Buffer zones for biodiversity of plants and arthropods: is there a compromise on width?
3 Results
- 3.1 Flora
- 3.2 Arthropods
- 3.3 The marginal gain of diversity at increased buffer width
- 3.4 Combined flora and arthropod analysis
3.1 Flora
3.1.1 Hedge
The hedgerows (Appendix B, Table B.3.) of the four fields, did not differ significantly with respect to species composition for woody plants (P=0.9457, one-way ANOVA) or for dominant herbs (P=0.7365; P=0.9010 and P=0.7532 respectively for each sampling run). However, despite the lack of statistical difference, the hedge in AM differed from the other three hedgerows by being dominated by roses (Rosa spp.) (see Table B.3 in appendix B).
3.1.2 Hedge bottom and field
All plant species present in the field and the hedge-bottom are presented in Appendix B, Tables B.1 and B.2 with the abundance given for each combination of distance and buffer zone width. Results of the statistical analysis on weed densities in the field are presented in Table 3.1. The densities of all recorded weeds in the field are presented in Fig. 3.1. The figure shows no change in number of weed plants with distance from the hedge, with a buffer width 0 m. At buffer 24, however, the number of weed plants increased with proximity to the hedge. Increasing buffer width resulted in higher number of weeds with distance from the hedgerow.
Table 3.1. Schematic summary of the statistical analyses on abundance of the wild flora in the field at the second sampling run in July. Monocots are all individuals of the monocotyledonous species , Dicots are all individuals of dicotyledonous species.
| Order | Family | Run² | Test results F(ndf,ddf)P¹ | |||
| Field³ | Distance4 | Buffer5 | Buffer × Distance6 |
|||
| Monocots | All | 2 | 21.31(3,14)*** | 5.52(4,11)* | 5.05(4,12)* | 1.99(16,52)* |
| Poaceae | 2 | 21.31(3,14)*** | 5.52(4,11)* | 5.05(4,12)* | 1.99(16,52)* | |
| Dicots | All | 2 | 13.36(3,12)*** | 6.77(4,11)** | 8.08(4,16)*** | 5.16(16,43)*** |
| Apiaceae | 2 | 51.15(3,16)*** | 4.49(4,7)* | 0.76(4,8) NS | 6.85(16,52)*** | |
| Asteraceae | 2 | 4.57(3,11)* | 15.54(4,15)*** | 3.08(4,55)* | 2.63(16,47)** | |
| Brassicaceae | 2 | 2.83(3.20)NS | 2.45(4,13)NS | 3.49(4,16)* | 3.90(16,51)*** | |
| Chenopodiaceae | 2 | 20.66(3,9)*** | 3.26(4,7)NS | 7.20(4,11)** | 4.99(16,55)*** | |
| Lamiaceae | 2 | 3.83(3,16)* | 7.93(4,13)** | 2.88(4,26)* | 1.55(16,51) NS | |
| Scrophulariaceae | 2 | 0.67(3,14)NS | 3.07(4,11)NS | 0.86(4,19) NS | 3.63(16,47)*** | |
| Violaceae | 2 | 9.94(3,16)*** | 0.91(4,11) NS | 2.06(4,11) NS | 3.33(16,45)*** | |
| All | All | 2 | 30.14(3,13)*** | 9.86(4,14)*** | 14.48(4,62)*** | 3.61(16,62)*** |
¹ NS not significant, *P < 0.05, **P < 0.01, ***P < 0.001, F is the F-value, ndf and ddf is the numerator and denominator degree of freedom used for testing the significance.
² The second sampling round was carried out from 24 June.
³ Effect of field (four fields were included in the experiment).
4 Effect of distance from field edge (sampling was carried out 2, 5, 9 and 18 m from the field edge).
5 Effect of buffer width (0, 4, 6, 12 and 24 m).
6 Effect of the combination of distance and buffer width.
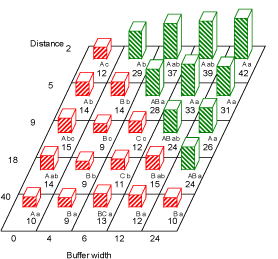
Fig. 3.1. Estimated total weed numbers (plant no. per m²) at the second sampling run (July)at the distances 2 ,5 ,9 ,18 and 40 m to the hedgerow at the buffer widths 0, 4, 6, 12 and 24 m. Within each buffer width, figures with the same capital letter are not significantly different (P=0.05). Within each distance, figures with the same lower case letter are not significantly different (P=0.05). Red bars (hatched from lower left to upper right) are numbers in areas treated with fertilizer and pesticides. Green bars (hatched from upper left to lower right) are non-treated area (buffer zone).
Monocotyledonous weeds (monocots)
For monocots (non-sensitive to the applied herbicide), there were significant effects of field, buffer zone and distance, as well as the interaction between buffer zone and distance (Table 3.1 and Fig. 3.2). There was a tendency towards more monocot weeds with increasing buffer width. The number of monocots seemed to decrease with distance from hedge. However the effect seemed to depend on the buffer width, and was only significant for some combinations of buffer width and distance – probably because of the low number of monocots and the dicot-selective herbicides used in the experimental period.
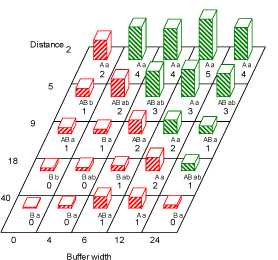
Fig. 3.2. Number of monocotyledoneous weed plants (no. per m²) at the second sampling run (late June-July)at the distances 2 ,5 ,9 ,18 and 40 m to the hedgerow at the buffer widths 0, 4, 6, 12 and 24 . Within each buffer width, figures with the same capital letter are not significantly different (P=0.05). Within each distance, figures with the same lower case letter are not significantly different (P=0.05). Red bars (hatched from lower left to upper right) are numbers in areas treated with fertilizer and pesticides. Green bars (hatched from upper left to lower right) are non-treated area (buffer zone).
Dicotyledonous weeds (dicots)
For dicots there were significant effects of field, distance, buffer zone and the interaction between distance and buffer zone (Table 3.1). The total number of dicots at the second sampling run seemed mainly to depend on whether the area was treated or not (Fig. 3.3). Buffer 4 was the narrowest buffer width to deliver significantly higher densities of dicots compared to treated field. Beyond distance 5 m the effect of buffer width was less clear but still revealing a tendency towards more dicots with increasing buffer width (Fig. 3.3).
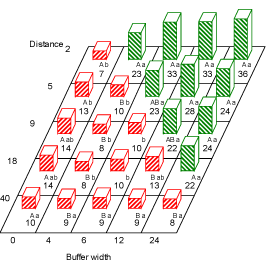
Fig. 3.3. Number of dicotyledoneous weeds (no. per m²) at the second sampling run (late June-July) at the distances 2, 5, 9, 18 and 40 m to the hedgerow at all the buffer widths: 0, 4, 6, 12 and 24 m. Within each buffer width, figures with the same capital letter are not significantly different (P=0.05). Within each distance, figures with the same lower case letter are not significantly different (P=0.05). Red bars (hatched from lower left to upper right) are numbers in areas treated with fertilizer and pesticides. Green bars (hatched from upper left to lower right) are non-treated area (buffer zone).
Weeds according to family
For all families, except Lamiaceae, a significant interaction between distance and buffer zone width (Table 3.1) was found. The effects of buffer width, distance from hedge and the interaction between those are visualised in Fig. 3. 4. For Apiaceae and Poaceae, the interaction seemed partly to be caused by an apparent missing effect of buffer widths for some distances. For Asteraceae, Chenopodiaceae and Scrophulariaceae the interaction was probably partly caused by very few weeds in some plots, and partly from the difference between treated and untreated areas. For Brasicaceae, the interaction seemed to be caused mainly by a difference between treated and untreated areas. For Lamiaceae, there was much higher number of weeds at distance 2 m than at the other distances. For Violaceae, a low number of weeds were found for buffer 0 at 2 m from the hedge. Otherwise the number of weeds seems to be relatively homogeneous over the area, but with a tendency to higher numbers in untreated areas than in treated areas.
The crop
The spring barley crop responded significantly to management with fertilization and pesticides. The crop cover, the crop height and the growth stage was smaller in the buffer zone than in the conventional field. The same number of crop plants had established in treated and non-treated areas (data not shown) (Table 3.2).
Table 3.2. Spring barley cover, height and growth stage (BBCH) at first (from 27 May) and second sampling run (from 6 July). Significant effects (one-way ANOVA) of management) are indicated as follows: † for P < 0.1; * for P <0.05; ** for P < 0.01 and *** for P < 0.001.
| Treatment | Cover (%) | Height (cm) | BBCH | |
| First run | + | 94 *** | 36 † | 22.5 * |
| First run | - | 26 | 27 | 19 |
| Second run | + | 80 ** | 72 * | 77 |
| Second run | - | 53 | 62 | 77 |
3.1.3 Buffer zone effects on floral biodiversity
Species richness and Shannon´s H in hedge bottom and field
In the analyses on plant densities above, it was not possible to include data from the hedge bottom because the data were sampled as percent ground cover, and data sampled in the field were a density per. m². However, as the number of species were recorded both in hedge bottom and field, it was possible to combine the data within the biodiversity analyses.
