Effects of reduced pesticide use on flora and fauna in agricultural fields
3 Arthropods
Farmland crop pests like Aphididae (aphids) and Oulema melanops (Cereal
leaf beetles) as well as their natural enemies, often referred to as beneficials, have
received much attention in pesticide research. The term "beneficial" when
applied to crop-dwelling arthropods is usually associated with the predatory species of
crop pests or with pollinators. There are however many other groups of species whose role
within the crops may be termed beneficial. These include the so-called chick-food group;
several orders or guilds of insects that are important components in the diet of other
farmland species, especially birds (Sotherton & Moreby 1992). In this chapter
attention will be paid to this group as well as on the traditional target species.
3.1 Pilot studies
Pilot experiments were conducted primarily to evaluate the time consumption and suitability of different sampling methods under the actual conditions and also to get an overview of the arthropod composition in general and in relation to reduced pesticide dosages.
Sampling of arthropods on plant foliage and on the ground below is an ongoing challenge to entomologists and each method has advantages and shortcomings. No single method of density or abundance estimation is suitable for all circumstances and different sampling methods are therefore relevant in order to obtain a reasonably efficient sampling depending on the target species.
The pilot experiments were carried out on the farm "Gjorslev" in 1996. Four sampling methods were evaluated in all three crops:
1. Coloured water traps, consisting of circular plastic bowls (diam. 200 mm, height 85 mm). A set of three traps in different colours was placed together on the ground and in each 1 litre of trapping fluid (water added 70% alcohol and ethylene glycol (3:1:1) with one drop of non-perfumed dish soap) to ensure drowning and preservation. In each plot five sets of traps were randomly placed avoiding the outer 20 metres of the plots to limit interference from boarder zones. The traps were serviced once a week from 9 May to 1 August in the cereals and 9 May to 10 October in beets. All catches were preserved in 70% alcohol until further treatment. The target arthropod groups were primarily flying insects and insects living in the canopy.
2. Pitfalls (plastic cups, diam. 110 mm, depth 135 mm) were buried pairwise in the ground 1 meter apart, connected by a 150 mm high steel barrier to improve the efficiency of the traps. The traps were partly filled with trapping fluid (see above). The number, distribution as well as sampling of the traps was performed as described above. The target group was epigaeic arthropods, in particular the adult carabids, a predominant and important arthropod family in the arable land.
3. Suction sampling. Different machinery has been used to sample by suction arthropods living on plant stems, in canopy and on the soil surface. The most widely used is the D-vac (Dietrick et al. 1959, Dietrick 1961). The advantages of vac-sampling is that it provides density estimates in contrast to water traps and pitfalls, which provide relative estimates since their sampling area is not defined. For the pilot experiments a modified hand carried vac-sampler was used. A few samples were taken in spring, but the machinery was not sufficiently powerful for this purpose and the sampling was cancelled.
4. Direct countings were conducted in all crops primarily to estimate Aphididae populations, but in beets also to monitor a severe attack of Autographa gamma (Silver Y’s). Sampling comprised mainly the Aphididae Rhopalosiphum padi (Bird-cherry oat aphid) and Sitobion avenae (English grain aphid) in the cereals and Aphis fabae (Black bean aphid) in beets. On each assessment day and in each plot, 100 randomly selected wheat and barley ears on a diagonal transect (Danielsen 1992) were inspected and the number of cereal ears with Aphididae was counted. In beets, sampling was done on weekly intervals in the period early July to late August. 50 plants per plot were inspected following in principle the same methodology as in the cereals. Four times during the season 21 randomly selected sugar beets plants were inspected for larvae of A. gamma (this was an ad hoc methodology).
All arthropods collected were identified to at least order. Coleoptera (beetles) were all identified at least to family with most emphasis on the pitfall collected Carabidae (ground beetles) among which imagines of larger species were identified to species. In the water traps the most abundant larger Diptera (two-winged) were identified to family. Lepidoptera (butterflies and moths) and Symphyta (sawflies) were not identified further with A. gamma as an exception. The water trap catches could be huge which automatically restricted the sorting to the higher level taxi.
3.1.2 Statistical analyses (pilot study)
Water traps and pitfalls
Data for all common groups of arthropods, depending on taxonomic level of identification, were analysed for possible dosage effects on the populations. Each crop was analysed separately since each type of crop was considered unique. Also, the data from water traps and pitfalls were analysed separately because of their different mode of action and their different target species, but the statistical method used was the same. The data on each relevant arthropod group from each set of traps on each sampling date were loge(x+1) transformed to normalise variance. Subsequently the data from the various sampling dates from each trap set were pooled to avoid that repeated measurements on the same spots provided dependent data (Stryhn 1996). The dependent variable (total number per trap group) was assumed to follow a Poisson distribution and was analysed by the GENMOD procedure in SAS/STAT using Likelihood ratio tests (SAS Institute 1990). The variable was analysed in relation to the class variable dosage only and corrections for over-dispersion were made. The replicates (sets of traps) were all from the same plot due to the experimental design, and this lack of "true" replications weakens the reliability of the test results.
Suction sampling
The sampling was insufficient for statistical analyses.
Direct counting
No statistical analyses were conducted. Percent infested plants on a given sampling date are presented.
3.1.3 Results (pilot study)
Water traps
The results from the water traps were unsatisfactory, variation in catch was very high and no results will be presented.
Pitfalls
It should be stated that pesticide effects on Carabidae and Staphylinidae (rove beetles) populations might be a result of treatments the previous year, during which some of the adult individuals, caught the current year, were at the larval stage. This, of course, weakens the possibility of obtaining significant results in a one-year pilot experiment. Only results of barley and beets are presented (Table 3.1), since no experimental sprayings were carried out in wheat. Significantly different numbers are found within a limited range of families and species of Carabidae and Staphylinidae. The strength of the results, however, in spite of the limitations of the method and statistical analysis, is a consistent picture. The significant differences found, all confirm the trend of higher catches in quarter dosage followed by half and normal dosages.
Table 3.1.
The estimated number of specific Carabidae (ground beetles)
and Staphylinidae (rove beetles) per pitfall group on Gjorslev. Numbers
are total catches from 9 May – 1 August 1996 in the barley, and 9 May – 10
October in beets. Estimates given are least squares means. Significant differences between
dosages (p<0.05, paired t-tests) are indicated by different letters. P-values of the
variable dosage are given (*: p<0.0.5, **: p<0.01, ***: p<0.001).
Crop |
Family |
Order / species |
Estimated number |
P-value |
||
Normal |
Half |
Quarter |
||||
Barley |
Carabidae |
Loricera pilicornis |
9.0 a |
8.4 a |
16.2 b |
0.0162 * |
Staphylinidae |
Philonthus spp. |
150.6 a |
152.4 a |
264.2 b |
0.0003 *** |
|
Larvae spp. |
18.8 a |
20.4 a |
49.8 b |
0.0186 * |
||
Beets |
Carabidae |
Harpalus rufipes |
14.0 ab |
6.0 a |
29.6 b |
0.0117 * |
Loricera pilicornis |
3.1 a |
6.6 ab |
12.7 b |
0.0213 * |
||
Carabus nemoralis |
0.2 a |
0.6 a |
3.4 b |
0.0162 * |
||
Direct countings
The number of A. gamma larvae per 21 plants per plot was counted 4 times during the season, and results are presented in Fig. 3.1. The counting on 16 July was carried out right before the application of dimethoate (see Appendix A for pesticide treatments). This application was not very effective and another application with Lambda-cyhalothrin was conducted 22 July. The latter application seemingly knocked down the population, and apparently even quarter dosage was enough to strongly diminish the population.
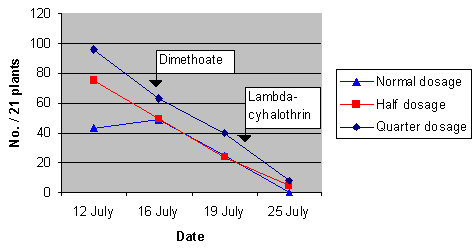
Fig. 3.1.
