiljøet og dets fauna. Disse sikkerhedsfaktorer kan være i størrelsesordenen 10-1000 i henhold til nationale vurderingsrammer og harmoniserede principper i pesticiddirektivet. Tages dette forhold i betragtning, vil flere af de anvendte eksponeringskoncentrationer ligge inden for, hvad der kan betragtes som miljørealistiske koncentrationer i pulse. Hvorvidt anvendelse af de værdier, der fremkommer ved standardtest med kontinuert eksponering, er tilstrækkeligt til at beskytte krebsdyr i vandmiljøet mod effekter af pulseksponeringer, er stadig et åbent spørgsmål. Ved sammenligning mellem de anvendte puls koncentrationer og effektkoncentrationer opnået ved standardtest vil sidstnævnte ligge klart lavest. Hermed er det nærliggende at konkludere, at anvendelse af data fra test med kontinuert eksponering vil være det mest konservative mht. at beskytte mod effekter i vandmiljøet. I den forbindelse skal det dog understreges, at de otte pesticider og to referencestoffer, der indgik i nærværende undersøgelse, udviste forskellige responsmønstre, som ikke umiddelbart kan relateres til stoffernes virkemekanismer. Desuden har undersøgelsens fokus, af økonomiske årsager, ligget på effekten af eksponeringens varighed. Dette forhold begrænser mulighederne for at konkludere mere håndfast på effekter af varierende koncentrationer. Derfor er det vanskeligt på baggrund af nærværende undersøgelser at konkludere om fx. PNEC-værdier (predicted no-effect concentration) fastsat på baggrund af standard test (med kontinuert eksponering) er over- eller underbeskyttende i forbindelse vurdering af pulspåvirkninger af vandmiljøet. Et meget væsentligt nyt resultat af undersøgelserne er dog, at det ikke er korrekt at antage, at der kun kan forventes akutte virkninger af pulseksponeringer. Kroniske effekter blev konstateret i pulstest med to af de undersøgte pesticider (dimethoat og pirimicarb) – endda ved pulse af kun 3 timers varighed. Hermed tilvejebringer pulstestene ny information, der ikke opnås ved de traditionelt anvendte standardtest. 5.3.5 Godkendelse og pulseksponeringUndersøgelsen viser desuden, at der efter eksponeringens ophør kan indtræde forsinkede negative effekter, hvor dyrene enten mister deres mobilitet eller dør, men også at der kan forekomme restitution med tiden. Dette er af stor betydning for godkendelse og risikovurdering af pesticider, såfremt man ønsker at inddrage effekter af pulseksponeringer. Umiddelbart vil man måske vurdere pulseksponeringer, som mindre problematiske end kontinuerte eksponeringer, idet organismernes eksponering i sagens natur er af kortere varighed. For de pesticider, der udviser forsinkede negative effekter efter en pulseksponering, er det forfatternes holdning, at det ikke vil være forsvarligt alene at anvende den korte varighed af pulseksponeringen som argument for at lempe på farlighedsvurderinger af pesticiderne. Dyrene er jo blevet svækket så meget af den kortvarigt høje eksponering, at effekter på mobiliteten indtræder selvom dyrene overføres til rent vand, fodres og får optimale livsbetingelser. Ud fra de gennemførte undersøgelser med pesticider, hvor genvinding af mobiliteten blev påvist efter overførsel til rent vand, er det ikke forsvarligt, på nuværende tidspunkt, at lade pulseksponeringernse korte varighed indgå som argumentation for at lempe på farlighedsvurderinger af pesticider. 5.3.6 Gentagne pulseDe pesticider, hvor pulsforsøg har vist, at eksponerede organismer kommer sig efter eksponeringen, kunne muligvis blive anset som mindre problematiske, hvis det vurderes, at eksponeringer hovedsagelig sker i form af pulse. Dette kunne gælde selv for stoffer, som har høj toksicitet vurderet på baggrund af standardtest med kontinuert eksponering (fx. pirimicarb og pyrazophos). Genvinding af mobiliteten efter pulseksponeringens ophør blev vist for seks af de undersøgte pesticider, men for to af disse seks blev der efterfølgende set alvorlige effekter på reproduktionen. For disse stoffer (dimethoat og pirimicarb) er det altså tydeligt, at den ”tilsyneladende restitution” (genvinding af mobilitet efter 24-48 timer i rent vand) dækker over, at dyrene er blevet så svækkede af pulseksponeringen, at det senere får betydning for reproduktionen. Den ”tilsyneladende restitution” kom også til udtryk ved de udførte forsøg med gentagne pulse af dimethoat, pirimicarb, pyrazophos og esfenvalerat. For tre af de fire stoffer havde den gentagne pulseksponering en effekt, der var mere markant end effekten af den indledende puls også selv om der sås et recovery i løbet af de 48 timer, der fulgte efter den indledende puls. Effekter som følge af gentagen pulseeksponering er et område, hvor manglen på viden er åbenlys, selvom der næppe hersker tvivl om, at netop er gentagen pulseksponering er meget sandsynlig i vandmiljøet.Vi står derfor overfor følgende spørgsmål, som yderligere test sandsynligvis ville kunne belyse: 1) I hvilken grad kommer dyrene sig efter den indledende eksponering? 2) Kunne effekten af eksponeringen være opstået uden den gentagne puls f.eks. som følge af en aldersbetinget følsomhed? 3) Ville samme mønster blive observeret, hvis intervallet mellem pulsene var blevet udvidet? På baggrund af nærværende projekt er anbefalingen derfor, at der igangsættes videregående undersøgelser af indflydelsen af gentagne pulseksponeringer med pesticider. 