For both Shannon’s H and number of weed species there were significant effects of both buffer width, distance to hedge, sampling time and interaction between these. The mid-field references at 40 m (all treated with pesticides and fertilizer) had a lower value than the mean of the other plots, as could be expected.
The number of weeds at sampling run 2 for buffer 4, 6 and 12 showed a rather steep decrease with increasing distance from the buffer zone margins and outwards, while buffer 24, with no records just outside the zone margin, showed a less steep decrease with distance – more equal to the general tendency at sampling run 1 (Fig. 3.5). For both sampling runs the biodiversity were generally larger for untreated than treated plots. Buffer 0 showed a steep decrease in plant numbers immediately outside its margins at both sample runs. The data used in the Fig. 3.5 are shown in Table 3.3. This table can also be used for pairwise comparisons of differences between buffer widths and distances.
Table 3.3. Estimated values of Shannon H and number of weed species for combinations of distance to hedge, buffer width and time.
| Distance, m | Buffer width, m | Shannon H | No of wild plant species | ||
| Run 1 | Run 2 | Run 1 | Run 2 | ||
| 0 | 0 | 1.38 | 1.42 | 5.60 | 6.00 |
| 4 | 1.02 | 1.22 | 3.88 | 5.08 | |
| 6 | 1.26 | 1.26 | 4.88 | 5.53 | |
| 12 | 1.21 | 1.32 | 4.75 | 5.58 | |
| 24 | 1.13 | 1.16 | 4.40 | 4.58 | |
| 2 | 0 | 0.66 | 0.49 | 2.73 | 2.15 |
| 4 | 0.90 | 1.31 | 3.70 | 5.83 | |
| 6 | 0.97 | 1.41 | 4.20 | 6.73 | |
| 12 | 1.06 | 1.38 | 4.63 | 6.75 | |
| 24 | 0.94 | 1.27 | 4.33 | 6.30 | |
| 5 | 0 | 0.49 | 0.61 | 2.10 | 2.38 |
| 4 | 0.66 | 0.43 | 2.55 | 1.98 | |
| 6 | 0.91 | 1.23 | 3.23 | 4.83 | |
| 12 | 1.02 | 1.30 | 3.70 | 5.90 | |
| 24 | 0.87 | 1.09 | 3.45 | 4.75 | |
| 9 | 0 | 0.51 | 0.41 | 2.13 | 1.80 |
| 4 | 0.52 | 0.35 | 2.10 | 1.65 | |
| 6 | 0.68 | 0.57 | 2.50 | 2.03 | |
| 12 | 0.91 | 1.25 | 3.10 | 5.20 | |
| 24 | 0.86 | 1.07 | 3.43 | 4.53 | |
| 18 | 0 | 0.42 | 0.38 | 1.85 | 1.73 |
| 4 | 0.42 | 0.43 | 1.63 | 1.68 | |
| 6 | 0.41 | 0.42 | 1.73 | 1.68 | |
| 12 | 0.45 | 0.47 | 2.23 | 2.05 | |
| 24 | 0.63 | 0.93 | 2.43 | 4.03 | |
| 40 | All | 0.41 | 0.40 | 1.78 | 1.55 |
| LSDa | Horizontal | 0.25 | 0.84 | ||
| LSDb | Other | 0.38 | 1.38 | ||
a) If the difference between the two sampling runs for the same plot (combination of buffer and distance) are larger than the LSD-value, then the parameter has changed significantly (at the 5% level) from run 1 to run 2.
b) If the difference between any pair of plots at the same sampling run are larger than the LSD-value then the variable are significantly different (at the 5% level) for those two plots. This LSD-value can similarly be used to compare a plot at run 1 with another plot at run 2.
Shannon´s biodiversity index modelled by a logistic function
In order to be able to interpolate the biodiversity index (Shannon´s H) to other distances than the measured, and to estimate the distance at which the biodiversity was reduced to half its value at the hedge, empirical models based on the logistic model was developed (see section 2.6.1 and Model 5 in Appendix F). For each sampling run, a full model with two parameters for each buffer zone (a parameter describing the distance at which the index is halved and the slope for each buffer zone) and a simplified model (with a common slope for all buffer zone) was estimated. The estimates of the parameters for both models and both sampling runs are shown in Table 3.4. The full model did not explain the data more sufficient than the simplified model (se the row AIC of Table 3.4) and therefore the simplified model, with a common slope (Model 5 of Appendix F) were applied for producing Fig. 3.6.
The biodiversity (Shannon´s H) at the hedge and in the middle of field was almost identical at both sampling runs (about 1.2 and 0.4, respectively) and the value in the field were for both sampling runs reduced to about one third of its value at the hedge. At sampling run 2, the effect of the different buffer width had an effect that reached further out into the field (almost 5 times further, the parameter β0) than at sampling run 1, and this seemed to be the most pronounced difference between the two sampling runs. The distances at which the biodiversity index was halved increased with buffer width but did not vary significantly from sampling run 1 to sampling run 2, although there seemed to be a steeper increase with buffer zones at sampling run 2 than at sampling run 1. For both buffer 12 and 24 at sampling run 1, the biodiversity index was halved at about 11 m from the hedge, whereas 13 m and 19 m, respectively, were needed to halve the number of species at buffer 12 and 24 sampling run 2. Part of this difference (although not significant) may have been caused by the larger number of species (mainly/partly because the plants had developed and more plants could be identified to species) at sampling run 2 than at sampling run 1.
Table 3.4. Estimated parameters of the logistic model (both Model 1 and 2 presented) for Shannon´s biodiversity index at each sampling run (time) separately. At the bottom, the halving distances db in m, (and its 95% confidence intervals) at which Shannon´s index has decreased by half of its value form the value of the hedge bottom for each bufferzone width. StdE = Standard Error of estimate.
| Time | Sampling run 1 | Sampling run 2 | ||||||
| Model | 1 (Full model) | 2 (Simplified model) | 1 (Full model) | 2 (Simplified model) | ||||
| Parametera | Estimate | StdE | Estimate | StdE | Estimate | StdE | Estimate | StdE |
| β0 | 2.02 | 1.50 | 2.05 | 2.24 | 9.96 | 13.09 | 3.46 | 5.33 |
| β4 | 1.41 | 1.02 | 10.15 | 129.5 | ||||
| β6 | 2.24 | 1.86 | 5.22 | 2.07 | ||||
| β12 | 4.98 | 5.75 | 7.45 | 4.20 | ||||
| β24 | 0.75 | 0.60 | 0.55 | 0.51 | ||||
| γfield | 0.46 | 0.12 | 0.45 | 0.11 | 0.43 | 0.06 | 0.41 | 0.06 |
| γhedge | 1.12 | 0.09 | 1.13 | 0.07 | 1.27 | 0.04 | 1.32 | 0.05 |
| δ0 | 0.17 | 0.45 | 0.15 | 0.42 | 0.15 | 0.38 | -0.32 | 0.99 |
| δ4 | 1.13 | 0.52 | 1.00 | 0.52 | 1.03 | 0.80 | 0.72 | 3.96 |
| δ6 | 1.91 | 0.39 | 1.89 | 0.35 | 2.04 | 0.23 | 1.87 | 0.14 |
| δ12 | 2.38 | 0.31 | 2.37 | 0.21 | 2.59 | 0.34 | 2.49 | 0.18 |
| δ24 | 2.39 | 0.46 | 2.35 | 0.73 | 2.93 | 0.08 | 3.48 | 1.29 |
| σA² | 0.011 | 0.011 | 0.010 | 0.008 | 0.006 | 0.006 | 0.008 | 0.006 |
| σD² | 0.065 | 0.010 | 0.063 | 0.009 | 0.049 | 0.007 | 0.045 | 0.007 |
| AIC | 38.7 | 43.1 | 10.1 | 10.2 | ||||
| d0 | 1.2 a (0.4-3.4) | 1.2 a (0.4-3.1) | 1.2 a (0.5-2.9) | 0.7 a (0.1-7.6) | ||||
| d4 | 3.1 ab (0.9-10.5) | 2.7 ac (0.8-9.2) | 2.8 abc (0.4-18.7) | 2.0 ab (0.0-24000) | ||||
| d6 | 6.7 ab (2.7-16.9) | 6.6 ac (2.9-15.3) | 7.7 bd (4.4-13.3) | 6.5 b (4.6-9.1) | ||||
| d12 | 10.8 b (5.2-22.3) | 10.7 bc (6.5-17.5) | 13.4 cd (6.0-29.9) | 12.1 b (7.9-18.5) | ||||
| d24 | 10.9 b (3.7-32.7) | 10.4 ab (1.9-58.0) | 18.8 c (15.6-22.6) | 32.6 b (1.5-695) | ||||
a The parameters with Greek letters are parameters of the statistical model (Model 5 of Appendix F): β0-β24 are the coefficients for the exponential effects. γfield and γhedge are the estimated biodiversity (Shannon´s H) in the field and hedge, respectively. δ0-δ24 are the constant effects of each buffer width. AIC is a measure for comparing model 1 and model 2 (a small value is best) (Akaike, 1974). The d0-d24 are estimates (with confidence limits) of the distance at which the biodiversity index (Shanons H) has been reduced to half it value at the hedge bottom. Halving distances followed by the same letter are not significant different (P≥0.05).