Estimated population densities of Autographa gamma larvae at
the three different dosage levels in sugar beets at Gjorslev 1996. The two relevant
insecticide treatments are marked with arrows.
In Table 3.2 results of the aphid countings in beets are given. Only the pirimicarb spraying had an effect, which was obviously dosage related. No insecticide sprayings were conducted in the cereal fields and only slightly differences were found in the Aphididae populations between the different dosage plots.
Tabel 3.2.
Mean percentages of sugar beets plants (Gjorslev, 1996) infested by the aphid
Aphis fabae. Dates of relevant insecticide applications are given.
Date |
Percentage of plants with Aphis fabae |
||
Normal dosage |
Half dosage |
Quarter dosage |
|
3 July |
14 |
8 |
16 |
9 July |
8 |
14 |
16 |
16 July |
Dimethoate |
||
22 July |
Karate |
||
23 July |
100 |
96 |
92 |
31 July |
100 |
100 |
92 |
4 August |
Pirimicarb |
||
8 August |
20 |
36 |
44 |
15 August |
12 |
24 |
76 |
22 August |
10 |
10 |
16 |
29 August |
0 |
4 |
6 |
3.1.4 Discussion (pilot study)
3.1.4.1 Evaluation of sampling methods
Water traps are easy to handle but suffered from the well-known problem of lacking density estimation. Besides insects blown into the traps only animals attracted by the colours and/or humidity may be caught, including species not related to the crop. The results obtained with this method are inconsistent.
Pitfall trapping. Pitfall trapping provided some interesting results. It is, however, very laborious to trap intensively at a large scale, and the subsequent sorting in the laboratory is often very demanding since catches may be high. Especially in beets there may be problems because of repeated weed hoeing. The lack of density estimates can be overcome by fencing the pitfalls thereby sampling a specific area only. This is, however, costly.
Suction sampling. Little experience was obtained due to the insufficient equipment. The method is however, widely used and accepted. The D-vac has proved to be a practical sampling device under a wide range of conditions. It is very effective at sampling most arthropod taxa, diurnally active in the vegetation layer and on the soil surface (Thomas & Marshall 1999). Suction machines may, however, not efficiently sample some life stages, e.g. juvenile stages of beneficials. Examples are larvae of Syrphidae (hooverflies) and Coccinellidae (ladybirds) (Sunderland et al. 1995). Furthermore the efficiency of suction samplers is likely to be influenced by vegetation structures and density. A significant limitation to the use of techniques involving suction samplers is that the habitat to be sampled must be dry (Sunderland et al. 1995), not only to avoid invertebrates getting stuck before reaching the collecting container, but also to ease the following sample treatment. Sampling is also often limited to daylight hours, and may therefore underestimate nocturnal species, which include most of the carabids found in agricultural habitats (Thiele 1977). The method, however, is rather independent of the activity of the sampled organism, and is therefore generally less prone to error (Thomas & Marshall 1999).
Field counting of ears with/without insects is a well-established binomial technique providing useful results for Aphididae and O. melanops. For easily recognisable A. gamma occurring in high numbers, counting the number per plant is a very useful method.
Generally, both pitfall trapping and direct counting proved useful, and it was decided to use both methods in the main phase of the study. The weakness of the pitfall sampling method in relation to obtaining absolute density estimates led, however, to the decision of downgrading the method and to use it in wheat only due to resource limitations.
Instead of pitfalls it was decided to focus on suction sampling as it provides the density estimates, which are relevant in population studies, and also enable comparisons with somehow similar investigations carried out in Denmark earlier. Based on the literature but taking the difficulties during the pilot phase into account, it was decided to develop a more powerful suction sampler than the well-known types. For practical reasons it was also decided to use suction sampling as the "back bone" sampling method throughout the investigations (see 3.2.1.1).
3.1.4.2 Discussion of results
The pitfall-sampled carabids as well as the Pirimor-sprayed Aphididae in beets showed a tendency towards higher densities at reduced pesticide dosages. The extent of the sampling combined with the lack of true replicates, however, obstruct the possibility to make any straightforward conclusions.
Regarding A. gamma there did not seem to be a pronounced dosage effect, due to the common slopes of the curves. It is, however, possible that the higher number of larvae recorded on beets plants at reduced dosages is caused by a higher weed density which may have imposed the pregnant females to lay their eggs there. The actual concentration of larvae especially in the plots half and quarter is higher than illustrated in Fig. 3.1, since larvae found on weed (not counted) contribute significant to the population.
3.2 Main studies 1997-1999
3.2.1.1 Suction sampling
In order to achieve comparable samples from a total area of at least 270 ha, it was decided to use a 4WD vehicle to transport the suction sampler between the sampling sites. The size of the suction sampler was therefore less important, opening possibilities to construct a more efficient suction sampler than the one used in the pilot studies and other commercially available models. Based on experiences from 1996 a new suction sampler was designed to extract insects from the crops at a higher level of efficiency (Fig. 3.2). Test of sampling efficiency was done with larger carabids known as difficult to extract by suction (Hald & Reddersen 1990). The test showed an 80% recapture efficiency of carabids, which was a higher rate than that of the vac-samplers (incl. the D-va®) with which it was compared. Further information will be published.
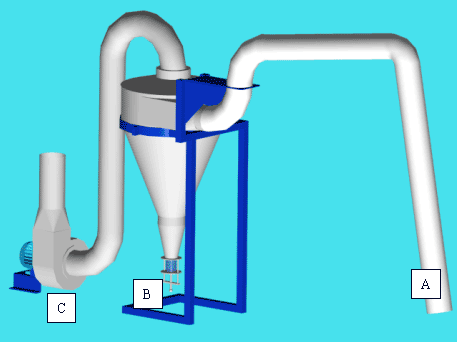
Fig. 3.2.
The suction sampler. A. Suction hose/pipe. B Collection jar. C. Fan providing
high speed air flow. The construction was mounted on a trailer with a 4WD as traction.
Many replicates are necessary to obtain useful density estimates. Within each plot, 15 sub-plots (sub-plot: ± 10 m from a marker stick) were selected. Sampling was restricted to the main crop area and plot margins were therefore excluded (minimum 20 m). Thereby interference between plots and effects from field edges was minimised.
The 15 sub-plots were evenly distributed along 3 tramlines in order to minimise crop damage. The length of the suction tubes limited sampling to within 1.5 meters from the tracks. The sampling of one field was always finished within 1 - 1.5 hour. If the sampling of a field had to be cancelled, e.g. because of rain, samples already collected were removed and a new set collected as soon as possible. All samples were collected between 10.00h and 21.00h under dry conditions. Each sample comprised 10 sub-samples of 10 seconds application of the vac-nozzle (total area 0.2 m2) in cereals. In sugar beets it was necessary to reduce the number of sub-samples to 5 (total area 0.1 m2) because large amounts of soil accumulated in the collecting jars due to the lesser plant cover compared to the cereals. Sampling in beets was only done within the rows since hardly any arthropods were observed at the almost bare field between the rows. The samples were labelled and placed directly into cooling bags. Later, the same day, they were frozen until further treatments.
Sampling was carried out 5-7 times during each season from mid May to mid August (Table 3.3). The first samples were taken in wheat at mid May, whereas sampling in barley and beets began by early June. The last samples were collected in sugar beets at mid August whereas sampling ended in early August in the cereals. If possible a sampling was conducted just before insecticide application and no samples were taken until about one week after spraying to make sure that killed arthropods were decomposed. The sampling order of the farms varied, as did the order of the fields sampled within the farms, whereas sampling within the fields always followed the same order.
Table 3.3.
Statistical information about the sampling in the experimental fields. For
each combination of crop and year, the number of sample visits is shown (average and
range). Also, the period during which the sample collections were performed is shown
together with the median date.