6 KonklusionHændelser som regnskyl, afstrømning fra marker eller udsprøjtning af pesticider kan føre til pulseksponering af vandlevende organismer i naturen. De økotoksikologiske test, der traditionelt anvendes i forbindelse med godkendelse og risikovurdering af pesticider, omfatter ikke effekter af pulse. Formålet med projektet var derfor at udvikle og afprøve en ny laboratoriebaseret testmetode til undersøgelse af toksiske effekter som følge af pulseksponeringer af ferskvandskrebsdyret Daphnia magna. Dette har ført til udarbejdelsen af en testprotokol, som kan anvendes i dens nuværende form eller som kan danne grundlag for en evt. standardisering af test til vurdering af pulseksponering. Generelt vurderes det, at de anvendte tests er lette at udføre indenfor en overskuelig tidsramme (3 dage for akut virkning og 21 dage for kronisk virkning), der anvendes kun få materialer ved testene, og Daphnia magna har været en velegnet testorganisme. Testene tager udgangspunkt i ISO/OECD-metoder med kontinuert eksponering, og denne opkobling til standardtest vurderes dels at give et effektivt testdesign og dels at være nødvendig for, at pulstest kan inkluderes i risikovurderinger af pesticider. Foruden akutte toksiciteststest til fastsættelse af immobilseringskoncentrationer (IC-værdier), blev de udviklede pulseksponeringstest anvendt til 3 typer forsøg: 1. Vurdering af effekter af enkeltstående pulseksponeringer for otte pesticider (Dimethoat, Pirimicarb, Chlorpyrifos, Esfenvalerat, Azoxystrobin, Pyrazophos, Diquat) og to referencestoffer (4-nitrophenol, 3,5-dichlorophenol). Der blev set på effekter på mobiliteten hos Daphnia magna umiddelbart efter pulseksponeringens ophør samt dyrenes evne til at restituere i løbet af en 48 timers posteksponeringsperiode. 2. Effekten af gentagne pulseksponeringer på mobiliteten. Disse forsøg blev gennemført for stofferne: Dimethoat, pirimicarb, esfenvalerat og pyrazophos. 3. Undersøgelse af langtidseffekter efter pulseksponeringen (størrelse, vægt og reproduktionsparametre). Disse test omfattede stofferne: Dimethoat, pirimicarb, esfenvalerat, chlorpyrifos, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol. Ved alle pulstest blev anvendt eksponeringstider (pulsvarigheder) fra 0,5-6 timer, og stofferne blev undersøgt i 2-5 forskellige koncentrationer valgt på baggrund af resultater opnået i standardtest med immobilisering som effektparameter. De indledende akutte toksicitetstest viste, at pyrazophos var det mest toksiske af de testede pesticider, med en IC50,24t-værdi på 0,62 μg/l, efterfulgt af esfenvalerat og chlorpyrifos. Referencestoffet 4-nitrophenol og herbicidet bromoxynil udviste mindst akut toksicitet med IC50,24t-værdier på hhv. 12,9 mg/l og 18,9 mg/l. Forsøgene med pulseksponering af Daphnia magna viste, at der efter eksponeringen for chlorpyrifos, pyrazophos og diquat var forsinkede negative effekter, hvor dyrene således mistede deres mobilitet eller døde efter at pulsen ophørte (ved overførsel til rent vand). For dimethoat, pirimicarb, esfenvalerat, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol blev det fundet, at dyrene genvandt deres mobilitet efter 24-48 timers ophold i rent vand. Der var hverken en forsinket negativ effekt eller tegn på restitution efter eksponeringen for azoxystrobin i løbet af en observationsperiode på 48 timer. Gentagne pulseksponeringer viste en mere markant effekt på mobiliteten hos Daphnia magna end enkeltstående eksponeringer. Selv grupper, der havde fået en kortvarende pulseksponering, og som tilsyneladende ikke blev påvirket af den indledende puls, viste ændringer i mobiliteten efter endnu en pulseksponering. Gentagne eksponeringer viste ligeledes et skift i responset, således at dyrene gik fra ikke at bevæge sig (immobile) efter første puls til at dø efter anden puls. Efter pulseksponering for dimethoat og pirimicarb blev der registreret en reduceret størrelse af moderdyerne. Hos de samme dyr blev der registreret både en forsinkelse og et samlet fald i antal unger produceret i løbet af 21 dage efter at eksponering ophørte. Antallet af unger faldt signifikant jo længere pulsen varede, men selv for pulse af 2 timers varighed kunne signifikante effekter observeres. Der blev fundet en signifikant sammenhæng mellem dyrenes længde efter 21 dage og antallet af unger. Det er muligt, at den nedsatte størrelse skyldes et mindre optag af føde efter pulsenes ophør, og at dette samlet set medfører en mindre produktion af unger, og men dette er ikke blevet undersøgt nærmere i dette projekt. I de udførte test med pesticiderne esfenvalerat, chlorpyrifos, bromoxynil, 4-nitrophenol eller 3,5-dichlorophenol blev der ikke set signifikante effekter på reproduktion efter pulseksponeringer, selvom dyr udsat for 3,5-dichlorophenol var mindre ved testens ophør (21 dage efter eksponering) end ikke-eksponerede dyr. På baggrund af de udførte test er konklusionen, at den type data, der er nødvendige for at kunne afgøre om forsinkede effekter af kortvarige eksponeringer indtræder, ikke opnås ved de test, der i dag anvendes ved risikovurdering af pesticider. De fundne effekter på reproduktion efter enkeltstående pulseksponeringer og effekterne af gentagne pulse er nemlig, så vidt vides, ikke tidligere er beskrevet i litteraturen. Resultaterne viser desuden, at ”overraskelser” findes selv for stoffer med virkemekanismer, hvor man, ud fra teoretiske overvejelser, ikke ville forvente forsinkede negative effekter (jf. den reversible virkning af acetylcholinesterase-hæmmere). 7 LitteraturAbel, P.D. (1980). Toxicity of g-hexachlorocyclohexane (lindane) to Gammarus pulex: Mortality in relation to concentration and duration of exposure. Freshwater Biology, 10, 251-259. Abel, P.D. & Gardner, S.M. (1986). Comparisons of median survival times and median lethal exposure times for Gammarus pulex exposed to cadmium, permethrin, and cyanide. Water Research, 20, 579-582. Andersen, T. H., Tjørnhøj, R., Wollenberger, L., Slothuus, T. & Baun, A. (2005). Acute and Chronic Effects of Pulse Exposure of Daphnia magna to dimethoate and pirimicarb. Submitted paper. Barry, M.J., Logan, D.C., Ahokas, J.T. & Holdway, D.A. (1995). Effects of algal food concentration on Toxicity of two agricultural pesticides on Daphnia carinata. Toxicology and Environmental Safety 32, 273-279. Beusen, J.-M. & Neven, B. (1989). Toxicity of dimethoate to Daphnia magna and freshwaterfish. Bulletin of Environmental Contamination and Toxicology 42, 126-133. Brent, R.N. & Herricks, E.E. (1998). Postexposure effects of brief cadmium, zinc and phenol exposures on freshwater organisms. Environmental Toxicology and Chemistry 17, 2091-2099. Brent, R.N. & Herricks E.E. (1999). A method for the toxicity assesment of wet weather events. Water Research 33[10], 2255-2264. Buhl, K.J., Hamilton, S.J. & Schmulbach, J.C. (1992). Acute toxicity of the herbicide bromoxynil to Daphnia magna. Environmental Toxicology and Chemistry 12, 1455-1468. Buhl, K.J., Hamilton, S.J. & Schmulbach, J.C. (1993). Chronic Toxicity of the bromoxynil formulation buctril to Daphnia magna exposed continuously and intermittently. Arch.Environ.Contam.Toxicol. 