At sampling run 1, a buffer width of 12 m was necessary in order to obtain a significantly higher halving distance compared to buffer 0 (Table 3.4). However, at sampling run 2 (were the wild flora had developed and more plants could be identified to species), a buffer width of 6 m was sufficient to get a significantly higher halving distance compared to buffer 0 (Table 3.4).
To get a significantly higher halving distance compared to buffer 6 at sampling run 2, a buffer width of 24 m was needed (Table 3.4).
3.1.4 Flowering in hedge-bottom and field
The percentages of flowering plants in the hedge bottom are presented in Table 3.5. There was no significant effect of buffer zones on the flowering percentages within the hedge bottom, but for the monocots (grasses) there seemed to be a tendency towards increased flowering at the widest buffer zones (12 and 24 m) compared to the more narrow buffers (0 – 6 m).
Table 3.5. Percent flowering plants in the hedge bottom in July (sampling run 2).
| Test taxa | Buffer 0 | Buffer 4 | Buffer 6 | Buffer 12 | Buffer 24 |
| All wild plants | 8 a¹ | 11 a | 11 a | 12 a | 11 a |
| Dicots | 15 a | 20 a | 23 a | 17 a | 24 a |
| Monocots | 4 a | 5 a | 3 a | 13 a | 13 a |
¹ Estimates within each row followed by the same letter are not significantly different (P≥0.05).
The flowering percentages of all plants in the field and the dicots in the field were significantly related to buffer width, distance to hedge and the interaction (Table 3.6). The dicots in the field area showed also a significant effect of field (Table 3.6).
Table 3.6. Schematic summary of the statistical effects on flowering percentages.
| Test taxa | Test results F(ndf,ddf)P¹ | |||
| Field | Buffer | Distance | Buffer ´ Distance | |
| All wild plants | 5.29(3,3)NS | 30.87(4,31)*** | 13.63(3,33)*** | 9.54(12,28)*** |
| Dicots | 19.51(3,3)* | 27.32(4,25)*** | 17.07(3,35)*** | 10.41(12,25)*** |
¹ NS not significant, *P < 0.05, **P < 0.01, ***P < 0.001, F is the F-value, ndf and ddf is the numerator and denominator degree of freedom used for testing the significance.
Within the field, the wild plants were flowering vividly in the buffer zones but not in the treated (fertilized and sprayed) field (Fig. 3.7).
3.2 Arthropods
3.2.1 Hedgerow
In hedgerow woody species, a total of 29,577 arthropods were sampled in beating trays. Only orders and families in which significant effects of buffer zone width were found are treated below. Arthropods sampled in hedgerow trees are presented in Appendix C, with sums of numbers collected in each buffer zone.
Araneae
Across hedgerow woody species, there were neither significant trends for the number of spider individuals versus buffer width nor the number of spider families versus buffer width.
Shannon´s H was significantly higher for buffer 0 when compared with all other buffers in period 1(t= 2.2, df=42, P=0.04 Fig. 3.8).
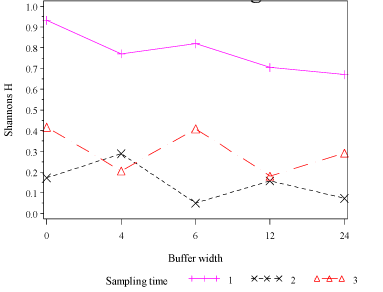
Fig. 3.8. Shannon´s H for Araneae in hedgerow trees in buffer widths 0, 4, 6, 12 and 24 m. For period 1, Araneae diversity was highest in buffer 0 (no buffer zone). In periods 2 and 3, after pesticide had been used, there were no significant differences.
In hawthorn, numbers of the family Araneidae were significantly affected by buffer width in period 3 (July) ((F=3.5, df=34, P=0.02). Tukeys test for pairwise comparison showed that there were significantly more spiders in buffer 24 than in buffer 12 (t=2.00, P=0.03). For other buffer widths, there is no clear trend indicating higher numbers or diversity with increasing buffer width (estimates for numbers in buffers 0, 4, 6, 12 and 24 were: 0.7, 0.1, 0.7, 0.2 and 1.1).
Hemiptera
There was no overall significant effect of buffer width on Hemiptera numbers or on Hemiptera species diversity in hedgerow trees, though for period 2, a trend towards more Hemiptera with wider buffers is seen(Fig. 3.9).
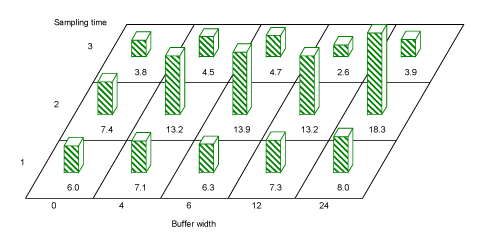
Fig. 3.9. Average Hemipteran numbers caught per sample in hedgerow trees in buffer widths 0, 4, 6, 12 and 24 m. A comparison of buffer 0 against all other buffers, showed that in period 2 there were significantly fewer Hemiptera in buffer 0. A pairwise comparison of Hemiptera numbers showed significantly more Hemiptera in buffer 24 than in buffer 0.
A comparison of buffer 0 against all other buffers, showed that in period 2 there were significantly fewer Hemiptera in buffer 0 (t=-2.52, df=17.3, P=0.02) than in buffers 4, 6, 12 or 24 m. A pairwise comparison of Hemiptera numbers in hedgerow woody species protected by different buffer widths, showed significantly more Hemiptera behind a 24 m buffer than behind a 0 m buffer (t=-2.67, df=14.2, P=0.02).
In blackthorn Hemiptera numbers were significantly affected by buffer at time 2 (P < 0.04) (estimates for buffers 0, 4, 6, 12 and 24: 10.2‚ 22.6‚ 16.6‚ 16.4 and 9.1). In hawthorn Hemiptera numbers were significantly higher in buffer 4 than 0 at time 2 (P=0.05)(estimates for buffers 0, 4, 6, 12 and 24: 14.3‚ 29.5‚ 32.7‚ 24.2 and 27.2).
Across tree species, buffer width significantly affected the number of aphids found within the hedgerows in period 1(May) and period 2 (June) (F=2.73, df=12, P=0.03 and F=4.84, df=11, P=0.02, respectively) (Fig. 3.10), with more aphids found where the buffer was wider. A pairwise comparison using Tukeys test showed significantly more aphids on hedgerow trees behind a buffer of 24 m than one of 0 m in Period 2 (estimate -1.2, df=12, P=0.004).
Hedgerow living aphids are mostly specialists on specific tree species. For example hazel is the only host of Corylobium avellana and Myzocallis coryli. Some winged specimens of Rhopalosiphum avenae were also found in the hedgerows. The trend of increasing numbers with increasing buffer width was also observed for the winged R. avenae (See Appendix C).
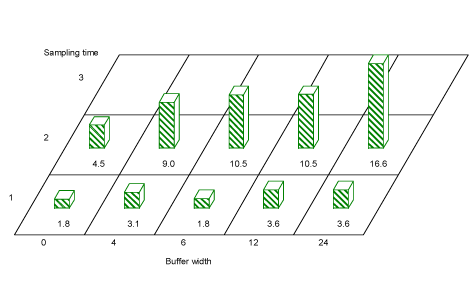
Fig. 3.10. Average aphid numbers caught per sample in hedgerow trees in buffer widths 0, 4, 6, 12 and 24 m. Both for period 1 in May (sampling time 1) and for period 2 in June (sampling time 2) there was a significant effect of buffer width on the number of aphids caught. For Period 3 (sampling time 3) there were too few aphids for a statistical analysis. The majority were tree living aphids, but a few Rhopalosiphum avenae were also caught.
The Heteroptera species number in buffer 0 versus all other buffer widths was 60% lower across sampling dates, with estimated species numbers of 0.4 at buffer 0 m, 0.7 at buffers 4, 6 and 12 and 0.8 at buffer 24, but the difference was not significant (df=42, P=0.14).