After defrosting the samples were sieved through 3 grids (2.0, 1.4 and 1.0 mm laboratory test sieves, Endecotts Ltd. London) to extract animals from soil and debris. The arthropods were hand-picked from the grids and transferred to 70% alcohol. All arthropods, except Aphididae and Collembola (spring-tails) found within the 3 grids, were collected but animals passing through all grids were ignored. The sample content was subsequently identified under binocular microscopes at 5 - 40 x magnification.
Araneae (spiders), Chilopoda (centripedes), Dermaptera (earwigs), Diplopoda (millipeds), Ephemeroptera (mayflies), Isopoda (woodlice), Orthoptera (grasshoppers) Opiliones (harvestmen), Plecoptera (stone flies), Psocoptera (booklice) and Thricoptera (caddis flies) were not identified further. Hemiptera were identified to at least family. Lepidoptera were identified to at least superfamily. Diptera were divided into 6 groups: 1. Asilidae (Robber flies) & Empidoidea, 2. Bibionidae (bibionid flies), 3. Syrphidae (Hoover flies), 4. Tipulidae (Crane flies), 5. Other Brachycera/Cyclorrhapha and 6. Other Nematocera. Neuroptera (lacewings) were identified to family whereas for Hymenoptera, parasitic wasps were identified as a group and the rest identified to family. Coleoptera were all identified at least to family but Carabidae imagines were identified to genus or species. All arthropods were separated into developmental stages except Aranae, Chilopoda, Dermaptera, Diplopoda and Opiliones for which life stage is not easily identified.
Collembola, an important detritivorous group of prey was not included because they require a comprehensive soil-sampling program in order to obtain reliable population estimates. Such a program was beyond the frame of this project.
Statistical analyses
Two variables were used to describe the data: 1. dry mass and 2. number of individuals. The dry mass is mainly important for arthropod – bird relations, and the numbers are mainly important when evaluating populations and predator – prey relations. Each type of data was analysed separately.
Dry mass was used as a measure of the available amount of arthropod food for birds. It is a variable relatively easy to measure making it widely used. Dry mass, however, does not take into account that the actual food quality may vary between the different arthropods. E.g., the food quality of carabids may be smaller than that of butterfly larvae due to their relatively higher cuticle content, which is of minor nutritious value.
Arthropod dry mass was estimated from the formula W = 0.0305 x L2.62 mg, where L is the length of the arthropod in mm (Rogers et al. 1977). When the arthropods were identified to species the length of adults used for the calculations was obtained from the literature (Fauna Entomologica Scandinavica) using the mean of the length intervals given. For the remaining arthropods, including juvenile stages, the length of the arthropods was obtained by measuring between 50 and 100 individuals of each relevant group (but sometimes less if occurrence was rare). Because the data were reasonable symmetrically distributed, simple arithmetic mean lengths were calculated. The individuals measured were selected randomly from all farms, crops, sampling dates and dosages. A restriction was that a maximum of 5 individuals from each relevant arthropod group was taken from one field at a certain sampling date, to ensure a representative length estimate, especially for juvenile life stages, which change in size over time. Changes in the mass of a given species because of change in life stage during a season were consequently not included in the analyses.
All arthropods are not equally important as food items for all bird species. Some species may be characterised as "important food items" because of the selective food choice of farmland birds. Their preferences are probably based on the abundance, size, availability, nutritious value and (lack of) defence/escape mechanisms of the arthropods. Skylark was the most common bird throughout the study period (Chapter 4) and several studies on its diet have been conducted, e.g. Elmegaard et al. (1994 , 1999). The food choice of this species was therefore selected as reference bird prey. The diet of Skylarks listed by Cramp & Simmons (1977-94) comprise almost all arthropods found, including even very small species like Collembola and small Ichneumonoidea (ichneumonid wasps). Such small species were not found to be relevant in a comprehensive study in Denmark of Skylark food references, because of their limited overall energy contribution (Elmegaard et al. 1999). They found that Carabidae dominated, accounting for 42% of the estimated food dry mass in faecal pellets. Lepidoptera imagines and larvae contributed with 19% and Heteroptera (bugs) 7%. The rest of the groups contributed less than 5% to the total food dry mass including Diptera, which represented < 3% in Elmegaard et al.’s results. No findings of Collembola or smaller Hymenoptera were reported although some may have been included in the < 2% "rest" group.
There is apparently no distinct line between "preferred" and "non-preferred" arthropods, making the "important food items" variable rather arbitrary. Focus, however, was put on Danish research with experiments by Elmegaard et al. (1999) as the main reference. The list of relevant arthropod food items selected in this experiment comprised Araneae and Coleoptera except Coccinellidae and Cantharidae. Others taxa included were Chilopoda, Dermaptera and Diplopoda. Among Diptera only Bibionidae and Tipulidae were relevant. Also Ephemeroptera and Hemiptera were included but among Hymenoptera only Symphyta were relevant. Finally, Lepidoptera, Orthoptera, Opiliones, Plecoptera and Trichoptera, were all on the list. Neuroptera were never identified in faecal pellets by Elmegaard and was therefore not included.
The experimental unit was a dosage plot (see chapter 1), each of which was represented by 15 samples per sampling date. These 15 samples were summarised to form the dependent variables "mg food item dry mass / 3 m2" or "mg total dry mass / 3m2" in the cereals (3 m2 equals 15 samples of 0.2 m2). In beets the sample area per plot was 1.5 m2 only. After summation, the data were loge(x+1) transformed to stabilise the variation. As described in 4.2.1.2., repeated sampling in the same plots may violate the required independence of data. To avoid this, a geometric mean of the data collected during the actual period (entire season, before/after insecticide spraying or other relevant periods within the sampling seasons) was used, leaving only one figure per combination of the variables period, farm, crop and dosage.
General Linear Models (GLM) (SAS Institute 1990) were used for the analyses of dry mass (mg food item dry mass or total dry mass / 3 m2) (1.5 m2 in beets), in order to determine a possible general effect of reduced dosages of pesticides on arthropods as a food resource. The dry mass was analysed in relation to the three class variables: year, farm, dosage as well as the interactions dosage´ farm and dosage´ year and the two numerical variables comprising the normal dose treatment intensity indices for insecticides (I-index) and herbicides (H-index). The model was extended with H-index and I-index (see Appendices A.2-A.4) to take into account that normal, half and quarter dosages were not reflecting uniform dosages between years and farms. Contradictory to the birds, arthropods may be directly affected by the actual dosage. By adding the treatment intensity indices the models were generally improved, as reflected by generally lower p-values of the models. Since sampling was restricted to the main crop area, it was not necessary to consider possible edge effects in the analyses.
In the population studies, data were separated into carnivore and non-carnivore groups, which were again separated into more specific taxonomic levels. If the juvenile stage was carnivorous, imagines were also recorded as carnivores e.g. Syrphidae. Overall the data sets were constructed as described above under dry mass analyses. The dependent variables in this case, however, were numbers of individuals / 3 m2. Data of many insect populations follow a Poisson distribution. To analyse such data, Generalised Linear Models (GENMOD) (SAS Institute 1990) with Likelihood ratio tests on loge(x+1) transformed data were used for the population analyses. The procedure can also be used for those arthropod groups not following a Poisson distribution since the difference between a Normal distribution and a Poisson distribution after loge(x+1) transformation with adjustment for over dispersion is limited. Therefore this procedure was used throughout the population analyses to make them comparable. For each analysis least squares means were estimated. When a significant dosage effect was found, t-tests were used to interpret pairwise dosage-differences.
A non-parametric test was performed to reveal if there were a significantly higher number of the most abundant arthropod groups improving under a reduced pesticide regime. The arthropod groups were divided into the six superior groups: Barley carnivores (n = 18), Barley non-carnivores (n = 17), Wheat carnivores (n = 18), Wheat non-carnivores (n = 18), Beets carnivores (n = 19) and Beets non-carnivores (n = 17)) (see Table 3.8-10). A Friedman test based on ranked data was conducted for each the superior groups using the FREQ procedure in SAS/STAT (SAS Institute 1990).