25, 152-159. Canton, J. H. Wegman, R. C. C, Van Oers, A., Tammer, A. H. M. Mathijssen-Spiekman, E. A. M & Vand der Broek, H. H. (1980). Environmental Toxicology Research with Dimethoate and Omethoate. Rep. no. 121/80. Nat. Inst. Public Health Environ. Hyg.: 6p (DUT) Reference number: 5180 (http://www.pesticideinfo.org/Index.html) Chemfinder 2004 (http://chemfinder.cambridgesoft.com/) Cold, A. & Forbes V. (2004). Consequences of a short pulse of pesticides exposure for survival and reproduction of Gammarus pulex. Aquatic Toxicology 67, 287-299. Connell, D. W. (1997). Basic Concepts of Environmental Chemistry, Lewis Publishers. Cornell University 2004. Information om bromoxynils virkningsmekanisme. (www.css.cornell.edu/WeedEco/bromoxynil.html) (besøgt januar 2005) Danmarks Statistik 2004 (http://www.danmarksstatistik.dk/) (besøgt oktober 2004) Dansk standardiseringsråd (1980). Vandundersøgelse Tørstof og gløderest. 1-4. Eckert, R., Randall, D. & Augustine, G. (1996). Animal Physiology 3. ed. W. H. Freeman and Company. Friis-Nielsen, J. (2002). Økotoksikologiske effekter ved pulsudledninger til recipienter – en ny metode til simulering af pulsudledninger. Eksamensprojekt Miljø & Ressourcer DTU, Danmarks Tekniske Universitet, Lyngby. Hammer-Wirtz, M. & Ratte, H.T. (2000). Offspring fitness in daphnia: is the daphnia reproduction test appropriate for extrapolating effects on the population level? Environmental Toxicology and Chemistry 19[7], 1856-1866. Hermes, J., Canton, H., Steyger, N. & Wegman, R. (1984). Joint Effects of a Mixture of 14 Chemicals on Mortality and Inhibition of Reproduction of Daphnia magna, Aquatic Toxicology 5[4] 315-322 Herricks, E.E., Brent, R. Milne, I & Johnsen, I. (1997). Assesing the response of aquatic organisms to short-term exposures to urban run off. Management on Aquatic Ecosystems 112-128. Holdway, D.A., Barry, M.J., Logan, D.C., Robertson, D., Young, V. & Ahokas, J.T. (1993). Toxicity of pulse-exposed fenvalerate and esfenvalerate to larval Australian crimson-spottet rainbow fish (Melanotaenia fluviatilis). Aquatic Toxicology 28, 169-187. Hosmer, A.J., Warren, L.W., Ward, T.J. (1998). Chronic toxicity of pulse-dosed fenoxycarb to Daphnia magna exposed to environmentally realistic concentrations. Environmental Toxicology and Chemistry, 17, 1860-1866. ISO (1989a). Water quality- Determination of the inhibition of the mobility of Daphnia magna Straus (Cladocera, Crustacea). ISO 6341. International Organization for Standardization, Geneva, Switzerland. ISO (1989b). Water quality- Freshwater algal growth inhibition test with Scenedesmus subspicatus and Selenastrum capricornutum. ISO 8692. International Organization for Standardization, Geneva, Switzerland ISO (1996). Water quality- Determination of the inhibitory effect of water samples on the light of Vibrio fischeri (Luminecent bacteria test) – Part 3 Method using freeze dried bacteria. ISO/DIS 11348-3. International Organization for Standardization, Geneva, Switzerland Jarvinen, A.W., Tanner, D.K., Kline, E.R. (1988). Toxicity of chlorpyrifos, endrin, or fenvalerate to fathead minnows following episodic or continuous exposure. Ecotoxicology and Environmental Safety, 15, 78-95. Jespersen, Å. & Lûtzen, J. (1996). Zoologisk Morfologi- form og funktion i dyreriget. G.E.C gads Forlag 1. udg. 1996. Key, P. B. & Fulton, M.H. (1993). Lethal and sublethal effects of chlorpyrifos exposure on adult and larval stages of the grass shrimp, Palaemonetes pugio J. Environ. Sci. Health, B82(5), 621-640. Kommissionen, (2003). Technical Guidance Document in support of Commission Directive 93/67/EEC on risk assessment for new notified substances and Commission Regulation (EC) No. 1488/94 on risk assessment for existing substances. Brussels, Belgium. Kooijman, S.A.L.M & Bedaux, J.J.M. (1996). The analysis of aquatic toxicity data. VU University Press, Amsterdam, The Netherlands. Kusk, K.O. (1996). Bioavailability and effect of pririmicarb on Daphnia magna in a laboratory freshwater/sediment system. Arch.Environ.Contam.Toxicol. 31, 252-255. Lauridsen, T.L., Friberg-Jensen, U. & Christoffersen, K. (2003). Effekter af cypermetrin, azoxystrobin og betazon på limniske invertebrater. Bekæmpelsesmiddelforskning fra Miljøstyrelsen [76], 1-97. Lauridsen, T.L. & Wiggers, L. (2001). Pesticider i Byrup Langsø. Vand og Jord, 154-159. Lawrence, E. (1995). Henderson´s Dictionary of Biological Terms 11. Ed. Longman Group 1995 Liess, M., Schulz, R., Liess, M.H.-D., Rother, B. & Kreuzig, R. (1999). Determination of insecticide contamination in agriculturel headwater streams. Water Research 33[1], 239-247. Maltby, L. (1995). Sensitivity of the crustaceans Gammarus pulex (L.) and Asellus aquaticus (L.) to short-term exposure of hypoxia and uionized ammonia observations and possible mechanisms. Water Research 29[3], 781-787. McCahon, C.P., Poulton M.J., Thomas, P.C., Xu Q., Pascoe, D. & Turner, C. (1990.) Lethal and sublethal toxicity of filed simulated farm waste episodes to several freshwater invertebrate species. Water Research 25[6], 661-671. Miljøstyrelsen (1998) Vejledning fra Miljøstyrelsen, nr. 5 (1998). Biologisk bedømmelse af Vandløbskvalitet. Miljø og Energiministeriet: Miljøstyrelsen, København. Miljøstyrelsen (2004). Bekæmpelsesmiddelstatistik 2003 (http://www.mst.dk/) Miljøstyrelsen (2005). Akutte og kroniske effekter af otte pesticider over for Daphnia magna. EU list of endpoints: Dimethoate (April 2005), Pirimicarb (August 2004), Chlorpyrifos (March 2005), Pyrazophos (ECCO64). Review report (6846/VI/97), Esfenvalerat (November 2000); Review report (7581/VI/97), Azoxystrobin (April 1998); Review report (SANCO/4347/2000), Bromoxynil (February 2004). Via: http://forum.europa.eu.int/Public/irc/sanco/Home/main. Møhlenberg F., Schlüter L., Gustavson K., Andersen, T.T., Forbes V., Gold A., Friberg N., Larsen, S.E. & Lauridsen, S.B. (2004). Effekt af bekæmpelsesmidler på flora og fauna i vandløb. Bekæmpelsesmiddelforskning fra Miljøstyrelsen [82], 