In blackthorn the numbers of Heteroptera were significantly affected by buffer width × period (F=3.86, df=31, P=0.01) (estimates for buffers 0, 4, 6, 12, 24 in period 1: 0.7‚ 0.6‚ 0.3‚ 0.6, 0.6 and in period 2: 0.3‚ 0.7‚ 0.7‚ 1.0, 0.9 and in period 3: 3.4‚ 2.3‚ 2.7‚ 0.7 and 3.0), likewise a highly significant effect of buffer width × period was found on the Shannon´s H for Heteroptera species diversity in blackthorn (F=8.08, df=13, P=0.0006).
A trend of higher number of Miridae, the most important family in the Heteroptera, with increasing buffer width was seen on roses in period 3 (estimates: 1.1‚ 1.7‚ 2.1‚ 2.3 and 4.4 respectively). However, since roses were only sampled in one field, AM (Andersmark), data cannot be statistically analysed.
Coleoptera
Overall, the order of Coleoptera was not significantly affected by buffer width either in numbers of individuals, species or diversity (Fig. 3.11).
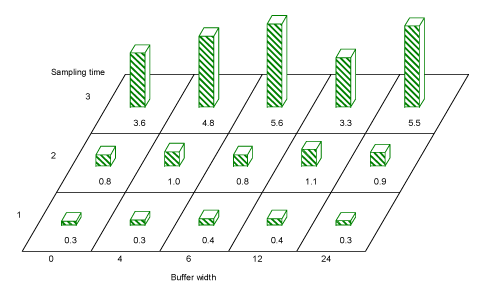
Fig. 3.11. Average Coleoptera numbers caught per sample in hedgerow trees in buffer widths 0, 4, 6, 12 and 24 m. Both for period 1 in May (sampling time 1) and for period 2 in June (sampling time 2) there was a significant effect of buffer width on the number of aphids caught. For Period 3 (sampling time 3) there were too few aphids for a statistical analysis.
However, a comparison of buffer width 0 m against all other buffer widths, found that in period 2 there were significantly fewer Coleoptera in hedgerow treatments without any buffer than with a buffer zone (t=-2.54, df=180, P=0.01). A pairwise comparison of Coleoptera numbers in hedgerow trees protected by different buffer widths, showed a significant difference between 0 m and 12 and 24 m (t=-2.28, P=0.02 and t=-2.54 , P=0.01, respectively, both df =180)
On the family level the effect of buffer width at period 3 was significant for Nitidulidae (F=.74, df=12, P=0.001) and Curculionidae (F=.33, df=12, P=0.049). There were significantly more Nitidulidae in buffers 6 m and 24 m than in buffer 0 m (Tukeys test for pairwise comparisons) (df=13, P=0.001 and df=13, P=0.006) (Fig. 3.12a). For Curculionidae there was no clear trend towards more individuals at increased buffer width (Fig. 3.12b). Curculionid diversity (Shannon´s H) at time 3 was less at buffer 0 than at other buffers (4, 6, 12 and 24 m), though not significantly so (df=45, P=0.07).
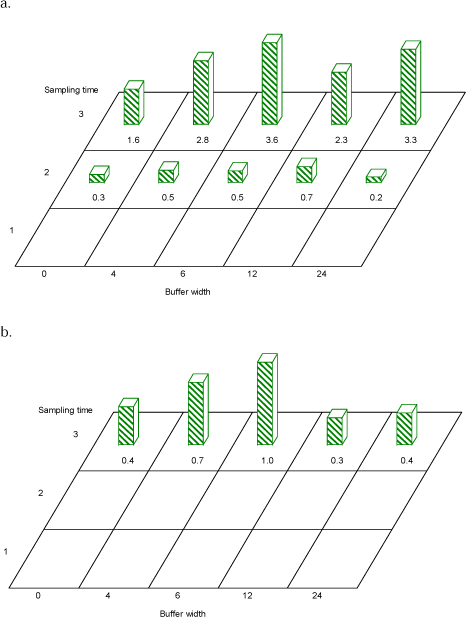
Fig. 3.12. Average numbers of a) Nitulidae and b) Curculionidae caught per sample in hedgerow trees in buffer widths 0, 4, 6, 12 and 24 m. In period 3 (July) there was a significant effect of buffer width on the number of Nitidulidae and Curculionidae caught. For Period 1 too few Nitidulidae were caught for a statistical analysis. Curculionid numbers could only be analysed for period 3.
On blackthorn there was a significant effect of buffer on Coccinellid numbers (F=3.56, df=15, P=0.03). In July 30 % more coccinellids were found in hedges with a buffer zone than without (buffer 0 compared to all treatments) (df=40.5, t=-2.07, P=0.04).
Chick-food
There were no significant effects of buffer width on the amount of chick- food available within the hedges.
The effect of woody species on arthropod abundance
There were significant differences among the numbers of individuals in the arthropod taxa found in the five species of hedgerow woody plants. For the arthropods which showed significant responses to buffer width at either order, suborder or family levels, differences in their number or diversity among woody species are listed below.
Araneae
In period 1 spider numbers varied significantly with the woody species sampled (F=3.34, df=4, 175.1, P=0.01) (Fig. 3.13a). Elderberry held fewest spiders, though not significantly different from rose (Tukeys test for pairwise comparisons, DF=168-179, P= 0.05). In period 2, the spider numbers in the different tree species did not differ significantly. Hawthorn held significantly more spiders than the other four woody species (Tukeys test for pairwise comparisons, df=168-179, P= 0.05). Among those hazel was superior to elderberry. Finally, in period 3 hawthorn and hazel both have significantly more spiders than elderberry (Fig. 3.13a)(Tukeys test for pairwise comparisons, df=169-179, P=0.05).
Hemiptera
For Hemiptera in period 1, numbers varied significantly with the woody species sampled (F=11.6, df=4, 179, P<0.0001). The number of Hemipterans in woody species could be ranked as follows: hawthorn > rose = hazel = blackthorn > elderberry (Tukeys test for pairwise comparisons, P=0.05)(Fig. 3.13b). In period 2 there was again a significant effect of woody species on Hemipteran numbers (F=2.52, df=4, 174.4, P=0.0429). The number of Hemipterans in woody species in period 2 could be ranked as follows: blackthorn > hawthorn > hazel = rose = elderberry (Tukeys test for pairwise comparisons, P≤0.05). In period 3 (F=3.40, df =4, 176.9, P=0.0105), blackthorn, hazel and hawthorn all had more Hemiptera than elderberry (Fig. 3.13b)(all differences given are Least square means, P<0.05 or less).
Coleoptera
In period 1, the Coleopteran numbers found were significantly different depending on tree species (F=6.2, df=4, 175.6, P<0.0001): most Coleopterans were found in hawthorn > hazel = rose = blackthorn > elderberry (Fig. 3.13c) (Tukeys test for pairwise comparisons, P=0.05). In period 2 (June) the Coleopteran numbers in the different tree species did not differ significantly. In period 3 (F=3.37, df =4, 172.3, P=0.01) rose and blackthorn had significantly more Coleoptera than hawthorn (Fig. 3.13c) (rose-elderberry: t=2.68, df =167.7, P=0.001, blacktorn-hawthorn: t=2.47, df =176.3, P=0.01).
3.2.2 Hedge bottom and field
3.2.2.1 Buffer width effects on avian species recorded by transect counts
A total of 3,029 Lepidoptera and Apidae observations were recorded during transect walks.
Effects on activity of Lepidoptera (butterflies) and Bombus (bumblebees)
The results of the statistical analysis on activity are presented in Tables 3.7-8. Only figures of Lepidoptera and Bombus are presented in this section. The activity of Apidae (bees) was low within the field which restricted the possibilities to carry out reliable statistical analyses. Bumblebee data were therefore only analysed on data sampled in period 3 (July) close to the field edge. More information on Apidae counts can be found in Table D.9 in Appendix D.
Lepidoptera activity (no. observed per 10 min.) was significantly affected by field, distance, buffer and distance × buffer (Table 3.7). The temperature, time and sampling day, together with relevant combinations of these, did also significantly affect the activity in either period 2 or 3 or both periods (Table 3.7). The activity was positively correlated with temperature and effect of sampling time during the day varied for the specific sampling dates (Table 3.8).
In period 2 (June to early July) a 16 times higher Lepidoptera activity was found 5 m from the field edge in buffer 24 compared to buffer 0. At 9 m, the activity was 12 times higher in buffer 24 compared to buffer 0 and 18 m from the field edge an up to 10 times higher activity was found in buffer 24 compared to the other treatments at similar distance in period 2 (Fig. 3.14).