3.2.1.2 Pitfall sampling
Carabids contribute significantly to the arthropod fauna. They are important both as beneficials (Lövei & Sunderland 1996) and as food items for birds (Elmegaard et al. 1999) making it important to estimate their density and therefore to sample them efficiently. Especially larger species, however, are not easy to sample by suction since they may be nocturnal and concealed in refugia during the day. Furthermore they may occur at very low densities at the soil surface (Lövei & Sunderland 1996).
Pitfalls where chosen because they are very suitable for catching carabids. Due to resource aspects, sampling was limited to winter wheat on three farms: Gjorslev, Oremandsgĺrd and Lekkende.
In 1998 and 1999 enclosed plots were established by surrounding 10 x 10 m areas with 60 cm high metal plates buried 20 cm into the ground. The barriers made it possible to obtain estimates of the actual density of carabids in the sampling period by catching nearly all carabids within the enclosures, which at the same time secured that no ground beetles from outside of barriers could reach the traps. 4 enclosures per plot were established at least 25 m from the plot edges in normal and quarter dosages treatments. The enclosures were equally spaced along a longitudinal gradient in an attempt to minimise variation. Also the barriers were placed at the same distance from field margins if possible to maximise comparability. The establishment in normal and quarter dosages only was due to resource limitations. The enclosures were established 18 – 26 May 1998 and 12 – 20 May 1999. At this time of year it was assumed that field invasion from the wintering sites was completed.
Nine pitfalls were placed within each enclosure in a way that aimed at ensuring maximum catches. Five were placed in the middle in the same pattern as number 5 on a dice separated by four 100 cm x 15 cm metal plates positioned between each peripheral trap and the central trap, a method that increase trapping efficiency. Furthermore one pitfall was placed in each corner of the enclosures since carabids are known to follow vertical edges. In 1998 the weekly samplings were carried out in the periods 1 July to 7 August at Gjorslev and Oremandsgĺrd and 17 June to 7 August at Lekkende. In 1999 the period was 23 June to 3 August at all 3 farms. Furthermore a pre-insecticide sampling was conducted in both years in mid June. After the five-day pre-insecticide sampling period, the pitfalls were closed with lids to temporary stop further sampling within the enclosures. One week after insecticide spraying the lids were removed again and sampling continued non-stop to the end of the season.
Catches were stored in glass containers with 70% alcohol until further treatment. In the laboratory adult carabids were counted and identified at least to genus.
Statistical analyses
The pre-insecticide catches were generally low and scattered, which weakened the possibility of making a reliable analysis on the pre-insecticide application data considering the low number of replicates. These data were therefore pooled with the post-insecticide spraying samples. Two variables were used to describe the data: 1. total dry mass and 2. number per genus. Due to competition among species of the carabid-family it seemed of less value to analyse the total number of carabids caught per barrier.
As described for the suction samples (see 3.2.1.1) the dry mass was estimated using the length of the beetles. Most lengths were obtained from the literature (Lindroth 1985 & 1986) but some beetle groups (Bembidion, Trechus and Amara) were identified to genus only. For those groups an arithmetic mean of the length of at least 50 randomly selected individuals per group were used for the dry mass estimations.
The experimental unit was the enclosure in which catches were collected 5-7 times during each season. These 7 samples were summarised to form the dependent variables "mg dry mass / 100 m2" and "number per genus / 100 m2" to avoid dependent data. Summation was followed by a loge(x+1) transformation to stabilise the variance.
General Linear Models (GLM, SAS/STAT) (SAS Institute 1990) were used for both the dry mass analyses and the population analyses in order to determine a possible general effect of reduced dosages of pesticides since the log-transformed data were assumed following a normal distribution. The dependent variables were analysed in relation to the 3 class variables: year, farm and dosage as well as the interactions dosage´ farm and dosage´ year. Stepwise model reduction was used. For each analysis least squares means for dry mass and population size were estimated.
G. polygoni (knotgrass beetles) is of particular interest because it is important for farmland birds, e.g. partridge (Perdix perdix) and it is a potential control agent for its host plants Polygonum convolvulus and Polygonum aviculare (Sotherton & Moreby 1992).
At Gjorslev a high numbers of G. polygoni - adults and larvae - were caught in the fenced pitfalls in 1999. The number of adults within each enclosure was counted in order to reveal a dosage effect on the beetles. The statistical analysis was conducted as described above, but the dependent variable "no. / 100 m2" was analysed in relation to the class variable dosage only.
3.2.1.3 Direct counts
The aphid counting was conducted as described in section 3.1.1, with the exception that 100 beets plants were inspected instead of 50 on each assessment day. If possible the first inspection was carried out immediately before insecticide application and the following about one week after application to get the full effect of the application. The overall aim was not to estimate crop damage, but to reveal possible effects on the populations.
Statistical analysis
Statistical analyses were not conducted on these data. Percent infested cereal ears / beets plants are presented in Appendix D.
3.2.1.4 Sweep net sampling of Miridae
In 1997 a severe attack of Miridae (mirid bugs) occurred in sugar beets on Gjorslev. The attack was restricted to the border zone. Two days after insecticide application 40 samples per plot were taken to evaluate the effects of reduced dosages. A sample comprised 10 standardised sweeps with a butterfly net (diam. 36 cm) with one sweep pr. row in the 10 outermost rows.
Statistical analysis
The basic sampling unit was considered to be one sample. The data followed a Poisson distribution and a log-linear model was fitted, using the GENMOD procedure. Log Likelihood ratio tests were used to estimate the difference in numbers per sample between treatments. Since sampling was carried out in one field and one year only, the replicates (n = 40) were all taken from the same dosage plot and should therefore be considered as pseudo-replicates, weakening the possibility to generalise the result.
3.2.2 Results
3.2.2.1 Suction samples
Dry mass
The results of the analyses of variance are summarised in Table 3.4. The factors year and farm always had significant effects and were consequently included in all the models. The factors dosage and I-index were significant in barley. In wheat and beets no significant effects of dosage was found, and the treatment intensity indices for insecticides and herbicides only occasionally proved significant which seemed incidental. With none of the tested models significant interactions were found.
The factors year and farm constituted, not surprisingly, an absolutely dominating part of the variation, leaving only a minor part to be explained by the factor dosage. Especially in the cereals year was dominating.
Table 3.4.
Schematic summary of the dry mass analyses based on dry mass means of the entire
sampling season. I-index / H-index. (treatment intensity indices for insecticides /
herbicides, for definition see section 1.1). Statistical significance is indicated as
follows: */+/–: 0.01<p<0.05, **/++/– –: 0.001<p<0.01,
***/+++/– – –: p<0.001. A +/¸ indicates if
the correlation is positive/negative.
|
Barley |
Wheat |
Beets |
|||
Bird prey d.w. |
Total |
Bird prey d.w. |
Total |
Bird prey d.w. |
Total |
|
Dosage |
** |
** |
|
|
|
|
Year |
*** |
*** |
*** |
*** |
* |
*** |
Farm |
*** |
*** |
** |
*** |
*** |
*** |
Dosage´ year |
|
|
|
|
|
|
Dosage´ farm |
|
|
|
|
|
|
I-index |
– – |
– |
|
– |
|
|
H-index |
|
|
+ |
|
|
|
The estimated mean dry masses are presented in Table 3.5. The estimates are means over the entire season.
Significant differences of the estimated mean dry masses between dosages were found only in barley with higher dry mass at quarter dosage than in normal and half dosages that mutually were not significant different. A 30% higher dry mass of food items was found at quarter dosage than in normal dosage. A corresponding 28% difference was found for the total dry mass.
Table 3.5.
The estimated mean dry masses of relevant bird prey (based on known skylark
prey (Elmegaard et al. 1999) and the estimated mean dry masses of all
arthropods collected in the three different crops. The numbers given are least squares
estimates of the mean dry mass, of 5-7 samples per year in the period late May - mid
August. In cereals the estimates are "mg/3 m2" and in beets
"mg/1.5 m2 plants". The estimates are followed by 95% confidence
intervals. P-values for test of significance of the factor "dosage" are given
(**: p<0.01). Significant differences (p<0.05) between the different dosages are
found by paired t-tests and are indicated by different letters.