1-134. Naddy, R. B., Johnson, K.A. & Klaine, S.J. (2000). Response of Daphnia magna to pulsed exposures of chlorpyrifos. Environmental Toxicology and Chemistry 19[2], 423-431. Naddy, R.B. & Klaine, S.J. (2001). Effect of pulse frequency and interval on the toxicity of chlorpyrifos to Daphnia magna. Chemosphere 45, 497-506. Nørum, U. & Bjerregaard, P. (2003). Ferskvandsinvertebrates bevægelsesadfærd som biomarkør for pesticideksponering og -effekt. Bekæmpelsesmiddelforskning fra Miljøstyrelsen [75], 3-64. OECD (1998). Guideline for testing of chemicals- Guideline 211: Daphnia magna Reproduction Test. Organisation of Economic Co-Operation and Development, Paris, France. Parsons, J.T. & Surgeoner, G.A. (1991). Effect of exposure time on the acute toxicities of permethrin, fenitrothion, carbaryl, and carbofuran to mosquito larvae. Environmental Toxicology and Chemistry, 10, 1219-1227. Phillips, P.J. & Bode, R.W. (2004). Pesticides in surface water runoff in south-eastern New York State, USA: seasonal and stormflow effects on concentrations. Pest. Manag. Sci., 60, 531-543. Raven, P. H., Evert, R. F & Eichhorn, S. E. (1992). Biology of Plants 5. Ed. Worth Publishers 1992 Reinert, K.H., Giddings, J.M., Judd, L. (2002). Effect analysis of time-varying or repeated exposures in aquatic ecological risk assessment of agrochemicals. Environmental Toxicology and Chemistry, 21 (9), 1977-1992. Rippen, G. (2004). Umweltchemikalien. CD-ROM. EcoMed. ISBN 3-609-58021-6. Sánchez, M., Ferrando, M.D., Sancho, E. & Andreu, E. (1999). Assesment of the toxicity of a pesticide with a two-generation reproduction test using Daphnia magna. Comparative Biochemistry and Physiology Part C 124, 247-252. Schulz, R. & Liess, M. (2000). Toxicity of esfenvalerat to caddisfly larvae: chronic effects of 1 vs. 10h pulse exposure with constant doses. Chemosphere 41, 1511-1517. Slothuus T., Jørgensen K.E., Nørum U. & Bjerregaard P. (2004). Kobber i dambrug. Vand & Jord 11, 93-97. Stenersen, J. (2004). Chemical Pesticides Mode of Action and Toxicology. CRC Press. Stephenson, R. R. (1982) Aquatic toxicology of cypermethrin. I Acute toxicity to some freshwater fish and invertebrates in laboratory tests. Aquatic Toxicology 2, 175-185. Stryer, L. (1995). Biochemistry. 4. Ed., W. H. Freeman and Company, New York. Styczen, M., Wiberg-Larsen, P. & Aagaard, A. (2003 ). Tag pulsen på pesticiderne i vandmiljøet. Vand og Jord 3, 84-87. Styczen, M. & Poulsen, R.N. (2005). Etablering af forvaltningsgrundlag for anvendelse af PestSurf. Bekæmpelsesmiddelforskning fra Miljøstyrelsen (in prep.) Timbrell, J. A. (1991). Principles of Biochemical Toxicology, 2. Ed. Taylor and Francis Ltd. Tomlin, C. D. S., (2003.) BCPE: The e-Pesticide Manual version 3.0 British Crop Protection Council. Toxnet (2005). Hazardous Substances Data Bank. http://toxnet.nlm.nih.gov/ (besøgt: januar 2005) Tucker, K.A. & Burton, G.A. (1999). Assessment of nonpoint-source runoff in a stream using in situ and laboratory approaches. Environmental Toxicology and Chemistry, 18 (12), 2797-2803. US-EPA (2004). Ecotox Database. www.epa.gov/ecotox (besøgt november 2004) Van der Hoeven, N. & Gerritsen, A. A. M (1997). Effects of Chlorpyrifos on Individual and Populations of Daphnia pulex in the Laboratory and Field. Environmental Toxicology and Chemistry 16[12] 2438-2447. Verschueren, K. (1996). Handbook of Environmental Data on Organic Chemicals. 3. ed. Van Nostrand Reinhold, 1996. Walker, C. H, Hopkin, S. P. Sibly, R. M. & Peakall, D. B. (1996). Principles of Ecotoxicology. Taylor and Francis Ltd. Wiberg-Larsen, P., Adamsen, N.B., Knudsen, J. & Larsen, F.G. (1991). Sprøjtegifte truer fynske vandløb. Vand og Miljø 7, 371-374. Wright, A. (1976). The Use of Recovery as a Criterion for Toxicity. Bulletin of Environmental Contamination and Toxicology 15[6], 747-749. Bilag 1 Testprotokol - Toksiske effekter af pulseksponering for pesticider - akutte og kroniske effekter hos Daphnia magna
Tina Slothuus, Signe Qualmann og Anders Baun Institut for Miljø & Ressourcer Danmarks Tekniske Universitet Kgs. Lyngby Juni 2005
ForordProtokollen beskriver fremgangsmåderne anvendt til bestemmelse af toksiske effekter af pulseksponeringer for pesticider. Metoderne er inddelt i 1) Kontinuerte eksponeringer, hvor der ses på akutte effekter af eksponeringer for et pesticid, i form af immobilisering og død. Testen udføres efter principperne beskrevet i ISO-6341 (1989a) og ISO-8692 (1989b). 2) Pulstest til bestemmelse af immobilisering efter en kortvarig eksponering (0,5-6t) for et pesticid samt 3) Reproduktionstest til bestemmelse af kroniske effekter efter eksponeringen for en puls af et pesticid. Testen udføres som en modificeret udgave af OECD Guideline nr 211 (1996). 1 ForsøgsorganismerSom forsøgsorganisme benyttes Daphnia magna, indsamlet i Langedammen ved Birkerød i 1978 og dyrket i laboratoriet siden. De dafnier, der anvendes i forsøgene, er <24 timer gamle. Dette sikres ved at oprense mediet senest 24 timer inden, forsøgene sættes i gang og fjerne alle organismer på nær moderdyrene (ISO, 1989a). 1.1 KulturholdDafniestamkulturen dyrkes i klimarum ved en temperatur på 20°C ± 2°C og en 12/12 timers lys/mørke periode (4,84-5,62 μEl*m-2*s-1). Kulturerne holdes i akvarieglas (18x13x18 cm) eller 1 liters bægerglas. Akvarierne fyldes med 1,6 l M7-medie (se afsnit 1.2), og bægerglassene fyldes med 800 ml M7 medie. Der anvendes højst 30 dyr pr. akvarie og højst 20 dyr til bægerglassene. Hvis der tilsættes flere dyr, kan konkurrencen mellem dyrene blive for stor, og produktionen af unger mindskes. Akvarieglassene overdækkes med plexiglas og bægerglas dækkes med urglas. Kulturerne holdes ca. 2 måneder inden de skiftes ud, for at holde produktionen af unger optimal. Dyrene får rent medie 2 gange ugentligt med 2-3 dages mellemrum. Når mediet skal skiftes hældes indholdet fra akvarierne, d.v.s medie og dafnier, over i en plastikbakke. Akvarierne skylles herefter i destilleret vand, så algerester fjernes fra siderne. 