In period 3 (July) a higher activity of Lepidoptera was found at all distances were a buffer zone was present (Fig. 3.14). 2 m from the edge, a significantly higher activity was estimated in buffer 12 with two times higher activity compared to buffer 0. At 5 m, butterflies were three to four times more active when a buffer zone was present. A similar pattern could be found at 9 and 18 m, and the relative difference became higher at increased distance (Fig. 3.14).
Among the various butterfly genera recorded in the present experiment, Pieris (whites) was sufficient numerous for a separate statistical analysis, and this genus responded positively to buffer zones (Table 3.7). For more information on the genus see Fig. D.1 and Table D.1 in Appendix D.
Buffer had no significant effect on bumblebee activity (Table 3.7). In Appendix D, Fig. D.2, the activity in period 3 is illustrated. For Bombus activity in relation to flower densities within the hedge bottom see section 3.3.1.
Effects on biodiversity of Lepidoptera and Bombus
As Lepidoptera and Bombus may be suitable bioindicators, and also identified to species in this study, they were used for estimating buffer zone effects on biodiversity. In total 13 species of Lepidoptera and four species of Bombus were observed. All species observations are found in Table D.9, Appendix D. The statistical analyses on biodiversity of Lepidoptera and Bombus are presented in Table 3.9.
There was a highly significant effect of buffer on species richness of Lepidoptera which could be estimated independently of the interactions distance × buffer and period × buffer, although the interaction with distance was nearly significant. The effect of buffer width on biodiversity estimated with Shannon’s H was also significant; an effect which also in this case was not affected by distance and sampling period (Table 3.9).
In Fig. 3.15 the biodiversity of Lepidoptera is illustrated in relation to buffer zones and distance to field edge. Within all three sampling periods, the average no. of Lepidoptera species seems to be correlated with the Shannon’s H diversity. The general trend is higher species richness and Shannon’s H in relation to increased buffer zone width and closeness to hedge (Fig. 3.15). A very clear effect of buffer width was found (independent of distance and time period) with significantly higher species richness in buffer 6, 12 and 24 m compared to buffer 0 and 4 m(for pair-wise comparisons see Table D.4 in Appendix D). A 55% higher species richness was estimated in buffer 6 compared to buffer 0 and a 45% higher species richness was estimated in buffer 24 compared to buffer 6 (Table D.5 in Appendix D). Furthermore, a buffer 6 significantly increased the species richness close to the hedge (0 – 4 m from the hedge indicated as dist. 2 see Fig. 3.15 and Table D.4 in Appendix D).
Estimated with Shannon´s H, the butterfly diversity was significantly higher in buffer 12 and 24 compared to buffer 0 and 4 and a significantly higher diversity (H-value) was also estimated in buffer 12 compared to buffer 6 (for more pair-wise comparisons see Table D.8 in Appendix D). Looking at the individual sampling periods, the clearest buffer effects was found in Period 3 (Fig. 3.15, Table D.8 in Appendix D). For more information on Shannon’s H estimates see Table D.6 in Appendix D.
These above results indicate, that butterfly may be suitable bioindicators for buffer zone effects (see section 3.4.2 and discussion).
There was no significant effect of buffer zones on biodiversity of bumblebees (Table 3.9), so no further information on this order is presented here. However, more information on Bombus can be found in Appendix D in Tables D.2 and D.9 and in section 3.4.1, where a relationship between bumblebees and flowering is presented.
3.2.2.2 Buffer width effects on herbaceous-dwelling arthropods
A total of 62,564 target arthropods from the sweep samples were identified to various taxonomical levels e.g. order, family, species and for some arthropods a distinction between stage, e.g. adult or juvenile, was made. This resulted in a total of 232 different Taxonomical Units (TU) from the sweep samples. The specific taxonomical levels to which the arthropods were identified are found in chapter 2 in Appendix D, Table D.20. The dominating arthropod groups not included were aphids, Diptera (except Syrphidae) and Collembola. Condensing the sweep data into a simple general format (in this case present or absent in the buffer plots across sampling periods) for the 232 TU, resulted in descriptive information not included in the statistical analyses presented in the following sections.
Of the 232 different TU identified, 197 were found in the hedge bottom and 71 of these were exclusively found here. In the field 161 TU were present. 153 of these could be found in the buffer areas, while 95 TU could be found in the treated areas. 35 TU were exclusively found in the field area with 27 TU found inside the buffer area, 3 in the treated area and 5 were common to both buffer and treated field area. This suggests that 27 TU have been gained by leaving a buffer in the field. The hedge bottom and the buffer area have 39 TU exclusively in common, suggesting that these 39 TU have successfully expanded their habitat area from the hedge bottom into the buffer area of the field. Considering the field area only, 66 TU are exclusively found in the buffer areas of the field, whereas 8 TU are solely found in the treated field area.
The TU of selected arthropods groups and total TU on buffer zone basis are given in Table 3.10
Table 3.10. Schematic summary of selected arthropod groups (TU) from the descriptive results on presence/absence data from sweep samples.
| Buffer 0 | Buffer 4 | Buffer 6 | Buffer 12 | Buffer 24 | |
| Chrysomelidae | 9 (0) | 10 (2) | 13 (1) | 13 (1) | 15 (3) |
| Cucujoidea | 10 (0) | 13 (3) | 12 (1) | 16 (2) | 15 (2) |
| Coleoptera | 58 (5) | 71 (12) | 81 (14) | 72 (9) | 68 (8) |
| Miridae | 16 (0) | 18 (0) | 21 (2) | 23 (4) | 21 (2) |
| Heteroptera | 26 (1) | 28 (0) | 31 (3) | 36 (6) | 33 (3) |
| Total TU | 125 (4) | 139 (12) | 153 (17) | 157 (15) | 150 (13) |
Numbers in () are Taxonomical Units exclusive to the particular buffer zone
The results in Table 3.10 indicate, that a maximum total number of TU caught by sweeps within the experimental plots may be reached with a 6 to 24 m buffer zone.
Effects on abundance of herbaceous-dwelling arthropod taxa
The results of the statistical analyses of individual arthropod taxa relatively abundant in sweep samples are summarized in Table 3.11. The analyses comprise 13 higher taxa (family or above) representing many more species (See Table D.20 in Appendix D). The taxa analysed all constitute important parts of the fauna in arable fields, and several have earlier been used to estimate effects of reducing agro-chemicals in arable crop edges (Frampton & Dorne 2007).
Table 3.11 shows that the abundances of all five orders analyzed were significantly affected by buffer width and the distance from hedge in at least two of the three sampling periods. In most cases significant effects were also found for the interaction between these two factors (buffer × distance). The majority of the sub-groups analyzed were also affected by buffer, distance and buffer × distance.
Only figures of the five orders analysed (Table 3.11) for abundance in relation to buffer width are presented in this section. Figures of the lower test taxa, which responded significantly to ‘buffer’ in at least one of the three time periods, can be found in chapter 2 in Appendix D, Figs. D.3-8. Many of the taxa included in Appendix D represents dominating sub-groups of the higher taxa presented here.
In Fig. 3.16 the buffer zone effects on Hemiptera (plant sucking insects such as true bugs) are presented. Aphids (which belong to this order) were not counted and therefore not included in this analysis. Hemipterans constitute an important part of the arthropod fauna both as beneficial and pests, and many species are important components of the chick-food diet for farmland birds.
At all three sampling periods, buffer zones increased the abundance of hemipterans in the hedge-bottom (Fig. 3.16). For obtaining a significant effect, a 24 m buffer was needed in period 1 but in periods 2 and 3, a 4 m buffer was sufficient for considerably higher abundance (between three and 11 times higher) in the hedge bottom compared to buffer 0 (Fig. 3.16). Within the field, the abundance of hemipterans was lower, but the general pattern was a significantly and several times higher abundance within the buffer zones at all distances and all sampling periods, especially after insecticide applications. In the field there was no significant effect of distance within buffer area.
Coleoptera (Fig. 3.17) is a very diverse order representing 28 beetle families in the present study (Table D.20 in Appendix D). This order includes among others the plant feeding Chrysomelidae and Curculionidea which later in this section are used to estimate biodiversity effects as many species are related to specific plants.
In period 2, there was a significantly and seven times higher abundance of coleopterans in the hedge-bottom at buffer 24 compared to buffer 0 (Fig. 3.17). However, in all parts of the hedge bottom guarded with a buffer strip there was a tendency towards higher abundance compared to buffer 0. In the field there were generally more coleopterans in buffer strips compared to treated field area. In period 3, more coleopterans were generally found within field buffer strips than in treated field area (Fig. 3.17). There was no effect of distance on abundance within the buffer areas in the field.
For Hymenoptera (the order comprised mainly of beneficial parasitic wasps (see Table D.20 in Appendix D), there was a tendency towards increased numbers in the hedge bottom (dist. 0) at increased buffer width at sampling period 1(Fig. 3.18).