Even though no significant differences between dosages were found in wheat and beets, there was a tendency towards higher dry masses at reduced dosages. In beets there was a 13% higher estimated food item dry mass at quarter dosage than in normal dosage. In wheat a correspondingly 7% higher dry mass was indicated. Similarly in wheat a 17% higher total biomass was revealed at quarter dosage than in normal dosage and in beets a 16% higher total dry mass. In barley and beets the estimated dry masses at half dosage were closer to the normal dosage estimates than to quarter dosage but in wheat the estimated dry mass at half dosage was closer to the dry mass at quarter dosage.
To reveal possible dosage effects limited to the post insecticide treatment period, statistical tests on the wheat data were conducted on the biomass data limited to the whole period after insecticide applications. The farm and year proved significant and no significant interactions were found. A test for a dosage effect on the bird prey biomass did not show a significant effect (p = 0.4973) (estimated means and 95% confidence intervals: Normal = 410.0 [ 339.4; 495.3] mg/3 m2, half = 459.7 [ 380.6; 555.3] mg/3 m2, quarter = 475.6 [ 393.7; 574.5] mg/3 m2). A similar test on the total biomass did neither reveal significant differences (p = 0.2354) (normal = 929.6 [ 790.4; 1093.2] mg/3 m2, half = 1047.7 [ 890.9; 1232.2] mg/3 m2, quarter = 1129.1 [ 960.1; 1327.9] mg/3 m2). Statistical tests for the 14 days period after insecticide applications did not show significant dosage effects on bird prey biomass (p = 0.5649) (normal = 458.7 [ 363.2; 579.3] mg/3 m2, half = 514.0 [ 407.0; 649.1] mg/3 m2, quarter = 543.8 [ 430.6; 686.6] mg/3 m2) or on total biomass (p = 0.6953) (normal = 1118.0 [ 933.3; 1339.4] mg/3 m2, half = 1179.3 [ 984.4; 1412.7] mg/3 m2, quarter = 1243.7 [ 1038.2; 1489.9] mg/3 m2).
In beets, tests for the corresponding periods after insecticide spraying were performed but only comprising data from farms which carried out a mid-summer insecticide application. Due to the lower number of replicates the treatment intensity indices for herbicides and insecticides were not included in the model. In all the models tested, the farm and year had a significant impact but no significant interactions were found. For the entire post-insecticide period, a nearly significant dosage effect (p = 0.0755) was found on the bird prey dry mass (estimated means and 95% confidence intervals: Normal = 220.9 [ 181.7; 268.7] mg/3 m2, half = 239.0 [ 196.5; 290.7] mg/3 m2, quarter = 288.5 [ 237.2; 350.8] mg/3 m2). The test for a dosage effect of the total biomass showed significance (p = 0.0273*) (normal = 378.5 [ 298.2; 480.4] mg/3 m2, half = 406.0 [ 319.9; 515.4] mg/3 m2, quarter = 555.5 [ 437.6; 723.5] mg/3 m2). Pairwise t-tests showed significant differences between normal and quarter dosage and between normal and half dosage. Tests for the 14-day period only after insecticide spraying did not reveal a significant dosage effect on bird prey biomass (p = 0.2445) (normal = 158.7 [ 123.5; 203.9] mg/3 m2, half = 175.6 [ 136.7; 225.5] mg / 3 m2, quarter = 203.2 [ 158.2; 261.0] mg/3 m2) but a significant effect was found on the total biomass (p = 0.0130*) (normal = 209.1 [ 157.6; 277.5] mg/3 m2, half = 237.3 [ 178.8; 314.9] mg/3 m2, quarter = 353.1 [ 266.1; 468.5] mg/3 m2). Pairwise t-tests showed significantly higher dry masses at quarter and half than at normal dosage.
The generally significant effect of the dominating factor year (table 3.4) may to a large extent be explained by climatic factors, which are important for arthropod population sizes. Also different spraying intensity between years and a possible accumulating pesticide effect through the experimental years, may have contributed to the significant effect. Estimates of the parameter differences of the factor year are presented in table 3.6.
Table 3.6.
Parameter estimates of the year-differences of the bird prey
dry mass analyses.
|
Bird prey dry mass |
||
Barley |
Wheat |
Beets |
|
b 1997 - |
-1.2283 |
-1.4326 |
-0.2271 |
b 1997 - |
-1.0739 |
-1.5250 |
0.1250 |
b 1998 - |
0.1544 |
-0.0924 |
0.3521 |
From table 3.6 it appears clearly that the estimated arthropod dry mass in the cereals in 1997 was overall low. This fact may be the main reason behind the significant effect of year in the analyses of cereals mentioned above the table. In beets the highest estimated arthropod dry mass was in 1998 followed by 1997 and 1999.
To evaluate the development in the amount of available important bird prey and the total arthropod dry mass during the sampling season, the relevant dry masses were estimated in period intervals for each of the three crops. Results are presented in Fig. 3.3 and 3.4. In barley and wheat the amount of important food items remained stable with a tendency to increase during the sampling season and peaking in July. In beets, however, there was a substantial and steady increase in the amount of available prey during the season. In barley there was a tendency towards higher bird prey biomass in quarter pesticide dosage throughout the season. After the period of insecticide treatments there was a higher dry mass at half than at normal dosage. In wheat there was a higher bird prey dry mass at half dosage in the first period, mid May to mid June, before insecticide spraying. During the rest of the season more biomass was found in quarter dosage followed by half and normal dosage. In beets there were only indications of a difference in the last period, 1 - 20 August, with more bird prey biomass found at quarter than at half and normal dosage.
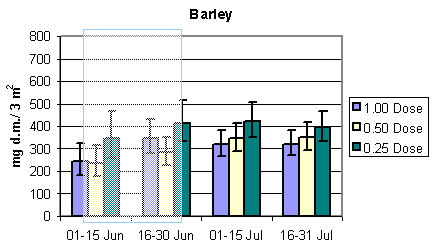
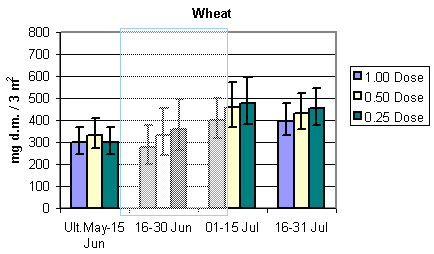
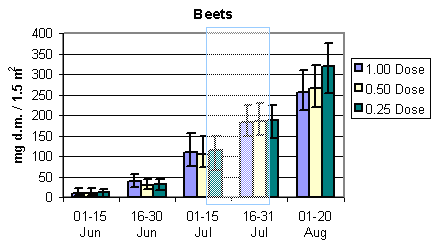
Fig. 3.3.
The development in the amount of important arthropod food for birds in the
three different crops. The shaded areas indicate periods of insecticide treatments. Notice
that data in barley and wheat are per 3 m2, whereas data in beets are per 1.5 m2
(75 plants). The error bars indicate 95% confidence intervals.
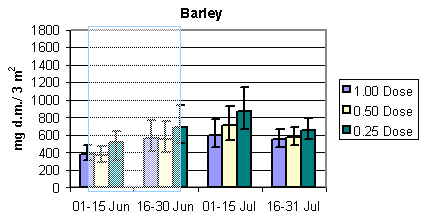
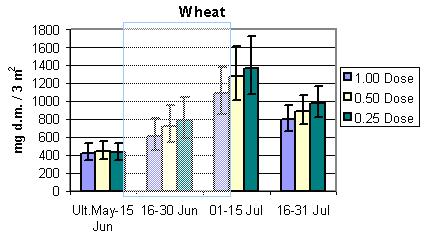
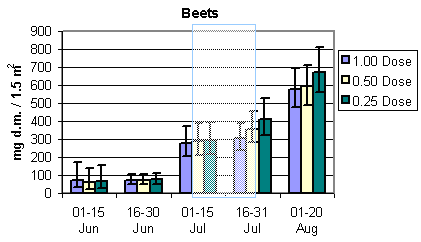
Fig. 3.4.