800 ml (400 ml for bægerglassene) rent M7 medie fyldes derefter op i de rene akvarier. Mediet med dafnierne filtreres herefter gennem et nylonfilter og 800 ml (400 ml for bægerglassene) af opløsningen hældes ligeledes over i akvarierne. Det nye medie består således af nyfremstillet M7 medie og ”gammelt” M7 medie i forholdet 1:1. Moderdyrene overføres til det nye medie med et nylonnet. Dafnierne får føde i form af grønalgen Pseudokirchneriella subcapitata (dyrkning m.v. beskrevet i afsnit 1.3), der tilføres akvarierne v.h.a. et pumpesystem 3 gange dagligt. Mængden af føde, der tilføres, skal svare til 1-2 mg C /10 dyr i.flg. OECD standarden (OECD, 1996). Hvis det vurderes at være nødvendigt, d.v.s. hvis der ikke produceres tilstrækkeligt med unger, kan der tilsættes en smule tørgær som ekstra foder. Tørgæret (nogle få korn) opløses i en smule M7-medie, inden det kommes i akvariet. 1.2 Fremstilling af M7-medieDet anvendte medium er kunstigt fremstillet ferskvand, såkaldt M7 medie fremstillet efter nedenstående forskrift (Tabel 1 og Tabel 2). De anvendte kemikalieopløsninger, blev fremstillet ved at afveje de angivne stofmængder, overføre disse til en 1-liters målekolbe og tilsætte Milli-Q vand til 1l-mærket. Til fremstillingen af M7 mediet blev der efterfølgende med en pipette udtaget de i Tabel 1 angivne volumener af stamopløsninger. Mediet blev ligeledes fremstillet med Milli-Q vand. Tabel 1: Kemikalier brugt til fremstilling af M7 medie
*Kemikalierne opløses i Milli-Q-vand **Vitaminerne skal opbevares koldt og tilsættes kort tid før anvendelse. Tabel 2. Fremstilling af stamopløsning II (M²)
*Kemikalierne opløses i Milli-Q-vand 1.3 Fremstilling af alge-medieVed dyrkning af alger til foder, overføres ca. 100 ml algesuspension (celletæthed: 4,7-6,1 ·106 celler/ml) til 5 liter nyt fremstillet algemedie fremstillet efter opskriften i tabel 3. Mediet skal have en temperatur på 20°C og pH 8,0 ± 0,2. Kulturen sættes på en magnetomrører, gennembobles med atmosfærisk luft, der først passerer et sterilfilter, inden det bobles gennem mediet, og dyrkes under konstant belysning (38 μEl*m-2*s-1) i 2-3 døgn, indtil algesuspension har en celletæthed på 4,7-6,1 ·106 celler/ml, hvilket afspejles i en mørk, grøn farve. Algekulturen opbevares mørkt ved en temperatur på 4°C, indtil den skal anvendes til foder. Flasken omrystes før brug, da algen bundfælder under opbevaringen. Det er ikke nødvendigt at vente til algesuspensionen opnår en temperatur på 20°C, inden den overføres til det nye algemedie. Algen bruges løbende, men vi vurderer, at den vil kunne holde sig mindst 2-3 uger i kølerum. Fodringen af dafnierne med et koncentreret medie betyder, at der ikke kommer for megen algemedie over i dafniekulturerne. Celletætheden måles på Coulter Counter Multisizer, og kulturens renhed undersøges ved mikroskopering hver 3. uge. Tabel 3: Fremstilling af algemedie
*Autoklaveres i 15 minutter ved 121ºC 2 Apparatur og materialer
3 Validitietskrav-referencetestStandard ISO testen for immobilisering (ISO, 1989a) udføres med kaliumdikromat (K2Cr2O7) som referencestof. I følge standarden skal IC50,24t ligge i intervallet 0,9-2,4 mg/l. I flg. ISO (1989a) kræves en iltmængde på ≥ 2 mg/l og en pH på 7,8 ± 0,2. Da M7 mediet ikke har en saltsammensætning, der stemmer 100% overens med ISO standarden, kan afvigelser i pH-værdien forventes, værdien bør dog ikke variere mere en 1,5 enhed. 4 Akut toksicitetstest - immobiliseringAlle akutte tests udføres i henhold til ISO(1989a). Der anvendes 4 replikater pr. koncentration og 6 replikater i kontrolgruppen. Hver replikat består af 25 ml testopløsning (M7 tilsat teststof/pesticid) og 5 juvenile Daphnia magna (<24 t). Testen udføres i bægerglas (100 ml), som dækkes med urglas, for at forhindre støv og andre partikler i at komme ned i glassene. Testen forløber i mørke og dyrene eksponeres kontinuert i 48 timer. Der skiftes ikke opløsning i løbet af de 48 timer. Under eksponeringen får dyrene ikke tilført føde i form af alger, da alger i opløsningen kan påvirke den stofmængde, der er tilgængelig til eksponeringen af dyrene. Testen varer kun 48 timer, da immobilisering ellers kan indtræffe p.g.a. fødemangel og ikke nødvendigvis p.g.a. kemikaliet, der testes. Der kontrolleres for immobilisering og dødelighed efter 0, 2, 4, 6, 24 og 48 timers eksponering. Testen er udvidet m.h.t. antallet af aflæsningstider, da disse vil være nyttige, når der skal udføres pulstests. Afvigelser fra ”normal” noteres ligeledes. Definitioner: Immobilitet: Dafnier, som ikke kan opretholde deres position i vandsøjlen (svømme) selv efter en forsigtig stimulering (15 sek.), regnes for immobile. Dafnierne kan have krampeagtige bevægelser. Døde dafnier vil indgå i det samlede antal immobile dafnier, men der skelnes mellem døde og immobile dyr ved observationerne. Dødelighed: Dafnier, der ikke har nogen form for bevægelse, som f.eks. bevægelse af antenner, selv 15 sekunder efter stimuli, regnes for døde (Naddy et al., 2000, OECD Guideline, 1996). I praksis blev dette overført til, at mobile dyr skal svømme i løbet af de 15 sekunder de observeres men ikke nødvendigvis i alle de 15 sekunder. Dette skyldes, at der ofte blev observeret dafnier, der ikke svømmede i en kort tidsperiode (opholdt sig typisk i bunden af glassene) for så pludselig at svømme livligt rundt. Resultaterne bruges til beregning af IC50, 24t og IC50, 48t. 5 Pulseksponering - ImmobiliseringProceduren for pulseksponering er illustreret i Figur 1. Dyrene eksponeres i grupper af 20 individer. Efter endt eksponering indfanges dyrene og overføres til et skyllekar og videre til observationsglas. Dette gøres v.h.a. en engangspipette af plast, som dyrene suges op i. Den overskydende væske sprøjtes derefter forsigtigt ud, indtil dyrene er ved åbningen af pipettespidsen. Herefter overføres dyrene til mediet i skyllekarret. Ved desuden at skifte mediet i skyllekarret hver gang en ny gruppe af dyr overføres, minimeres mængden af pesticid, der kommer over i de endelige observationsglas. Dyrene indfanges i skyllekarret v.h.a. et fintmasket net, hvorfra overskydende væske fjernes ved at lægge det imod en papirsserviet. Herefter fordeles dyrene i 4 grupper pr. eksponeringstid. Hver replikat består således af 5 juvenile Daphnia magna (<24t) (se Tabel 4). Tabel 4: Pulseksponering af Daphnia magna ved fastholdt teststofkoncentration.