In the field however, the Hymenoptera abundance in period 2 and 3 was several times higher in the buffer zones compared to treated field at all distances (Fig. 3.18). Only in buffer 24 there was an effect of distance on abundance of Hymenoptera within buffer strips in the field (Fig. 3.18) with a higher abundance at distance 2 m compared to 24 m. There was no buffer effect on abundance outside the buffer strips.
For Diptera, only the family Syrphidae (hoverflies) was counted and the effect of buffer zones was very similar to the effects on Hymenoptera. There was no buffer effect on abundance within the hedge bottom (although there was a weak tendency towards higher abundance in the hedge bottom at increased buffer width in period 3). In the field however there were several times higher abundances within the buffers at all distances. As for Hymenoptera, there was no buffer effect on abundance of Syrphidae outside the buffer strips (Fig. 3.19).
For Thysanoptera (trips) buffer zones of 6, 12 or 24 m increased the abundance in the hedge bottom, and most markedly in period 3 (Fig. 3.20). In the field there were several times higher abundances within the buffers than outside them at all distances. There was no buffer effect on abundance outside the buffer strips.
Effects on biodiversity of herbaceous-dwelling arthropods
Among the sweep-caught arthropods, two taxa, Heteroptera and Coleoptera, with specific plant preferences, were used to estimate biodiversity effects of buffer width. Both taxa responded significantly to buffer zones in terms of abundance (Table 3.11.). Among the coleopterans, Chrysomelidae and Curculionidea were included. The species of these two families have specific plant preferences. Therefore, a high diversity of these species may also indicate high plant diversity. The statistical analyses are presented in Table 3.12.
Buffer width had a highly significant effect on the species richness of heteropterans but the size of the effect depended both on the distance from hedge and sampling period (Table 3.12). In period 1, there were no significant differences between buffer zones (Fig. 3.21, Table D.13 in Appendix D.). In period 2, significantly more Heteroptera species (P<0.05) were found in buffer 6 and 24 compared to buffer 0 at the hedge bottom (distance 0 m) (Fig. 3.21). At 2 m more species were estimated at buffer 12 and 24 compared to buffer 0 (P<0.05). At 5 m more species were found at buffer 6, 12 and 24 compared to buffer 0, and at 9 m more species were caught in buffer 12 and 24 compared to buffer 0. Buffer 4 did not increase the number of species significantly at any distance compared to buffer 0 in period 2 (Fig. 3.21, Table D.13 in Appendix D). In period 3, there were no differences in species richness within the hedge bottom. At distance 2 m, significantly more Heteroptera species were found in buffer 4, 6, 12 and 24 compared to buffer 0 (Fig. 3.17). At 5 m, more species were estimated in buffer 6, 12 and 24 compared to buffer 0. At 9 m, more species were found in buffer 12 and 24 compared to buffer 0 and at 18 m more species were found at buffer 24 compared to buffer 0. For more pair-wise comparisons of species richness of heteropterans at combinations of sampling period, buffer width and distance see Table D.13 in Appendix D.
The biodiversity of heteropterans measured by Shannon’s H index (H-value) was quite similar to species richness with buffer width having a highly significant effect on the H-value (Table 3.12 and Fig. 3.21). In period 1, there were no significant differences between any buffer width at any distance (Fig. 3.21, Table D.15 in Appendix D). In period 2, a significantly higher H-value (P<0.05) was found in buffer 24 compared to buffer 0 and 4 at the hedge bottom (distance 0 m) (Fig. 3.21) Buffer widths less than 24 m did not increase the H-value significantly at distance 0 in period 2. In period 3, there were no differences in species richness within the hedge bottom. At distance 2 m, a significantly higher H-value was found in buffer 4, 6, 12 and 24 compared to buffer 0 (Fig. 3.21). At 5 m, higher H-values were estimated in buffer 6, 12 and 24 compared to buffer 0. At 9 m, the H-value was higher in buffer 12 and 24 compared to buffer 0, and at 18 m, a higher H-value was found at buffer 24 compared to buffer 0. For more pair-wise comparisons of Shannon’s H for heteropterans at combinations of sampling period, buffer width and distance see Table D.15 in Appendix D.
In summary - in sampling periods 2 and 3 (after insecticide applications) significantly higher biodiversity of heteropterans (measured both as species richness and Shannon’s H) was generally found within buffer zones compared to treated field area at all distances.
In Fig. 3.22 the pooled biodiversity of the Coleoptera families Chrysomelidae and Curculionidea is illustrated in relation to sampling period, buffer width and distance to field edge.
There were no significant differences on species richness of selected Coleoptera families in period 1 (Fig. 3.22). In period 2, there was no difference in species richness in the hedge bottom (Appendix D Table D.17). At 2 m, species richness at buffer 0 was significant lower than the other buffer zones. At 5 m, species richness at buffer 0 and 4 were significantly lower compared to buffer 6, 12 and 24. At increased distance, the smaller buffer widths became more similar to buffer 0 and 4. The results in period 3 were quite similar to period 2. In period 3, there was no effect on species richness in the hedge bottom either, and generally the results were comparable to period 2 (for all comparisons in periods 2 and 3 see Table D.17 in Appendix D).
The biodiversity measured by Shannon’s H in period 1 revealed a significantly higher biodiversity at the hedge bottom at buffer 0, 6, 12 compared to buffer 24 (Fig. 3.22) (in line with the lower plant diversity at buffer 4 and 24 – see Table 3.3, section 3.1.2). In period 2, there were no significant differences. In period 3, the diversity was higher at the hedge bottom at buffer 6 and 24 compared to buffer 0 and 4. At 2 m, a higher diversity was found for buffer 6 and 12 compared to buffer 0. At 5 m there was significantly difference between buffer 0 and 24. Significantly differences were also found at 9 m for buffer 4 versus buffer 12 and 24 and between buffer 6 and 24 (for more information on the comparisons of Shannon’s diversity H see Table D.19 in Appendix D).
In Tables D.16 & 18 in Appendix D, the 95% confidence limits of estimated species richness and Shannon’s H biodiversity of Chrysomelidae and Curculionidea are presented.
Chick-food in sweep net samples
Buffer width significantly affected the quantity of chick-food estimated from sweep net data in periods 2 and 3 (Table 3.13).
Table 3.13. Schematic summary of the statistical analyses on important chick-food arthropods (see section 2.4.2.2) caught by sweep netting.
| Per.² | Test results F(ndf,ddf)P¹ | ||||
| Field³ | Distance4 | Buffer5 | Buffer × Distance6 |
||
| Chick-food | 1 | 1.70(3, 15)NS | 13.16(4, 12)*** | 0.08(4, 11)NS | 1.52(16, 40)NS |
| 2 | 8.05(3, 13)** | 0.26(4, 13)NS | 11.40(4, 48)*** | 0.75(16, 47)NS | |
| 3 | 3.49(3, 11)NS | 19.23(4, 11)*** | 90.00(4, 363)*** | 11.02(16, 369)*** | |
¹ NS not significant, *P < 0.05, **P < 0.01, ***P < 0.001. F is the F-value, ndf and ddf is the numerator and denominator degree of freedom used for testing the significance.
² Three sampling periods (Per.): 1. After herbicide application (May), 2. After first insecticide application (June), 3. After second insecticide application (July).
³ Effect of field (four fields were included in the experiment).
4 Effect of distance from field edge (sampling was carried out 0, 2, 5, 9 and 18 m from the field edge).
5 Effect of buffer width (0, 4, 6, 12 and 24 m).
6 Effect of the combination of distance and buffer width (in total there were 5 × 5 = 25 combinations).
In period 1(after herbicide application in May) there was no significant effect of buffer width on available chick-food. A considerable amount of chick-food was only found in the hedge bottom. In period 2 (after insecticide application) significant more food prey was estimated with up to nine times more available food in both hedge bottom bordering a buffer zone and the buffer zones. In period 3 (after the second insecticide application), the overall trend was similar to period 2, but within the field the relative difference between buffer zones and treated field was markedly higher with up 60 times higher food-mass in buffer area. There was no significantly effect of distance within the buffer strips (Fig. 3.23).
3.2.2.3 Buffer width effects on epigaeic arthropods
A total of 25,179 arthropods were identified from pitfall samples.
Epigaeic (ground-dwelling) arthropods, primarily Araneae (spiders), Carabidae (ground beetles) and Staphylinidae (rove beetles) are normally relatively abundant in agricultural fields. Many species are important beneficials preying on agricultural pests and may be of economic importance for the farmers (Östman 2003). A high density and diversity are therefore considered important, although a few species may act as crop pests.
Effects on individual epigaeic arthropod taxa
In Table 3.14, the statistical analyses on abundance in relation to buffer width are presented.