The development in the total arthropod dry mass in the three different crops.
The shaded areas indicate periods of insecticide treatments. Notice that data in barley
and wheat are per 3 m2, whereas data in beets are per 1.5 m2 (75
plants) The error bars indicate 95% confidence intervals.
Overall the total dry mass of arthropods was roughly about twice the amount of the bird prey dry mass. Generally, the development during the season of the total dry mass was similar to bird prey dry mass with two exceptions: 1. The increase during the season in the total dry mass in wheat was much steeper compared to the bird prey dry mass. 2. Contrary to the bird prey dry mass in beets, there seemed to be a higher total dry mass at reduced dosages for the period 16 – 31 July.
The composition of the biomass, including estimates and statistical tests for dosage effects on the most dominant orders is presented in Table 3.7. In the cereals the dominant order across dosages were Coleoptera (about 50%) followed by Diptera (30-40%) whereas the rest of the abundant orders were more evenly distributed. In beets Diptera dominated with about 50% of the dry mass and Coleoptera with 20%. Lepidoptera contributed here 10%, which was remarkably higher than in the cereals. Significant differences between dosages were found in barley for the predominant taxa Coleoptera and the undefined rest group "others". There was, however, in all crops and for all orders except Araneae a general trend towards higher biomass in quarter dosage compared to normal dosage. The dry mass estimates of half dosage did not reveal a uniform trend.
Table 3.7.
Estimated dry masses of arthropods separated into the most important orders.
The numbers given are least squares estimates of the mean dry mass of 5-7 samples per year
in the period late May - mid August. In cereals the estimates are "mg/3 m2"
and in beets "mg/1.5 m2". The estimates are followed by 95%
confidence intervals in square brackets. P-values for test of significance of the factor
"dosage" are given (*: p<0.0.5, **: p<0.01, ***: p<0.001). Significant
differences (p<0.05) between the different dosages are found by paired t-tests and are
indicated by different letters.
Population estimates
In Tables 3.8 – 3.10 density estimates of the most common arthropod groups are presented. The results of the analyses of variance of each factor for each arthropod group except dosage are excluded, since it would be too comprehensive to present. Other arthropod groups than those listed were found but their relevance to this study were considered minor, or their abundance was too low to be relevant. Aphididae are considered the most important insect pest and special emphasis will be put on their predators. Polyphagous predators like Carabidae, Staphylinidae and Araneae have often been considered of special importance because they are abundant throughout the season. Emphasis will also be put on the specialised aphid predators Syrphidae, Chrysopidae and Coccinellidae, as well as on species considered of special importance as food items. Regarding the non-carnivores, focus will be on the potential pests as well as on groups considered important food items for birds.
Table 3.8.
Barley. Mean densities of the most common carnivore and non-carnivore groups
(no./3 m2). Estimates given are least squares means with 95% confidence limits
in square brackets per sampling. Significant differences between dosages (p<0.05,
paired t-tests) are indicated by different letters. P-values for test of the factor dosage
are given (ns: P>0.05, *: p<0.05, **: p<0.01, ***: p<0.001). Abbreviations:
Col.= Coleoptera, Dip.= Diptera, Hem.= Hemiptera,
Hym.= Hymenoptera, Neu.= Neuoptera, Img.= Imagines,
Lar.= Larvae, Nym.= Nymphs, Pup.= Pupae.
Results of the population analyses in barley are presented in Table 3.8. Dosage had a significant effect on nine groups, of which seven were juvenile groups. Dermaptera, which was significantly affected by dosage, was not divided into imagines and juveniles and the only group purely of imagines, which responded significantly to dosage, was "other dipterans". The groups showing significant responses were almost evenly divided between carnivores and non-carnivores. It is noticeable that all aphid-specific juvenile predator groups Coccinellidae, Syrphidae and Chrysopidae were significantly affected by dosage with the highest densities at quarter dosage.
The population analyses of carnivores in wheat (Table 3.9) revealed results quite similar to those found in barley. The two aphid specific predators Coccinellidae and Syrphidae responded significantly to dosage but in contrast to barley the significant effect was for adult Coccinellidae. The estimates for the larvae, however, also indicated an effect with twice as high estimates at quarter than in normal dosage. Also for Dermaptera there was an effect in wheat whereas contra to barley a dosage effect was found for Carabidae instead of Staphylinidae.
Table 3.9.
Wheat. Mean densities of the most common carnivore and non-carnivore groups
(no./3 m2). Estimates given are least squares means with 95% confidence limits
in square brackets per sampling. Significant differences between dosages (p<0.05,
paired t-tests) are indicated by different letters. P-values for test of the factor dosage
are given (ns: p>0.05, *: p<0.05, **: p<0.01, ***: p<0.001). Abbreviations:
Col.= Coleoptera, Dip.= Diptera, Hem.= Hemiptera,
Hym.= Hymenoptera, Neu.= Neuoptera, Img.= Imagines,
Lar.= Larvae, Nym.= Nymphs, Pup.= Pupae.
An equal number of carnivore and non-carnivore groups revealed a significant dosage effect. The dosage effects on non-carnivores in wheat were on other groups than in barley. In wheat significant dosage effect were found for larvae/pupae of Diptera, Auchenorrhyncha and adults and nymphs of Miridae, the last one considered important bird prey.
Table 3.10.
Beets. Mean densities of the most common carnivore and non-carnivore groups
(no./1.5 m2). Estimates given are least squares means with 95% confidence
limits in square brackets per sampling. Significant differences between dosages
(p<0.05, paired t-tests) are indicated by different letters. P-values for test of the
factor dosage are given (ns: p>0.05, *: p<0.05, **: p<0.01, ***: p<0.001).
Abbreviations: Col.= Coleoptera, Dip.= Diptera, Hem.= Hemiptera,
Hym.= Hymenoptera, Neu.= Neuoptera, Img.= Imagines,
Lar.= Larvae, Nym.= Nymphs, Pup.= Pupae.
In beets (Table 3.10) the number of groups significantly affected was about evenly distributed between carnivores (5) and non-carnivores (4). The carnivore group comprised the generalists Staphylinidae and Dermaptera as well as the aphid specific specialists, Coccinellidae and parasitoid wasps, which were a broad ranged group comprising a lot of specialists. The non-carnivores affected significantly by dosage were all true herbivores: Adult Curculionidae, Meligethes (pollen beetles), Miridae and Auchenorrhyncha, all considered relevant bird prey.
A non-parametric Friedman test confirmed that a significantly higher number of the most common carnivore groups did improve at quarter dosage in all three crops (Table 3.11). Regarding the non-carnivores, significantly more groups benefited from quarter dosages in barley and beets, but significant differences were not found in wheat. Half dosage did not show any uniform pattern. For the groups "barley non-carnivores", "wheat carnivores" and "beets non-carnivores" the effect of half dosage was in between quarter and full dosage without being significantly different. For "barley carnivores" and "beet carnivores" the effect of half was nearer normal dosage.
Table 3.11.
Results of a non-parametric test performed on the data in Tables 3.8 - 10 to
elucidate if a significant number of arthropod groups benefited from reduced dosages of
pesticide applications. P-values for test of the factor dosage are given (ns: P>0.05,
*: P<0.05, **: P<0.01, ***: P<0.001). Pairwise tests were used to reveal
significant differences (p<0.05) between dosages.