Ved hvert forsøg anvendes 5 forskellige eksponeringstider (se Tabel 4) og 2-3 koncentrationer af teststoffet. Eksponeringskoncentrationerne udvælges på baggrund af gennemførte ISO-test for immobilisering, ved at beregne den koncentration, der vil have en effekt på 90% af dyrene, en såkaldt IC90,24t-værdi. Denne værdi er valgt ud fra den antagelse, at dyrene skal udsættes for en højere koncentration under pulstestene end under den kontinuerte eksponering under akuttestene, hvis der skal ses en effekt, da pulstestene er af meget kortere varighed (maks. 6 timer) sammenlignet med akuttestene. Ligeledes er der taget hensyn til dyrenes reaktion overfor pesticidet efter 6 timers eksponering under akuttestene- hvilket vil svare til en 6 timers puls. På baggrund af dette kan koncentrationen sænkes/øges yderligere. Der anvendes en kontrolgruppe bestående af 30 dyr, som fordeles i 6 replikater. Kontrolgruppen håndteres på tilsvarende måde som gruppen, der får en puls af 0,5 time, da det forventes, at der vil være en større stressfaktor hvis dyrene i kontrolgruppen håndteres gentagende gange i korte tidsintervaller. Antallet af immobile og døde dyr registreres 0 timer, 24 timer og 48 timer efter pulsens ophør. I posteksponeringsperioden får dyrene føde i form af algen Pseudokirchneriella subcapitata . Dyrene får en føde mængde svarende til 0,1-0,2 mg C/dag (C = carbon). Føden tilføres umiddelbart efter overførslen af dyrene til rent vand. Resultaterne bruges til beregning af PEt-IT50-værdier (Post Exposure Inhibition Time 50). Værdierne angiver den pulsvarighed, der kræves for at 50% af dyrene er immobile til tiden t (i dette tilfælde t= 0, 24 og 48 timer).
Figur 1: Pulseksponering af dafnier - akutte effekter. 5.1 Beregning af tilsætning af algemængde under postobservationsperiodenTælletal: V (målt på en koncentreret algeopløsning på Culter Counter) Fortyndingsfaktor: x Celler pr. ml algeopløsning er derfor V*x = W celler/ml (NB! Der skal tages højde for den volumen der er målt på. Er der f.eks. målt på 0,5 ml skal der ganges med 2 osv.) Algen Pseudokirchneriella subcapitata har ca. 1*10-8 mg C pr. celle (Halling-Sørensen et al., 1996). Ved at multiplicere de to værdier fås algekoncentrationen i mg C/ml og fodervoluminen kan beregnes: (W celler/ml) * (1*10-8 mg C/ celle) = X mg C/ml OECD sætter mængden af carbon (C), der skal være tilgængelig for hver dafnie pr. dag, til 0,1-0,2 mg C. Der skal derfor tilsættes mellem Y og Z ml/dafnie/dag (0,1 mg C/dag / X mg C/ml ) = Y ml/dag (0,2 mg C/dag / X mg C/ml ) = Z ml/dag EksempelTælletal: 30369 (målt på en koncentreret algeopløsning på Culter Counter) Fortyndingsfaktor: 200 Volumen: 0,5 ml Celler pr. ml algeopløsning er derfor 30369*200*2= 1,2*107 celler/ml D.v.s. (1,2*107 celler/ml) * (1*10^-8 mg C/ celle) = 0,12 mg C/ml Der skal derfor tilsættes 0,83-1,67 ml/dafnie/dag da (0,1mg C/dag / 0,12 mg C/ml ) = 0,83 ml/dag (0,2 mg C/dag / 0,12 mg C/ml ) = 1,67 ml/dag 6 Kronisk toksicitet – ReproduktionAnbefalingerne i OECD (1996) er blevet fulgt, d.v.s. kravet om max. 20 % døde moderdyr blandt de eksponerede dyr, kravet om min. 60 unger pr. moderdyr i kontrolgruppen, en pH-værdi mellem 6 og 9 og som ikke varierer mere end 1,5 enheder, og et O2-niveau over 3 mg/l. Desuden er der lavet visse modifikationer, da OECD guiden er designet som en forlænget akuttest, hvor dyrene går i en fast stofkoncentration. I modsætning hertil er pulstesten for kroniske effekter udført således, at dyrene eksponeres for teststoffet i en given tidsperiode, hvorefter de igen kommer over i rent medie. Kravet om 10 testdyr pr. koncentration er overført til 10 testdyr pr. eksponeringstid ved fastholdt eksponeringskoncentration. De 10 dyr eksponeres i en fælles beholder. Gruppen af dyr, der skal have en puls af 6 timers varighed, eksponeres som den første og 2 timer senere eksponeres gruppen, der skal have en puls af 4 timers varighed o.s.v., indtil alle grupper er blevet eksponeret (på denne måde afsluttes alle grupperne på samme tid efter 21 dage). Dernæst indsamles dyrene v.h.a. en pipette og overføres til et kar med rent M7 medie, der fungerer som skyllekar. Så meget som muligt af den overskydende væske i pipetten fjernes, inden dyrene overføres til karret. Fra skyllekarene overføres dyrene til et observationsglas ved at indfange dem med et fintmasket net, hvorfra vandet kan dryppe af eller nettet duppes mod en papirsserviet. Herved minimeres risikoen for kontaminering af rentvandsglassene, der fungerer som observationglas. I posteksponeringsperioden går dyrene enkeltvis i 100 ml bægerglas. Dyrene, der indgår i kontrolgrupperne, eksponeres for rent M7 medie på samme måde som dyrene, der eksponeres for et kemikalie. D.v.s. de bliver også overført til et nyt medie og ”skyllet”. På denne måde sikres det, at variationer ikke skyldes forskelle i håndteringen af organismerne (jf. anbefalinger af Naddy & Klaine, 2001). Reproduktionen hos Daphnia magna følges efter en pulseksponering (0,5t, 1t; 2t; 4t; og 6t ) og 21 dage frem. I posteksponeringsperioden frem til de 21 dage får dyrene føde tilsat i form af algekulturen Pseudokirchneriella subcapitata. Mængden af føde svarer til ca 0,1-0,2 mg C/ dag (0,1 mg C den første uge siden 0,2 mg C). Dyrene går ved 20 °C ± 2°C og en 12/12 timers lys/mørke periode (4,84-5,62 μEl*m-2*s-1). Der måles ilt og pH ved hvert medieskift. I OECD’s guideline anbefales det, at mediet med kemikalie skiftes tre gange ugentligt for at sikre, at niveauet af kemikalie holdes på et fast niveau (OECD, 1996). Dyrene i pulstesten for kroniske effekter kommer over i et rent medie uden kemikalie efter få timer, og mediet skiftes derfor kun to gange ugentligt på samme måde som beskrevet for stamkulturen med moderdyr (se afsnit 1.1.1). Følgende parametre kontrolleres under testene og ved testenes afslutning:
7 ReferencerHalling-Sørensen, B., Nyholm, N. & Baun, A. (1995) Algal Toxicity tests with volatile and hazardous compounds in air tight test flasks with CO2 enriched headspace. Chemosphere, vol. 8, pp. 1513-1525. ISO (1989a). Water quality- Determination of the inhibition of the mobility of Daphnia magna Straus (Cladocera, Crustacea). ISO 6341. International Organization for Standardization, Geneva, Switzerland. ISO (1989b). Water quality- Freshwater algal growth inhibition test with Scenedesmus subspicatus and Selenastrum capricornutum. ISO 8692. International Organization for Standardization, Geneva, Switzerland Naddy, R. B., Johnson, K. A., Klaine, S. J. (2000). Response of Daphnia magna to pulsed exposures of chlorpyrifos. Evironmental Toxicology and Chemistry, Vol. 19, No 2, pp. 423-431. Naddy, R. B., Klaine, S. J. (2001). Effect of pulse frequency and interval on the toxicity of chlorpyrifos to Daphnia magna. Chemosphere, Vol. 45, pp. 497-506. OECD Guidelines for testing of chemicals (1996). Proposal for updated guideline 211. Daphnia magna Reproduction test. Organization for Economic Cooperation and Development, Paris, France. Bilag 2 Målinger af ilt og pH2.1 Ilt- og pH-målinger foretaget under akuttest Iltmålinger og pH målinger foretaget under testene fremgår af tabel 1-3. Tabel 1: O2 og pH i testopløsningerne målt i forbindelse med akuttestene.
* Forsøget blev udført som beskrevet i ISO standarden (1989a). Det anvendte medie er imidlertid fremstillet som beskrevet i OECD standarden (1996) og krav til ilt og pH er derfor angivet for denne standard. 2.2 Ilt- og pH-målinger foretaget under akutte pulstest Tabel 2: O2 og pH i testopløsningerne målt i forbindelse med pulstestene.
b: ingen DMSO kontrol * Krav til pH og O2-koncentrationen i M7 medie angivet i OECD standarden (1996). 2.3 Ilt- og pH-målinger foretaget under kroniske pulstest Tabel 3: Validitetskrav i flg. OECD (O2 og pH) (1996) samt resultater fra de kroniske pulstests.
* Krav til pH og O2-koncentrationen i M7 medie angivet i OECD standarden (1996). ** Parametrene blev målt ved hvert medieskift. Den laveste og højeste målte værdi ud af samtlige målinger er angivet. Bilag 3 Data for ISO-test (akut)3.1 Koncentrations-Responskurver for akuttest.
Figur 1: Dosis-Respons kurve for akuttest med pirimicarb, esfenvalerat, diquat, chlorpyrifos og pyrazophos. Det var ikke muligt at estimere en kurve for diquat (48t).
Figur 2: Dosis-Respons kurve for akuttest med dimethoat, azoxystrobin, bromoxynil, 4-nitrophenol og 3,5-dichlorophenol. 3.2 Beregninger fra ToxCalc (akuttest) For at sikre at bilag ikke opnåede en uoverskuelig størrelse, er data fra beregninger i ToxCalc udeladt, hvis disse udelukkende eller til en stor del bestod af ekstrapolerede data. I de givne tilfælde vil dette være angivet. 3.2.1 Dimethoat Tabel 4: EC-værdier beregnet for dimethoat (E = Immobilitet). 3.2.2 Pirimicarb Tabel 5: EC-værdier beregnet for pirimicarb (E = Immobilitet). 3.2.3 Chlorpyrifos Tabel 6: EC-værdier beregnet for chlorpyrifos (E = Immobilitet). Tabel 7: EC- værdier beregnet for esfenvalerat (test 2 (0,05-10,0μg/l))) (E = Immobilitet). Tabel 8: EC-værdier beregnet for esfenvalerat (test 3 (0,05-10,0μg/l))) (E = Immobilitet). 3.2.5 Azoxystrobin Tabel 9: EC- værdier beregnet for azoxystrobin (E = Immobilitet). 3.2.6 Pyrazophos Tabel 10: EC-værdier beregnet pyrazophos (E = Immobilitet). 3.2.8 Bromoxynil Tabel 11: EC-værdier beregnet for bromoxynil (E = Immobilitet). 3.2.9 4-nitrophenol Tabel 12: EC- værdier beregnet for 4-nitrophenol (E = Immobilitet). 3.2.10 3,5-dichlorophenol Tabel 13: EC-værdier beregnet for 3,5-dichlorophenol (E = Immobilitet). 3.2.11 Kaliumdikromat Tabel 14: EC-værdier beregnet for kaliumdikromat (E = Immobilitet). Bilag 4 Pulstest – akutte effekter4.1 Data fra akutte pulstests beregnet v.h.a. ToxCalc For at sikre at bilag ikke opnåede en uoverskuelig størrelse, er data fra beregninger i ToxCalc udeladt, hvis disse udelukkende eller til en stor del bestod af ekstrapolerede data. I de givne tilfælde vil dette være angivet. 4.2 Fordelingen af mobile, immobile og døde dafnier som funktion af tiden (akutte pulstest) Dimethoate og pirimicarb var de første to pesticider der blev testet. Under forsøgene med disse stoffer blev der ikke skelnet mellem død og immobil og der er derfor ikke lavet en grafisk fremstilling for disse. 4.2.1 Chlorpyrifos
Figur 1: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for chlorpyrifos (2,5 μg/l og 3 μg/l). Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.2 Esfenvalerat
Figur 2: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 20, μg/l og 30 μg/l esfenvalerat. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.3 Pyrazophos
Figur 3: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 2,5-20 μg/l pyrazophos. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.4 Azoxystrobin
Figur 4: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 10,5-12,5 μg/l azoxystrobin. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.5 Diqua 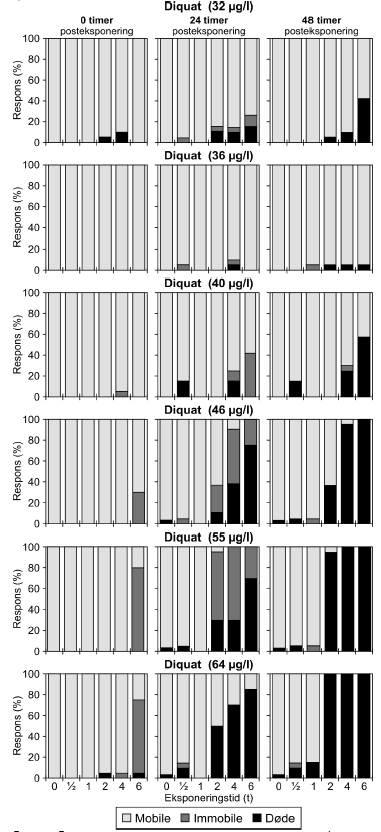