Only figures of the higher taxa Araneae (spiders) and Carabidae (ground beetles), which both responded significantly to buffer (Table 3.14), are presented in this section. Figures of the remaining test taxa, which responded significantly to ‘buffer’ in least at one of the three periods (see Table 3.14), can be found in chapter 3 in Appendix D, Figs. D.10-12.
In periods 1 and 2, the presence of a buffer zone did not affect the Araneae activity in the hedge bottom significantly, although there was a tendency towards higher abundance at increased buffer width (Fig. 3.24).
In period 1, there were in some cases significantly higher Araneae activity outside the buffer zones at the distances 2 and 5 m from the field edge (this may be due to a denser and higher crop outside the buffer zones more suitable to spiders – see Table 3.2). In period 2 (after the first insecticide application) the activity of Araneae was several times higher within the buffer zones compared to treated field (Fig. 3.24). In period 3, the activity was generally higher in hedge bottom protected by a buffer zone (Fig. 3.24). In the field the activity of Araneae was always significantly higher in buffer zones than in the treated field. Distance from field edge within buffer zones did generally not affect the activity level of Araneae within any of the three sampling periods. Probably caused by general population cycles of Araneae, there was a drop in abundance within the hedge bottom in periods 2 and 3 (June – July).
Buffer zones did not affect the activity of Carabidae in period 1 (Table 3.14, Fig. 3.25). In period 2, there was a tendency towards higher activity at 2 and 5 m within the buffer zones (Fig. 3.25). 9 m from the edge there was a significantly higher activity in buffer 12 and 24 than at buffer 0. In period 3, significantly higher carabid activity was estimated in the buffer zones 2 m from the edge compared to buffer 0. At the higher distances, the general pattern was a tendency towards more carabids when a buffer zone was present (Fig. 3.25). At 9 m, there was a significantly higher abundance at buffer 12 and 24 compared to buffer 0. At 18 m, the carabid abundance at buffer 24 was significantly higher that at buffer 0 and 4.
There was a tendency towards higher carabid activity up to 200 m into the treated field from the nearest buffer edge (Fig. 3.25).
Staphylinid abundance was not affected significantly by buffer width, but significant interactions between buffer width and distance were found for some combinations of sampling run and subfamily/genus (Table 3.14).
Buffer zone effects on biodiversity of epigaeic arthropods
As buffer zones had a positive effect on the abundance of Araneae and Carabidae in this trial (Figs. 3.24 and 3.25), these two taxa were used to estimate effects of buffer zones on biodiversity of epigaeic arthropods. The results of the statistical analyses are presented in Table 3.15.
For Araneae families, there was a highly significant effect of buffer and buffer × sampling period on family richness (“family” because Araneae were only identified to this taxonomic level). Biodiversity estimated with Shannon’s H was not affected significantly by buffer zones (Table 3.15), maybe because the family Linyphiidae was very dominating (see Table D.24 in Appendix D).
In Fig 3.26, the biodiversity of Araneae families is presented. In period 1 there were no significant differences. In period 2 and 3 there were no differences on family richness at the hedge bottom. In period 2, differences in family richness were only found from 5 m and outwards. At 5 m, buffer 12 and 24 had a significantly higher family richness compared to buffer 0. At 9 m, buffer 24 had a significantly higher family richness compared to buffer 0, 4 and 6. 18 m from the field edge, buffer 24 had a higher family richness compared to buffer 0, 4 and 12. Most significant differences were found in period 3. In this period, significant differences between the buffer zones started from distance 2 m and outwards. Within the five distances, buffer area had always a significantly higher family richness, with the exception that there was no difference between buffer 12 and 24 at distance 18 m (Fig. 3.26) (for more specific information on significant effects of family richness see Table D.23 in Appendix D).
There were no significant effects of buffer or buffer × distance on the biodiversity of Carabids (Table 3.15). In period 2 and 3, however, there was a tendency towards a higher species richness and biodiversity measured with Shannon’s H-value within the field when a buffer zone was present (Fig. 3.27).
3.3 The marginal gain of diversity at increased buffer width
Estimating the accumulated number of species at increased distances from the hedge is a simple method to provide information on how much more biodiversity that can be gained by widening buffer zones. The method can be used to establish the buffer zone width, where gains (defined as new species) do not increase further when widening of the buffer zones. Another method is to estimate the power form of the species-area relationship - called SPAR by Rosenweig (2003) or SAR by Desmet & Cowling (2004). Such a power equation can be used to interpolate or extrapolate the effect on biodiversity of any given buffer width. For more information on the models, see section 2.6.3.2 and Appendix F – models 13 and 15). In the two sub-sections below, the results of both methods are presented.
Wild plants were included as test organisms for biodiversity effects of buffer zone width, taking species – area relationships into considerations. Among the Arthropods, Heteroptera (true bugs), herbivorous coleopterans (leaf beetles and weevils), Carabidae (ground beetles) and Lepidoptera (butterflies) were selected. These taxa were relatively abundant in the present experiment and Heteroptera and Carabidae had the highest species richness among the test taxi. Heteroptera is a relatively immobile but important part of the fauna in many crops, and due to their sensitivity to ecological factors they may be good bioindicators (Fauvel 1999). The herbivorous Coleopterans, Chrysomelidae and Curculinoidea, are possible suitable bioindators with medium dispersal ability. Carabidae are species rich and abundant in arable sites. They are less dependent on plants and relatively mobile compared to Heteroptera and the herbaceous-dwelling beetles. Some carabid species are bound to or prefer the field boundary, other species hibernate in field edges vegetation and disperse into the field during spring and some species hibernate within the field during winter (Kromp 1999, Fournier & Loreau 1999). As carabids are affected by agricultural cultivation e.g. by weediness and field boundaries, they are considered of bioindicative value for cultivation impacts (Kromp 1999). Lepidoptera is a well studied taxa, wich has been under a huge pressure in the arable land. Lepidoptera serve as a general bioindicator (Thomas 2005). Lepidoptera are highly mobile compared to the other test taxa. This mobility may cause species richness to be more dependent on changes at landscape scale rather than at a local scale (Rundlöf et al 2008). However, if they respond on a local scale they may be considered a strong indicator of habitat changes. The test taxa (wild plants, Heteroptera, herbivorous coleopterans, Carabidae and Lepidoptera) have different habitat requirements and this, in combination with dispersal ability, makes them suitable taxa for studying general distance-buffer width interactions on biodiversity, also at a local scale.
3.3.1 Accumulated number of species at increased distance to hedge in relation to buffer width
The analyses were carried out on the July data (data from the last sampling rounds), where the experimental plots had received the full chemical treatments. The results of the statistical analyses are presented in Table 3.16.
Table 3.16. Schematic summary of the statistical analyses on accumulated species richness at increased distance from the hedge of taxa selected as bioindicators. The analyses were carried out on the July data.
| Test taxa | Fixed effect | Test results F(ndf,ddf)P¹ | |||||
| Distance 0 | Distance 0-2 m | Distance 0-5 m | Distance 0-9 m | Distance 0-18 m | |||
| Wild plants | Buffer | 1.01(4, 12)NS | 5.30(4, 12)* | 1.93(4, 12)NS | 2.23(4, 12)NS | 3.16(4, 12)(0.05) | |
| Field | 10.26(3, 12)** | 10.15(3, 12)** | 4.90(3, 12)* | 7.56(3, 12)** | 5.74(3, 12)* | ||
| Heteroptera | Buffer | 2.83(4, 12)NS | 6.81(4, 12)** | 6.99(4, 12)** | 6.05(4, 12)** | 6.05(4, 12)** | |
| Field | 13.14(3, 12)*** | 6.47(3, 12)** | 6.50(3, 12)** | 4.17(3, 12)* | 4.17(3, 12)* | ||
| Herbivorous | Buffer | 0.59(4, 11)NS | 4.29(4, 12)* | 8.57(4, 12)** | 4.27(4, 12)* | 2.87(4, 12)(P=0.07) | |
| coleopterans² | Field | 7.13(3, 11)** | 5.72(3, 12)* | 7.54(3, 12)** | 2.52(3, 12)NS | 1.43(3, 12)NS | |
| Carabidae | Buffer | 0.33(4, 12)NS | 1.14(4, 12)NS | 1.50(4, 12)NS | 0.36(4, 12)NS | 0.49(4, 12)NS | |
| Field | 6.63(3, 12)** | 3.78(3, 12)* | 4.43(3, 12)* | 4.10(3, 12)* | 5.24(3, 12)* | ||
| Lepidoptera | Buffer | -³ | 2.45(4, 12)NS | 1.40(4, 12)NS | 1.08(4, 12)NS | 1.17(4, 12)NS | |
| Field | - | 3.35(3, 12)NS | 1.89(3, 12)NS | 1.06(3, 12)NS | 0.93(3, 12)NS | ||
¹ NS not significant, *P < 0.05, **P < 0.01, ***P < 0.001. F is the F-value, ndf and ddf is the numerator and denominator degree of freedom used for testing the significance.