Crop |
Food preference |
No. of groups |
p-value dosage |
Difference |
Barley |
Carnivores |
18 |
0.0017** |
1/4 > 1/2, 1/1 |
Non-carnivores |
17 |
0.006** |
1/4 > 1/1 |
|
Wheat |
Carnivores |
18 |
0.0032** |
1/4 > 1/1 |
Non-carnivores |
18 |
0.1603ns |
- |
|
Beets |
Carnivores |
19 |
0.0005*** |
1/4 > 1/2, 1/1 |
Non-carnivores |
17 |
0.0027** |
1/4 > 1/1 |
3.2.2.2 Pit-falls
The estimated total carabid dry mass differed significantly between quarter and normal dosages (Table 3.12). The difference in dry mass was about 25%. As for the suction samples the factors farm and especially year constituted a dominating par of the variation. A significantly higher carabid dry mass was found in 1999 accordingly to the parameter estimates (not presented).
Table 3.12.
Estimated total dry mass of adult carabids caught in 100 m2
enclosures in winter wheat in the periods 1/7- 7/8 1998 and 31/5-3/8 1999. Estimates given
are least squares means with 95% confidence limits in square brackets per sampling.
P-value for test of the factor dosage is given (*: p<0.05).
Normal dosage |
Quarter dosage |
p-value |
|
Mg dry mass / 100 m2 |
7738.7 [ 6642.3; 9015.9] |
9648.0 [ 8281.2; 11240.3] |
0.0456 * |
The genus Pterostichus dominated across dosages, contributing about 40% to the total dry mass of adult carabids (Fig. 3.5). Also Loricera, Calathus, Agonum and Harpalus contributed significantly to the carabid biomass.
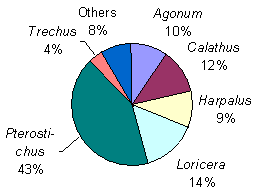
Fig. 3.5.
The composition (by dry mass) of the carabid beetle fauna caught in fenced
pitfalls 1998-99.
In table 3.13 the results of statistical analyses of possible dosage effects on the most abundant genera are presented. The results of the analyses of variance of each factor for each group except dosage are excluded, since it would be to too comprehensive to present. The population of the larger carabid Pterostichus increased significantly at reduced pesticide applications but the two genera Bembidion and Synuchus were significantly more abundant at normal dosage. Two other genera, Loricera and Demetrias, also seemed affected by dosage. The populations of both seemed to increase at reduced dosages. The conclusion of these two genera, however, was complicated by the significant interaction farm´ dosage, which revealed that a dosage effect was not found on all farms.
Juvenile stages may be more sensitive than the adult individuals. It was, however, not possible to count all larvae caught in the pit-falls but a pesticide effect on the larva, whether direct (lethal) or indirect (sublethal, changed microclimate or altered food supply), was fund by suction sampling in wheat (Table 3.9).
Table 3.13.
Mean numbers of carabids caught in pit-falls within 100 m2
enclosures in wheat in the periods 16/6-5/8 1998 and 31/5-3/8 1999. Estimates given are
least squares means with 95% confidence limits in square brackets per sampling. P-values
for test of dosage are given (ns: p>0.05, *: p<0.05, **: p<0.01, ***:
p<0.001).
Genus |
Normal dosage |
Quarter dosage |
p-value |
||
Agonum |
140.3 |
[ 118.9; 165.5] |
123.5 |
[ 104.6; 145.8] |
0.2787 ns |
Amara |
25.1 |
[ 20.9; 30.0] |
22.4 |
[ 18.7; 26.9] |
0.3961 ns |
Bembidion |
56.8 |
[ 44.3; 72.7] |
39.4 |
[ 30.7; 50.5] |
0.0416 * |
Calathus |
51.4 |
[ 38.7; 68.2] |
76.5 |
[ 57.7; 101.4] |
0.0507 ns |
Carabus |
0.4 |
[ 0.1; 0.8] |
0.8 |
[ 0.5; 1.3] |
0.0919 ns |
Clivina |
13.2 |
[ 8.2; 20.8] |
16.3 |
[ 10.2; 25.5] |
0.5235 ns |
Demetrias |
5.2 |
[ 3.9; 6.9] |
8.7 |
[ 6.6; 11.3] |
0.2924 ns |
Harpalus |
36.6 |
[ 30.0; 44.7] |
33.2 |
[ 27.1; 40.5] |
0.4833 ns |
Loricera |
139.7 |
[ 123.1; 158.5] |
218.1 |
[ 192.2; 247.4] |
0.1149 ns |
Nebria |
0.5 |
[ 0.2; 0.9] |
0.5 |
[ 0.2; 0.9] |
0.9578 ns |
Notiophilus |
9.0 |
[ 6.6; 12.0] |
8.9 |
[ 6.5; 11.9] |
0.9848 ns |
Pterostichus |
62.1 |
[ 44.4; 86.6] |
100.7 |
[ 72.2; 140.3] |
0.0444 * |
Stomis |
7.3 |
[ 5.1; 10.3] |
8.9 |
[ 6.3; 12.5] |
0.4074 ns |
Synuchus |
17.9 |
[ 13.2; 24.3] |
9.8 |
[ 7.1; 13.5] |
0.0085 ** |
Trechus |
273.5 |
[ 217.9; 343.3] |
214.5 |
[ 170.8; 269.3] |
0.1349 ns |
Others |
0.9 |
[ 0.5; 1.3] |
0,9 |
[ 0.6; 1.4] |
0.8562 ns |
Gastrophysa
A significant dosage effect was found on the number of Gastrophysa polygoni (chrysomelid beetles) caught in pitfalls (Table 3.14). Least squares estimates revealed much higher catches at quarter dosage than at normal dosage. It is unclear whether the effect was due to reduced insecticide spraying or to higher occurrence of its host plants Polygonum convolvus or P. aviculare or to a combination of various factors.
Table 3.14.
Estimated numbers of Gastrophysa polygoni adults (total
no./100m2) in wheat on Gjorslev 1999. Estimates given are least squares means
with 95% confidence limits in square brackets per sampling. P-value for test of the factor
dosage is given (*: p<0.05).
|
Normal dosage |
Quarter dosage |
p-value |
Number/100m2 |
35.4 [ 8.0; 138.6 ] |
588.2 [ 148.3; 2324.1 ] |
0.0121* |
3.2.2.3 Direct counts
Generally a dosage effect on Aphididae was found in all three crops. The aphid specific insecticide Pirimor (pirimicarb) proved more effective than pyrethoids and Dimethoate. The efficiency of the insecticide applications, however, was highly variable. A table with results is presented in Appendix D.
3.2.2.4 Sweep net sampling
The results are presented in Table 3.15, which shows a significantly higher occurrence of Miridae in quarter dosage. Since the estimates are not absolute due to the sweep net sampling method, it may be more relevant to look at the ratios between the estimated numbers. The results showed that it could be expected to find between 3 – 17 times more Miridae at quarter dosage compared to normal dosage, and between 2 - 8 times more in half compared to normal dosage.
Table 3.15.
The estimated number of Miridae (Mirid-bugs) per sample (10
standardised sweeps in the outer 10 rows) after insecticide application in beets, Gjorslev
11 July 1997. Estimates given are least squares means with 95% confidence limits in square
brackets per sampling. Significant differences between dosages (p<0.05, paired t-tests)
are indicated by different letters. The p-value for test of the factor dosage is given
(***: p< 0.001).
Normal dosage |
Half dosage |
Quarter dosage |
p-value |
0.15 a [ 0.84; 1.51] |
0.28 a [ 0.15; 0.50] |
1.13 b [ 0.84; 1.51] |
<0.0001*** |
3.2.3 Discussion
In a tri-trophic context the insect part had a dual aim. One was to research if, and to what extent, reduced dosages of pesticides, insecticides and herbicides, affected the amount of available arthropod food for the farmland birds. The other was to explore if and how much pesticides affected the populations of specific taxa of arthropods. Of special importance were populations of "beneficials", especially predators of crop pests also being important food items for birds. Dosage effects on the most important crop pests, Aphididae (aphids), were roughly estimated by counting tillers/plants with aphids. The overall aim was not to estimate crop damage, but only to reveal effects on the populations.