Figur 5: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for diquat (32, 36, 40, 46, 55 og 64 μg/l). Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.6 Bromoxynil
Figur 6: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 24, 25 og 26 mg/l bromoxynil. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.7 4-nitrophenol
Figur 7: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 14, 15 og 16 mg/l 4-nitrophenol. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. 4.2.8 3,5-dichlorophenol
Figur 8: Fordeling af mobile, immobile og døde dafnier (angivet i %) efter 0t, 24t og 48t posteksponering for 2,9, 3,1 og 3,3 mg/l 3,5-dichlorophenol. Ved hver eksponering blev anvendt 20 dyr, mens der var 30 dyr i kontrolgrupperne. Bilag 5 Gentagne pulseksponeringer5.1. Gentagne pulseksponeringer 5.1.1 Dimethoat
Figur 9: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med dimethoat 10 mg/l.
Figur 10: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med dimethoat 20 mg/l. 5.1.2. Pirimicarb
Figur 11: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med pirimicarb (40 μg/l).
Figur 12: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med pirimicarb (70 μg/l). 5.1.3 Esfenvalerat
Figur 13: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med esfenvalerat (10 μg/l). 5.1.4 Pyrazophos
Figur 14: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med pyrazophos (2,5 μg/l).
Figur 15: Fordelingen af Mobile, Immobile og Døde dafnier til tiderne t= 0, 24 og 48 timer efter gentagne pulseksponeringer med pyrazophos (5 μg/l). Bilag 6 Pulstest – kroniske effekter6.1 Effekt af pulseksponering på reproduktion 6.1.2 Dimethoat Tabel 45: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 30 mg/l dimethoat (forsøg 1).
Tabel 46: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 30 mg/l dimethoat (forsøg 2).
Tabel 47: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper (forsøg 1) efter pulseksponering for 30 mg/l dimethoat.
Tabel 48: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper (forsøg 2) efter pulseksponering for 30 mg/l dimethoat.
Tabel 49: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter eksponering for 30 mg/l dimethoat (forsøg 2)
6.1.2 Pirimicarb Tabel 50: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 100 μg/l pirimicarb.
Tabel 51: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for 100 μg/l pirimicarb.
Tabel 52: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter eksponering for 100 μg/l pirimicarb
6.1.3 Chlorpyrifos Tabel 53: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 1,2 μg/l chlorpyrifos.
Tabel 54: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for 1,2 μg/l chlorpyrifos.
Tabel 55: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter pulseksponering for 1,2 μg/l chlorpyrifos
6.1.4 Esfenvalerat Tabel 56: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 0,1 μg/l esfenvalerat.
Tabel 57: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for 0,1 μg/l esfenvalerat.
Tabel 58: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter pulseksponering for 0,1 μg/l esfenvalerat.
Der blev foretaget flere gentagne forsøg med esfenvalerat (20 μg/l, 10 μg/l, 5 μg/l og 1 μg/l). I alle testene var dødeligheden høj allerede efter få dage (tabel 7). Under testen udført med koncentrationerne 5 μg/l og 1 μg/l opstod der ligeledes problemer med algen som blev brugt til foder. Der opstod en markant vækst af blågrønalger i stort set alle algeopløsninger og vi havde derfor en mangel på foder. Vi forsøgte at bruge algen på trods af væksten, men forsøget blev afsluttet da algen satte sig fast på dyrene hvorved deres bevægelse og måske også deres iltoptagelse er blevet påvirket. Tabel 59: % immobile og % døde dafnier efter 0, 48 og 96 timer (1 μg/l: 0 og 48 timer samt 6 dage) efter eksponering for esfenvalerat (1, 5 og 20 μg/l)
(n =10 (5μg/l, 2t: n=9)) Tabel 60: Beregnede PEt –IT50 værdier for pulseksponering med esfenvalerat – kronisk test (1 μg/l og 5 μg/l) pulstest (20 μg/l og 30 μg/l)
Tilsvarende beregnninger kunne ikke foretages for forsøget med 0,1μg/l, da ingen af grupperne reagerede i løbet af de første 48 timer efter eksponeringen. Tabel 61: Reproduktionsdata for dafnier eksponeret for esfenvalerat (0,1 μg/l).
* signifikant forskellig fra kontrollen a) kun et dyr 6.1.5. Bromoxynil Tabel 62: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 20 mg/l bromoxynil
Tabel 63: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for 20 mg/l bromoxynil.
Tabel 64: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter pulseksponering for 20 mg/l bromoxynil
6.1.6 4-nitrophenol Tabel 65: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for henholdsvis 2 mg og 8 mg 4-nitrophenol.
Tabel 66: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for henholdsvis 2 mg/l og 8 mg/l 4-nitrophenol.
Tabel 67: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter eksponering for henholdsvis 2 mg/l og 8 mg/l 4-nitrophenol.
6.1.7 3,5-dichlorophenol Tabel 68: Gennemsnitlig antal afkom (dag 14 og dag 21) efter pulseksponering for 2,5 mg/l 3,5-dichlorophenol.
Tabel 69: % Døde moderdyr efter 14 og 21 dage fordelt over eksponeringsgrupper efter pulseksponering for 2,5 mg/l 3,5-dichlorophenol
Tabel 70: Gennemsnitlig længde og tørvægt af overlevende moderdyr (dag 21) efter eksponering for henholdsvis 3,5-dichlorophenol.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||