² The herbivorous families Chrysomelidae and Curculinoidea only.
³ Lepidoptera were not recorded at distance 0 specifically, but in the edge zone (hedge – 4 m within the field).
The accumulated species richness of wild plants estimated at distance 0-18 m (the species richness for the entire plot areas – see Fig. 2.2) was significantly lower at buffer 0 compared to buffer 6, 12 and 24 (P<0.05) (Fig. 3.28 and Table E.5 in Appendix E). Furthermore, a buffer zone wider than 6 m did not lead to significantly more weed species (Table E.5 in Appendix E). This indicates, that a buffer zone width of 6 m is needed in order to increase the biodiversity in terms of species richness of wild plants in the field, but also that a buffer width higher than 6 m will not significantly increase the species richness of wild plants. There was no overall significant buffer effect on weed species richness at the intermediate accumulated distances 0-5 m and 0-9 m, although some pair-wise significant differences between narrow and wider buffer zones could be identified (see Tables E.3-4 in Appendix E). Buffer zones had no effect on species richness within the hedge bottom (Fig. 3.28).
Buffer width had also a significant effect on the accumulated species richness of Heteroptera (Table 3.16, Fig.3.28). The accumulated species richness estimated at distance 0-18 m (the species richness for the entire plot areas) showed that species richness at buffer 0 was significantly lower than at any other buffer width. The other buffer zone widths were not significantly different (P≥0.05), indicating that a buffer zone wider than 4 m did not lead to significantly more Heteroptera species (Table E.10 in Appendix E). The accumulated number of heteropteran species at buffer 0 was also significant lower compared to the other buffer widths at the intermediate accumulated distances (P<0.05). Furthermore, a buffer width of 4 m or wider, resulted in higher species richness of heteropterans within the hedge bottom compared to buffer 0 (Table E.6 in Appendix E).
The analysis of the pooled species richness of the herbivorous coleopteran families Curculinoidea and Chrysomelidae revealed, that when the entire plot area (0-18 m) was analysed, a buffer width of 6 m was needed in order to secure a significantly higher species richness of these coleopterans compared to plots without buffer zones (P<0.05) (Fig. 3.28, Table E.15 in Appendix E). The other buffer widths were not significantly different (P≥0.05), indicating that a buffer zones wider than 6 m did not lead to more species (Table E.15 in Appendix E). Furthermore, buffer zones did not result in significantly higher species richness of herbivorous coleopterans within the hedge bottom compared to buffer 0 (Table E.11 in Appendix E), although there was a tendency towards higher species richness in the hedge bottom when a buffer zone was present (Fig. 3.28).
For both Carabidae and Lepidoptera the accumulated number of species did not depend significantly (P≥0.05) on the width of buffer zones at any of the 4 accumulated distances in this analysis (Table 3.16) (Tables E.20 and E.24 in Appendix E). However, there was a general tendency towards higher species richness of both Carabidae and Lepidoptera at increased buffer width (Fig. 3.28). Furthermore, buffer zones had no effect on species richness of Carabidae within the hedge bottom (Fig. 3.28, Table E.16 in Appendix E). For Lepidoptera, a buffer width of 6 m significantly increased the species richness close to the hedge (0-4 m indicated as distance 2 in Fig. 3.28, see also Table E.21 in Appendix E).
3.3.2 Species-Area Relationship (SPAR)
For convenience of reading, the SPAR model is presented below, but for more information on the power equation and its use please see section 2.6.3.2.
Click here to see: The SPAR model
The species richness data used for parameterization were the sum of species across all sampling times and across all sampling areas (hedgerow, hedge bottom and field). The parameter estimates of a and ß (Table 3.17) can be used as estimates for a-diversity and ß-diversity (Pollnac et al. 2009). a-diversity (as estimated by the y-intercept) indicates the plot-scale diversity in the hedge bottom (measured as species richness) for the experimental plots containing the various buffer width. ß-diversity is a measure of the change in species richness across spatial scales (distance in plot units).
For wild plants (including distance 0) the model did not describe the data satisfactory, as the model systematically overestimated the number of species in the hedge and systematically underestimated the number of species at distances 2 and 5 m from the hedge (Table 3.17 and Fig. 3.29). The reason for that is most probably, that the species present in the hedge bottom and in the field are very different (few species are present both in the hedge bottom and in the field).
When distance 0 was excluded, the model fitted much better, but although significant effects between buffer widths were found, they were all small (Table 3.17 and Fig. 3.29). There seemed to be a tendency towards increased a-diversity with increased buffer width and towards increased ß-diversity when a buffer was present for buffer widths higher than 4 m.
For heteropterans (true bugs), a buffer width of 4 m was enough to secure a significantly higher a-diversity (species diversity within the hedge bottom) compared to non-buffered hedge bottom. A buffer 24, however, gave a markedly higher a-diversity (Table 3.17 and Fig. 3.29). The ß-diversity of buffer 12 and 24 was significantly higher than at buffer 4, meaning that widening the buffer zones may offer more niches for heteropteran species leading to increase species richness (Table 3.17). Overall, the estimated species diversities at distance 18 m (the species richness of the entire plots) were very similar at buffer 4, 6 and 12 and noticeably higher at buffer 24 compared to buffer 0 (Fig. 3.29).
For the herbivorous coleopterans (weevils and leaf beetles) there was no consistent trend for the a-diversity (species richness within the hedge bottom). Although a-diversity was significantly higher for buffer 4 and 6 compared to buffer 0, buffer 12 and 24 were not significantly different from buffer 0 (Table 3.17). Hence, the effects can best be described as a tendency towards higher a-diversity when the hedge bottom was bordered by a buffer zone (Fig. 3.29). The ß-diversity increased with increased buffer widths, indicating that more niches are offered for the herbivorous beetles at increased buffer widths (Table 3.17).
Neither the a-diversity nor ß-diversity of Carabidae (ground beetles) differed significantly between the buffer zones, but there was a tendency towards increased species richness when a buffer zone was present, and increased diversity at increased buffer widths (Table 3.17, Fig. 3.29).
For Lepidoptera, wider buffer widths (up to 6 m) significantly increased the a-diversity (species richness close to the hedge; 0-4 m indicated as distance 2 in Fig. 3.29) compared to buffer 0. Buffers wider then 6 m did not further increase the a-diversity. There was no general pattern in the estimated ß-diversity values for butterflies. Two single species observations in buffer 12 at distance 9 m may have led to the relatively high ß-diversity observed for this buffer width (see Appendix D, Table D. 9)
3.4 Combined flora and arthropod analysis
3.4.1 Activity of Lepidoptera (butterflies) and Bombus in relation to flower and host plant abundance
The activity of both butterfly species belonging to the genus Pieris (whites) and of bumblebees was significantly and positively correlated to flower density of thistle (Cirsium and Carduus) in hedge bottom (Tables 3.18-19). There was also a strong indication of a positive relationship between host-plant ground cover (Brassica cover) and the activity of Pieris (Tables 3.18-19). The activity of Pieris increased with thistle flowers in the hedge bottom and with host-plants up to a 6% cover.
The activity of bumblebees increased strongly with the number of flowers generally in the field and with thistle flowers (up to 15 flowers) in the hedge bottom (Tables 3.18-19).
Click here to see: Table 3.19. Estimated effects of the covariates (see Table 3.18).
3.4.2 Lepidoptera (butterflies) as indicator for biodiversity gains
As Thomas (2005) revealed that species richness of butterflies is a suitable bioindicator for terrestrial environmental changes, focus was on this taxa and we carried out statistical analyses (see section 2.6.3.3) on the relationship between butterflies and other test taxa used in the previous sections for estimating biodiversity effects of buffer zones. Furthermore, in search for other suitable bioindicators than butterflies, additional combinations of the test taxa were also included in the analyses (see Table 3.20). The analyses were carried out on July data (sampling run 2 for plants and sampling period 3 for arthropods).
There was an indication of a positive relationship between species richness of dicots (wild plants) and Lepidoptera (butterflies) but there were no significant indications of relationships between dicots and the other test taxa (Table 3.20). Furthermore, the species richness of Lepidoptera correlated positively also to the species richness of Heteroptera (true bugs), Carabidae (ground beetles) and herbivorous coleopterans (leaf beetles and weevils), the latter however only on treated field area (Table 3.20). The above results suggest that butterfly species richness is a suitable bioindicator for biodiversity effects of buffer zones, as it correlates to the majority of the taxa used to estimate effect of buffer zones on biodiversity.
The analyses were all carried out after removing effects of field and treatment. This was done in order to avoid relationships caused by differences between fields and treatment. If the relationships had been estimated across treatments, a significant and positive relationship would have been found for (almost) all pairs of taxa shown above.
Version 1.0 November 2009, © Danish Environmental Protection Agency