Possible pesticides effects could be either direct (lethal) or indirect (sublethal, changed microclimate or altered food supply). Due to the complexity of this experiment and the complexity in general it is complicated (if at all possible) to reveal the relative importance of the actual mechanism(s) causing significant findings. However, the most likely causes for effects found will briefly be discussed here and more deeply in chapter 7 and further analyses of correlation between arthropods and weed are presented and discussed in chapter 6 and 7.
Overall there was a general tendency towards more arthropod biomass at reduced dosages of pesticides (Table 3.5). There was considerable difference between the findings in the three experimental crops. In barley, a significantly higher dry mass was revealed at quarter dosage than at half and normal dosages. In wheat and beet no overall significant differences between dosages were found, but in beets a higher total arthropod dry mass was revealed at the reduced dosages after insecticide application. In barley a 30% higher total dry mass and food item dry mass was estimated between quarter and normal dosages. In wheat and beet the corresponding differences indicated were never more than about the half of that in barley. The dry mass estimates for half dosage was mostly in between the two other dosages but sometimes the estimate was nearer quarter and other times it was closer to normal dosage.
Possible reasons for the pronounced dosage effects found in barley were, that the insecticides were applied earlier and barley was a more open crop compared to wheat, allowing pesticides to penetrate deeper into the canopy thereby improving their effects (see also 7.2). Furthermore insecticides were applied more often in barley than in beets and broader ranged products were used in barley. In beets, weed hoeing was always conducted at half and quarter dosage. At Oremandsgĺrd and Gjorslev weed hoeing in normal dosage plots was carried out once per season irrespective of the number of herbicide applications. Nordfeld had done similarly at one instance (1998), while the farms Lekkende and Nřbřllegĺrd have never used weed hoeing in normal dosage plots (see Appendix B). Generally it may be assumed that soil-tilling has a negative impact on arthropods (Holm et al. unpubl.). Reasons for this could be disturbance and altered micro-climate, maybe shading the effects of reduced pesticide dosages.
Wheat had the highest arthropod biomass followed by that of barley and beets. A possible reason could be, that winter wheat is an early established, higher and denser crop probably providing a more favourable environment throughout the season. Furthermore, in wheat no soil tilling was conducted in the spring probably in favour of especially soil-dwelling arthropods. In beets the arthropod dry masses were always lower during the season when comparing with the corresponding periods in the cereals; especially at the beginning of the season. This is most likely due to the canopy development, which affects the microclimate. In beets, the long period of bare soil in early half of the season creates a rather harsh microclimate, which however changes with ongoing crop development towards being shadowy and humid. As mentioned for wheat, in winter cereals, the less extreme conditions already established in early spring creates more favourable conditions for most relevant arthropods. It should be noticed, that the arthropod estimates for beets, which had the lowest dry mass estimates until the end of the season, actually would be lower if sampling had covered not only the crop plants but also the almost bare field between the rows.
When comparing the dry mass fluctuation between years in beets with the climatic data presented in section 1.2.8 it seems likely that the relatively high precipitation in July 1998 had benefited the arthropod populations. In the cereals, in which arthropod populations are established earlier due to the crop phenology, the relatively cold May in 1997 may have suppressed the populations permanently that year. It is also possible that the relatively higher catches in the cereals in 1998 and 1999 are due to an accumulated effect of reduced pesticide dosages. It is, however, not possible to analyse such an effect isolated. Between-year differences of the amount of pesticides applied are limited and do apparently not explain the fluctuations (Table 1.1, Appendix A.2-A.4).
A non-parametric test (Friedman test) confirmed that numbers of the most common arthropod groups did increase under a reduced pesticide regime, but with the group "wheat non-carnivores" as an exception. There was a clear effect of quarter dosage, whereas there was no general effect of half dosage.
There was a tendency towards, that most affected arthropod carnivores in all three crops were aphid specific, often at juvenile stages. The populations of Dermaptera (earwigs) were higher at quarter dosage in all three crops, and they are also known as important aphid predators (Sunderland & Vickerman 1980). It is possible that this was due to prey removal, rather than a direct lethal effect on the predators. On the other hand the specific aphid predators are very exposed to insecticides due to their location high in the canopy. Furthermore the juvenile stages have limited mobility making them good indicators of pesticide effects compared to the often highly mobile adults having the ability to re-colonise quickly. The increase of the aphid specific predators responding significantly to the reduced dosages of pesticides was in the range of 20% - 175%, most pronounced for Coccinellidae larvae in barley. The populations, however, were probably still too low to have a significant impact on the aphid populations. Among the non-carnivores, it was generally not the same non-carnivore groups, which were significantly affected by dosage in the three crops, however the number of groups significantly affected was the same (4).
A dosage effect on Aphididae was found in al three crops. Aphids are considered the most important crop pests and they are the main targets of a majority of the insecticide applications. The aphid specific insecticide Pirimicarb, which is considered less harmful to most arthropod predators, proved more effective than pyrethoids and Dimethoate in all three crops. The absolutely lowest damage threshold in barley and wheat is a 30% ear infestation at the most vulnerable growth stages (Nielsen et al. 2000). Therefore, the insecticide applications in both cereals in 1997 and in wheat in 1998 could probably have been omitted. The other insecticide sprayings in the cereals seemed justified. Despite a high variation in the efficiency of the applications, which blurs the overall picture, quarter dosage seemed close to the required minimum. In beets all the Pirimicarb applications proved efficient, even at quarter dosage, contrary to the other insecticides.
With fenced pitfalls a significantly higher estimated dry mass (25%) of the important carabids was found at quarter dosage in wheat. The results obtained on carabids using fenced pitfalls are not reflected in the suction samples probably because carabids are poorly extracted by suction. It was not possible to conduct corresponding experiments in barley and beets due to resource limitations (and the ongoing weed hoeing in beets). It is, however, most possible that the results obtained in wheat could be found in barley and beets too. This is an interesting hypothesis since Carabidae is a very dominating family within Coleoptera, which already constitute a significant part of the dry mass of the suctions samples. A significant effect for carabids in barley and beets could therefore turn the overall tendency even more towards a clear dosage effect. The most abundant genus Pterostichus responded positively to reduced dosages with 62% higher density at quarter dosage. The most abundant Pterostichus species was P. malanarius which is a medium to large sized species. It is a widely studied species, known as an important predator of many crop pests.
Because the pre-insecticide catches were insufficient for statistical analysis, it was not possible to distinguish between insecticide or herbicide effects on the adult populations. Furthermore the life cycle of carabids, with larval stages in the soil having different emergence periods and consequently population fluctuations difficult to access, complicates the conclusions. However, since larger carabids generally are not very sensitive to insecticides at the applied dosages, weed cover may play a key role in the differences found between dosages. P. melanarius is nocturnal and prefers probably a dense plant cover as found at quarter dosage, whereas e.g. the most abundant Bembidion species are diurnal and may therefore prefer the less dense plant cover found at normal dosage (see 7.2). The pesticide effects found on the adult beetles may also be due to lethal effects on the larvae, especially on those with a pronounced epigaeic activity. A species, which apparently was affected by a differentiated spraying regime, was Loricera pilicornis, which has epigaeic activity during the period of insecticide spraying (Traugott 1998). It is possible that the significantly higher catches in 1999 were due to an accumulated effect of reduced dosages, but it cannot be documented statistically.
The significantly higher numbers of Gastrophysa polygoni (knotgrass beetle) at quarter dosage (estimated number 15 – 16 times higher, but with high variation) on Gjorslev 1999 is in line with the results of Kjćr & Jepson (1995) who found increased populations at reduced field rates of dimethoate. G. polygoni is found on the aerial parts of its host plants and is therefore directly exposed to the spraying droplets. This probably makes it very sensitive to the pyrethoid spraying (Tau-fluvalinat) conducted. It may also be because of an increased host plant resource although this was not verified. Also when herbicides do not kill the host plants, but only limit their growth and quality, the abundance of G. polygoni may be severely reduced (Sotherton 1982, Kjćr & Elmegaard 1996).
Even isolated it is a very interesting case since earlier research have documented G. polygonum (as well as its host plants) as a key factor for the partridge Perdix perdix (Sotherton 1982).